Named Series: Twenty Years of Brain, Behavior, and Immunity Brain-immune communication pathways Ning Quan a , William A. Banks b, * a Institute of Behavioral Medicine, Ohio State University, USA b GRECC, Veterans Affairs Medical Center-St. Louis and Saint Louis University School of Medicine, Division of Geriatrics, Department of Internal Medicine, USA Received 23 April 2007; received in revised form 13 May 2007; accepted 15 May 2007 Abstract Communication between the central nervous and immune systems lies at the heart of the neuroimmune axis. We trace here some of the major conceptual hurtles which were raised, first against the acceptance of a neuroimmune axis and later in understanding it. We review the major concepts formulated and established during the last two decades and focus on four pathways that have been proposed as important in communication: the neural route, circumventricular organs, blood–brain barrier transport of cytokines, and secretions from BBB cells. These and other pathways have established the existence of a neuroimmune axis, but raise new questions on how they act and interact with one another. Published by Elsevier Inc. Keywords: Neuroimmune; CNS; Immune system; Vagus; Afferent nerves; Blood–brain barrier; Circumventricular organ 1. Introduction The concept that the brain and immune systems com- municate lies at the heart of neuroimmunology. Indeed, the current power and future hope of this field lies in the strength it derives from the realization that the three great systems, the nervous, the endocrine, and the immune, by which the body communicates and regulates also commu- nicate with one another. The last 20 years has witnessed the birth and the transformation of the field of neuroim- mune communication. Of course, neuroimmunology existed long before that. But early neuroimmunologists generally treated the immune and the nervous systems as separate entities. Only disease conditions involving infec- tion or immunological pathology within the nervous sys- tem were studied. These conditions were considered as a consequence of a breach of the insulation that isolated these two systems, thereby pathological in nature. The concept that neuroimmune communication as an integrated part of a physiological supra-system that included both the immune and the nervous system distin- guished the new neuroimmunology from its predecessor. This novel perspective met with considerable resistance and skepticism from its parent fields of neuroscience and immunology. For many traditional immunologists, meddling with the intricately organized immunological processes by the whim of neurotransmitters and neuro- peptides seemed counterintuitive and an unneeded level of complexity. For the neuroscience of the time, the immune system also seemed an unlikely target to regulate. The nervous system was characterized by a speed of communication through a hardwired network of immobile neurons, whereas immune cells were built to move and roam, engulf pathogens and secret antibodies. For the nervous system to receive input from immune cells, one had to envision nerve terminals ‘‘synapsed’’ onto the mobile immune cells. This was certainly heresy at the time. In addition, most of the known immunological activities could be demonstrated in vitro without the help of neu- rons. Therefore, the need for the nervous system to respond to signals from the immune system was not obvious. The progress made in the field of neuroimmune communication has certainly cleared these hurdles to establish that these 0889-1591/$ - see front matter Published by Elsevier Inc. doi:10.1016/j.bbi.2007.05.005 * Corresponding author. Fax: +1 314 289 6374. E-mail address: [email protected] (W.A. Banks). www.elsevier.com/locate/ybrbi Brain, Behavior, and Immunity 21 (2007) 727–735 BRAIN, BEHAVIOR, and IMMUNITY

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

BRAIN,
www.elsevier.com/locate/ybrbi
Brain, Behavior, and Immunity 21 (2007) 727–735
BEHAVIOR,and IMMUNITY
Named Series: Twenty Years of Brain, Behavior, and Immunity
Brain-immune communication pathways
Ning Quan a, William A. Banks b,*
a Institute of Behavioral Medicine, Ohio State University, USAb GRECC, Veterans Affairs Medical Center-St. Louis and Saint Louis University School of Medicine, Division of Geriatrics,
Department of Internal Medicine, USA
Received 23 April 2007; received in revised form 13 May 2007; accepted 15 May 2007
Abstract
Communication between the central nervous and immune systems lies at the heart of the neuroimmune axis. We trace here some ofthe major conceptual hurtles which were raised, first against the acceptance of a neuroimmune axis and later in understanding it. Wereview the major concepts formulated and established during the last two decades and focus on four pathways that have been proposedas important in communication: the neural route, circumventricular organs, blood–brain barrier transport of cytokines, and secretionsfrom BBB cells. These and other pathways have established the existence of a neuroimmune axis, but raise new questions on how they actand interact with one another.Published by Elsevier Inc.
Keywords: Neuroimmune; CNS; Immune system; Vagus; Afferent nerves; Blood–brain barrier; Circumventricular organ
1. Introduction
The concept that the brain and immune systems com-municate lies at the heart of neuroimmunology. Indeed,the current power and future hope of this field lies in thestrength it derives from the realization that the three greatsystems, the nervous, the endocrine, and the immune, bywhich the body communicates and regulates also commu-nicate with one another. The last 20 years has witnessedthe birth and the transformation of the field of neuroim-mune communication. Of course, neuroimmunologyexisted long before that. But early neuroimmunologistsgenerally treated the immune and the nervous systems asseparate entities. Only disease conditions involving infec-tion or immunological pathology within the nervous sys-tem were studied. These conditions were considered as aconsequence of a breach of the insulation that isolatedthese two systems, thereby pathological in nature.
The concept that neuroimmune communication as anintegrated part of a physiological supra-system that
0889-1591/$ - see front matter Published by Elsevier Inc.
doi:10.1016/j.bbi.2007.05.005
* Corresponding author. Fax: +1 314 289 6374.E-mail address: [email protected] (W.A. Banks).
included both the immune and the nervous system distin-guished the new neuroimmunology from its predecessor.This novel perspective met with considerable resistanceand skepticism from its parent fields of neuroscience andimmunology. For many traditional immunologists,meddling with the intricately organized immunologicalprocesses by the whim of neurotransmitters and neuro-peptides seemed counterintuitive and an unneeded levelof complexity. For the neuroscience of the time, theimmune system also seemed an unlikely target to regulate.The nervous system was characterized by a speed ofcommunication through a hardwired network of immobileneurons, whereas immune cells were built to move androam, engulf pathogens and secret antibodies. For thenervous system to receive input from immune cells, onehad to envision nerve terminals ‘‘synapsed’’ onto themobile immune cells. This was certainly heresy at the time.In addition, most of the known immunological activitiescould be demonstrated in vitro without the help of neu-rons. Therefore, the need for the nervous system to respondto signals from the immune system was not obvious. Theprogress made in the field of neuroimmune communicationhas certainly cleared these hurdles to establish that these
ning
Highlight
ning
Highlight
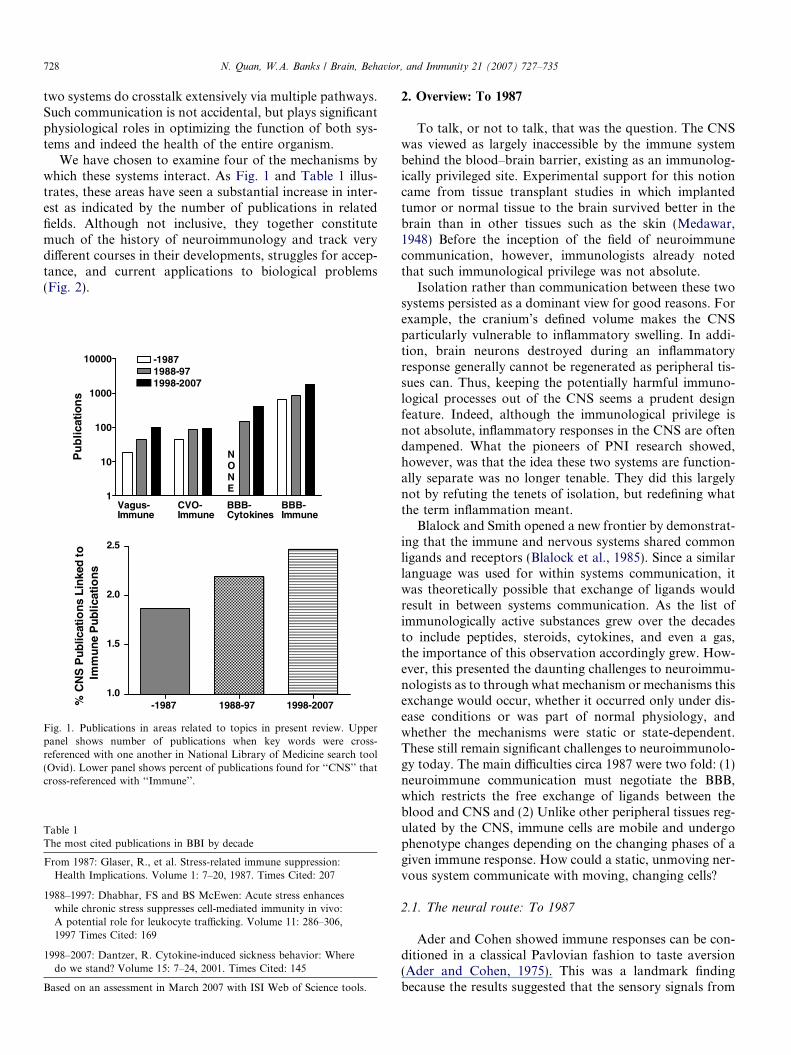
728 N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735
two systems do crosstalk extensively via multiple pathways.Such communication is not accidental, but plays significantphysiological roles in optimizing the function of both sys-tems and indeed the health of the entire organism.
We have chosen to examine four of the mechanisms bywhich these systems interact. As Fig. 1 and Table 1 illus-trates, these areas have seen a substantial increase in inter-est as indicated by the number of publications in relatedfields. Although not inclusive, they together constitutemuch of the history of neuroimmunology and track verydifferent courses in their developments, struggles for accep-tance, and current applications to biological problems(Fig. 2).
1
10
100
1000
10000 -19871988-971998-2007
NONE
Vagus-Immune
BBB-Cytokines
CVO-Immune
BBB-Immune
Pu
blic
atio
ns
-1987 1988-97 1998-20071.0
1.5
2.0
2.5
% C
NS
Pu
blic
atio
ns
Lin
ked
to
Imm
une
Pub
licat
ion
s
Fig. 1. Publications in areas related to topics in present review. Upperpanel shows number of publications when key words were cross-referenced with one another in National Library of Medicine search tool(Ovid). Lower panel shows percent of publications found for ‘‘CNS’’ thatcross-referenced with ‘‘Immune’’.
Table 1The most cited publications in BBI by decade
From 1987: Glaser, R., et al. Stress-related immune suppression:Health Implications. Volume 1: 7–20, 1987. Times Cited: 207
1988–1997: Dhabhar, FS and BS McEwen: Acute stress enhanceswhile chronic stress suppresses cell-mediated immunity in vivo:A potential role for leukocyte trafficking. Volume 11: 286–306,1997 Times Cited: 169
1998–2007: Dantzer, R. Cytokine-induced sickness behavior: Wheredo we stand? Volume 15: 7–24, 2001. Times Cited: 145
Based on an assessment in March 2007 with ISI Web of Science tools.
2. Overview: To 1987
To talk, or not to talk, that was the question. The CNSwas viewed as largely inaccessible by the immune systembehind the blood–brain barrier, existing as an immunolog-ically privileged site. Experimental support for this notioncame from tissue transplant studies in which implantedtumor or normal tissue to the brain survived better in thebrain than in other tissues such as the skin (Medawar,1948) Before the inception of the field of neuroimmunecommunication, however, immunologists already notedthat such immunological privilege was not absolute.
Isolation rather than communication between these twosystems persisted as a dominant view for good reasons. Forexample, the cranium’s defined volume makes the CNSparticularly vulnerable to inflammatory swelling. In addi-tion, brain neurons destroyed during an inflammatoryresponse generally cannot be regenerated as peripheral tis-sues can. Thus, keeping the potentially harmful immuno-logical processes out of the CNS seems a prudent designfeature. Indeed, although the immunological privilege isnot absolute, inflammatory responses in the CNS are oftendampened. What the pioneers of PNI research showed,however, was that the idea these two systems are function-ally separate was no longer tenable. They did this largelynot by refuting the tenets of isolation, but redefining whatthe term inflammation meant.
Blalock and Smith opened a new frontier by demonstrat-ing that the immune and nervous systems shared commonligands and receptors (Blalock et al., 1985). Since a similarlanguage was used for within systems communication, itwas theoretically possible that exchange of ligands wouldresult in between systems communication. As the list ofimmunologically active substances grew over the decadesto include peptides, steroids, cytokines, and even a gas,the importance of this observation accordingly grew. How-ever, this presented the daunting challenges to neuroimmu-nologists as to through what mechanism or mechanisms thisexchange would occur, whether it occurred only under dis-ease conditions or was part of normal physiology, andwhether the mechanisms were static or state-dependent.These still remain significant challenges to neuroimmunolo-gy today. The main difficulties circa 1987 were two fold: (1)neuroimmune communication must negotiate the BBB,which restricts the free exchange of ligands between theblood and CNS and (2) Unlike other peripheral tissues reg-ulated by the CNS, immune cells are mobile and undergophenotype changes depending on the changing phases of agiven immune response. How could a static, unmoving ner-vous system communicate with moving, changing cells?
2.1. The neural route: To 1987
Ader and Cohen showed immune responses can be con-ditioned in a classical Pavlovian fashion to taste aversion(Ader and Cohen, 1975). This was a landmark findingbecause the results suggested that the sensory signals from

To 87
88-97
98-07
Lymph Node
SympatheticAb
Aff Vagu s
Cholinergic(AntiInflamm)
PrimarySensory
Neural CVO
Pineal
Environ Cues(Light)
Melatonin
Blood
VLT
Cytokines
Fever
VLT
IL-1
PG
IL-1
Tanycytes(Barrier,Secretory?)
CVOImmuneCell
Blood
LPS
NeuronalRelay
BBB SecretoryINTACT
ImmuneCells
Albumin
IL-1, -6, TNF, FGF,IL-2, MIPSol Rec, INF-a
Cytokines
CSFDrainage
NO, PG, IL-1, -6, TGF-B LPS
COX-2
PG Fever
Blood
Brain
Cytokine Transporters
IL-2
IL-6
Cytokine Secretion
(Diurnal, CNS Injury, Reg. Var.) MODULATORS (LPS, HIV, Cocaine)
Fig. 2. Diagrams of some major advances, proposed and established, in four areas of neuroimmunology. Neural panels. To 1987: Sympathetic outflow tolymph nodes connected the central nervous and immune systems. 1988–97: Afferent vagal fibers found to mediate effects of intraperitoneal LPS on CNS.1998–2007: Other afferent pathways (e.g., primary sensory nerves) found to relay information from peripheral tissues to CNS. Cholinergic outflow maymediate anti-inflammatory effects. CVO panels. To 1987: Secretion of melatonin by pineal illustrates how environmental cues can regulate immune cellfunctioning. Circulating cytokines ability to modulate temperature linked to OVLT. 1988–97: IL-1 acting within OVLT releases prostaglandins which, inturn, induce fever. 1998–2007: CVO to adjacent brain tissue leakage shown to be restricted by tanycytic layer. LPS induces immune cells residing in CVOto locally secrete IL-1 which then acts at nerve terminals in CVO which project to distant brain regions. Sympathetic projections modulate CVO secretionsof cytokines. BBB panels. To 1987: Early concepts visualized BBB as either intact or disrupted, as in multiple sclerosis. Blood-to-brain transfer of cells andproteins was thought to be unregulated mechanisms and to occur only in the disrupted disease state. 1988–1997: Some cytokines shown to be transportedby saturable system across the BBB, whereas others were not. 1998–2007: Cytokine transport across the BBB shown to influence CNS, transportersmodulated, first saturable brain-to-blood transporter (for IL-2) established, and other cytokines found to enter blood from the CNS with the brain toblood drainage of CSF. Cytokine transporters variable and modulated (e.g., by diurnal rhythm, CNS injury, brain regions). Secretory panels. To 1987: Noliterature on cytokine secretion from BBB cells found. 1988–1997: BBB cells shown to secrete nitric oxide, prostaglandin, IL-1, IL-6, and TGF-beta. LPSfever mediated by induction of COX-2 at brain endothelium with subsequent BBB secretion of prostaglandins. 1998–2007: BBB shown to have polarizedsecretion (e.g. IL-6 preferentially secreted from luminal membrane of brain endothelial cells). Cytokine secretion inducible and modulated (e.g., by LPS,HIV, and cocaine).
N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735 729
immune activity and from taste can be integrated in theCNS. Later, Besedovsky et al. (1979), showed that primaryantibody responses can be regulated by the activity of thesympathetic nervous system and David Felten’s groupshowed that lymphoid organs were innervated by the sym-pathetic nerve fibers (Felten et al., 1985). The importantfinding by Felten et al. was that besides the classic nerveendings on blood vessels, nerve terminals were found clo-sely associated with lymphocytes. Thus, lymphocytes maydirectly receive neural inputs from the autonomic nervoussystem and the concept of immunoregulation by the auto-nomic system was born.
2.2. Circumventricular organs: To 1987
Not surprisingly, scientists first looked for cracks in theBBB. Circumventricular organs (CVOs) are regions of thebrain in which the capillary bed does not form a BBB, butinstead the vessels are leaky. Thus, circulating substancescould reach these areas of the brain which might be incommunication with other regions. Alternatively, sub-stances produced in CVOs or within brain but secreted intoblood via the CVOs could link brain to immune system.Just such secretory pathways had been established in the1970’s for arginine vasopressin and oxytocin, substancesproduced behind the BBB but secreted into the circulationby the neurohypophysis, a CVO.
Early work implicated the pineal gland as a tissue bridg-ing the immune, central nervous, and endocrine systems(Guerrero and Reiter, 1992). The largest of the CVOs,the pineal, secretes melatonin, a substance with effects onantibody response and the development of uveitis. Thismeant the pineal was functioning as an endocrine tissue.Melatonin is present in both CSF and serum and so seemedan ideal ligand for communicating simultaneously with theCNS and peripheral tissues. Melatonin secretion is linkedto photoperiod and so could mediate an environmentalinfluence on the neuroimmunoendocrine system. Immunecells were also demonstrated in the pineal.
Blatteis et al. were the first to show lesioning a CVOclose to the thermoregulatory center of the hypothalamusblocked cytokine induced fever (Blatteis et al., 1983), sug-gesting leakage of cytokines into the CVO, and perhapsbeyond, as a mechanism of neuroimmune communication.The problem with interpreting such lesion studies is that itis not clear whether the procedure prevented the assumedcytokine leakage into the brain or it significantly impairedthe brain regions important for the generation of feverinduced by peripheral cytokines.
2.3. The blood–brain barrier transport of cytokines: To 1987
That the BBB was involved with the immune system wasevidenced by BBB disruption in some disease states, such

730 N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735
as multiple sclerosis and CNS infections. An increase inleakiness of the BBB as an indicator of disease was appre-ciated and serum/CSF ratios for albumin and immuno-globulins were developed for diagnostic purposes. Theneuroimmune literature seems to have assigned only twostates to the BBB: disrupted or not disrupted. The notionthat a disrupted or intact BBB could have participated inCNS-immune communications was not developed fromthis line of work. Cytokines were considered too large tocross the BBB, although it was hypothesized that theymight mediate disruption of the BBB. However, Habifet al. in 1975 found that human leukocyte interferoninjected into the circulation of monkeys appeared in theCSF (Habif et al., 1975) and that interferon injected intothe CSF appeared in blood.
2.4. Secretions of BBB cells: To 1987
Although a modern view would consider many sub-stances secreted by BBB cells, such as nitric oxide and pros-taglandins to be immune modulators, this review willconcentrate on cytokine secretion. No publications priorto 1987 were found for this area.
3. Overview: 1988–1997
Cytokines came to dominate thinking during this era.The ability of both the CNS and immune cells to secreteand respond to them provided a mechanism by which thetwo systems could be linked. Injection of interleukin-1(IL-1) into the brain produced almost all of the behav-ioral, neuroendocrine, and autonomic responses that ani-mals typically display when they are infected. Thisfostered the idea that all of these responses were a coor-dinated, adaptive behavior, termed sickness behavior.Such coordination only made sense in a neuroimmunecontext. Additionally, sickness behavior could be blockedby centrally injected IL-1 receptor antagonist. This meantthat circulating IL-1 was somehow translated into CNSIL-1. The induction of cytokine gene expression (Banet al., 1992; Gatti and Bartfai, 1993) and bioactivity(Quan et al., 1994) was found in the brain after peripheralchallenge of bacterial endotoxin lipopolysaccharide (LPS),suggesting that the IL-1 acting in brain was produced bythe brain. A second anchor to the neuroimmune pathwaysis the production of prostaglandins. These substancescame to be viewed as neuroimmune agents. Prostaglan-dins, for example, were found to both stimulate and bereleased by cytokines.
The concept of the CNS as an immune privileged areaalso began to change. Widner et al., for example, cautionedagainst the assumptions of a generalized immune privilegeand suggested that protection for intracerebrally trans-planted tissue is precarious. His studies found that oncethe host brain was sensitized, immunological rejection ofthe grafted tissue was almost certain (Widner and Brundin,1988).
3.1. The neural route: 1988–1997
A central pathway was demonstrated by Ericsson et al.,who showed that peripheral IL-1 may first stimulate cate-cholamine neurons of the ventrolateral medulla which thenproject to other sites of autonomic control, including theparaventricular nucleus of the hypothalamus (PVN) (Erics-son et al., 1994). What took neuroanatomists by surpriseduring this period was the discovery that direct neuraltransmission via the vagus nerve is a critical neuroimmunecommunication pathway. The first demonstration of thisneural route of neuroimmune communication was pub-lished in 1994 (Bluthe et al., 1994; Watkins et al., 1994).The advantages of using a neural route to communicateimmune information to the brain are the speed and the factthat signals traveling this route will directly activate specifictargets of the brain without the interference of the BBB.Skeptics of the theory pointed out that it is not likely thatvagal afferents can stimulate brain cytokine and Cox-2 pro-duction in a manner that matches the observed neuroana-tomical patterns observed in previous studies (Quan andHerkenham, 2002) In addition, new sensor elements specif-ically responsive to immune signals at the terminals ofvagal afferents are required for this mechanism to be func-tional. Such sensors were found by Goehler et al., whoshowed that glomus cells of the vagal paraganglia bindIL-1ra (Goehler et al., 1997). Numerous vagotomy studiesfollowed the initial breakthrough confirming that the intactvagus plays an important role in mediating a vast array ofeffects induced by peripheral inflammatory challengesincluding hyperalgesia, fever, anorexia, taste aversions,increased levels of plasma corticosteroid, and brain norepi-nephrine changes. These studies left no doubt vagal affer-ents constitute a critical, maybe even major,neuroimmune pathway under physiological conditions(Watkins et al., 1995).
3.2. Circumventricular organs: 1988–1997
Breder et al. (1988) formally showed that IL-1 was takenup by CVOs from the circulation. This conceptually sup-ported the earlier work of Blatteis et al. Breder et al. pro-posed that IL-1 simply leaked out of the CVO to act atadjacent brain tissue. However, CVOs are generally sur-rounded by a tanycytic barrier that limits leakage, and so thishypothesis has been largely dismissed. Arimura and workersproposed that IL-1 works at its receptors in CVOs to relay itssignal to areas behind the BBB (Komaki et al., 1992).
3.3. The blood–brain barrier transport of cytokines:
1988–1997
Thirteen years after the original publication by Habifet al., workers returned once more to the question ofwhether cytokines could cross the BBB. Breder et al. (Bre-der, Dinarello, and Saper, 1988) in their paper on CVOsconcluded that IL-1 did not cross the BBB. However, they

N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735 731
actually did not examine this issue, but like most at thistime just assumed that cytokines were too large to crossthe BBB. In 1989, the first evidence for a saturable trans-port system across the BBB for a cytokine, IL-1 alpha,was published (Banks et al., 1989). The presence of trans-porters could explain how molecules the size of cytokinescould cross the BBB. IL-1 beta and IL-ra were later foundto use the same or a related transporter. IL-6 and TNFwere found to have their own transporters. Leptin, latercategorized as a cytokine, was also found to cross theBBB by a saturable transporter. IL-2, MIP, interferon-alpha, and soluble cytokine receptors were found not tocross the BBB. Although most of the work in this area dur-ing this decade was done by the lab of Banks and Kastin,others independently found that FGF and IL-6 could crossthe BBB. Reichlin and coworkers showed that the non-sat-urable efflux of CNS-derived cytokines that occurs with thereabsorption of CSF can produce substantial levels of cyto-kines in blood (Chen et al., 1997).
The transporters were found to be affected by diseasestates. Pan et al. found in 1996 that the saturable transportsystem for TNF was enhanced during the clinical phase ofEAE (Pan et al., 1996). Later, spinal cord injury was shownto modulate TNF transport in a complex temporal andregional pattern which was unrelated to BBB disruption.
3.4. Secretions of BBB cells: 1988–1997
Brain endothelial cells were found to secrete nitric oxide,prostaglandins, IL-1, IL-6, and TGF-beta (Fabry et al.,1993). The induction by LPS of Cox-2 in brain blood ves-sels correlated with the febrile response (Cao et al., 1997).
4. Overview: 1998–2007
This decade saw confirmation of the discovered neuro-immune pathways and extension of their actions to newfunctions. Increasingly, views such as sickness behaviorbeing adaptive rather than pathologic, involvement ofTNF in physiologic as well as pathologic sleep, immune cellsurveillance of the CNS occurring under normal condi-tions, and secretion and response being a part of normalcellular function subtly changed neuroimmunology froma field primarily focused on pathology to one increasinglyfocused on physiology. With the concept of bi-directionalcommunication between the CNS and immune system wellestablished, questions of how that communication actuallyoccurs became increasingly of interest. How, for example,does the neuroimmune axis distinguish between signalsoriginating from the peripheral immune system butintended for the CNS from those originating in the CNSand intended for the peripheral immune system?
4.1. The neural route: 1998–2007
Evidence supporting the afferent vagal pathway accu-mulated significantly during this period. There were also
discrepant reports produced by different laboratories.One prominent example is the mediation of febrile responseby the vagus. Although earlier studies showed that feverinduced by intraperitoneal injection of IL-1 or LPS wasblocked by vagotomy, later studies showed that vagotomyonly blocked fever induced by low doses, but not highdoses, of these pyrogens (Hansen et al., 2001; Romanovskyet al., 1997). In yet another study, febrile responses to evenlow doses of IL-1 could not be blocked by vagotomy(Luheshi et al., 2000). These results suggest that neuroim-mune communication is not mediated by a single dominantpathway. There may be conditions under which one of theneuroimmune pathways is preferentially activated suchthat the activity of another may be masked. Despite suchfunctional complexity, elements of the immune-vagus-brain pathway hardwiring were continually added to theliterature. Peripheral IL-1 was shown to activate nodoseganglia neurons, cells which express receptors for bothIL-1 and prostaglandin E2 (Ek et al., 1998). After periph-eral injection of LPS, dendritic cells near the vagus nerveexpress IL-1 (Goehler et al., 1999). The medullary visceralzone was identified as a neural station relating vagal signalsto the PVN (Yang et al., 2000). These studies showed acti-vation of appropriate vagal related targets after peripheralimmune challenge, further solidifying this immune sensorypathway.
Two major advances demonstrated that the concept ofneural involvement could be expanded beyond that ofvagal afferents. Roth et al. showed that whereas vagotomyblocked fever induced by intraperitoneally injected LPS, itdid not block fever induced by intramuscularly injectedLPS. However, in a local subcutaneous inflammationmodel, fever was blocked by local anesthetic. Therefore,primary sensory never fibers may be able to relate inflam-matory signals to the brain (Roth and De Souza, 2001).Similarly, fever induced by injecting LPS into the soft pal-ate could be prevented by severing the glossopharyngealnerve. Thus, the inflammatory status of the entire bodymay be monitored through the various afferent neuralpathways. Another exciting addition to the neuroimmunecommunication is the discovery of the cholinergic anti-inflammatory pathway by Tracey and co-workers. Stimula-tion of vagal efferents powerfully inhibited the productionof TNF and protected animals from LPS induced endo-toxic shock (Borovikova et al., 2000). These researchersfound that the nicotinic acetylcholine a7 receptor mediatesthese anti-inflammatory effects (Wang et al., 2003). Thus,the nervous system can sense immune activities throughoutthe soma and regulate the function of the immune systemvia both sympathetic and parasympathetic outflows.Doubts linger, however, about the cholinergic hypothesisbecause acetylcholine a7 receptor have been found in sym-pathetic postganglionic neurons (Zhous et al., 2001).Therefore, it is possible that sympathetic, rather than para-sympathetic, outflow exerts anti-inflammatory effects.
Work in this period also attempted to discover thepoints where different neuroimmune pathways may

732 N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735
converge. After all, if similar illness behavior outcomes canbe induced by these pathways, should the diverse circuitseventually affect a common central regulator? One idea isthat activation of all of the ascending pathways leads tothe production of IL-1 in the brain. Evidence for andagainst vagally mediated CNS production of IL-1 has beenreported (for review see Dantzer et al., 2000). An unsatisfy-ing aspect of this proposal is that if neural transmission ofthe immune signal to the brain needs to induce brain to re-synthesize the peripheral messenger IL-1, then some of theadvantage of the speed of neural transmission is negated.In addition, unpublished observations from the Quan labshow that fever induced by peripheral, localized inflamma-tion does not require CNS production of IL-1.
Another critical issue is where is IL-1 produced andwhere does it act in the brain? Peripheral LPS inducesIL-1 mRNA expression highlights CVOs, choroid plexus,meninges, and blood vessels, suggesting the ventricles andthe CSF flow may be the conduit of IL-1 propagation inthe brain (Quan et al., 1998). In adult brain, the functionalIL-1 receptor, IL-1R1, has been found on cerebral bloodvessels, and choroid plexus (Cunningham et al., 1992),but conspicuously missing from neurons of the hypotha-lamic regions controlling relevant autonomic and neuroen-docrine responses (French et al., 1999). In addition,peripheral LPS induced expression of IjBa, a marker ofcytokine-induced cellular activation via the classical NF-jB pathway, was located in non-neuronal cells of the brain,especially in endothelial cells (Quan et al., 1997). Thesefindings suggest that in the brain, IL-1 may act on neuronsindirectly by downstream signaling molecules, e.g. theprostaglandins and nitric oxide. Another possibility is thatIL-1 may act on neurons through non-canonical signalingpathways. Bartfai’s group has made great progress show-ing that ceremide can mediate rapid neuronal effectsinduced by IL-1, independent of transcriptional activation(Sanchez-Alavez et al., 2006).
4.2. Circumventricular organs: 1998–2007
Work by Maness et al. confirmed entry of IL-1 into CVOsdid not leak into adjacent brain tissues and Peruzzo et al.(2000) confirmed the tanycytic barrier at the CVOs. Thus,the relay hypothesis of Arimura was favored over the leakagehypothesis. Goehler et al. showed that LPS stimulated IL-1release from immune cells within the area postrema. ThisIL-1 may, in turn, stimulate the neural elements present atthe CVO which project to and from areas of the CNS behindthe BBB. Work in the pineal showed that cytokine releasecould control melatonin secretion and that cytokine secre-tion itself can be altered by norepinephrine and IL-1.
4.3. The blood–brain barrier transport of cytokines:
1998–2007
The number of cytokines examined for BBB transportexpanded greatly during this period. As examples, brain-
derived neurotrophic factor, neurotrophins, ciliary neuro-trophic factor, EGF, leukemia inhibitory factor, and LIFwere found to cross by saturable transport systems. Cyto-kine-induced neutrophil chemoattactant-1 crosses by thenon-saturable mechanism of transmembrane diffusion,the largest substance known to use this mechanism. TGFbeta1 and glial cell derived neurotrophic factor were foundto not cross the BBB. IL-2 has a saturable transport systemdirected in the brain-to-blood direction only, the only cyto-kine so far know to have a saturable efflux system. Reichlinand coworkers further showed that the brain can be a sub-stantial source of blood-borne cytokines, especially afterneuroimmune stimulation (Chen et al., 2000).
TNF transport was found to be mediated by the TNFreceptors (Pan and Kastin, 2002). This contrasts to the sit-uation for IL-1 and EGF, where the transporters are notidentified, but known to not be the same proteins as thereceptors.
Work continued to show that the transporters for cyto-kines are not static, but adapted to or affected by physio-logical and disease states. The rates at which cytokinesare transported across the BBB vary as a function ofgenetic strain, age, time of day, and CNS region (bothwithin brain and spinal cord). Traumatic and ischemicinjuries to brain or spinal cord also affect transport rates.
As pharmacokinetic evidence for dynamic and variedtransport of cytokines accumulated, the question arose asto what, if any, role these transporters played in neuroimmu-nology. As already discussed, multiple mechanisms areestablished by which circulating cytokines can affect CNSfunctioning without directly crossing the BBB, includingafferent nerve stimulation, action at circumventricularorgans, disruption and other alterations in BBB function,and induction of BBB secretions. In 2001, it was shown thatthe impairment in learning induced by intravenously injectedIL-1 largely depended on its ability to cross the BBB to enterthe posterior division of the septum (Banks et al., 2001).Most recently, damage to dopamine neurons in the substan-tia nigra by peripherally administered LPS has been shownto depend on peripherally-released TNF crossing the BBBto induce release of CNS stores of TNF (Qin et al., 2007).
4.4. Secretions of BBB cells: 1998–2007
Although the number of publications in this area arestill few, they constitute an important and growing litera-ture. The induction by LPS of COX-2 in brain blood ves-sels correlated with the febrile response (Cao et al., 1997)and subsequent work from several laboratories has shownthat prostaglandins released from brain endothelial cellsmediates the fever induced by LPS (Inoue et al., 2002).Table 2 lists the cytokines thus far found to be secretedby BBB cells. Secretion was found to be both constitutiveand inducible or modified by LPS, other cytokines,cocaine, HIV-1 related proteins, and other immunoactivesubstances (Hofman et al., 1999; Reyes et al., 1999). Boththe endothelial cells forming the vascular BBB and ependy-

Table 2Cytokines secreted from BBB (choroid plexus ependymal or brainendothelial) cells
IL-1IL-3IL-6IL-8IL-10Endothelin-1GM-CSFMCP-1Monokine induced by IFN-gamma (MIG)NGFRANTESTGF-betaTNF
N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735 733
mal cells forming the blood-cerebrospinal fluid barrier, butnot human umbilical cord cells, are capable of cytokinesecretion. Secretion from monolayer cultures of brainendothelial cells were found to be polarized; that is, somesubstances, such as IL-6, were preferentially secreted fromeither the luminal or abluminal cell membrane surface(Verma et al., 2006). IL-6 secretion from the luminal sur-face was also found to be more robust when LPS wasapplied to the abluminal in comparison to the luminalsurface.
5. Beyond 2007: Future directions for BBB transport of
cytokines
Research in the last twenty years has clearly demon-strated that neuroimmune communication exists via multi-ple pathways. Both neural and humoral pathways areinvolved in relating information between the nervous andthe immune systems. Afferent pathways are able to provideinflammatory signals to the brain and efferent transmissionis able to profoundly modulate both innate and adaptiveimmunity. Within the neuroimmune system, a signalingmolecule can arise from multiple sources to act at multipletargets. These achievements have laid an important foun-dation for our understanding of the neuroimmune systemas an interconnected complex. Interestingly, the findingthat the vagal, BBBs, and circumventricular pathways allcome together in a single anatomical location makes thedorsal vagal complex a very interesting area for thoseinterested in the integration of neuroimmune concepts(Banks et al., 2004; Marvel et al., 2003).
The accumulated findings also call for reevaluation andrethinking of the current paradigms and assumptions inregards to the structure of neuroimmune communication.As examples:
If the primary sensory nerve fibers can transmit inflam-matory signal, how is the immune signal coded so that theCNS can distinguish the immune information from othersensory signals such as pain, temperature, or pressure?
IL-1 and prostaglandin receptors have been found inperipheral tissue, in vagal ganglia, in spinal cord, and in
brain. The assumption that these mediators need to act inthe brain to trigger immune response induced CNS effectsmay not be correct. IL-1 and prostaglandins may well acton other points of the pathway to induce sickness behaviorand regulate other neuroimmune phenomena. Much of themodeling of the neuroimmune pathways has been basedupon peripheral LPS induced effects. In reality, many infec-tions cause illness behavior without detectable levels ofLPS and cytokines in the circulation. Therefore, localinflammation may engage neuroimmune pathways thatare independent of either blood or brain cytokineexpression.
The linear pathway model assumes that a step-by-stepcascade of signaling molecules are induced, passing theinformational batons between the two systems. This modelneeds revision, and will likely be replaced by a network-likemodel in which signaling molecules are connected to oneanother in multiple ways and at multiple levels. The rela-tions of one signaling molecule to another will likely bestate-dependent.
Using IL-1 as the central messenger for peripheralimmune signal also brings into focus an importantdilemma regarding the role of IL-1 in the brain. IL-1may present two conflicting meanings: (1) IL-1 is producedas a consequence of CNS infection; in this case, IL-1 mayplay the traditional immunological role of inducing infiltra-tion of leukocytes to the brain. (2) IL-1 is present in thebrain as a consequence of peripheral inflammation; in thiscase, brain IL-1 should serve primarily as a signaling mol-ecule that activates central neural circuits related to illnessbehaviors. The fact that IL-1 receptors are highly expressedin the blood vessels, meninges, and choroid plexus, siteswhere typically central inflammatory responses are mani-fested, reminds neurobiologists that besides the neuromod-ulatory role, IL-1 has not forgotten its duty as aninflammatory cytokine in the brain. Interestingly, intrace-rebrally injected IL-1 can induce leukocyte infiltration intothe brain, but intravenous IL-1 does not. Further, periph-eral LPS injection can prevent central IL-1 induced leuko-cyte infiltration (Ching et al., 2006) suggesting that biologymust carefully distinguish the two meanings of central IL-1such that the presence of central IL-1 is not always inter-preted as a signal for central inflammation and that centralinflammation signaling may even be suppressed in the faceof severe peripheral inflammation, probably to prevent therecruitment of highly activated leukocytes from enteringthe brain. Therefore, the effects of central IL-1 are contextdependent. How neuromodulatory actions of central IL-1are modulated by other factors remains to be clarified.
Work so far indicates that transport across the BBBoccurs for many cytokines. The extent to which such trans-port is modulated, adaptive, and responsive to disease andinjury suggests that transport is important in many realmsinvolving communication between the immune and ner-vous systems. However, work to date is largely based onpharmacokinetic methods and raises, but cannot in isola-tion answer, key questions. One such question is what

734 N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735
effects does blood-to-brain passage of cytokines exert onthe CNS and immune system? An important extension ofthis question is what role do these transporters play in neu-roprotection and neuroregeneration after CNS injury?Some of the most promising therapeutics being tested fortreatment of CNS injury and degenerative diseases arecytokines that have transport systems across the BBB(Xiang et al., 2005). Effective delivery of these substancesto the CNS may depend more on producing enzymaticallystable, long-lasting analogs than overcoming the BBB. Assuch, another question is whether the cytokine transporterscan be used to deliver therapeutics to the CNS. Finally, theCNS as a source of peripheral cytokines needs furtherinvestigation.
The ability of the BBB to secrete cytokines indicates thatbrain endothelial and ependymal cells are in cross talk withother elements of the neurovascular unit and neuroimmuneaxis. The ability of a wide range of immunoactive sub-stances, including cytokines themselves, to regulate BBBsecretion means that BBB cells can be activated in a man-ner analogous to microglia, astrocytes, and immune cells.Polarized responses provide a type of neuroimmune axiscommunication unique to the BBB: because the BBB isthe only cell type which is simultaneously in both theperiphery and the CNS, polarized stimulation and secre-tion allows it to receive input from one side of the neuroim-mune axis and produce a secreted response into the otherside.
6. Conclusions
Understanding how the immune and central nervoussystems communicate with one another is central to under-standing the interactions between brain, behavior, andimmunity. We examined here four major pathways bywhich such communications can occur: neural routes, cir-cumventricular organs, cytokine transport across theBBB, and secretion of immune-active substances by thecells which constitute the BBBs. All these areas have seentremendous development in the last 10 years. All faceimportant future questions for which the tools and tech-niques for addressing them are available. Additional chal-lenges to neuroimmunology include the discovery ofother mechanisms by which the immune, nervous, andendocrine systems communicate. The ultimate challengemay be to elucidate how these various mechanisms of com-munication interact with one another, under what circum-stances one mechanism may come to dominate or recederelative to the others, and what conditions control theseshifting interactions among mechanisms of brain-bodycommunication.
Acknowledgments
This work was supported by R01 NS40098 (N.Q.), R01AI059089 (N.Q.), VA Merit Review (W.A.B.), R21
DA019396 (W.A.B.), R01 NS050547 (W.A.B.), R01NS051334 (W.A.B.).
References
Ader, R., Cohen, N., 1975. Behaviorally conditioned immunosuppression.Psychosom. Med. 37, 333–350.
Ban, E., Haour, F., Lenstra, R., 1992. Brain interleukin 1 gene expressioninduced by peripheral lipopolysaccharide administration. Cytokine 4,48–54.
Banks, W.A., Kastin, A.J., Durham, D.A., 1989. Bidirectional transportof interleukin-1 alpha across the blood–brain barrier. Brain Res. Bull.23, 433–437.
Banks, W.A., Farr, S.A., La Scola, M.E., Morley, J.E., 2001. Intravenoushuman interleukin-1a impairs memory processing in mice: dependenceon blood–brain barrier transport into posterior division of the septum.J. Pharmacol. Exp. Ther. 299, 536–541.
Banks, W.A., Niehoff, M.L., Zalcman, S., 2004. Permeability of the mouseblood–brain barrier to murine interleukin-2: predominance of asaturable efflux system. Brain Behav. Immun. 18, 434–442.
Besedovsky, H.O., de Rey, A., Sorkin, E., Da Prada, M., Keller, H.H.,1979. Immunoregulation mediated by the sympathetic nervous system.Cell Immunol. 48, 346–355.
Blalock, J.E., Bost, K.I., Smith, E.M., 1985. Neuroendocrine peptidehormones and their receptors in the immune system. Production,processing and action. J. Neuroimmunol. 10, 31–40.
Blatteis, C.M., Bealer, S.L., Hunter, W.S., Llanos, Q.J., Ahokas, R.A.,Mashburn Jr., T.A., 1983. Suppression of fever after lesions of theanteroventral third ventricle in guinea pigs. Brain Res. Bull. 11,519–526.
Bluthe, R.M., Walter, V., Parnet, P., Laye, S., Lestage, J., Verrier, D.,Poole, S., Stenning, B.E., Kelley, K.W., Dantzer, R., 1994. Lipopoly-saccharide induces sickness behaviour in rats by a vagal mediatedmechanism. C R Acad. Sci. III 317, 499–503.
Borovikova, L.V., Ivanova, S., Zhang, M., Yang, H., Botchkina, G.I.,Watkins, L.R., Wang, H., Abumrad, N., Eaton, J.W., Tracey, K.J.,2000. Vagus nerve stimulation attenuates the systemic inflammatoryresponse to endotoxin. Nature 405, 458–462.
Breder, C.D., Dinarello, C.A., Saper, C.B., 1988. Interleukin-1 immuno-reactive innervation of the human hypothalamus. Science 240,321–324.
Cao, C., Watanabe, Y., Yamagata, K., Matsumura, K., 1997. Involve-ment of cyclooxygenase-2 in LPS-induced fever and regulation of itsmRNA by LPS in the rat brain. Am. J. Physiol. 272, R1712–R1725.
Chen, G., Castro, W.L., Chow, H.H., Reichlin, S., 1997. Clearance of125 I-labelled interleukin-6 from brain into blood following intracerebro-ventricular injection in rats. Endocrinology 138, 4830–4836.
Chen, G., McCuskey, R.S., Reichlin, S., 2000. Blood interleukin-6 andtumor necrosis factor-a elevation after intracerebroventricular injec-tion of Escherichia coli endotoxin in the rat is determined by twoopposing factors: peripheral induction by LPS transferred from brainto blood and inhibition of peripheral response by a brain-mediatedmechanism. nim 8, 59–69.
Ching, S., Zhang, H., Lai, W., Quan, N., 2006. Peripheral injection oflipopolysaccharide prevents brain recruitment of leukocytes inducedby central injection of interleukin-1. Neuroscience 137, 717–726.
Cunningham Jr., E.T., Wada, E., Carter, D.B., Tracey, D.E., Battey, J.F.,De Souza, E.B., 1992. In situ histochemical localization of type 1interleukin-1 receptor messenger RNA in the central nervous system,pituitary, and adrenal gland of the mouse. J. Neurosci. 12,1101–1114.
Dantzer, R., Konsman, J.P., Bluthe, R.M., Kelley, K.W., 2000. Neuraland humoral pathways of communication from the immune system tothe brain: parallel or convergent? Auton. Neurosci. 85, 60–65.
Ek, M., Kurosawa, M., Lundeberg, T., Ericsson, A., 1998. Activation ofvagal afferents after intravenous injection of interleukin-1 beta: role ofendogenous prostaglandins. Neuroscience 18, 9471–9479.

N. Quan, W.A. Banks / Brain, Behavior, and Immunity 21 (2007) 727–735 735
Ericsson, A., Kovacs, K.J., Sawchenko, P.E., 1994. A functionalanatomical analysis of central pathways subserving the effects ofinterleukin-1 on stress-related neuroendocrine neurons. J. Neurosci.14, 897–913.
Fabry, Z., Fitzsimmons, K.M., Herlein, J.A., Moninger, T.O., Dobbs,M.B., Hart, M.N., 1993. Production of the cytokines interleukin 1 and6 by murine brain microvessel endothelium and smooth musclepericytes. J. Neuroimmunol. 47, 23–34.
Felten, D.L., Felten, S.Y., Carlson, S.L., Olschowka, J.A., Livnat, S.,1985. Noradrenergic and peptidergic innervation of lymphoid tissue. J.Immunol. 135, 755s–765s.
French, R.A., VanHoy, R.W., Chizzonite, R., Zachary, J.F., Dantzer, R.,Parnet, P., Bluthe, R.M., Kelley, K.W., 1999. Expression andlocalization of p80 and p68 interleukin-1 receptor proteins in thebrain of adult mice. J. Neuroimmunol. 93, 194–202.
Gatti, S., Bartfai, T., 1993. Induction of tumor necrosis factor-alphamRNA in the brain after peripheral endotoxin treatment: compar-ison with interleukin-1 family and interleuin-6. Brain Res. 624,291–294.
Goehler, L.E., Relton, J.K., Dripps, D., Kiechle, R., Tartaglia, N., Maier,S.F., Watkins, L.R., 1997. Vagal paraganglia bind biotinylatedinterleukin-1 receptor antagonist: a possible mechanism for immune-to-brain communication. Brain Res. Bull. 43, 357–364.
Goehler, L.E., Gaykema, R.P., Nguyen, K.T., Lee, J.E., Tilders, F.J.,Maier, S.F., Watkins, L.R., 1999. Interleukin-1 beta in immune cells ofthe abdominal vagus nerve: a link between the immune and nervoussystems? J. Neurosci. 19, 2799–2806.
Guerrero, J.M., Reiter, R.J., 1992. A brief survey of pineal gland-immunesystem interrelationships. Endocr. Res. 18, 91–113.
Habif, D.V., Lipton, R., Cantell, K., 1975. Interferon crosses the blood-cerebrospinal fluid barrier in monkeys. Proc. Soc. Exp. Biol. Med. 149,287–289.
Hansen, M.K., O’Connor, K.A., Goehler, L.E., Watkins, L.R., Maier,S.F., 2001. The contribution of the vagus nerve in interleukin-1 beta-induced fever is dependent on dose. Am. J. Physiol. Regul. Integr.Comp. Physiol. 280, R929–R934.
Hofman, F., Chen, P., Incardona, F., Zidovetzki, R., Hinton, D.R., 1999.HIV-tat protein induces the production of interleukin-8 by humanbrain-derived endothelial cells. J. Neuroimmunol. 94, 28–39.
Inoue, W., Matsumura, K., Yamagata, K., Takemiya, T., Shiraki, T.,Kobayashi, S., 2002. Brain-specific endothelial induction of prosta-glandin E(2) synthesis enzymes and its temporal relation to fever.Neurosci. Res. 44, 51–61.
Komaki, G., Arimura, A., Koves, K., 1992. Effect of intravenous injectionof IL-1b on PGE2 levels in several brain areas as determined bymicrodialysis. Am. J. Physiol. 262, E246–E251.
Luheshi, G.N., Bluthe, R.M., Rushforth, D., Mulcahy, N., Konsman,J.P., Goldbach, M., Dantzer, R., 2000. Vagotomy attenuates thebehavioral but not the pyrogenic effects of interlekin-1 in rats. Auton.Neurosci. 85, 127–132.
Marvel, F.A., Chen, C.-C., Badr, N., Gaykema, R.P.A., Goehler, L.E.,2003. Reversible inactivation of the dorsal vagal complex blockslipopolysaccharide-induced social withdrawal and c-Fos expression incentral autonomic nuclei. Brain Behav. Immun. 18, 123–134.
Medawar, P., 1948. Immunity to homologous grafted skin. III. The fate ofskin homografts transplanted to the brain and to the anterior chamberof the eye. Br. J. Exp. Pathol. 29, 58–69.
Pan, W., Kastin, A.J., 2002. TNFa transport across the blood–brainbarrier is abolished in receptor knockout mice. Exp. Neurol. 174,193–200.
Pan, W., Banks, W.A., Kennedy, M.K., Gutierrez, E.G., Kastin, A.J.,1996. Differential permeability of the BBB in acute EAE: enhancedtransport of TNF-a. Am. J. Physiol. 271, E636–E642.
Peruzzo, B., Pastor, F.E., Blazquez, J.L., Schobitz, K., Pelaez, B., Amat,P., Rodriguez, E.M., 2000. A second look at the barriers of the medialbasal hypothalamus. Exp. Brain Res. 132, 10–26.
Qin, L., Wu, X., Block, M.L., Liu, Y., Breese, G.R., Hong, J.S., Knapp,D.J., Crews, F.T., 2007. Systemic LPS causes chronic neuroinflam-mation and progressive neurodegeneration. GLIA 55, 453–462.
Quan, N., Herkenham, M., 2002. Connecting cytokines and brain: areview of current issues. Histol. Histopathol. 46, 178–192.
Quan, N., Sundar, S.K., Weiss, J.M., 1994. Induction of interleukin-1 invarious brain regions after peripheral and central injections oflipopolysaccharide. J. Neuroimmunol. 49, 125–134.
Quan, N., Whiteside, M., Kim, L., Herkenham, M., 1997. Induction ofinhibitory factor jBa mRNA in the centralnervous system afterperipheral lipopolysaccharide administration: an in situ hybridizationhistochemistry study in the rat. Proc. Natl. Acad. Sci. USA 94,10985–10990.
Quan, N., Whiteside, M., Herkenham, M., 1998. Time course andlocalization patterns of interleukin-1b messenger RNA expression ibrain and pituitary after peripheral administration of lipopolysaccha-ride. Neuroscience 83, 281–293.
Reyes, T.M., Fabry, Z., Coe, C.L., 1999. Brain endothelial cell productionof a neuroprotective cytokine, interleukin-6, in response to noxiousstimuli. Brain Res. 851, 215–220.
Romanovsky, A.A., Simon, C.T., Szekely, M., Kulchitsky, V.A., 1997.The vagus nerve in the thermoregulatory response to systemicinflammation. Am. J. Physiol. 273, R407–R413.
Roth, J., De Souza, G.E., 2001. Fever induction pathways: evidence fromresponses to systemic or local cytokine formation. Braz. J. Med. Biol.Res. 34, 301–314.
Sanchez-Alavez, M., Tabarean, I.V., Behrens, M.M., Bartfai, T., 2006.Ceramide mediates the rapid phase of febrile response to IL-1beta.Proc. Natl. Acad. Sci. USA 103, 2904–2908.
Verma, S., Nakaoke, R., Dohgu, S., Banks, W.A., 2006. Release ofcytokines by brain endothelial cells: a polarized response to lipopoly-saccharide. Brain Behav. Immun. 20, 449–455.
Wang, H., Yu, M., Ochani, M., Amella, C.A., Tanovic, M., Susarla, S.,Li, J.H., Wang, H., Yang, H., Ulloa, L., Al-Abed, Y., Czura, C.J.,Tracey, K.J., 2003. Nicotinic acetycholine receptor alpha7 subunit isan essential regulator of inflammation. Nature 421, 384–388.
Watkins, L.R., Wiertelak, E.P., Goehler, L.E., Mooney-Heiberger, K.,Martinez, J., Furness, L., Smith, K.P., Maier, S.F., 1994. Neurocir-cuitry of illness-induced hyperalgesia. Brain Res. 639, 283–299.
Watkins, L.R., Maier, S.F., Goehler, L.E., 1995. Cytokine-to-braincommunication: a review & analysis of alternative mechanisms. LifeSci. 57, 1011–1026.
Widner, H., Brundin, P., 1988. Immunological aspects of grafting in themammalian central nervous system. A review and speculative synthe-sis. Brain Res. 472, 287–324.
Xiang, S., Pan, W., Kastin, A.J., 2005. Strategies to create a regeneratingenvironment for the injured spinal cord. Curr. Pharmaceut. Design 11,1267–1277.
Yang, Z.J., Rao, Z.R., Ju, G., 2000. Evidence for the medullary visceralzone as a neural station of neuroimmunomodulation. Neurosci. Res.38, 237–247.
Zhous, Y., Deneris, E., Zigmond, R.E., 2001. Nicotinic acetylcholinereceptor subunit proteins alpha7 and beta4 decrease in the superiorcervical ganglion after axotomy. J. Neurobiol. 46, 178–192.
Related Documents











