ECOGRAPHY 24: 659–670. Copenhagen 2001 Methods for comparison of biotic regionalizations: the case of pteridophytes in the Iberian Peninsula Ana L. Ma ´ rquez, Raimundo Real and J. Mario Vargas Ma ´rquez, A. L., Real, R. and Vargas, J. M. 2001. Method s for compar ison of biotic region alizat ions: the case of pteri dophytes in the Iberian Peninsula . – Ecograph y 24: 659–670. We made several regionalizations of the Iberian Peninsula based on the distributions of the pteridophyte flora to see whether the regionalization depended on the type and sca le of lat tic e or the simil ari ty inde x cons ide red. We use d five types of lat tic e in which the sca le also var ied: river basi ns, mountains and plains , nat ural regions, physiog raphic and geologic al region s, and admini strat ive provinces; and two similar- ity indices: those of Jaccard and of Baroni-Urbani and Buser. The regionalizations varied according to the type of lattice, the grain size, and the similarity index used. To assess the different regionalizations we used four methods: 1) the coefficient of variation of the size of sites in each lattice, 2) the bestblock method, which considers as the best lattice that which maximizes the number of matches between presences ove r all pair wis e sit e compar isons, 3) the Mantel tes t, to ass ess the statis tic al significance of the regionalizations obtained, and 4) mapability, which considers the most contiguou s region alizat ion to be the best. The best regionali zation accordin g to our four criteria was that based on administrative provinces and Jaccard’s index. This yielde d a small central region and three large regions: northern, wester n, and easte rn. A. L. Ma ´ rqu ez ( rrgimenez@uma .es ) , R. Real and J . M . Vargas, Dept de Biol ogı ´a Animal , Fac. de Ciencias, Uni 6. de Ma ´ laga , E -29071 Ma ´ laga, Spain. A maj or obj ect ive of des cri pti ve bio geo graphy is to simplify the complex patterns of contemporary species’ distributions by classifying areas based on their biotic composition. That areas of the world can be character- ized on the basis of the presence of certain taxa and the absence of others has long been recognised by biogeo- gra phe rs (Buffo n 1761), and is a dee ply ent ren che d compon ent of muc h con tempor ary bio geogra phi cal thin king. In addit ion, class ifying geogr aphic al areas into groups with different species composition is valu- abl e for nat ure con servat ion pla nni ng (Br own et al. 1993), so as to ensure that all groups are represented in the selection of natural reserves (Margules 1986), and to evalua te the biolo gic al resources of an area in a regional and global context (Carey et al. 1995). How- ever, currently administrators are often confused by the number of different classifications of their country pro- posed by ecologists and phytogeographers. In addition, as Thaler and Plowright (1973) pointed out, phytogeo- graph ers have disa greed about wheth er floristic areas repres ent real ent iti es in nature or whe the r the y are simp ly conve nient , subj ective constru cts that lack any objective and unambiguous basis. Operat ionall y, it is poss ible to identi fy bi ot ic bou nda ries when a gro up of are as with similar bio ta sha res fewer spe cie s than expect ed at ran dom wit h ot her group of areas wi th si mi lar bi ot a. Then the Operational Geographic Units (OGUs; Crovello 1981) of thi s ter rit ory can be gro upe d int o bio tic reg ion s (Re al et a l. 1 99 2a , Myk l esta d an d Bi rk s 19 93, McLau ghlin 1994). Clus terin g metho ds repr esent one approach to this problem (Birks 1987, Legendre 1990), but onl y whe n a pro bab ili sti c pro ced ure all ows the researcher to distinguish between statistically significant clust ers and those indis tingu ishab le from random ex- pecta tion (Ma ´rquez et al. 1997). Area clust ers may be Accepted 6 March 2001 Copyright © ECOGRAPHY 2001 ISSN 0906-759 0 Print ed in Ireland – all rights res erved ECOGRAPHY 24:6 (2001) 659

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 1/12
ECOGRAPHY 24: 659–670. Copenhagen 2001
Methods for comparison of biotic regionalizations: the case of
pteridophytes in the Iberian Peninsula
Ana L. Marquez, Raimundo Real and J. Mario Vargas
Marquez, A. L., Real, R. and Vargas, J. M. 2001. Methods for comparison of bioticregionalizations: the case of pteridophytes in the Iberian Peninsula. – Ecography 24:659–670.
We made several regionalizations of the Iberian Peninsula based on the distributionsof the pteridophyte flora to see whether the regionalization depended on the type andscale of lattice or the similarity index considered. We used five types of lattice inwhich the scale also varied: river basins, mountains and plains, natural regions,physiographic and geological regions, and administrative provinces; and two similar-ity indices: those of Jaccard and of Baroni-Urbani and Buser. The regionalizationsvaried according to the type of lattice, the grain size, and the similarity index used.To assess the different regionalizations we used four methods: 1) the coefficient of variation of the size of sites in each lattice, 2) the bestblock method, which considersas the best lattice that which maximizes the number of matches between presencesover all pairwise site comparisons, 3) the Mantel test, to assess the statisticalsignificance of the regionalizations obtained, and 4) mapability, which considers themost contiguous regionalization to be the best. The best regionalization according toour four criteria was that based on administrative provinces and Jaccard’s index. Thisyielded a small central region and three large regions: northern, western, and eastern.
A. L. Marquez ( [email protected] ) , R. Real and J . M . Vargas, Dept de BiologıaAnimal , Fac. de Ciencias, Uni 6. de Malaga, E -29071 Malaga, Spain.
A major objective of descriptive biogeography is to
simplify the complex patterns of contemporary species’
distributions by classifying areas based on their biotic
composition. That areas of the world can be character-
ized on the basis of the presence of certain taxa and the
absence of others has long been recognised by biogeo-graphers (Buffon 1761), and is a deeply entrenched
component of much contemporary biogeographical
thinking. In addition, classifying geographical areas
into groups with different species composition is valu-
able for nature conservation planning (Brown et al.
1993), so as to ensure that all groups are represented in
the selection of natural reserves (Margules 1986), and
to evaluate the biological resources of an area in a
regional and global context (Carey et al. 1995). How-
ever, currently administrators are often confused by the
number of different classifications of their country pro-
posed by ecologists and phytogeographers. In addition,
as Thaler and Plowright (1973) pointed out, phytogeo-
graphers have disagreed about whether floristic areas
represent real entities in nature or whether they are
simply convenient, subjective constructs that lack any
objective and unambiguous basis.
Operationally, it is possible to identify bioticboundaries when a group of areas with similar biota
shares fewer species than expected at random with
other group of areas with similar biota. Then the
Operational Geographic Units (OGUs; Crovello 1981)
of this territory can be grouped into biotic regions
(Real et al. 1992a, Myklestad and Birks 1993,
McLaughlin 1994). Clustering methods represent one
approach to this problem (Birks 1987, Legendre 1990),
but only when a probabilistic procedure allows the
researcher to distinguish between statistically significant
clusters and those indistinguishable from random ex-
pectation (Marquez et al. 1997). Area clusters may be
Accepted 6 March 2001
Copyright © ECOGRAPHY 2001ISSN 0906-7590Printed in Ireland – all rights reserved
ECOGRAPHY 24:6 (2001) 659

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 2/12
considered as biotic regions when a group of OGUs is
statistically significant. Legendre (1987) advocated con-
straining the clustering to respect spatial contiguities.
However, in some cases, biotic regions may reflect a
climatic or ecological pattern that may be not strictly
contiguous in space, and then they may be better
detected with clusters only defined by the similarity of
their species composition. Then, area clusters can be
related to environmental variables by techniques such
as regression or canonical variates analysis (Carey et al.
1995, Olivero et al. 1998) to investigate possible causes
of the regionalization pattern.
However, to analyse biogeographic patterns, any ter-
ritory must be subdivided initially into different types
of OGUs (Crovello 1981, Kenkel et al. 1989, Mandrak
1995). This establishes a lattice that influences the bio-
geographic regions subsequently obtained, because any
biotic boundary necessarily has to follow along the
edges of the sites used to partition the study area.
Cressie (1993) defined a lattice as a countable collection
of spatial sites that are spatially regular or irregular,
represent points or regions, and are indices for continu-
ous or discrete variables. In biogeographic regionaliza-
tion the concept of lattice corresponds to any kind of
area subdivision, irrespective of its size and the criteria
used to define it, and the sites correspond to the OGUs.
The lattices can be natural (e.g., watersheds, Vargas etal. 1998), arbitrary (e.g., latitude-longitude cells, Schall
and Pianka 1977, or equal-area quadrats, Marquez et
al. 1997). The sites can have the same size when the
lattice is regular, such as those based on grid-squares,
or different sizes when the lattice is irregular, such as
those made according to, for example, physical, geolo-
gical, or political viewpoints. Variation in size and
shape may affect the species richness of each site and
hence the results of the subsequent regionalization.
Square equal-area quadrats are considered not to be
subject to these problems as quadrat size and shape are
uniform (Mandrak 1995), although coastal quadrats
may vary in area when terrestrial organisms are stud-
ied, and mountainous areas have more surface areathan flat areas in a ‘‘equal-area quadrat’’.
There are also problems in defining the scale of the
grain used (Jardine 1972, Birks 1987, Carey et al. 1995).
Scale is dependent on both the extent of the study area
and the size of organism studied (Wiens 1989). If too
coarse a lattice is used, biotic regions will be corre-
spondingly coarse. Conversely, the use of too fine a
lattice may fail to reveal major biotic regions and
generate many spurious small regions where recordings
may have been uneven. Results obtained using small
sites may be different from those obtained using larger
sites, although these different patterns might be interre-
lated (Blondel 1985, Vargas 1993). However, this does
not mean that there are no criteria for comparing theresults obtained using different types of sites or differ-
ent scales (Levin 1992). These criteria, however, need
proper definition and testing.
Ferns could provide a valuable source of biogeo-
graphical information that is distinct from, or at least
complementary to, that provided by other types of
plant (Marquez et al. 1997). Pteridophytes are a group
of species with a great dispersal capacity due to the
lightness of their spores (Jermy 1984). However, most
fern species are restricted in distribution and only a
limited number have wide ranges (Kato and Iwatsuki
1983), because their distribution is highly related to
climate and substratum (Barrington 1993, Given 1993,
Kornas 1993). The 113 Iberian pteridophyte species are
well recorded and well understood taxonomically in theIberian Peninsula (Salvo et al. 1984, Castroviejo et al.
1986), and are an important number of species in a
European context.
In this paper we have used several criteria to select
the best biotic regionalization among several partitions
obtained using a quantitative method of classification.
With this aim, we have used fern distributions in the
Iberian Peninsula a) to establish whether the Iberian
Peninsula may be divided into pteridogeographic re-
gions or whether the pteridophyte species distributions
follow a mainly gradual pattern; b) to verify whether
the resulting regionalization patterns are different ac-
cording to the type of lattice, the number of sites
considered, and the similarity index used; and c) toapply several criteria to select the best regionalization
pattern.
Material and methods
Study area
The Iberian Peninsula is situated on the south-western
end of Europe, and presents a stronger peninsular
condition than other large Mediterranean peninsulas
because its isthmus is relatively narrow. In addition, the
presence of the Pyrenees in the isthmus hinders the
spread of propagules between the Iberian Peninsula andthe rest of the European continent. The interchange of
biota with Africa is also hindered by the Straits of
Gibraltar. Climatically, the Iberian Peninsula is placed
in the southern band of the temperate zone.
Species distribution and Operational Geographic
Units (OGUs)
The distribution of the 113 species of pteridophytes
were obtained from Salvo et al. (1984), supplemented
with new records published later.
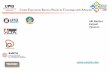
We considered five types of OGUs in the Iberian
Peninsula: river basins, mountains and plains, naturalregions, physiographic and geological regions, and ad-
ministrative provinces (Anon. 1980) (Fig. 1). The
province lattice is political-administrative, although is
660 ECOGRAPHY 24:6 (2001)

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 3/12
partially based on natural partitions, whereas the other
four lattices have natural borders. Natural regions and
physiographic and geological regions are partially
based on vegetation, and so some circular reasoning
may be involved in their use as phytogeographic re-
gions (Birks 1987, Real and Ramırez 1992, Vargas
1993).
Classification analysis
Initially we constructed a presence-absence matrix forthe 113 species in each lattice considered in the area.
We thus had five presence-absence matrices, in which
OGU size and number varied: river basins (11×113),
mountains and plains (12×113), natural regions (17×
113), physiographical regions (22×113), and adminis-
trative provinces (56×113).
We applied two similarity indices to each pair of
OGUs: Jaccard’s (1908) index and Baroni-Urbani and
Buser’s (1976). The Jaccard index considers only double
presences, whereas the Baroni-Urbani and Buser index
incorporates double absences. Jaccard values may be
considered as absolute measures of biotic similarity,
because they are not affected by other species absent
from the two OGUs compared, while the Baroni-Ur-
bani and Buser coef ficient measures similarity in thecontext of the Iberian Peninsula, because species absent
from the two OGUs but present in other Iberian OGUs
increase the similarity between the two OGUs analysed
Fig. 1. The five differentlattices considered in the study(Anon. 1980).
ECOGRAPHY 24:6 (2001) 661

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 4/12
(Real et al. 1992b). However, the possibility that two
OGUs are considered similar exclusively because of
double absences is avoided in the Baroni-Urbani and
Buser index by multiplying double absences by double
presences.
We used UPGMA (Unweighted Pair-Group
Method using Arithmetic Averages) as our clustering
method (Sneath and Sokal 1973). Agglomerative tech-
niques are preferred to divisive techniques because the
former maximizes the similarity within a group, which
defines biotic regions. We represented the results as
dendrograms.To test the groups obtained in any association
analysis several authors use a null model based on a
Monte Carlo randomization procedure (Connor and
Simberloff 1979, Oden et al. 1993). However, this
model uses the observed data matrix to generate a
null distribution, and so observed and null distribu-
tions lack statistical independence (Grant and Abbot
1980). Notwithstanding, it is feasible to determine all
the possible distributions of N species in any of the
OGUs compared for each value of N, and so an
exact randomization test (Sokal and Rohlf 1981, p.
788) can be performed to determine whether an ob-
served value of the Jaccard index or the Baroni-Ur-
bani and Buser index is significantly different fromthose expected at random. We obtained the statistical
significance of the observed similarity values, accord-
ing to the exact randomization tests in Real and Var-
gas (1996) for the Jaccard index, and in
Baroni-Urbani and Buser (1976) for the Baroni-Ur-
bani and Buser index. In this way, the similarity ma-
trices were transformed into matrices of statistical
significance (Marquez et al. 1997) that consisted of
pluses, for similarity values significantly higher than
those expected at random, minuses, for similarity val-
ues significantly lower than those expected at random,
and zeros, for similarity values compatible with ran-
dom expectation.
Our null hypothesis is that biotic boundaries donot exist, so that we started searching for boundaries
at the level of the dendrogram with the lowest simi-
larity value, and then moved to the higher similarity
nodes, finishing when no more significant boundaries
appeared. This divisive procedure does not contradict
the use of an agglomerative classification, because sig-
nificant boundaries divide the space into biotic re-
gions that require not only the existence of a biotic
boundary, but also a biotic af finity between OGUs in
the region, which is favoured by the agglomerative
method.
We tested at every node of the dendrogram for the
existence of significant biotic boundaries between the
OGUs separated by the node using the method of McCoy et al. (1986), as modified by Real et al.
(1992b) for application to dendrograms, as indicated
in Appendix 1.
To determine which lattice and similarity index best
define the biotic regions we used four methods: 1)
Coef ficient of variation of the OGU’s size in each
lattice (Sokal and Rohlf 1981). Differences in the size
of the OGUs within a lattice produce area-related
distortions, such as differences in the number of spe-
cies in different parts of the study area. So, the lattice
with the lowest coef ficient of variation of the OGUs’
size is considered the best one. This method selects
the best lattice regardless of the distribution data. 2)
Bestblock (Phipps 1975), which considers the best lat-
tice to be that which maximizes the number of matches between presences over all pairwise OGU
comparisons. This method selects the best lattice after
taking into account the distribution data regardless of
the regionalization obtained. 3) We used the Mantel
test of matrix comparison (Mantel 1967) to compare
each original similarity matrix with each matrix of
hypothetical similarities according to the resulting re-
gionalizations. The matrices of hypothetical similari-
ties were obtained by considering all the similarities
within a biotic region to be 1, all the similarities be-
tween OGUs separated by weak boundaries to be the
random expectation (0.33 for Jaccard’s index and 0.5
for Baroni-Urbani and Buser’s index), and all thesimilarities between OGUs separated by strong
boundaries to be 0. Only significant regionalizations
were considered. The most significant regionalization
is that which is most consistent with the original simi-
larities and is thus considered the best. 4) Mapability
(Harper 1978). Contiguity is preferred to discontiguity
of the biotic regions obtained using each similarity
index in each lattice. We measured the mapability in
terms of the minimum number of quadrats UTM
50×50 km necessary to join together the parts of all
fragmented regions. The lower this number of
quadrats is, the more mapable the regionalization is.
We applied the two former criteria to the five lat-
tices and the two latter criteria to the ten regionaliza-tions resulting from using each similarity index in
each lattice.
Results
We found significant biotic boundaries for all types
of OGU using both similarity indices. All the region-
alization patterns obtained using the Jaccard index
(Table 1) and most of the regionalizations obtained
using Baroni-Urbani and Buser’s index (Table 2) were
statistically significant according to the Mantel test of
matrix comparison (Table 3). Only the regionalizationobtained using mountains and plains and the Baroni-
Urbani and Buser index was found to be non-signifi-
cant (p\0.01) using the Mantel test.
662 ECOGRAPHY 24:6 (2001)

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 5/12
Table 1. Statistically significant boundaries between groups of OGUs formed by UPGMA using the Jaccard similarity index.DW\0 and significant GW indicate weak boundary between the groups; DS\0 and significant GS indicate a strong boundarybetween the groups. N.S.: p\0.05; *: pB0.05; **: pB0.01; ***: pB0.005.
Biotic boundaryGroups set up byUPGMA
StrongWeak
Group Group DW DW DW GW p DS GS pA B (A×A) (B×B)
River basins 3 –5 9 –8 0.206 0.168 N.S0.187 9.074 *** −0.501 0Mountains and plains 2 –9 4 0.562 0.627 0.595 20.249 *** −0.079 0 N.S.
2 –5 10 –9 0.153 0.153 N.S.0.153 8.573 *** −0.555 0Natural regions 2 –13 4 –9 0.219 N.S.0.209 0.214 24.756 *** −0.457 0
Physiographical regions 7 –2 13 –14 0.372 0.571 0.472 28.758 *** −0.099 0.781 N.S.7 –17 4 –2 0.239 0.280 N.S.0.259 28.422 *** −0.349 0.606
N.S.7 –16 20 –17 0.201 0.222 0.211 16.107 *** −0.486 0Administrative 16 –54 27 0.189 0.707 0.448 27.311 *** 0.183 19.153 ***provinces 16 –40 14 –54 0.439 0.356 ***0.397 208.640 *** 0.011 80.326
14 –24 49 –54 0.387 ***0.348 0.367 176.987 *** −0.049 18.98649 –43 37 –54 0.522 0.592 0.557 45.891 *** −0.033 0 N.S.14 –12 23 –24 0.254 0.191 N.S.0.222 38.099 *** −0.351 1.365
N.S.14 –9 12 0.320 0.392 0.356 8.727 *** −0.316 023 –33 21 –10 0.407 0.389 0.398 13.508 *** −0.300 0 N.S.14 –15 20 –9 0.202 0.162 N.S.0.182 6.812 ** −0.505 0
N.S.37 –36 42 –54 0.126 0.126 0.126 6.328 * 0.581 020 –29 2 –9 0.080 0.080 0.080 5.555 N.S.* −0.627 0
Table 2. Statistical significant boundaries between groups of OGUs formed by UPGMA using the Baroni-Urbani and Buser
similarity index. DW\0 and significant GW indicate weak boundary between the groups; DS\0 and significant GS indicatea strong boundary between the groups. N.S.: p\0.05; *: pB0.05; **: pB0.01; ***: pB0.005.
Groups set up by Biotic boundaryUPGMA
StrongWeak
Group Group DW DW DW GW p DS GS pA B (A×A) (B×B)
River basins 3 –5 9 –8 0.116 0.116 0.116 6.98 ** −0.591 0 N.SMou ntains and p lain s 2 –4 1 –5 0.307 0.307 0.307 24.015 *** −0.400 0 N.SNatural regions 6 –14 5 –11 0.114 0.114 N.S.0.114 22.100 *** −0.593 0Physiographical regions 7 –14 4 –2 0.140 0.147 N.S.0.144 43.864 *** −0.551 0.007Administrative 16 –12 27 0.513 – 0.575 94.044 *** 0.089 28.972 N.S.
16 –40 49 –12 0.193provinces 0.008 ***0.10 1 16 1.90 9 ** * −0.456 23.21449 –54 20 –12 0.096 0.091 0.093 89.621 *** −0.611 0 N.S.20 –24 12 0.099 0.099 N.S.0.099 9.930 *** −0.608 0
16 –56 N.S.18 –40 0.062 0.072 0.067 9.620 *** −0.635 0
Table 3. Results of the application of the four criteria used to determine which is the best regionalization. CV: Coef ficient of variation of the OGUs’ size; TM: Total matches between presences in all pair-wise comparisons; SQ: Number of squares 50×50km necessary to join together each fragmented region.
Mantel testOGU Systems Coef ficient Best Mapabilityblockof variation
JaccardJaccard Baroni-Urbani Baroni-Urbaniand Buser and Buser
SQCV TM z p(z obsBz exp) z p(z obsBz exp ) S Q
River basins 0.481 20 983 10.17 0.002 5.33 0.002 0 0Mountains and plains 0.445 21 754 13.72 00.001 3.91 0.014 0
Natural regions 0.360 26 885 24.92 0.001 12.36 0.001 00Physiographical regions 0.447 28 977 60.75 00.001 20.06 0.001 1Administrative 0 30.359 47 881 544.9 0.001 212.9 0.001
provinces
ECOGRAPHY 24:6 (2001) 663

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 6/12
Area classification using the Jaccard similarityindex
Figure 2 shows the biotic boundaries found using each
lattice and the Jaccard similarity index. The lattices of
river basins and natural regions present only a weak
biotic boundary that divides the Iberian Peninsula into
a northern and a southern region, whereas two or more
boundaries are obtained using the other lattices (Fig. 2,
Table 1).
In the administrative provinces lattice, three strong
boundaries divide the Iberian Peninsula into four
supraregions: northern supraregion, western suprare-gion, eastern supraregion, and a small central suprare-
gion. The western and eastern supraregions are divided
by weak biotic boundaries into four and three regions,
respectively (Fig. 2).
Area classification using the Baroni-Urbani andBuser similarity index
Figure 3 shows the biotic boundaries found using
each lattice and the Baroni-Urbani and Buser index.
In four lattices (river basins, mountains and plains,
natural regions, and physiographical regions) there are
only weak biotic boundaries (Table 2) that divide theIberian Peninsula into northern and southern regions,
although using mountains and plains the regionaliza-
tion was not statistically significant (Table 3). However,
using the provinces there are three big regions
(northern, western, and eastern) and three small regions
(Fig. 3).
Which lattice defines best the biotic regions?
The administrative provinces have the lowest coef ficient
of variation for the size of the sites (Table 3). That
means that using the provinces the Iberian territory is
more evenly distributed than using the other lattices.
According to the bestblock method, the administrative
provinces lattice maximizes the number of positive
matches over all pairwise species comparisons (Table
3). The significance values associated with the Mantel
text are lower for river basins and nonsignificant for
mountains and plains when using the Baroni-Urbani
and Buser’s index, whereas the other regionalizationsall have the same high probability value (Table 3). The
regions obtained from physiographical regions using
the Jaccard index and from administrative provinces
using the Baroni-Urbani and Buser index are frag-
mented (Table 3, Figs 2 and 3), whereas the other
lattices yield continuous regions.
Therefore, the only regionalization that obtains the
best scores according to the four criteria is the one
obtained using administrative provinces and the Jac-
card index (Fig. 2).
DiscussionAlthough there is no absolute regionalization of the
Iberian Peninsula according to the distribution of pteri-
dophytes, the concept of the biotic region is not an
useless artefact. Regionalizations vary according to the
type of OGU, the grain size, and the similarity index
used. In this way, any regionalization must indicate
Fig. 2.
664 ECOGRAPHY 24:6 (2001)

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 7/12
Fig. 2. Dendrograms andbiotic regions forpteridophytes in the IberianPeninsula using the Jaccardsimilarity index on eachlattice considered. Numbers of the units as in Fig. 1. W:weak boundary; S: strongboundary; *: pB0.05;**: pB0.01; ***: pB0.005.
those characteristics, which are inseparable from the
pattern obtained. This is especially true given the ex-
ploratory nature of current approaches to biotic re-
gionalization (Carey et al. 1995). However, the
biogeographic regions and boundaries are useful for
understanding the mechanisms underlying the distribu-tion of organisms, provided that each regionalization
reflects a distribution pattern that is statistically signifi-
cant and may be related to environmental conditions,
although detected through the filters of the OGUs, the
scale of grain, and the similarity index.
The indices of similarity
Oden et al. (1993) modified the method of Womble
(1951) to detect intervals of marked change in categori-
cal variables between contiguous sites. When the vari-
ECOGRAPHY 24:6 (2001) 665

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 8/12
able analysed is the presence or absence of species this
method could be used to obtain biotic boundaries and
regions. However, the basic statistic that results from
the method of Oden et al. (1993) is the complement of
the simple matching coef ficient, which is not recom-
mended for biogeographical regionalizations, since re-
gions may be defined only on the basis of shared
absences, even with no single species in common (Ba-
roni-Urbani and Buser 1976).
We opted for using the Jaccard and Baroni-Urbani
and Buser similarity indices to estimate the associations
between the OGUs and, later, to test the statisticalsignificance of such associations, because they are com-
monly used and are somewhat complementary. The
Jaccard index only takes into account the double pres-
ences of species in the two OGUs compared, and the
similarity between two OGUs is not affected by other
species present in other OGUs in the study area. The
Baroni-Urbani and Buser index takes into account dou-
ble absences, which offers potentially useful informa-
tion because if a species is absent from two OGUs, it
may be due to ecological or historical factors impacting
in a similar way on the species in the context of the area
studied. If the pattern obtained considering the similar-
ities between each pair of OGUs only (using the Jac-
card index) is maintained when the similarities take intoaccount all the study area (using the Baroni-Urbani and
Buser index), then the lattice used may be considered
suitable for revealing a consistent distribution pattern
of the species.
The regionalizations obtained using the two indices
are identical when based on river basins and on natural
regions, partially similar using physiographical regions
and provinces, and very different using mountains and
plains. The differences seem to be more related to the
type of OGU than to the scale of grain. It may be that
when using river basins and natural regions the distri-
bution of pteridophytes is very well defined, because the
double absences do not modify the overall relationship
pattern obtained using only the presences, whereas
mountains and plains are very unsuitable units for
recording the distribution of pteridophytes, so that
double absences greatly alter the relationship pattern
between the OGUs, with physiographical regions and
provinces in an intermediate position.The number of boundaries obtained using the Ba-
roni-Urbani and Buser coef ficient tends to be lower
than that obtained using the Jaccard index, probably
because the double absences considered in the Baroni-
Urbani and Buser index tend to homogenize the re-
gions. This may be seen especially in the south, that has
a lower number of species than in the north. Perhaps
this tendency should be taken into account when a
researcher decides which index to use to reveal a re-
gionalization pattern, according to the underlying
knowledge and assumptions.
The criteria to select the best regionalization
Even when using an numerical method of classification
and a probabilistic procedure to assess the groups of
OGUs obtained, several regionalizations may result
from the use of different types of OGUs or different
similarity indices, so that several criteria are needed to
compare the suitability of the regionalizations obtained.
Fig. 3.
666 ECOGRAPHY 24:6 (2001)

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 9/12
Fig. 3. Dendrograms and bioticregions for pteridophytes in theIberian Peninsula using theBaroni-Urbani and Busersimilarity index on each latticeconsidered. Numbers of theunits as in Fig. 1. W: weakboundary; S: strong boundary;*: pB0.05; **: pB0.01;***: pB0.005.
We used two criteria prior to the classification proce-
dure, and two criteria after the classification. The co-
ef ficient of variation (Sokal and Rohlf 1981) indicated
that the size of each OGU in the provinces is more
similar than in the other lattices and is thus to be
preferred independently of the distribution data. The
‘‘bestblock’’ method (Phipps 1975) showed that admin-
istrative provinces provide the best lattice for establish-ing the biotic boundaries of the Iberian Peninsula
according to its pteridoflora (Table 3), because the finer
scale of the partition produced more units, and because
the presences are more equitably distributed through
the OGUs, thus producing more matches of presences
in pairwise comparisons. The regionalization obtained
using provinces and the Jaccard index was one of five
regionalizations that were suitable according to both
the Mantel test and the mapability criterion, because it
had the highest statistical significance and was totally
mapable. Mantel and mapability tests were appliedto the final regionalization patterns, and showed
whether the patterns were in accordance with the origi-
nal similarities between OGUs and with the within-
ECOGRAPHY 24:6 (2001) 667

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 10/12
region continuity recommendable for a biotic region,
respectively.
The higher number of units in the provinces lattice is
not the only cause of this partitioning being selected.
These four criteria are not biased towards partitionings
with a large number of units nor more isodiametrically-
shaped units. The coef ficient of variation selects a parti-
tioning of more equal-sized units, but not more circular
in shape, and irrespective of the number of units. The
Phipps algorithm not always chooses the partitioning
with the greatest number of units (see Phipps 1975).
Although the provinces lattice has more units it is notmore likely to produce statistically significant results
after using Mantel’s test, and, in fact, the partitioning
obtained using provinces has the same high significance
level than other partitionings. As regards the criterion
of mapability, a partitioning based on more units is
more likely to produce fragmented regions, although
this is not the case for provinces when using Jaccard’s
index.
Administrative provinces as operational geographicunits
In the Iberian Peninsula, provinces provide the latticethat yields the most appropriate regionalization accord-
ing to our four statistical criteria when using Jaccard’s
index, and is thus the regionalization to be preferred in
attempting to identify the biogeographical processes
affecting the distribution of pteridophytes. One reason
for this may be that provinces are administrative units
of ancient origin and are not completely arbitrary. This
is likely to be true elsewhere, at least in Europe, where
administrative units are partially based on natural par-
titions perceived by local people, as is the case, for
example, with vice-counties in the British Isles (Baroni-
Urbani and Collingwood 1976).
Different types of arbitrary areas within a study
region can be used, such as countries, counties,provinces, river basins, latitude/longitude blocks, or
equal size U.T.M. squares (Birks 1987). In the last
decade regular geographical units, like networks of
regular squares, have been commonly used (e.g. Mart ın
and Gurrea 1990, Lausini and Nimis 1991, Myklestad
and Birks 1993, Carey et al. 1995, Moreno-Saiz et al.
1998). However, regular squares have artificial borders
and, therefore, may include several fragments of differ-
ent geographic or environmental units, so hindering the
search for biogeographical processes that may only act
on natural units (Palomo and Antunez 1992). The
division in provinces is also arbitrary, but the adminis-
trative borders between provinces follow, in part, natu-
ral boundaries such as geographical ranges, so that theymay be more appropriate to assess the environmental
explanations for species distributions. In addition,
provinces may be a suitable type of OGU for analysing
human influence on natural distributions, because in
most countries human activities follow political-admin-
istrative borders.
Acknowledgements – We thank H. J. B. Birks, S. P. McLaugl-hin and P. L. Nimis for revisions of the manuscript and theirvaluable suggestions. We are also grateful to M. A. Rendon, J.C. Guerrero and J. Olivero for their help with the statisticalprocedures.
References
Anon. 1980. Divisiones territoriales de Espana. – CEOMAT,Madrid.
Baroni-Urbani, C. and Buser, M. W. 1976. Similarity of binary data. – Syst. Zool. 25: 251 – 259.
Baroni-Urbani, C. and Collingwood, C. A. 1976. A numericalanalysis of the distribution of British Formicidae (Hy-menoptera, Aculeata). – Verhandl. Naturf. Ges. Basel 85:51 – 91.
Barrington, D. 1993. Ecological and historical factors in fernbiogeography. – J. Biogeogr. 20: 275 – 280.
Birks, H. J. B. 1987. Recent methodological development inquantitative descriptive biogeography. – Ann. Zool. Fenn.24: 165 – 178.
Blondel, J. 1985. Biogeographie. Approche ecologique et evo-lutive. – Masson.
Brown, A., Horsfield, D. and Thompson, D. B. A. 1993. Anew biogeographical classification of the Scottish Uplands.I. Descriptions of vegetation blocks and their spatial varia-
tion. – J. Ecol. 81: 207 – 230.Buffon, G. L. L. 1761. Histoire Naturelle, Generale et Parti-
culiere. Vol. 9. – Imprimerie Royale, Paris.Carey, P. D. et al. 1995. An environmentally defined biogeo-
graphical zonation of Scotland designed to reflect speciesdistributions. – J. Ecol. 83: 833 – 845.
Castroviejo, S. et al. 1986. Flora Iberica. Plantas vasculares dela Penınsula Iberica y Baleares. – Serv. Publ. C.S.I.C.,Madrid, pp. 2 – 158.
Connor, E. F. and Simberloff, D. 1979. The assembly of species communities: chance or competition? – Ecology601: 1132 – 1140.
Cressie, N. A. C. 1993. Statistics for spatial data. – JohnWiley.
Crovello, T. J. 1981. Quantitative biogeography: an overview. – Taxon 30: 563 – 575.
Given, D. R. 1993. Changing aspects of endemism and endan-germent in Pteridophyta. – J. Biogeogr. 20: 293 – 302.
Grant, P. R. and Abbot, I. 1980. Interspecific competition,island biogeography and null hypotheses. – Evolution 34:332 – 341.
Harper, C. W. Jr 1978. Groupings by locality in communityecology and paleoecology: tests of significance. – Lethaia11: 251 – 257.
Jaccard, P. 1908. Nouvelles recherches sur la distributionflorale. – Bull. Soc. Vaud. Sci. Nat. 44: 223 – 270.
Jardine, N. 1972. Computational methods in the study of plant distributions. – In: Valentine, D. H. (ed.), Taxon-omy, phytogeography and evolution. Academic Press, pp.381 – 393.
Jermy, A. C. 1984. Origin and distribution of Pteridophytes inthe Mediterranean area. – Webbia 38: 397 – 416.
Kato, M. and Iwatsuki, K. 1983. Phytogeographic relation-ships of pteridophytes between temperate North Americaand Japan. – Ann. Miss. Bot. Gard. 70: 724 – 733.
Kenkel, N. C., Juhasz-Nagy, P. and Podani, J. 1989. On
sampling procedures in population and community ecol-ogy. – Vegetatio 83: 195 – 207.Kornas, J. 1993. The significance of historical factors and
ecological preference in the distribution of African pterio-phytes. – J. Biogeogr. 20: 281 – 286.
668 ECOGRAPHY 24:6 (2001)

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 11/12
Lausini, D. and Nimis, P. L. 1991. Ecological phytogeographyof the southern Yukon territory (Canada). – In: Nimis, P.L. and Crovello, T. J. (eds), Quantitative approaches tophytogeography. Kluwer Academic Publ., pp. 35 – 122.
Legendre, P. 1987. Constrained clustering. – In: Legendre, P.and Legendre, L. (eds), Developments in numerical ecol-ogy. Springer, pp. 289 – 307.
Legendre, P. 1990. Quantitative methods and biogeographicanalysis. Evolutionary biogeography of the marine algae of the North Atlantic. – In: Garbary, D. J. and South, G. R.(eds), NATO ASI Series. Vol. G22. Springer, pp. 9 – 34.
Levin, S. A. 1992. The problem of pattern and scale inecology. – Ecology 73: 1943 – 1967.
Mandrak, N. E. 1995. Biogeographic patterns of fish species
richness in Ontario lakes in relation to historical andenvironmental factors. – Can. J. Fish. Aquat. Sci. 52:1462 – 1474.
Mantel, N. 1967. The detection of disease clustering and ageneralized regression approach. – Cancer Res. 27: 209 –220.
Margules, C. R. 1986. Conservation evaluation in practice. –In: Usher, M. B. (ed.), Wildlife conservation evaluation.Chapman and Hall, pp. 298 – 314.
Marquez, A. L. et al. 1997. On identifying common distribu-tion patterns and their causal factors, a probabilisticmethod applied to pteridophytes in the Iberian Peninsula. – J. Biogeogr. 24: 613 – 631.
Martın, J. and Gurrea, P. 1990. The peninsular effect in theIberian butterflies (Lepidoptera: Papilionoidea and Hespe-rioidea). – J. Biogeogr. 17: 85 – 96.
McCoy, E. D., Bell, S. S. and Walters, K. 1986. Identifyingbiotic boundaries along environmental gradients. – Ecol-
ogy 67: 749 – 759.McLaughlin, S. P. 1994. Floristic plant geography: the classifi-
cation of floristic areas and floristic elements. – Progr.Phys. Geogr. 18: 185 – 208.
Moreno-Saiz, J. C., Castro-Parga, I. and Sainz-Ollero, H.1998. Numerical analyses of distributions of Iberian andBalearic endemic monocotyledons. – J. Biogeogr. 25: 179 –194.
Myklestad, A. and Birks, H. J. B. 1993. A numerical analysisof the distribution patterns of Salix L. species in Europe. –J. Biogeogr. 20: 1 – 32.
Oden, N. L. et al. 1993. Categorical wombling: detectingregions of significant change in spatially located categoricalvariables. – Geographical Anal. 25: 315 – 336.
Olivero, J., Real, R. and Vargas, J. M. 1998. Distribution of breeding, wintering, and resident water-birds in Europe:biotic regions and the macroclimate. – Ornis Fenn. 75:153 – 175.
Palomo, L. J. and Antunez, A. 1992. Los atlas de distribucionde especies. – In: Vargas, J. M., Real, R. and Antunez, A.(eds), Objetivos y metodos biogeograficos. Aplicaciones enherpetologıa. Monogr. Herpetol. 2. Valencia, pp. 39 – 50.
Phipps, J. B. 1975. Bestblock: optimizing grid size in biogeo-graphic studies. – Can. J. Bot. 53: 1447 – 1452.
Real, R. and Ramırez, J. M. 1992. Fundamento filosofico delos objetivos y metodos de la Biogeograf ıa. – In: Vargas, J.M., Real, R. and Antunez, A. (eds), Objetivos y metodosbiogeograficos. Aplicaciones en herpetologıa. Monogr.Herpetol. 2. Valencia, pp. 221 – 230.
Real, R. and Vargas, J. M. 1996. The probabilistic basis of Jaccard’s index of similarity. – Syst. Biol. 45: 380 – 385.
Real, R., Guerrero, J. C. and Ramırez, J. M. 1992a. Identifica-cion de fronteras bioticas significativas para los anfibios enla cuenca hidrografica del Sur de Espana. – Donana ActaVertebrata 19: 53 – 70.
Real, R., Vargas, J. M. and Guerrero, J. C. 1992b. Analisisbiogeografico de clasificacion de areas y de especies. – In:Vargas, J. M., Real, R. and Antunez, A. (eds), Objetivos ymetodos biogeograficos. Aplicaciones en herpetologıa.Monogr. Herpetol. 2. Valencia, pp. 73 – 84.
Salvo, A. E., Cabezudo, B. and Espana, L. 1984. Atlas de lapteridoflora Iberica y Balear. – Acta Bot. Malacit. 9:105 – 129.
Schall, J. J. and Pianka, E. R. 1977. Species densities of reptiles and amphibians on the Iberian Peninsula. – Do-nana Acta Vertebrata 4: 27 – 34.
Sneath, P. H. A. and Sokal, R. R. 1973. Numerical taxonomy.The principles and practices of numerical classification. –Freeman.
Sokal, R. R. and Rohlf, F. J. 1981. Biometry. The principlesand practice of statistics in biological research. – Freeman.
Thaler, G. R. and Plowright, R. C. 1973. An examination of the floristic zone concept with special reference to thenorthern limit of the Carolinian zone in southern Ontario. – Can. J. Bot. 51: 1765 – 1789.
Vargas, J. M. 1993. Siete pecados capitales en Biogeograf ıa. –Zool. Baetica 4: 39 – 56.
Vargas, J. M., Real, R. and Guerrero, J. C. 1998. Biogeo-graphical regions of the Iberian Peninsula based on fresh-water fish and amphibian distributions. – Ecography 21:371 – 382.
Wiens, J. A. 1989. Spatial scaling in ecology. – Funct. Ecol. 3:385 – 397.
Harper, W. H. 1978. Differential systematics. – Science 114:315 – 322.
Appendix 1. Calculation of parameters used to test theexistence of significant biotic boundaries.
For each dendrogram node we established a submatrix of significant similarities that only included the two groups of
OGUs separated by that node, which we named group A andgroup B, respectively. This submatrix was divided into threezones: zone A×A and zone B×B, which corresponded to thesignificant similarities between OGUs of group A and betweenOGUs of group B, respectively; and zone A×B, correspond-ing to the significant similarities between the two groups of OGUs.
We call Pp(A×A) the number of pluses within zone A×Adivided by the total number of pairwise OGU comparisons inzone A×A. So, Pp(A×A) is the proportion of pluses in zoneA×A. We call Psp(A×A) the number of OGUs in group Athat have at least one plus divided by the total number of OGUs in group A. We can then compute d1(A×A) asfollows: if the number of pluses in A×A is zero, then d1(A×A)=0; otherwise,
d1(A×A)=Pp(A×A)×Psp(A×A)
(Pp(A×A))2
+(Psp(A×A))2
The values of d1(A×A) range from 0 to 0.707, estimating towhat extent similarities higher than expected at random (+)predominate within zone A×A.
We define Pm(A×A) and Psm(A×A) as the proportion of minuses in zone A×A and the proportion of OGUs in groupA with at least one minus, respectively, and these are com-puted in the same way as Pp(A×A) and Psp(A×A), buttaking into account the minuses. We then define d2(A×A) inthe following way: if the number of minuses in zone A×A iszero, then d2(A×A)=0; otherwise,
d2(A×A)=Pm(A×A)×Psm(A×A)
(Pm(A×A))2+(Psm(A×A))2
The values of d2(A×A) range from 0 to 0.707, estimating to
what extent similarities lower than expected at random (−)predominate within zone A×A. We d efine Pp(A×B) andPsp(A×B) in a similar way to Pp(A×A) and Psp(A×A),but with reference to zone A×B. So, d3 is zero when thenumber of pluses in A×B is zero; otherwise,
ECOGRAPHY 24:6 (2001) 669

8/2/2019 Biotics Regionalizations Methods
http://slidepdf.com/reader/full/biotics-regionalizations-methods 12/12
d3=Pp(A×B)×Psp(A×B)
(Pp(A×B))2+(Psp(A×B))2
The values of d3 range from 0 to 0.707, estimating to whatextent similarities higher than expected at random (+) pre-dominate within zone A×B.
The parameter DW(A×A) measures to what extent thesimilarities that are higher than expected (+) tend to be inzones A×A but not in A×B (see McCoy et al. 1986), whereDW(A×A)=d1(A×A)−d2(A×A)−d3.
Similarly, DW(B×B)=d1(B×B)−d2(B×B)−d3, whered1(B×B) and d2(B×B) are calculated as d1(A×A) andd2(A×A), but computing the pluses and minuses in zoneB×B. The average of DW(A×A) and DW(B×B), named
DW, measures to what extent similarities that are higher thanexpected (+) tend to be in either zones A×A o r B×B butnot in A×B.
We define d4 in the same way as d3, but compute theminuses in A×B. We then compute the parameter DS=d4−d3−d2(A×A)−d2(B×B), which gives a measure o f whether the similarities that are lower than expected (−) tendto be located in A×B, but not in A×A or B×B.
The statistical significance of a node was assessed using aG-test of independence (Sokal and Rohlf 1981, McCoy et al.1986) of the distribution of the signs ‘‘+’’, ‘‘−’’ and ‘‘0’’ inthe three zones of the submatrix, and so we obtained theparameters GW, for weak boundaries, and GS, for strongboundaries. If similarities higher than expected (+) tendsignificantly to be in zones A×A or B×B, but not in A×B,that is, if DW\0 and GW is statistically significant, thenthere is at least a weak biotic boundary between both groupsof OGUs. In this case, if DW(A×A)\0 then the group of OGUs A constitutes a biotic region, and the same applies forDW(B×B) and the group of OGUs B, because then a groupof OGUs with similar biota shares a number of species com-patible with random expectation with another group of OGUsthat also has a similar biota. If similarities significantly lower
than expected (−) tend to be located in A×B, but not inA×A or B×B, that is, if DS\0 and GS is significant, thena strong biotic boundary exists between the groups of OGUs.In this case a group of OGUs shares fewer species thanexpected at random with another group of OGUs. The areasdelimited by strong boundaries are biotic supraregions. Asupraregion can consist of several biotic regions separated byweak boundaries.
670 ECOGRAPHY 24:6 (2001)
Related Documents