CLONING AND STEM CELLS Volume 6, Number 3, 2004 © Mary Ann Liebert, Inc. Birth of African Wildcat Cloned Kittens Born from Domestic Cats MARTHA C. GÓMEZ, 1,2 C. EARLE POPE, 1 ANGELICA GIRALDO, 1,2 LESLIE A. LYONS, 3 REBECCA F. HARRIS, 1 AMY L. KING, 1 ALEX COLE, 1 ROBERT A. GODKE, 2 and BETSY L. DRESSER 1,4 ABSTRACT In the present study, we used the African Wildcat (Felis silvestris lybica) as a somatic cell donor to evaluate the in vivo developmental competence, after transfer into domestic cat re- cipients, of cloned embryos produced by the fusion of African Wildcat (AWC) fibroblast cell nuclei with domestic cat cytoplasts. Cloned embryos were produced by fusion of a single AWC somatic cell to in vivo or in vitro enucleated domestic cat cytoplasts. When the two sources of oocytes were compared, fusion rate was higher using in vivo–matured oocytes as recipient cytoplasts, but cleavage rate was higher after reconstruction of in vitro–matured oocytes. To determine the number of reconstructed embryos required per domestic cat recip- ient to consistently establish pregnancies, AWC cloned embryos were transferred within two groups: recipients (n 24) receiving 25 embryos and recipients (n 26) receiving 30 em- bryos. Twelve recipients (46.2%) receiving 30 embryos were diagnosed to be pregnant, while no pregnancies were established in recipients receiving 25 NT embryos. Also, to determine the influence of length of in vitro culture on pregnancy rate, we compared oviductal transfer on day 1 and uterine transfer on day 5, 6, or 7. Pregnancy rates were similar after transfer of embryos on day 1 (6/12; 50.0%), day 5 (4/9; 44.4%), or day 6 (2/5; 40.0%) to synchronous recip- ients, but the number of fetuses developing after transfer of embryos on day 1 (n 17), ver- sus day 5 (n 4) or day 6 (n 3) was significantly different. Of the 12 pregnant recipients, nine (75%) developed to term and fetal resorption or abortion occurred in the other three (25%) from day 30 to 48 of gestation. Of a total of 17 cloned kittens born, seven were stillborn, eight died within hours of delivery or up to 6 weeks of age, and two are alive and healthy. Peri- natal mortality was due to lung immaturity at premature delivery, placental separation and bacterial septicemia. Subsequent DNA analysis of 12 cat-specific microsatellite loci confirmed that all 17 kittens were clones of the AWC donor male. These AWC kittens represent the first wild carnivores to be produced by nuclear transfer. 247 INTRODUCTION T HE SURVIVAL OF MOST SPECIES in the Felidae family is considered as threatened or endan- gered. In fact, 36 of 37 species of this family are at risk of extinction. The population decline can be attributed to poaching and human destruction of natural ecosystems. As populations decrease, 1 Audubon Center for Research of Endangered Species, New Orleans, Louisiana 70131 USA. 2 Department of Animal Sciences, LSU Agricultural Center, Baton Rouge, Louisiana 70803 USA. 3 School of Veterinary Medicine, University of California Davis, Davis, California 95616 USA. 4 Department of Biological Sciences, University of New Orleans, New Orleans, Louisiana 70148 USA.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CLONING AND STEM CELLSVolume 6, Number 3, 2004© Mary Ann Liebert, Inc.
Birth of African Wildcat Cloned Kittens Born fromDomestic Cats
MARTHA C. GÓMEZ,1,2 C. EARLE POPE,1 ANGELICA GIRALDO,1,2 LESLIE A. LYONS,3REBECCA F. HARRIS,1 AMY L. KING,1 ALEX COLE,1 ROBERT A. GODKE,2
and BETSY L. DRESSER1,4
ABSTRACT
In the present study, we used the African Wildcat (Felis silvestris lybica) as a somatic celldonor to evaluate the in vivo developmental competence, after transfer into domestic cat re-cipients, of cloned embryos produced by the fusion of African Wildcat (AWC) fibroblast cellnuclei with domestic cat cytoplasts. Cloned embryos were produced by fusion of a singleAWC somatic cell to in vivo or in vitro enucleated domestic cat cytoplasts. When the twosources of oocytes were compared, fusion rate was higher using in vivo–matured oocytes asrecipient cytoplasts, but cleavage rate was higher after reconstruction of in vitro–maturedoocytes. To determine the number of reconstructed embryos required per domestic cat recip-ient to consistently establish pregnancies, AWC cloned embryos were transferred within twogroups: recipients (n � 24) receiving �25 embryos and recipients (n � 26) receiving �30 em-bryos. Twelve recipients (46.2%) receiving �30 embryos were diagnosed to be pregnant, whileno pregnancies were established in recipients receiving �25 NT embryos. Also, to determinethe influence of length of in vitro culture on pregnancy rate, we compared oviductal transferon day 1 and uterine transfer on day 5, 6, or 7. Pregnancy rates were similar after transfer ofembryos on day 1 (6/12; 50.0%), day 5 (4/9; 44.4%), or day 6 (2/5; 40.0%) to synchronous recip-ients, but the number of fetuses developing after transfer of embryos on day 1 (n � 17), ver-sus day 5 (n � 4) or day 6 (n � 3) was significantly different. Of the 12 pregnant recipients,nine (75%) developed to term and fetal resorption or abortion occurred in the other three (25%)from day 30 to 48 of gestation. Of a total of 17 cloned kittens born, seven were stillborn, eightdied within hours of delivery or up to 6 weeks of age, and two are alive and healthy. Peri-natal mortality was due to lung immaturity at premature delivery, placental separation andbacterial septicemia. Subsequent DNA analysis of 12 cat-specific microsatellite loci confirmedthat all 17 kittens were clones of the AWC donor male. These AWC kittens represent the firstwild carnivores to be produced by nuclear transfer.
247
INTRODUCTION
THE SURVIVAL OF MOST SPECIES in the Felidaefamily is considered as threatened or endan-
gered. In fact, 36 of 37 species of this family areat risk of extinction. The population decline canbe attributed to poaching and human destructionof natural ecosystems. As populations decrease,
1Audubon Center for Research of Endangered Species, New Orleans, Louisiana 70131 USA.2Department of Animal Sciences, LSU Agricultural Center, Baton Rouge, Louisiana 70803 USA.3School of Veterinary Medicine, University of California Davis, Davis, California 95616 USA.4Department of Biological Sciences, University of New Orleans, New Orleans, Louisiana 70148 USA.
5241_e03_p247-258 9/30/04 9:34 AM Page 247

insularization eventually results in subpopula-tion extinctions caused by detrimental geneticand demographic effects (Nowell and Jackson,1996a).
Domestic cats are a useful research model todevelop assisted reproductive technologies forthe conservation of endangered felids. During thelast two decades, the feasibility of oocyte recov-ery (in vivo maturation), in vitro maturation(IVM), in vitro fertilization (IVF), in vitro embryoculture (IVC) and embryo transfer (ET) to recip-ients has been demonstrated (Pope, 2000; Luvoni,2000). The domestic cat can also serve as a suc-cessful recipient of embryos from closely relatedsmall non-domestics cats as shown by the birthof Indian desert cat (Pope et al., 1993) and AfricanWildcat kittens (Pope et al., 2000) after transfer ofIVF-derived embryos.
Recently, the feasibility of producing viable do-mestic cat offspring by nuclear transfer was dem-onstrated (Shin et al., 2002). Regarding endan-gered felids, nuclear transfer is a potentiallyvaluable technique for assuring the continuationof species with few remaining numbers of ani-mals. In fact, cloning could maintain genetic vari-ability in endangered felids when there are fewanimals in a founder population by conservingthe maximum number of alleles for future breed-ing projects (Seidel, 2001).
Inter-species nuclear transfer involves thetransfer of a donor cell nucleus of one species intoan enucleated oocyte of another species. Since alimited number of endangered felid oocytes areavailable, the ooplasm of the domestic cat oocytecan be used as the recipient cytoplast of a somaticcell nucleus of an endangered cat. Recently, wehave shown that domestic cat ooplasm is a com-patible host for somatic cell nuclei from an AWCand the reconstructed embryos do undergo de-velopment in vitro (Gómez et al., 2003). Althoughthe feasibility of inter-species nuclear transfer hasbeen demonstrated only three live endangeredmammals have been produced: gaur (Lanza et al.,2000), mouflon (Loi et al., 2001), and banteng(Jansen et al., 2004) after transfer of NT embryosinto domestic ruminant recipients.
Incomplete nuclear reprogramming is a majorconstraint to the in vivo developmental potentialof cloned embryos (Kikyo and Wolffe, 2000).Most cloned blastocysts, regardless of species, arenot able to maintain fetal development to term(Colman, 2000). Therefore, to maximize preg-
nancy rates in species such as the pig, up to 62reconstructed embryos are transferred into eachrecipient (Polejaeva et al., 2000). In the domesticcat, we reported that more pregnancies occurredafter uterine transfer of �12 IVF/IVC-derivedembryos (Pope et al., 1993). In vivo developmen-tal potential of cloned embryos may be even moresusceptible to the stressful environment of in vitroculture. Although in vitro development of cat em-bryos, as measured by blastocyst developmentand number of cells per embryo, has improved(Pope et al., 1999; Gómez et al., 2001, 2003), thepresence of perturbations affecting embryo via-bility such as nutritional imbalances, oxidativestress, and defective genomic imprinting must beacknowledged (Leese, 2002).
In the present study, we used the African Wild-cat (Felis silvestris lybica) as a somatic cell donorto evaluate the in vivo developmental compe-tence, after transfer into domestic cat recipients,of cloned embryos produced by the fusion ofAWC fibroblast cell nuclei with domestic cat cy-toplasts. Specifically, our primary goals were to(1) determine the minimal number of recon-structed embryos required per recipient to estab-lish pregnancy; (2) evaluate the influence of invitro culture interval on establishment and main-tenance of fetal development in recipients aftertransfer of reconstructed embryos; and (3) evalu-ate factors affecting pre- and post-natal fetal sur-vival and describe characteristics of the kittens.
MATERIALS AND METHODS
Animals
Domestic cats used as oocyte donors and em-bryo recipients were group-housed in environ-mentally controlled rooms with a 14-h/10-hlight/dark cycle at 20–26°C. The antibody de-fined cats were purchased from Liberty Research,Inc. (Waverly, NY). The AWC male was housedin a separate room under the same conditions.The rooms were cleaned, and cats were fed oncedaily (Science Diet, Hill Pet Nutrition). Fresh wa-ter was available at all times. All animal proce-dures were approved by the Institutional AnimalCare and Use Committee of the Audubon Centerfor Research of Endangered Species as requiredby the Health Research Extension Act of 1985(Public Law 99-158).
GÓMEZ ET AL.248
5241_e03_p247-258 9/30/04 9:34 AM Page 248

Chemicals
Chemicals were purchased from Sigma Chem-ical Co. (St. Louis, MO), unless otherwise stated.
Establishment of adult AWC cell line
The cell line was generated from skin tissue col-lected by biopsy from a 3-year-old male AWC.Collected tissue was finely cut into 1-mm2 piecesand plated in a 75-cm2 tissue-culture flask (Nunc,Denmark) containing 7 mL of Dulbecco ModifiedEagles Medium (DMEM) supplemented with 50�g/mL of gentamicin and 10% (v/v) fetal bovineserum (FBS; Hyclone, Logan, UT) and cultured at 38°C in 5% CO2/air. After 7–10 days of incu-bation, monolayer outgrowths with fibroblastic-like morphology were disaggregated with 2.5mg/mL of pronase, re-suspended in DMEM with10% FBS and 10% (v/v) dimethyl sulfoxide, andcooled at 1.0°C/min to �80°C (Mr. Frosty; Nal-gene, Rochester, NY) before storage in liquid ni-trogen (LN2). To prepare fibroblasts as donor cellsfor NT, cells were thawed and cultured in DMEMcontaining 10% FBS for 1–3 passages. Then, whencells reached 80% confluency, FBS was reducedto 0.5% and cells were cultured for an additional5 days (serum starvation) before cryopreserva-tion and storage in LN2.
Oocyte maturation
For in vitro maturation, cumulus–oocyte com-plexes (COC) were recovered from ovaries of do-mestic cats within 2–5 h after ovariohysterec-tomy. Selected COCs were cultured in modifiedTCM-199 containing 1 IU/mL hCG, 0.5 IU/mLeCG, 10 �g/mL epidermal growth factor, and 3mg/mL BSA (Fraction V, fatty acid free; Sero-logical Proteins, Kankakee, IL) for 24 h in 5% CO2,5% O2 and 90% N2 at 38°C (Gómez et al., 2003).
To obtain in vivo–matured oocytes, domesticcats were treated with a total of 3–4 IU of porcine-follicle stimulating hormone (FSH, Sioux Bio-chemical, Sioux Center, IA) given in decreasingdaily doses (s.c.) for 4 days, followed by 3 IU ofporcine-luteinizing hormone (im; LH, Sioux Bio-chemical) on day 5. At 24–26 h after LH injection,oocytes were collected by laparoscopic aspirationof mature ovarian follicles (Gómez et al., 2000).In vivo–matured oocytes were used for NT im-mediately after aspiration.
Cumulus cells of in vivo– and in vitro–maturedoocytes were removed by vortexing in 1 mg/mLof hyaluronidase (type IV) for 5 min, followed bygentle mechanical pipetting. Denuded oocyteswere placed in Tyrode’s solution (Irvine Scien-tific, Santa Ana, CA) with 1% MEM nonessentialamino acids, 3 mg/mL BSA, 15 mM NaHCO3,0.36 mM pyruvate, 2.2 mM calcium lactate, 1 mMglutamine, and 50 �g/mL gentamicin (IVC-1medium) at 38°C in 5% CO2 until further use.
Nuclear transfer
The protocol for nuclear transfer has been de-scribed previously (Gómez et al., 2003). Briefly,denuded metaphase II oocytes were incubated for 15 min at 38°C in Ca2�-free and Mg2�-freemodified Tyrode’s salt solution supplementedwith 1% MEM nonessential amino acids, 3mg/mL BSA, 30 mM NaHCO3, 0.36 mM pyru-vate, 1 mM glutamine, 50 �g/mL gentamicin(ECM medium), 20 �g/mL Hoechst 33342 and 20�g/mL cytochalasin B (CCB). After incubation,oocytes were enucleated in ECM medium inwhich supplemental NaHCO3 was reduced to 15mM and to which 15 mM Hepes, 20 �g/mL CCBand 2 mg/mL sucrose were added. The first po-lar body and �10% of the underlying cytoplasmwere drawn into an enucleation pipette (20 �m,OD). Removal of the metaphase II plate was con-firmed by brief exposure to epifluorescence mi-croscopy.
A vial containing AWC fibroblast cells syn-chronized in G0/G1 phase by serum starvation(Gómez et al., 2003) was thawed prior to beingused for NT, and a single AWC fibroblast cell wasintroduced into the perivitelline space of eachenucleated oocyte. For fusion, each NT couplet,in a solution of 0.3 M mannitol and 0.1 mM Mg2�,was placed between two stainless-steel electrodesspaced 120 �m apart (LF-101; Nepa Gene, Tokyo,Japan) that were controlled by micromanipula-tors. Membrane fusion was induced by applyinga 3-sec AC pre-pulse of 20V, 1 MHz followed bytwo 30-�sec DC pulses of 240V/mm at intervalsof 0.5 sec. Following the fusion pulses, coupletswere washed and cultured in IVC-1 medium sup-plemented with 7.8 mM calcium lactate (IVC-1 �Ca2�) for 30 min., at which time fusion was eval-uated visually by confirming the presence or ab-sence of the donor cell in the perivitelline space.
BIRTH OF AFRICAN WILDCAT CLONED KITTENS 249
5241_e03_p247-258 9/30/04 9:34 AM Page 249

After 2–3 h of culture in IVC-1 � Ca2�, activa-tion was induced by placing fused couplets be-tween two electrodes in a fusion chamber con-taining 3 mL of a solution of 0.3 M mannitol, 0.1mM Mg2�, and 0.05 mM Ca2� and exposing themto two 60-�sec DC pulses of 120 V/mm. Then,couplets were incubated in 30-�L droplets ofIVC-1 medium supplemented with 10 �g/mL cy-cloheximide and 5 �g/mL CCB at 38°C in 5% CO2in air for 4 h.
Embryo culture
After activation, reconstructed embryos werecultured in 500 �L of IVC-1 medium in 5% CO2,5% O2, and 90% N2 at 38°C. Some couplets weretransferred to the oviducts of recipient females onday 1. The remaining couplets were cultured un-til day 2 or 3, at which time, after uncleaved cou-plets were recorded and removed, embryos wereplaced into 500 �L of fresh IVC-1 medium con-taining 1% MEM essential amino acids (IVC-1 �EAA). Then, on day 5, the number of morulaewas recorded and all embryos were moved intoTyrode’s solution containing 1% NEAA, 2% EAA,10% FBS, and the same supplements as in IVC-1medium (IVC-2 medium) and cultured untiltransfer to a recipient.
Embryo transfer
Cloned embryos were transferred into theoviducts or uteri of 50 gonadotropin-treated do-mestic short hair (DSH) recipients. Day 1 em-bryos were transferred to the oviducts of recipi-ents, some of which had served as oocyte donorson day 0 (n � 2) and others of which served asrecipients only after induction of ovulation (n �14) using a lower dose of FSH (1–3 IU) and ahigher dose of LH (5 IU) than the oocyte donorsreceived. For oviductal embryo transfer, the ab-dominal cavity was visualized via a 5-mm endo-scope inserted just above the umbilicus, and a 16gauge � 6.35 cm stainless steel trocar/cannulawas inserted transabdominally near the midline�5 cm below the umbilicus. Then, a 14.5-cm tomcat catheter (3.5 French, Sherwood Medical, St.Louis, MO) was passed through the 16-gaugecannula and directed through the infundibulumoverlaying the ovary and into the upper am-pullary portion of the oviduct. A 50-cm length ofsterile polyethylene tubing (PE 10, no. 427400,Becton Dickinson, Sparks, MD) containing the
embryos near the lower tip was threaded throughthe tom cat catheter, and the embryos were ex-pelled into the oviduct using positive pressurefrom a 1-mL syringe.
The recipients of day 5, 6, and 7 embryos weregonadotropin-treated females that had under-gone laparoscopic oocyte aspiration 5–7 days pre-viously. Briefly, one uterine horn was exterior-ized through a 1.5-cm mid-ventral incision andpunctured with a sterile 16-gauge round-tipped,short bevel trocar �1 cm from the anterior tip.Embryos were aspirated in �50 �L of IVC-2medium into a 14.5-cm tom cat catheter using a1-mL plastic syringe. The catheter was thenthreaded 4–5 cm into the uterine lumen before de-positing the embryos (Pope et al., 1994).
Pregnancy detection and parturition
Each recipient was checked for pregnancy sta-tus using abdominal ultrasonography on day21–23 after ovulation or oocyte aspiration. Preg-nant recipients were monitored by ultrasono-graphy on a weekly basis until day 60, at whichtime daily ultrasonographic examinations weremade to check for fetal movement and heart beat.Pregnant recipients were allowed to completegestation, at which time kittens were deliveredby Caesarean section, either (1) as an interventionafter onset of vaginal bleeding and/or occurrenceof labor symptoms or (2) on a pre-scheduled daybefore symptoms of parturition were seen. In aneffort to reduce the onset of premature parturi-tion, some pregnant cats (n � 3) were orally ad-ministered altrenogest (0.08 mg/kg, Regu-Mate,Hoechst; Kustritz, 2001) daily from days 55 to65–67 of gestation. Also, in an attempt to reduceperinatal kitten mortality (n � 7) cats weretreated with four injections (im), 12 h apart, of betamethasone (0.1 mg/kg, CelestoneSoluspan,Schering) starting at varying times after day 60 ofgestation.
Microsatellite analysis
To determine the clonal status of all kittens,DNA was extracted using standard phenol/chlo-roform techniques from the fibroblast cells andblood of the AWC donor, blood of the recipientdam, umbilical cord blood of live kittens, orspleen of dead kittens. Standard sodium hydrox-ide isolation was used to isolate DNA from cheekswabs of live kittens. Feline derived microsatel-
GÓMEZ ET AL.250
5241_e03_p247-258 9/30/04 9:34 AM Page 250
lite loci were amplified as previously reported byRaymond et al. (1999) using standard ABI fluo-rescent chemistries on an ABI 377. The resultswere analyzed using the software STRand(Hughes).
Statistical analyses
The Chi square test was used to test the dif-ferences between treatments. Statistical differ-ence was set at the p � 0.05 level.
RESULTS
Embryo production after NT
A total of 2,565 in vivo (n � 1751) and in vitro(n � 814) matured domestic cat oocytes were me-chanically enucleated and electrically fused tofrozen-thawed AWC fibroblasts. Fusion effi-ciency was higher when in vivo–matured oocytes(1696/1751; 96.8%) were used as recipient cyto-plasts as compared with in vitro–matured oocytes(736/814, 90.4%, p � 0.05). In contrast, cleavagerate of embryos reconstructed with in vitro–ma-tured oocytes (435/511, 85.1%) was higher thanthat of embryos reconstructed with in vivo–ma-tured oocytes (944/1194, 79.1%, p � 0.05). Somereconstructed couplets (n � 727) were not in-cluded in the cleavage rate evaluation becausethey were transferred to domestic cat recipientson day 1 before cleavage occurred.
Embryo transfer and pregnancies
To determine the number of reconstructed em-bryos that should be transferred per recipient to
consistently establish pregnancies, AWC clonedembryos were transferred into domestic cat re-cipients within two groups: (1) recipients (n � 24)that received fewer than 25 embryos (mean �19.4; range � 8–25), and (2) recipients (n � 26)that received more than 30 embryos (mean �41.7; range � 31–59). Also, to determine the in-fluence of length of in vitro culture on pregnancyrate we compared the transfer of day 1 embryosinto the oviduct (n � 588) with transfer of day 5(n � 467), day 6 (n � 355), and day 7 (n � 142)embryos into the uterus. Embryos were at the1–2-cell stage on day 1, morulae on day 5 and day6, and morulae and blastocysts on day 7 of cul-ture.
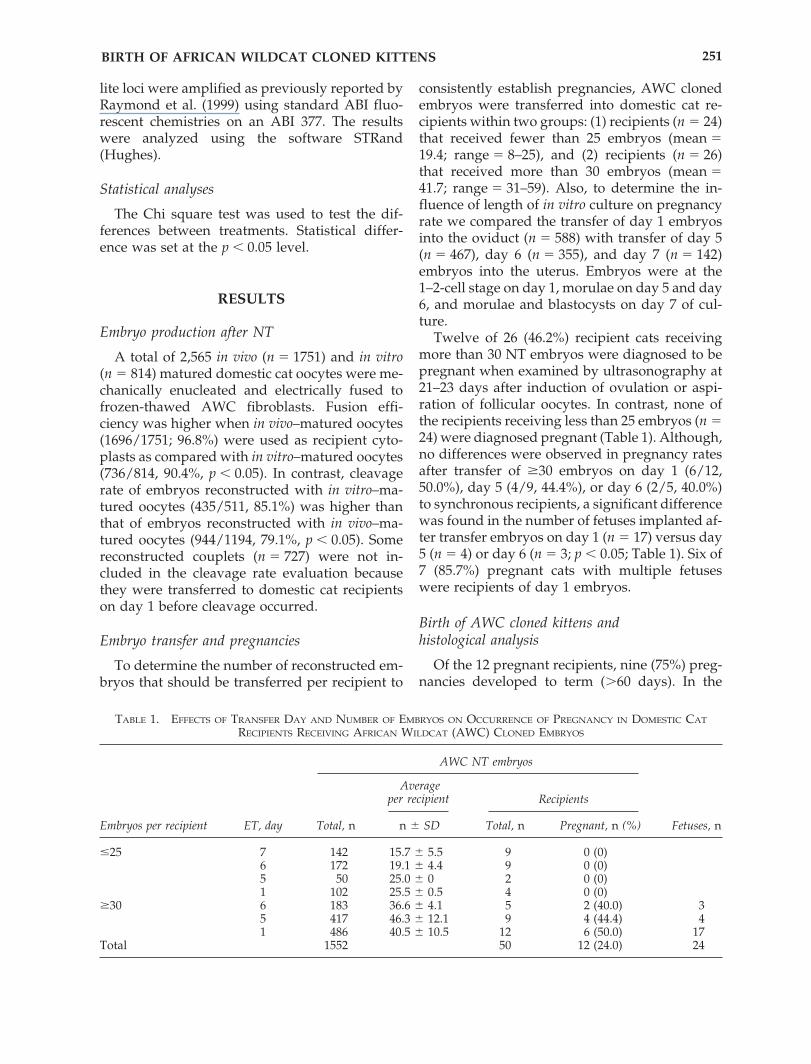
Twelve of 26 (46.2%) recipient cats receivingmore than 30 NT embryos were diagnosed to bepregnant when examined by ultrasonography at21–23 days after induction of ovulation or aspi-ration of follicular oocytes. In contrast, none ofthe recipients receiving less than 25 embryos (n �24) were diagnosed pregnant (Table 1). Although,no differences were observed in pregnancy ratesafter transfer of �30 embryos on day 1 (6/12,50.0%), day 5 (4/9, 44.4%), or day 6 (2/5, 40.0%)to synchronous recipients, a significant differencewas found in the number of fetuses implanted af-ter transfer embryos on day 1 (n � 17) versus day5 (n � 4) or day 6 (n � 3; p � 0.05; Table 1). Six of7 (85.7%) pregnant cats with multiple fetuseswere recipients of day 1 embryos.
Birth of AWC cloned kittens and histological analysis
Of the 12 pregnant recipients, nine (75%) preg-nancies developed to term (�60 days). In the
BIRTH OF AFRICAN WILDCAT CLONED KITTENS 251
TABLE 1. EFFECTS OF TRANSFER DAY AND NUMBER OF EMBRYOS ON OCCURRENCE OF PREGNANCY IN DOMESTIC CAT
RECIPIENTS RECEIVING AFRICAN WILDCAT (AWC) CLONED EMBRYOS
AWC NT embryos
Averageper recipient
Embryos per recipient ET, day Total, n n � SD Total, n Pregnant, n (%) Fetuses, n
25 7 142 15.7 � 5.5 9 0 (0)6 172 19.1 � 4.4 9 0 (0)5 50 25.0 � 0 2 0 (0)1 102 25.5 � 0.5 4 0 (0)
�30 6 183 36.6 � 4.1 5 2 (40.0) 35 417 46.3 � 12.1 9 4 (44.4) 41 486 40.5 � 10.5 12 6 (50.0) 17
Total 1552 50 12 (24.0) 24
Recipients
5241_e03_p247-258 9/30/04 9:34 AM Page 251
three (25%) other pregnancies, a total of five fe-tuses were present at day 21–23, four of whichwere reabsorbed between days 30 and 40, and onewhich was spontaneously aborted on day 48 ofgestation (Table 2).
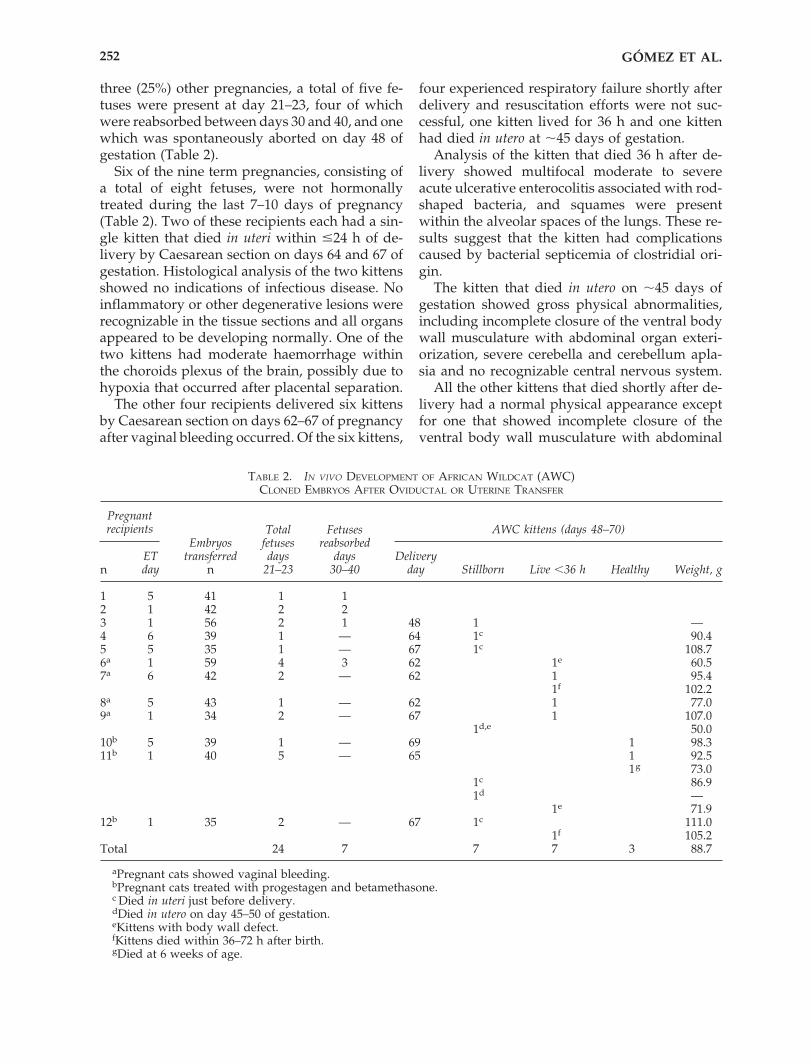
Six of the nine term pregnancies, consisting ofa total of eight fetuses, were not hormonallytreated during the last 7–10 days of pregnancy(Table 2). Two of these recipients each had a sin-gle kitten that died in uteri within 24 h of de-livery by Caesarean section on days 64 and 67 ofgestation. Histological analysis of the two kittensshowed no indications of infectious disease. Noinflammatory or other degenerative lesions wererecognizable in the tissue sections and all organsappeared to be developing normally. One of thetwo kittens had moderate haemorrhage withinthe choroids plexus of the brain, possibly due tohypoxia that occurred after placental separation.
The other four recipients delivered six kittensby Caesarean section on days 62–67 of pregnancyafter vaginal bleeding occurred. Of the six kittens,
four experienced respiratory failure shortly afterdelivery and resuscitation efforts were not suc-cessful, one kitten lived for 36 h and one kittenhad died in utero at �45 days of gestation.
Analysis of the kitten that died 36 h after de-livery showed multifocal moderate to severeacute ulcerative enterocolitis associated with rod-shaped bacteria, and squames were presentwithin the alveolar spaces of the lungs. These re-sults suggest that the kitten had complicationscaused by bacterial septicemia of clostridial ori-gin.
The kitten that died in utero on �45 days of gestation showed gross physical abnormalities,including incomplete closure of the ventral bodywall musculature with abdominal organ exteri-orization, severe cerebella and cerebellum apla-sia and no recognizable central nervous system.
All the other kittens that died shortly after de-livery had a normal physical appearance exceptfor one that showed incomplete closure of theventral body wall musculature with abdominal
GÓMEZ ET AL.252
TABLE 2. IN VIVO DEVELOPMENT OF AFRICAN WILDCAT (AWC) CLONED EMBRYOS AFTER OVIDUCTAL OR UTERINE TRANSFER
Total FetusesEmbryos fetuses reabsorbed
ET transferred days days Deliveryn day n 21–23 30–40 day Stillborn Live �36 h Healthy Weight, g
1 5 41 1 12 1 42 2 23 1 56 2 1 48 1 —4 6 39 1 — 64 1c 90.45 5 35 1 — 67 1c 108.76a 1 59 4 3 62 1e 60.57a 6 42 2 — 62 1 95.4
1f 102.28a 5 43 1 — 62 1 77.09a 1 34 2 — 67 1 107.0
1d,e 50.010b 5 39 1 — 69 1 98.311b 1 40 5 — 65 1 92.5
1.g 73.01c 86.91d —
1e 71.912b 1 35 2 — 67 1c 111.0
1f 105.2Total 24 7 7 7 3 88.7
aPregnant cats showed vaginal bleeding.bPregnant cats treated with progestagen and betamethasone.c Died in uteri just before delivery.dDied in utero on day 45–50 of gestation.eKittens with body wall defect.fKittens died within 36–72 h after birth.gDied at 6 weeks of age.
Pregnantrecipients AWC kittens (days 48–70)
5241_e03_p247-258 9/30/04 9:34 AM Page 252

organ exteriorization. Histological evaluation ofthe lungs of the kittens revealed variable collapseof alveolar spaces and presence of epithelialsquames within the lumen of alveolar spaces, in-dicating that alveoli were not inflated and thelungs were immature. Thus, prenatal and earlyneonatal mortality appears to have been primar-ily due to lung immaturity resulting from thespontaneous initiation of premature parturition.
Three pregnant cats that were hormonallytreated during the last 10–12 days of gestation de-livered eight kittens by Caesarean section. Thefour kittens that survived were born on 6 August2003, 15 November 2003, and 12 January 2004 at69, 65, and 67 days of gestation, respectively(Table 2).
Of the four kittens that did not survive, twodied in utero within hours of delivery on days 64and 66. One kitten that died within a few min-utes after delivery had incomplete closure of theventral body wall musculature with abdominalorgan exteriorization. Another kitten died in uteroon �50 days of gestation and was being resorbedat delivery.
Pathology analysis of one kitten that died inutero on day 64 revealed diffuse adrenal conges-tion and mild nephrosis. Necrosis of the renaltubular epithelium could be induced by endo-toxic shock. The hypoxia may have been causedby cardiovascular collapse and release of nephro-toxins before death. Therefore, prenatal deathwas possibly due to hypoxia that occurred afterplacenta separation and is probably analogous tothe prenatal death occurring in pregnant recipi-
ents that were not hormonally treated during thefinal stage of gestation.
Two kittens, currently 7 and 3 months of age(Fig. 1) are healthy and developing normally. Ofthe other two kittens, one died at 56 h after birthand one died at 6 weeks of age. Histological anal-ysis of the kitten that died at 56 h after birth in-dicated a diffuse moderate septic and supurativepneumonia with abnormally high bacterialcounts in several organs. The bacteria may havegained entry through the urachus and extendedinto the urinary bladder.
The kitten that died at 6 weeks of age had acongenital megaesophagus and acute supurativepneumonia. The primary cause of death waspneumonia, likely a result of aspiration of for-mula during bottle-feeding.
As shown in Table 2, the birth average weightof live kittens (92.3 g; range � 71.9–105.0 g) wassimilar to the average weight of kittens that diedduring the peri- and post-parturient period (87.4g; range � 50.0–111.0 g). Phenotypically, allcloned kittens (n � 17) were African Wildcats.Subsequent DNA analysis of 12 cat-specific mi-crosatellite loci performed with “blind” testing atthe School of Veterinary Medicine, University ofCalifornia, Davis, confirmed that they were iden-tical to DNA of the AWC donor male (“Jazz”;Table 3).
Placental analysis
A total of 11 placentae were analyzed. Macro-scopic analyses showed that five placentae had
BIRTH OF AFRICAN WILDCAT CLONED KITTENS 253
AB
FIG. 1. African wildcat cloned kittens (A) “Ditteaux” born on 6th August 2003, at two months after birth. (B) “Miles”and Otis” born on 15th November 2003, at seven days after birth.
5241_e03_p247-258 9/30/04 9:34 AM Page 253

patches of serous-lacy tissue replacing normalplacental tissue. Microscopic analysis of these fiveplacentae showed that areas of the placental ep-ithelium were reduced in thickness and hadshorter and thicker papillary projections ormerely a thin band of edematous fibrous con-nective tissue. These observations indicate theocurrence of multifocal placental atrophy with fi-brous tissue replacement. Curiously, these ab-normal placentae allowed development of kittensto term, including two kittens that survived. Theplacenta of a kitten that died in uteri before de-livery by Caesarean section showed excessive ab-normal mineralization and intermittent serosan-guineous exudates. These abnormalities were notpresent in all the placental sections analyzed;some sections were histologically normal.
DISCUSSION
In the present study, we demonstrated thatAWC kittens can be produced after transfer ofembryos derived by fusion of adult somatic cellsfrom one species with enucleated oocytes of aclosely related species (domestic cat). We haveshown that, under our current laboratory condi-tions, transfer of at least 30 NT embryos per re-cipient was required for establishing a preg-nancy. Also, the length of in vitro culture affectedthe number of embryos that successfully under-went fetal development. The primary causes ofperi-natal mortality were placental separation,
lung immaturity at premature delivery and bac-terial septicemia.
We obtained pregnancies only in recipientsthat received �30 NT embryos. A possible ex-planation for the high number of embryos re-quired for producing pregnancies is that most ofthe reconstructed embryos had inadequate nu-clear remodeling or incomplete reprogramming(Gómez et al., 2003). We previously demonstratedin AWC reconstructed embryos that 18% werefragmented and unable to undergo chromatin re-modeling, 38% arrested at the 8–10-cell stages and44% and 24% reached the morula and blastocystsstages, respectively (Gómez et al., 2003). In thisstudy, NT embryos were reconstructed from thesame AWC somatic cell line and oocytes weresimilar in type and source to those used in pre-vious NT experiments. Then, from the meannumber of embryos transferred on day 1 (40.5),day 5 (46.3), and day 6 (36.6), we can estimate thatat least 10 embryos should develop to the blasto-cyst stage. Based on these values, we suggest that,using our current methods, to consistently estab-lish pregnancies in domestic cats receiving AWCNT-derived embryos, at least 30 embryos shouldbe transferred per recipient. If it is assumed thatNT-derived cat blastocysts may be less develop-mentally competent than IVF-derived cat blasto-cysts, these numbers directly correlate with theminimal number of IVF derived embryos re-quired to reliably establish pregnancies in do-mestic cat recipients.
Domestic cat recipients receiving day-7 blasto-cysts (mean � 7.4) and morulae (mean � 8.3) onday 7 post-oocyte retrieval did not establish preg-nancies. During this portion of the study we weretransferring an average of 10 blastocysts per cat.We do not think that the longer in vitro cultureinterval was the primary reason for the develop-mental failure because we have produced do-mestic cat kittens after transfer of either 10 or 11IVF-derived day-7 blastocysts per recipient (Popeet al., 2003). Further experiments should deter-mine if pregnancies can be established after trans-ferring AWC cloned blastocysts.
In sheep, more lambs were produced afterearly transfer (day 2) of IVF embryos as comparedto later transfer at day 6 (Walmsley et al., 2004).Similarly, in the present study, a higher percent-age of NT embryos developed into fetuses whenthe transfer was done after a shorter period of cul-ture (day 1, n � 17/486, 3.5%) as compared withembryos that were cultured for longer periods
GÓMEZ ET AL.254
TABLE 3. ANALYSIS OF FELINE GENETIC MARKERS
Feline Cheek Umbilicalmarkers Fibroblasts Blood cells cord
FCA075 132/134 132/134 132/134 132/134FCA097 140/146 140/146 140/146 140/146FCA201 153/155 153/155 153/155 153/155FCA224 160/178 160/178 160/178 160/178FCA229 158/162 158/162 158/162 158/162FCA293 179/187 179/187 179/187 179/187FCA305 191/201 191/201 191/201 191/201FCA441 158/166 158/166 158/166 158/166FCA018 233/233 233/233 233/233 233/233FCA145 174/174 174/174 174/174 174/174FCA651 135/135 135/135 135/135 135/135FCA674 148/148 148/148 148/148 148/148
Values represent the sizes of the two alleles of the amplified microsatellites markers.
AWC donor“Jazz”
AWC cloned kittens(n � 17)
5241_e03_p247-258 9/30/04 9:34 AM Page 254

(day 5, n � 4/417, 1.0%; day 6, n � 3/183, 1.6%).From these results, we suggest that the develop-mental potential of some NT embryos is “res-cued” by transferring them to recipient cats as un-cleaved couplets and two-cell embryos; therebyminimizing the detrimental effects of in vitro cul-ture (Leese, 2002). Although more embryos im-planted after transfer on day 1, fetal mortality byday 48 of gestation also increased (6/17; 35.3%)as compared with embryos transferred on day 5(1/4; 25.0%) or day 6 (0/3; 0%). Also, three kit-tens that were born after embryo transfer on day1 had incomplete closure of the ventral body wallmusculature with abdominal organ exterioriza-tion. Because few fetuses were evaluated, we can-not conclude that the increase in fetal mortalityor the physical abnormalities were directly re-lated to the shorter period of culture. However,we can speculate that losses occurring after trans-fer on day 1 are not found after the transfer ofembryos on day 5 or 6 because the developmen-tal potential of such embryos is sufficiently re-duced by the extended in vitro culture intervalthat they are incapable of further development inutero.
Although embryo viability was demonstrated,embryo survival after transfer was quite modest(1.0–3.5%) and was similar to the overall effi-ciency of cloning reported in other species(1.0–4.0%; Wilmut and Paterson, 2003). In addi-tion to incomplete reprogramming of the differ-entiated nucleus, another factor that may con-tribute to low cloning efficiency is variation inoocyte quality. Both in vivo–matured and invitro–matured oocytes have been used as recipi-ent cytoplasts for production of cloned animals.For example, in sheep, higher blastocyst de-velopment rates, pregnancy rates and embryosurvival were reported when in vivo–maturedoocytes were used as donor cytoplasts (Wells etal., 1997). In contrast, in goats no differences werefound in fusion rates, embryo development andin the number of cloned kids produced fromoocytes harvested from FSH-stimulated donoranimals or from in vitro–matured oocytes recov-ered from abbatoir ovaries (Reggio et al., 2001).In pigs, in vivo–matured oocytes have been usedextensively as cytoplast recipients (Miyoshi et al.,2003). Walker et al. (2002) reported that invitro–matured pig oocytes could also be used toproduce cloned piglets; however, a direct com-parison between the two sources of cytoplasts hasnot been done. In the present study, we found
higher fusion rates with in vivo–matured cyto-plasts and higher cleavage rates with in vitro–ma-tured cytoplasts. In a previous study, we foundno differences in blastocyst development in vitrowhen in vivo–matured and in vitro–matured catoocytes were used as recipient cytoplasts of AWCcells (Gómez et al., 2003). A primary purpose ofthe present study was to find out if AWC kittenscould be produced by transfer of NT embryos todomestic cat recipients. Because we were unableto produce NT pregnancies after transferring 17 embryos per recipient in a previous study(Gómez et al., 2003), in the present study in or-der to transfer �25 embryos per recipient, formost transfers it was necessary to combine re-constructed embryos derived from both invitro–matured and in vivo–matured domestic catcytoplasts. The one domestic cat recipient receiv-ing NT embryos derived from in vivo–maturedcytoplasts did establish pregnancy and producea live kitten. We do not know if the other NT kit-tens originated from embryos derived from invivo–matured or in vitro–matured cytoplasts. Cer-tainly, the comparative developmental compe-tence of in vitro–matured versus in vivo–maturedcat cytoplasts requires investigation.
Several developmental abnormalities occur inpregnancies from somatic cell nuclear trans-fer–derived embryos, including a high rate ofabortion during early gestation and a number ofperinatal complications. One complication we en-countered in some cats was the occurrence ofvaginal bleeding by day 62–67 of pregnancy with-out overt signs of labor. Although the kittenswere delivered within the normal gestation pe-riod of domestic cats (62–71 days; Herron, 1977)and African Wildcats (56–63 days; Nowell andJackson, 1996b), the kittens died shortly after de-livery from respiratory failure, a condition usu-ally associated with premature delivery.
In domestic cats, serum progesterone levels be-gin a slow decline by day 50 of pregnancy, pro-gressing to a precipitous decrease during the lastweek prior to parturition (Verhage et al., 1976).Although we did not measure serum progester-one levels in pregnant recipients, it seems possi-ble that, since the recipient domestic cats weregestating AWC cloned fetuses, feto-placental en-docrine signaling may have been inadequate. Asa result, progesterone levels may have shown anabnormally early decrease, starting a cascade ofpre-parturient events that resulted in vaginalbleeding. Consequently, with the last three preg-
BIRTH OF AFRICAN WILDCAT CLONED KITTENS 255
5241_e03_p247-258 9/30/04 9:34 AM Page 255

nant recipients, in an effort to reduce perinatalkitten mortality, a synthetic progestagen (al-trenogest) was orally administered. Altrenogesthas been shown to maintain gestation in ovariec-tomized mares and dogs (Hinrichs et al., 1986;Eilts et al., 1994). Since pregnancy was success-fully maintained (until elective Caesarean sec-tion) in cats treated with the synthetic progesta-gen, luteal insufficiency may be implicated in theoccurrence of premature vaginal bleeding. Ma-ternal administration of betamethasone is widelyused to improve fetal lung maturation (Senat,2002; Celik et al., 2002). Recipients administeredboth synthetic progestagen and betamethasoneduring the latter part of gestation produced a to-tal of five live kittens. Thus, while the improve-ment in fetal survival rate cannot be conclusivelyattributed to the treatment combination of pro-gestagen and betamethasone, the present resultsdo provide at least circumstantial evidence oftheir effectiveness.
Placental abnormalities are a major cause ofmortality in cloned animals and likely contributeto a number of perinatal complications in farmanimals (Cibelli et al., 2002). Abnormal placentaehave been reported in cloned mice, cattle andsheep (Shimozawa et al., 2003; Hill et al., 2000;Hartwich et al., 2003) and may be a result of aber-rant gene expression (Suemizu et al., 2003). In ourstudy, histological analysis of placental tissue re-vealed a reduction in thickness and multifocal at-rophy. In fact, some kittens died from hypoxia af-ter placental separation, with respiratory failureas a secondary consequence. Accordingly, it isplausible that placental anomalies/dysfunctionwere either directly or indirectly responsible forthe perinatal deaths of the cloned kittens. To im-prove success rates and reduce neonatal deaths,basic studies are needed to identify factors caus-ing abnormal placentation.
Developmental anomalies occur with a greaterfrequency in cloned animals than in the averagepopulation. Diagnostic pathology of clonedlambs that died after birth revealed severalcloning-related abnormalities, some of whichwere analogous to congenital human diseases(Rhind et al., 2003). For example, four of eightcloned lambs exhibited fetal overgrowth and abody wall defect. These abnormalities are linkedto Beckwith-Wiedemann syndrome (B-WS), a hu-man genetic syndrome characterized by abdom-inal wall defects, visceromegaly, macroglossia,pre- and postnatal overgrowth and neonatal
hypoglycemia. This disorder involves the dereg-ulation of several genes, including insulin-likegrowth factor, type 2 (IGF2) and cyclin-depen-dent kinase inhibitor 1C (CDKN1C; Murrel et al.,2004). The product of IGF2 is a major fetal growthfactor, and is associated with overgrowth in fe-tuses derived from sheep embryos cultured invitro before transfer to recipients (Young et al.,2001). Recently, in humans, an association wasfound between babies produced by in vitro tech-nology and the incidence of B-WS (DeBaun et al.,2003). In our study, a major abnormality found inthree cloned kittens was incomplete closure ofventral body wall musculature with exterioriza-tion of abdominal organs; however, pathology as-sociated with overgrowth was not observed.Likewise, knockout mice for CDKN1C have beenfound to manifest an anterior wall defect but donot exhibit fetal overgrowth or other features ofB-WS (Fitzpatrick et al., 2002). Whether body wallpathology is caused by gene deregulation or ismerely a consequence of another wall anomaly isnot known.
The influence of possible mitochondrial het-eroplasmy and/or a deleterious mtDNA effect onneonatal death of the cloned AWC kittens cannotbe discounted. Indeed, in future studies the pre-cise mitochondrial inheritance pattern in AWCcloned kittens will be evaluated to determine itsinfluence on survival rate.
The effectiveness of our cloning procedure wasshown by the birth of healthy AWC kittens. Thesebirths represent the first wild carnivores to beproduced by NT and further expand the grow-ing list of species in which cloned offspring havebeen produced. Our success may be attributableto improvements in the embryo culture system(Gómez et al., 2003), refinements made to the nu-clear transfer procedure (Gómez et al., 2003), in-creasing the number of embryos transferred, re-ducing the culture interval before transfer andmodification of current protocols successfullyused in other species to manage pregnancy andimprove viability of offspring (Ptak et al., 2002).
While there are major limitations to currentcloning technologies, knowledge about nucleartransfer procedures is advancing rapidly andtechnology is improving. Researchers should beaccessible to the possibilities offered by nucleartransfer as one of the assisted reproduction tech-niques potentially useful for the conservation ofendangered species, which in some cases, may of-fer the prospect of species continuation rather
GÓMEZ ET AL.256
5241_e03_p247-258 9/30/04 9:34 AM Page 256

than extinction. Future research should focus ondefining and understanding the mechanisms re-sponsible for the abnormalities, so that efficiencyis improved and neonatal death rate reduced. Weexpect that advances in understanding the mo-lecular events for reprogramming of the donorgenome will contribute to clarifying the deriva-tion of developmental defects in cloned embryos.
ACKNOWLEDGMENTS
We are grateful to Stella Sullivan for her ex-cellent cat care and hand rearing some of thecloned kittens. In addition, we thank Dr. KarenMiller, Dr. Steve Miller, Barbara Vincent and theAudubon Nature Institute veterinary staff fortheir help with the surgery procedures; JackieCoulon for coordinating and collecting ovariesfrom the veterinary clinics; and the staff and in-terns from the Species Survival Center for the 24h per day cat observations towards the end ofeach pregnancy.
REFERENCES
Celik, C., Acar, A., Cicek, N., et al. (2002). Corticosteroidtreatment for prevention of prematurity complications.Arch. Gynecol. Obstet. 267, 90–94.
Cibelli, J.B., Campbell, K.H., Seidel, G.E., et al. (2002). Thehealth profile of cloned animals. Nat. Biotechnol. 20,13–14.
Colman, A. (2000). Somatic cell nuclear transfer in mam-mals: progress and applications. Cloning 1, 185–200.
Debaun, M.R., Niemitz, E.L., and Feinberg, A.P. (2003).Association of in vitro fertilization with Beckwith-Wiedemann syndrome and epigenetic alterations ofLIT1 and H19. Am. J. Hum. Genet. 72, 156–160.
Eilts, B.E., Paccamonti, D.L., Hosgood, G., et al. (1994).The use of Ally-trenbolone as a progestational agent tomaintain pregnancy in ovariectomized bitches. Theri-ogenology 24, 1237–1245.
Fitzpatrick, G.V., Soloway, P.D., and Higgins, M.J. (2002).Regional loss of imprinting and growth deficiency inmice with a targeted deletion of KvDMR1. Nat. Genet.32, 426–431.
Gómez, M.C., Pope, C.E., Harris, R.F., et al. (2000). Birthsof kittens produced by intracytoplasmic sperm injec-tion of domestic cat oocytes matured in vitro. Reprod.Fertil. Dev. 12, 423–433.
Gómez, M.C., Jenkins, J.A., Giraldo, A., et al. (2003). Nu-clear transfer of synchronized African wildcat somaticcells into enucleated domestic cat oocytes. Biol. Reprod.69, 1032–1041.
Gómez, M.C., Pope, C.E., Harris, R.F., et al. (2003). De-
velopment of in vitro matured, in vitro fertilized do-mestic cat embryos following cryopreservation, cultureand transfer. Theriogenology 60, 239–251.
Hartwich, K.M., Walker, S.K., Jurisevic, A.M., et al. (2003).Evidence of impaired placental function in cloned fetalsheep during late gestation. Theriogenology 59, 256 [ab-str].
Herron, M.A. (1977). Feline reproduction. Vet. Clin. NorthAm. 7, 715–722.
Hill, J.R., Burghardt, R.C., Jones, K., et al. (2000). Evidencefor placental abnormality as the major cause of mor-tality in first-trimester somatic cell cloned bovine fe-tuses. Biol. Reprod. 63, 1787–1794.
Hinrichs, K., Sertich, P.L., and Kenny R.M. (1986). Use ofaltrenogest to prepare ovariectomized mares as embryotransfer recipients. Theriogenology 26, 455–460.
Janssen, D.L., Edwards, M.L., Koster, J.A., et al. (2004).Postnatal management of chryptordich banteng calvescloned by nuclear transfer utilizing frozen fibroblastcultures and enucleated cow ova. Reprod. Fertil. Dev.16, 224(abst).
Kikyo, N., and Wolffe, A.P. (2000). Reprogramming nu-clei: insights from cloning, nuclear transfer and het-erokaryons. J. Cell Sci. 113, 11–20.
Kustritz, R. (2001). Use of supplemental progesterone inmanagement of canine pregnancy. In: Recent Advancesin Small Animal Reproduction. P.W. Concannon, G. En-gland, and J. Verstegen, eds. (Ithaca: International Vet-erinary Information Service), p. A1220.0401.
Lanza, R.P., Cibelli, J.B., Diaz, F., et al. (2000). Cloning ofan endangered species (Bos gaurus) using interspeciesnuclear transfer. Cloning 2, 79–90.
Leese, H.J. (2002). Quiet please, do not disturb: a hy-pothesis of embryo metabolism and viability. BioEssay24, 845–849.
Loi, P., Ptak, G., Fulka, J., Jr., et al. (2001). Genetic rescueof an endangered mammal by cross-species nucleartransfer using post-mortem somatic cells. Nat. Biotech-nol. 19, 962–964.
Luvoni, G.C. (2000). Current progress on assisted repro-duction in dog and cats: in vitro embryo production.Reprod. Nutr. Dev. 40, 505–512.
Miyoshi, K., Rzucidlo, S.J., Pratt, S.L., et al. (2003). Im-provements in cloning efficiencies may be possible byincreasing uniformity in recipient oocyte and donorcells. Biol. Reprod. 68, 1079–1086.
Murrell, A., Heeson, S., Cooper, W.N., et al. (2004). An as-sociation between variants in the IGF2 gene and Beck-with Wiedemann syndrome: interaction between geno-type and epygenotype. Hum. Mol. Genet. 13, 247–255.
Nowell, P., and Jackson, P. (1996a). Cats and habitat loss.In Status Survey and Conservation Action Plan: Wild Cats(Gland, Switzerland, IUCN), pp. 149–179.
Nowell, K., and Jackson, P. (1996b). African wildcatgroup. In Status Survey and Conservation Action Plan:Wild Cats (Gland, Switzerland, IUCN), pp. 32–35.
Polejaeva, I.A., Chen, S.H., Vaught, T.D., et al. (2000).Cloned pigs produced by nuclear transfer from adultsomatic cells. Nature 407, 86–90.
Polge, C., Rowson, L.E., and Chang, M.C. (1966). The ef-
BIRTH OF AFRICAN WILDCAT CLONED KITTENS 257
5241_e03_p247-258 9/30/04 9:34 AM Page 257

fect of reducing the number of embryos during earlystages of gestation on the maintenance of pregnancy inthe pig. J. Reprod. Fertil. 12, 395–397.
Pope, C.E. (2000). Embryo technology in conservation ef-forts for endangered felids. Theriogenology 53, 163–174.
Pope, C.E., Keller, G.L., and Dresser, B.L. (1993). In vitrofertilization in domestic and non-domestic cats includ-ing sequences of early nuclear events, development invitro, cryopreservation and successful intra- and inter-species embryo transfer. J. Reprod. Fertil. 47, 189–201.
Pope, C.E., McRae, M.A., Plair, B.L., et al. (1994). Suc-cessful in vitro and in vivo development of in vitro fer-tilized two- to four-cell cat embryos following cryo-preservation, culture and transfer. Theriogenology 42,513–25.
Pope, C.E., Schmid, R., and Dresser, B.L. (1999). In vitrodevelopment of cat embryos produced by in vitro fer-tilization is enhanced by addition of cysteine to the mat-uration medium and a reduced O2 atmosphere. Theri-ogenology 51, 291(abst).
Pope, C.E., Gómez, M.C., Mikota, S.K., et al. (2000). De-velopment of in vitro–produced African Wildcat (Felissilvestris) embryos after cryopreservation and transferinto domestic cat recipients. Biol. Reprod. 62, 321(abst).
Pope, C.E., Gómez, M.C., King, A.L., et al. (2003). Em-bryos produced in vitro after recovery of oocytes fromcat ovaries stored at 4°C for 24 to 48 hours retain thecompetence to develop into live kittens after transfer torecipients. Theriogenology 59, 308(abst).
Ptak, G., Clinton, M., Tischner, M., et al. (2002). Improv-ing delivery and offspring viability of in vitro–producedand cloned sheep embryos. Biol. Reprod. 67, 1719–1725.
Raymond, M.M., David, V.A., Lyons, L.A., et al. (1999).A genetic linkage map of microsatellites in the domes-tic cat (Felis catus). Genomics 57, 9–23.
Rhind, S.M., King, T.J., Harkness, L.M., et al. (2003).Cloned lambs—lessons from pathology. Nat. Biotech-nol. 21, 744–745.
Seidel, G.E., Jr. (2001). Cloning, transgenesis, and geneticvariance in animals. Cloning Stem Cells 3, 251–256.
Senat, M.V. (2002). Corticosteroid for fetal lung matura-tion: indication and treatment protocols. J. Gynecol. Ob-stet. Biol. Reprod. 31, 672–676.
Shimozawa, N., Tajima, S., Azuma, N., et al. (2003). His-tological study of the hypertrophic placentas and openeyelids observed in cloned fetuses. J. Reprod. Dev. 49,221–226.
Shin, T., Kraemer, D., Pryor, J., et al. (2002). A cat clonedby nuclear transplantation. Nature 415, 859.
Suemizu, H., Aiba, K., Yoshikawa, T., et al. (2003). Ex-pression profiling of placentomegaly associated withnuclear transplantation of mouse ES cells. Dev. Biol.253, 36–53.
Verhage, H.G., Beamer, N.B., and Brenner, R.M. (1976).Plasma levels of estradiol and progesterone in the catduring polyestrus, pregnancy and pseudopregnancy.Biol. Reprod. 14, 579–585.
Walker, S.C., Shin, T., Zaunbrecher, G.M., et al. (2002). Ahighly efficient method for porcine cloning by nucleartransfer using in vitro–matured oocytes. Cloning StemCells 4, 105–112.
Walmsley, S.E., Buckrell, B.C., Buschbeck, C., et al. (2004).Rate of abnormalities in lambs from in vitro–producedembryos transferred on day 2 compared with day 6postfertilization. Theriogenology 62, 195–206.
Wells, D.N., Misica, P.M., Day, T.A., et al. (1997). Pro-duction of cloned lambs from an established embryoniccell line: acomparison between in vivo– and in vitro–ma-tured cytoplasts. Biol. Reprod. 57, 385–393.
Wilmut, I., and Paterson, L. (2003). Somatic cell nucleartransfer. Oncol. Res. 13, 303–307.
Young, L.E., Fernandes, K., McEvoy, T.G., et al. (2001).Epigenetic change in IGF2R is associated with fetalovergrowth after sheep embryo culture. Nat. Genet. 27,153–154.
Address reprint requests to:Dr. Martha C. Gómez
Audubon Center for Research of Endangered Species
New Orleans, LA 70131
E-mail: [email protected]
GÓMEZ ET AL.258
5241_e03_p247-258 9/30/04 9:34 AM Page 258
Related Documents











