Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009) doi: 10.1111/j.1463-6395.2008.00344.x © 2009 The Authors 344 Journal compilation © 2009 The Royal Swedish Academy of Sciences Abstract Lebedev, O.A., Mark-Kurik, E., KaratajutE-Talimaa, V.N., Lukßeviçs, E. and Ivanov, A. 2009. Bite marks as evidence of predation in early vertebrates. — Acta Zoologica (Stockholm), 90 (Suppl. 1): 344–356 Study of lifetime bite traces on agnathans and fish (or gnathostomes) from Ukraine, Estonia, Latvia and north-western and central European Russia reveals evidence of predator–prey relationships in communities of Devonian age. Numerous bite traces on skeletal parts of agnathan pteraspidiforms and psammosteiforms, placoderm arthrodires and antiarchs and sarcopterygian porolepiforms and osteolepiforms are described. Evidence of healing shows that prey organisms responded to predation by reconstruction of damaged skeletal elements. Ichthyofaunistic analysis is used to establish possible predators. The most probable predators in the Middle and Late Devonian communities are sarcopterygian porolepiforms and osteolepiforms. Predatory tetrapods become evident during the Famennian. Global analysis of aquatic predators during the Silurian–Devonian interval shows a gradual increase in species numbers with time. During the Late Silurian, only ischnacantid acanthodians, early osteichthyans and sarcopterygians are known to belong to this trophic group. By the end of the Devonian this list is complemented by chondrichthyans, arthrodires, porolepiform, osteolepiform, struniiform and rhizodontiform sarcopterygians and tetrapods. Only Devonian agnathans show no predatory groups. In sarcopterygians, predatory dentitions, which developed according to more or less the same pattern, show little change during the Devonian. Oleg A. Lebedev, Palaeontological Institute of the Russian Academy of Sciences, Profsoyuznaya St. 123, Moscow, 117997, Russia. E-mail: [email protected] Blackwell Publishing Ltd Bite marks as evidence of predation in early vertebrates Oleg A. Lebedev, 1 Elga Mark-Kurik, 2 Valentina N. KaratajutE-Talimaa, 3 Ervins Lukßeviçs 4 and Alexander Ivanov 5 1 Palaeontological Institute of the Russian Academy of Sciences, Profsoyuznaya St. 123, Moscow, 117997, Russia; 2 Institute of Geology, Tallinn University of Technology, Ehitajate tee 5, Tallinn, 19086, Estonia; 3 Institute of Geology and Geography, T. Sevçenkos 13, Vilnius LT-03223, Lithuania; 4 Department of Geology, University of Latvia, Rainis Boulevard 19, Riga LV-1586, Latvia; 5 Department of Palaeontology, St Petersburg University, 16 Liniya 29, St Petersburg 199178, Russia Keywords: Vertebrates Devonian predation bite marks Accepted for publication: 8 June 2008 Introduction Study of predator–prey relationships is one of the most important topics in both neo- and palaeoecology. The materials used for this research in palaeoecology are stomach content, coprolites and bite marks. All of these information sources have their benefits and disadvantages. Evidence from stomach content is often questioned because it is not always clear whether skeletal elements found in the abdominal region have been swallowed or whether this is just a superposition of parts of two organisms within the same bedding plane during burial. Coprolites are always separated from the organism that produced them and we can only guess to which animal they are related. Bite marks only seldom bear parts of the skeleton of the predator; if this happens, it is the most important and unequivocal evidence of predator–prey relationships in the community. Fossil vertebrate materials offer numerous traces of lifetime damage. However, investigation of bite traces in the Devonian agnathans and fish is uncommon (Obruchev and

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Acta Zoologica
(Stockholm)
90
(Suppl. 1): 344–356 (May 2009) doi: 10.1111/j.1463-6395.2008.00344.x
© 2009 The Authors
344
Journal compilation © 2009 The Royal Swedish Academy of Sciences
Abstract
Lebedev, O.A., Mark-Kurik, E., Karataj
u
t
E
-Talimaa, V.N., Luk
ß
evi
ç
s, E. andIvanov, A. 2009. Bite marks as evidence of predation in early vertebrates. —
Acta Zoologica
(Stockholm),
90
(Suppl. 1): 344–356
Study of lifetime bite traces on agnathans and fish (or gnathostomes) fromUkraine, Estonia, Latvia and north-western and central European Russiareveals evidence of predator–prey relationships in communities of Devonianage. Numerous bite traces on skeletal parts of agnathan pteraspidiforms andpsammosteiforms, placoderm arthrodires and antiarchs and sarcopterygianporolepiforms and osteolepiforms are described. Evidence of healing showsthat prey organisms responded to predation by reconstruction of damagedskeletal elements. Ichthyofaunistic analysis is used to establish possiblepredators. The most probable predators in the Middle and Late Devoniancommunities are sarcopterygian porolepiforms and osteolepiforms. Predatorytetrapods become evident during the Famennian. Global analysis of aquaticpredators during the Silurian–Devonian interval shows a gradual increase inspecies numbers with time. During the Late Silurian, only ischnacantidacanthodians, early osteichthyans and sarcopterygians are known to belong tothis trophic group. By the end of the Devonian this list is complemented bychondrichthyans, arthrodires, porolepiform, osteolepiform, struniiform andrhizodontiform sarcopterygians and tetrapods. Only Devonian agnathansshow no predatory groups. In sarcopterygians, predatory dentitions, whichdeveloped according to more or less the same pattern, show little changeduring the Devonian.
Oleg A. Lebedev, Palaeontological Institute of the Russian Academy of Sciences, Profsoyuznaya St. 123, Moscow, 117997, Russia. E-mail: [email protected]
Blackwell Publishing Ltd
Bite marks as evidence of predation in early vertebrates
Oleg A. Lebedev,
1
Elga Mark-Kurik,
2
Valentina N. Karataj
u
t
E
-Talimaa,
3
Ervins Luk
ß
evi
ç
s
4
and Alexander Ivanov
5
1
Palaeontological Institute of the Russian Academy of Sciences, Profsoyuznaya St. 123, Moscow, 117997, Russia;
2
Institute of Geology, Tallinn University of Technology, Ehitajate tee 5, Tallinn, 19086, Estonia;
3
Institute of Geology and Geography, T.
S
ev
ç
enkos 13, Vilnius LT-03223, Lithuania;
4
Department of Geology, University of Latvia, Rainis Boulevard 19, Riga LV-1586, Latvia;
5
Department of Palaeontology, St Petersburg University, 16 Liniya 29, St Petersburg 199178, Russia
Keywords:
Vertebrates Devonian predation bite marks
Accepted for publication:
8 June 2008
Introduction
Study of predator–prey relationships is one of the mostimportant topics in both neo- and palaeoecology. Thematerials used for this research in palaeoecology are stomachcontent, coprolites and bite marks. All of these informationsources have their benefits and disadvantages. Evidencefrom stomach content is often questioned because it is notalways clear whether skeletal elements found in theabdominal region have been swallowed or whether this is just
a superposition of parts of two organisms within the samebedding plane during burial. Coprolites are always separatedfrom the organism that produced them and we can only guessto which animal they are related. Bite marks only seldom bearparts of the skeleton of the predator; if this happens, it is themost important and unequivocal evidence of predator–preyrelationships in the community.
Fossil vertebrate materials offer numerous traces oflifetime damage. However, investigation of bite traces in theDevonian agnathans and fish is uncommon (Obruchev and

Acta Zoologica
(Stockholm)
90
(Suppl. 1): 344–356 (May 2009)
Lebedev
et al.
•
Predation in early vertebrates
© 2009 The AuthorsJournal compilation © 2009 The Royal Swedish Academy of Sciences
345
Mark-Kurik 1965; Mark-Kurik 1966; Lebedev 1993; Capasso
et al
. 1996; Karataj
u
t
E
-Talimaa
et al
. 2007). The psammosteidmonograph by Obruchev and Mark-Kurik (1965) includesfigures of nine specimens, mostly from the Estonian collec-tion, with various bite marks or scratches. These specimenshave been described more fully in a paper by Mark-Kurik(1966). Halstead Tarlo (1965) also noticed bite marks intwo branchial plates of the psammosteid
Psammolepisvenyukovi
from Estonia. Analysis of bite marks and subsequentpathological changes makes it possible to study the rela-tionships with predatory members of the community andaids studies of skeletal tissue remodelling. Every discoverytherefore is of great importance.
Identification and description of injury traces can be prob-lematic because other pathological conditions or problemswith preservation can lead to inconclusive results. For thiswork, the following criteria have been used to determinewhether a structure should be classified as an injury trace:(1) the location of the damage (i.e. on a bony element
accessible to a predator, that is, on the dermal bone of ahead, shoulder girdle or a non-overlapped scale surface);
(2) the presence of regenerated tissue (confirmation of thelifetime nature of the lesion);
(3) the presence of damage traces or their repair on the
internal
surface of the bony element;(4) the specific outline of the injury trace.
Only damage that satisfies all these criteria can be con-sidered reliable evidence of predation. An indisputable case,for example, might be of multiple puncta located in a rowmatching a known predator’s dentition pattern.
Materials and Methods
The collections of the Institute of Geology, Tallinn Universityof Technology, include about 30 specimens of MiddleDevonian fish including agnathans with bite marks and/orscratches (reference numbers: 98, 100, 116, 117 and 257).The specimens come from the Aruküla, Burtnieki and, mostcommonly, the Gauja Formations of Estonia and Latvia. TheJõksi (Kalmetumägi) exposure on the Piusa River, south-eastEstonia, yielded the richest material for evidence of predationon psammosteid heterostracans. Previously describedspecimens (Mark-Kurik 1966; Lebedev 1993) have beenredescribed and reinterpreted in this paper. An onlinecatalogue of collections of the Institute of Geology (Tallinn,Estonia) is accessible at http://sarv.gi.ee/. It includes the newspecimen numbers (used in this paper) as well as the old ones(with prefix Pi) published earlier.
Larnovaspis kneri
(Lankester, 1868) from the ChortkivRegional Stage (Lower Devonian, Lochkovian) of the Gorodoklocality (left bank of the Seret River, Podolia, Ukraine) ishoused in the Institute of Geology and Geography, Lithuania,Vilnius (LGGI). The concretion, which contained thespecimen LGGI 5-800 was chemically prepared, providingaccess to the visceral side of the armour.
Materials kept in the Palaeontological Institute of theRussian Academy of Sciences (PIN) are from the Middle-Upper Devonian sections in the Leningrad and Tula Regions.
Fish remains with bite marks from the holdings of theNatural History Museum of Latvia are from the UpperDevonian Ogre Formation (Frasnian) on the left bank of theImula River, and from the Ketleri Formation (Famennian)on the left bank of the Ciecere River close to Pavari hamlet.
The specimens of Middle-Late Devonian psammosteidsand Late Devonian arthrodires from the Leningrad andPskov Regions (Russia) are housed in the PalaeontologicalMuseum of St Petersburg University (PM SPU).
X-ray tomography (X-ray MicroCT) scanning wasperformed using a SkyScan 1172 (Kontich, Belgium).
Results
Lower Devonian examples
The earliest bite trace is found on the dorsal shield of thepteraspidiform heterostracan
Larnovaspis kneri
(Lankester,1868) (Fig. 1A–C).
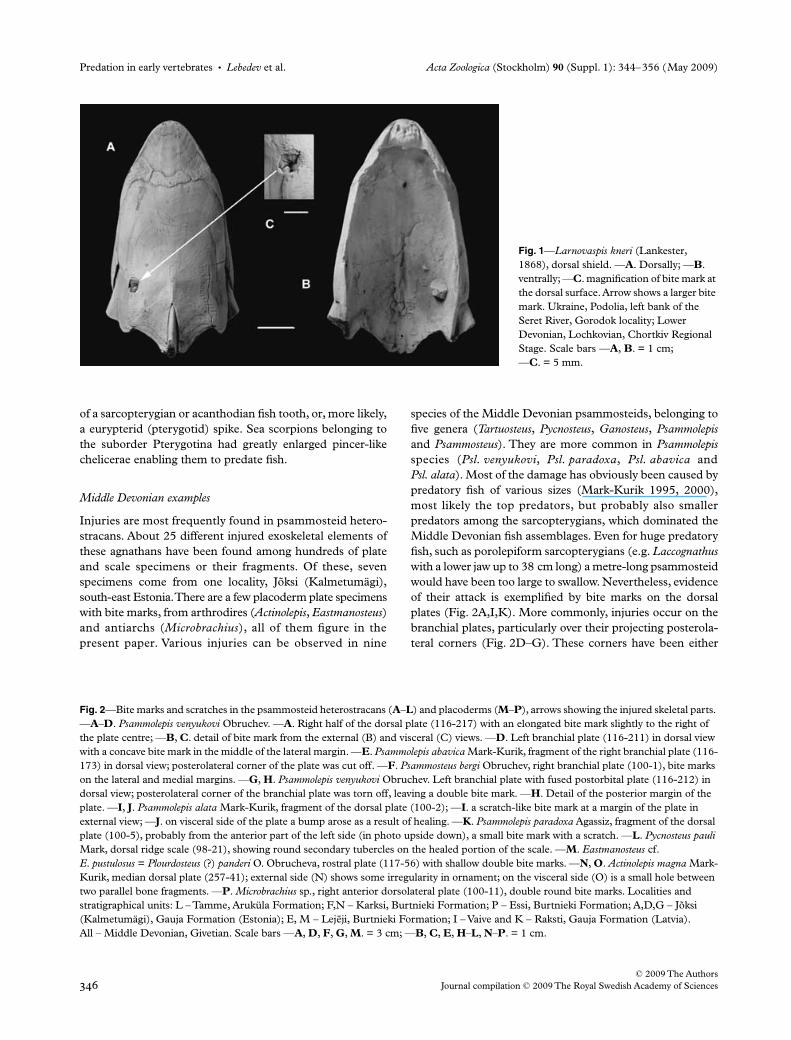
The left posterior quarter of the dorsal plate bears a smallpit anteriorly, in which the external dentine layer is missing.The second trace is larger and is located approximately5 mm posterolaterally from the first; it is an ovoid breach5 mm wide, 6 mm long and up to 3 mm deep (Fig. 1C). Theanterior part of the pit bottom is a gently sloping surfaceinterrupted transversely by an abrupt swelling and a crack.This area bears regular ornamentation of longitudinaldentine ridges with four seismo-sensory grooves transectingthem. The posterior part of this area has short, irregularlydisposed dentine ridges, which formed during the woundhealing. The lateral and posterior walls of the rupture arevertical and the plate edges have regenerated with short andcurved dentine ridges, which are wider than the straightlongitudinal ones ornamenting the rest of the dorsal plate.These secondary ridges are especially clearly expressed at themesial wall of the breach. The bottom of the posterior part ofthe pit is formed from two small sheets. The lateral sheet isslightly lifted and is ornamented by very narrow longitudinaldentine ridges. The mesial sheet is concave, bears narrowtransverse ridges and is fused to the mesial edge of the breach.
The internal structure of the damaged area confirms themechanical origin of this trace during the animal’s lifetime(Fig. 1B), that is by a predator’s stroke rather than ecto-parasitic action. The anterior small breach is marked with threesplinters of broken basal layer, which are closely spacedso that no newly formed tissue shows between them. Thevisceral side of the larger damaged area is formed from twolarge transversely directed bone splinters. The predatorystroke was directed anterodorsally, and the mesio-posteriorcorner houses part of the hitting spur. The outline of thisobject is a rounded-hexagonal; its diameter is about 1.0 mmand a cross-section shows small pores. This might be the tip
Predation in early vertebrates
•
Lebedev
et al.
Acta Zoologica
(Stockholm)
90
(Suppl. 1): 344–356 (May 2009)
© 2009 The Authors
346
Journal compilation © 2009 The Royal Swedish Academy of Sciences
of a sarcopterygian or acanthodian fish tooth, or, more likely,a eurypterid (pterygotid) spike. Sea scorpions belonging tothe suborder Pterygotina had greatly enlarged pincer-likechelicerae enabling them to predate fish.
Middle Devonian examples
Injuries are most frequently found in psammosteid hetero-stracans. About 25 different injured exoskeletal elements ofthese agnathans have been found among hundreds of plateand scale specimens or their fragments. Of these, sevenspecimens come from one locality, Jõksi (Kalmetumägi),south-east Estonia. There are a few placoderm plate specimenswith bite marks, from arthrodires (
Actinolepis
,
Eastmanosteus
)and antiarchs (
Microbrachius
), all of them figure in thepresent paper. Various injuries can be observed in nine
species of the Middle Devonian psammosteids, belonging tofive genera (
Tartuosteus
,
Pycnosteus
,
Ganosteus
,
Psammolepis
and
Psammosteus
). They are more common in
Psammolepis
species (
Psl. venyukovi
,
Psl. paradoxa
,
Psl. abavica
and
Psl. alata
). Most of the damage has obviously been caused bypredatory fish of various sizes (Mark-Kurik 1995, 2000),most likely the top predators, but probably also smallerpredators among the sarcopterygians, which dominated theMiddle Devonian fish assemblages. Even for huge predatoryfish, such as porolepiform sarcopterygians (e.g.
Laccognathus
with a lower jaw up to 38 cm long) a metre-long psammosteidwould have been too large to swallow. Nevertheless, evidenceof their attack is exemplified by bite marks on the dorsalplates (Fig. 2A,I,K). More commonly, injuries occur on thebranchial plates, particularly over their projecting posterola-teral corners (Fig. 2D–G). These corners have been either
Fig. 1—Larnovaspis kneri (Lankester, 1868), dorsal shield. —A. Dorsally; —B. ventrally; —C. magnification of bite mark at the dorsal surface. Arrow shows a larger bite mark. Ukraine, Podolia, left bank of the Seret River, Gorodok locality; Lower Devonian, Lochkovian, Chortkiv Regional Stage. Scale bars —A, B. = 1 cm; —C. = 5 mm.
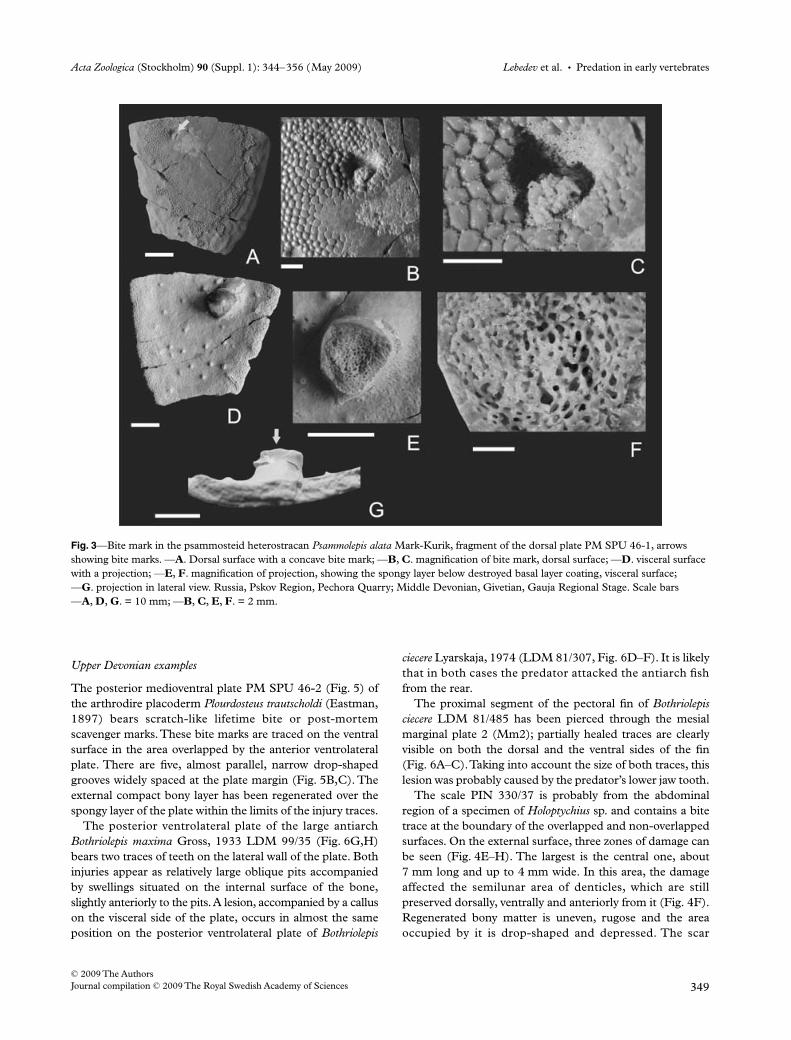
Fig. 2—Bite marks and scratches in the psammosteid heterostracans (A–L) and placoderms (M–P), arrows showing the injured skeletal parts. —A–D. Psammolepis venyukovi Obruchev. —A. Right half of the dorsal plate (116-217) with an elongated bite mark slightly to the right of the plate centre; —B, C. detail of bite mark from the external (B) and visceral (C) views. —D. Left branchial plate (116-211) in dorsal view with a concave bite mark in the middle of the lateral margin. —E. Psammolepis abavica Mark-Kurik, fragment of the right branchial plate (116-173) in dorsal view; posterolateral corner of the plate was cut off. —F. Psammosteus bergi Obruchev, right branchial plate (100-1), bite marks on the lateral and medial margins. —G, H. Psammolepis venyukovi Obruchev. Left branchial plate with fused postorbital plate (116-212) in dorsal view; posterolateral corner of the branchial plate was torn off, leaving a double bite mark. —H. Detail of the posterior margin of the plate. —I, J. Psammolepis alata Mark-Kurik, fragment of the dorsal plate (100-2); —I. a scratch-like bite mark at a margin of the plate in external view; —J. on visceral side of the plate a bump arose as a result of healing. —K. Psammolepis paradoxa Agassiz, fragment of the dorsal plate (100-5), probably from the anterior part of the left side (in photo upside down), a small bite mark with a scratch. —L. Pycnosteus pauli Mark, dorsal ridge scale (98-21), showing round secondary tubercles on the healed portion of the scale. —M. Eastmanosteus cf. E. pustulosus = Plourdosteus (?) panderi O. Obrucheva, rostral plate (117-56) with shallow double bite marks. —N, O. Actinolepis magna Mark-Kurik, median dorsal plate (257-41); external side (N) shows some irregularity in ornament; on the visceral side (O) is a small hole between two parallel bone fragments. —P. Microbrachius sp., right anterior dorsolateral plate (100-11), double round bite marks. Localities and stratigraphical units: L – Tamme, Aruküla Formation; F,N – Karksi, Burtnieki Formation; P – Essi, Burtnieki Formation; A,D,G – Jõksi (Kalmetumägi), Gauja Formation (Estonia); E, M – Lejeji, Burtnieki Formation; I – Vaive and K – Raksti, Gauja Formation (Latvia). All – Middle Devonian, Givetian. Scale bars —A, D, F, G, M. = 3 cm; —B, C, E, H–L, N–P. = 1 cm.

Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009) Lebedev et al. • Predation in early vertebrates
© 2009 The AuthorsJournal compilation © 2009 The Royal Swedish Academy of Sciences 347

Predation in early vertebrates • Lebedev et al. Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009)
© 2009 The Authors348 Journal compilation © 2009 The Royal Swedish Academy of Sciences
cut off or torn off (Fig. 2E,G). In the former case, the cornerhas a regular concave shape, as if cut with scissors, in thelatter case it has an irregular zigzag configuration.
Three types of trauma in psammosteids have been found:(1) Injuries on the dorsal plates. Three specimens (Fig. 2A,I,K)
suggest that predators grasped psammosteids in theirjaws, clasping the carapace with their fangs, but thenabandoned their prey, evidently being unable to swallowit because of its large size (such Psammolepis species asPsl. venyukovi and Psl. paradoxa with dorsoventrallyflattened bodies could be about 85 cm long and almost50 cm wide). In the case of Psl. venyukovi (Fig. 2A–C) itcan be estimated that the bite mark on the dorsal platewas approximately 20 cm from the right anterolateralmargin of the carapace. The grip was so strong that theplate broke into pieces and was recemented byregenerated tissue (Fig. 2C). In the second case, thestrike on the dorsal plate of Psl. alata (Fig. 2I,J) appearsto have been less powerful. The scratch-like tooth markwas shallow, and healed to leave a smooth bump on thevisceral side (Fig. 2J). The third bite mark on the platefragment of Psl. paradoxa (Fig. 2K), is small and round:it has not penetrated the plate and continues backwardas a scratch. All cases were the results of attacks from thefront, either from the right (Fig. 2A) or from the left(Fig. 2K) according to the position of the particularinjuries on the carapace of the above-mentionedpsammosteid species.
(2) Injuries over the branchial plates are found on fourspecimens (Fig. 2D–G). Most typically are missingposterolateral corners of the plates, either cut off inPsl. abavica (Fig. 2E) and Psl. paradoxa (see Obruchevand Mark-Kurik 1965; pl. 44; fig. 1) or torn off as inPsl. venyukovi (Fig. 2G) as their configuration afterattack shows (see above). On the Psammosteus bergibranchial plate the tooth marks are present on both freemargins and are situated on the same oblique line(Fig. 2F). The predator appears to have taken the plateinto its mouth but been unable to detach the posteriorportion. Two more injuries are evident on various partsof the branchial plates of Psl. venyukovi. A round,concave bite mark occurs approximately in the middle ofthe lateral margin of the left plate (Fig. 2D). A deepirregular mark occurs at the anterior end of specimen116-212 (Obruchev and Mark-Kurik 1965; pl. 52;fig. 1) from which the posterolateral corner has beentorn off (Fig. 2G).
(3) Several types of scratches, that are mostly linear damageon the external surface of the psammosteid exoskeletalplates, occur more often in the specimens that bear otherinjuries, for example, on the ventral side of the branchialplate specimen 116-211 (Fig. 2D). Some scratches werefound on the ridge scales, examples being Pycnosteus pauli(Fig. 2L) and Pycnosteus tuberculatus (Obruchev andMark-Kurik 1965; pl. 30; fig. 2). Secondary tubercles
have healed both the scratches and the more serioustraumas. These tubercles are usually simpler in shape, or,in the case of ridge scales, are similar to those of theskeletal plates (Fig. 2L).
Three placoderm specimens exhibit injuries on theirdermal plates (Fig. 2M–P). The rostral plate of Eastmanosteuscf. Eastmanosteus pustulatus (Fig. 2M) bears shallow and some-what irregular double bite marks. The position of the injuryon the fish head permits us to suppose that the predatorattacked the arthrodire from the front and sideways. There aretwo shallow conical pits, the anterior one being smaller, con-taining irregularly shaped tubercles. The larger (posterior)depression shows at least two rows of smaller tubercles,situated at the pit edges, rather than at the pit bottom.
The same kind of attack was suffered by Actinolepis magna(Fig. 2N,O). The anterior margin of the median dorsal plate(immediately behind the skull) bears a tooth mark that ishardly visible on the external side of the plate, and is recognizedby an irregular arrangement of tubercles. However, thevisceral side shows a round pit between two elongated bonefragments (Fig. 2O). The small antiarch Microbrachius sp.was probably attacked from the side, indicated by doubleround bite marks on the anterior dorsolateral plate of thetrunk shield (Fig. 2P).
A bite mark, possibly resulting from a sarcopterygian orarthrodire attack, was found on the dorsal plate of thepsammosteid heterostracan Psammolepis alata Mark-Kurik,1965 (PM SPU 46-1). The posterior fragment of the platebears a trace from a deep tooth penetration (Fig. 3). A roundconcave mark on the dorsal surface has been partly healedwith secondary dentine tubercles smaller than the regularornamentation tubercles (Fig. 3A–C). A high projection,triangular in cross-section, has been formed at the visceralsurface as a result of this healing (Fig. 3D–G). The projec-tion is composed of a thin lamellar basal layer externally andspongy aspidin layer on the interior. The spongy layer of thisprojection is denser in comparison to that of the plate andbears small vascular spaces and numerous short aspidintrabeculae (Fig. 3F).
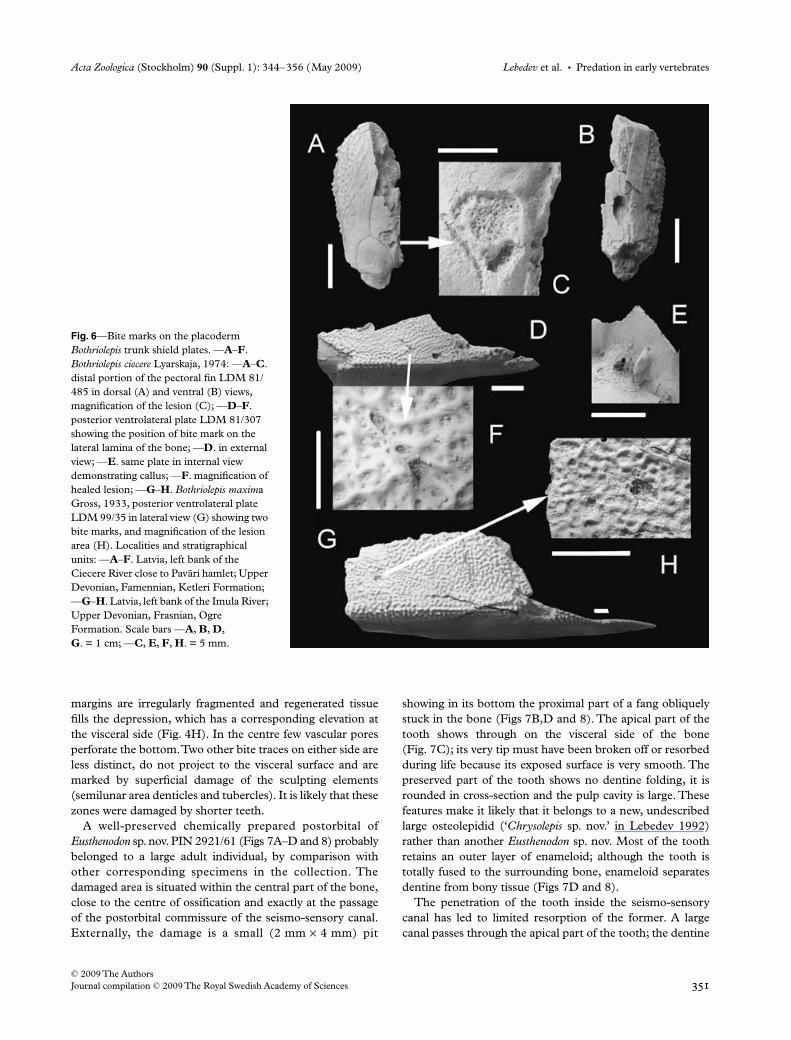
The scale PIN 1737/47 (Fig. 4A–D) comes from the bodyflank (?) of Glyptolepis sp. A bite trace occurs at the boundaryof the overlapped and non-overlapped surfaces within thesemilunar denticle area. Externally, the scale (Fig. 4A,C)bears an L-shaped depression about 5 mm long, 3 mm wideand 2 mm deep. The horseshoe-shaped denticles are partlyregenerated at the pit bottom, and the surrounding tissueseems to have suffered little deterioration. The visceral sur-face (Fig. 4B,D) exhibits a large elongated callus about10 mm long and up to 5 mm wide. No bone splinter edgesare visible as they evidently became obliterated during heal-ing. The callus surface is uneven but smoother than the sur-rounding intact visceral scale surface. On this latter surface,within the damaged area, are imprints of branching bloodvessels (Fig. 4B), which obviously provided an additionalblood supply for more intensive regeneration.
Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009) Lebedev et al. • Predation in early vertebrates
© 2009 The AuthorsJournal compilation © 2009 The Royal Swedish Academy of Sciences 349
Upper Devonian examples
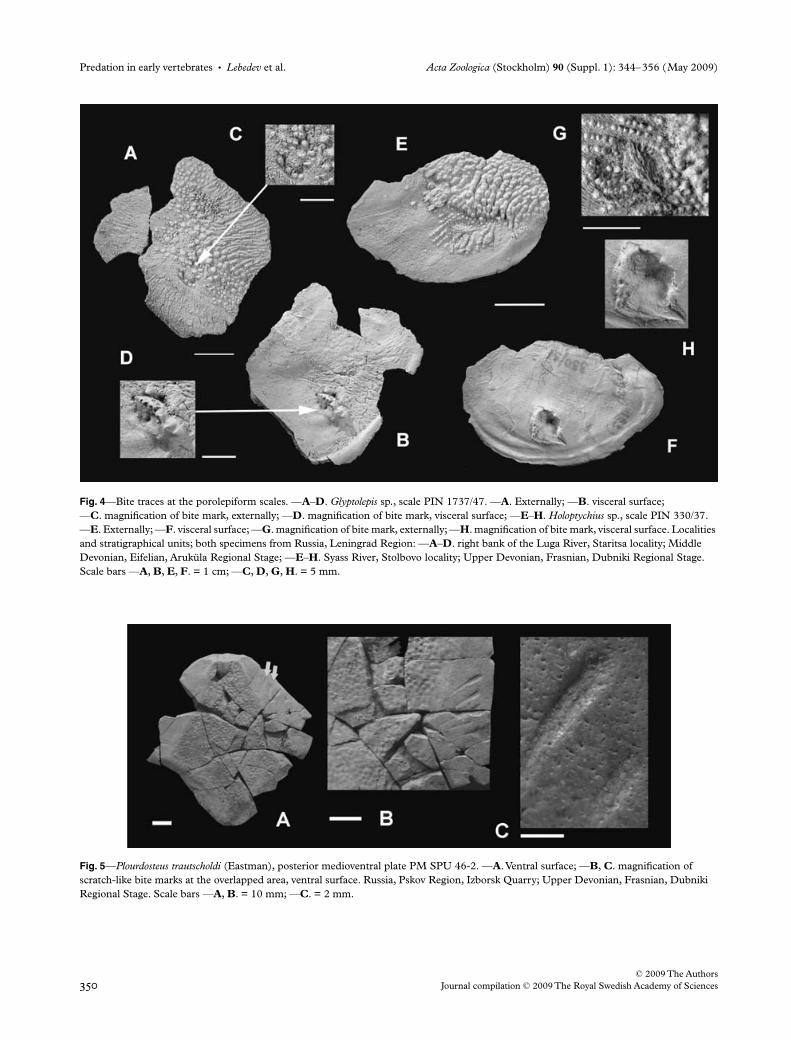
The posterior medioventral plate PM SPU 46-2 (Fig. 5) ofthe arthrodire placoderm Plourdosteus trautscholdi (Eastman,1897) bears scratch-like lifetime bite or post-mortemscavenger marks. These bite marks are traced on the ventralsurface in the area overlapped by the anterior ventrolateralplate. There are five, almost parallel, narrow drop-shapedgrooves widely spaced at the plate margin (Fig. 5B,C). Theexternal compact bony layer has been regenerated over thespongy layer of the plate within the limits of the injury traces.
The posterior ventrolateral plate of the large antiarchBothriolepis maxima Gross, 1933 LDM 99/35 (Fig. 6G,H)bears two traces of teeth on the lateral wall of the plate. Bothinjuries appear as relatively large oblique pits accompaniedby swellings situated on the internal surface of the bone,slightly anteriorly to the pits. A lesion, accompanied by a calluson the visceral side of the plate, occurs in almost the sameposition on the posterior ventrolateral plate of Bothriolepis
ciecere Lyarskaja, 1974 (LDM 81/307, Fig. 6D–F). It is likelythat in both cases the predator attacked the antiarch fishfrom the rear.
The proximal segment of the pectoral fin of Bothriolepisciecere LDM 81/485 has been pierced through the mesialmarginal plate 2 (Mm2); partially healed traces are clearlyvisible on both the dorsal and the ventral sides of the fin(Fig. 6A–C). Taking into account the size of both traces, thislesion was probably caused by the predator’s lower jaw tooth.
The scale PIN 330/37 is probably from the abdominalregion of a specimen of Holoptychius sp. and contains a bitetrace at the boundary of the overlapped and non-overlappedsurfaces. On the external surface, three zones of damage canbe seen (Fig. 4E–H). The largest is the central one, about7 mm long and up to 4 mm wide. In this area, the damageaffected the semilunar area of denticles, which are stillpreserved dorsally, ventrally and anteriorly from it (Fig. 4F).Regenerated bony matter is uneven, rugose and the areaoccupied by it is drop-shaped and depressed. The scar
Fig. 3—Bite mark in the psammosteid heterostracan Psammolepis alata Mark-Kurik, fragment of the dorsal plate PM SPU 46-1, arrows showing bite marks. —A. Dorsal surface with a concave bite mark; —B, C. magnification of bite mark, dorsal surface; —D. visceral surface with a projection; —E, F. magnification of projection, showing the spongy layer below destroyed basal layer coating, visceral surface; —G. projection in lateral view. Russia, Pskov Region, Pechora Quarry; Middle Devonian, Givetian, Gauja Regional Stage. Scale bars —A, D, G. = 10 mm; —B, C, E, F. = 2 mm.
Predation in early vertebrates • Lebedev et al. Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009)
© 2009 The Authors350 Journal compilation © 2009 The Royal Swedish Academy of Sciences
Fig. 4—Bite traces at the porolepiform scales. —A–D. Glyptolepis sp., scale PIN 1737/47. —A. Externally; —B. visceral surface; —C. magnification of bite mark, externally; —D. magnification of bite mark, visceral surface; —E–H. Holoptychius sp., scale PIN 330/37. —E. Externally; —F. visceral surface; —G. magnification of bite mark, externally; —H. magnification of bite mark, visceral surface. Localities and stratigraphical units; both specimens from Russia, Leningrad Region: —A–D. right bank of the Luga River, Staritsa locality; Middle Devonian, Eifelian, Aruküla Regional Stage; —E–H. Syass River, Stolbovo locality; Upper Devonian, Frasnian, Dubniki Regional Stage. Scale bars —A, B, E, F. = 1 cm; —C, D, G, H. = 5 mm.
Fig. 5—Plourdosteus trautscholdi (Eastman), posterior medioventral plate PM SPU 46-2. —A. Ventral surface; —B, C. magnification of scratch-like bite marks at the overlapped area, ventral surface. Russia, Pskov Region, Izborsk Quarry; Upper Devonian, Frasnian, Dubniki Regional Stage. Scale bars —A, B. = 10 mm; —C. = 2 mm.
Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009) Lebedev et al. • Predation in early vertebrates
© 2009 The AuthorsJournal compilation © 2009 The Royal Swedish Academy of Sciences 351
margins are irregularly fragmented and regenerated tissuefills the depression, which has a corresponding elevation atthe visceral side (Fig. 4H). In the centre few vascular poresperforate the bottom. Two other bite traces on either side areless distinct, do not project to the visceral surface and aremarked by superficial damage of the sculpting elements(semilunar area denticles and tubercles). It is likely that thesezones were damaged by shorter teeth.
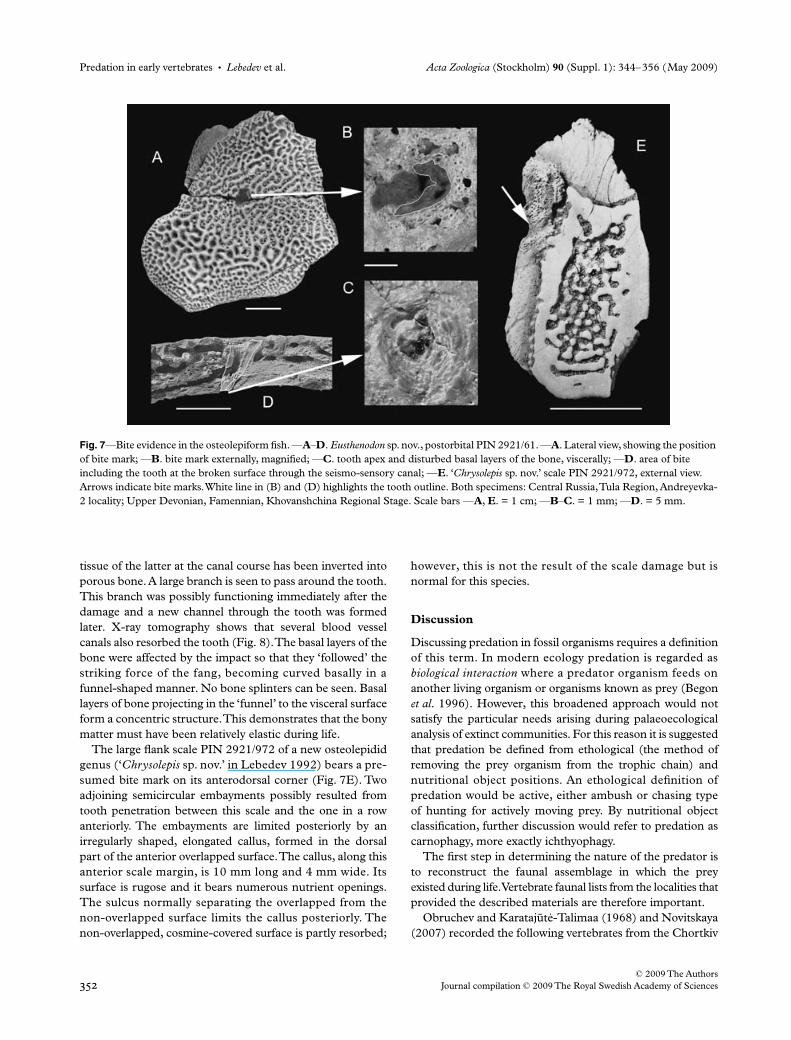
A well-preserved chemically prepared postorbital ofEusthenodon sp. nov. PIN 2921/61 (Figs 7A–D and 8) probablybelonged to a large adult individual, by comparison withother corresponding specimens in the collection. Thedamaged area is situated within the central part of the bone,close to the centre of ossification and exactly at the passageof the postorbital commissure of the seismo-sensory canal.Externally, the damage is a small (2 mm × 4 mm) pit
showing in its bottom the proximal part of a fang obliquelystuck in the bone (Figs 7B,D and 8). The apical part of thetooth shows through on the visceral side of the bone(Fig. 7C); its very tip must have been broken off or resorbedduring life because its exposed surface is very smooth. Thepreserved part of the tooth shows no dentine folding, it isrounded in cross-section and the pulp cavity is large. Thesefeatures make it likely that it belongs to a new, undescribedlarge osteolepidid (‘Chrysolepis sp. nov.’ in Lebedev 1992)rather than another Eusthenodon sp. nov. Most of the toothretains an outer layer of enameloid; although the tooth istotally fused to the surrounding bone, enameloid separatesdentine from bony tissue (Figs 7D and 8).
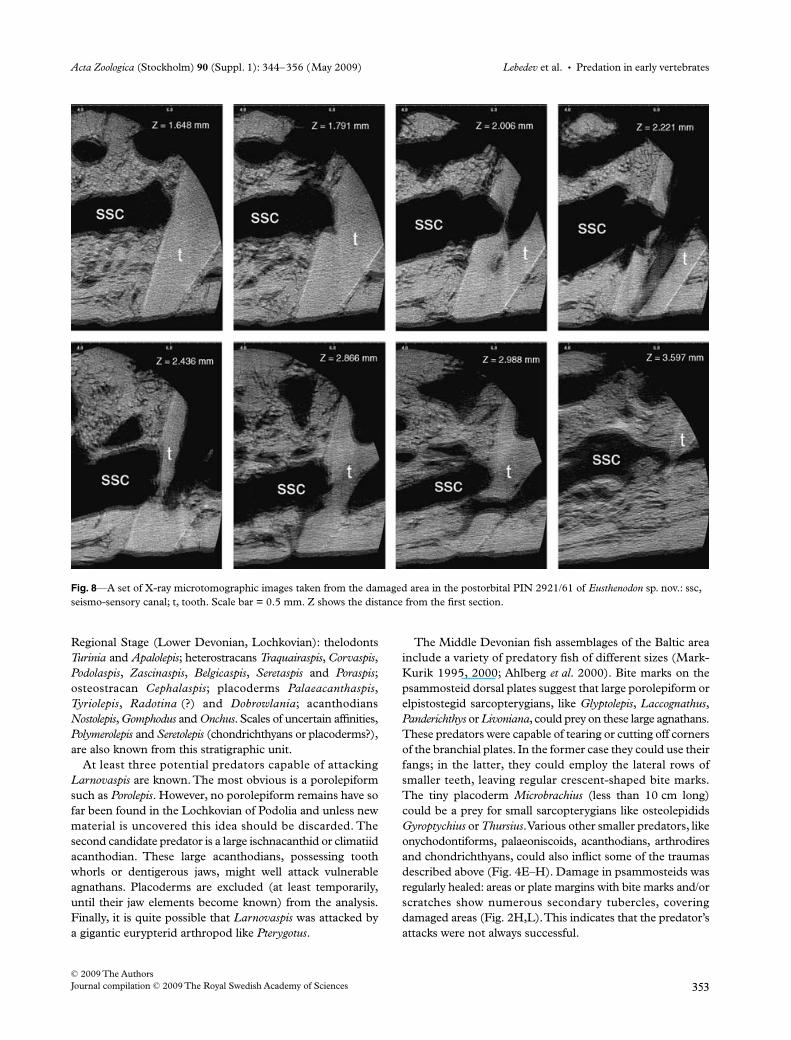
The penetration of the tooth inside the seismo-sensorycanal has led to limited resorption of the former. A largecanal passes through the apical part of the tooth; the dentine
Fig. 6—Bite marks on the placoderm Bothriolepis trunk shield plates. —A–F. Bothriolepis ciecere Lyarskaja, 1974: —A–C. distal portion of the pectoral fin LDM 81/485 in dorsal (A) and ventral (B) views, magnification of the lesion (C); —D–F. posterior ventrolateral plate LDM 81/307 showing the position of bite mark on the lateral lamina of the bone; —D. in external view; —E. same plate in internal view demonstrating callus; —F. magnification of healed lesion; —G–H. Bothriolepis maxima Gross, 1933, posterior ventrolateral plate LDM 99/35 in lateral view (G) showing two bite marks, and magnification of the lesion area (H). Localities and stratigraphical units: —A–F. Latvia, left bank of the Ciecere River close to Pavari hamlet; Upper Devonian, Famennian, Ketleri Formation; —G–H. Latvia, left bank of the Imula River; Upper Devonian, Frasnian, Ogre Formation. Scale bars —A, B, D, G. = 1 cm; —C, E, F, H. = 5 mm.
Predation in early vertebrates • Lebedev et al. Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009)
© 2009 The Authors352 Journal compilation © 2009 The Royal Swedish Academy of Sciences
tissue of the latter at the canal course has been inverted intoporous bone. A large branch is seen to pass around the tooth.This branch was possibly functioning immediately after thedamage and a new channel through the tooth was formedlater. X-ray tomography shows that several blood vesselcanals also resorbed the tooth (Fig. 8). The basal layers of thebone were affected by the impact so that they ‘followed’ thestriking force of the fang, becoming curved basally in afunnel-shaped manner. No bone splinters can be seen. Basallayers of bone projecting in the ‘funnel’ to the visceral surfaceform a concentric structure. This demonstrates that the bonymatter must have been relatively elastic during life.
The large flank scale PIN 2921/972 of a new osteolepididgenus (‘Chrysolepis sp. nov.’ in Lebedev 1992) bears a pre-sumed bite mark on its anterodorsal corner (Fig. 7E). Twoadjoining semicircular embayments possibly resulted fromtooth penetration between this scale and the one in a rowanteriorly. The embayments are limited posteriorly by anirregularly shaped, elongated callus, formed in the dorsalpart of the anterior overlapped surface. The callus, along thisanterior scale margin, is 10 mm long and 4 mm wide. Itssurface is rugose and it bears numerous nutrient openings.The sulcus normally separating the overlapped from thenon-overlapped surface limits the callus posteriorly. Thenon-overlapped, cosmine-covered surface is partly resorbed;
however, this is not the result of the scale damage but isnormal for this species.
Discussion
Discussing predation in fossil organisms requires a definitionof this term. In modern ecology predation is regarded asbiological interaction where a predator organism feeds onanother living organism or organisms known as prey (Begonet al. 1996). However, this broadened approach would notsatisfy the particular needs arising during palaeoecologicalanalysis of extinct communities. For this reason it is suggestedthat predation be defined from ethological (the method ofremoving the prey organism from the trophic chain) andnutritional object positions. An ethological definition ofpredation would be active, either ambush or chasing typeof hunting for actively moving prey. By nutritional objectclassification, further discussion would refer to predation ascarnophagy, more exactly ichthyophagy.
The first step in determining the nature of the predator isto reconstruct the faunal assemblage in which the preyexisted during life. Vertebrate faunal lists from the localities thatprovided the described materials are therefore important.
Obruchev and KaratajutE-Talimaa (1968) and Novitskaya(2007) recorded the following vertebrates from the Chortkiv
Fig. 7—Bite evidence in the osteolepiform fish. —A–D. Eusthenodon sp. nov., postorbital PIN 2921/61. —A. Lateral view, showing the position of bite mark; —B. bite mark externally, magnified; —C. tooth apex and disturbed basal layers of the bone, viscerally; —D. area of bite including the tooth at the broken surface through the seismo-sensory canal; —E. ‘Chrysolepis sp. nov.’ scale PIN 2921/972, external view. Arrows indicate bite marks. White line in (B) and (D) highlights the tooth outline. Both specimens: Central Russia, Tula Region, Andreyevka-2 locality; Upper Devonian, Famennian, Khovanshchina Regional Stage. Scale bars —A, E. = 1 cm; —B–C. = 1 mm; —D. = 5 mm.
Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009) Lebedev et al. • Predation in early vertebrates
© 2009 The AuthorsJournal compilation © 2009 The Royal Swedish Academy of Sciences 353
Regional Stage (Lower Devonian, Lochkovian): thelodontsTurinia and Apalolepis; heterostracans Traquairaspis, Corvaspis,Podolaspis, Zascinaspis, Belgicaspis, Seretaspis and Poraspis;osteostracan Cephalaspis; placoderms Palaeacanthaspis,Tyriolepis, Radotina (?) and Dobrowlania; acanthodiansNostolepis, Gomphodus and Onchus. Scales of uncertain affinities,Polymerolepis and Seretolepis (chondrichthyans or placoderms?),are also known from this stratigraphic unit.
At least three potential predators capable of attackingLarnovaspis are known. The most obvious is a porolepiformsuch as Porolepis. However, no porolepiform remains have sofar been found in the Lochkovian of Podolia and unless newmaterial is uncovered this idea should be discarded. Thesecond candidate predator is a large ischnacanthid or climatiidacanthodian. These large acanthodians, possessing toothwhorls or dentigerous jaws, might well attack vulnerableagnathans. Placoderms are excluded (at least temporarily,until their jaw elements become known) from the analysis.Finally, it is quite possible that Larnovaspis was attacked bya gigantic eurypterid arthropod like Pterygotus.
The Middle Devonian fish assemblages of the Baltic areainclude a variety of predatory fish of different sizes (Mark-Kurik 1995, 2000; Ahlberg et al. 2000). Bite marks on thepsammosteid dorsal plates suggest that large porolepiform orelpistostegid sarcopterygians, like Glyptolepis, Laccognathus,Panderichthys or Livoniana, could prey on these large agnathans.These predators were capable of tearing or cutting off cornersof the branchial plates. In the former case they could use theirfangs; in the latter, they could employ the lateral rows ofsmaller teeth, leaving regular crescent-shaped bite marks.The tiny placoderm Microbrachius (less than 10 cm long)could be a prey for small sarcopterygians like osteolepididsGyroptychius or Thursius. Various other smaller predators, likeonychodontiforms, palaeoniscoids, acanthodians, arthrodiresand chondrichthyans, could also inflict some of the traumasdescribed above (Fig. 4E–H). Damage in psammosteids wasregularly healed: areas or plate margins with bite marks and/orscratches show numerous secondary tubercles, coveringdamaged areas (Fig. 2H,L). This indicates that the predator’sattacks were not always successful.
Fig. 8—A set of X-ray microtomographic images taken from the damaged area in the postorbital PIN 2921/61 of Eusthenodon sp. nov.: ssc, seismo-sensory canal; t, tooth. Scale bar = 0.5 mm. Z shows the distance from the first section.

Predation in early vertebrates • Lebedev et al. Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009)
© 2009 The Authors354 Journal compilation © 2009 The Royal Swedish Academy of Sciences
The faunal list from the Late Devonian Stolbovo localityand a number of smaller ones of the Frasnian DubnikiRegional Stage includes psammosteid agnathan Psammosteus,acanthodians Persacanthus, Acanthodes, arthrodires Holonema,Gyroplacosteus, Plourdosteus, Eastmanosteus, antiarchs Asterolepisand Bothriolepis, porolepiform sarcopterygian Holoptychius,tristichopterid Jarvikina, osteolepidid Latvius (?), elpisto-stegid Parapanderichthys, dipnoans Dipterus and Rhinodipterus.Of those, only the large predators Holoptychius, Jarvikina andParapanderichthys might attack Holoptychius and leave thebite traces on the scale described above, because the victimfish was itself very large (Fig. 4E–H).
The fish assemblage from the Imula fossil site includesfewer taxa than the complete vertebrate list from the OgreFormation (Upper Devonian, Frasnian) (Lyarskaja andLukßeviçs 1992) and includes: heterostracan Psammosteus,antiarch Bothriolepis, acanthodian Devononchus, sarcoptery-gians Holoptychius, Platycephalichthys, and an undetermineddipnoan. Only the large porolepiform Holoptychius orosteolepiform Platycephalichthys might have been capable ofattacking the placoderm Bothriolepis maxima, which couldgrow to a very large size (up to 1 m long).
The Latvian Ciecere locality (Upper Devonian, Famennian)contains a rather diversified assemblage of vertebrates(Ahlberg et al. 1994; Lukßeviçs and Zupinß 2004). Amongpossible predators, the porolepiforms Holoptychius andVentalepis, an osteolepiform Cryptolepis, and a tetrapodVentastega could be possible attackers of Bothriolepis.Glyptopomus (?) from the same assemblage is known fromseveral small fragments and is unlikely to have been largeenough to feed on placoderms.
In the Famennian Andreyevka-2 locality the acanthodiansDevononchus, Acanthodei gen. et sp. nov. 1, 2, chondrichthyansEugeneodontidae (?) gen. et sp. nov., antiarchs Remigolepisand Bothriolepis, dipnoan Andreyevichthys, porolepiformHoloptychius, osteolepiform Eusthenodon, ‘Chrysolepis’;struniiform Strunius; actinopterygian Moythomasia; andtetrapod Tulerpeton are known (Lebedev 1992). Of these fish,subadult or early adult Eusthenodon might be attacked by alarge osteolepidid ‘Chrysolepis’, rather than another Eusthen-odon, on the evidence of penetrated tooth morphology. Oneof the two osteolepiforms, or the tetrapod Tulerpeton mightalso be capable of producing the damage to the scale belongingto ‘Chrysolepis’.
It is noteworthy that all the skeletal parts described herebelonged to adult individuals. The regenerated tissues overthem are apparently well developed, so that the prey wasprobably attacked at the early adult or subadult stage a longtime before its death, as new bony matter formation mighthave taken from several months to years. Predation on largepiscivorous fish by other large piscivorous fish happens inmodern basins in cases of food shortage, as well as aggressionof adults to younger individuals regardless of their owntrophic specialization. If predators hunt unusual prey, theirattack mode may not be suitable for this prey, so their
attempts are unlikely to be successful, and evidence of theattacks in the form of punctures and other trauma may bepreserved in fossil material. In contrast, hunting of theirusual prey would normally lead to success and consumptionof the victim, so that no bite marks would be preservedfor the palaeontological record. Some coprolites (in thecollections of the Institute of Geology, Tallinn University ofTechnology) contain tiny osteolepiform scales, demonstratingthat these sarcopterygians were prey for larger fish, even othersarcopterygians.
Analysis of aquatic consumers during the Silurian–Devonian interval carried out above shows a gradualincrease of the number of types of predators with time. Nobite traces are known from the Silurian and we may onlysuggest that ischnacanthiforms with dentigerous jaws arethe earliest vertebrate predators, based upon jaw morphol-ogy. The earliest known dentigerous ischnacanthiform,Xylacanthus, comes from the Wenlock or Ludlow ofCanada and Onchus from the same time interval of Czechia.Large ischnacanthiform dentigerous jaws are known fromthe end of the Late Silurian of England, the Baltic, Greenland,Canada, Nevada and Bolivia (Burrow 2004). Two pred-atory osteichthyans, Lophosteus and Andreolepis, are smallbut bear conical teeth and are known from the Late Silurianof the Baltic (Gross 1971; Janvier 1978). The Late Silurianof China has yielded one more presumed sarcopterygian,Psarolepis, which possesses well-developed fangs and lateralrows of smaller teeth (Zhu and Schultze 1997; Zhu et al.1999).
The Early Devonian shows an increase in predatorswhich, as well as ischnacanthiform acanthodians andosteichthyans, include chondrichthyans. Arthrodires,appearing at the beginning of the Early Devonian, haveincreased in number by the end of this period. Starting fromthe Middle Devonian some arthrodires are thought to bepredatory (Dennis-Bryan and Miles 1981; Davidson andTrewin 2005). Osteolepiform rhipidistians like Tristichop-terus, Koharalepis, Vorobjevaia, Kenichthys and a rhizodon-tiform Notorhizodon appear during the Middle Devonian;these groups attains maximum diversity and size during theLate Devonian. The largest Devonian osteolepiforms andrhizodontiforms and the Carboniferous genera Megalichthysand Rhizodus are crown predators in their communities.Large struniiforms exist during the Givetian–Frasnian.The earliest tetrapods known from the Upper Frasnian–Famennian join the aquatic vertebrate assemblage as piscivo-rous predators.
The following predatory vertebrates become known bythe end of the Devonian: acanthodians, chondrichthyans,arthrodires, porolepiform, osteolepiform, struniiform andrhizodontiform sarcopterygians and tetrapods. This expan-sion of predatory vertebrate groups might result from anincrease in the number of ecotopes suitable for vertebratedwelling and a corresponding growth in the number of preytaxa. No predatory agnathans are known from the geological

Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009) Lebedev et al. • Predation in early vertebrates
© 2009 The AuthorsJournal compilation © 2009 The Royal Swedish Academy of Sciences 355
record (if the parasitic and possibly scavenging Cyclostomata arenot considered). Within acanthodians only ischnacanthiformsare mainly predatory. Apart from brachythoracid arthrodires,no predatory placoderms are known. Reliably identifiedchondrichthyans do not demonstrate predatory-typedentitions before the Givetian. Within sarcopterygians,dipnoans, including Diabolepis, show phytophagous orsclerophagous dentitions right from their first records in theLochkovian.
A special place is occupied by porolepiforms, osteolepi-forms, struniiforms, rhizodontiforms and actinistians, inwhich predatory dentitions developed according more or lessto the same pattern and showed little change throughout theDevonian. No other trophic specializations are known withinthese fish taxa.
Conclusions
– Analysis of bite marks and subsequent pathologicalchanges enables the study of the relationships withpredatory members of the community and aids studies ofskeletal tissue remodelling.
– Criteria used to determine whether a structure shouldbe classified as an injury trace are worked out: thelocation of the damage; the presence of regeneratedtissue; the presence of damage traces or their repair onthe internal surface of the bony element; the specificoutline of the injury trace. Only damage which satisfiesall these criteria can be considered reliable evidence ofpredation.
– Lower, Middle and Upper Devonian specimens bearinglifetime traces of bites belonging to heterostracans,placoderms and sarcopterygians are described andinterpreted.
– Vertebrate faunal lists from the localities from whichmaterials are described are presented and analysed.Possible predators are suggested.
– Analysis of aquatic consumers during the Silurian–Devonian interval shows a gradual increase of the numberof predator types with time. This expansion of predatoryvertebrate groups might result from an increase in thenumber of ecotopes suitable for vertebrate dwelling and acorresponding growth in the number of prey taxa.
Acknowledgements
E. Mark-Kurik would like to thank G. Baranov, Institute ofGeology at Tallinn University of Technology, for the photo-graphs of the specimens and for kind help with the composi-tion of Fig. 2. The X-ray MicroCT scanning was performedby Dr A. V. Pakhnevich (Palaeontological Institute of theRAS, Moscow). Useful comments and advice during manu-script preparation were given by two anonymous referees, towhom we are greatly indebted. One of them carried outthoughtful linguistic correction.
References
Ahlberg, P. E., Lukßeviçs, E. and Lebedev, O. 1994. The first tetrapodfinds from the Devonian (Upper Famennian) of Latvia. – Philosophical Transactions of the Royal Society of London B 343: 303–328.
Ahlberg, P. E., Lukßeviçs, E. and Mark-Kurik, E. 2000. A near-tetrapod from the Baltic Middle Devonian. – Palaeontology 43:533–548.
Begon, M., Townsend, C. and Harper, J. 1996. Ecology: Individuals,Populations and Communities, pp. 3–1068. Blackwell Science,London.
Burrow, C. J. 2004. Acanthodian fishes with dentigerous jaw bones:the Ischnacanthiformes and Acanthodopsis. In Young, G. C. (Ed.):Lower Vertebrates from the Palaeozoic. First International Palaeonto-logical Congress, Sydney, Australia, July 2002, Proceedings ofSymposium 6 (Palaeozoic Vertebrates). – Fossils and Strata 50: 8–22.
Capasso, L., Bacchia, F., Rabottini, N., Rotschield, B. M. and Mariani-Constantini, R. 1996. Fossil evidence of intraspecific aggressivebehaviour of Devonian giant fishes (Arthrodira, Dinichthyidae). – Journal of Paleopathology 8: 153–160.
Davidson, R. G. and Trewin, N. H. 2005. Unusual preservation ofthe internal organs of acanthodian and actinopterygian fish in theMiddle Devonian of Scotland. – Scottish Journal of Geology 41:129–134.
Dennis-Bryan, K. and Miles, R. S. 1981. A pachyosteomorpharthrodire from Gogo, Western Australia. – Zoological Journal ofthe Linnean Society 73: 213–258.
Gross, W. 1971. Lophosteus superbus Pander: Zähne, Zahnknochenund besondere Schuppenformen. – Lethaia 4: 131–152.
Halstead Tarlo, L. B. 1965. Psammosteiformes (Agnatha) – a reviewwith descriptions of new material from the Lower Devonian ofPoland. II. Systematic part. – Palaeontologica Polonica 15: 1–168,19 pls.
Janvier, P. 1978. On the oldest known teleostome fish Andreolepishedei Gross (Ludlow of Gotland), and the systematic position ofthe lophosteids. – Proceedings of the Academy of Sciences of theEstonian SSR, Geology 27: 88–95, 2 pls.
KaratajutE-Talimaa, V., Lebedev, O., Lukßeviçs, E. and Ivanov, A.2007. Evidence of predation and parasitism in the skeletons of theDevonian vertebrates. In Blom, H. and Brazeau, M. D. (Eds):40th Anniversary Symposium on Early Vertebrates/Lower Vertebrates.Abstracts Volume, Uppsala, Sweden, August 13–16, 2007. – Ichthyolith Issues Special Publication 10: 53–54.
Lebedev, O. A. 1992. The latest Devonian, Khovanian vertebratecommunity of Andreyevka-2 locality, Tula Region, Russia. InMark-Kurik, E. (Ed.): Fossil Fishes as Living Animals. – Academia1, pp. 265–272. Academy of Sciences of Estonia, Tallinn.
Lebedev, O. A. 1993. On the lifetime damages in the skeletons of theDevonian fishes. In Vyalov, O. S. and Fedonkin, M. A. (Eds):Trace Fossils of Extinct Organisms, pp. 75–86. Nauka Publishers,Moscow (in Russian).
Lukßeviçs, E. and Zupinß, I. 2004. Sedimentology, fauna, andtaphonomy of the Pavari site, Late Devonian of Latvia. – LatvijasUniversitåtes Raksti, Sérija Zemes un Vides zinåtnes 679: 99–119.
Lyarskaja, L. A. and Lukßeviçs, E. 1992. Distribution of agnathansand fishes in Silurian and Devonian deposits of Latvia. In Sorokin,V. (Ed.): Palaeontology and Stratigraphy of Phanerozoic of Latvia andBaltic Sea, pp. 46–62. Zinatne, Riga (in Russian).
Mark-Kurik, E. 1966. On some injuries of the exoskeleton ofpsammosteids (Agnatha). In R. F. Hecker (Ed.): Organisms andEnvironment in the Geological Past, pp. 55–60. Nauka, Moscow (inRussian).

Predation in early vertebrates • Lebedev et al. Acta Zoologica (Stockholm) 90 (Suppl. 1): 344–356 (May 2009)
© 2009 The Authors356 Journal compilation © 2009 The Royal Swedish Academy of Sciences
Mark-Kurik, E. 1995. Trophic relations of Devonian fishes. – Geobios, Mémoire Spécial 19: 121–123.
Mark-Kurik, E. 2000. The Middle Devonian fishes of the BalticStates (Estonia, Latvia) and Belarus. – Courier ForschungsinstitutSenckenberg 223: 309–324.
Novitskaya, L. I. 2007. Evolution of taxonomic diversity of agnathanvertebrates at the generic and species levels (Heterostraci: ordersCyathaspidiformes, Pteraspidiformes. – Paleontologicheskiy Zhurnal3: 33–46 (in Russian).
Obruchev, D. V. and Mark-Kurik, E. J. 1965. Devonian Psammosteids(Agnatha, Psammosteidae) of the USSR. Institute of Geology,
Academy of Sciences of the Estonian SSR, Tallinn (in Russian,English summary).
Obruchev, D. V. and KaratajutE-Talimaa, V. N. 1968. Vertebratefaunas and correlation of the Ludlovian and Lower Devoniandeposits of East Europe. In Obruchev, D. V. (Ed.): Essays on thePhylogeny and Systematics of Fossil Fishes and Agnathans, pp. 63–70.Nauka, Moscow (in Russian).
Zhu, M. and Schultze, H.-P. 1997. The oldest sarcopterygian fish. – Lethaia 30: 239–304.
Zhu, M., Yu, X.-B. and Janvier, P. 1999. A primitive fossil fish shedslight on the origin of bony fishes. – Nature 397: 607–610.
Related Documents











