Branched-Chain Amino Acid Catabolism: Unique Segregation of Pathway Enzymes in Organ Systems and Peripheral Nerves Andrew J. Sweatt 1 Mac Wood 1 Agus Suryawan 1 Reidar Wallin 2 Mark C Willingham 3 Susan M. Hutson 1 1 Department of Biochemistry Wake Forest University School of Medicine Medical Center Blvd. Winston-Salem NC 27157 2 Department of Internal Medicine Section on Rheumatology Wake Forest University School of Medicine Medical Center Blvd. Winston-Salem NC 27157 3 Department of Pathology Wake Forest University School of Medicine Medical Center Blvd., Winston-Salem NC 27157 Running Title: Branched-Chain Amino Acid Catabolic Enzymes Contact Information: Andrew J. Sweatt Department of Biochemistry Wake Forest University School of Medicine Medical Center Blvd. Winston-Salem NC 27157 Tel 336-713-7201 FAX 336-716-7671 e-mail [email protected] Copyright (c) 2003 by the American Physiological Society. Articles in PresS. Am J Physiol Endocrinol Metab (September 9, 2003). 10.1152/ajpendo.00276.2003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Branched-Chain Amino Acid Catabolism: Unique Segregation of Pathway Enzymes in Organ Systems and Peripheral Nerves
Andrew J. Sweatt1
Mac Wood1
Agus Suryawan1
Reidar Wallin2
Mark C Willingham3
Susan M. Hutson1
1 Department of Biochemistry Wake Forest University School of Medicine Medical Center Blvd. Winston-Salem NC 27157
2 Department of Internal Medicine Section on Rheumatology
Wake Forest University School of Medicine Medical Center Blvd. Winston-Salem NC 27157
3 Department of Pathology Wake Forest University School of Medicine
Medical Center Blvd., Winston-Salem NC 27157
Running Title: Branched-Chain Amino Acid Catabolic Enzymes
Contact Information: Andrew J. Sweatt Department of Biochemistry Wake Forest University School of Medicine Medical Center Blvd. Winston-Salem NC 27157
Tel 336-713-7201 FAX 336-716-7671
e-mail [email protected]
Copyright (c) 2003 by the American Physiological Society.
Articles in PresS. Am J Physiol Endocrinol Metab (September 9, 2003). 10.1152/ajpendo.00276.2003

2
ABSTRACT
We have examined the localization of the first two enzymes in the branched-chain
amino acid catabolic pathway – the branched-chain aminotransferase (BCAT) isozymes
(mitochondrial BCATm and cytosolic BCATc) and the branched-chain -keto-acid
dehydrogenase (BCKD) enzyme complex. Antibodies specific for BCATm or BCATc were
used to immunolocalize the respective isozymes in cryosections of rat tissues. BCATm was
expressed in secretory epithelia throughout the digestive tract, with the most intense
expression in the stomach. BCATm was also strongly expressed in secretory cells of the
exocrine pancreas, uterus, and testis, as well as in the transporting epithelium of convoluted
tubules in kidney. In muscle, BCATm was located in myofibrils. Liver, as predicted, was not
immunoreactive for BCATm. Unexpectedly, BCATc was localized in elements of the
autonomic innervation of the digestive tract, as well as in axons in the sciatic nerve. The
distributions of BCATc and BCATm did not overlap. BCATm-expressing cells also
expressed the second enzyme of the BCAA catabolic pathway, BCKD. In selected monkey
and human tissues examined by immunoblot and/or immunohistochemistry, BCATm and
BCATc were distributed in patterns very similar to those found in the rat. The results show
that BCATm is in a position to regulate BCAA availability as protein precursors and anabolic
signals in secretory portions of the digestive and other organ systems. The unique expression
of BCATc in neurons of the peripheral nervous system, without co-expression of BCKD,
raises new questions about the physiological function of this BCAT isozyme.
Keywords not in title: digestive system, human, leucine, monkey, rat

1
INTRODUCTION
In the body, the nutritionally indispensable branched chain amino acids (BCAAs)
serve a number of important metabolic functions. BCAAs are key nitrogen donors for the
synthesis of the metabolically significant dispensable amino acids glutamine and alanine.
Glutamine is an important energy substrate for the gastrointestinal tract (38). Glutamine and
alanine are also the major carriers of nitrogen from amino acid oxidation in skeletal muscle
to the liver (7, 20, 33, 48, 56). In the central nervous system, BCAAs are thought to
participate in an intercellular shuttle between neurons and astroglia that provides nitrogen for
synthesis of the excitatory amino acid glutamate (3, 4, 31, 39, 40, 64). In addition to the role
of BCAAs in nitrogen metabolism, the BCAA leucine serves as an anabolic nutritional
signal. Leucine stimulates protein synthesis in selected tissues via activation of the
ribosomal protein S6 kinase-1 (12, 19, 42, 61). Furthermore, high physiologic concentrations
of leucine stimulate secretion of insulin, and it has been postulated that this effect occurs in
part via activation of glutamate dehydrogenase (43, 52).
The initial reaction in the degradation of most indispensable amino acids is essentially
irreversible, with excess amino acids being oxidized primarily in the liver. Thus, the major
fate of indispensable amino acids in peripheral tissues is synthesis into tissue proteins.
Catabolism of the BCAAs differs markedly from that of the other indispensable amino acids,
in that the first step is reversible. In addition, BCAA catabolic enzymes appear to be
distributed throughout the body, including in tissues of the digestive tract (18, 28, 36). The
physiological significance of BCAA metabolism in tissues other than skeletal muscle and
brain is not understood.

2
The first step in breakdown of the BCAAs is the reversible transfer of the -amino
group to -ketoglutarate to form glutamate and the respective branched chain -keto-acids in
a reaction catalyzed by the branched chain aminotransferase (BCAT) isozymes
(mitochondrial BCATm and cytosolic BCATc; reviewed in 23, 35). The next and first
irreversible step in BCAA catabolism is the oxidative decarboxylation of the branched chain
-keto-acid products of the transamination reaction. This step is catalyzed by the
mitochondrial branched chain -keto-acid dehydrogenase multi-enzyme complex (BCKD),
which contains multiple copies of three enzymes: a branched chain -keto-acid
decarboxylase (E1); a dihydrolipoyl transacylase (E2); and a dihydrolipoyl dehydrogenase
(E3). Activity of BCKD is regulated by phosphorylation/dephosphorylation of the E1
subunit, (21, 49). Based on observed tissue-specific differences in the activity of BCAT and
the BCKD complex, it is thought that oxidation of BCAA involves extensive movement of
metabolites between tissues (10, 24, 28, 30, 34, 53, 56).
The distribution of the BCAT isozymes in different tissues has been determined from
measurements of enzyme activity and by Western blot analysis of BCAT proteins. In the rat,
BCATm is found in most tissues, with very high BCAT activity found in the stomach,
pancreas, and salivary glands (36, 56). BCATc appears to have more limited expression than
BCATm. BCATc activity has been identified in rat brain, ovary, and placenta (18, 35). In
the rat, BCKD activity is found in most organs, with highest activities occurring in the liver
and kidney (56). In other organs, including the stomach, intestine and brain, BCKD activities
are up to two orders of magnitude lower than for liver and kidney (56).
The localization of the BCAA catabolic enzymes to particular cell types within a
tissue has been investigated only for cells of the central nervous system. Immunostaining of
cell cultures derived from rat brain revealed that BCAT isozymes are differentially

3
expressed, with BCATm expressed in astroglia, and BCATc expressed in neurons (4, 31, 40).
In contrast, BCKD appeared to be expressed in both cell types (3). In this study, we report
on the immunolocalization of BCAA catabolic enzymes to specific cell types in tissues
known to have BCAA catabolic activities or to express mRNA coding for the catabolic
enzymes. Particular attention was focused on tissues of the digestive tract, in which we
demonstrate expression of BCATc in peripheral nerves, without concomitant expression of
other BCAA catabolic enzymes.

4
MATERIALS AND METHODS
Tissues. Rat tissues (salivary, esophagus, stomach, pancreas, duodenum, jejunum,
ileum, colon, skeletal muscle, kidney, pancreas, ovary, uterus, testis, liver) were removed
from male and female Sprague Dawley or Long-Evans rats (225-250 g) and either flash
frozen in liquid nitrogen and stored at -80oC or cryo-embedded in OCT Compound (Sakura
Finetek, Torrance, CA) by freezing in liquid nitrogen and stored at -80oC. Monkey tissues
(brain, skeletal muscle, adipose, liver, kidney, pancreas, jejunum, stomach) were obtained
from monkeys involved in a study of the effect of dietary cholesterol on lipoprotein
metabolism (50). Portions of the monkey tissues had been used previously to measure tissue
BCAT and BCKD activities (56). Experimental procedures involving animals were
approved by the Institutional Animal Care and Use Committee of the Wake Forest University
School of Medicine. Human tissues (brain, muscle, adipose, liver, kidney, pancreas,
jejunum, heart) were collected as described in a previously published study (56). Specimens
were obtained from patients undergoing surgical procedures. All samples were obtained
from tissues that would have been discarded following pathological examination during
surgery. Stomach was obtained from a single organ donor who had given consent for use of
the tissue for research purposes. All tissues were flash frozen and stored in liquid nitrogen
until analyzed. The protocol for human tissues was approved by the Institutional Review
Board at Wake Forest University School of Medicine.
Antibodies. For identification of BCATm in rat tissues, a polyclonal antibody raised
in rabbits against purified human recombinant BCATm was used. Characterization and
affinity purification of this antibody have been described previously (40). With the exception
of rat stomach, where three different BCATc antibodies were tested to verify the localization

5
of BCATc (see Fig. 4), an antibody raised in rabbits against purified human recombinant
BCATc was used for the immmunolocalization of BCATc in rat tissues. Affinity purification
of this antibody is described below. The other two BCATc antibodies, used with rat
stomach, were an immunoaffinity purified rabbit anti-rat BCATc peptide antibody, directed
against the first 50 amino acids of the rat enzyme (40), and an IgG fraction of an antiserum
that was raised in rabbits against purified rat brain BCATc protein (18). These two BCATc
antibodies and the BCATm antibody have been used previously to identify BCATc and
BCATm in rat tissues or rat brain primary cell cultures by immunoblotting and/or by
immunohistochemistry (3, 4, 18, 31, 40). Antiserum for localization of BCKD was generated
against the purified E2 subunit of the rat liver BCKD complex, and was the gift of Dr.
Yoshiharu Shimomura (Nagoya Institute of Technology, Nagoya, Japan). An IgG fraction of
the E2 antiserum has been used previously to identify BCKD in primary cell cultures derived
from rat brain (3, 25).
For preparation of the affinity-purified human BCATc and E2 antibodies, the antisera
were made 50% saturated with ammonium sulfate, and centrifuged for 10 min at 10,000 xg.
The supernatants were discarded, and the pellets were washed in PBS. Subsequently, the
protein pellets were dissolved in PBS, followed by dialysis against PBS, and loaded onto an
affinity resin having human BCATc or rat liver BCKD complex as a ligand. The columns
were washed with PBS and the fraction of antigen-specific antibodies was eluted in 0.1 M
sodium acetate buffer (pH 4.0) containing 4 M urea and 0.5 M NaCl. The affinity purified
antibodies were dialyzed overnight against 50% glycerol/water at 4oC. The dialyzed
antibodies were aliquoted and stored at –80oC. Human BCATc-Sepharose and rat liver
BCKD-Sepharose were prepared by coupling the purified human recombinant BCATc or

6
purified rat liver BCKD complex to Affigel 10 support (Bio-Rad, Richmond, CA) according
to the manufacturer’s directions.
Immunoblotting. Tissues for SDS-PAGE/immunoblot were pulverized with mortar
and pestle while submerged in liquid nitrogen. Proteins were extracted from tissue powders
by three rounds of freeze-thaw-sonication in 25 mM HEPES (pH 7.4) containing 0.4%
CHAPS, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 10 ug/ml leupeptin, 5 mM benzamidine,
and 1 mM diisopropyl fluorophosphate. Proteins in aliquots of tissue extracts (10–60 µg
protein) were separated by SDS-PAGE using 10% gels. Purified recombinant human
BCATc and BCATm proteins (11) were included as standards. Proteins were transferred to
Immobilon P membranes. Membranes were blocked with 5% nonfat milk/PBS or 1%
BSA/PBS and incubated with immunoaffinity-purified rabbit anti-human BCATm (0.3-0.5
µg/ml), anti-rat peptide BCATc (0.4-0.6 µg/ml), or anti-human BCATc (0.4-0.6 µg/ml)
antibodies. The immunoreactive protein bands were visualized using the Enhanced
Chemiluminescence (ECL; for monkey and human tissue blots) or ECL Plus (for rat tissue
blots) detection system according to the manufacturer’s instructions (Amersham Biosciences,
Piscataway NJ), and detected on X-ray film (Amersham Biosciences). Immunoreactive
band intensities were analyzed for film exposures producing signals below saturation (bands
were translucent). Band intensities were quantified in scanned images of the film
(ImageQuant Software, Amersham Biosciences) and are reported as arbitrary units/µg protein
loaded. For quantification of BCATm in monkey tissues, band intensities for individual
samples from each tissue were compared to intensities of a series of purified recombinant
human BCATm or human BCATc standards (2-8 ng purified BCAT). Extract protein
concentrations were adjusted so that band intensities were within the linear range of the
BCAT standards.

7
Immunohistochemistry. Frozen sections, 6-8 µm in thickness, were collected
directly on slides or collected on adhesive tape and transferred to adhesive-coated slides
using a UV-crosslinking system (Instrumedics CryoJane System, Hackensack, NJ). Sections
were fixed by immersion in acetone (10 min), followed by lyophilization and storage at 25oC.
Sections were rehydrated by immersion in PBS for 10 min. Non-specific binding sites were
blocked by treatment with 1% BSA/PBS for 15 min. Sections were incubated with the
primary antibodies diluted to 5-10 µg/ml in BSA/PBS for 30-60 min, and rinsed three times
with PBS before incubation with horseradish peroxidase- or FITC-conjugated secondary
antibodies (Jackson ImmunoResearch Laboratories, West Grove, PA). After washing with
PBS, color was developed for HRP-conjugates using diaminobenzidine/H2O2. Controls
consisted of incubations of sections with secondary antibodies only, or with primary
antibodies that had been preincubated overnight with a 10-fold excess of competing antigen,
followed by secondary antibodies. In some experiments, immunolabeled sections were
counterstained with hematoxylin, a nuclear stain, to aid in the identification of cell types. To
visualize myelin in sciatic nerve, cryosections were fixed with 4% formaldehyde in PBS, and
stained with Oil Red O by standard methods (22). Tissues were viewed with a Zeiss
AxioPlan 2 microscope, and images were obtained using an AxioCam digital camera and
AxioVision imaging software (Carl Zeiss USA, Thornwood, NY). Images were adjusted and
assembled using Adobe Photoshop 6.0 (Adobe Systems Incorporated, San Jose CA).
RNA Extraction and RT PCR. RT-PCR was used to determine if BCATc mRNA was
present in tissues outside the brain that exhibited BCATc-specific immunostaining in nervous
elements. Flash-frozen tissue that had been powdered under liquid nitrogen (100-300 mg)
and stored at -80 oC was used for preparation of total RNA. Total RNA was extracted from
the frozen tissue powder using TriZol reagent according to manufacturer’s instructions (Life

8
Technologies, Rockville, MD). First-strand cDNA was synthesized using oligo(dT) primer
and SuperScript II reverse transcriptase (Gibco BRL, Gaithersburg MD) with 10µg RNA per
sample. The integrity of the cDNA was confirmed by PCR using primers for
glyceraldehyde-3-phosphate-dehydrogenase (G3PDH): sense primer: 5’-
CCTTCATTGACCTCAACTACATGG-3’; antisense primer: 5’-
TCCACCACCCTGTTGCTGTAGC-3’. Rat BCATc was amplified with the following
sequences: sense primer 5’-TCATGGCCTACTTGTCCCGG-3’; antisense primer 5’-
CCATTAGGGCAACTCCAGTGT-3’. The predicted PCR product of 1241 bp includes all
1236 nucleotides of the BCATc coding sequence - ATG start through TAA stop (26), plus
five nucleotides derived from the ends of the primers.
For PCR, cDNA (1.5 µl) was added to each reaction mixture (50 µl total volume)
containing 10 mM Tris-HCl (pH 9.0), 50 mM KCl, 1.5 mM MgCl2 , 0.2 mM deoxynucleotide
triphosphates , and 0.2 µM of each primer. After an initial heat-denaturation step, 0.5 µl Taq
polymerase (5 unit/µl) was added to each reaction. An amplification program of denaturation
(94 C, 1 min), annealing (60 C, 2 min) and extension (72 C, 1 min) was used for 34 cycles
followed by a final elongation step at 72 C for 10 min. A second round of PCR was
conducted by adding 1.5 µl of the first PCR reaction to a fresh reaction mixture. Aliquots of
each reaction mixture (10 µl) were analyzed on a 2% agarose gel followed by staining with
ethidium bromide.

9
RESULTS
Expression of BCATm and BCATc in rat tissues. Immunoblotting with affinity-
purified hBCATm antibodies showed ubiquitous but variable expression of BCATm protein
in rat digestive tissues. To provide an estimate of the relative amounts of BCATm in
digestive tissues, it was necessary to load different amounts of extract protein for each tissue
on gels used for Western blotting (see Fig. 1A). Immunoreactive band intensities were then
converted to intensity per µg protein for each tissue, and the value for pancreas was taken as
100%. As shown in Fig. 1B, there was a 20-fold range of BCATm protein concentrations in
the digestive system tissues. Consistent with previous reports (18, 24, 35), the highest
relative concentrations of BCATm were found in pancreas and stomach. The other parts of
the upper digestive tract (esophagus, duodenum) and the salivary glands had BCATm
concentrations that were 24-40% of the pancreas. The lowest concentrations of BCATm
were found in jejunum, ileum, and colon which had 5-8% of the pancreatic levels. In tissues
outside the digestive system and as shown previously (24, 56), heart and kidney had the
highest BCATm concentrations, and all other tissues examined contained BCATm, including
testis, spleen, uterus, lung, kidney, and thymus (data not shown).
In the rat, BCATc protein has been detected by immunological methods in brain,
ovary, and placenta (18). As shown in Fig. 1C, BCATc protein was not detected by
immunoblotting in whole tissue extracts of the BCATm-expressing tissues of the digestive
system. Similar results were obtained with other BCATm-expressing tissues (data not
shown). Thus, the immunoblot results are consistent with the current view that, in the rat,
BCATc is not expressed outside the brain and female reproductive tissues. In other species,
it has been reported that BCATc is expressed in a wider range of tissues (5, 17). Therefore,

10
we used RT-PCR to determine if BCATc mRNA was detectable in tissues of the digestive
system. After a single round of PCR (34 cycles), a 1241 bp band corresponding to the
BCATc mRNA was observed in several digestive tissues (ileum, jejunum, colon; data not
shown). As shown in Fig. 1D, when an aliquot of the first round PCR reaction was used in a
second round of PCR amplification, a BCATc mRNA-derived band was observed in all
tissues of the digestive system, although at almost undetectable levels in the esophagus. Low
concentrations of BCATc mRNA were also found in testis, kidney, spleen, thymus, heart,
lung, and liver (data not shown). The results raised the possibility that BCATc could be
expressed at low levels in rat tissues or in a selected cell type within a tissue.
Localization of BCATm protein in rat tissues. BCATm immunoreactivity was found
in epithelial cells in all portions of the digestive tract (Fig. 2). Controls for stomach and
colon (no secondary antibody) are shown in Fig. 2G and H. In the submandibular salivary
gland, labeling was strongest in the serous-secreting components of the secretory epithelium;
mucus-secreting epithelia were not stained (Fig. 2A). Throughout the stomach, the heaviest
immunolabeling for BCATm was observed in the epithelial cells of the middle and deep
zones of the gastric mucosa (Fig. 2B). Parietal cells, which have abundant peripherally
located mitochondria, were stained intensely for BCATm (Fig. 2B and inset). Chief cells
were also labeled. The superficial zone of the mucosa, including much of the mucus–
secreting epithelium, was not labeled (not shown). The longitudinal and circular smooth
muscle layers, as well as the thin muscularis mucosae underlying the gastric mucosa showed
light immunolabeling. Light BCATm staining of the smooth muscle was seen throughout the
digestive tract.
In the small intestine, labeling of the duodenum was principally of cells in the deep
zone of the mucosa, in particular, those lining the crypts (Fig. 2C). Labeling in the jejunum

11
and ileum was diffuse and much lighter than in the duodenum, with the highest
concentrations of label again found in the crypts (Fig. 2D & E). Cell types in the small
intestinal crypts include lysozyme-secreting Paneth cells, though some endocrine and stem
cells are also present and may be immunolabeled. Immunoreactivity for BCATm was not
seen in the absorptive cells lying in the more superficial zone of the small intestine mucosa.
The colon was labeled in much the same pattern as the duodenum, with the crypt epithelium
being most heavily labeled (Fig. 2F). The epithelial cells of the crypt are precursor cells for
the absorptive, mucus, and enteroendocrine cells of the more superficial mucosal epithelium.
BCATm also showed cell-specific localization in tissues outside the digestive system
(Fig. 3). In Fig. 3, BCATm-specific immunostaining is seen as a brown color and the cell
nuclei are stained with hematoxylin (blue/purple color). In skeletal muscle (Fig. 3A) and
heart (data not shown) BCATm was found in myofibrils. In kidney, BCATm was located in
the cortex. Intense labeling with the BCATm antibody was seen in epithelial cells lining
short tubule segments in the cortex (Fig. 3B). The smaller lumina of these tubules are
indicative of distal convoluted tubules, as are the more apical positions of the nuclei of the
immunoreactive epithelial cells. Less intense staining was observed in cells of the
glomerulus. Immunoreactive cells were seen in interstitial capillaries (Fig. 3B). These cells,
which may be endothelial cells, were not labeled in the controls (data not shown). BCATm-
specific immunostaining was not evident in proximal tubules and collecting ducts. In the
pancreas, there was intense immunolabeling for BCATm in acinar cells of the exocrine
pancreas, whereas immunolabeling was weak in the Islets of Langerhans and in the cells
lining the intercalated ducts (Fig. 3C). Light staining was observed in cells at the periphery
and in cells in the interior of the islets. BCATm-specific immunostaining was not observed
in the immunoadsorbed control shown in Fig. 3I. BCATm was also found in the lung, where

12
it was localized to the bronchiolar and alveolar epithelium (data not shown). Consistent with
earlier reports (32), BCATm was not seen in sections of liver (compare Fig. 3G & H).
All tissues of the reproductive tract of male and female rats that were examined
expressed BCATm. In ovarian follicles, labeling for BCATm was seen in the secretory cells
that make up the theca interna (arrows in Fig. 3D). The follicular epithelial cells
(granulosum) and stromal fibroblasts (theca externa) were unlabeled. BCATm was localized
in the uterus to secretory epithelial cells in the deep portions of endometrial glands (Fig. 3E).
Surrounding smooth muscle was not labeled. In the testis, BCATm was restricted to the
interstitial tissues, with the most intense labeling in Leydig cells (Fig. 3F). Seminiferous
tubules did not show appreciable BCATm-specific immunoreactivity.
Localization of BCATc in rat tissues. The observation that low levels of BCATc
mRNA were present in rat tissues raised the possibility that BCATc protein might also be
present in these tissues, albeit at levels too low to be detected easily by immunoblotting of
whole tissue extracts. Therefore, several different BCATc-specific antibodies were used to
look for BCATc-specific staining in rat stomach (see Methods). Rat stomach was chosen
because it expresses the BCATc mRNA, and the high concentration of BCATm in this tissue
would provide a good test for the specificity of the BCATc antibodies. All three anti-BCATc
antibodies exhibited a similar staining pattern. Cells and processes of the neural elements of
the stomach exhibited intense staining for BCATc (Fig. 4B-D). Intense BCATc-specific
labeling was found in the myenteric (Auerbach’s) plexus lying between the circular and
longitudinal smooth muscle layers. Immunolabeling was also observed in fine processes of
the submucosal (Meissner’s) plexus (not shown). Because the affinity purified anti-human
BCATc antibody showed high specificity and low background (Fig. 4D), it was used to
localize BCATc in other rat tissues.

13
BCATc-specific immunostaining was found in neural elements of all gastrointestinal
tract tissues that were examined (Fig. 5). In the salivary gland (Fig. 5A), bundles and finer
processes of immunoreactive neurons were seen in the vascular/connective tissue beds.
BCATc-specific staining was also seen in nerves in the pancreas (Fig. 5B). From the
stomach to the colon, BCATc was immunolocalized in the nerves and ganglia of the
myenteric nerve plexus (Fig. 5C-G). It appeared that cell bodies as well as neuronal
processes were immunolabeled. In these and all other tissues, BCATc immunostaining was
not evident in non-neuronal cells. Salivary gland and jejunum controls are shown in Fig. 5H
and 5I, respectively.
To determine if BCATc was also present in nerves outside the digestive tract, rat
sciatic nerve was examined for BCATc-specific immunoreactivity. Nerve-specific
localization of BCATc was confirmed in cross-sections of the sciatic nerve (Fig. 6).
Labeling within the cross-sectioned nerve appeared as well-defined small circular and
polygonal profiles (Fig. 6A). Companion sections of the sciatic nerve were stained with the
lipid stain Oil Red O to visualize the myelin sheath surrounding the nerve axons (Fig. 6B).
Comparison of Fig. 6A and B suggests that BCATc immunoreactivity is restricted to the
nerve axons and not found in the myelin sheath. As the nerve enters the muscle
compartment, immunoreactivity for BCATc could also be seen in tangentially and cross-
sectioned axons (Fig. 6C).
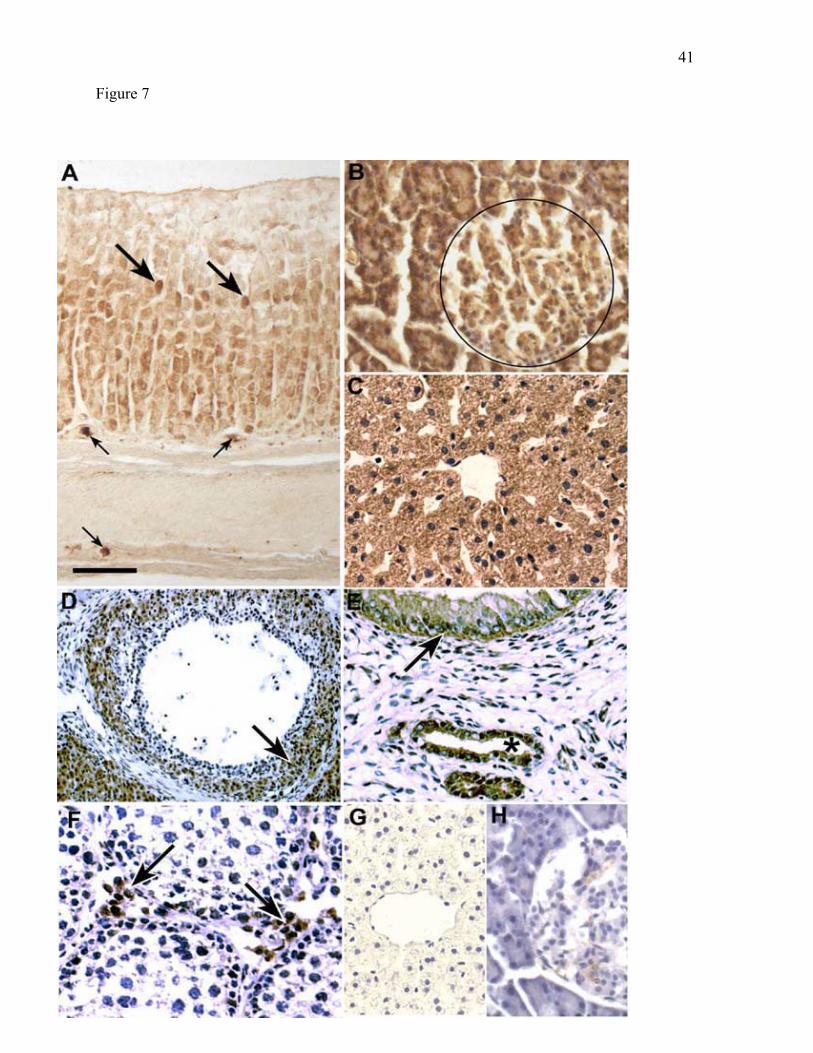
Localization of BCKD in rat tissues. In general, the distribution of the E2 subunit of
BCKD in rat tissues paralleled that of BCATm (Fig. 7). BCKD E2-specific staining was
found in the epithelial cells of the middle and deep zones of the gastric mucosa (compare Fig.
7A and Fig 2B). As seen with BCATm, cells with parietal cell morphology exhibited intense
E2 immunostaining. There was no specific labeling for BCKD E2 in the myenteric nerve

14
plexus. E2 was present in the exocrine and endocrine pancreas, though there was less
difference in intensity of immunolabeling between acinar cells and islets than was observed
with BCATm. (compare Fig. 7B & Fig. 3C). BCKD immunoreactivity was the same as
observed with BCATm in skeletal muscle and heart, whereas in kidney BCKD E2-specific
staining was found in the proximal as well as in the distal convoluted tubules (data not
shown). Consistent with reports showing that rat liver has the highest concentration of
BCKD activity and protein (21, 56, 59), intense BCKD E2-specific staining was found in
liver hepatocytes (Fig. 7C). Immunoadsorbed liver and pancreas controls are shown in Fig
7G and 7H, respectively.
In the female reproductive tract, immunoreactivity for BCKD E2 was seen in cells
composing the granulosa and theca interna in the ovary (Fig 7D) and in the epithelial cells of
the uterine endometrium (Fig. 7E). As observed with BCATm, BCKD E2 was found in the
Leydig cells of the testis and not in the seminiferous tubule (Fig. 7F). Thus, the distribution
of BCKD is very similar to that of BCATm.
Expression of BCAT isozymes in selected human and nonhuman primate tissues.
Immunoblotting of African Green monkey and human tissues with isozyme-specific affinity-
purified anti-BCAT antibodies confirmed the ubiquitous expression of BCATm and
expression of BCATc in monkey and human brain tissue (Fig. 8). BCATm protein levels
varied considerably both in monkey and human tissues. The human recombinant proteins
were used as standards to estimate the level of the BCAT proteins in the monkey tissues.
The results indicated that the highest concentration of BCATm protein is found in pancreas
and kidney followed by muscle, stomach, and jejunum. Some degradation of BCATm can be
observed in the pancreas extracts. Significant concentrations of BCATc protein were only
observed in brain tissue. In monkey brain BCATc represented about 80% of total BCAT

15
protein (BCATm ~20%) which is similar to what has been reported for rat brain (24, 35, 36).
BCATm was near the limits of detection in liver and subcutaneous fat.
The human BCATm and rat BCKD antibodies were used to examine the cellular
localization of these proteins in available monkey tissues. In most tissues the localization of
these enzymes was similar to what was observed in rat tissues. An exception was monkey
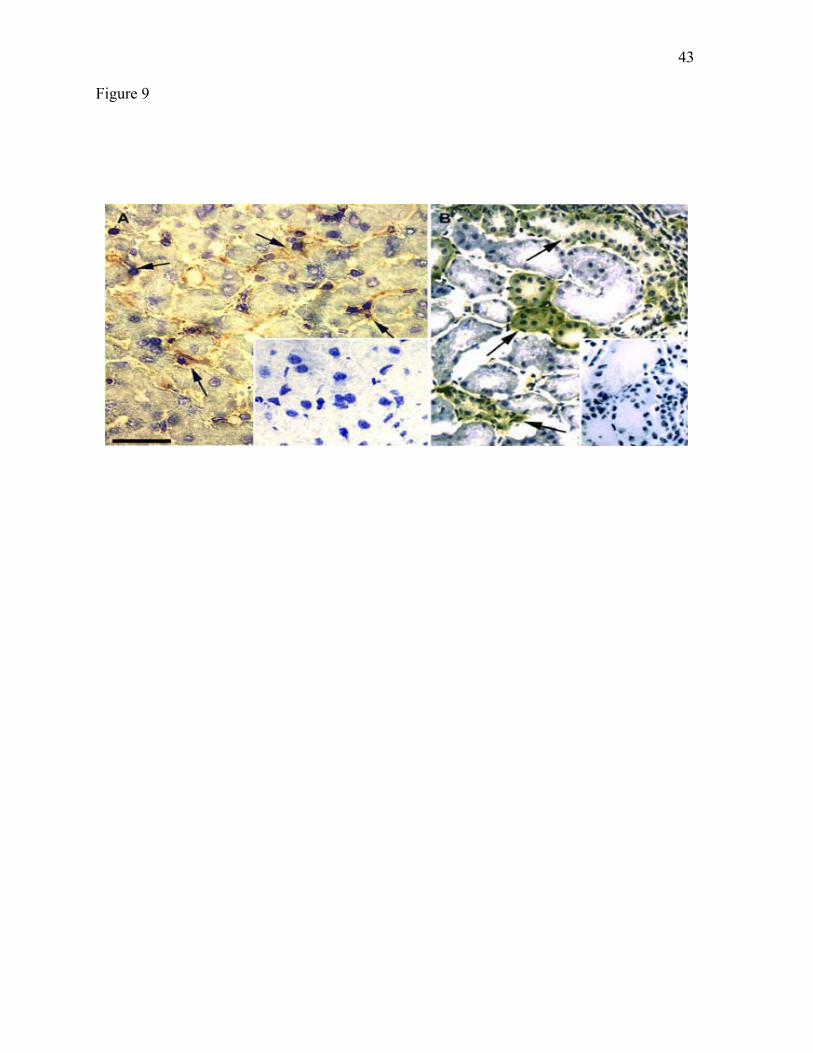
liver, which like human liver, has measurable BCAT activity (56). As shown in Fig. 9A,
BCATm immunoreactivity was not found in hepatocytes. Immunoreactivity appeared to be
localized in cells that have the location and morphology of Kupffer cells. As in the rat,
BCKD E2 immunoreactivity was found in hepatocytes. For other tissues (stomach, pancreas,
kidney, skeletal muscle, and heart), the localization of BCATm immunoreactivity paralleled
the localization of this protein in the rat (data not shown). As in the rat, distal convoluted
tubules of monkey kidney were labeled, though the glomerulus was not immunoreactive for
BCATm. For BCKD, the labeling pattern in liver, pancreas, skeletal and heart muscle was
the same as in the corresponding rat tissues (data not shown).
DISCUSSION
A novel finding of the present work is the consistent localization of BCAA catabolic
enzymes in secretory epithelial cells. This is particularly marked for BCATm, which appears
along the course of the digestive tract in cells that secrete either salivary components, acid,
enzymes, or enzyme precursors. BCATm was not found in absorptive cells or in goblet cells,
which secrete mucus components. In other organ systems, BCATm is found in protein- and
steroid- secreting cells (e.g., ovarian endometrium, testicular Leydig cells, respectively) as

16
well as in transporting epithelia (kidney distal convoluted tubules). The BCATm-containing
cells are variously derived from ecto- endo-and mesoderm. For the most part, these cells
have in common only their position at an interface with the lumen of an organ (i.e.,
epithelial). Second, our results show that with a few exceptions, BCATm is expressed with
BCKD, the second enzyme of the BCAA catabolic pathway.
The preferential localization of BCATm in secretory epithelial cells raises interesting
questions about the function(s) of the BCAA in these cells. For example, high concentrations
of BCATm and BCKD are found in the pancreatic acini that synthesize and secrete proteins,
rather than in the islet, where leucine and -ketoisocaproate (KIC, the transamination product
of leucine) are known to stimulate insulin secretion (43, 60, 61). The high concentrations of
BCATm in the pancreatic acini could function to provide KIC as a signaling molecule to the
islet. Alternatively, BCATm activity in the acini may serve to dampen leucine signaling in
the islet. KIC release from pancreas has not been measured directly. However, even though
differences did not reach statistical significance, portal KIC concentrations were 6.4 M
higher than arterial KIC concentrations in fed dogs (40.6 versus 34.2 M; reference 63). The
portal vein receives blood from the stomach, pancreas, and intestine. Leucine stimulates
protein synthesis in several tissues (15, 42, 46, 60). If leucine or KIC is a nutrient signal in
the exocrine pancreas, then the high concentrations of BCATm may regulate the availability
of the active metabolite.
In the gastrointestinal tract, BCAA catabolic enzymes are found in the secretory
epithelial cells rather than in the absorptive epithelial cells of the intestinal mucosa.
Therefore, only a small fraction of enterally fed BCAAs would be expected to be oxidized by
the intestinal epithelium during the absorptive process. This interpretation is consistent with
results from studies using leucine and KIC tracers in which it has been shown that rates of

17
nitrogen transfer (BCAA transamination) are higher than rates of oxidation (BCKD step) and
that oxidation by the gut is limited (2, 44, 45, 62, 63). For example, Matthews and
coworkers (44, 45) calculated that <2% of nasogastrically delivered leucine and about 5% of
KIC were oxidized on the first pass in humans. Yu et al. (63) measured metabolism of
arterially delivered leucine across the midgut (duodenum to transverse colon) and liver in
dogs. The gut accounted for only 4% of whole body leucine oxidation in fasted dogs and for
13% in fed dogs.
Across species, stomach has one of the highest specific activities of BCAT (35, 56).
While a number of studies have examined leucine metabolism across the splanchnic bed,
where amino acids derived from hydrolysis in the small intestine are plentiful (2, 13, 16, 44,
45, 47, 62, 63), information on the stomach is limited. Amino acid transporters that transport
BCAA have been identified in the stomach. Sobrevia et al (55) have reported that an “L”
type Na+-independent transport system is present in the basolateral surface of the oxyntic
glands, which contain both parietal and chief cells. This transporter is likely to mediate
BCAA uptake by the gastric epithelium from the arterial circulation. Another amino acid
transporter, the ATB0+ protein, is located on the luminal surface of lung epithelial cells, and
is also expressed in mouse stomach (54). This indicates that free BCAA, if available in the
stomach, can also be taken up at the luminal surface of the gastric epithelium. On the other
hand, mRNA encoding a recently cloned peptide transport protein was expressed in the small
intestine but not in stomach or colon in several animal species (8). Thus, it is likely that most
stomach BCAA metabolism is derived from arterial BCAAs. However, since stomach
metabolism of lumen-derived free BCAAs is possible, it could affect conclusions about
BCAA requirements and the proportion of BCAA metabolism that occurs in the digestive
system.

18
Generally, liver is the primary site of catabolism of indispensable amino acids. It is
also likely that liver is the primary site of BCAA oxidation in the rat (28, 53, 56). What sets
BCAAs apart from other indispensable amino acids is that nitrogen transfer via
transamination is largely extrahepatic. For example, in the rat BCATm is not found in liver,
but liver has the highest concentration and activity of BCKD (20,21,56). On the other hand,
nonhuman primates and humans have measurable liver BCAT activity, but it is still less than
6% of total body capacity (17, 56), and BCATm protein is near the limits of detection in both
human and monkey liver (Fig. 8). In the present study, BCATm is found in monkey liver in
what appears to be the phagocytic Kupffer cells, which make up a small proportion of the
cells in the liver. In contrast, BCKD is found in the far more numerous liver hepatocytes in
monkey and rat (Fig 7C). Thus, monkey liver is an example of a tissue where BCKD is
expressed in a cell without BCATm. Whether or not this is true in human liver remains to be
determined. Nevertheless, it is still likely that, as in the rat, under most conditions liver is a
primary site of BCAA oxidation in humans and nonhuman primates. This hypothesis is
supported by a case report showing that after transplantation of a normal liver to a patient
with Maple Syrup Urine Disease (BCKD defect), the patient was able to tolerate a diet with a
normal protein content (58).
For BCATc, the brain is the primary site of expression in human, monkey, sheep, and
rat (14, 56; see also Fig. 8), and in rat, BCATc is found in neurons in culture (3, 25).
However, the expression of BCATc in rat peripheral nerves was unexpected. There is
evidence that BCATc has a more widespread distribution in other animals. By separating
BCATc from BCATm activity using DEAE-cellulose chromatography of crude tissue
homogenates, Goto et al (17) concluded that BCATc activity represents a variable but
significant proportion of total BCAT activity in most human tissues. In a more recent study

19
involving, BCATc-specific antibodies were used to immunoprecipitate BCATc from sheep
tissue homogenates (5). Significant activity attributed to BCATc was found in sheep muscle,
along with BCATc mRNA (5). If the rat is representative of other animals, then the BCATc
measured in these studies may be localized in nervous elements of peripheral tissues of these
animals as well.
Although we have not yet investigated the types of nerves that express BCATc, in the
rat, this isozyme is found in neural elements supplying the salivary gland and pancreas, as
well as in Auerbach’s and Meissner’s plexuses in the gut wall. These nerves are all parts of
the autonomic nervous system, supplying motor neurons that innervate glands and smooth
muscle in the digestive system. In the stomach and intestine, some of the BCATc
immunolabeling may be associated with the interstitial cells of Cajal. These cells are closely
associated with enteric ganglia and smooth muscle cells, and have some neuronal
characteristics (51). In the sciatic nerve, the nonuniform distribution of BCATc
immunoreaction in the axons (see Fig. 6) may reflect the association of BCATc with a
particular class of neurons or with a specific neuronal component that is unevenly distributed
in the axoplasm. BCATc is a target of the neuroactive drug gabapentin (25), which is used
widely to treat neuropathic pain (1). We have also found BCATc in spinal cord neurons (AJ
Sweatt, MA Garcia-Espinosa, R Wallin, SM Huston, unpublished observations). It is
possible that inhibition of BCATc by gabapentin may contribute to its efficacy in the
treatment of neuropathic pain.
In the central nervous system, the BCAT isozymes are thought to participate in a
shuttle that provides amino nitrogen for de novo synthesis of excitatory neurotransmitter
glutamate in brain and in the retina (25, 40, 64). In this scheme, nitrogen is shuttled between
BCATm in astroglia and BCATc in glutamatergic and/or -aminobutyric acid (GABAergic)

20
neurons. In the peripheral nervous system, including the autonomic division innervating the
gastrointestinal tract, the principal neurotransmitters are acetylcholine, the catecholamines
(norepinephrine and epinephrine), and neuroactive peptides. For these neurotransmitter
systems, the function of a nitrogen shuttle operating between neurons and surrounding glia is
not clear, although -aminobutyric acid is synthesized in tissues outside the brain. An
alternative function for BCATc might be to regulate (or attenuate) the anabolic signal
provided by leucine (12, 19, 42, 61). The apparent absence of BCKD in the peripheral
nerves indicates that KIC produced from leucine by BCATc in neurons could not be
metabolized, and is likely released. Finally, it should be noted that for both BCATm and
BCATc, non-enzymatic roles in intracellular signaling may be possible. Recent work has
shown that a splice variant of BCATm with an internal deletion of 12 amino acids is
expressed in colon carcinoma cells (41). This isoform of BCATm binds to the thyroid
hormone receptor and enhances its effects on nuclear transcription activity (41).
Furthermore, BCATm has a redox-active CXXC center (9) and has been found to associate
with BCKD and other proteins (27). Identifying the constituents of neurons with which
BCATc might interact may shed light on its role in the peripheral nervous system.

21
ACKNOWLEDGEMENTS
The work reported here was supported by grants DK 34738 and NS 38641 from the U.S.
National Institutes of Health and grant 98-35200-6067 from the U.S. Department of
Agriculture (SMH).
Current Address for Agus Suryawan:
U.S. Department of Agriculture/ Agriculture Research Service Children’s Nutrition Research Center Department of Pediatrics Baylor College of Medicine Houston TX 77030

22
DISCLOSURE STATEMENT
The authors of the paper, “Branched-Chain Amino Acid Catabolism: Unique Segregation of
Pathway Enzymes in Organ Systems and Peripheral Nerves,” have no potential conflicts of
interest in submitting this paper to the APS for consideration for publication.

23
REFERENCES
1. Backonja M, and Glanzman RL. Gabapentin dosing for neuropathic pain: evidence from
randomized, placebo-controlled clinical trials. Clin Ther 25: 81-104., 2003.
2. Biolo G, and Tessari P. Splanchnic versus whole-body production of alpha-ketoisocaproate
from leucine in the fed state. Metabolism 46: 164-167, 1997.
3. Bixel M, Shimomura Y, Hutson S, and Hamprecht B. Distribution of key enzymes of
branched-chain amino acid metabolism in glial and neuronal cells in culture. J Histochem
Cytochem 49: 407-418, 2001.
4. Bixel MG, Hutson SM, and Hamprecht B. Cellular distribution of branched-chain amino
acid aminotransferase isoenzymes among rat brain glial cells in culture. J Histochem
Cytochem 45: 685-694, 1997.
5. Bonfils J, Faure M, Gibrat JF, Glomot F, and Papet I. Sheep cytosolic branched-chain
amino acid aminotransferase: cDNA cloning, primary structure and molecular modelling and
its unique expression in muscles. Biochim Biophys Acta 1494: 129-136, 2000.
6. Burrin DG, Stoll B, Chang X, van Goudever JB, Fujii H, Hutson S, and Reeds PJ.
Parenteral nutrition results in impaired lactose digestion and hexose absorption when enteral
feeding is initiated in infant pigs. Am J Clin Nutr : In Press, 2003.
7. Chang TW, and Goldberg AL. The metabolic fates of amino acids and the formation of
glutamine in skeletal muscle. J Biol Chem 253: 3685-3693, 1978.
8. Chen H, Wong EA, and Webb KE, Jr. Tissue distribution of a peptide transporter mRNA
in sheep, dairy cows, pigs, and chickens. J Anim Sci 77: 1277-1283, 1999.

24
9. Conway ME, Yennawar N, Wallin R, Poole LB, and Hutson SM. Identification of a
peroxide-sensitive redox switch at the CXXC motif in the human mitochondrial branched
chain aminotransferase. Biochemistry 41: 9070-9078, 2002.
10. Cree TC, Hutson SM, and Harper AE. Gas-liquid chromatography of alpha-keto acids:
quantification of the branched-chain-alpha-keto acids from physiological sources. Anal
Biochem 92: 159-163, 1979.
11. Davoodi J, Drown PM, Bledsoe RK, Wallin R, Reinhart GD, and Hutson SM.
Overexpression and characterization of the human mitochondrial and cytosolic branched-
chain aminotransferases. J Biol Chem 273: 4982-4989, 1998.
12. Dumont FJ, and Su Q. Mechanism of action of the immunosuppressant rapamycin. Life Sci
58: 373-395, 1996.
13. Elango R, Pencharz PB, and Ball RO. The branched-chain amino acid requirement of
parenterally fed neonatal piglets is less than the enteral requirement. J Nutr 132: 3123-3129,
2002.
14. Faure M, Hayes H, Bledsoe RK, Hutson SM, and Papet I. Assignment of the gene of
mitochondrial branched chain aminotransferase (BCAT2) to sheep chromosome band 14q24
and to cattle and goat chromosome bands 18q24 by in situ hybridization. Cytogenet Cell
Genet 83: 96-97, 1998.
15. Flaim KE, Peavy DE, Everson WV, and Jefferson LS. The role of amino acids in the
regulation of protein synthesis in perfused rat liver. I. Reduction in rates of synthesis
resulting from amino acid deprivation and recovery during flow-through perfusion. J Biol
Chem 257: 2932-2938, 1982.

25
16. Gelfand RA, Glickman MG, Jacob R, Sherwin RS, and DeFronzo RA. Removal of
infused amino acids by splanchnic and leg tissues in humans. Am J Physiol 250: E407-413,
1986.
17. Goto M, Shinno H, and Ichihara A. Isozyme patterns of branched-chain amino acid
transaminase in human tissues and tumors. Gann 68: 663-667, 1977.
18. Hall TR, Wallin R, Reinhart GD, and Hutson SM. Branched chain aminotransferase
isoenzymes. Purification and characterization of the rat brain isoenzyme. J Biol Chem 268:
3092-3098, 1993.
19. Hara K, Yonezawa K, Weng QP, Kozlowski MT, Belham C, and Avruch J. Amino acid
sufficiency and mTOR regulate p70 S6 kinase and eIF-4E BP1 through a common effector
mechanism. J Biol Chem 273: 14484-14494, 1998.
20. Harper AE. Thoughts on the role of branched-chain alpha-keto acid dehydrogenase complex
in nitrogen metabolism. Ann N Y Acad Sci 573: 267-273, 1989.
21. Harris RA, Hawes JW, Popov KM, Zhao Y, Shimomura Y, Sato J, Jaskiewicz J, and
Hurley TD. Studies on the regulation of the mitochondrial alpha-ketoacid dehydrogenase
complexes and their kinases. Adv Enzyme Regul 37: 271-293, 1997.
22. Humason GL. Animal Tissue Techniques. San Francisco: W. H. Freeman and Company,
1979.
23. Hutson S. Structure and function of branched chain aminotransferases. Prog Nucleic Acid
Res Mol Biol 70: 175-206, 2001.
24. Hutson SM. Subcellular distribution of branched-chain aminotransferase activity in rat
tissues. J Nutr 118: 1475-1481, 1988.

26
25. Hutson SM, Berkich D, Drown P, Xu B, Aschner M, and LaNoue KF. Role of branched-
chain aminotransferase isoenzymes and gabapentin in neurotransmitter metabolism. J
Neurochem 71: 863-874, 1998.
26. Hutson SM, Bledsoe RK, Hall TR, and Dawson PA. Cloning and expression of the
mammalian cytosolic branched chain aminotransferase isoenzyme. J Biol Chem 270: 30344-
30352, 1995.
27. Hutson SM, Conway ME, Fujii H, and Wallin R. Discovery of a regulated metabolon
involving key enzymes of the leucine catabolic pathway. Experimental Biology 2003 (FASEB
Annual Meeting), San Diego CA, 2003. Abstract 524.8.
28. Hutson SM, Cree TC, and Harper AE. Regulation of leucine and alpha-ketoisocaproate
metabolism in skeletal muscle. J Biol Chem 253: 8126-8133, 1978.
29. Hutson SM, and Hall TR. Identification of the mitochondrial branched chain
aminotransferase as a branched chain alpha-keto acid transport protein. J Biol Chem 268:
3084-3091, 1993.
30. Hutson SM, and Harper AE. Blood and tissue branched-chain amino and alpha-keto acid
concentrations: effect of diet, starvation, and disease. Am J Clin Nutr 34: 173-183, 1981.
31. Hutson SM, Lieth E, and LaNoue KF. Function of leucine in excitatory neurotransmitter
metabolism in the central nervous system. J Nutr 131: 846S-850S, 2001.
32. Hutson SM, Wallin R, and Hall TR. Identification of mitochondrial branched chain
aminotransferase and its isoforms in rat tissues. J Biol Chem 267: 15681-15686, 1992.
33. Hutson SM, and Zapalowski C. Relationship of branched-chain amino acids to skeletal
muscle gluconeogenic amino acids. In: Metabolism and clinical implications of branched-
chain amino and keto-acids, edited by M. Walser and J. R. Williamson. New York:
Elsevier/North Holland, 1981, p. 245-250.

27
34. Hutson SM, Zapalowski C, Cree TC, and Harper AE. Regulation of leucine and alpha-
ketoisocaproic acid metabolism in skeletal muscle. Effects of starvation and insulin. J Biol
Chem 255: 2418-2426, 1980.
35. Ichihara A. Aminotransferases of branched-chain amino acids. In: Transaminases, edited by
P. Christen and D. E. Metzler. New York: John Wiley & Sons, 1985, p. 430-438.
36. Ichihara A, Noda C, and Goto M. Transaminase of brainched chain amino acids. X. High
activity in stomach and pancreas. Biochem Biophys Res Commun 67: 1313-1318, 1975.
37. Kholodilov NG, Neystat M, Oo TF, Hutson SM, and Burke RE. Upregulation of cytosolic
branched chain aminotransferase in substantia nigra following developmental striatal target
injury. Brain Res Mol Brain Res 75: 281-286, 2000.
38. Labow BI, and Souba WW. Glutamine. World J Surg 24: 1503-1513, 2000.
39. LaNoue KF, Berkich DA, Conway M, Barber AJ, Hu LY, Taylor C, and Hutson S. Role
of specific aminotransferases in de novo glutamate synthesis and redox shuttling in the retina.
J Neurosci Res 66: 914-922, 2001.
40. Lieth E, LaNoue KF, Berkich DA, Xu B, Ratz M, Taylor C, and Hutson SM. Nitrogen
shuttling between neurons and glial cells during glutamate synthesis. J Neurochem 76: 1712-
1723, 2001.
41. Lin HM, Kaneshige M, Zhao L, Zhang X, Hanover JA, and Cheng SY. An isoform of
branched-chain aminotransferase is a novel co-repressor for thyroid hormone nuclear
receptors. J Biol Chem 276: 48196-48205, 2001.
42. Lynch CJ, Patson BJ, Anthony J, Vaval A, Jefferson LS, and Vary TC. Leucine is a
direct-acting nutrient signal that regulates protein synthesis in adipose tissue. Am J Physiol
Endocrinol Metab 283: E503-513, 2002.

28
43. Malaisse WJ, Hutton JC, Carpinelli AR, Herchuelz A, and Sener A. The stimulus-
secretion coupling of amino acid-induced insulin release: metabolism and cationic effects of
leucine. Diabetes 29: 431-437, 1980.
44. Matthews DE, Harkin R, Battezzati A, and Brillon DJ. Splanchnic bed utilization of
enteral alpha-ketoisocaproate in humans. Metabolism 48: 1555-1563, 1999.
45. Matthews DE, Marano MA, and Campbell RG. Splanchnic bed utilization of leucine and
phenylalanine in humans. Am J Physiol 264: E109-118, 1993.
46. May ME, and Buse MG. Effects of branched-chain amino acids on protein turnover.
Diabetes Metab Rev 5: 227-245, 1989.
47. Metges CC, El-Khoury AE, Selvaraj AB, Tsay RH, Atkinson A, Regan MM, Bequette
BJ, and Young VR. Kinetics of L-[1-(13)C]leucine when ingested with free amino acids,
unlabeled or intrinsically labeled casein. Am J Physiol Endocrinol Metab 278: E1000-1009,
2000.
48. Odessey R, Khairallah EA, and Goldberg AL. Origin and possible significance of alanine
production by skeletal muscle. J Biol Chem 249: 7623-7629, 1974.
49. Reed LJ, Damuni Z, and Merryfield ML. Regulation of mammalian pyruvate and
branched-chain alpha-keto acid dehydrogenase complexes by phosphorylation-
dephosphorylation. Curr Top Cell Regul 27: 41-49, 1985.
50. Rudel L, Deckelman C, Wilson M, Scobey M, and Anderson R. Dietary cholesterol and
downregulation of cholesterol 7 alpha-hydroxylase and cholesterol absorption in African
green monkeys. J Clin Invest 93: 2463-2472, 1994.
51. Sanders KM. A case for interstitial cells of Cajal as pacemakers and mediators of
neurotransmission in the gastrointestinal tract. Gastroenterology 111: 492-515, 1996.

29
52. Sener A, Somers G, Devis G, and Malaisse WJ. The stimulus-secretion coupling of amino
acid-induced insulin release. Biosynthetic and secretory responses of rat pancreatic islet to L-
leucine and L-glutamine. Diabetologia 21: 135-142, 1981.
53. Shinnick FL, and Harper AE. Branched-chain amino acid oxidation by isolated rat tissue
preparations. Biochim Biophys Acta 437: 477-486, 1976.
54. Sloan JL, Grubb BR, and Mager S. Expression of the amino acid transporter ATB 0+ in
lung: possible role in luminal protein removal. Am J Physiol Lung Cell Mol Physiol 284:
L39-49, 2003.
55. Sobrevia L, Medina V, Reinicke K, and Bravo I. Uptake of L-leucine and L-phenylalanine
across the basolateral cell surface in isolated oxyntic glands. Biochim Biophys Acta 1106:
257-263, 1992.
56. Suryawan A, Hawes JW, Harris RA, Shimomura Y, Jenkins AE, and Hutson SM. A
molecular model of human branched-chain amino acid metabolism. Am J Clin Nutr 68: 72-
81, 1998.
57. Wallin R, Hall TR, and Hutson SM. Purification of branched chain aminotransferase from
rat heart mitochondria. J Biol Chem 265: 6019-6024, 1990.
58. Wendel U, Saudubray JM, Bodner A, and Schadewaldt P. Liver transplantation in maple
syrup urine disease. Eur J Pediatr 158 Suppl 2: S60-64, 1999.
59. Wohlhueter RM, and Harper AE. Coinduction of rat liver branched chain alpha-keto acid
dehydrogenase activities. J Biol Chem 245: 2391-2401, 1970.
60. Xu G, Kwon G, Cruz WS, Marshall CA, and McDaniel ML. Metabolic regulation by
leucine of translation initiation through the mTOR-signaling pathway by pancreatic beta-
cells. Diabetes 50: 353-360, 2001.

30
61. Xu G, Kwon G, Marshall CA, Lin TA, Lawrence JC, Jr., and McDaniel ML. Branched-
chain amino acids are essential in the regulation of PHAS-I and p70 S6 kinase by pancreatic
beta-cells. A possible role in protein translation and mitogenic signaling. J Biol Chem 273:
28178-28184, 1998.
62. Yu YM, Burke JF, Vogt JA, Chambers L, and Young VR. Splanchnic and whole body L-
[1-13C,15N]leucine kinetics in relation to enteral and parenteral amino acid supply. Am J
Physiol 262: E687-694, 1992.
63. Yu YM, Wagner DA, Tredget EE, Walaszewski JA, Burke JF, and Young VR.
Quantitative role of splanchnic region in leucine metabolism: L-[1-13C,15N]leucine and
substrate balance studies. Am J Physiol 259: E36-51, 1990.
64. Yudkoff M, Daikhin Y, Grunstein L, Nissim I, Stern J, and Pleasure D. Astrocyte
leucine metabolism: significance of branched-chain amino acid transamination. J Neurochem
66: 378-385, 1996.
65. Zielke HR, Huang Y, Baab PJ, Collins RM, Jr., Zielke CL, and Tildon JT. Effect of
alpha-ketoisocaproate and leucine on the in vivo oxidation of glutamate and glutamine in the
rat brain. Neurochem Res 22: 1159-1164, 1997.

31
FIGURE LEGENDS
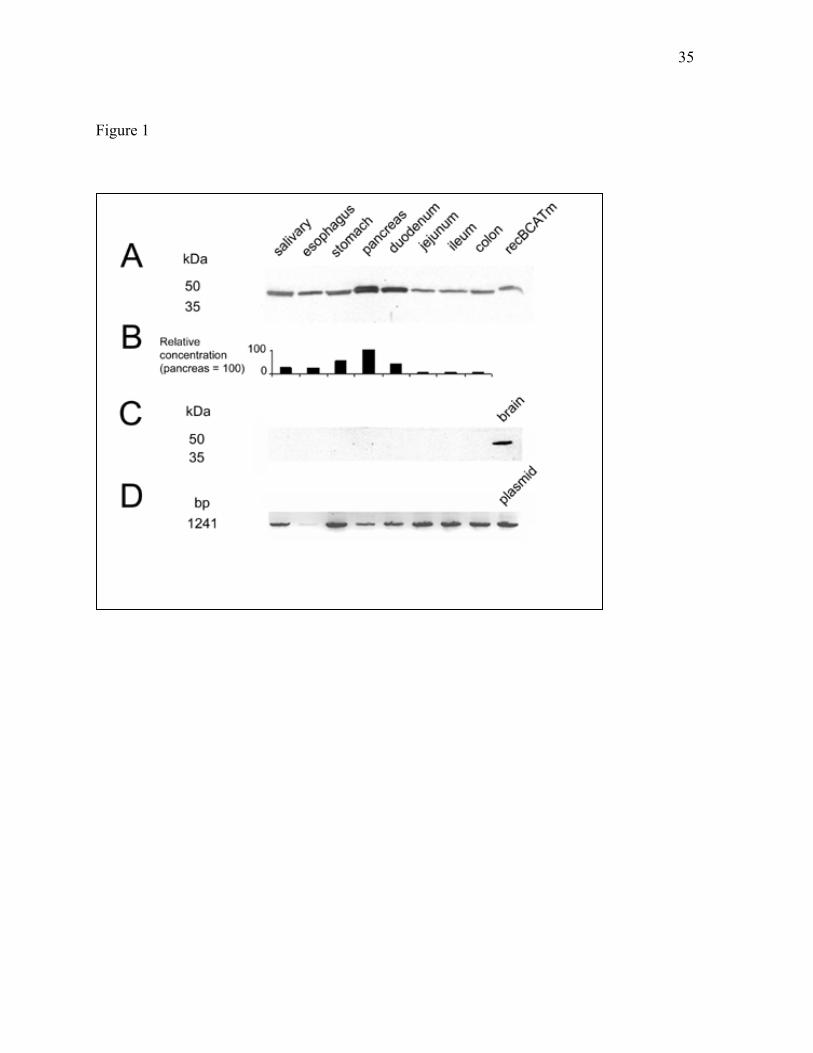
Figure 1. BCATm is detectable by immunoblot throughout the rat gastrointestinal tract,
while BCATc is detectable only by RT-PCR. A: Immunoblot of BCATm. Amounts of
tissue extract loaded onto the gel ranged from 10-60 µg protein/lane. Recombinant human
BCATm (8 ng) was used as a standard . B: Relative tissue concentrations of BCATm.
Immunoreactive band intensity per µg protein loaded was calculated for each tissue, and is
presented as a percentage of the value for pancreas, which had the highest concentration
(taken as 100%). C: Immunoblot for BCATc. Forty µg of tissue extract protein was loaded
on the gel, except for brain, where 20 µg was used. D: RT-PCR for BCATc mRNA. cDNA
was reverse-transcribed from mRNA from rat gastrointestinal tissues, used as the template
for PCR with BCATc-specific primers, and reamplified in second round of PCR (see
Methods). Lanes are in the same order as for the immunoblot, except for the positive control,
for which a BCATc-encoding plasmid was used as the PCR template.
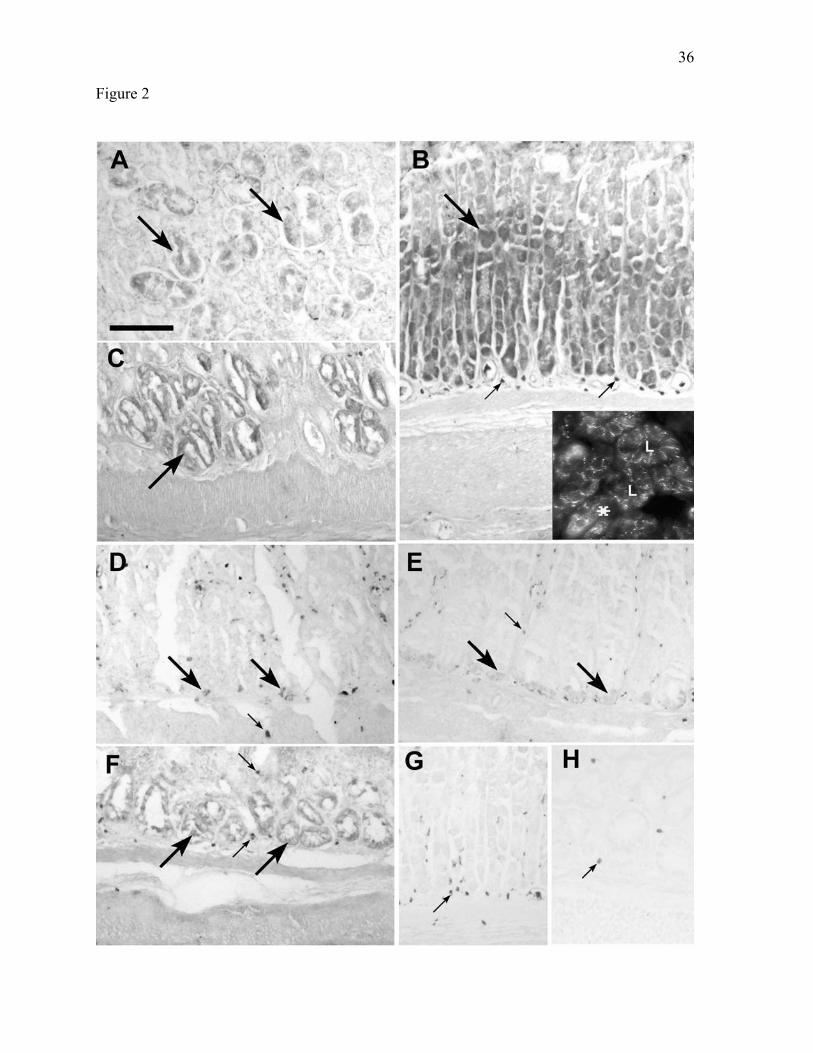
Figure 2. BCATm is present in epithelial cells in all portions of the rat gastrointestinal tract.
A: Salivary gland. B: Stomach. C: Duodenum. D: Jejunum. E: Ileum. F: Colon. G: Stomach
control (secondary antibody only). H: Colon control. Images in panels B-H are oriented
such that the mucosa is at the top, while the smooth muscle layers are at the bottom of each
panel. In all tissues, eosinophils display non-immunologically based reactivity (small
arrows). Labeling for BCATm is most prominent over serous-secreting portions of the
salivary gland, and over the middle (neck) and deeper zones, including crypts, of the gastric
mucosa (large arrows). In panel B, inset shows immunofluorescent localization of BCATm
in the deep zone of the gastric epithelium. Glands are seen in longitudinal (*) and cross-

32
section (L indicates lumen of gland). Immunolabel is strongest in the peripheral cytoplasm
of the epithelial cells. Magnification for all panels is X250. Scale bar: 100 µm.
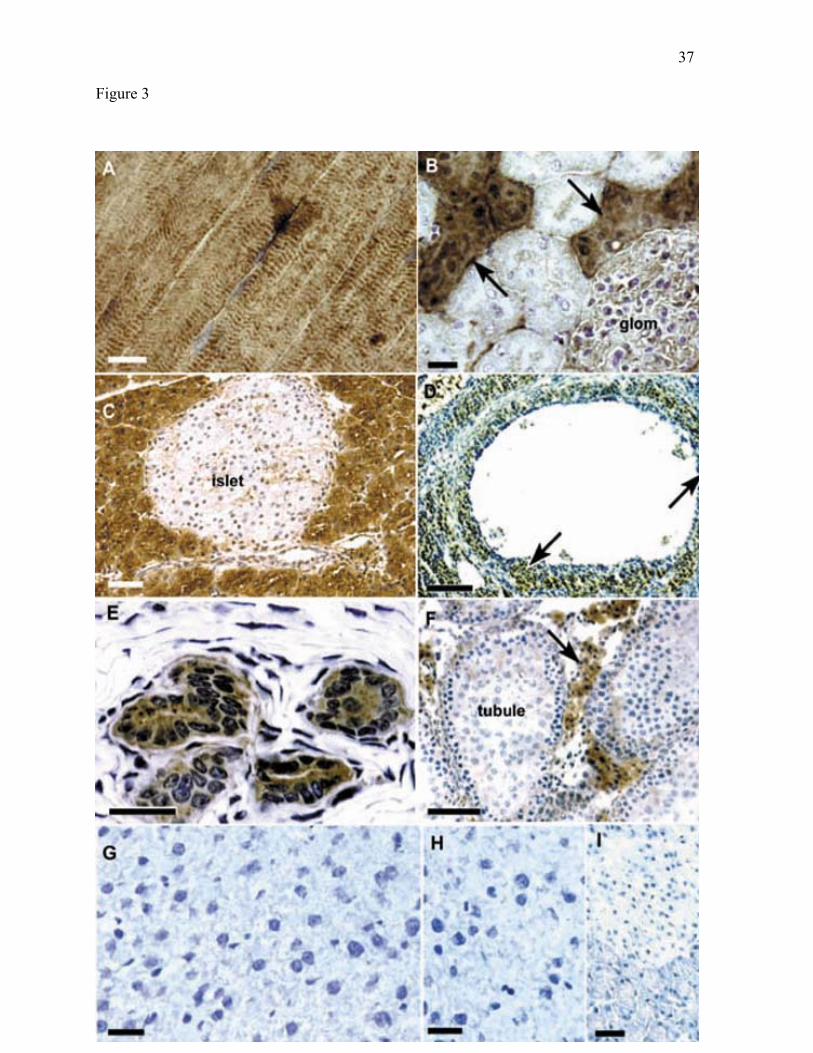
Figure 3. BCATm is present in specific cell types in rat tissues. In this figure and other
color figures, cell nuclei were counterstained with hematoxylin (blue/purple) to allow
identification of cell types in the tissues. Specific immunolabel appears brown.
A: Skeletal muscle, X600. B: Kidney, X440. Arrows indicate immunoreactive distal
convoluted tubules. Occasional immunoreactive cells are seen in the interstitial capillaries.
(glom: glomerulus) C: Pancreas, X280 (islet: Islet of Langerhans). D: Ovary, X170.
Arrows indicate immunoreactive theca interna of an antral follicle. E: Uterus, X1040. F:
Testis, X200. Arrows indicate immunoreactive interstitial Leydig cells (tubule: seminiferous
tubule). G: Liver, X560. H: Liver control, X520. Primary antibody (5-10 ug/ml) was
preadsorbed with recombinant BCATm. I: Pancreas, X440. Immunoadsorbed control. Scale
bar is 50 um for panel C, 100 um for panel F, and 25 µm for all other panels.
Figure 4. BCATc antibodies identify BCATc in nervous elements in rat stomach. A:
Control (secondary antibody only); B: Anti-rat BCATc N-terminal peptide antibody. C:
Anti-rat BCATc, IgG fraction. D: Anti-recombinant human BCATc antibody. Deep zone of
gastric mucosa is at top of each panel, while smooth muscle layers are at the bottom of each
panel. All three antibodies react with elements lying between the inner circular and outer
longitudinal muscle layers. The immunoreactive structures are the ganglia and neural fibers
of the intrinsic innervation of the gut (Auerbach’s plexus, large arrows). In panel D,
additional labeling of fine innervating fibers is seen within the circular muscle layer. Small

33
arrows indicate eosinophils showing non-immunological background activity. Magnification
for all panels is X140. Scale bar is 100 µm.
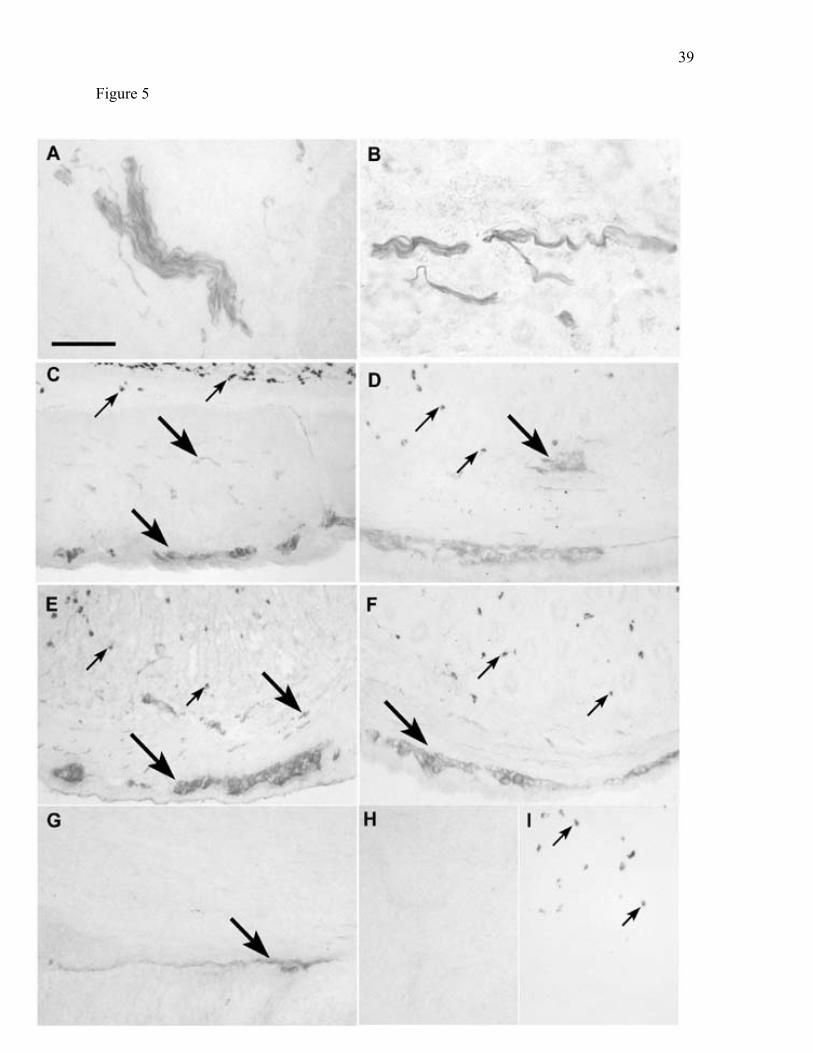
Figure 5. BCATc is localized in nerves of the rat gastrointestinal tract tissues. Cryosections
were probed with affinity-purified anti-recombinant human BCATc, followed by secondary
antibody and peroxidase substrate as described in the Methods. A: Salivary gland. B:
Pancreas. C: Stomach. D: Duodenum. E: Jejunum. F: Ileum. G: Colon. H: Salivary gland
control (secondary antibody only). I: Jejunum control. Labeling for BCATc is restricted to
nervous elements lying between lobes of the salivary gland and to nerves and ganglia in
Auerbach’s plexus in the gastrointestinal tract (large arrows). Small arrows indicate
eosinophil background activity. Magnification is X500 for panels A and B, and X250 for all
other panels. Scale bar is 50 µm for panels A and B, and 100 µm for all other panels.
Figure 6. BCATc is expressed in axons in the sciatic nerve. A: Rat sciatic nerve was
immunolabeled with affinity-purified anti-recombinant human BCATc. Arrows indicate
immunoreactive axons. Arrowheads correspond to structures stained in Panel. B. B: Sciatic
nerve was stained with Oil Red O to reveal lipid-rich elements (myelin, indicated by
arrowheads). C: Rat gastrocnemius was labeled with anti-recombinant human BCATc. N-
nerve; m – muscle. Magnification for all panels is X920. Scale bar is 25 µm.
Figure 7. Branched chain -keto acid dehydrogenase (BCKD) is expressed ubiquitously in
rat tissues. The E2 subunit of branched chain -keto acid dehydrogenase (BCKD) was
immunolocalized in selected rat tissues. In all tissues except stomach, cell nuclei were
counterstained with hematoxylin. A: Stomach. Large arrows indicate immunoreactive

34
epithelial cells in the neck region of the gastric mucosa. Eosinophils display background
activity (small arrows). B: Pancreas. Circle shows approximate boundary of an Islet of
Langerhans. C: Liver. D: Ovary. Arrow indicates immunoreactive cells of theca interna.
E: Uterus. Arrow indicates immunoreactive epithelial cells lining endometrial cavity and
asterisk indicates lumen of an endometrial gland with immunoreactive epithelium. F: Testis.
Arrow indicates immunoreactive interstitial Leydig cells. G, H: Respective liver and
pancreas controls (primary antibody immunoadsorbed with purified BCKD complex).
Magnification in panels A and E is X250, in panel D is X125, and in all other panels is X500.
Scale bar is 100 µm for A and E, 200 µm for D, and 50 µm for all other panels.
Figure 8. In the monkey and the human, BCAT isozymes show tissue distributions similar to
those in the rat. Extracts of monkey and human tissues were analyzed by immunoblot with
antibodies directed against recombinant human BCATm or BCATc. Recombinant human
BCATm and BCATc (20 ng each) were used as standards. Human tissues are in the same
order as monkey tissues, except for heart, which was used instead of stomach. Tissue
extracts were loaded at 30 µg protein/lane. Inset table shows concentrations of BCATc and
BCATm in monkey tissues, as determined by densitometry in separate blots (see Methods).
Figure 9. In the monkey, BCATm is localized to Kupffer cells in liver and to distal
convoluted tubules in kidney. In these sections, nuclei were counterstained with
hematoxylin. A: Liver. Arrows indicate immunoreactive Kupffer cells in the liver sinusoidal
lining. Inset shows control section exposed to immunoadsorbed antibody. B: Kidney.
Arrows indicate immunoreactive epithelium of convoluted tubules. Inset shows control
section. Magnification for both panels is X230. Scale bar is 100 µm.
35
Figure 1
36
Figure 2
37
Figure 3

38
Figure 4
39
Figure 5

40
Figure 6
41
Figure 7

42
Figure 8
43
Figure 9
Related Documents











