Braz J Med Biol Res 39(11) 2006 Blastocyst-endometrium interaction: intertwining a cytokine network 1 Semillero de Investigación SIMBIOSIS, 2 Grupo Reproducción - Corporación Biogénesis, Universidad de Antioquia, Medellín, Colombia W.A. Castro-Rendón 1,2 , J.F. Castro-Álvarez 1,2 , C. Guzmán-Martinez 2 and J.C. Bueno-Sanchez 1,2 Abstract The successful implantation of the blastocyst depends on adequate interactions between the embryo and the uterus. The development of the embryo begins with the fertilized ovum, a single totipotent cell which undergoes mitosis and gives rise to a multicellular structure named blastocyst. At the same time, increasing concentrations of ovarian steroid hormones initiate a complex signaling cascade that stimulates the differentiation of endometrial stromal cells to decidual cells, preparing the uterus to lodge the embryo. Studies in humans and in other mammals have shown that cytokines and growth factors are produced by the pre-implantation embryo and cells of the reproduc- tive tract; however, the interactions between these factors that con- verge for successful implantation are not well understood. This review focuses on the actions of interleukin-1, leukemia inhibitory factor, epidermal growth factor, heparin-binding epidermal growth factor, and vascular endothelial growth factor, and on the network of their interactions leading to early embryo development, peri-implantatory endometrial changes, embryo implantation and trophoblast differen- tiation. We also propose therapeutical approaches based on current knowledge on cytokine interactions. Correspondence W.A. Castro-Rendón Grupo Reproducción Sede de Investigación Universitaria Cll 62 #52-59, Laboratorio 534 Universidad de Antioquia Medellín Colombia Fax: +57-4-210-6470 E-mail: [email protected] Received November 3, 2005 Accepted June 21, 2006 Key words • Embryo implantation • Trophoblast • Cytokines • Prostaglandins • Aspirin Introduction Some decades ago cytokines were known as intercellular messengers with a unique function in the immune system. Subse- quently, experimental evidence showed that they participate in many events of immu- noendocrine interaction, characteristic of complex signaling networks. Thus, the sig- nals from both the embryo and the mother during early embryo development converge around the actions of leukemia inhibitory factor (LIF), interleukin-1 (IL-1) and colony- stimulating factors (CSF), among others (1), supported by an adequate hormonal microen- vironment and the recognition of the fetus by the maternal immune system. The embryo plays an active role in the modulation of the immunoendocrine activity during endome- trial implantation, particularly by participat- ing in (in the murine model) or enhancing (as in humans) the endometrial inflammatory reaction known as decidualization, thus regu- lating the production of maternal soluble factors that might be necessary for the inva- sion of the trophoblast for embryo lodging Brazilian Journal of Medical and Biological Research (2006) 39: 1373-1385 ISSN 0100-879X Review

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1373
Braz J Med Biol Res 39(11) 2006
Blastocyst-endometrium interactions
Blastocyst-endometrium interaction:intertwining a cytokine network
1Semillero de Investigación SIMBIOSIS,2Grupo Reproducción - Corporación Biogénesis,Universidad de Antioquia, Medellín, Colombia
W.A. Castro-Rendón1,2,J.F. Castro-Álvarez1,2,
C. Guzmán-Martinez2
and J.C. Bueno-Sanchez1,2
Abstract
The successful implantation of the blastocyst depends on adequateinteractions between the embryo and the uterus. The development ofthe embryo begins with the fertilized ovum, a single totipotent cellwhich undergoes mitosis and gives rise to a multicellular structurenamed blastocyst. At the same time, increasing concentrations ofovarian steroid hormones initiate a complex signaling cascade thatstimulates the differentiation of endometrial stromal cells to decidualcells, preparing the uterus to lodge the embryo. Studies in humans andin other mammals have shown that cytokines and growth factors areproduced by the pre-implantation embryo and cells of the reproduc-tive tract; however, the interactions between these factors that con-verge for successful implantation are not well understood. This reviewfocuses on the actions of interleukin-1, leukemia inhibitory factor,epidermal growth factor, heparin-binding epidermal growth factor,and vascular endothelial growth factor, and on the network of theirinteractions leading to early embryo development, peri-implantatoryendometrial changes, embryo implantation and trophoblast differen-tiation. We also propose therapeutical approaches based on currentknowledge on cytokine interactions.
CorrespondenceW.A. Castro-Rendón
Grupo Reproducción
Sede de Investigación Universitaria
Cll 62 #52-59, Laboratorio 534
Universidad de Antioquia
Medellín
Colombia
Fax: +57-4-210-6470
E-mail: [email protected]
Received November 3, 2005
Accepted June 21, 2006
Key words• Embryo implantation• Trophoblast• Cytokines• Prostaglandins• Aspirin
Introduction
Some decades ago cytokines were knownas intercellular messengers with a uniquefunction in the immune system. Subse-quently, experimental evidence showed thatthey participate in many events of immu-noendocrine interaction, characteristic ofcomplex signaling networks. Thus, the sig-nals from both the embryo and the motherduring early embryo development convergearound the actions of leukemia inhibitoryfactor (LIF), interleukin-1 (IL-1) and colony-
stimulating factors (CSF), among others (1),supported by an adequate hormonal microen-vironment and the recognition of the fetus bythe maternal immune system. The embryoplays an active role in the modulation of theimmunoendocrine activity during endome-trial implantation, particularly by participat-ing in (in the murine model) or enhancing (asin humans) the endometrial inflammatoryreaction known as decidualization, thus regu-lating the production of maternal solublefactors that might be necessary for the inva-sion of the trophoblast for embryo lodging
Brazilian Journal of Medical and Biological Research (2006) 39: 1373-1385ISSN 0100-879X Review

1374
Braz J Med Biol Res 39(11) 2006
W.A. Castro-Rendón et al.
into the endometrium (2). These typicalchanges of decidualization can be inducedwith oil, air bubbles or concanavalin A andhave also been observed in deciduomata (3)and ectopic pregnancies.
The decidual inflammatory reaction dif-fers from the classical description of inflam-mation that occurs in response to injury. Inparticular, in decidua, the recruitment ofleukocytes is mainly dominated by naturalkiller (NK) cells that in other contexts arerecognized as cytotoxic cells which partici-pate in the antitumoral and antiviral immuneresponse. On the other hand, the transforma-tion of the uterine stromal cells into decidualcells is demonstrated by prolactin produc-tion, increased IL-15 concentrations and ac-tivation of the arachidonic acid pathway.These factors enhance the decidual reactionin response to embryo invasion. In general,the decidual environment is able to regulateembryo differentiation, mainly the tropho-ectodermic layer, which is dependent onepithelial contact and extracellular matrixproteins from the stroma in addition to cyto-kines (4). These trophoectodermic/tropho-blast cell subpopulations are producers ofhormones and cytokines whose endocrineaction can cross the maternal-fetal interfaceand directly influence the systemic physi-ological changes that characterize gestation.
In this intricate network of interactionscytokines act as intermediary links of thematernal-fetal relationship including theevents related to decidualization, implanta-tion, placentation, embryogenesis, and fetalgrowth. The aim of this review is to identifycertain basic aspects of cytokine actions inmaternal-fetal interaction and to suggest newpossibilities for immunoendocrine regula-tion, which could be useful to understandsome disorders of early pregnancy in hu-mans and in other animal species.
Early embryo development
The establishment of the pre-implantatory
embryo begins when the oocyte is fertilizedby a spermatozoid. When the penetration ofthe male gamete into the oocyte is con-cluded, maternal and paternal pronuclei mi-grate to the center of the cell and the nuclearmembranes fuse to allow the recombinationof the genetic material. This is the first stageof embryo development, named zygote. Sub-sequently mitotic divisions of the zygotegive rise to the stages of two, four and eightcells, until the morula (5). Then, the morulaundergoes a process of cell polarization,where the outermost cells form the trophec-toderm (external embryo layer). Simulta-neously, the expression and activation ofNa+/K+ ATPase pumps transport sodium andwater towards the interior of the morula thatgenerates a cavity known as the blastocele(6). Meanwhile, a group of pluripotent cellsdenoted the inner cell mass is established ina pole of the embryo, between the trophecto-derm and the blastocele. These changes giverise to the blastocyst, the final step of earlyembryo development.
Previous studies have provided informa-tion about the regulatory effects of the IL-1system in human pre-implantatory embryos,as well as in embryos of other species (Table1). The IL-1 system is composed of twoagonists (IL-1α and IL-1ß), one inhibitordesignated IL-1 receptor antagonist (IL-l ra),and two receptors known as IL-1 receptorstype I (IL-1R tI) and type II. Interestingly,only IL-1R tI transduces a signal in responseto IL-1 (7). Single blastomeres from humanpreimplantation embryos cultured in vitroexpress the mRNAs for IL-1ß, IL-1R tI andIL-1 ra (7). In bovine embryos the additionof IL-1ß after in vitro fertilization enhancesthe recovery of blastocysts under conditionsof culture at high density (25-30 embryos/drop) but not at low density (10 embryos/drop). Thus, IL-1ß can modulate embryodevelopment towards the blastocyst stage(8).
Human embryos cultured in vitro pro-duce high concentrations of IL-1α and IL-

1375
Braz J Med Biol Res 39(11) 2006
Blastocyst-endometrium interactions
Table 1. Cytokines, prostaglandins, and growth factors implicated in embryo-endometrium interaction.
Compounds Biological actions Reference
LIF Blastocyst implantation Stewart et al., 1992 (40)Early embryogenesis as an embryotrophin Lavranos et al., 1995 (16)Decidualization Chen et al., 2000 (41)
LIFR Decidualization and placentation Ni et al., 2002 (31)IL-1 Successful implantation Sheth et al., 1991 (9)
Induces LIF production Arici et al., 1995 (43)Induces COX-2 Huang et al., 1998 (26)Endometrial changes McMaster et al., 1992 (23)GM-CSF and CSF-1 production Garcia-Lloret et al., 1994 (57)
IL-1 ra Failure of implantation Simon et al., 1994 (49)IL-1R tI Embryo development Kruessel et al., 1997 (10)EGF Blastocyst differentiation Kaye, 1997 (1)
Angiogenic factor Moller et al., 2001 (37)Trophoblastic differentiation Kliman et al., 1990 (55)Induces LIF production Arici et al., 1995 (43)
HB-EGF Blastocyst growth, zona-hatching and Das et al., 1994 (39)trophoblast outgrowth Leach et al., 1999 (36)
Implantation reaction Wang et al., 1994 (46)IL-15 Decidual marker Okada et al., 2000 (19)IGFBP-1 Decidual marker Dimitriadis et al., 2002 (20)TNF-α Endometrial changes von Wolff et al., 1999 (24)
GM-CSF and CSF-1 production Garcia-Lloret et al., 1994 (57)PGE2 Endometrial changes Jabbour et al., 2003 (33)
Multiple female reproductive failures Lim et al., 1997 (28)Reese et al., 2001 (29)
Differentiation of NK cells Linnemeyer et al., 1993 (35)VEGF Angiogenic factor Moller et al., 2001 (37)GM-CSF Trophoblastic differentiation Garcia-Lloret et al., 1994 (57)CSF-1 Trophoblastic differentiation Garcia-Lloret et al., 1994 (57)
LIF = leukemia inhibitory factor; LIFR = LIF receptor; IL-1 = interleukin-1; IL-1 ra = IL-1 receptor antagonist; IL-1R tI = IL-1 receptor type I; EGF = epidermal growth factor; HB-EGF = heparin binding-EGF; IL-15 =interleukin-15; IGFBP-1 = insulin-like growth factor-binding protein 1; TNF-α = tumor necrosis factor alpha;PGE2 = prostaglandin E2; VEGF = vascular endothelial growth factor; GM-CSF = granulocyte-macrophagecolony-stimulating factor; CSF-1 = colony stimulating factor-1.
1ß (>60 and >80 pg/mL, respectively), andthe presence of these cytokines has beencorrelated with successful implantation aftertransfer to the uterine cavity (9). Kruessel etal. (10) demonstrated the expression of IL-1ß, IL-1R tI and IL-1 ra mRNA by RT-PCRin single-cell preimplantation mouse em-bryos. They did not detect the correspondingmRNAs at the stage of 2-cell-embryos, andonly IL-1 ra was expressed at the 8-cell-stage in a few embryos. Thus, the stagewhere IL-1ß, IL-1R tI and IL-1 ra could bedetected was the compact morula stage.Therefore, this expression was increased byongoing maturation and reached its highest
level at the stage of hatched blastocyst (10).The aim of assisted reproductive tech-
niques and embryo culture is to obtain em-bryos of good quality with the ability toimplant into the uterus. In this context, co-culture systems of embryos on cell mono-layers enhance early embryo development.Different types of cells such as uterine fibro-blasts, human tubular cells, endometrial epi-thelial cells, and Vero cells (a cell line de-rived from green rhesus monkey kidney)have been used successfully. These mono-layers produce embryotrophic factors thatmake it possible to obtain a higher yield ofblastocysts. Some studies suggest that low

1376
Braz J Med Biol Res 39(11) 2006
W.A. Castro-Rendón et al.
concentrations of matrigel in embryo cul-tures enhance blastocyst formation and invitro hatching (11). Matrigel is an extracel-lular matrix whose main structural compo-nents are proteins and growth factors that arealso present in the endometrial extracellularmatrix such as fibronectin, collagen type IV,heparan sulfate proteoglycans, entactin, ni-dogen, transforming growth factor ß, fibro-blast growth factor, and plasminogen tissueactivator, among others. These compoundsconstitute a culture medium similar to theuterine microenvironment that is appropri-ate for embryo growth, viability, and forcytokine secretion. For example, the amountof the highly secreted IL-1α in Quinn’s HTFmedium enriched with matrigel can be cor-related with the embryo’s ability to be im-planted (11). Another explanation for thiseffect could be that these monolayers elimi-nate toxic metabolites generated by the em-bryo and mimic the environment of the ovi-duct (12). It is possible that matrigel in-creases IL-1ß and IL-1 ra and the expressionof membrane receptors such as IL-1R tI, butthis possibility has not yet been tested.
Before embryo implantation the blasto-cyst must escape from the zona pellucida(ZP), a thick extracellular cover composedof three glycoproteins: ZP1, ZP2, ZP3. Thisevent is known as hatching, a dual processwhere the ZP is degraded by enzymes inboth the trophectoderm and the uterus, fa-cilitating blastocyst protrusion from the ZP.In humans, hatching occurs prior to implan-tation between the 5th and 7th day post-ovulation (13) and at 3.5 days post-mating inmice.
In humans and mice, hatching appears tobe regulated by some cytokines, and thebeneficial effect of LIF on embryo develop-ment, in vitro, has been very well docu-mented (Table 1). LIF is a glycoprotein witha remarkable range of biological activities invarious tissues; it is a highly glycosylatedsingle chain polypeptide whose action ismediated through a LIF receptor-glycopro-
tein 130-kDa complex (LIFR-gp130). There-fore, signal transduction and its biologicalactivity require heterodimerization of thesetwo low affinity, components LIFR andgp130, which generate a high-affinity bind-ing site (14). The group of Tsai (15) con-cluded that culture media supplemented withrecombinant human LIF (r-hLIF) enhancesthe rate of two-cell embryos reaching thestage of hatched blastocysts. Thus, the regu-latory effect of LIF is associated with bothearly embryo development and viability:when early-stage embryos are cultured with1,000 IU/mL of r-hLIF the rate of blastocystformation and trophoectodermic prolifera-tion is enhanced (16).
The peri-implantatory endometrialchanges
The endometrium is regulated by cyclicchanges that modify the uterine microenvi-ronment for embryo implantation. In hu-mans the menstrual cycle has been describedin two consecutive phases: proliferative orovarian and secretory or luteal. This view ofthe cycle is supported by the hypothalamic-hypophysial-gonadal axis that gives rise tothe ovarian and endometrial cycles. The be-ginning of the ovarian phase is mediated bythe action of follicle-stimulating hormonewhich acts on several follicles that enter thematuration process during this cycle and ongrowth of dominant follicles. The granulosaand theca cells that surround the oocyte syn-thesize estrogens; these hormones influencethe proliferation of the endometrial cells thatis characteristic of the endometrial prolifera-tive phase (17).
A positive feedback occurs in the hypo-thalamic-hypophysial-gonadal axis whenestrogens are increased in mid-cycle, sud-denly releasing luteinizing hormone and pro-moting ovulation. Granulosa and theca cellsform the corpus luteum, an endocrine struc-ture that produces progesterone and estro-gen, hormones that are necessary to generate

1377
Braz J Med Biol Res 39(11) 2006
Blastocyst-endometrium interactions
the phenotypic changes characteristic of thenext phase of the endometrial cycle, i.e., theluteal or secretory phase. The action of thesehormones is to transform the epithelial andstromal cells into a secretory phenotype,enhancing synthesis of vasoactive com-pounds such as prostaglandins (PGs) andcytokines related to the communication be-tween mother and embryo (17,18).
In the secretory phase, progesterone in-duces the morphological and phenotypicchanges of the endometrial cells that areassociated with the production of cytokinesand inflammatory mediators (18). Changesoccurring during the secretory phase in thestroma are known as decidualization, an in-flammatory process characterized by themorphological transformation of the stromalcells from small spindle-shaped cells to largeplump decidual cells. Decidual cells secreteprolactin, IL-15 and insulin-like growth fac-tor-binding protein 1 (19-21). Spiral arteriesand endometrial glands undergo character-istic changes during this phase: widening ofthe walls of the blood vessels and the musclelayer of the arteries and increase of the se-cretion of inflammatory mediators and cyto-kines by the endometrial gland cells (22).
Regulation of endometrial changes is notexclusively due to ovarian hormones: dur-ing the proliferative phase, for instance,mediators that induce an inflammatory pro-cess in the endometrium such as IL-1 (23),tumor necrosis factor alpha (TNF-α) (24)and PGs are produced (25). This inflamma-tory process is characterized by edema andangiogenesis of the endometrial tissue, leu-kocyte recruitment and production of cyto-kines that could be participating in endome-trium remodeling and blastocyst-endome-trium interaction (23).
IL-1 production during the endometrialproliferative phase induces the release ofvasoactive lipids such as PGs (26), whosesynthesis is regulated by the cyclooxygenase(COX) enzyme, which presents two isoforms:COX-1 and COX-2. COX-1 is constitutively
expressed and located in the gastric mucosa,while COX-2 presents an inducible expres-sion related to inflammatory processes (27).The actions of PGs are not clearly under-stood, since COX-2 null mice showed asignificant decrease of the weight of theuterus when decidualization was induced byintraluminal oil injection (28). The sameobservations were made when celecoxib andaspirin (selective inhibitors of COX-2 andCOX-1, respectively) were used in wild-type mice (29). Based on these results, oneof the mechanisms implicated in deciduali-zation could be the action of PGE2 and pros-tacyclin (PGI2) through production of cyto-kines and growth factors (30).
The enzymatic function of COX is tooxygenate arachidonic acid in cell mem-branes and generate PGH, which is the sub-strate used by tissue-specific isomerases pro-ducing different kinds of PGs (27). Usingimmunohistochemistry, Milne et al. (25)observed the production of PGE synthetaseand PGE2 in the endometrial epithelium,stroma and endothelium during differentphases of the menstrual cycle and also foundthat these compounds decrease during thelate secretory phase. In the murine endome-trium it has been documented that there isexpression and production of both cytosolicand microsomal PGE synthetase isoformsproduced by COX-1 and COX-2, respec-tively. mRNA expression and protein syn-thesis of these enzymes increase on day 5post-ovulation, when murine endometrialdecidualization begins (31,32).
The pre-implantatory endometrium re-quires PGs during decidualization, especiallyfor the vascular changes generated by itsaction as a vasodilator. Furthermore, newfunctions of PGE2 related to cell prolifera-tion in the glandular epithelium have beendescribed: epithelial cells stimulated withPGE2 significantly increased BrdU incorpo-ration (33), indicating that glandular epithe-lial cell division is facilitated by PGE2, whichmay also exert these actions on other cell

1378
Braz J Med Biol Res 39(11) 2006
W.A. Castro-Rendón et al.
populations.Another peculiarity of the early pregnant
uterus is the recruitment of immune cellsfrom peripheral blood, principally NK cells,which differentiate into another subpopula-tion distinct from circulating NK cells, i.e.,uNK cells. The latter grow in size and granu-larity and, conversely, decrease their cyto-toxic ability (34). Linnemeyer and Pollack(35) showed that NK cell differentiation couldbe induced by PGE2 in a concentration-de-pendent manner. Thus, PGE2 could be re-sponsible for inducing these endometrialfunctions but the mechanisms for the induc-tion of differentiation of these cells throughPGE2 have not been elucidated.
Increased production of growth factorssuch as epidermal growth factor (EGF), hep-arin binding-epidermal growth factor (HB-EGF) and vascular endothelial growth factor(VEGF) during both phases of the menstrualcycle has been observed in the endometrium(36,37). VEGF and its receptor have beendetected in endometrial cells and blood ves-sels during the proliferative and secretoryphases with sustained protein expression ofVEGF during both phases, but the number ofreceptors increases during the secretory phase(37). EGF has been found at high concentra-tions in epithelial cells, and its receptor isalso increased in the secretory phase at thelevel of epithelia, stroma and blood vessels(37). HB-EGF production is enhanced in theendometrial stroma during both the prolif-erative and secretory phases in glandularepithelium and endometrial blood vessels(36). In this complex network a balancedcell production of inflammatory mediatorsand cytokines provides an adequate endome-trial environment for interaction betweenthe embryo and endometrium.
Embryo implantation:blastocyst-endometrium interaction
Embryo implantation can be defined asthe process by which the embryo attaches to
the endometrium (38), invades the decidual-ized stroma and reaches the maternal mi-crovasculature (13). Hence, the endometriumrequires structural modifications induced byprogesterone and estrogens, pivotal changesthat facilitate the three consecutive phases ofimplantation: apposition, attachment and in-vasion (38). This morphological and func-tional transformation of the endometriumaround embryo implantation generates a pe-riod known as implantation window (13). Inthis context changes involve glycocalix modi-fications in the epithelial cells of the luminalsurface, growth factor secretion and adhe-sion molecule expression related to blasto-cyst adherence and stromal invasion.
The murine blastocyst moves down theoviduct and falls into the uterine cavity andat that time it orientates its mural trophecto-derm towards the epithelial surface. In hu-mans, the hatched blastocyst opposes itspolar trophectoderm to the surface of theepithelium on the fifth day of gestation, aprocess that occurs on the fourth day inmice. Das et al. (39) proposed that the im-plantation process begins as a kind of dialogbetween the embryo and the endometrium,prior to active penetration into the decidua.They observed a high level of HB-EGF by insitu hybridization in the luminal epitheliumthat surrounds the embryo, at the site ofimplantation, whereas HB-EGF was absentfrom pseudopregnant mice. Another findingrelated to blastocyst growth and hatchingwas obtained with HB-EGF, which enhancesthe cell number per blastocyst by almost40% and leads to a 2-fold increase in thepercentage of hatched embryos. Anothercytokine, LIF, is necessary at this stage, asdemonstrated in LIF-deficient females,whose embryos normally develop to the blas-tocyst stage but fail to implant (40). Implan-tation and normal pregnancy can occur whenthese embryos are transferred from LIF-de-ficient mothers to wild-type pseudopregnantrecipients, or when an exogenous supply ofLIF is provided to the LIF knock-out fe-
1379
Braz J Med Biol Res 39(11) 2006
Blastocyst-endometrium interactions
males (41). Based on this approach, Vogiagiset al. (42) examined the estrous cycle andearly pregnancy in ewes and observed anincrease in immunoreactive LIF at the timeof blastocyst hatching in the luminal andglandular epithelium. In humans, LIF is ex-pressed throughout the cycle with a strikingincrease in LIF mRNA levels in the middleand late secretory phases, the period duringwhich embryo implantation occurs (43). Inthe murine uterus, from days one to eight ofpregnancy, the expression of LIFR and gp130mRNAs was detected by in situ hybridiza-tion, reaching the highest levels on days 5and 4, respectively (44). During early mouseembryogenesis the mRNAs of LIF, LIFRand gp130 are undetectable in 1- or 2-cellembryos, but are present in the blastocyststage (45). This pattern of expression isstrongly suggestive of a paracrine action ofLIF between the embryo and the uterus dur-ing the implantation window.
Other evidence suggests that hormoneactions are mediated by cytokines during theperi-implantation period. For instance, thenidatory surge of estrogens stimulates im-plantation in mice through the action of sec-ondary factors such as LIF (41). Up-regula-tion of uterine LIF expression is coincidentwith high levels of estradiol (E2) at ovulationand implantation. Thus, LIF can replace thenidatory effect of E2 in ovariectomized fe-male mice, inducing implantation and de-cidualization (41). Based on these findings,enhanced uterine expression of LIF could bemediated by a direct action of E2, but werather postulate that such LIF expression canbe induced by HB-EGF, which increases thelevel of LIF by stimulation of E2 (46) (Figure1). In this context the action of LIF can bemediated also by other cytokines: an in vitrostudy with decidual human cells showedhow the addition of 20 µL rIL-1 or E2 to theculture medium increased LIF production ina time-dependent manner (47). In anotherassay, Arici et al. (43) demonstrated that theaddition of 0.01-10 ng EGF ß1 increased the
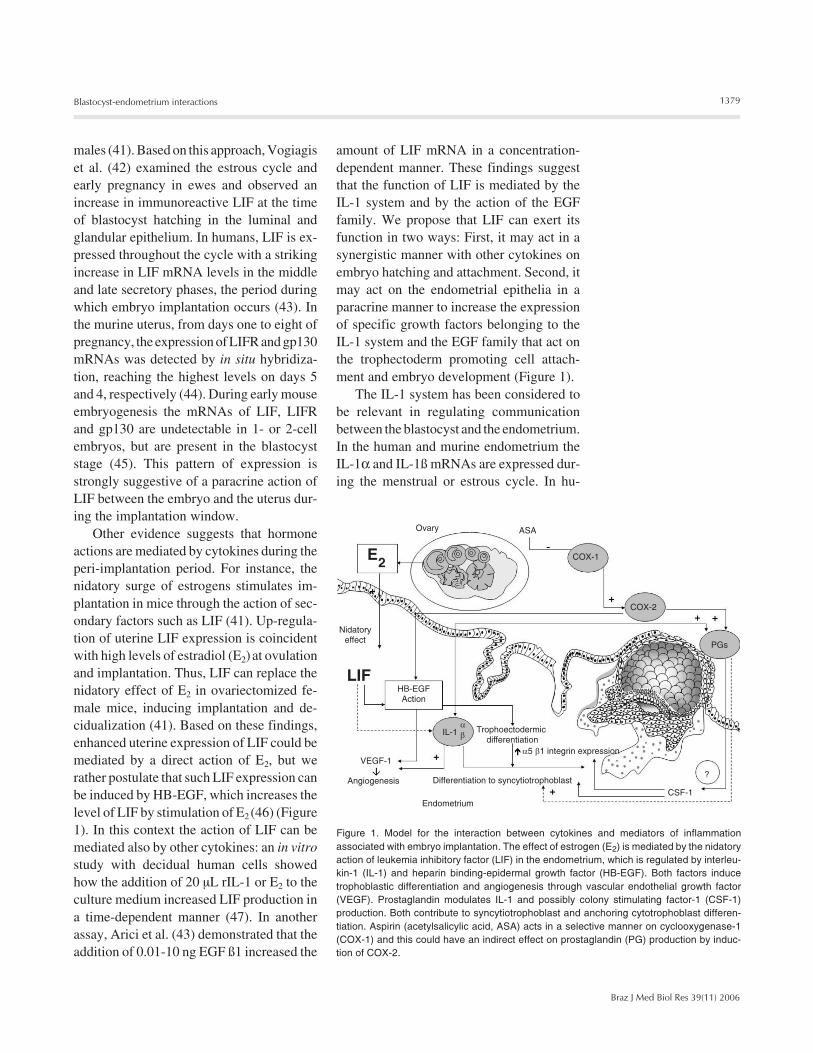
amount of LIF mRNA in a concentration-dependent manner. These findings suggestthat the function of LIF is mediated by theIL-1 system and by the action of the EGFfamily. We propose that LIF can exert itsfunction in two ways: First, it may act in asynergistic manner with other cytokines onembryo hatching and attachment. Second, itmay act on the endometrial epithelia in aparacrine manner to increase the expressionof specific growth factors belonging to theIL-1 system and the EGF family that act onthe trophectoderm promoting cell attach-ment and embryo development (Figure 1).
The IL-1 system has been considered tobe relevant in regulating communicationbetween the blastocyst and the endometrium.In the human and murine endometrium theIL-1α and IL-1ß mRNAs are expressed dur-ing the menstrual or estrous cycle. In hu-
Figure 1. Model for the interaction between cytokines and mediators of inflammationassociated with embryo implantation. The effect of estrogen (E2) is mediated by the nidatoryaction of leukemia inhibitory factor (LIF) in the endometrium, which is regulated by interleu-kin-1 (IL-1) and heparin binding-epidermal growth factor (HB-EGF). Both factors inducetrophoblastic differentiation and angiogenesis through vascular endothelial growth factor(VEGF). Prostaglandin modulates IL-1 and possibly colony stimulating factor-1 (CSF-1)production. Both contribute to syncytiotrophoblast and anchoring cytotrophoblast differen-tiation. Aspirin (acetylsalicylic acid, ASA) acts in a selective manner on cyclooxygenase-1(COX-1) and this could have an indirect effect on prostaglandin (PG) production by induc-tion of COX-2.

1380
Braz J Med Biol Res 39(11) 2006
W.A. Castro-Rendón et al.
mans, IL-1α, IL-1ß, IL-1R tI, and IL-1 ra arealso expressed. This secretion of IL-1α, IL-1ß and IL-1 ra changes in the endometriumaccording to the levels of steroid hormones,an event that is clearly visible when IL-1ßreaches a peak around the time of implanta-tion. The IL-1R tI mRNA is expressed atincreased concentrations in human endome-trial epithelium during the luteal phase (48).In mice it was demonstrated that IL-1 rainterferes with embryonic attachment; IL-1ra injected during the peri-implantation pe-riod induces failure of implantation (49).
The pre-ovulatory increase of 17-ß estra-diol stimulates both proliferation and differ-entiation of endometrial epithelial cells (17).In particular, 17-ß estradiol promotes anincrease of apical protein density and a de-crease of electronegative charges at the siteof implantation. In some species, glycocalixchanges are mediated by mucin 1 (MUC-1)expression on the epithelial surface in theendometrium, which overlaps putative bind-ing sites of trophectoderm cells, mainly gly-coconjugates and heparan sulfate-associatedproteoglycans (50). For instance, MUC-1, acomponent of the apical glycocalix, is alarge glycoprotein (>200 kDa) with an ex-tracellular domain rich in proline, serine andthreonine residues. This protein is abun-dantly present on the apical glandular epi-thelial surface, and in the midsecretory im-plantation phase; it is also secreted into thegland lumen and in the uterine cavity. Over-expression of MUC-1 is able to induce stericinhibition of the adhesive properties of cul-tured cells and it is known that this mightencourage metastatic spread. In mice andother species there is a distinctive reductionin MUC-1 protein expression during im-plantation, but not in humans (50), in whomdown-regulation of this molecule may per-mit the expression of proteins such as hepa-ran sulfate binding proteins that mediateembryo attachment.
Blastocyst attachment in humans occursin the midsecretory phase. Aplin et al. (51)
characterized the luminal epithelium at thetime of implantation and observed a con-tinuous staining pattern of MUC-1 betweendays 20 and 24 of the normal cycle (pre-dicted period of the receptive phase). Thismeans that the antiadhesive properties ofMUC-1 detected in other species may not bevalid in humans. These antiadhesive proper-ties of MUC-1 have been shown to be due tothe large extracellular domain, but it is clearfrom Aplin’s work that immunoreactivityagainst sulfated glycosylation in apical epi-thelia is maintained during the proliferativephase and is increased in the early secretoryphase. Thus, on day 20, apical staining witha 5D4 mAb, that recognizes keratan sulfatechains, was reduced in the luminal epithe-lium. Based on this observation, we proposethat both the glycosylation with sulfatedchains in the core protein of MUC-1 and theelectronegativity of this molecule are alteredduring uterine receptivity. The antiadherence/adherence balance is modulated in the endo-metrium by the attached embryo, which couldinduce indirect actions through enzymes suchas metalloprotease and heparinase. In thisway, the group of Simon (52) showed byimmunofluorescence that MUC-1 was ab-sent in endometrial epithelial cells subjacentand adjacent to the attached embryo, sug-gesting an active regulation by the embryoin the receptivity of the human endometrium(52).
On the other hand, synthesis of heparansulfate in trophoectodermic cells increases4- to 5-fold at the murine peri-implantationstage, which indicates that the embryo ex-presses proteoglycans necessary for adhe-sive interactions with the endometrium, suchas syndecan and perlecan (53). The latter, aswell as other proteoglycans (fibronectin andlaminin) are expressed on the external sur-face of the trophectoderm and could interactwith the extracellular matrix (54) in the de-cidualized endometrium. These compoundscould also bind to members of the EGFfamily (amphiregulin, HB-EGF, among oth-
1381
Braz J Med Biol Res 39(11) 2006
Blastocyst-endometrium interactions
ers, improving attachment and growth of theimplantatory blastocyst.
Effect of growth factors ontrophoblast differentiation
The decidua is thought to be a source ofimportant growth factors and cytokines forplacental development. Growth factors regu-late PG synthesis in the endometrium, pro-moting the invasion, proliferation and changesrelated to endometrial vascular permeabil-ity. Enhanced PG production by the actionof EGF has been shown in rat endometrialstromal cells. EGF acts through the phos-phorylation of membrane proteins as a mito-gen promoting DNA and RNA synthesis. Aspregnancy progresses, the increased produc-tion of EGF enhances trophoblast differen-tiation (55).
Granulocyte-macrophage CSF (GM-CSF) is produced by murine decidual cellsin response to invasive trophoblast. Wegmann(56) documented the expression of GM-CSF,CSF-1, TNF-α, IL-1, among other factors,in the murine placenta using a panel of cDNAprobes. This pattern of cytokine expressionmay involve growth regulatory signals forthe placenta (56). Garcia-Lloret et al. (57)demonstrated that both CSF-1 and GM-CSFstimulate the differentiation of the cytotro-phoblast to the syncytiotrophoblast. Thismorphological differentiation was indirectlymeasured by the increased production ofhuman placental lactogen (hPL) and humanchorionic gonadotropin (hCG). Additionally,they found that GM-CSF and CSF-1 produc-tion was controlled by IL-1 and TNF-α infibroblasts derived from the villous stromaof the placenta. These cytokines could playan important role in the development andfunction of the human placenta (57).
Our group has studied the effect of aspi-rin on the production of GM-CSF and IL-3by peripheral blood mononuclear cells (PBMC)and the effect of these cytokines on tropho-blast differentiation. Briefly, PBMC were
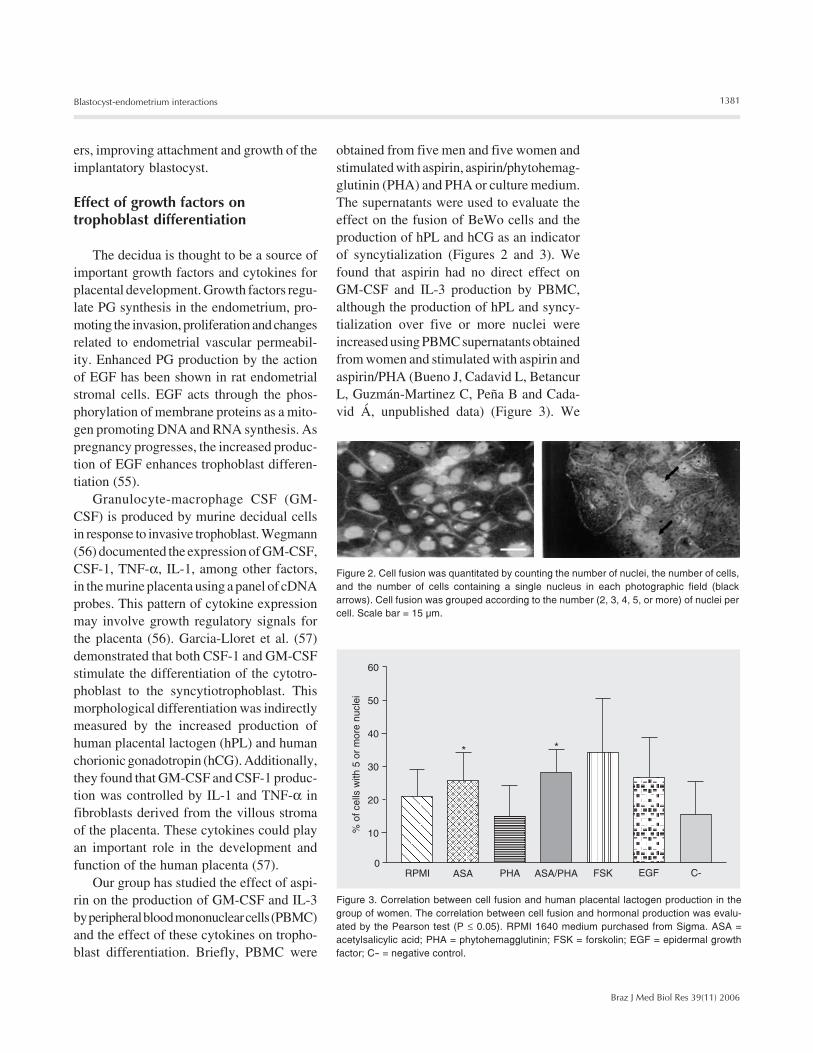
obtained from five men and five women andstimulated with aspirin, aspirin/phytohemag-glutinin (PHA) and PHA or culture medium.The supernatants were used to evaluate theeffect on the fusion of BeWo cells and theproduction of hPL and hCG as an indicatorof syncytialization (Figures 2 and 3). Wefound that aspirin had no direct effect onGM-CSF and IL-3 production by PBMC,although the production of hPL and syncy-tialization over five or more nuclei wereincreased using PBMC supernatants obtainedfrom women and stimulated with aspirin andaspirin/PHA (Bueno J, Cadavid L, BetancurL, Guzmán-Martinez C, Peña B and Cada-vid Á, unpublished data) (Figure 3). We
Figure 2. Cell fusion was quantitated by counting the number of nuclei, the number of cells,and the number of cells containing a single nucleus in each photographic field (blackarrows). Cell fusion was grouped according to the number (2, 3, 4, 5, or more) of nuclei percell. Scale bar = 15 µm.
Figure 3. Correlation between cell fusion and human placental lactogen production in thegroup of women. The correlation between cell fusion and hormonal production was evalu-ated by the Pearson test (P ≤ 0.05). RPMI 1640 medium purchased from Sigma. ASA =acetylsalicylic acid; PHA = phytohemagglutinin; FSK = forskolin; EGF = epidermal growthfactor; C- = negative control.

1382
Braz J Med Biol Res 39(11) 2006
W.A. Castro-Rendón et al.
conclude that the trophoblast differentiationinduced by the effect of aspirin cannot becorrelated with GM-CSF or IL-3 produc-tion. Perhaps other cytokines could be in-duced by aspirin to improve syncytializa-tion. Moreover, since it is possible that aspi-rin can induce some effect on growth factorsecretion by the trophoblast per se, we haveperformed some experiments in which aspi-rin directly induced an increase in tropho-blast syncitialization, maybe through the au-tocrine action of growth factors produced bythese cells.
Therapeutical approaches
A recently developed r-hLIF (or emfiler-min) will be tested in human clinical trials.The experimental evidence in animals hasdemonstrated the physiological role of LIFin implantation, but the implications of theuse of r-hLIF in human reproductive medi-cine are not clear. LIF has a wide spectrumof action and therefore has been used formany diseases; e.g., in a phase I study in-cluding patients with advanced lung cancer,a biological effect of r-hLIF was demon-strated by the increase of blood progenitorcells, C-reactive protein levels, and hemopoi-etic recovery after chemotherapy (58). Onthe basis of reports from animal and humanstudies, we agree with the statement madeby SERONO in its website (59) that LIFcould be used as a safe therapeutical alterna-tive and as a potent inducer of human em-bryonic implantation.
For the adaptive modifications of theendometrial environment, steroid hormonesare a key factor, particularly E2 that exerts itsactions on the production of LIF in the endo-metrium. On this basis, we propose that thenidatory surge of LIF during the implanta-tion window could induce a change of theelectronegative charges on the epithelial sur-face and the remodeling of the glycocalix tohome the embryo. It is possible that glycosy-lation of MUC-1 changes these electronega-
tive charges produced by keratan sulfatechains in the glycoprotein structure and fi-nally enhances the exposure of heparan sul-fate-binding proteins in the epithelium. LIFcould play an important role in these modifi-cations, improving the attachment of theembryo to the endometrium and ensuringtrophoectodermic differentiation towards thefibronectin-binding phenotype influenced byHB-EGF.
We believe that the differentiation of NKcells and other endometrial cells such asstromal cells may be induced by PGE2. Atthe same time this could amplify the produc-tion of other cytokines that participate indecidualization and embryo implantation.Furthermore, PG production could also acton trophoblast proliferation and differentia-tion. The therapeutic use of aspirin prior toimplantation could be useful for successfulpregnancy because it has been demonstratedthat aspirin can inhibit the action of COX-1in platelets and up-regulate COX-2 produc-tion from other sources. This stimulus po-tentiates PGE2 production that could finallyenhance trophoblast differentiation and in-crease the vasodilatory action necessary foradequate blood perfusion of the embryo.
Although therapeutic strategies based oncytokine targets have been used in manymodels and in diseases such as cancer, theresults reported are not completely conclu-sive in clinical trials, maybe due to the com-plexity of the cytokine network implicatedin each disease. Alternative pathways couldbe activated when a cytokine is blocked orwhen it is used as an inducer of a specificimmune response. New therapeutical ap-proaches must focus on drugs able to inducethe production of several cytokines, in orderto maintain an adequate profile of cytokinesthat support the desired response. Aspirincould be one such compound: it is an ancientdrug with unexplored actions on cytokineproduction, for instance on IL-3 productionby mononuclear cells. We did not establishwhat kind of cytokine modulate the syncy-

1383
Braz J Med Biol Res 39(11) 2006
Blastocyst-endometrium interactions
tialization of the BeWo cells throughoutacetylsalicylic acid or what kind of mediators,e.g., PGE2 or leukotrienes, were indirectlyimplicated in this immune modulation but it ispossible that more than one cytokine wasinvolved in these effects. The probable induc-tion by aspirin of cytokine production by tro-phoblastic cells cannot be ruled out in thiscontext: the trophoblast has an important rolein the maternal-fetal dialogue and aspirin couldmodulate bidirectionally this interaction.
In other animal models such as the pig,the immunomodulator effect of PGE2 hasbeen shown on growth factor and cytokineproduction by lymphocytes during gestation(60). In humans, this same action on thelymphocytes and other cells such as NKcells could be stimulated by PGE2 and couldinduce the production of cytokines such asthose of the CSF family. Thus, implantationand trophoblast invasion could be mediatedby CSF-1, GM-CSF and other cytokinesproduced by cells localized in the decidua.
Finally, IL-1ß induces COX-2 synthesis(26) and LIF production (47) by human en-dometrial stromal cells and in decidual cellsin vitro. These results strongly suggest thatPGs may control blastocyst implantation
through a paracrine effect of the IL-1. On theother hand, they could suggest a synergisticeffect of both IL-1 and LIF on blastocystimplantation (Figure 1).
Detailed information about the cellularand molecular mechanisms associated withthe action of cytokines on stromal cells andon the blastocyst during implantation willhave a significant impact on the understand-ing of the fundamental causes of implanta-tion failure. Moreover, the participation ofthe trophoblast in the paracrine dialoguebetween mother and fetus must be under-stood in more detail due to its pivotal actionon embryo and fetus development.
Acknowledgments
The authors are grateful for the help andencouragement of Professors Angela Cadavid(coordinator of the Reproduction Group at theUniversity of Antioquia), Juan GuillermoMaldonado (coordinator of the CENTAUROGroup at the University of Antioquia), Anne-Lise Haenni (from the Institute Jacques Monod- France), and Jorge Ossa (from the Universityof Antioquia).
References
1. Kaye PL. Preimplantation growth factor physiology. Rev Reprod1997; 2: 121-127.
2. Kimber SJ. Cell biology of implantation and plancentation. Introduc-tion. Semin Cell Dev Biol 2000; 11: 61-65.
3. Shaw TJ, Murphy CR. Leucocyte involvement in lectin-induced de-ciduomata formation. Cell Biol Int 1995; 19: 577-584.
4. Pollheimer J, Knofler M. Signalling pathways regulating the invasivedifferentiation of human trophoblasts: a review. Placenta 2005; 26(Suppl A): S21-S30.
5. Cooke S, Quinn P, Kime L, Ayres C, Tyler JP, Driscoll GL. Improve-ment in early human embryo development using new formulationsequential stage-specific culture media. Fertil Steril 2002; 78: 1254-1260.
6. Watson AJ. The cell biology of blastocyst development. Mol ReprodDev 1992; 33: 492-504.
7. Krussel JS, Simon C, Rubio MC, Pape AR, Wen Y, Huang HY, et al.Expression of interleukin-1 system mRNA in single blastomeresfrom human preimplantation embryos. Hum Reprod 1998; 13: 2206-2211.
8. Paula-Lopes FF, de Moraes AA, Edwards JL, Justice JE, HansenPJ. Regulation of preimplantation development of bovine embryosby interleukin-1beta. Biol Reprod 1998; 59: 1406-1412.
9. Sheth KV, Roca GL, al-Sedairy ST, Parhar RS, Hamilton CJ, al-Abdul JF. Prediction of successful embryo implantation by measur-ing interleukin-1-alpha and immunosuppressive factor(s) in preim-plantation embryo culture fluid. Fertil Steril 1991; 55: 952-957.
10. Kruessel JS, Huang HY, Wen Y, Kloodt AR, Bielfeld P, Polan ML.Different pattern of interleukin-1 beta- (IL-1 beta), interleukin-1 re-ceptor antagonist- (IL-1ra) and interleukin-1 receptor type I- (IL-1RtI) mRNA-expression in single preimplantation mouse embryos atvarious developmental stages. J Reprod Immunol 1997; 34: 103-120.
11. Lazzaroni L, Fusi FM, Doldi N, Ferrari A. The use of Matrigel at lowconcentration enhances in vitro blastocyst formation and hatching ina mouse embryo model. Fertil Steril 1999; 71: 1133-1137.
12. Ouhibi N, Hamidi J, Guillaud J, Menezo Y. Co-culture of 1-cellmouse embryos on different cell supports. Hum Reprod 1990; 5:737-743.

1384
Braz J Med Biol Res 39(11) 2006
W.A. Castro-Rendón et al.
13. Norwitz ER, Schust DJ, Fisher SJ. Implantation and the survival ofearly pregnancy. N Engl J Med 2001; 345: 1400-1408.
14. Senturk LM, Arici A. Leukemia inhibitory factor in human reproduc-tion. Am J Reprod Immunol 1998; 39: 144-151.
15. Tsai HD, Chang CC, Hsieh YY, Lo HY, Hsu LW, Chang SC. Recom-binant human leukemia inhibitory factor enhances the developmentof preimplantation mouse embryo in vitro. Fertil Steril 1999; 71: 722-725.
16. Lavranos TC, Rathjen PD, Seamark RF. Trophic effects of myeloidleukaemia inhibitory factor (LIF) on mouse embryos. J Reprod Fertil1995; 105: 331-338.
17. Chabbert-Buffet N, Bouchard P. The normal human menstrual cycle.Rev Endocr Metab Disord 2002; 3: 173-183.
18. Rider V. Progesterone and the control of uterine cell proliferationand differentiation. Front Biosci 2002; 7: d1545-d1555.
19. Okada S, Okada H, Sanezumi M, Nakajima T, Yasuda K, Kanzaki H.Expression of interleukin-15 in human endometrium and decidua.Mol Hum Reprod 2000; 6: 75-80.
20. Dimitriadis E, Salamonsen LA, Robb L. Expression of interleukin-11during the human menstrual cycle: coincidence with stromal celldecidualization and relationship to leukaemia inhibitory factor andprolactin. Mol Hum Reprod 2000; 6: 907-914.
21. King A. Uterine leukocytes and decidualization. Hum Reprod Up-date 2000; 6: 28-36.
22. Fazleabas AT, Strakova Z. Endometrial function: cell specificchanges in the uterine environment. Mol Cell Endocrinol 2002; 186:143-147.
23. McMaster MT, Newton RC, Dey SK, Andrews GK. Activation anddistribution of inflammatory cells in the mouse uterus during thepreimplantation period. J Immunol 1992; 148: 1699-1705.
24. von Wolff M, Classen-Linke I, Heid D, Krusche CA, Beier-Hellwig K,Karl C, et al. Tumour necrosis factor-alpha (TNF-alpha) in humanendometrium and uterine secretion: an evaluation by immunohisto-chemistry, ELISA and semiquantitative RT-PCR. Mol Hum Reprod1999; 5: 146-152.
25. Milne SA, Perchick GB, Boddy SC, Jabbour HN. Expression, local-ization, and signaling of PGE(2) and EP2/EP4 receptors in humannonpregnant endometrium across the menstrual cycle. J Clin Endo-crinol Metab 2001; 86: 4453-4459.
26. Huang JC, Liu DY, Yadollahi S, Wu KK, Dawood MY. Interleukin-1beta induces cyclooxygenase-2 gene expression in cultured en-dometrial stromal cells. J Clin Endocrinol Metab 1998; 83: 538-541.
27. Majerus PW. Prostaglandins: critical roles in pregnancy and coloncancer. Curr Biol 1998; 8: R87-R89.
28. Lim H, Paria BC, Das SK, Dinchuk JE, Langenbach R, Trzaskos JM,et al. Multiple female reproductive failures in cyclooxygenase 2-deficient mice. Cell 1997; 91: 197-208.
29. Reese J, Zhao X, Ma WG, Brown N, Maziasz TJ, Dey SK. Compara-tive analysis of pharmacologic and/or genetic disruption of cyclo-oxygenase-1 and cyclooxygenase-2 function in female reproductionin mice. Endocrinology 2001; 142: 3198-3206.
30. Kimber SJ. Leukaemia inhibitory factor in implantation and uterinebiology. Reproduction 2005; 130: 131-145.
31. Ni H, Sun T, Ding NZ, Ma XH, Yang ZM. Differential expression ofmicrosomal prostaglandin E synthase at implantation sites and indecidual cells of mouse uterus. Biol Reprod 2002; 67: 351-358.
32. Ni H, Sun T, Ma XH, Yang ZM. Expression and regulation of cytoso-lic prostaglandin E synthase in mouse uterus during the peri-implan-tation period. Biol Reprod 2003; 68: 744-750.
33. Jabbour HN, Boddy SC. Prostaglandin E2 induces proliferation ofglandular epithelial cells of the human endometrium via extracellular
regulated kinase 1/2-mediated pathway. J Clin Endocrinol Metab2003; 88: 4481-4487.
34. Pollack SB, Linnemeyer PA. Natural killer cells in the nonpregnantmurine uterus. Nat Immun 1996; 15: 34-40.
35. Linnemeyer PA, Pollack SB. Prostaglandin E2-induced changes inthe phenotype, morphology, and lytic activity of IL-2-activated natu-ral killer cells. J Immunol 1993; 150: 3747-3754.
36. Leach RE, Khalifa R, Ramirez ND, Das SK, Wang J, Dey SK, et al.Multiple roles for heparin-binding epidermal growth factor-like growthfactor are suggested by its cell-specific expression during the hu-man endometrial cycle and early placentation. J Clin EndocrinolMetab 1999; 84: 3355-3363.
37. Moller B, Rasmussen C, Lindblom B, Olovsson M. Expression of theangiogenic growth factors VEGF, FGF-2, EGF and their receptors innormal human endometrium during the menstrual cycle. Mol HumReprod 2001; 7: 65-72.
38. Cavagna M, Mantese JC. Biomarkers of endometrial receptivity - areview. Placenta 2003; 24 (Suppl B): S39-S47.
39. Das SK, Wang XN, Paria BC, Damm D, Abraham JA, Klagsbrun M,et al. Heparin-binding EGF-like growth factor gene is induced in themouse uterus temporally by the blastocyst solely at the site of itsapposition: a possible ligand for interaction with blastocyst EGF-receptor in implantation. Development 1994; 120: 1071-1083.
40. Stewart CL, Kaspar P, Brunet LJ, Bhatt H, Gadi I, Kontgen F, et al.Blastocyst implantation depends on maternal expression of leu-kaemia inhibitory factor. Nature 1992; 359: 76-79.
41. Chen JR, Cheng JG, Shatzer T, Sewell L, Hernandez L, Stewart CL.Leukemia inhibitory factor can substitute for nidatory estrogen andis essential to inducing a receptive uterus for implantation but is notessential for subsequent embryogenesis. Endocrinology 2000; 141:4365-4372.
42. Vogiagis D, Fry RC, Sandeman RM, Salamonsen LA. Leukaemiainhibitory factor in endometrium during the oestrous cycle, earlypregnancy and in ovariectomized steroid-treated ewes. J ReprodFertil 1997; 109: 279-288.
43. Arici A, Engin O, Attar E, Olive DL. Modulation of leukemia inhibitoryfactor gene expression and protein biosynthesis in human endome-trium. J Clin Endocrinol Metab 1995; 80: 1908-1915.
44. Ni H, Ding NZ, Harper MJ, Yang ZM. Expression of leukemia inhibi-tory factor receptor and gp130 in mouse uterus during early preg-nancy. Mol Reprod Dev 2002; 63: 143-150.
45. Nichols J, Davidson D, Taga T, Yoshida K, Chambers I, Smith A.Complementary tissue-specific expression of LIF and LIF-receptormRNAs in early mouse embryogenesis. Mech Dev 1996; 57: 123-131.
46. Wang XN, Das SK, Damm D, Klagsbrun M, Abraham JA, Dey SK.Differential regulation of heparin-binding epidermal growth factor-like growth factor in the adult ovariectomized mouse uterus byprogesterone and estrogen. Endocrinology 1994; 135: 1264-1271.
47. Sawai K, Matsuzaki N, Okada T, Shimoya K, Koyama M, Azuma C,et al. Human decidual cell biosynthesis of leukemia inhibitory factor:regulation by decidual cytokines and steroid hormones. Biol Reprod1997; 56: 1274-1280.
48. Simon C, Mercader A, Frances A, Gimeno MJ, Polan ML, Remohi J,et al. Hormonal regulation of serum and endometrial IL-1 alpha, IL-1beta and IL-1ra: IL-1 endometrial microenvironment of the humanembryo at the apposition phase under physiological and supra-physiological steroid level conditions. J Reprod Immunol 1996; 31:165-184.
49. Simon C, Frances A, Piquette GN, el Danasouri I, Zurawski G, DangW, et al. Embryonic implantation in mice is blocked by interleukin-1

1385
Braz J Med Biol Res 39(11) 2006
Blastocyst-endometrium interactions
receptor antagonist. Endocrinology 1994; 134: 521-528.50. Braga VM, Gendler SJ. Modulation of Muc-1 mucin expression in
the mouse uterus during the estrus cycle, early pregnancy andplacentation. J Cell Sci 1993; 105 ( Pt 2): 397-405.
51. Aplin JD, Meseguer M, Simon C, Ortiz ME, Croxatto H, Jones CJ.MUC1, glycans and the cell-surface barrier to embryo implantation.Biochem Soc Trans 2001; 29: 153-156.
52. Meseguer M, Aplin JD, Caballero-Campo P, O’Connor JE, MartinJC, Remohi J, et al. Human endometrial mucin MUC1 is up-regu-lated by progesterone and down-regulated in vitro by the humanblastocyst. Biol Reprod 2001; 64: 590-601.
53. Carson DD, Tang JP, Julian J. Heparan sulfate proteoglycan(perlecan) expression by mouse embryos during acquisition of at-tachment competence. Dev Biol 1993; 155: 97-106.
54. Smith SE, French MM, Julian J, Paria BC, Dey SK, Carson DD.Expression of heparan sulfate proteoglycan (perlecan) in the mouseblastocyst is regulated during normal and delayed implantation. DevBiol 1997; 184: 38-47.
55. Kliman HJ, Feinberg RF, Haimowitz JE. Human trophoblast-en-
dometrial interactions in an in vitro suspension culture system.Placenta 1990; 11: 349-367.
56. Wegmann T. The role of cytokine cross-talk in preventing abortion.Res Immunol 1990; 141: 185-188.
57. Garcia-Lloret MI, Morrish DW, Wegmann TG, Honore L, Turner AR,Guilbert LJ. Demonstration of functional cytokine-placental interac-tions: CSF-1 and GM-CSF stimulate human cytotrophoblast differ-entiation and peptide hormone secretion. Exp Cell Res 1994; 214:46-54.
58. Gunawardana DH, Basser RL, Davis ID, Cebon J, Mitchell P, Un-derhill C, et al. A phase I study of recombinant human leukemiainhibitory factor in patients with advanced cancer. Clin Cancer Res2003; 9: 2056-2065.
59. SERONO site. http://www.seronousa.com/news/archives/June3003.jsp. Accessed March 16, 2006.
60. Li TK, Fox BS. Effect of prostaglandin E2 (PGE2) on IL-3/granulo-cyte-macrophage colony-stimulating factor production by T helpercells. Mode of stimulation and presence of costimulation can deter-mine response to PGE2. J Immunol 1993; 150: 1680-1690.
Related Documents











