Article Biotic and Abiotic Determinants of Soil Organic Matter Stock and Fine Root Biomass in Mountain Area Temperate Forests—Examples from Cambisols under European Beech, Norway Spruce, and Silver Fir (Carpathians, Central Europe) Anna Zielonka 1, * , Marek Drewnik 1 , Lukasz Musielok 1 , Marcin K. Dyderski 2 , Dariusz Struzik 1 , Grzegorz Smulek 1 and Katarzyna Ostapowicz 1,3 Citation: Zielonka, A.; Drewnik, M.; Musielok, L.; Dyderski, M.K.; Struzik, D.; Smulek, G.; Ostapowicz, K. Biotic and Abiotic Determinants of Soil Organic Matter Stock and Fine Root Biomass in Mountain Area Temperate Forests—Examples from Cambisols under European Beech, Norway Spruce, and Silver Fir (Carpathians, Central Europe). Forests 2021, 12, 823. https://doi.org/10.3390/f12070823 Academic Editor: Bartosz Adamczyk Received: 16 May 2021 Accepted: 16 June 2021 Published: 22 June 2021 Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affil- iations. Copyright: © 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https:// creativecommons.org/licenses/by/ 4.0/). 1 Faculty of Geography and Geology, Institute of Geography and Spatial Management, Jagiellonian University, Gronostajowa 7, 30387 Kraków, Poland; [email protected] (M.D.); [email protected] (L.M.); [email protected] (D.S.); [email protected] (G.S.); [email protected] (K.O.) 2 Institute of Dendrology, Polish Academy of Sciences, Parkowa 5, 62035 Kórnik, Poland; [email protected] 3 Department of Environmental Science, Policy & Management, University of California Berkeley, Mulford Hall, Berkeley, CA 94720, USA * Correspondence: [email protected] Abstract: Forest ecosystems significantly contribute to the global organic carbon (OC) pool, exhibiting high spatial heterogeneity in this respect. Some of the components of the OC pool in a forest (woody aboveground biomass (wAGB), coarse root biomass (CRB)) can be relatively easily estimated using readily available data from land observation and forest inventories, while some of the components of the OC pool are very difficult to determine (fine root biomass (FRB) and soil organic matter (SOM) stock). The main objectives of our study were to: (1) estimate the SOM stock; (2) estimate FRB; and (3) assess the relationship between both biotic (wAGB, forest age, foliage, stand density) and abiotic factors (climatic conditions, relief, soil properties) and SOM stocks and FRB in temperate forests in the Western Carpathians consisting of European beech, Norway spruce, and silver fir (32 forest inventory plots in total). We uncovered the highest wAGB in beech forests and highest SOM stocks under beech forest. FRB was the highest under fir forest. We noted a considerable impact of stand density on SOM stocks, particularly in beech and spruce forests. FRB content was mostly impacted by stand density only in beech forests without any discernible effects on other forest characteristics. We discovered significant impacts of relief-dependent factors and SOM stocks at all the studied sites. Our biomass and carbon models informed by more detailed environmental data led to reduce the uncertainty in over- and underestimation in Cambisols under beech, spruce, and fir forests for mountain temperate forest carbon pools. Keywords: forest characteristics; fine roots biomass; soil organic matter; Cambisols; mountain temperate forests; Carpathians 1. Introduction Forest ecosystems significantly contribute to global organic carbon (OC) sequestra- tion [1,2], storing 80% of the aboveground OC pool and 40% of the soil organic carbon (SOC) pool [3,4]. In the case of forests, a very high heterogeneity of the OC stock was noted, which was explained via both abiotic factors—such as topographic effects [5–9], morphogenetic processes [10], and soil properties [11–14]—and biotic factors—such as the tree species composition [15–20], silviculture treatment [10,12] and stand age [21,22]. The quantification of the OC pool in forests that cover large areas of land is important not only from a research point of view but more importantly in the context of expected changes due to climate change and deforestation observed all over the world [23–26]. Forests 2021, 12, 823. https://doi.org/10.3390/f12070823 https://www.mdpi.com/journal/forests

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Article
Biotic and Abiotic Determinants of Soil Organic Matter Stockand Fine Root Biomass in Mountain Area TemperateForestsmdashExamples from Cambisols under European BeechNorway Spruce and Silver Fir (Carpathians Central Europe)
Anna Zielonka 1 Marek Drewnik 1 Łukasz Musielok 1 Marcin K Dyderski 2 Dariusz Struzik 1Grzegorz Smułek 1 and Katarzyna Ostapowicz 13
Citation Zielonka A Drewnik M
Musielok Ł Dyderski MK Struzik
D Smułek G Ostapowicz K Biotic
and Abiotic Determinants of Soil
Organic Matter Stock and Fine Root
Biomass in Mountain Area Temperate
ForestsmdashExamples from Cambisols
under European Beech Norway
Spruce and Silver Fir (Carpathians
Central Europe) Forests 2021 12 823
httpsdoiorg103390f12070823
Academic Editor Bartosz Adamczyk
Received 16 May 2021
Accepted 16 June 2021
Published 22 June 2021
Publisherrsquos Note MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations
Copyright copy 2021 by the authors
Licensee MDPI Basel Switzerland
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https
creativecommonsorglicensesby
40)
1 Faculty of Geography and Geology Institute of Geography and Spatial Management Jagiellonian UniversityGronostajowa 7 30387 Krakoacutew Poland marekdrewnikujedupl (MD) lmusielokujedupl (ŁM)darekstruzikstudentujedupl (DS) grzegorzsmulekujedupl (GS)katarzynaostapowiczujedupl (KO)
2 Institute of Dendrology Polish Academy of Sciences Parkowa 5 62035 Koacuternik Polandmdyderskimanpoznanpl
3 Department of Environmental Science Policy amp Management University of California BerkeleyMulford Hall Berkeley CA 94720 USA
Correspondence annazielonkadoctoralujedupl
Abstract Forest ecosystems significantly contribute to the global organic carbon (OC) pool exhibitinghigh spatial heterogeneity in this respect Some of the components of the OC pool in a forest (woodyaboveground biomass (wAGB) coarse root biomass (CRB)) can be relatively easily estimated usingreadily available data from land observation and forest inventories while some of the components ofthe OC pool are very difficult to determine (fine root biomass (FRB) and soil organic matter (SOM)stock) The main objectives of our study were to (1) estimate the SOM stock (2) estimate FRB and(3) assess the relationship between both biotic (wAGB forest age foliage stand density) and abioticfactors (climatic conditions relief soil properties) and SOM stocks and FRB in temperate forestsin the Western Carpathians consisting of European beech Norway spruce and silver fir (32 forestinventory plots in total) We uncovered the highest wAGB in beech forests and highest SOM stocksunder beech forest FRB was the highest under fir forest We noted a considerable impact of standdensity on SOM stocks particularly in beech and spruce forests FRB content was mostly impactedby stand density only in beech forests without any discernible effects on other forest characteristicsWe discovered significant impacts of relief-dependent factors and SOM stocks at all the studiedsites Our biomass and carbon models informed by more detailed environmental data led to reducethe uncertainty in over- and underestimation in Cambisols under beech spruce and fir forests formountain temperate forest carbon pools
Keywords forest characteristics fine roots biomass soil organic matter Cambisols mountaintemperate forests Carpathians
1 Introduction
Forest ecosystems significantly contribute to global organic carbon (OC) sequestra-tion [12] storing 80 of the aboveground OC pool and 40 of the soil organic carbon(SOC) pool [34] In the case of forests a very high heterogeneity of the OC stock wasnoted which was explained via both abiotic factorsmdashsuch as topographic effects [5ndash9]morphogenetic processes [10] and soil properties [11ndash14]mdashand biotic factorsmdashsuch as thetree species composition [15ndash20] silviculture treatment [1012] and stand age [2122] Thequantification of the OC pool in forests that cover large areas of land is important not onlyfrom a research point of view but more importantly in the context of expected changes dueto climate change and deforestation observed all over the world [23ndash26]
Forests 2021 12 823 httpsdoiorg103390f12070823 httpswwwmdpicomjournalforests
Forests 2021 12 823 2 of 27
The total OC pool in a forest is partitioned between OC accumulated in abovegroundbiomass (AGB) belowground biomass (BGB) and the soil organic matter (SOM) stock [9]The estimation of AGB for a forest is relatively easy Effective models for calculatingAGB for forests based on stand characteristics (ie tree species age diameter at breastheight) have already been introduced [27] Similarly remote sensing data provide globalinformation on AGB for forests with high accuracy [28ndash31] In contrast the estimation ofBGB and the SOM stock is more difficult and expensive in terms of time and money [53233]Therefore a model that allows for a reliable estimation of both BGB and SOC stocks basedon readily available data remains to be developed [22]
In the case of BGB the quantity of coarse root biomass (CRB) can be estimated quitereliably using the already developed allometric models noted above [27] while in the caseof fine root biomass (FRB) there is no proven reliable method Fine roots are difficult tostudy as they have a short lifespan (less than one to nine years) [34] significant seasonalvariability [1835] and a high rate of decomposition [36] In addition the estimation ofSOC stocks based on biotic and abiotic factors is difficult because it is necessary to takeinto account events occurring both in the present day and in the past since the amountof SOC depends on the balance between the accumulation and decomposition of organicmatter at present but also on the initial quantity [10] However it should be emphasizedthat the field measurements of both SOC stocks as well as biomass accumulated in litterare accurate [510323337] while measurements of FRB involve a considerable degree ofuncertainty [3839]
The estimation of both the BGB and SOM stock is also challenging due to theirfunctional and not fully understood linkages including the seasonal effects of living rootson organic matter turnover [40ndash42] Generally due to slow OC and nutrient turnoverthe coarse root distribution represented by CRB may be more important for long-termecosystem productivity [4344] however fine roots also indirectly determine the size ofthe OC pool in forest ecosystems [45ndash48] Fine roots may be considered a vital element ofthe physical connection between trees and soil [48] and by releasing various substancesinto the surrounding soil they control the decomposition of dead organic matter whichthen leads to the allocation of OC [49] Additionally special attention should be paid toFRB due to its important role in the biogeochemical cycle which is an important part ofthe ecophysiology of trees [50]
As noted earlier tree species directly affect AGB and CRB but may also indirectly affectFRB and SOC stocks by altering the dynamics of soil organic matter decomposition [5152]Tree species are thought to influence microbiological decomposition mainly by varyingthe content of lignin nitrogen and alkaline and acid cations [155354] Conifers have ahigher lignin content in their needles (25 to 33 dry weight) than broadleaf trees do inleaves (20 to 25 dry weight) [55] resulting in different litter quality which later affectsdecomposition rates As a result there are different SOC stocks under different tree speciesFor example in temperate mountain forests a higher SOC stock in organic soil horizons(ie litter consisting of different fresh non-decomposed or slightly decomposed organicmaterial) was observed in comparison with that in the mineral part of soil under coniferousspecies [5] while in soil found under broadleaved species more SOC is stored in themineral part of the soil [56] From all the above-mentioned studies it may be concludedthat in the case of forests taking into account the tree species is very important in relationto the estimation of both FRB and SOM
As previous studies have shown remote sensing data provide global information onAGB found in forests with a rather high degree of accuracy [28ndash31] while allometric modelsbased on detailed data from forest inventories [27] provide information on AGB and FRB inforests with a very high degree of accuracy Thus if we knew the relationship between AGBand both the BGB and SOM stocks the elucidation of these relationships would improvefuture global OC pool models for forests Such knowledge would significantly contributeto better use of the capabilities of global Earth observation missions (eg GEDI BIOMASS)to determine global and regional OC pools in forests The authors of [28] highlighted the
Forests 2021 12 823 3 of 27
urgent need to link soil organic carbon stock data with aboveground biomass data on thebasis of Earth observation missions yielding forest inventory data
The determination of a relationship between AGB and the less accurately identifiedSOC stock and FRB requires the field and laboratory research on study plots for whichvery detailed forest inventory data are available [57] A potentially broad set of abiotic andbiotic factors should be taken into account in these investigations [102256]
We hypothesized that it may be possible to estimate both the SOC stock and FRBusing easily available data obtained from national forest inventories (NFI) characterizingstandsmdashnotably woody aboveground biomass (wAGB) in mountain temperate forestsin the Polish Carpathians In this regard we aimed to both determine and evaluate therelationship between selected forest stand characteristics ie wAGB forest age standdensity foliage (coming from NFI) and both FRB and SOM stocks in mountain areatemperate forests in the Western Carpathians consisting primarily of the most commonstand species ie European beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and silver fir (Abies alba Mill) We decided to take into account available abiotic factors(ie elevation hillslope exposure hillslope gradient mean annual air temperature (MAT)mean annual precipitation (MAP) soil properties) as they all affect the species-specificpatterns of aboveground biomass accumulation in mountain area temperate forests [58]Considering above- and belowground OC pools the main objectives of our study were(1) to estimate SOM stocks (2) estimate FRB and (3) assess the relationship between bothabiotic and biotic factors and the SOM stock and FRB in the studied soils
2 Materials and Methods21 Study Area and Study Plot Selection
The study area is located in the Western Carpathians (Central Europe) The climatein the study area is moderately cold with average monthly summertime temperaturesvarying from 16 C in the foothills to 8 C atop the highest peaks The average monthlywinter temperature stays below 0 C reachingminus8 C at the highest elevations The averageannual air temperature reaches 7 C [59] Our study sites differed slightly in terms ofclimate characteristics (Table 1) and differed strongly in terms of tree species typemdashtheselected sites were covered with beech spruce fir and mixed stands The present-dayforest structure in the Polish part of the Western Carpathians is mainly the product ofcenturies of forest usemdashwith significant forest gains occurring since around 1850 Howeverover time primeval beech and firndashbeech forests were replaced by spruce plantationsat most of the study sites [60ndash62] Beech and firndashbeech forests occupy a much smallerarea today
Table 1 Characteristics of the investigated study sites
Study Site CoordinatesElevation Range
MinndashMax(m asl)
HillslopeGradient Range
MinndashMax( )
HillslopeExposure Range
MinndashMax( )
Mean AnnualAir
Temperature(C)
Mean AnnualPrecipitation
(mm)
1 4926prime54rdquo N 1903prime05rdquo E 701ndash808 13ndash24 4ndash165 51 11272 4911prime30rdquo N 2228prime12rdquo E 940ndash1067 11ndash21 276ndash316 46 10683 4938prime01rdquo N 1858prime36rdquo E 768ndash887 15ndash25 218ndash319 52 11034 4934prime28rdquo N 1941prime09rdquo E 706ndash753 2ndash22 44ndash340 55 9785 4929prime27rdquo N 2036prime35rdquo E 575ndash658 21ndash26 59ndash79 63 10216 4937prime44rdquo N 1928prime30rdquo E 836ndash937 16ndash20 32ndash358 48 11347 4925prime10rdquo N 2201prime56rdquo E 602ndash624 8ndash16 100ndash266 61 870
Our study was carried out on selected NFI plots which are regular grids of studysites located 4 km from each other featuring L-shaped trailsmdashplots part of the ICP Forestsystem (httpsicp-forestsnet (accessed on 15 May 2020)) The above study plots wereestablished for the purpose of the assessment of forest damage across the European UnionA total of 32 plots out of 2450 plots located in the Western Carpathians were selected forstudy purposes (Figure 1) all of which had to meet the following conditions (1) the studied
Forests 2021 12 823 4 of 27
plots had to represent very similar soils ie loamy texture medium depth Cambisolsdeveloped from cover-beds originating in flysch rocks [6364] and (2) all the plots had to besituated in a similar relief position (ie linear hillslope) thus the areal drainage conditionscould be expected to be quite similar (digital elevation model produced as part of Polandrsquosnational GIS project) The selected study plots were located at seven study sites (Table 1)and composed of five circular plots each (r = 12 m) The study plots were located 200 maway from each other in a regular pattern Together all the study sites formed an L-shapedtrail (Figure 1) We excluded three study plots due to the presence of young forest resultingin a lack of accurate tree stand measurements decreasing the accuracy of the biomassassessment [21]
Figure 1 Study area on a map of Europe (A) distribution of study sites (B) (numbers) and distribu-tion of study plots (numbers with letters eg 6A 6B) at each study site (C)
22 Field Survey
Field sampling was conducted in 2017 (June to November) For each study plot a soilprofile was described according to [65] and then sampled (total of 32 soil profiles) Onesoil pit at each study site (plot labeled lsquoCrsquo according to the A to E lettering at Figure 1)was excavated to the lithic contact (seven reference soils) while in the other study plotssoil pits were excavated to approximately 50 cm (25 soil profiles) Rock fragment (coarseparticles diameter gt 2 mm) content in each soil horizon was estimated using standardcharts (templates) commonly used in soil surveys [65] Soil profiles were not established inclose proximity of tree stems (up to 5 m as this distance represented averaged conditionsin the investigated study plots)
A core sampler (100 cm3 in stony soils 635 cm3) was used to collect undisturbedsoil samples (which also incorporated fine roots) using 10 cm intervals (0 to 10 cm 10 to20 cm 20 to 30 cm 30 to 40 cm) at each of the 32 study sites Each sample was collectedin triplicate In addition bulk soil samples (disturbed samples ca 1 kg) from selectedgenetic soil horizons were collected Bulk samples from littermdashdefined as the surface layerof the forest floor for the purpose of the current paper consisting of different fresh non-decomposed or slightly decomposed organic material with recognizable tissuesmdashwerecollected using a metal frame 10 cm x 10 cm taking into account the representativenessof the sampling point Every soil sample was then placed in a sterile polyethylene bag
Forests 2021 12 823 5 of 27
(Whirl-Pakreg) and stored at ~4 C before laboratory analysis The investigated soils wereclassified according to the WRB [66]
23 Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
Litter samples were air-dried crushed and homogenized for further analysis Fromundisturbed soil samples (taken using the core sampler) fine roots were hand-picked anddried to constant mass at 70 C and weighed with an accuracy of 01 g Prior to dryinglive and dead fine roots were distinguished by their color elasticity and degree of barkcohesion [3839] Based on an analysis of 30 of randomly selected samples a share of10 (in mass) of dead fine roots was determined Therefore for the purpose of furtheranalysis it was assumed that 90 of the mass of fine roots in a given sample are live rootsand these values were selected for subsequent models This approach is consistent withfindings by [186768] all of whom demonstrated that in the middle and at the end of thegrowing season the amount of live fine roots is very large in total FRB Dead fine roots wereclassified as non-humic substances in soil organic matter We used the volume of the coresampler as a sample volume for calculations (Equation (1)) Soil material from undisturbedsoil samples (without live fine roots) was dried at 105 C and weighed Then the bulkdensity (BD) was calculated according to [32] The calculation procedure according to [32]excludes coarse particles (Oslash gt 2 mm) as they are not a component of soil bulk density(Equation (1))
BD =masssampleminusmassparticles gt 2 mm
volumesampleminus massparticlesgt2 mmdensityparticlesgt2 mm
(1)
Disturbed bulk soil samples (taken separately in parallel) were air-dried gentlycrushed and sieved through a 2 mm mesh steel sieve Rock fragments (Oslash gt 2 mm) weredetermined by weighing Based on this measurement the content of rock fragmentsdetermined during fieldwork (see above) was calibrated Soil properties for fine soil(Oslash lt 2 mm) and litter subsamples were determined The soil and litter pH were measuredpotentiometrically in distilled water (125) [69] using a pH meter (Elmetron CPI-505) with aglass electrode (Elmetron ERH-11) The concentration of total carbon (TC) and nitrogen (N)was determined in triplicate (and then averaged) using dry combustion gas chromatographwith a CHN analyzer (Elementar vario MICRO cube elemental analyzer) Due to theabsence of carbonates in the studied soils it was assumed that TC corresponds to SOC [70]The SOC stock was herein calculated according to [32] The soil texture was determinedusing the combined sieving and hydrometer method [71]
Due to the need to carry out an analysis for comparable values for further analysisthe studied SOC was converted into SOM using the conversion factor 1724 [72] as precisedata (obtained via experimental research) allowing the conversion of wAGB CRB andFRB into the OC pool remain unavailable
24 Stand Characteristics and Coarse Root Biomass Estimation
We obtained data on tree species and DBH (diameter at breast height) for all treeswith DBH gt 7 cm This approach omits small trees and shrubs however their biomassyields only a small contribution to total biomass as biomass increases exponentially withDBH [21] These data had been previously collected as part of NFI by the Polish ForestResearch Institute and were verified in the course of our fieldwork in 2017 (June to October)The wAGB was calculated using biomass equations based on meta-analysis (Equation (2))for European tree species [27] basing on DBH (cm)
ln (AGB) = ln (β0) + β1ln(d) + ε (2)
where AGBmdashwoody aboveground biomass (kg) β0 β1mdashmodel parameters (coefficients)beechmdashβ0 = minus16594 β1 = 23589 sprucemdashβ0 = minus18865 β1 = 23034 firmdashβ0 = minus23958β1 = 24497 other coniferous treesmdashβ0 = minus16173 β1 = 23177 other deciduous treesmdash
Forests 2021 12 823 6 of 27
β0 = minus11972 β1 = 23417 εmdashan error that is assumed to be normally distributed with amean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
Additionally we calculated the stand density (ind ha-1) The biomass of foliage wascalculated using models provided by [73] for beech and spruce forests (Equation (3))
M = a times Xb (3)
where Mmdashfoliage biomass (Mg ha-1) a bmdashmodel parameters sprucemdasha = 44073b = 03544 beechmdasha = 08813 b = 04151 Xmdashstand basal area (m2 ha-1) Model parametersfor fir forest were obtained from [74] a = minus307447 b = 281850
The biomass of coarse roots (CRB diameter of root gt 2 mm) was calculated for beechspruce and fir separately using general equations (Equation (4)) [27]
ln (CRB) = ln (β0) + β1ln (d) + ε (4)
where CRBmdashcoarse root biomass (kg) β0 β1mdashmodel parameters sprucemdashβ0 = minus37387β1 = 24323 beechmdashβ0 = minus33713 β1 = 23424 firmdashβ0 = minus40287 β1 = 24957 otherspeciesmdashβ0 = minus31677 β1 = 22796 εmdashan error that is assumed to be normally distributedwith a mean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
25 Data Analysis
We employed machine learning techniques to determine the effects of selected bioticforest stand characteristics and abiotic factors on FRB and SOM stocks (Table 2) Owingto a large number of predictions and interactions among them machine learning andexplanatory tools [75] enable the generation of a clear description of the meaning of eachgiven prediction and its partial dependence From different machine learning algorithmswe selected a random forest algorithmmdasha technique based on multiple decisions or treeclassifications which increases model stability and accuracy [76] The random forestmodel has been successfully used for vegetation modeling [77] We used R software [78](version 402 ldquoTaking Off Againrdquo R Core Team 2019) for data analysis All models weredeveloped using the caret package [79] for model development and the DALEX packagefor the visual explanation and exploration of predictive models [75] To understand howthe said models work we used variable importance and partial dependence plots Wequantified the variable importance using the drop-out loss function ie loss-decreaseroot mean square error (RMSE) This function informs us how much the model RMSEincreases when a particular variable is shuffled within a given dataset Literally it showshow much model quality we lose when we replace a particular variable with its randompermutations [75] The higher drop in losses in the RMSE indicates the higher importanceof the variable for proper prediction The partial dependence plots show the change in themodel output along with an increasing value of a particular predictor assuming all othervariables at the constant (mean) level [75] Literally this is the prediction of the modelusing all variables at the constant level and manipulating only the variable of interestSuch an approach simulates the experimental rule ceteris paribus which means lsquoall equalwithout the single manipulatedrsquo and allows to see how FRB and SOM stocks vary alongeach of the predictors without changing other variables These marginal predictions allowone to make conclusions about the explicit impact of a particular variable
Forests 2021 12 823 7 of 27
Table 2 List of predictors
Abbreviation Predictor Unit
wAGB Woody aboveground biomass Mg haminus1
StDens Stand density ind haminus1
foliage Foliage biomass Mg haminus1
age Age of dominant species yearslitter Litter mass Mg haminus1
pH_lit Litter pH (in H2O) dimensionlessCN_lit Organic CarbonNitrogen ratio in litter dimensionless
MAT Mean annual air temperature CMAP Mean annual precipitation mmelev Elevation m asl
slope Hillslope gradient
aspect Hillslope exposure
SOM Soil Organic Matter stock Mg haminus1
CN Organic CarbonNitrogen ratio in soil dimensionlessClay Clay content Silt Silt content pH Soil pH (in H2O) dimensionlessCR Coarse roots biomass Mg haminus1
FRB Fine roots biomass Mg haminus1
mean values at 0 to 40 cm depth
To assess the FRB and SOM we developed separate regression models for eachexamined dominant tree species stand and training models to decrease RMSE usingthe caret package [79] To demonstrate the modelrsquos quality we provided RMSE andr2mdashcoefficient of determination indicating the proportions of variability explained bythe model
3 Results31 Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
The analyzed forest stand characteristics varied between study plots and betweentree species (Tables 3ndash5) Similar findings were also obtained for climatic and morphologicpredictors as well as the herein analyzed soil property predictors (Table A1) Both themorphology and properties of the investigated soils were very similar (Figure A1) Thetexture was loamy throughout the studied soil profiles (silt loam sandy loam and loam)(Table S1 from Supplementary Materials) The soil structure was subangular or angularblocky transitioning into massive in the bottom part of the soil In addition the soilconsistence changed gradually with the depth of the soil profile from soft in the uppermosthorizons to very hard in the lowermost part of the soil (Table 6) The gradual decreasein SOC content with soil depth occurred in all the study plots (Figure 2) According totree species groups the studied plotsrsquo soil texture varied slightlymdashthe highest silt and claycontent was observed under fir-dominated forests while the lowest silt content was underbeech forest and clay content under spruce forest (Table A1)
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 2 of 27
The total OC pool in a forest is partitioned between OC accumulated in abovegroundbiomass (AGB) belowground biomass (BGB) and the soil organic matter (SOM) stock [9]The estimation of AGB for a forest is relatively easy Effective models for calculatingAGB for forests based on stand characteristics (ie tree species age diameter at breastheight) have already been introduced [27] Similarly remote sensing data provide globalinformation on AGB for forests with high accuracy [28ndash31] In contrast the estimation ofBGB and the SOM stock is more difficult and expensive in terms of time and money [53233]Therefore a model that allows for a reliable estimation of both BGB and SOC stocks basedon readily available data remains to be developed [22]
In the case of BGB the quantity of coarse root biomass (CRB) can be estimated quitereliably using the already developed allometric models noted above [27] while in the caseof fine root biomass (FRB) there is no proven reliable method Fine roots are difficult tostudy as they have a short lifespan (less than one to nine years) [34] significant seasonalvariability [1835] and a high rate of decomposition [36] In addition the estimation ofSOC stocks based on biotic and abiotic factors is difficult because it is necessary to takeinto account events occurring both in the present day and in the past since the amountof SOC depends on the balance between the accumulation and decomposition of organicmatter at present but also on the initial quantity [10] However it should be emphasizedthat the field measurements of both SOC stocks as well as biomass accumulated in litterare accurate [510323337] while measurements of FRB involve a considerable degree ofuncertainty [3839]
The estimation of both the BGB and SOM stock is also challenging due to theirfunctional and not fully understood linkages including the seasonal effects of living rootson organic matter turnover [40ndash42] Generally due to slow OC and nutrient turnoverthe coarse root distribution represented by CRB may be more important for long-termecosystem productivity [4344] however fine roots also indirectly determine the size ofthe OC pool in forest ecosystems [45ndash48] Fine roots may be considered a vital element ofthe physical connection between trees and soil [48] and by releasing various substancesinto the surrounding soil they control the decomposition of dead organic matter whichthen leads to the allocation of OC [49] Additionally special attention should be paid toFRB due to its important role in the biogeochemical cycle which is an important part ofthe ecophysiology of trees [50]
As noted earlier tree species directly affect AGB and CRB but may also indirectly affectFRB and SOC stocks by altering the dynamics of soil organic matter decomposition [5152]Tree species are thought to influence microbiological decomposition mainly by varyingthe content of lignin nitrogen and alkaline and acid cations [155354] Conifers have ahigher lignin content in their needles (25 to 33 dry weight) than broadleaf trees do inleaves (20 to 25 dry weight) [55] resulting in different litter quality which later affectsdecomposition rates As a result there are different SOC stocks under different tree speciesFor example in temperate mountain forests a higher SOC stock in organic soil horizons(ie litter consisting of different fresh non-decomposed or slightly decomposed organicmaterial) was observed in comparison with that in the mineral part of soil under coniferousspecies [5] while in soil found under broadleaved species more SOC is stored in themineral part of the soil [56] From all the above-mentioned studies it may be concludedthat in the case of forests taking into account the tree species is very important in relationto the estimation of both FRB and SOM
As previous studies have shown remote sensing data provide global information onAGB found in forests with a rather high degree of accuracy [28ndash31] while allometric modelsbased on detailed data from forest inventories [27] provide information on AGB and FRB inforests with a very high degree of accuracy Thus if we knew the relationship between AGBand both the BGB and SOM stocks the elucidation of these relationships would improvefuture global OC pool models for forests Such knowledge would significantly contributeto better use of the capabilities of global Earth observation missions (eg GEDI BIOMASS)to determine global and regional OC pools in forests The authors of [28] highlighted the
Forests 2021 12 823 3 of 27
urgent need to link soil organic carbon stock data with aboveground biomass data on thebasis of Earth observation missions yielding forest inventory data
The determination of a relationship between AGB and the less accurately identifiedSOC stock and FRB requires the field and laboratory research on study plots for whichvery detailed forest inventory data are available [57] A potentially broad set of abiotic andbiotic factors should be taken into account in these investigations [102256]
We hypothesized that it may be possible to estimate both the SOC stock and FRBusing easily available data obtained from national forest inventories (NFI) characterizingstandsmdashnotably woody aboveground biomass (wAGB) in mountain temperate forestsin the Polish Carpathians In this regard we aimed to both determine and evaluate therelationship between selected forest stand characteristics ie wAGB forest age standdensity foliage (coming from NFI) and both FRB and SOM stocks in mountain areatemperate forests in the Western Carpathians consisting primarily of the most commonstand species ie European beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and silver fir (Abies alba Mill) We decided to take into account available abiotic factors(ie elevation hillslope exposure hillslope gradient mean annual air temperature (MAT)mean annual precipitation (MAP) soil properties) as they all affect the species-specificpatterns of aboveground biomass accumulation in mountain area temperate forests [58]Considering above- and belowground OC pools the main objectives of our study were(1) to estimate SOM stocks (2) estimate FRB and (3) assess the relationship between bothabiotic and biotic factors and the SOM stock and FRB in the studied soils
2 Materials and Methods21 Study Area and Study Plot Selection
The study area is located in the Western Carpathians (Central Europe) The climatein the study area is moderately cold with average monthly summertime temperaturesvarying from 16 C in the foothills to 8 C atop the highest peaks The average monthlywinter temperature stays below 0 C reachingminus8 C at the highest elevations The averageannual air temperature reaches 7 C [59] Our study sites differed slightly in terms ofclimate characteristics (Table 1) and differed strongly in terms of tree species typemdashtheselected sites were covered with beech spruce fir and mixed stands The present-dayforest structure in the Polish part of the Western Carpathians is mainly the product ofcenturies of forest usemdashwith significant forest gains occurring since around 1850 Howeverover time primeval beech and firndashbeech forests were replaced by spruce plantationsat most of the study sites [60ndash62] Beech and firndashbeech forests occupy a much smallerarea today
Table 1 Characteristics of the investigated study sites
Study Site CoordinatesElevation Range
MinndashMax(m asl)
HillslopeGradient Range
MinndashMax( )
HillslopeExposure Range
MinndashMax( )
Mean AnnualAir
Temperature(C)
Mean AnnualPrecipitation
(mm)
1 4926prime54rdquo N 1903prime05rdquo E 701ndash808 13ndash24 4ndash165 51 11272 4911prime30rdquo N 2228prime12rdquo E 940ndash1067 11ndash21 276ndash316 46 10683 4938prime01rdquo N 1858prime36rdquo E 768ndash887 15ndash25 218ndash319 52 11034 4934prime28rdquo N 1941prime09rdquo E 706ndash753 2ndash22 44ndash340 55 9785 4929prime27rdquo N 2036prime35rdquo E 575ndash658 21ndash26 59ndash79 63 10216 4937prime44rdquo N 1928prime30rdquo E 836ndash937 16ndash20 32ndash358 48 11347 4925prime10rdquo N 2201prime56rdquo E 602ndash624 8ndash16 100ndash266 61 870
Our study was carried out on selected NFI plots which are regular grids of studysites located 4 km from each other featuring L-shaped trailsmdashplots part of the ICP Forestsystem (httpsicp-forestsnet (accessed on 15 May 2020)) The above study plots wereestablished for the purpose of the assessment of forest damage across the European UnionA total of 32 plots out of 2450 plots located in the Western Carpathians were selected forstudy purposes (Figure 1) all of which had to meet the following conditions (1) the studied
Forests 2021 12 823 4 of 27
plots had to represent very similar soils ie loamy texture medium depth Cambisolsdeveloped from cover-beds originating in flysch rocks [6364] and (2) all the plots had to besituated in a similar relief position (ie linear hillslope) thus the areal drainage conditionscould be expected to be quite similar (digital elevation model produced as part of Polandrsquosnational GIS project) The selected study plots were located at seven study sites (Table 1)and composed of five circular plots each (r = 12 m) The study plots were located 200 maway from each other in a regular pattern Together all the study sites formed an L-shapedtrail (Figure 1) We excluded three study plots due to the presence of young forest resultingin a lack of accurate tree stand measurements decreasing the accuracy of the biomassassessment [21]
Figure 1 Study area on a map of Europe (A) distribution of study sites (B) (numbers) and distribu-tion of study plots (numbers with letters eg 6A 6B) at each study site (C)
22 Field Survey
Field sampling was conducted in 2017 (June to November) For each study plot a soilprofile was described according to [65] and then sampled (total of 32 soil profiles) Onesoil pit at each study site (plot labeled lsquoCrsquo according to the A to E lettering at Figure 1)was excavated to the lithic contact (seven reference soils) while in the other study plotssoil pits were excavated to approximately 50 cm (25 soil profiles) Rock fragment (coarseparticles diameter gt 2 mm) content in each soil horizon was estimated using standardcharts (templates) commonly used in soil surveys [65] Soil profiles were not established inclose proximity of tree stems (up to 5 m as this distance represented averaged conditionsin the investigated study plots)
A core sampler (100 cm3 in stony soils 635 cm3) was used to collect undisturbedsoil samples (which also incorporated fine roots) using 10 cm intervals (0 to 10 cm 10 to20 cm 20 to 30 cm 30 to 40 cm) at each of the 32 study sites Each sample was collectedin triplicate In addition bulk soil samples (disturbed samples ca 1 kg) from selectedgenetic soil horizons were collected Bulk samples from littermdashdefined as the surface layerof the forest floor for the purpose of the current paper consisting of different fresh non-decomposed or slightly decomposed organic material with recognizable tissuesmdashwerecollected using a metal frame 10 cm x 10 cm taking into account the representativenessof the sampling point Every soil sample was then placed in a sterile polyethylene bag
Forests 2021 12 823 5 of 27
(Whirl-Pakreg) and stored at ~4 C before laboratory analysis The investigated soils wereclassified according to the WRB [66]
23 Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
Litter samples were air-dried crushed and homogenized for further analysis Fromundisturbed soil samples (taken using the core sampler) fine roots were hand-picked anddried to constant mass at 70 C and weighed with an accuracy of 01 g Prior to dryinglive and dead fine roots were distinguished by their color elasticity and degree of barkcohesion [3839] Based on an analysis of 30 of randomly selected samples a share of10 (in mass) of dead fine roots was determined Therefore for the purpose of furtheranalysis it was assumed that 90 of the mass of fine roots in a given sample are live rootsand these values were selected for subsequent models This approach is consistent withfindings by [186768] all of whom demonstrated that in the middle and at the end of thegrowing season the amount of live fine roots is very large in total FRB Dead fine roots wereclassified as non-humic substances in soil organic matter We used the volume of the coresampler as a sample volume for calculations (Equation (1)) Soil material from undisturbedsoil samples (without live fine roots) was dried at 105 C and weighed Then the bulkdensity (BD) was calculated according to [32] The calculation procedure according to [32]excludes coarse particles (Oslash gt 2 mm) as they are not a component of soil bulk density(Equation (1))
BD =masssampleminusmassparticles gt 2 mm
volumesampleminus massparticlesgt2 mmdensityparticlesgt2 mm
(1)
Disturbed bulk soil samples (taken separately in parallel) were air-dried gentlycrushed and sieved through a 2 mm mesh steel sieve Rock fragments (Oslash gt 2 mm) weredetermined by weighing Based on this measurement the content of rock fragmentsdetermined during fieldwork (see above) was calibrated Soil properties for fine soil(Oslash lt 2 mm) and litter subsamples were determined The soil and litter pH were measuredpotentiometrically in distilled water (125) [69] using a pH meter (Elmetron CPI-505) with aglass electrode (Elmetron ERH-11) The concentration of total carbon (TC) and nitrogen (N)was determined in triplicate (and then averaged) using dry combustion gas chromatographwith a CHN analyzer (Elementar vario MICRO cube elemental analyzer) Due to theabsence of carbonates in the studied soils it was assumed that TC corresponds to SOC [70]The SOC stock was herein calculated according to [32] The soil texture was determinedusing the combined sieving and hydrometer method [71]
Due to the need to carry out an analysis for comparable values for further analysisthe studied SOC was converted into SOM using the conversion factor 1724 [72] as precisedata (obtained via experimental research) allowing the conversion of wAGB CRB andFRB into the OC pool remain unavailable
24 Stand Characteristics and Coarse Root Biomass Estimation
We obtained data on tree species and DBH (diameter at breast height) for all treeswith DBH gt 7 cm This approach omits small trees and shrubs however their biomassyields only a small contribution to total biomass as biomass increases exponentially withDBH [21] These data had been previously collected as part of NFI by the Polish ForestResearch Institute and were verified in the course of our fieldwork in 2017 (June to October)The wAGB was calculated using biomass equations based on meta-analysis (Equation (2))for European tree species [27] basing on DBH (cm)
ln (AGB) = ln (β0) + β1ln(d) + ε (2)
where AGBmdashwoody aboveground biomass (kg) β0 β1mdashmodel parameters (coefficients)beechmdashβ0 = minus16594 β1 = 23589 sprucemdashβ0 = minus18865 β1 = 23034 firmdashβ0 = minus23958β1 = 24497 other coniferous treesmdashβ0 = minus16173 β1 = 23177 other deciduous treesmdash
Forests 2021 12 823 6 of 27
β0 = minus11972 β1 = 23417 εmdashan error that is assumed to be normally distributed with amean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
Additionally we calculated the stand density (ind ha-1) The biomass of foliage wascalculated using models provided by [73] for beech and spruce forests (Equation (3))
M = a times Xb (3)
where Mmdashfoliage biomass (Mg ha-1) a bmdashmodel parameters sprucemdasha = 44073b = 03544 beechmdasha = 08813 b = 04151 Xmdashstand basal area (m2 ha-1) Model parametersfor fir forest were obtained from [74] a = minus307447 b = 281850
The biomass of coarse roots (CRB diameter of root gt 2 mm) was calculated for beechspruce and fir separately using general equations (Equation (4)) [27]
ln (CRB) = ln (β0) + β1ln (d) + ε (4)
where CRBmdashcoarse root biomass (kg) β0 β1mdashmodel parameters sprucemdashβ0 = minus37387β1 = 24323 beechmdashβ0 = minus33713 β1 = 23424 firmdashβ0 = minus40287 β1 = 24957 otherspeciesmdashβ0 = minus31677 β1 = 22796 εmdashan error that is assumed to be normally distributedwith a mean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
25 Data Analysis
We employed machine learning techniques to determine the effects of selected bioticforest stand characteristics and abiotic factors on FRB and SOM stocks (Table 2) Owingto a large number of predictions and interactions among them machine learning andexplanatory tools [75] enable the generation of a clear description of the meaning of eachgiven prediction and its partial dependence From different machine learning algorithmswe selected a random forest algorithmmdasha technique based on multiple decisions or treeclassifications which increases model stability and accuracy [76] The random forestmodel has been successfully used for vegetation modeling [77] We used R software [78](version 402 ldquoTaking Off Againrdquo R Core Team 2019) for data analysis All models weredeveloped using the caret package [79] for model development and the DALEX packagefor the visual explanation and exploration of predictive models [75] To understand howthe said models work we used variable importance and partial dependence plots Wequantified the variable importance using the drop-out loss function ie loss-decreaseroot mean square error (RMSE) This function informs us how much the model RMSEincreases when a particular variable is shuffled within a given dataset Literally it showshow much model quality we lose when we replace a particular variable with its randompermutations [75] The higher drop in losses in the RMSE indicates the higher importanceof the variable for proper prediction The partial dependence plots show the change in themodel output along with an increasing value of a particular predictor assuming all othervariables at the constant (mean) level [75] Literally this is the prediction of the modelusing all variables at the constant level and manipulating only the variable of interestSuch an approach simulates the experimental rule ceteris paribus which means lsquoall equalwithout the single manipulatedrsquo and allows to see how FRB and SOM stocks vary alongeach of the predictors without changing other variables These marginal predictions allowone to make conclusions about the explicit impact of a particular variable
Forests 2021 12 823 7 of 27
Table 2 List of predictors
Abbreviation Predictor Unit
wAGB Woody aboveground biomass Mg haminus1
StDens Stand density ind haminus1
foliage Foliage biomass Mg haminus1
age Age of dominant species yearslitter Litter mass Mg haminus1
pH_lit Litter pH (in H2O) dimensionlessCN_lit Organic CarbonNitrogen ratio in litter dimensionless
MAT Mean annual air temperature CMAP Mean annual precipitation mmelev Elevation m asl
slope Hillslope gradient
aspect Hillslope exposure
SOM Soil Organic Matter stock Mg haminus1
CN Organic CarbonNitrogen ratio in soil dimensionlessClay Clay content Silt Silt content pH Soil pH (in H2O) dimensionlessCR Coarse roots biomass Mg haminus1
FRB Fine roots biomass Mg haminus1
mean values at 0 to 40 cm depth
To assess the FRB and SOM we developed separate regression models for eachexamined dominant tree species stand and training models to decrease RMSE usingthe caret package [79] To demonstrate the modelrsquos quality we provided RMSE andr2mdashcoefficient of determination indicating the proportions of variability explained bythe model
3 Results31 Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
The analyzed forest stand characteristics varied between study plots and betweentree species (Tables 3ndash5) Similar findings were also obtained for climatic and morphologicpredictors as well as the herein analyzed soil property predictors (Table A1) Both themorphology and properties of the investigated soils were very similar (Figure A1) Thetexture was loamy throughout the studied soil profiles (silt loam sandy loam and loam)(Table S1 from Supplementary Materials) The soil structure was subangular or angularblocky transitioning into massive in the bottom part of the soil In addition the soilconsistence changed gradually with the depth of the soil profile from soft in the uppermosthorizons to very hard in the lowermost part of the soil (Table 6) The gradual decreasein SOC content with soil depth occurred in all the study plots (Figure 2) According totree species groups the studied plotsrsquo soil texture varied slightlymdashthe highest silt and claycontent was observed under fir-dominated forests while the lowest silt content was underbeech forest and clay content under spruce forest (Table A1)
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
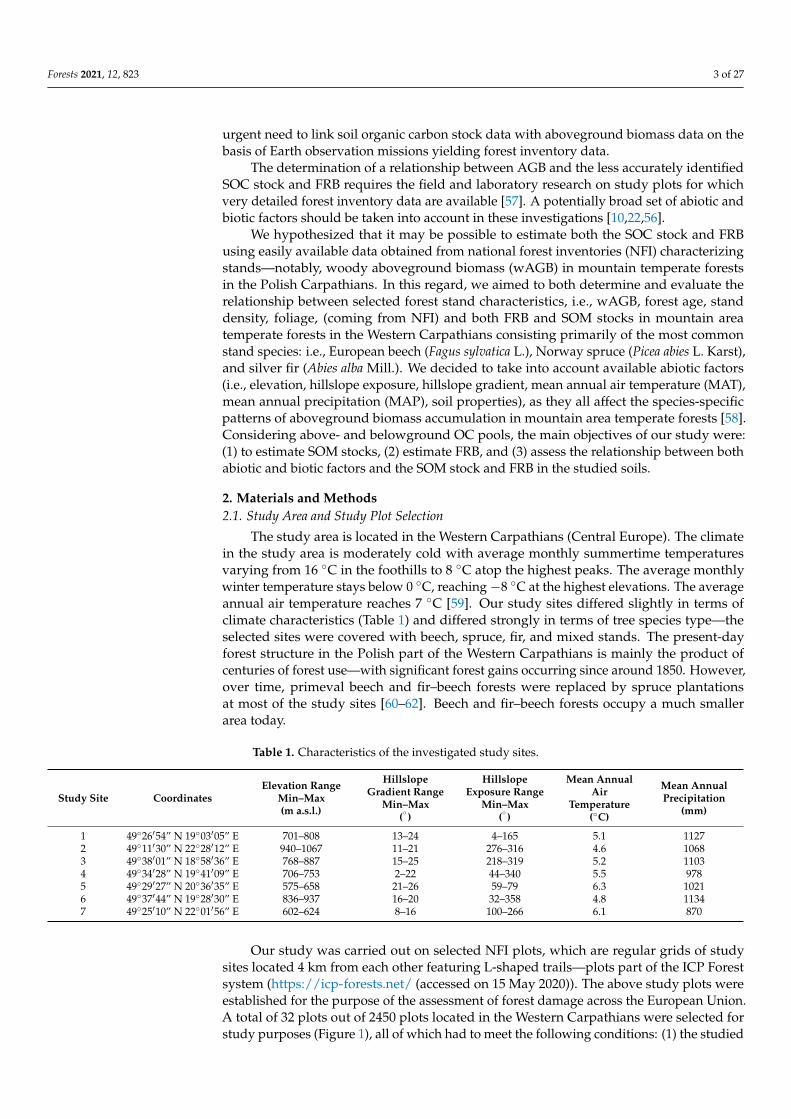
Forests 2021 12 823 3 of 27
urgent need to link soil organic carbon stock data with aboveground biomass data on thebasis of Earth observation missions yielding forest inventory data
The determination of a relationship between AGB and the less accurately identifiedSOC stock and FRB requires the field and laboratory research on study plots for whichvery detailed forest inventory data are available [57] A potentially broad set of abiotic andbiotic factors should be taken into account in these investigations [102256]
We hypothesized that it may be possible to estimate both the SOC stock and FRBusing easily available data obtained from national forest inventories (NFI) characterizingstandsmdashnotably woody aboveground biomass (wAGB) in mountain temperate forestsin the Polish Carpathians In this regard we aimed to both determine and evaluate therelationship between selected forest stand characteristics ie wAGB forest age standdensity foliage (coming from NFI) and both FRB and SOM stocks in mountain areatemperate forests in the Western Carpathians consisting primarily of the most commonstand species ie European beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and silver fir (Abies alba Mill) We decided to take into account available abiotic factors(ie elevation hillslope exposure hillslope gradient mean annual air temperature (MAT)mean annual precipitation (MAP) soil properties) as they all affect the species-specificpatterns of aboveground biomass accumulation in mountain area temperate forests [58]Considering above- and belowground OC pools the main objectives of our study were(1) to estimate SOM stocks (2) estimate FRB and (3) assess the relationship between bothabiotic and biotic factors and the SOM stock and FRB in the studied soils
2 Materials and Methods21 Study Area and Study Plot Selection
The study area is located in the Western Carpathians (Central Europe) The climatein the study area is moderately cold with average monthly summertime temperaturesvarying from 16 C in the foothills to 8 C atop the highest peaks The average monthlywinter temperature stays below 0 C reachingminus8 C at the highest elevations The averageannual air temperature reaches 7 C [59] Our study sites differed slightly in terms ofclimate characteristics (Table 1) and differed strongly in terms of tree species typemdashtheselected sites were covered with beech spruce fir and mixed stands The present-dayforest structure in the Polish part of the Western Carpathians is mainly the product ofcenturies of forest usemdashwith significant forest gains occurring since around 1850 Howeverover time primeval beech and firndashbeech forests were replaced by spruce plantationsat most of the study sites [60ndash62] Beech and firndashbeech forests occupy a much smallerarea today
Table 1 Characteristics of the investigated study sites
Study Site CoordinatesElevation Range
MinndashMax(m asl)
HillslopeGradient Range
MinndashMax( )
HillslopeExposure Range
MinndashMax( )
Mean AnnualAir
Temperature(C)
Mean AnnualPrecipitation
(mm)
1 4926prime54rdquo N 1903prime05rdquo E 701ndash808 13ndash24 4ndash165 51 11272 4911prime30rdquo N 2228prime12rdquo E 940ndash1067 11ndash21 276ndash316 46 10683 4938prime01rdquo N 1858prime36rdquo E 768ndash887 15ndash25 218ndash319 52 11034 4934prime28rdquo N 1941prime09rdquo E 706ndash753 2ndash22 44ndash340 55 9785 4929prime27rdquo N 2036prime35rdquo E 575ndash658 21ndash26 59ndash79 63 10216 4937prime44rdquo N 1928prime30rdquo E 836ndash937 16ndash20 32ndash358 48 11347 4925prime10rdquo N 2201prime56rdquo E 602ndash624 8ndash16 100ndash266 61 870
Our study was carried out on selected NFI plots which are regular grids of studysites located 4 km from each other featuring L-shaped trailsmdashplots part of the ICP Forestsystem (httpsicp-forestsnet (accessed on 15 May 2020)) The above study plots wereestablished for the purpose of the assessment of forest damage across the European UnionA total of 32 plots out of 2450 plots located in the Western Carpathians were selected forstudy purposes (Figure 1) all of which had to meet the following conditions (1) the studied
Forests 2021 12 823 4 of 27
plots had to represent very similar soils ie loamy texture medium depth Cambisolsdeveloped from cover-beds originating in flysch rocks [6364] and (2) all the plots had to besituated in a similar relief position (ie linear hillslope) thus the areal drainage conditionscould be expected to be quite similar (digital elevation model produced as part of Polandrsquosnational GIS project) The selected study plots were located at seven study sites (Table 1)and composed of five circular plots each (r = 12 m) The study plots were located 200 maway from each other in a regular pattern Together all the study sites formed an L-shapedtrail (Figure 1) We excluded three study plots due to the presence of young forest resultingin a lack of accurate tree stand measurements decreasing the accuracy of the biomassassessment [21]
Figure 1 Study area on a map of Europe (A) distribution of study sites (B) (numbers) and distribu-tion of study plots (numbers with letters eg 6A 6B) at each study site (C)
22 Field Survey
Field sampling was conducted in 2017 (June to November) For each study plot a soilprofile was described according to [65] and then sampled (total of 32 soil profiles) Onesoil pit at each study site (plot labeled lsquoCrsquo according to the A to E lettering at Figure 1)was excavated to the lithic contact (seven reference soils) while in the other study plotssoil pits were excavated to approximately 50 cm (25 soil profiles) Rock fragment (coarseparticles diameter gt 2 mm) content in each soil horizon was estimated using standardcharts (templates) commonly used in soil surveys [65] Soil profiles were not established inclose proximity of tree stems (up to 5 m as this distance represented averaged conditionsin the investigated study plots)
A core sampler (100 cm3 in stony soils 635 cm3) was used to collect undisturbedsoil samples (which also incorporated fine roots) using 10 cm intervals (0 to 10 cm 10 to20 cm 20 to 30 cm 30 to 40 cm) at each of the 32 study sites Each sample was collectedin triplicate In addition bulk soil samples (disturbed samples ca 1 kg) from selectedgenetic soil horizons were collected Bulk samples from littermdashdefined as the surface layerof the forest floor for the purpose of the current paper consisting of different fresh non-decomposed or slightly decomposed organic material with recognizable tissuesmdashwerecollected using a metal frame 10 cm x 10 cm taking into account the representativenessof the sampling point Every soil sample was then placed in a sterile polyethylene bag
Forests 2021 12 823 5 of 27
(Whirl-Pakreg) and stored at ~4 C before laboratory analysis The investigated soils wereclassified according to the WRB [66]
23 Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
Litter samples were air-dried crushed and homogenized for further analysis Fromundisturbed soil samples (taken using the core sampler) fine roots were hand-picked anddried to constant mass at 70 C and weighed with an accuracy of 01 g Prior to dryinglive and dead fine roots were distinguished by their color elasticity and degree of barkcohesion [3839] Based on an analysis of 30 of randomly selected samples a share of10 (in mass) of dead fine roots was determined Therefore for the purpose of furtheranalysis it was assumed that 90 of the mass of fine roots in a given sample are live rootsand these values were selected for subsequent models This approach is consistent withfindings by [186768] all of whom demonstrated that in the middle and at the end of thegrowing season the amount of live fine roots is very large in total FRB Dead fine roots wereclassified as non-humic substances in soil organic matter We used the volume of the coresampler as a sample volume for calculations (Equation (1)) Soil material from undisturbedsoil samples (without live fine roots) was dried at 105 C and weighed Then the bulkdensity (BD) was calculated according to [32] The calculation procedure according to [32]excludes coarse particles (Oslash gt 2 mm) as they are not a component of soil bulk density(Equation (1))
BD =masssampleminusmassparticles gt 2 mm
volumesampleminus massparticlesgt2 mmdensityparticlesgt2 mm
(1)
Disturbed bulk soil samples (taken separately in parallel) were air-dried gentlycrushed and sieved through a 2 mm mesh steel sieve Rock fragments (Oslash gt 2 mm) weredetermined by weighing Based on this measurement the content of rock fragmentsdetermined during fieldwork (see above) was calibrated Soil properties for fine soil(Oslash lt 2 mm) and litter subsamples were determined The soil and litter pH were measuredpotentiometrically in distilled water (125) [69] using a pH meter (Elmetron CPI-505) with aglass electrode (Elmetron ERH-11) The concentration of total carbon (TC) and nitrogen (N)was determined in triplicate (and then averaged) using dry combustion gas chromatographwith a CHN analyzer (Elementar vario MICRO cube elemental analyzer) Due to theabsence of carbonates in the studied soils it was assumed that TC corresponds to SOC [70]The SOC stock was herein calculated according to [32] The soil texture was determinedusing the combined sieving and hydrometer method [71]
Due to the need to carry out an analysis for comparable values for further analysisthe studied SOC was converted into SOM using the conversion factor 1724 [72] as precisedata (obtained via experimental research) allowing the conversion of wAGB CRB andFRB into the OC pool remain unavailable
24 Stand Characteristics and Coarse Root Biomass Estimation
We obtained data on tree species and DBH (diameter at breast height) for all treeswith DBH gt 7 cm This approach omits small trees and shrubs however their biomassyields only a small contribution to total biomass as biomass increases exponentially withDBH [21] These data had been previously collected as part of NFI by the Polish ForestResearch Institute and were verified in the course of our fieldwork in 2017 (June to October)The wAGB was calculated using biomass equations based on meta-analysis (Equation (2))for European tree species [27] basing on DBH (cm)
ln (AGB) = ln (β0) + β1ln(d) + ε (2)
where AGBmdashwoody aboveground biomass (kg) β0 β1mdashmodel parameters (coefficients)beechmdashβ0 = minus16594 β1 = 23589 sprucemdashβ0 = minus18865 β1 = 23034 firmdashβ0 = minus23958β1 = 24497 other coniferous treesmdashβ0 = minus16173 β1 = 23177 other deciduous treesmdash
Forests 2021 12 823 6 of 27
β0 = minus11972 β1 = 23417 εmdashan error that is assumed to be normally distributed with amean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
Additionally we calculated the stand density (ind ha-1) The biomass of foliage wascalculated using models provided by [73] for beech and spruce forests (Equation (3))
M = a times Xb (3)
where Mmdashfoliage biomass (Mg ha-1) a bmdashmodel parameters sprucemdasha = 44073b = 03544 beechmdasha = 08813 b = 04151 Xmdashstand basal area (m2 ha-1) Model parametersfor fir forest were obtained from [74] a = minus307447 b = 281850
The biomass of coarse roots (CRB diameter of root gt 2 mm) was calculated for beechspruce and fir separately using general equations (Equation (4)) [27]
ln (CRB) = ln (β0) + β1ln (d) + ε (4)
where CRBmdashcoarse root biomass (kg) β0 β1mdashmodel parameters sprucemdashβ0 = minus37387β1 = 24323 beechmdashβ0 = minus33713 β1 = 23424 firmdashβ0 = minus40287 β1 = 24957 otherspeciesmdashβ0 = minus31677 β1 = 22796 εmdashan error that is assumed to be normally distributedwith a mean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
25 Data Analysis
We employed machine learning techniques to determine the effects of selected bioticforest stand characteristics and abiotic factors on FRB and SOM stocks (Table 2) Owingto a large number of predictions and interactions among them machine learning andexplanatory tools [75] enable the generation of a clear description of the meaning of eachgiven prediction and its partial dependence From different machine learning algorithmswe selected a random forest algorithmmdasha technique based on multiple decisions or treeclassifications which increases model stability and accuracy [76] The random forestmodel has been successfully used for vegetation modeling [77] We used R software [78](version 402 ldquoTaking Off Againrdquo R Core Team 2019) for data analysis All models weredeveloped using the caret package [79] for model development and the DALEX packagefor the visual explanation and exploration of predictive models [75] To understand howthe said models work we used variable importance and partial dependence plots Wequantified the variable importance using the drop-out loss function ie loss-decreaseroot mean square error (RMSE) This function informs us how much the model RMSEincreases when a particular variable is shuffled within a given dataset Literally it showshow much model quality we lose when we replace a particular variable with its randompermutations [75] The higher drop in losses in the RMSE indicates the higher importanceof the variable for proper prediction The partial dependence plots show the change in themodel output along with an increasing value of a particular predictor assuming all othervariables at the constant (mean) level [75] Literally this is the prediction of the modelusing all variables at the constant level and manipulating only the variable of interestSuch an approach simulates the experimental rule ceteris paribus which means lsquoall equalwithout the single manipulatedrsquo and allows to see how FRB and SOM stocks vary alongeach of the predictors without changing other variables These marginal predictions allowone to make conclusions about the explicit impact of a particular variable
Forests 2021 12 823 7 of 27
Table 2 List of predictors
Abbreviation Predictor Unit
wAGB Woody aboveground biomass Mg haminus1
StDens Stand density ind haminus1
foliage Foliage biomass Mg haminus1
age Age of dominant species yearslitter Litter mass Mg haminus1
pH_lit Litter pH (in H2O) dimensionlessCN_lit Organic CarbonNitrogen ratio in litter dimensionless
MAT Mean annual air temperature CMAP Mean annual precipitation mmelev Elevation m asl
slope Hillslope gradient
aspect Hillslope exposure
SOM Soil Organic Matter stock Mg haminus1
CN Organic CarbonNitrogen ratio in soil dimensionlessClay Clay content Silt Silt content pH Soil pH (in H2O) dimensionlessCR Coarse roots biomass Mg haminus1
FRB Fine roots biomass Mg haminus1
mean values at 0 to 40 cm depth
To assess the FRB and SOM we developed separate regression models for eachexamined dominant tree species stand and training models to decrease RMSE usingthe caret package [79] To demonstrate the modelrsquos quality we provided RMSE andr2mdashcoefficient of determination indicating the proportions of variability explained bythe model
3 Results31 Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
The analyzed forest stand characteristics varied between study plots and betweentree species (Tables 3ndash5) Similar findings were also obtained for climatic and morphologicpredictors as well as the herein analyzed soil property predictors (Table A1) Both themorphology and properties of the investigated soils were very similar (Figure A1) Thetexture was loamy throughout the studied soil profiles (silt loam sandy loam and loam)(Table S1 from Supplementary Materials) The soil structure was subangular or angularblocky transitioning into massive in the bottom part of the soil In addition the soilconsistence changed gradually with the depth of the soil profile from soft in the uppermosthorizons to very hard in the lowermost part of the soil (Table 6) The gradual decreasein SOC content with soil depth occurred in all the study plots (Figure 2) According totree species groups the studied plotsrsquo soil texture varied slightlymdashthe highest silt and claycontent was observed under fir-dominated forests while the lowest silt content was underbeech forest and clay content under spruce forest (Table A1)
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 4 of 27
plots had to represent very similar soils ie loamy texture medium depth Cambisolsdeveloped from cover-beds originating in flysch rocks [6364] and (2) all the plots had to besituated in a similar relief position (ie linear hillslope) thus the areal drainage conditionscould be expected to be quite similar (digital elevation model produced as part of Polandrsquosnational GIS project) The selected study plots were located at seven study sites (Table 1)and composed of five circular plots each (r = 12 m) The study plots were located 200 maway from each other in a regular pattern Together all the study sites formed an L-shapedtrail (Figure 1) We excluded three study plots due to the presence of young forest resultingin a lack of accurate tree stand measurements decreasing the accuracy of the biomassassessment [21]
Figure 1 Study area on a map of Europe (A) distribution of study sites (B) (numbers) and distribu-tion of study plots (numbers with letters eg 6A 6B) at each study site (C)
22 Field Survey
Field sampling was conducted in 2017 (June to November) For each study plot a soilprofile was described according to [65] and then sampled (total of 32 soil profiles) Onesoil pit at each study site (plot labeled lsquoCrsquo according to the A to E lettering at Figure 1)was excavated to the lithic contact (seven reference soils) while in the other study plotssoil pits were excavated to approximately 50 cm (25 soil profiles) Rock fragment (coarseparticles diameter gt 2 mm) content in each soil horizon was estimated using standardcharts (templates) commonly used in soil surveys [65] Soil profiles were not established inclose proximity of tree stems (up to 5 m as this distance represented averaged conditionsin the investigated study plots)
A core sampler (100 cm3 in stony soils 635 cm3) was used to collect undisturbedsoil samples (which also incorporated fine roots) using 10 cm intervals (0 to 10 cm 10 to20 cm 20 to 30 cm 30 to 40 cm) at each of the 32 study sites Each sample was collectedin triplicate In addition bulk soil samples (disturbed samples ca 1 kg) from selectedgenetic soil horizons were collected Bulk samples from littermdashdefined as the surface layerof the forest floor for the purpose of the current paper consisting of different fresh non-decomposed or slightly decomposed organic material with recognizable tissuesmdashwerecollected using a metal frame 10 cm x 10 cm taking into account the representativenessof the sampling point Every soil sample was then placed in a sterile polyethylene bag
Forests 2021 12 823 5 of 27
(Whirl-Pakreg) and stored at ~4 C before laboratory analysis The investigated soils wereclassified according to the WRB [66]
23 Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
Litter samples were air-dried crushed and homogenized for further analysis Fromundisturbed soil samples (taken using the core sampler) fine roots were hand-picked anddried to constant mass at 70 C and weighed with an accuracy of 01 g Prior to dryinglive and dead fine roots were distinguished by their color elasticity and degree of barkcohesion [3839] Based on an analysis of 30 of randomly selected samples a share of10 (in mass) of dead fine roots was determined Therefore for the purpose of furtheranalysis it was assumed that 90 of the mass of fine roots in a given sample are live rootsand these values were selected for subsequent models This approach is consistent withfindings by [186768] all of whom demonstrated that in the middle and at the end of thegrowing season the amount of live fine roots is very large in total FRB Dead fine roots wereclassified as non-humic substances in soil organic matter We used the volume of the coresampler as a sample volume for calculations (Equation (1)) Soil material from undisturbedsoil samples (without live fine roots) was dried at 105 C and weighed Then the bulkdensity (BD) was calculated according to [32] The calculation procedure according to [32]excludes coarse particles (Oslash gt 2 mm) as they are not a component of soil bulk density(Equation (1))
BD =masssampleminusmassparticles gt 2 mm
volumesampleminus massparticlesgt2 mmdensityparticlesgt2 mm
(1)
Disturbed bulk soil samples (taken separately in parallel) were air-dried gentlycrushed and sieved through a 2 mm mesh steel sieve Rock fragments (Oslash gt 2 mm) weredetermined by weighing Based on this measurement the content of rock fragmentsdetermined during fieldwork (see above) was calibrated Soil properties for fine soil(Oslash lt 2 mm) and litter subsamples were determined The soil and litter pH were measuredpotentiometrically in distilled water (125) [69] using a pH meter (Elmetron CPI-505) with aglass electrode (Elmetron ERH-11) The concentration of total carbon (TC) and nitrogen (N)was determined in triplicate (and then averaged) using dry combustion gas chromatographwith a CHN analyzer (Elementar vario MICRO cube elemental analyzer) Due to theabsence of carbonates in the studied soils it was assumed that TC corresponds to SOC [70]The SOC stock was herein calculated according to [32] The soil texture was determinedusing the combined sieving and hydrometer method [71]
Due to the need to carry out an analysis for comparable values for further analysisthe studied SOC was converted into SOM using the conversion factor 1724 [72] as precisedata (obtained via experimental research) allowing the conversion of wAGB CRB andFRB into the OC pool remain unavailable
24 Stand Characteristics and Coarse Root Biomass Estimation
We obtained data on tree species and DBH (diameter at breast height) for all treeswith DBH gt 7 cm This approach omits small trees and shrubs however their biomassyields only a small contribution to total biomass as biomass increases exponentially withDBH [21] These data had been previously collected as part of NFI by the Polish ForestResearch Institute and were verified in the course of our fieldwork in 2017 (June to October)The wAGB was calculated using biomass equations based on meta-analysis (Equation (2))for European tree species [27] basing on DBH (cm)
ln (AGB) = ln (β0) + β1ln(d) + ε (2)
where AGBmdashwoody aboveground biomass (kg) β0 β1mdashmodel parameters (coefficients)beechmdashβ0 = minus16594 β1 = 23589 sprucemdashβ0 = minus18865 β1 = 23034 firmdashβ0 = minus23958β1 = 24497 other coniferous treesmdashβ0 = minus16173 β1 = 23177 other deciduous treesmdash
Forests 2021 12 823 6 of 27
β0 = minus11972 β1 = 23417 εmdashan error that is assumed to be normally distributed with amean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
Additionally we calculated the stand density (ind ha-1) The biomass of foliage wascalculated using models provided by [73] for beech and spruce forests (Equation (3))
M = a times Xb (3)
where Mmdashfoliage biomass (Mg ha-1) a bmdashmodel parameters sprucemdasha = 44073b = 03544 beechmdasha = 08813 b = 04151 Xmdashstand basal area (m2 ha-1) Model parametersfor fir forest were obtained from [74] a = minus307447 b = 281850
The biomass of coarse roots (CRB diameter of root gt 2 mm) was calculated for beechspruce and fir separately using general equations (Equation (4)) [27]
ln (CRB) = ln (β0) + β1ln (d) + ε (4)
where CRBmdashcoarse root biomass (kg) β0 β1mdashmodel parameters sprucemdashβ0 = minus37387β1 = 24323 beechmdashβ0 = minus33713 β1 = 23424 firmdashβ0 = minus40287 β1 = 24957 otherspeciesmdashβ0 = minus31677 β1 = 22796 εmdashan error that is assumed to be normally distributedwith a mean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
25 Data Analysis
We employed machine learning techniques to determine the effects of selected bioticforest stand characteristics and abiotic factors on FRB and SOM stocks (Table 2) Owingto a large number of predictions and interactions among them machine learning andexplanatory tools [75] enable the generation of a clear description of the meaning of eachgiven prediction and its partial dependence From different machine learning algorithmswe selected a random forest algorithmmdasha technique based on multiple decisions or treeclassifications which increases model stability and accuracy [76] The random forestmodel has been successfully used for vegetation modeling [77] We used R software [78](version 402 ldquoTaking Off Againrdquo R Core Team 2019) for data analysis All models weredeveloped using the caret package [79] for model development and the DALEX packagefor the visual explanation and exploration of predictive models [75] To understand howthe said models work we used variable importance and partial dependence plots Wequantified the variable importance using the drop-out loss function ie loss-decreaseroot mean square error (RMSE) This function informs us how much the model RMSEincreases when a particular variable is shuffled within a given dataset Literally it showshow much model quality we lose when we replace a particular variable with its randompermutations [75] The higher drop in losses in the RMSE indicates the higher importanceof the variable for proper prediction The partial dependence plots show the change in themodel output along with an increasing value of a particular predictor assuming all othervariables at the constant (mean) level [75] Literally this is the prediction of the modelusing all variables at the constant level and manipulating only the variable of interestSuch an approach simulates the experimental rule ceteris paribus which means lsquoall equalwithout the single manipulatedrsquo and allows to see how FRB and SOM stocks vary alongeach of the predictors without changing other variables These marginal predictions allowone to make conclusions about the explicit impact of a particular variable
Forests 2021 12 823 7 of 27
Table 2 List of predictors
Abbreviation Predictor Unit
wAGB Woody aboveground biomass Mg haminus1
StDens Stand density ind haminus1
foliage Foliage biomass Mg haminus1
age Age of dominant species yearslitter Litter mass Mg haminus1
pH_lit Litter pH (in H2O) dimensionlessCN_lit Organic CarbonNitrogen ratio in litter dimensionless
MAT Mean annual air temperature CMAP Mean annual precipitation mmelev Elevation m asl
slope Hillslope gradient
aspect Hillslope exposure
SOM Soil Organic Matter stock Mg haminus1
CN Organic CarbonNitrogen ratio in soil dimensionlessClay Clay content Silt Silt content pH Soil pH (in H2O) dimensionlessCR Coarse roots biomass Mg haminus1
FRB Fine roots biomass Mg haminus1
mean values at 0 to 40 cm depth
To assess the FRB and SOM we developed separate regression models for eachexamined dominant tree species stand and training models to decrease RMSE usingthe caret package [79] To demonstrate the modelrsquos quality we provided RMSE andr2mdashcoefficient of determination indicating the proportions of variability explained bythe model
3 Results31 Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
The analyzed forest stand characteristics varied between study plots and betweentree species (Tables 3ndash5) Similar findings were also obtained for climatic and morphologicpredictors as well as the herein analyzed soil property predictors (Table A1) Both themorphology and properties of the investigated soils were very similar (Figure A1) Thetexture was loamy throughout the studied soil profiles (silt loam sandy loam and loam)(Table S1 from Supplementary Materials) The soil structure was subangular or angularblocky transitioning into massive in the bottom part of the soil In addition the soilconsistence changed gradually with the depth of the soil profile from soft in the uppermosthorizons to very hard in the lowermost part of the soil (Table 6) The gradual decreasein SOC content with soil depth occurred in all the study plots (Figure 2) According totree species groups the studied plotsrsquo soil texture varied slightlymdashthe highest silt and claycontent was observed under fir-dominated forests while the lowest silt content was underbeech forest and clay content under spruce forest (Table A1)
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 5 of 27
(Whirl-Pakreg) and stored at ~4 C before laboratory analysis The investigated soils wereclassified according to the WRB [66]
23 Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
Litter samples were air-dried crushed and homogenized for further analysis Fromundisturbed soil samples (taken using the core sampler) fine roots were hand-picked anddried to constant mass at 70 C and weighed with an accuracy of 01 g Prior to dryinglive and dead fine roots were distinguished by their color elasticity and degree of barkcohesion [3839] Based on an analysis of 30 of randomly selected samples a share of10 (in mass) of dead fine roots was determined Therefore for the purpose of furtheranalysis it was assumed that 90 of the mass of fine roots in a given sample are live rootsand these values were selected for subsequent models This approach is consistent withfindings by [186768] all of whom demonstrated that in the middle and at the end of thegrowing season the amount of live fine roots is very large in total FRB Dead fine roots wereclassified as non-humic substances in soil organic matter We used the volume of the coresampler as a sample volume for calculations (Equation (1)) Soil material from undisturbedsoil samples (without live fine roots) was dried at 105 C and weighed Then the bulkdensity (BD) was calculated according to [32] The calculation procedure according to [32]excludes coarse particles (Oslash gt 2 mm) as they are not a component of soil bulk density(Equation (1))
BD =masssampleminusmassparticles gt 2 mm
volumesampleminus massparticlesgt2 mmdensityparticlesgt2 mm
(1)
Disturbed bulk soil samples (taken separately in parallel) were air-dried gentlycrushed and sieved through a 2 mm mesh steel sieve Rock fragments (Oslash gt 2 mm) weredetermined by weighing Based on this measurement the content of rock fragmentsdetermined during fieldwork (see above) was calibrated Soil properties for fine soil(Oslash lt 2 mm) and litter subsamples were determined The soil and litter pH were measuredpotentiometrically in distilled water (125) [69] using a pH meter (Elmetron CPI-505) with aglass electrode (Elmetron ERH-11) The concentration of total carbon (TC) and nitrogen (N)was determined in triplicate (and then averaged) using dry combustion gas chromatographwith a CHN analyzer (Elementar vario MICRO cube elemental analyzer) Due to theabsence of carbonates in the studied soils it was assumed that TC corresponds to SOC [70]The SOC stock was herein calculated according to [32] The soil texture was determinedusing the combined sieving and hydrometer method [71]
Due to the need to carry out an analysis for comparable values for further analysisthe studied SOC was converted into SOM using the conversion factor 1724 [72] as precisedata (obtained via experimental research) allowing the conversion of wAGB CRB andFRB into the OC pool remain unavailable
24 Stand Characteristics and Coarse Root Biomass Estimation
We obtained data on tree species and DBH (diameter at breast height) for all treeswith DBH gt 7 cm This approach omits small trees and shrubs however their biomassyields only a small contribution to total biomass as biomass increases exponentially withDBH [21] These data had been previously collected as part of NFI by the Polish ForestResearch Institute and were verified in the course of our fieldwork in 2017 (June to October)The wAGB was calculated using biomass equations based on meta-analysis (Equation (2))for European tree species [27] basing on DBH (cm)
ln (AGB) = ln (β0) + β1ln(d) + ε (2)
where AGBmdashwoody aboveground biomass (kg) β0 β1mdashmodel parameters (coefficients)beechmdashβ0 = minus16594 β1 = 23589 sprucemdashβ0 = minus18865 β1 = 23034 firmdashβ0 = minus23958β1 = 24497 other coniferous treesmdashβ0 = minus16173 β1 = 23177 other deciduous treesmdash
Forests 2021 12 823 6 of 27
β0 = minus11972 β1 = 23417 εmdashan error that is assumed to be normally distributed with amean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
Additionally we calculated the stand density (ind ha-1) The biomass of foliage wascalculated using models provided by [73] for beech and spruce forests (Equation (3))
M = a times Xb (3)
where Mmdashfoliage biomass (Mg ha-1) a bmdashmodel parameters sprucemdasha = 44073b = 03544 beechmdasha = 08813 b = 04151 Xmdashstand basal area (m2 ha-1) Model parametersfor fir forest were obtained from [74] a = minus307447 b = 281850
The biomass of coarse roots (CRB diameter of root gt 2 mm) was calculated for beechspruce and fir separately using general equations (Equation (4)) [27]
ln (CRB) = ln (β0) + β1ln (d) + ε (4)
where CRBmdashcoarse root biomass (kg) β0 β1mdashmodel parameters sprucemdashβ0 = minus37387β1 = 24323 beechmdashβ0 = minus33713 β1 = 23424 firmdashβ0 = minus40287 β1 = 24957 otherspeciesmdashβ0 = minus31677 β1 = 22796 εmdashan error that is assumed to be normally distributedwith a mean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
25 Data Analysis
We employed machine learning techniques to determine the effects of selected bioticforest stand characteristics and abiotic factors on FRB and SOM stocks (Table 2) Owingto a large number of predictions and interactions among them machine learning andexplanatory tools [75] enable the generation of a clear description of the meaning of eachgiven prediction and its partial dependence From different machine learning algorithmswe selected a random forest algorithmmdasha technique based on multiple decisions or treeclassifications which increases model stability and accuracy [76] The random forestmodel has been successfully used for vegetation modeling [77] We used R software [78](version 402 ldquoTaking Off Againrdquo R Core Team 2019) for data analysis All models weredeveloped using the caret package [79] for model development and the DALEX packagefor the visual explanation and exploration of predictive models [75] To understand howthe said models work we used variable importance and partial dependence plots Wequantified the variable importance using the drop-out loss function ie loss-decreaseroot mean square error (RMSE) This function informs us how much the model RMSEincreases when a particular variable is shuffled within a given dataset Literally it showshow much model quality we lose when we replace a particular variable with its randompermutations [75] The higher drop in losses in the RMSE indicates the higher importanceof the variable for proper prediction The partial dependence plots show the change in themodel output along with an increasing value of a particular predictor assuming all othervariables at the constant (mean) level [75] Literally this is the prediction of the modelusing all variables at the constant level and manipulating only the variable of interestSuch an approach simulates the experimental rule ceteris paribus which means lsquoall equalwithout the single manipulatedrsquo and allows to see how FRB and SOM stocks vary alongeach of the predictors without changing other variables These marginal predictions allowone to make conclusions about the explicit impact of a particular variable
Forests 2021 12 823 7 of 27
Table 2 List of predictors
Abbreviation Predictor Unit
wAGB Woody aboveground biomass Mg haminus1
StDens Stand density ind haminus1
foliage Foliage biomass Mg haminus1
age Age of dominant species yearslitter Litter mass Mg haminus1
pH_lit Litter pH (in H2O) dimensionlessCN_lit Organic CarbonNitrogen ratio in litter dimensionless
MAT Mean annual air temperature CMAP Mean annual precipitation mmelev Elevation m asl
slope Hillslope gradient
aspect Hillslope exposure
SOM Soil Organic Matter stock Mg haminus1
CN Organic CarbonNitrogen ratio in soil dimensionlessClay Clay content Silt Silt content pH Soil pH (in H2O) dimensionlessCR Coarse roots biomass Mg haminus1
FRB Fine roots biomass Mg haminus1
mean values at 0 to 40 cm depth
To assess the FRB and SOM we developed separate regression models for eachexamined dominant tree species stand and training models to decrease RMSE usingthe caret package [79] To demonstrate the modelrsquos quality we provided RMSE andr2mdashcoefficient of determination indicating the proportions of variability explained bythe model
3 Results31 Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
The analyzed forest stand characteristics varied between study plots and betweentree species (Tables 3ndash5) Similar findings were also obtained for climatic and morphologicpredictors as well as the herein analyzed soil property predictors (Table A1) Both themorphology and properties of the investigated soils were very similar (Figure A1) Thetexture was loamy throughout the studied soil profiles (silt loam sandy loam and loam)(Table S1 from Supplementary Materials) The soil structure was subangular or angularblocky transitioning into massive in the bottom part of the soil In addition the soilconsistence changed gradually with the depth of the soil profile from soft in the uppermosthorizons to very hard in the lowermost part of the soil (Table 6) The gradual decreasein SOC content with soil depth occurred in all the study plots (Figure 2) According totree species groups the studied plotsrsquo soil texture varied slightlymdashthe highest silt and claycontent was observed under fir-dominated forests while the lowest silt content was underbeech forest and clay content under spruce forest (Table A1)
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 6 of 27
β0 = minus11972 β1 = 23417 εmdashan error that is assumed to be normally distributed with amean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
Additionally we calculated the stand density (ind ha-1) The biomass of foliage wascalculated using models provided by [73] for beech and spruce forests (Equation (3))
M = a times Xb (3)
where Mmdashfoliage biomass (Mg ha-1) a bmdashmodel parameters sprucemdasha = 44073b = 03544 beechmdasha = 08813 b = 04151 Xmdashstand basal area (m2 ha-1) Model parametersfor fir forest were obtained from [74] a = minus307447 b = 281850
The biomass of coarse roots (CRB diameter of root gt 2 mm) was calculated for beechspruce and fir separately using general equations (Equation (4)) [27]
ln (CRB) = ln (β0) + β1ln (d) + ε (4)
where CRBmdashcoarse root biomass (kg) β0 β1mdashmodel parameters sprucemdashβ0 = minus37387β1 = 24323 beechmdashβ0 = minus33713 β1 = 23424 firmdashβ0 = minus40287 β1 = 24957 otherspeciesmdashβ0 = minus31677 β1 = 22796 εmdashan error that is assumed to be normally distributedwith a mean of 0 and standard deviation dmdashDBH (cm) (for more details see [27])
25 Data Analysis
We employed machine learning techniques to determine the effects of selected bioticforest stand characteristics and abiotic factors on FRB and SOM stocks (Table 2) Owingto a large number of predictions and interactions among them machine learning andexplanatory tools [75] enable the generation of a clear description of the meaning of eachgiven prediction and its partial dependence From different machine learning algorithmswe selected a random forest algorithmmdasha technique based on multiple decisions or treeclassifications which increases model stability and accuracy [76] The random forestmodel has been successfully used for vegetation modeling [77] We used R software [78](version 402 ldquoTaking Off Againrdquo R Core Team 2019) for data analysis All models weredeveloped using the caret package [79] for model development and the DALEX packagefor the visual explanation and exploration of predictive models [75] To understand howthe said models work we used variable importance and partial dependence plots Wequantified the variable importance using the drop-out loss function ie loss-decreaseroot mean square error (RMSE) This function informs us how much the model RMSEincreases when a particular variable is shuffled within a given dataset Literally it showshow much model quality we lose when we replace a particular variable with its randompermutations [75] The higher drop in losses in the RMSE indicates the higher importanceof the variable for proper prediction The partial dependence plots show the change in themodel output along with an increasing value of a particular predictor assuming all othervariables at the constant (mean) level [75] Literally this is the prediction of the modelusing all variables at the constant level and manipulating only the variable of interestSuch an approach simulates the experimental rule ceteris paribus which means lsquoall equalwithout the single manipulatedrsquo and allows to see how FRB and SOM stocks vary alongeach of the predictors without changing other variables These marginal predictions allowone to make conclusions about the explicit impact of a particular variable
Forests 2021 12 823 7 of 27
Table 2 List of predictors
Abbreviation Predictor Unit
wAGB Woody aboveground biomass Mg haminus1
StDens Stand density ind haminus1
foliage Foliage biomass Mg haminus1
age Age of dominant species yearslitter Litter mass Mg haminus1
pH_lit Litter pH (in H2O) dimensionlessCN_lit Organic CarbonNitrogen ratio in litter dimensionless
MAT Mean annual air temperature CMAP Mean annual precipitation mmelev Elevation m asl
slope Hillslope gradient
aspect Hillslope exposure
SOM Soil Organic Matter stock Mg haminus1
CN Organic CarbonNitrogen ratio in soil dimensionlessClay Clay content Silt Silt content pH Soil pH (in H2O) dimensionlessCR Coarse roots biomass Mg haminus1
FRB Fine roots biomass Mg haminus1
mean values at 0 to 40 cm depth
To assess the FRB and SOM we developed separate regression models for eachexamined dominant tree species stand and training models to decrease RMSE usingthe caret package [79] To demonstrate the modelrsquos quality we provided RMSE andr2mdashcoefficient of determination indicating the proportions of variability explained bythe model
3 Results31 Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
The analyzed forest stand characteristics varied between study plots and betweentree species (Tables 3ndash5) Similar findings were also obtained for climatic and morphologicpredictors as well as the herein analyzed soil property predictors (Table A1) Both themorphology and properties of the investigated soils were very similar (Figure A1) Thetexture was loamy throughout the studied soil profiles (silt loam sandy loam and loam)(Table S1 from Supplementary Materials) The soil structure was subangular or angularblocky transitioning into massive in the bottom part of the soil In addition the soilconsistence changed gradually with the depth of the soil profile from soft in the uppermosthorizons to very hard in the lowermost part of the soil (Table 6) The gradual decreasein SOC content with soil depth occurred in all the study plots (Figure 2) According totree species groups the studied plotsrsquo soil texture varied slightlymdashthe highest silt and claycontent was observed under fir-dominated forests while the lowest silt content was underbeech forest and clay content under spruce forest (Table A1)
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 7 of 27
Table 2 List of predictors
Abbreviation Predictor Unit
wAGB Woody aboveground biomass Mg haminus1
StDens Stand density ind haminus1
foliage Foliage biomass Mg haminus1
age Age of dominant species yearslitter Litter mass Mg haminus1
pH_lit Litter pH (in H2O) dimensionlessCN_lit Organic CarbonNitrogen ratio in litter dimensionless
MAT Mean annual air temperature CMAP Mean annual precipitation mmelev Elevation m asl
slope Hillslope gradient
aspect Hillslope exposure
SOM Soil Organic Matter stock Mg haminus1
CN Organic CarbonNitrogen ratio in soil dimensionlessClay Clay content Silt Silt content pH Soil pH (in H2O) dimensionlessCR Coarse roots biomass Mg haminus1
FRB Fine roots biomass Mg haminus1
mean values at 0 to 40 cm depth
To assess the FRB and SOM we developed separate regression models for eachexamined dominant tree species stand and training models to decrease RMSE usingthe caret package [79] To demonstrate the modelrsquos quality we provided RMSE andr2mdashcoefficient of determination indicating the proportions of variability explained bythe model
3 Results31 Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
The analyzed forest stand characteristics varied between study plots and betweentree species (Tables 3ndash5) Similar findings were also obtained for climatic and morphologicpredictors as well as the herein analyzed soil property predictors (Table A1) Both themorphology and properties of the investigated soils were very similar (Figure A1) Thetexture was loamy throughout the studied soil profiles (silt loam sandy loam and loam)(Table S1 from Supplementary Materials) The soil structure was subangular or angularblocky transitioning into massive in the bottom part of the soil In addition the soilconsistence changed gradually with the depth of the soil profile from soft in the uppermosthorizons to very hard in the lowermost part of the soil (Table 6) The gradual decreasein SOC content with soil depth occurred in all the study plots (Figure 2) According totree species groups the studied plotsrsquo soil texture varied slightlymdashthe highest silt and claycontent was observed under fir-dominated forests while the lowest silt content was underbeech forest and clay content under spruce forest (Table A1)
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
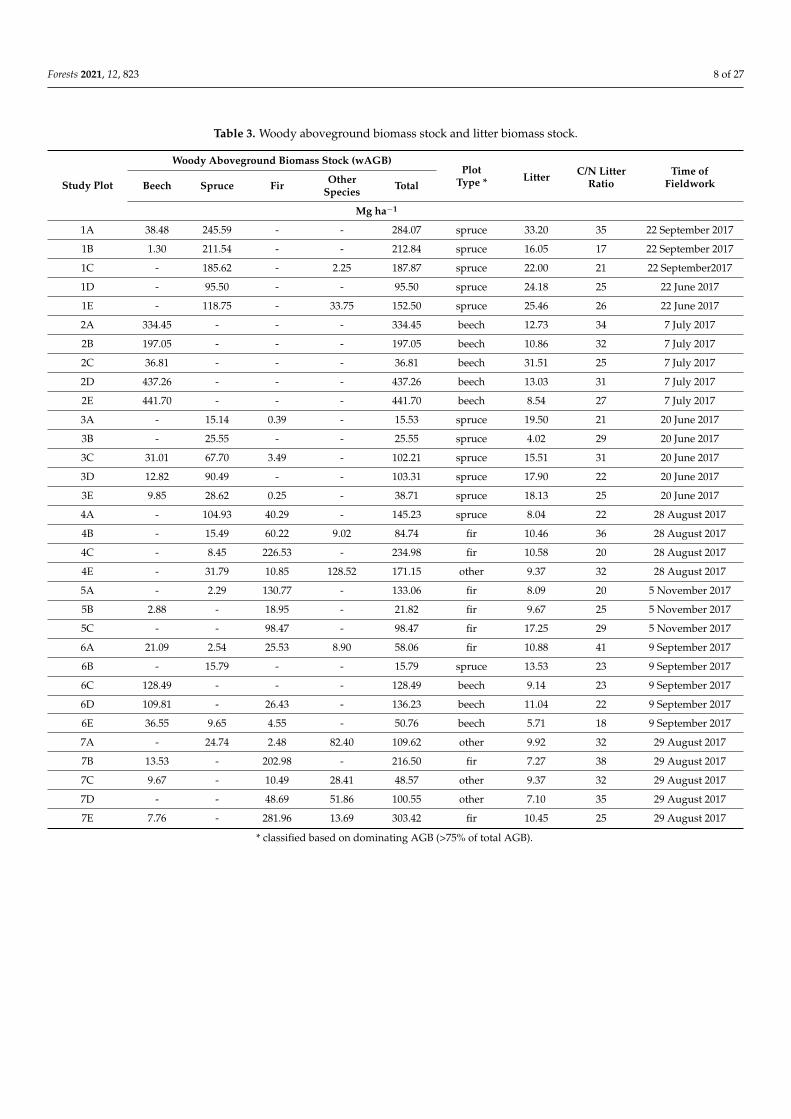
Forests 2021 12 823 8 of 27
Table 3 Woody aboveground biomass stock and litter biomass stock
Study Plot
Woody Aboveground Biomass Stock (wAGB)Plot
Type Litter CN LitterRatio
Time ofFieldworkBeech Spruce Fir Other
Species Total
Mg haminus1
1A 3848 24559 - - 28407 spruce 3320 35 22 September 2017
1B 130 21154 - - 21284 spruce 1605 17 22 September 2017
1C - 18562 - 225 18787 spruce 2200 21 22 September2017
1D - 9550 - - 9550 spruce 2418 25 22 June 2017
1E - 11875 - 3375 15250 spruce 2546 26 22 June 2017
2A 33445 - - - 33445 beech 1273 34 7 July 2017
2B 19705 - - - 19705 beech 1086 32 7 July 2017
2C 3681 - - - 3681 beech 3151 25 7 July 2017
2D 43726 - - - 43726 beech 1303 31 7 July 2017
2E 44170 - - - 44170 beech 854 27 7 July 2017
3A - 1514 039 - 1553 spruce 1950 21 20 June 2017
3B - 2555 - - 2555 spruce 402 29 20 June 2017
3C 3101 6770 349 - 10221 spruce 1551 31 20 June 2017
3D 1282 9049 - - 10331 spruce 1790 22 20 June 2017
3E 985 2862 025 - 3871 spruce 1813 25 20 June 2017
4A - 10493 4029 - 14523 spruce 804 22 28 August 2017
4B - 1549 6022 902 8474 fir 1046 36 28 August 2017
4C - 845 22653 - 23498 fir 1058 20 28 August 2017
4E - 3179 1085 12852 17115 other 937 32 28 August 2017
5A - 229 13077 - 13306 fir 809 20 5 November 2017
5B 288 - 1895 - 2182 fir 967 25 5 November 2017
5C - - 9847 - 9847 fir 1725 29 5 November 2017
6A 2109 254 2553 890 5806 fir 1088 41 9 September 2017
6B - 1579 - - 1579 spruce 1353 23 9 September 2017
6C 12849 - - - 12849 beech 914 23 9 September 2017
6D 10981 - 2643 - 13623 beech 1104 22 9 September 2017
6E 3655 965 455 - 5076 beech 571 18 9 September 2017
7A - 2474 248 8240 10962 other 992 32 29 August 2017
7B 1353 - 20298 - 21650 fir 727 38 29 August 2017
7C 967 - 1049 2841 4857 other 937 32 29 August 2017
7D - - 4869 5186 10055 other 710 35 29 August 2017
7E 776 - 28196 1369 30342 fir 1045 25 29 August 2017
classified based on dominating AGB (gt75 of total AGB)
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
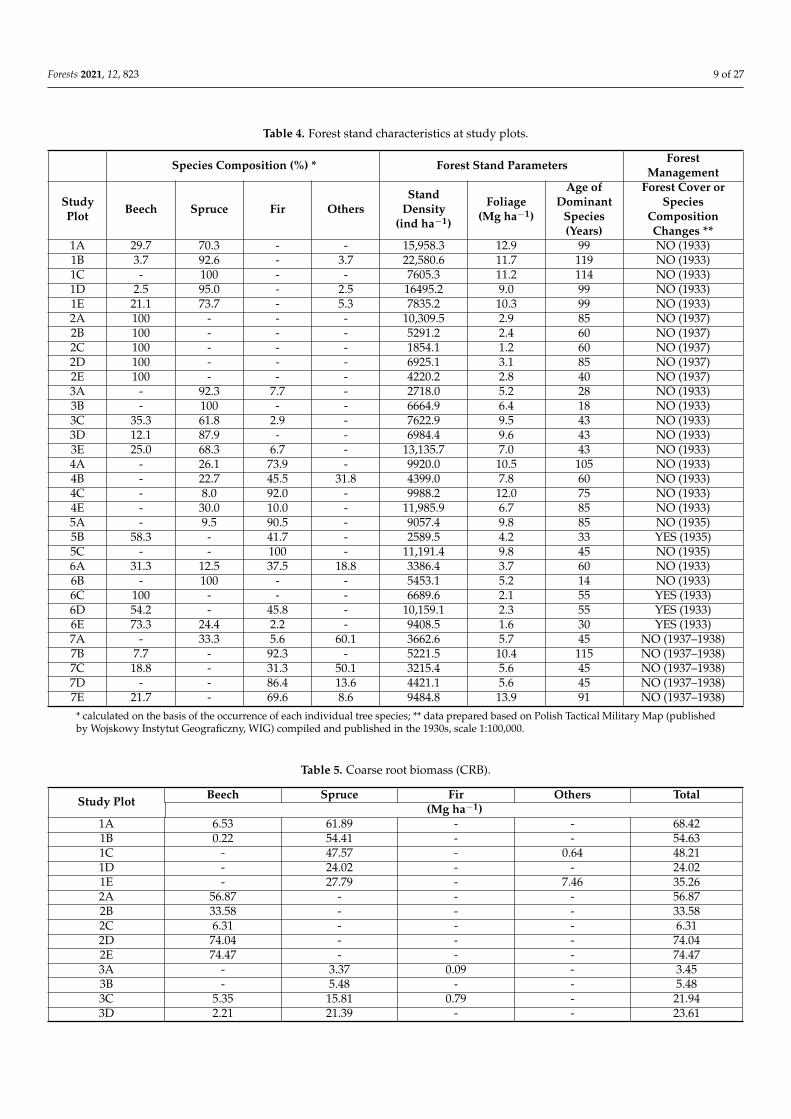
Forests 2021 12 823 9 of 27
Table 4 Forest stand characteristics at study plots
Species Composition () Forest Stand Parameters ForestManagement
StudyPlot Beech Spruce Fir Others
StandDensity
(ind haminus1)
Foliage(Mg haminus1)
Age ofDominant
Species(Years)
Forest Cover orSpecies
CompositionChanges
1A 297 703 - - 159583 129 99 NO (1933)1B 37 926 - 37 225806 117 119 NO (1933)1C - 100 - - 76053 112 114 NO (1933)1D 25 950 - 25 164952 90 99 NO (1933)1E 211 737 - 53 78352 103 99 NO (1933)2A 100 - - - 103095 29 85 NO (1937)2B 100 - - - 52912 24 60 NO (1937)2C 100 - - - 18541 12 60 NO (1937)2D 100 - - - 69251 31 85 NO (1937)2E 100 - - - 42202 28 40 NO (1937)3A - 923 77 - 27180 52 28 NO (1933)3B - 100 - - 66649 64 18 NO (1933)3C 353 618 29 - 76229 95 43 NO (1933)3D 121 879 - - 69844 96 43 NO (1933)3E 250 683 67 - 131357 70 43 NO (1933)4A - 261 739 - 99200 105 105 NO (1933)4B - 227 455 318 43990 78 60 NO (1933)4C - 80 920 - 99882 120 75 NO (1933)4E - 300 100 - 119859 67 85 NO (1933)5A - 95 905 - 90574 98 85 NO (1935)5B 583 - 417 - 25895 42 33 YES (1935)5C - - 100 - 111914 98 45 NO (1935)6A 313 125 375 188 33864 37 60 NO (1933)6B - 100 - - 54531 52 14 NO (1933)6C 100 - - - 66896 21 55 YES (1933)6D 542 - 458 - 101591 23 55 YES (1933)6E 733 244 22 - 94085 16 30 YES (1933)7A - 333 56 601 36626 57 45 NO (1937ndash1938)7B 77 - 923 - 52215 104 115 NO (1937ndash1938)7C 188 - 313 501 32154 56 45 NO (1937ndash1938)7D - - 864 136 44211 56 45 NO (1937ndash1938)7E 217 - 696 86 94848 139 91 NO (1937ndash1938)
calculated on the basis of the occurrence of each individual tree species data prepared based on Polish Tactical Military Map (publishedby Wojskowy Instytut Geograficzny WIG) compiled and published in the 1930s scale 1100000
Table 5 Coarse root biomass (CRB)
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
1A 653 6189 - - 68421B 022 5441 - - 54631C - 4757 - 064 48211D - 2402 - - 24021E - 2779 - 746 35262A 5687 - - - 56872B 3358 - - - 33582C 631 - - - 6312D 7404 - - - 74042E 7447 - - - 74473A - 337 009 - 3453B - 548 - - 5483C 535 1581 079 - 21943D 221 2139 - - 2361
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
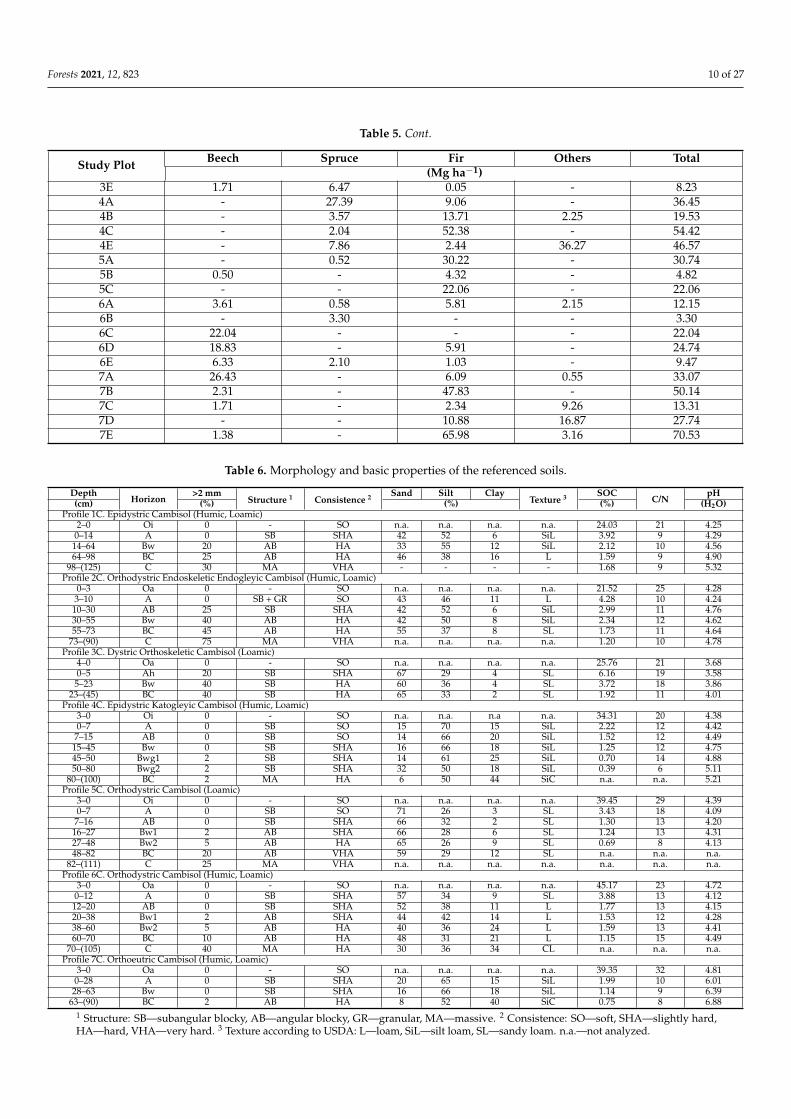
Forests 2021 12 823 10 of 27
Table 5 Cont
Study Plot Beech Spruce Fir Others Total(Mg haminus1)
3E 171 647 005 - 8234A - 2739 906 - 36454B - 357 1371 225 19534C - 204 5238 - 54424E - 786 244 3627 46575A - 052 3022 - 30745B 050 - 432 - 4825C - - 2206 - 22066A 361 058 581 215 12156B - 330 - - 3306C 2204 - - - 22046D 1883 - 591 - 24746E 633 210 103 - 9477A 2643 - 609 055 33077B 231 - 4783 - 50147C 171 - 234 926 13317D - - 1088 1687 27747E 138 - 6598 316 7053
Table 6 Morphology and basic properties of the referenced soils
DepthHorizon
gt2 mmStructure 1 Consistence 2 Sand Silt Clay
Texture 3 SOCCN
pH(cm) () () () (H2O)
Profile 1C Epidystric Cambisol (Humic Loamic)2ndash0 Oi 0 - SO na na na na 2403 21 425
0ndash14 A 0 SB SHA 42 52 6 SiL 392 9 42914ndash64 Bw 20 AB HA 33 55 12 SiL 212 10 45664ndash98 BC 25 AB HA 46 38 16 L 159 9 490
98ndash(125) C 30 MA VHA - - - - 168 9 532Profile 2C Orthodystric Endoskeletic Endogleyic Cambisol (Humic Loamic)
0ndash3 Oa 0 - SO na na na na 2152 25 4283ndash10 A 0 SB + GR SO 43 46 11 L 428 10 42410ndash30 AB 25 SB SHA 42 52 6 SiL 299 11 47630ndash55 Bw 40 AB HA 42 50 8 SiL 234 12 46255ndash73 BC 45 AB HA 55 37 8 SL 173 11 464
73ndash(90) C 75 MA VHA na na na na 120 10 478Profile 3C Dystric Orthoskeletic Cambisol (Loamic)
4ndash0 Oa 0 - SO na na na na 2576 21 3680ndash5 Ah 20 SB SHA 67 29 4 SL 616 19 358
5ndash23 Bw 40 SB HA 60 36 4 SL 372 18 38623ndash(45) BC 40 SB HA 65 33 2 SL 192 11 401
Profile 4C Epidystric Katogleyic Cambisol (Humic Loamic)3ndash0 Oi 0 - SO na na na na 3431 20 4380ndash7 A 0 SB SO 15 70 15 SiL 222 12 442
7ndash15 AB 0 SB SO 14 66 20 SiL 152 12 44915ndash45 Bw 0 SB SHA 16 66 18 SiL 125 12 47545ndash50 Bwg1 2 SB SHA 14 61 25 SiL 070 14 48850ndash80 Bwg2 2 SB SHA 32 50 18 SiL 039 6 511
80ndash(100) BC 2 MA HA 6 50 44 SiC na na 521Profile 5C Orthodystric Cambisol (Loamic)
3ndash0 Oi 0 - SO na na na na 3945 29 4390ndash7 A 0 SB SO 71 26 3 SL 343 18 409
7ndash16 AB 0 SB SHA 66 32 2 SL 130 13 42016ndash27 Bw1 2 AB SHA 66 28 6 SL 124 13 43127ndash48 Bw2 5 AB HA 65 26 9 SL 069 8 41348ndash82 BC 20 AB VHA 59 29 12 SL na na na
82ndash(111) C 25 MA VHA na na na na na na naProfile 6C Orthodystric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 4517 23 4720ndash12 A 0 SB SHA 57 34 9 SL 388 13 41212ndash20 AB 0 SB SHA 52 38 11 L 177 13 41520ndash38 Bw1 2 AB SHA 44 42 14 L 153 12 42838ndash60 Bw2 5 AB HA 40 36 24 L 159 13 44160ndash70 BC 10 AB HA 48 31 21 L 115 15 449
70ndash(105) C 40 MA HA 30 36 34 CL na na naProfile 7C Orthoeutric Cambisol (Humic Loamic)
3ndash0 Oa 0 - SO na na na na 3935 32 4810ndash28 A 0 SB SHA 20 65 15 SiL 199 10 60128ndash63 Bw 0 SB SHA 16 66 18 SiL 114 9 639
63ndash(90) BC 2 AB HA 8 52 40 SiC 075 8 6881 Structure SBmdashsubangular blocky ABmdashangular blocky GRmdashgranular MAmdashmassive 2 Consistence SOmdashsoft SHAmdashslightly hardHAmdashhard VHAmdashvery hard 3 Texture according to USDA Lmdashloam SiLmdashsilt loam SLmdashsandy loam namdashnot analyzed
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
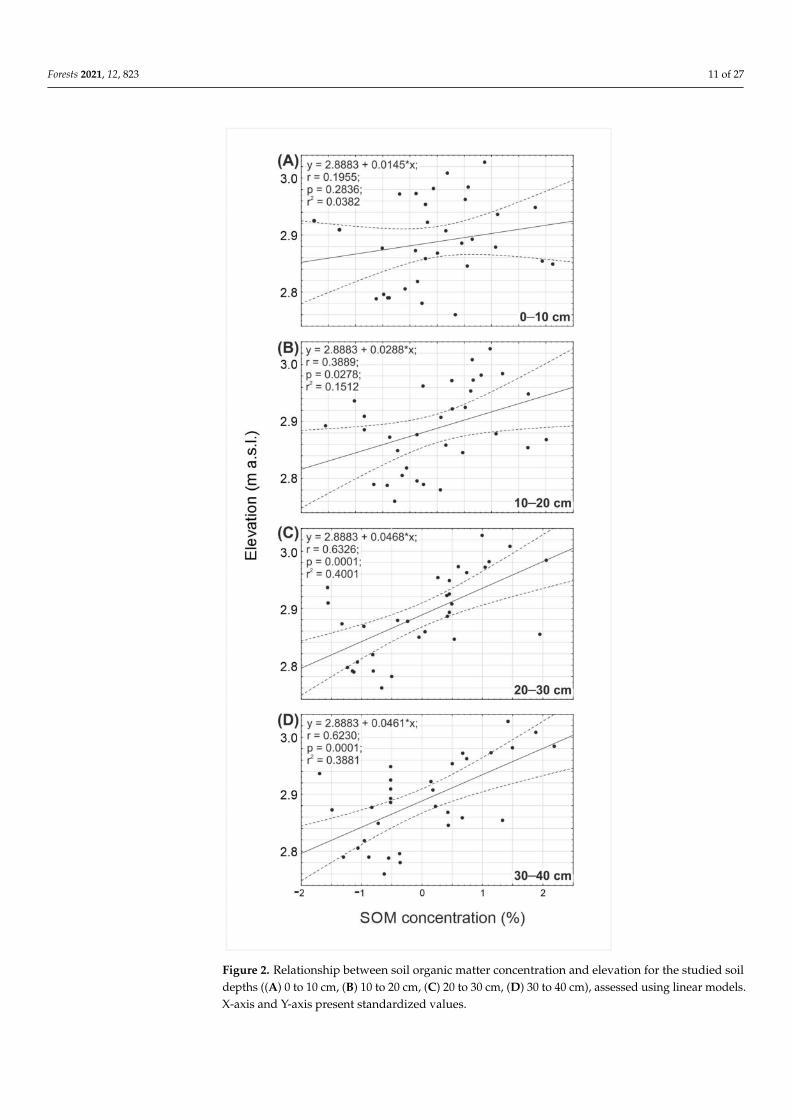
Forests 2021 12 823 11 of 27
Figure 2 Relationship between soil organic matter concentration and elevation for the studied soildepths ((A) 0 to 10 cm (B) 10 to 20 cm (C) 20 to 30 cm (D) 30 to 40 cm) assessed using linear modelsX-axis and Y-axis present standardized values
Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 12 of 27
The FRB and SOM stock varied between the analyzed tree species forest types butwith no significant differences at the 0 to 40 cm depth examined in the study (Table 7)However some differences in FRB content were identified for individual soil layers Forexample in beech stands the FRB content was relatively low in the uppermost part of thesoil (mean 14 Mg haminus1 plusmn SD 12 Mg haminus1 in the 0 to 10 cm layer) and decreased with depth(mean 06 Mg haminus1 plusmn SD 08 Mg haminus1 in the 30 to 40 cm layer) (Figure 3) The FRB in soilsunder spruce forest was much higher than that under beech especially in the 0 to 10 cmsoil layer (Figure 3) It also strongly decreased with the soil layer from 17 Mg haminus1 in the0 to 10 cm layer (mean) to 04 Mg haminus1 in the 30 to 40 cm layer (mean) In fir forest theamount of FRB was rather close to values determined under spruce forest The highest FRBvalues noted across all the sampled soil layers of 0 to 40 cm (Table 7) were detected underfir forest (mean 65 Mg haminus1 plusmn SD 49 Mg haminus1) Slightly lower values were typical forother types of forest (beech and spruce forest) The proportion of FRB in the total biomassstock (wAGB BGB SOM) equaled a maximum of 36 and 17 and 89 in spruce beechand fir forest respectively The highest OC content in fine roots was noted in beech forestsand the lowest in spruce forests (Table 8)
Table 7 FRB and SOM stocks (0 to 40cm) at each plot type
Plot Type Mean Max Min Q1 Q3(Mg haminus1)
beechFRB 32 55 12 12 41SOM 1629 2133 1285 1476 1701
spruce FRB 34 102 00 15 39SOM 1423 2245 566 1242 1626
firFRB 65 138 12 23 110SOM 955 1433 786 793 976
Q1 Q3mdashfirst quartile third quartile
Figure 3 Distribution of fine roots biomass stock at the studied soil depths for the analyzed tree stands expressed usingboxplots The box range denotes the interquartile range line within the boxmdashthe median whiskers indicate the minimumand maximum within range of three times interquartile ranges Points indicate outliers The different letters above the barsindicate statistically significant differences
Table 8 C content () and CN ratio range of FR at each plot type
Plot Type Mean Max Min Q1 Q3
beechC () 470 568 374 405 519CN 46 63 29 39 55
spruce C () 342 478 281 309 342CN 40 52 28 31 47
firC () 407 503 265 322 495CN 40 51 23 32 50
Q1 Q3mdashfirst quartile third quartile
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
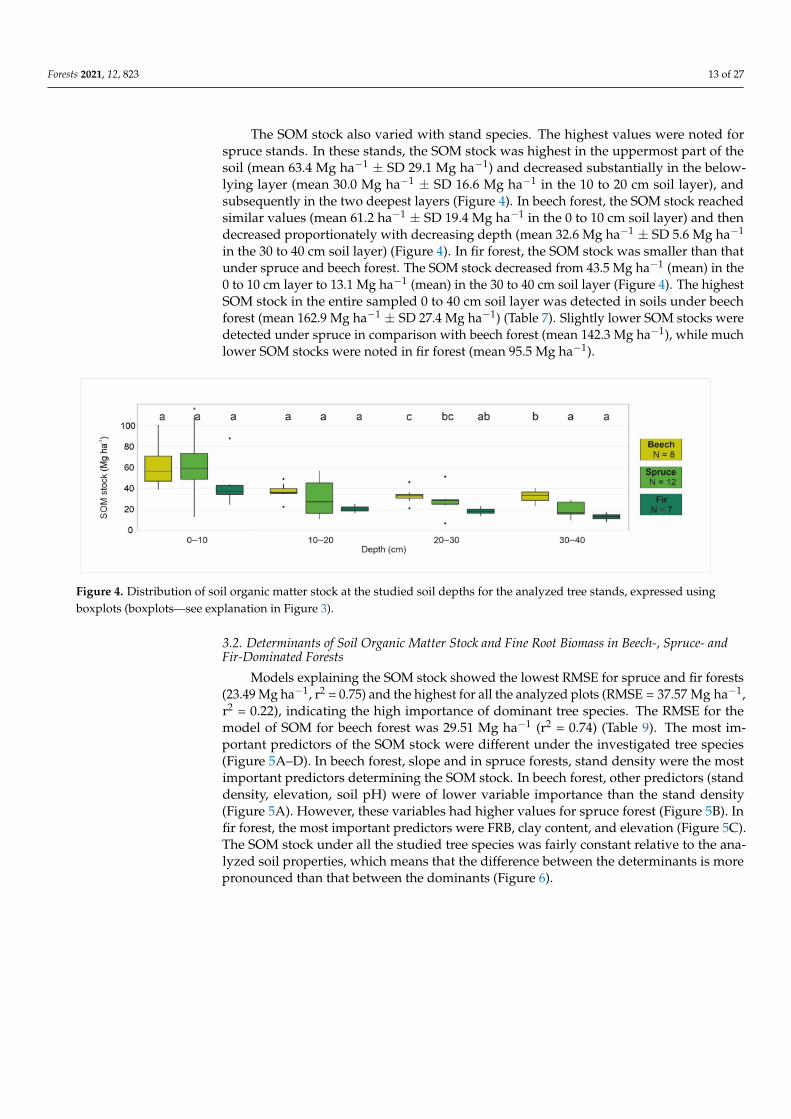
Forests 2021 12 823 13 of 27
The SOM stock also varied with stand species The highest values were noted forspruce stands In these stands the SOM stock was highest in the uppermost part of thesoil (mean 634 Mg haminus1 plusmn SD 291 Mg haminus1) and decreased substantially in the below-lying layer (mean 300 Mg haminus1 plusmn SD 166 Mg haminus1 in the 10 to 20 cm soil layer) andsubsequently in the two deepest layers (Figure 4) In beech forest the SOM stock reachedsimilar values (mean 612 haminus1 plusmn SD 194 Mg haminus1 in the 0 to 10 cm soil layer) and thendecreased proportionately with decreasing depth (mean 326 Mg haminus1 plusmn SD 56 Mg haminus1
in the 30 to 40 cm soil layer) (Figure 4) In fir forest the SOM stock was smaller than thatunder spruce and beech forest The SOM stock decreased from 435 Mg haminus1 (mean) in the0 to 10 cm layer to 131 Mg haminus1 (mean) in the 30 to 40 cm soil layer (Figure 4) The highestSOM stock in the entire sampled 0 to 40 cm soil layer was detected in soils under beechforest (mean 1629 Mg haminus1 plusmn SD 274 Mg haminus1) (Table 7) Slightly lower SOM stocks weredetected under spruce in comparison with beech forest (mean 1423 Mg haminus1) while muchlower SOM stocks were noted in fir forest (mean 955 Mg haminus1)
Figure 4 Distribution of soil organic matter stock at the studied soil depths for the analyzed tree stands expressed usingboxplots (boxplotsmdashsee explanation in Figure 3)
32 Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- andFir-Dominated Forests
Models explaining the SOM stock showed the lowest RMSE for spruce and fir forests(2349 Mg haminus1 r2 = 075) and the highest for all the analyzed plots (RMSE = 3757 Mg haminus1r2 = 022) indicating the high importance of dominant tree species The RMSE for themodel of SOM for beech forest was 2951 Mg haminus1 (r2 = 074) (Table 9) The most im-portant predictors of the SOM stock were different under the investigated tree species(Figure 5AndashD) In beech forest slope and in spruce forests stand density were the mostimportant predictors determining the SOM stock In beech forest other predictors (standdensity elevation soil pH) were of lower variable importance than the stand density(Figure 5A) However these variables had higher values for spruce forest (Figure 5B) Infir forest the most important predictors were FRB clay content and elevation (Figure 5C)The SOM stock under all the studied tree species was fairly constant relative to the ana-lyzed soil properties which means that the difference between the determinants is morepronounced than that between the dominants (Figure 6)
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
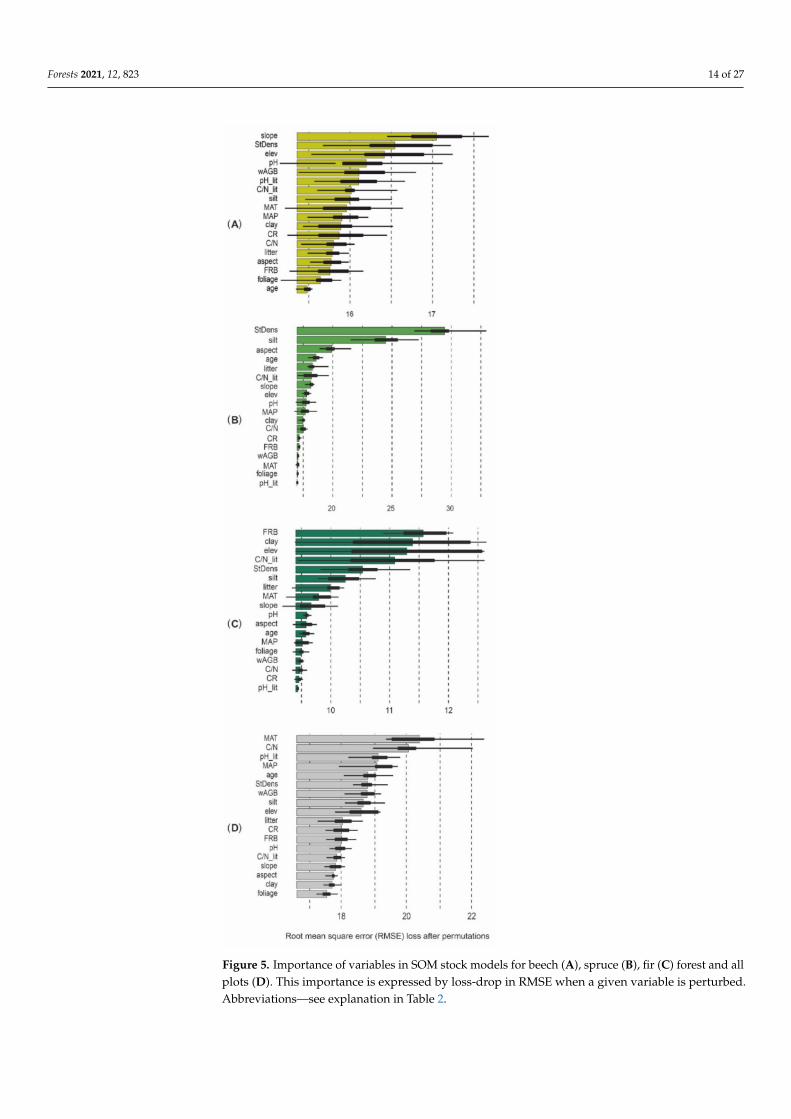
Forests 2021 12 823 14 of 27
Figure 5 Importance of variables in SOM stock models for beech (A) spruce (B) fir (C) forest and allplots (D) This importance is expressed by loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
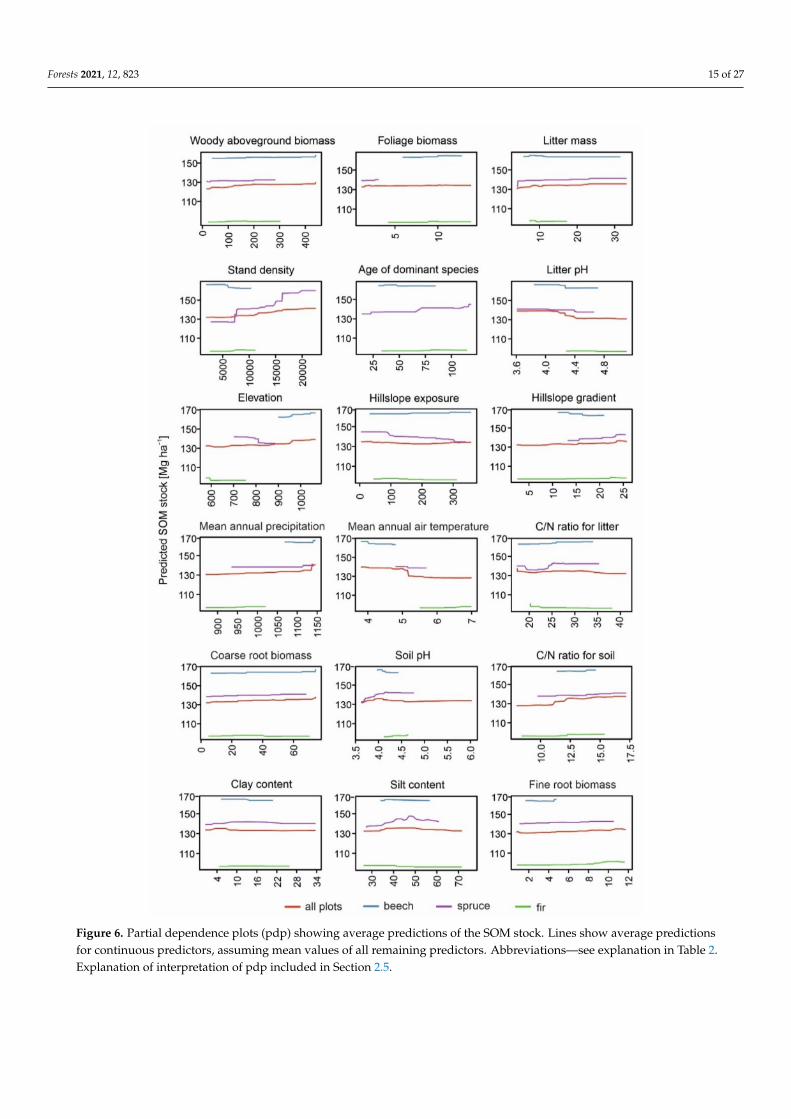
Forests 2021 12 823 15 of 27
Figure 6 Partial dependence plots (pdp) showing average predictions of the SOM stock Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
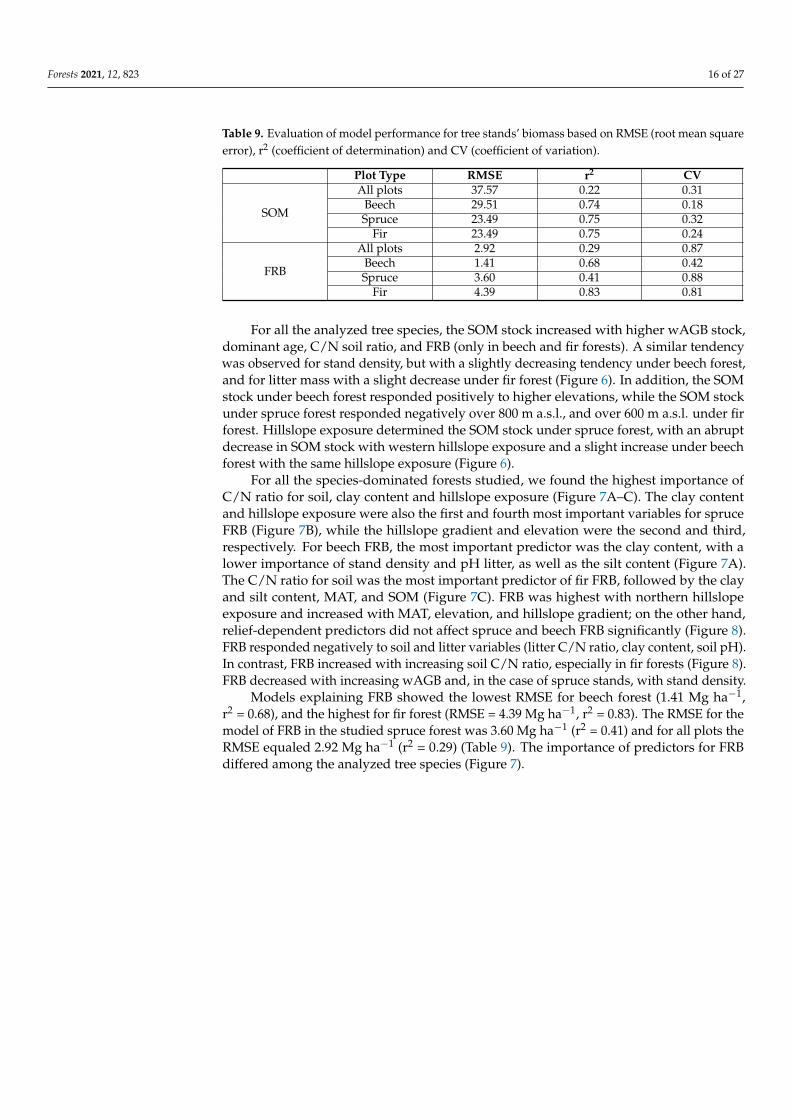
Forests 2021 12 823 16 of 27
Table 9 Evaluation of model performance for tree standsrsquo biomass based on RMSE (root mean squareerror) r2 (coefficient of determination) and CV (coefficient of variation)
Plot Type RMSE r2 CV
SOM
All plots 3757 022 031Beech 2951 074 018Spruce 2349 075 032
Fir 2349 075 024
FRB
All plots 292 029 087Beech 141 068 042Spruce 360 041 088
Fir 439 083 081
For all the analyzed tree species the SOM stock increased with higher wAGB stockdominant age CN soil ratio and FRB (only in beech and fir forests) A similar tendencywas observed for stand density but with a slightly decreasing tendency under beech forestand for litter mass with a slight decrease under fir forest (Figure 6) In addition the SOMstock under beech forest responded positively to higher elevations while the SOM stockunder spruce forest responded negatively over 800 m asl and over 600 m asl under firforest Hillslope exposure determined the SOM stock under spruce forest with an abruptdecrease in SOM stock with western hillslope exposure and a slight increase under beechforest with the same hillslope exposure (Figure 6)
For all the species-dominated forests studied we found the highest importance ofCN ratio for soil clay content and hillslope exposure (Figure 7AndashC) The clay contentand hillslope exposure were also the first and fourth most important variables for spruceFRB (Figure 7B) while the hillslope gradient and elevation were the second and thirdrespectively For beech FRB the most important predictor was the clay content with alower importance of stand density and pH litter as well as the silt content (Figure 7A)The CN ratio for soil was the most important predictor of fir FRB followed by the clayand silt content MAT and SOM (Figure 7C) FRB was highest with northern hillslopeexposure and increased with MAT elevation and hillslope gradient on the other handrelief-dependent predictors did not affect spruce and beech FRB significantly (Figure 8)FRB responded negatively to soil and litter variables (litter CN ratio clay content soil pH)In contrast FRB increased with increasing soil CN ratio especially in fir forests (Figure 8)FRB decreased with increasing wAGB and in the case of spruce stands with stand density
Models explaining FRB showed the lowest RMSE for beech forest (141 Mg haminus1r2 = 068) and the highest for fir forest (RMSE = 439 Mg haminus1 r2 = 083) The RMSE for themodel of FRB in the studied spruce forest was 360 Mg haminus1 (r2 = 041) and for all plots theRMSE equaled 292 Mg haminus1 (r2 = 029) (Table 9) The importance of predictors for FRBdiffered among the analyzed tree species (Figure 7)
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
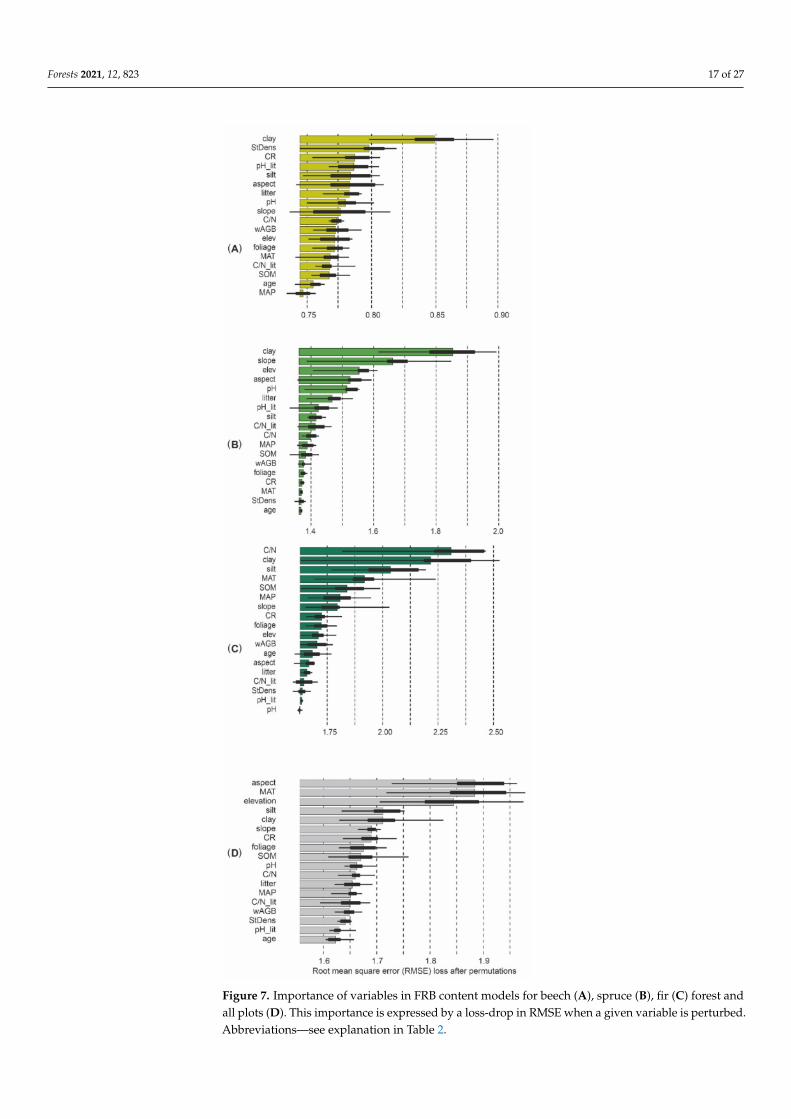
Forests 2021 12 823 17 of 27
Figure 7 Importance of variables in FRB content models for beech (A) spruce (B) fir (C) forest andall plots (D) This importance is expressed by a loss-drop in RMSE when a given variable is perturbedAbbreviationsmdashsee explanation in Table 2
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
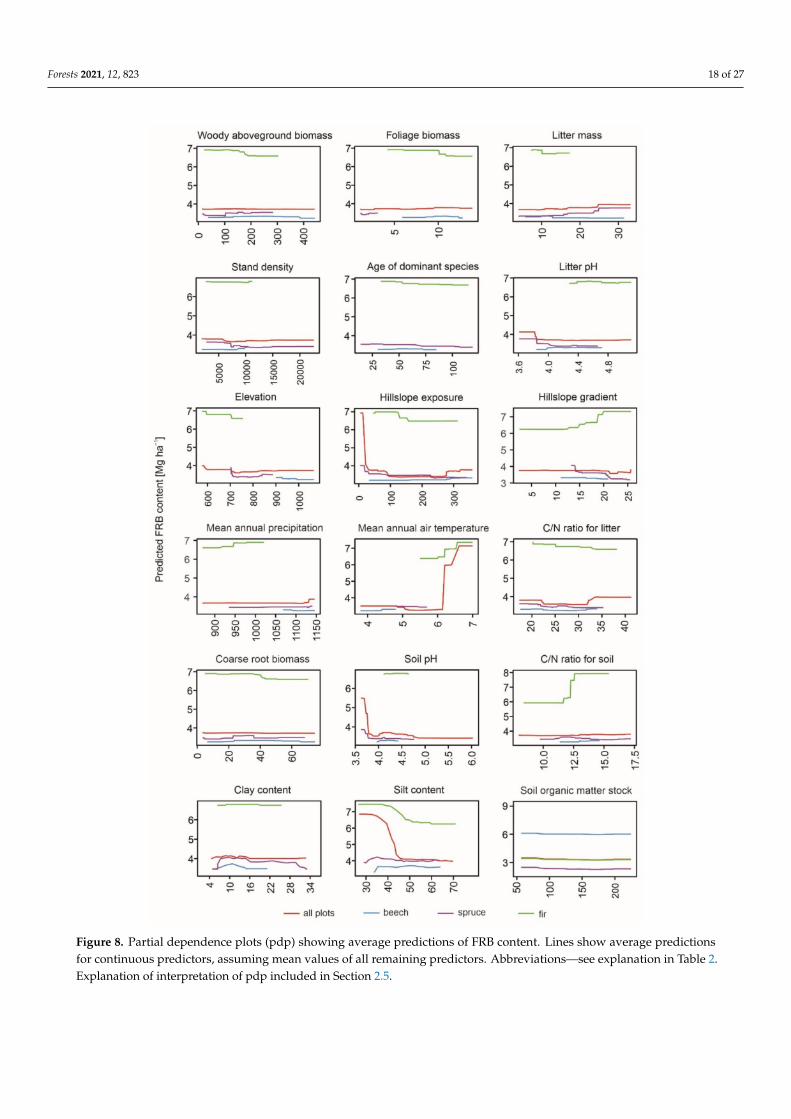
Forests 2021 12 823 18 of 27
Figure 8 Partial dependence plots (pdp) showing average predictions of FRB content Lines show average predictionsfor continuous predictors assuming mean values of all remaining predictors Abbreviationsmdashsee explanation in Table 2Explanation of interpretation of pdp included in Section 25
Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 19 of 27
4 Discussion41 Relationships Between Selected Forest Characteristics and Fine Root Biomass
In our study foliage biomass and stand density were variables with a slight im-pact on FRB (Figure 8) The absence of this relationship confirms the results of previousstudies conducted in various conditions focused on the determination of the relation-ship between aboveground metrics (eg DBH basal area) and FRB both at the tree- andstand-level [1880ndash82] High accuracy was achieved only for a few sites [83] In addition inthe case of the use of stand characteristics obtained from Earth Observation missions toevaluate FRB in the boreal and cool-temperate forestlands in Canada at the stand level aclose relationship was not determined [84] This may be explained by the fact that suchmodels did not reflect temporal and spatial heterogeneity in FRB in various ecosystems [83]Nevertheless considering the significance of FRB in the productivity and biogeochemicalcycles of ecosystems efforts are still pursued to yield reliable estimates using alreadyavailable predictors even though this represents only a small proportion of belowgroundbiomass [188085]
In our study the stand density was found to be a highly important predictor of FRB inbeech forest which confirms a physiological link between FRB that has to take up waterand nutrients the sapwood area that transports the said nutrients and foliage mass wherewater is transpired This is indirectly consistent with the findings of [86] who found thatFRB is stand age-dependent It is noteworthy that the FRB distribution in the soil should berelated to nutrient availability [8788] soil bulk density and pH [8589] and the thickness oforganic soil horizons [90] These findings were confirmed in our study which underscoresthe importance of soil properties affecting FRB (Figure 8) The soils in our study wereselected for their similarity at the study plot scale thus in this case only very detailedmicroscale studies would allow for a better understanding of the reason for any differences
One reason for the absence of a close relationship between the selected forest predictors(foliage biomass stand density wAGB) and FRB in our study may be the impact of site-specific factors (eg soil biological activity present-day and historical natural disturbances)determining FRB production which results in the inapplicability of the same models todifferent study sites and especially those on a larger scale [35] A possible explanation forthe weak effect of the selected forest predictors on FRB in our study may be the fact that theturnover rate of particulate organic matter (to which fine roots belong) may be more thanan order of magnitude faster than that of mineral-associated organic matter as determinedby [91] for subalpine soils in the Swiss Alps It may therefore be assumed that a smallshare of dead fine roots transforms into SOM but most of them are already decomposed
The lower impact of the forest predictors on FRB discussed in our study may be theeffect of the methodology of NFI data collection In the course of forest inventory work(our database) trees with DBH lt 7 cm and herbaceous plants were not taken into accountDue to their small contribution to the overall ABG pool this drawback may be ignoredhowever it is not known whether this may also be possible in the case of FRB
At the same time the FRB sampling methodology used according to [809293] mayhave an effect on the results obtained Collecting root samples in a core samplermdashintriplicatemdashonce in the period of June to November may not take into account the spatialand temporal diversity of fine roots found across the studied plots [18] On the other handthe findings in [94] emphasized the advantages of soil-core methods (cylinders) for thevalidation of other FRB estimation methods therefore it may be assumed that the mostaccurate method was chosen for our research work In the described shallow organicsoil horizons (Table 6 2 to 4 cm) roots were not detected during field description In alllikelihood root biomass found in very deep organic horizons typical of soils in cold areas(ie Finland Sweden Canada) or typical of organic soils (peatlands) will be significantFollowing a review of the abovementioned sources and the results discussed here it maybe concluded that special FRB determination methods should be developed
Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 20 of 27
42 Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
The SOC stock is usually the result of a balance between organic matter input andorganic matter decomposition [1316] and depends on the amount of organic matter previ-ously found in the soil as a result of past soilndashvegetation relationships [10] In our studywe did not find a major impact of the wAGB stock on the SOM stock (Figure 7) Howevera high importance of stand density was noted in finding relationships between stand char-acteristics and the SOM stock Most likely factors other than the present vegetation havehad a decisive impact on the SOM stock in our study (Figure 6) as well as that in [2295]
The key issue here may be organic matter persistence Some studies have shown thepresence of the effect of individual factors on SOM which may vary between differenttree species [19] For example a significant effect of beech forests on higher SOM stockwas noted [1796] which may be explained by the high amount of foliage and litter massdocumented in our study (Table 5) The findings in [97] emphasized possible differences infoliar nutrient concentrations between beech and conifer stands which may affect litterdecomposition rates among species and affect SOC stocks as a consequence In addition thetissue composition of roots varies between tree species and may be a factor that determinesSOC [98] Despite a large number of studies the identification of all mechanisms of treespecies composition effects on SOM and SOC stocks remains challenging [999] and requiresmore complex studies
Many studies [9091100] suggest that SOM found in soils in mountain temperateforests is dominated by organic matter formed hundreds or even thousands of years agoIn our study we found indirect evidence of this pattern as a higher correlation betweenelevation and the SOM stock in the deeper parts of the soil versus the subsurface wasdocumented (Figure 3) The authors of [16] as well as [5] used the example of forest soils inthe Sudety Mountains in Poland to show that past conversion from beech to spruce forestmay have resulted in an absence of significant differences in soil properties (among otherproperties of the SOM stock) between these two species Here we hypothesize that in atime sequence of spruce forests in our study that have replaced deciduous forests a newequilibrium had not had enough time to establish itself as explicitly suggested in [10] It isalso quite possible that in the case of deciduous forests the relationship between selectedstand characteristics and the SOM stock is always stronger as documented in this study aswell as in several other studies [101102]
It is not at all obvious whether differences in the SOM stock between existing conif-erous and deciduous forests provide a reliable basis for predicting changes in the SOMstock following conversion as we do not know how conversion-sensitive the previouslyaccumulated organic components are [1316103] Only some studies [22] have been ableto show that SOM stock growth may be observed in forests with a 10- to 30-year succes-sion [22] Finally the past is likely more important as it includes the occurrence of naturalphenomena and human impact determining what may be observed in the soil today [104]
43 Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
Many studies have shown an effect of precipitation and temperature on fine root pro-duction (fine root mortality) based on seasonal studies [18105] A variety of interpretationsof the relationship between MAT MAP and FRB have been formulated depending onlocal conditions in study areas In mountain temperate forests this trend may also varybetween tree species due to ecological adaptation sensitivity (eg drought tolerance) [106]and elevation
Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 21 of 27
In our study FRB decreased with soil layer depth (Figure 4) which was consistent withother research studies [8587106] In addition a significant impact of climatic conditions(especially MAT) on FRB was observed (Figure 7D) In other studies climatic factors alsoserved as important regulators of the rate of growth of fine roots [107108] Similarlyto FRB the SOM stock was more strongly affected by climatic factors in our study asshown by the vital role of elevation (Figure 7D) Many studies emphasize the crucial role ofclimatic variation as a factor influencing SOM and SOC stocks [516109110] as lower airtemperatures and greater air moisture may lead to less decomposition of organic matterand thus more biomass accumulation in the soil The same explanation is often applied toother mountain area temperate forests [570111] This tendency was observed for all ourstudy sites without distinguishing between tree species stands However in some casesclimatic differences related to tree species-specific stand composition were identified asimportant factors [9112] Studies on the rate of microbial decomposition in soils occurringin mountain areas including the effect of elevation are relatively rarely conducted Suchresearch was carried out by [111113114] who state that the rate of microbial decompositionof cellulose in soil found in the Carpathians depends on elevationmdashthe higher the elevationthe slower the rate of microbial decomposition however this relationship is not absoluteand may be strongly modified due to changes in the arrangement of the soilndashplant factor(including soil pH)
Other factors associated with relief (ie hillslope exposure and hillslope gradient) alsohad a considerable impact on FRB and SOM stocks in our study (Figures 6 and 8) A greaterimpact of hillslope exposure (in comparison with hillslope gradient) on FRB was observed(Figure 7D) which determines the availability of light regulating plant growth and sitetemperature A similar conclusion was drawn by [115] Additionally [116] emphasizedthe effect of the hillslope gradient on FRB which declines down the hillslope gradient andwas observed to some extent in our study (Figure 8) Finally our study demonstrates thatabiotic factors such as the hillslope gradient and exposure are important in determining thesize of FRB and SOM stocks It should be noted that due to the similarity of the studied soilsand predictable soil depth-related variations in their key properties (pH soil structure SOCcontent) (Table 6) the effects of different soil chemical properties [117] soil erosion [10]and drainage conditions [17118] on the SOM stock may be ruled out
5 Conclusions
Our study provides new data on SOM stocks and FRB for mountain temperate ecosys-tems and discusses their determinants Our hypothesis was partially confirmed at the endof our study It is possible to estimate the SOM stock in mountain temperate forests in theWestern Carpathians consisting primarily of the most common stand speciesmdashEuropeanbeech (Fagus sylvatica L) Norway spruce (Picea abies L Karst) and silver fir (Abies albaMill)mdashusing forest inventory data The estimation error is smaller if the species composi-tion of the forest as well as the impact of abiotic factors are taken into account which wasclearly shown by the root mean square error values
Referring to the specific objectives of our study we found the largest wAGB andSOM stocks in beech-dominated forests We also observed a considerable impact of standdensity (as one of the analyzed variables) on SOM stocks particularly in beech forests Wedetermined significant relationships between relief-dependent factors and SOM stocks forall the sites studied Soil properties (ie soil texture pH CN) affected FRB more stronglythan the SOM stock Thus we have shown that the SOM stock and FRB are affected for themost part by abiotic factors (relief-dependent factors climatic conditions) as well as soilproperties For this reason we recommend accounting for soil climate relief and forestmanagement history in models used to estimate belowground OC stocks However themost important source of differences is the dominant tree species
Accounting for all the above predictors may increase the accuracy of predictions basedon select conversion factors or aboveground-to-belowground biomass ratios and maybetter reflect the site-specific variability of stored OC In this study we showed that FRB
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
Forests 2021 12 823 22 of 27
(as a link between plant and soil) and SOM stocks can be estimated using forest inventorydata supported by various other environmental variables This approach can improve theaccuracy of estimating OC fluxes in forest ecosystems and a forestrsquos ability to sequester OCOur results may be used to decrease the uncertainties associated with the least studied partof the forest ecosystemmdashfine roots and soil
Supplementary Materials The following are available online at httpswwwmdpicomarticle103390f12070823s1 Table S1 Detailed soil properties for each of the studied plots
Author Contributions Conceptualization AZ MD and KO methodology AZ MD ŁM andMKD formal analysis AZ DS and GS investigation AZ data curation AZ ŁM and MKDwritingmdashoriginal draft preparation AZ writingmdashreview and editing MD ŁM MKD DS GSand KO visualization AZ and MD supervision MD and KO funding acquisition AZ andKO All authors have read and agreed to the published version of the manuscript
Funding This research study was funded by the Polish National Science Centre (RS4FOR Project For-est change detection and monitoring using passive and active remote sensing data (No 201519BST1002127) and via Project No UJIGiGPKDSC004779 The study was partially supported bythe Institute of Dendrology of the Polish Academy of Sciences
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement Not applicable
Acknowledgments We wish to thank Ewa Grabska and Katarzyna Staszynska for discussionsand their comments on the manuscript Special thanks are directed to Karolina Piech KarolinaKrzyzanowska Marcin Szwagrzyk and Mateusz Stolarczyk for their assistance with fieldwork andlaboratory analysis AZ gratefully acknowledges Dirk Pflugmacher for his constructive discussionsand dedication in the course of the biomass research effort Jakub Nowosad is acknowledged fordiscussions on data analysis Language editing was carried out by Greg Zebik and Wojciech Zielonka
Conflicts of Interest The authors declare no conflict of interest
Appendix A
Table A1 Descriptive statistics (mean min max Q1 Q3) for studied variables including the division into forest types
Spruce Beech FirPredictor Mean Min Max Q1 Q3 Mean Min Max Q1 Q3 Mean Min Max Q1 Q3
age 7333 1400 11900 3925 10050 5875 3000 8500 5125 6625 7200 3300 11500 5250 8800StDens 1024781 271802 2258058 690453 1384136 685718 185414 1030953 502346 959620 741884 258952 1119139 481029 973649foliage 206 107 310 148 248 989 590 1284 873 1183 969 420 1390 879 1118AGB 11493 1553 28407 3542 16134 22034 3681 44170 10906 36015 15614 2182 30342 9160 22574litter 1736 402 3320 1364 2254 1282 571 3151 899 1281 1054 727 1725 888 1052
CN_lit 2490 1732 3527 2148 2676 2640 1751 3396 2260 3136 2775 2024 3821 2256 3266pH_lit 427 361 466 416 440 426 384 472 422 428 462 428 511 439 476
CR 2775 330 6842 754 3939 3769 631 7447 1890 6116 3603 482 7053 2080 5228FRB 341 001 1022 155 391 324 122 547 250 409 648 122 1380 230 1102SOM 14086 5662 22451 12420 16261 16293 12854 21326 14760 17013 9555 7860 14333 7932 9759CN 1389 973 1720 1234 1609 1276 1136 1465 1243 1308 1187 840 1540 1078 1291pH 408 362 477 382 419 418 396 443 406 431 439 411 464 424 461
aspect 19392 437 34016 11348 28115 27325 3197 35852 27688 31771 13914 4460 31247 6785 20094slope 1997 1317 2553 1732 2254 1718 1110 2087 1574 1909 1458 245 2575 662 2213
elevation 77810 70114 88725 72083 81829 96318 89951 106685 93267 97880 65412 57564 75332 61476 70189MAP 110600 93500 114000 110300 113100 110825 106800 114600 106800 113400 94200 87000 102100 90450 96800MAT 514 478 574 499 523 451 381 478 449 478 625 554 697 591 664clay 525 3275 1079 675 1138 475 1975 947 669 1106 525 2500 1346 700 1738silt 2800 5925 4033 3269 4575 3575 6425 4841 4406 5169 2650 7100 4982 3888 6025
Abbreviations explained in Table 2
Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 23 of 27
Figure A1 Exemplary reference soil profiles Numbers refers to study plots and study sites distribu-tion presented at Figure 1 Soil profile depths 1Cmdash125 cm 4Cmdash130 cm 6Cmdash140 cm 3Cmdash105 cm7Cmdash150 cm 5Cmdash105 cm
References1 Nave LE Domke GM Hofmeister KL Mishra U Perry CH Walters BF Swanston CW Reforestation can sequester
Table Proc Natl Acad Sci USA 2018 115 2776ndash2781 [CrossRef]2 Pan Y Birdsey RA Fang J Houghton R Kauppi PE Kurz WA Phillips OL Shvidenko A Lewis SL Canadell JG
et al A large and persistent carbon sink in the worldrsquos forests Science 2011 333 988ndash993 [CrossRef]3 Dixon RK Brown S Houghton RA Solomon AM Trexler MC Wisniewski J Carbon pools and flux of global forest
ecosystems Science 1994 263 185ndash190 [CrossRef] [PubMed]4 Lal R Soil Carbon Sequestration Impacts on Global Climate Change and Food Security Science 2004 304 1623ndash1627 [CrossRef]5 Bojko O Kabala C Organic carbon pools in mountain soilsmdashSources of variability and predicted changes in relation to climate
and land use changes Catena 2017 149 209ndash220 [CrossRef]6 Dieleman WIJ Venter M Ramachandra A Krockenberger AK Bird MI Soil carbon stocks vary predictably with altitude
in tropical forests Implications for soil carbon storage Geoderma 2013 204 59ndash67 [CrossRef]7 Liu S Zhang ZB Li DM Hallett PD Zhang GL Peng XH Temporal dynamics and vertical distribution of newly-derived
carbon from a C3C4 conversion in an Ultisol after 30-yr fertilization Geoderma 2019 337 1077ndash1085 [CrossRef]8 Schepaschenko D Moltchanova E Shvidenko A Blyshchyk V Dmitriev E Martynenko O See L Kraxner F Improved
Estimates of Biomass Expansion Factors for Russian Forests Forests 2018 9 312 [CrossRef]9 Vesterdal L Clarke N Sigurdsson BD Gundersen P Do tree species influence soil carbon stocks in temperate and boreal
forests For Ecol Manag 2013 309 4ndash18 [CrossRef]10 Wasak K Drewnik M Land use effects on soil organic carbon sequestration in calcareous Leptosols in former pastureland-a
case study from the Tatra Mountains (Poland) Solid Earth 2015 6 1103ndash1115 [CrossRef]11 Baritz R Seufert G Montanarella L Van Ranst E Carbon concentrations and stocks in forest soils of Europe For Ecol Manag
2010 260 262ndash277 [CrossRef]12 De Vos B Cools N Ilvesniemi H Vesterdal L Vanguelova E Carnicelli S Benchmark values for forest soil carbon stocks in
Europe Results from a large scale forest soil survey Geoderma 2015 251 33ndash46 [CrossRef]13 Jandl R Lindner M Vesterdal L Bauwens B Baritz R Hagedorn F Johnson DW Minkkinen K Byrne KA How
strongly can forest management influence soil carbon sequestration Geoderma 2007 137 253ndash268 [CrossRef]
Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 24 of 27
14 Lehtonen A Palviainen M Ojanen P Kalliokoski T Noumljd P Kukkola M Penttilauml T Maumlkipaumlauml R Leppaumllammi-KujansuuJ Helmisaari H-S Modelling fine root biomass of boreal tree stands using site and stand variables For Ecol Manag 2016 359361ndash369 [CrossRef]
15 Hobbie SE Ogdahl M Chorover J Chadwick O Oleksyn J Zytkowiak R Reich P Tree Species Effects on Soil OrganicMatter Dynamics The Role of Soil Cation Composition Ecosystems 2007 10 999ndash1018 [CrossRef]
16 Galka B Labaz B Bogacz A Bojko O Kabala C Conversion of Norway spruce forests will reduce organic carbon pools inthe mountain soils of SW Poland Geoderma 2014 213 287ndash295 [CrossRef]
17 Gruba P Socha J Błonska E Lasota J Effect of variable soil texture metal saturation of soil organic matter (SOM) and treespecies composition on spatial distribution of SOM in forest soils in Poland Sci Total Environ 2015 521 90ndash100 [CrossRef][PubMed]
18 Konocircpka B Differences in fine root traits between norway spruce (Picea abies [L] Karst) and european beech (Fagus sylvaticaL)mdashA case study in the Kysuckeacute Beskydy Mts J For Sci 2009 55 556ndash566 [CrossRef]
19 Mueller K Eissenstat D Hobbie S Oleksyn J Jagodzinski A Reich B Chadwick O Chorover J Tree species effectson coupled cycles of carbon nitrogen and acidity in mineral soils at a common garden experiment Biogeochemistry 2012 111601ndash614 [CrossRef]
20 Ransedokken Y Asplund J Ohlson M Nybakken L Vertical distribution of soil carbon in boreal forest under European beechand Norway spruce Eur J For Res 2019 138 353ndash361 [CrossRef]
21 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Cysewska A Wierczynska S Maciejczyk K How do treestand parameters affect young Scots pine biomassmdashAllometric equations and biomass conversion and expansion factors ForEcol Manag 2018 409 74ndash83 [CrossRef]
22 Jones IL DeWalt SJ Lopez OR Bunnefeld L Pattison Z Dent DH Above- and belowground carbon stocks are decoupledin secondary tropical forests and are positively related to forest age and soil nutrients respectively Sci Total Environ 2019 697133987 [CrossRef]
23 Baumann M Gasparri I Piquer-Rodriacuteguez M Gavier Pizarro G Griffiths P Hostert P Kuemmerle T Carbon emissionsfrom agricultural expansion and intensification in the Chaco Glob Chang Biol 2017 23 1902ndash1916 [CrossRef]
24 Erb KH Kastner T Plutzar C Bais ALS Carvalhais N Fetzel T Gingrich S Haberl H Lauk C NiedertscheiderM et al Unexpectedly large impact of forest management and grazing on global vegetation biomass Nature 2018 553 73ndash76[CrossRef] [PubMed]
25 Hansen MC High-Resolution Global Maps of Journal Science 2013 850 850ndash854 [CrossRef]26 Tyukavina A Baccini A Hansen MC Potapov PV Stehman SV Houghton RA Krylov AM Turubanova S Goetz
SJ Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012 Environ Res Lett 2015 10 74002[CrossRef]
27 Forrester DI Tachauer IHH Annighoefer P Barbeito I Pretzsch H Ruiz-Peinado R Stark H Vacchiano G Zlatanov TChakraborty T et al Generalized biomass and leaf area allometric equations for European tree species incorporating standstructure tree age and climate For Ecol Manag 2017 396 160ndash175 [CrossRef]
28 Duncanson L Armston J Disney M Avitabile V Barbier N Calders K Carter S Chave J Herold M Crowther TWet al The Importance of Consistent Global Forest Aboveground Biomass Product Validation Surv Geophys 2019 40 979ndash999[CrossRef]
29 Matasci G Hermosilla T Wulder MA White JC Coops NC Hobart GW Zald HSJ Large-area mapping of Canadianboreal forest cover height biomass and other structural attributes using Landsat composites and lidar plots Remote Sens Environ2018 209 90ndash106 [CrossRef]
30 Schwieder M Leitatildeo PJ Pinto JRR Teixeira AMC Pedroni F Sanchez M Bustamante MM Hostert P Landsatphenological metrics and their relation to aboveground carbon in the Brazilian Savanna Carbon Balance Manag 2018 13 7[CrossRef]
31 Wulder MA White JC Nelson RF Naeligsset E Oslashrka HO Coops NC Hilker T Bater CW Gobakken T Lidar samplingfor large-area forest characterization A review Remote Sens Environ 2012 121 196ndash209 [CrossRef]
32 Don A Schumacher J Scherer-Lorenzen M Scholten T Schulze ED Spatial and vertical variation of soil carbon at twograssland sitesmdashImplications for measuring soil carbon stocks Geoderma 2007 141 272ndash282 [CrossRef]
33 Leifeld J Koumlgel-Knabner I Soil organic matter fractions as early indicators for carbon stock changes under different land-useGeoderma 2005 124 143ndash155 [CrossRef]
34 Matamala R Gonzagravelez-Meler MA Jastrow JD Norby RJ Schlesinger WH Impacts of fine root turnover on forest NPP andsoil C sequestration potential Science 2003 302 1385ndash1387 [CrossRef] [PubMed]
35 Fineacuter L Helmisaari HS Lotildehmus K Majdi H Brunner I Boslashrja I Eldhuset T Godbold D Grebenc T Konocircpka B et alVariation in fine root biomass of three European tree species Beech (Fagus sylvatica L) Norway spruce (Picea abies L Karst)and Scots pine (Pinus sylvestris L) Plant Biosyst 2007 141 394ndash405 [CrossRef]
36 Silver WL Miya RK Global patterns in root decomposition Comparisons of climate and litter quality effects Oecologia 2001129 407ndash419 [CrossRef]
37 Don A Scholten T Schulze ED Conversion of cropland into grassland Implications for soil organic-carbon stocks in two soilswith different texture J Plant Nutr Soil Sci 2009 172 53ndash62 [CrossRef]
Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 25 of 27
38 Leuschner C Backes K Hertel D Schipka F Schmitt U Terborg O Runge M Drought responses at leaf stem and fine rootlevels of competitive Fagus sylvatica L and Quercus petraea (Matt) Liebl trees in dry and wet years For Ecol Manag 2001 14933ndash46 [CrossRef]
39 Persson H The distribution and productivity of fine roots in boreal forests Plant Soil 1983 71 87ndash101 [CrossRef]40 Finzi AC Abramoff RZ Spiller KS Brzostek ER Darby BA Kramer MA Phillips RP Rhizosphere processes are
quantitatively important components of terrestrial carbon and nutrient cycles Glob Chang Biol 2015 21 2082ndash2094 [CrossRef]41 Han M Sun L Gan D Fu L Zhu B Root functional traits are key determinants of the rhizosphere effect on soil organic
matter decomposition across 14 temperate hardwood species Soil Biol Biochem 2020 151 108019 [CrossRef]42 Sun L Ataka M Kominami Y Yoshimura K Relationship between fine-root exudation and respiration of two Quercus species
in a Japanese temperate forest Tree Physiol 2017 37 1011ndash1020 [CrossRef]43 Mao R Zeng DH Li L Fresh root decomposition pattern of two contrasting tree species from temperate agroforestry systems
Effects of root diameter and nitrogen enrichment of soil Plant Soil 2011 347 115ndash123 [CrossRef]44 Zhang X Wang W The decomposition of fine and coarse roots Their global patterns and controlling factors Sci Rep 2015 5
[CrossRef] [PubMed]45 Clemmensen KE Bahr A Ovaskainen O Dahlberg A Ekblad A Wallander H Stenlid J Finlay RD Wardle DA
Lindahl BD Roots and associated fungi drive long-term carbon sequestration in boreal forest Science 2013 340 1615ndash1618[CrossRef]
46 Raich JW Clark DA Schwendenmann L Wood TE Aboveground tree growth varies with belowground carbon allocationin a tropical rainforest environment PLoS ONE 2014 9 e100275 [CrossRef] [PubMed]
47 Rasse DP Rumpel C Dignac MF Is soil carbon mostly root carbon Mechanisms for a specific stabilisation Plant Soil 2005269 341ndash356 [CrossRef]
48 Kozłowski TT Pallardy SG Physiology of Woody Plants Academic Press San Diego CA USA 199749 Canarini A Kaiser C Merchant A Richter A Wanek W Root Exudation of Primary Metabolites Mechanisms and Their
Roles in Plant Responses to Environmental Stimuli Front Plant Sci 2019 10 157 [CrossRef]50 McCormack ML Dickie IA Eissenstat DM Fahey TJ Fernandez CW Guo D Helmisaari HS Hobbie EA Iversen
CM Jackson RB et al Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphereprocesses New Phytol 2015 207 505ndash518 [CrossRef]
51 Franklin J Serra-Diaz JM Syphard AD Regan HM Global change and terrestrial plant community dynamics Proc NatlAcad Sci USA 2016 113 3725ndash3734 [CrossRef]
52 Wang X Tang C Severi J Butterly CR Baldock JA Rhizosphere priming effect on soil organic carbon decomposition underplant species differing in soil acidification and root exudation New Phytol 2016 211 864ndash873 [CrossRef]
53 Błonska E Lasota J Gruba P Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation toother properties of soil derived from loess and glaciofluvial sand Ecological Res 2016 31 655ndash664 [CrossRef]
54 Horodecki P Nowinski M Jagodzinski AM Advantages of mixed tree stands in restoration of upper soil layers on postminingsites A five-year leaf liter decomposition experiment Land Degrad Dev 2019 30 3ndash13 [CrossRef]
55 Sjoumlstroumlm E Westermark U Chemical composition of wood and pulps Basic constituents and their distribution In AnalyticalMethods in Wood Chemistry Pulping and Papermaking Sjoumlstroumlm E Aleacuten R Eds Springer Berlin Germany 1999 pp 1ndash19
56 Mayer M Prescott C Abaker W Augusto L Ceacutecillon L Ferreira G James J Jandl R Katzensteiner K Laclau JP et alInfluence of forest management activities on soil organic carbon stocks A knowledge synthesis For Ecol Manag 2020 466[CrossRef]
57 Schepaschenko D Shvidenko A Usoltsev V Lakyda P Luo Y Vasylyshyn R Lakyda I Myklush Y See L McCallum Iet al A dataset of forest biomass structure for Eurasia Sci Data 2017 4 sdata201770 [CrossRef] [PubMed]
58 Dyderski MK Pawlik Ł Spatial distribution of tree species in mountain national parks depends on geomorphology and climateFor Ecol Manag 2020 474 118366 [CrossRef]
59 Wypych A Ustrnul Z Schmatz DR Long-term variability of air temperature and precipitation conditions in the PolishCarpathians J Mt Sci 2018 15 237ndash253 [CrossRef]
60 Grabska E Hostert P Pflugmacher D Ostapowicz K Forest stand species mapping using the sentinel-2 time series RemoteSens 2019 11 1197 [CrossRef]
61 Matiakowska J Szabla K Beskidy bez lasu Magazyn Społeczno-Kulturalny ldquoSlaskrdquo Goacuternoslaskie Towarzystwo LiterackieKatowice Poland 2007
62 Turnock D Ecoregion-based conservation in the Carpathians and the land-use implications Land Use Pol 2002 19 47ndash63[CrossRef]
63 Kacprzak A Szymanski W Wojcik-Tabol P The role of flysch sandstones in forming the properties of cover deposits andsoilsmdashExamples from the Carpathians Z Geomorphol 2015 59 [CrossRef]
64 Skiba S Drewnik M Mapa gleb obszaru Karpat w granicach Polski Rocz Bieszcz 2003 11 15ndash2065 FAO Guidelines for Soil Description 4th ed Food and Agriculture Organization of the United Nations (FAO) Rome Italy 200666 IUSS Working Group WRB World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and
Creating Legends for Soil Maps World Soil Resources Reports No 106 FAO Rome Italy 2014 Update 2015
Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 26 of 27
67 Anderson LJ Comas LH Lakso AN Eissenstat DM Multiple risk factors in root survivorship A four-year study inConcord grape New Phytol 2003 158 489ndash501 [CrossRef]
68 Eissenstat DM Yanai RD The ecology of root lifespan Adv Ecol Res 1997 27 1ndash60 [CrossRef]69 Thomas GW Soil pH and Soil Acidity In Methods of Soil Analysis Part 3 Chemical Methods Sparks DL Page AL Helmke
PA Loeppert RH Eds SSSA and ASA Madison WI USA 1996 pp 475ndash49070 Egli M Sartori G Mirabella A Giaccai D The effects of exposure and climate on the weathering of late Pleistocene and
Holocene Alpine soils Geomorphology 2010 114 466ndash482 [CrossRef]71 Gee GW Bauder JW Particle-Size Analysis In Methods of Soil Analysis Part 1 Physical and Mineralogical Methods Klute A Ed
ASA-SSSA Madison WI USA 1986 pp 427ndash44572 Pribyl D A critical review of conventional SOC to SOM conversion factor Geoderma 2010 156 75ndash83 [CrossRef]73 Jagodzinski AM Dyderski MK Horodecki P Differences in biomass production and carbon sequestration between highland
and lowland stands of Picea abies (L) H Karst and Fagus sylvatica L For Ecol Manag 2020 474 118329 [CrossRef]74 Jagodzinski AM Dyderski MK Gesikiewicz K Horodecki P Tree and stand level estimations of Abies alba Mill aboveground
biomass Ann For Sci 2019 76 56 [CrossRef]75 Biecek P DALEX Explainers for Complex Predictive Models in R J Mach Learn Res 2018 19 8476 Breiman L Random forests Mach Learn 2001 45 5ndash32 [CrossRef]77 Hengl T Walsh MG Sanderman J Wheeler I Harrison SP Prentice IC Global mapping of potential natural vegetation
An assessment of machine learning algorithms for estimating land potential PeerJ 2018 6 e5457 [CrossRef]78 R Core Team R A Language and Environment for Statistical Computing R Foundation for Statistical Computing Vienna
Austria 201679 Kuhn M Building Predictive Models in R Using the caret Package J Stat Soft 2008 28 1ndash26 [CrossRef]80 Addo-Danso SD Prescott CE Smith AR Methods for estimating root biomass and production in forest and woodland
ecosystem carbon studies A review For Ecol Manag 2016 359 332ndash351 [CrossRef]81 Jurasinski G Jordan A Glatzel S Mapping soil CO2 efflux in an old-growth forest using regression kriging with estimated fine
root biomass as ancillary data For Ecol Manag 2012 263 101ndash113 [CrossRef]82 Mooney HA Chu C Seasonal carbon allocation in Heteromeles arbutifolium a California evergreen shrub Oecologia 1974 14
295ndash306 [CrossRef]83 Helmisaari H Derome J Noumljd P Kukkola M Fine root biomass in relation to site and stand characteristics in Norway spruce
and Scots pine stands Tree Physiol 2007 27 1493ndash1504 [CrossRef]84 Chen W Zhang Q Cihlar J Bauhus J Price DT Estimating fine-root biomass and production of boreal and cool temperate
forests using aboveground measurements A new approach Plant Soil 2004 265 31ndash46 [CrossRef]85 Kirfel K Heinze S Hertel D Leuschner C Effects of bedrock type and soil chemistry on the fine roots of European beechmdashA
study on the belowground plasticity of trees For Ecol Manag 2019 444 256ndash268 [CrossRef]86 Jagodzinski AM Zioacutełkowski J Warnkowska A Prais H Tree Age Effects on Fine Root Biomass and Morphology over
Chronosequences of Fagus sylvatica Quercus robur and Alnus glutinosa Stands PLoS ONE 2016 11 e0148668 [CrossRef]87 Jobbagy EG Jackson RB The vertical distribution of soil organic carbon and its relation to climate and vegetation Ecol Appl
2000 10 423ndash436 [CrossRef]88 Vogt KA Vogt DJ Palmiotto PA Boon P OrsquoHara J Asbjornsen H Review of root dynamics in forest ecosystems grouped
by climate climatic forest type and species Plant Soil 1995 187 159ndash219 [CrossRef]89 Leuschner C Ellenberg H Ecology of Central European Forests Vegetation Ecology of Central Europe Volume I Springer Cham
Switzerland 201790 Schenk HJ Jackson RB The global biogeography of roots Ecol Monogr 2002 72 311ndash328 [CrossRef]91 Leifeld J Zimmerman M Fuhrer J Conen F Storage and turnover of carbon in grassland soils along an elevation gradiend in
the Swiss Alps Glob Chang Biol 2009 15 668ndash679 [CrossRef]92 Makkonen K Helmisaari H Assessing fine-root biomass and production in a Scots pine standmdashComparison of soil core and
root ingrowth core methods Plant Soil 1999 210 43ndash50 [CrossRef]93 Yuan ZY Chen HYH Indirect Methods Produce Higher Estimates of Fine Root Production and Turnover Rates than Direct
Methods PLoS ONE 2012 7 e48989 [CrossRef]94 Day FP Schroeder RE Stover DB Brown ALP Butnor JR Dilustro J Hungate BA Dijkstra P Duval BD Seiler TJ
et al The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem New Phytol 2013 200 778ndash787 [CrossRef][PubMed]
95 Augusto L Dupouey JL Ranger J Effects of tree species on understory vegetation and environmental conditions in temperateforests Ann For Sci 2003 60 823ndash831 [CrossRef]
96 Gurmesa GA Schmidt IK Gundersen P Vesterdal L Soil carbon accumulation and nitrogen retention traits of four treespecies grown in common gardens For Ecol Manag 2013 309 47ndash57 [CrossRef]
97 Cremer M Kern NV Prietzel J Soil organic carbon and nitrogen stocks under pure and mixed stands of European beechDouglas fir and Norway spruce For Ecol Manag 2017 367 30ndash40 [CrossRef]
Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-

Forests 2021 12 823 27 of 27
98 Angst G Mueller K Eissenstat D Trumbore S Freeman K Hobbie S Chorover J Oleksyn J Reich P Mueller C Soilorganic carbon stability in forests Distinct effects of tree species identity and traits Glob Chang Biol 2019 25 1529ndash1546[CrossRef]
99 Koumlgel-Knabner I Guggenberger G Kleber M Kandeler M Kalbitz K Scheu S Eusterhues K Leinweber P Organo-mineralassociations in temperate soils Integrating biology mineralogy and organic matter chemistry J Plant Nutr Soil Sci 2008 17161ndash82 [CrossRef]
100 Luumltzow MV Koumlgel-Knabner I Ekschmitt K Matzner E Guggenberger G Marschner B Flessa H Stabilization of organicmatter in temperate soils Mechanisms and their rele- vance under different soil conditionsmdashA review Eur J Soil Sci 2006 57426ndash445 [CrossRef]
101 Lei P Scherer-Lorenzen M Bauhus J Belowground facilitation and competition in young tree species mixtures For EcolManag 2012 265 191ndash200 [CrossRef]
102 Yimer F Ledin S Abdelkadir A Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetationin the Bale Mountains Ethiopia Geoderma 2006 135 335ndash344 [CrossRef]
103 Matos ES Freese D Slazak A Bachmann U Veste M Huumlttl RF Organic-carbon and nitrogen stocks and organic-carbonfractions in soil under mixed pine and oak forest stands of different ages in NE Germany J Plant Nutr Soil Sci 2010 173 654ndash661[CrossRef]
104 Targulian VO Krasilnikov PV Soil system and pedogenic processes Self-organization time scales and environmentalsignificance Catena 2007 71 373ndash381 [CrossRef]
105 Gaul D Hertel D Leuschner C Effects of experimental soil frost on the fine-root system of mature Norway spruce J PlantNutr Soil Sci 2008 171 690ndash698 [CrossRef]
106 Leuschner C Hertel D Schmid I Koch O Muhs A Holscher D Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility Plant Soil 2004 258 43ndash56 [CrossRef]
107 Norby RJ Jackson RB Root dynamics and global change Seeking an ecosystem perspective New Phytol 2000 147 3ndash12[CrossRef]
108 Yuan ZY Chen HYH Fine Root Biomass Production Turnover Rates and Nutrient Contents in Boreal Forest Ecosystemsin Relation to Species Climate Fertility and Stand Age Literature Review and Meta-Analyses Crit Rev Plant Sci 2010 29204ndash221 [CrossRef]
109 Britton AJ Helliwell RC Lilly A Dawson L Fisher JM Coull M Ross J An integrated assessment of ecosystem carbonpools and fluxes across an oceanic alpine toposequence Plant Soil 2011 345 287ndash302 [CrossRef]
110 Djukic I Zehetner F Tatzber M Gerzabek MH Soil organic-matter stocks and characteristics along an alpine elevationgradient J Plant Nut Soil Sci 2010 173 30ndash38 [CrossRef]
111 Drewnik M The effect of environmental conditions on the decomposition rate of cellulose in mountain soils Geoderma 2006 132116ndash130 [CrossRef]
112 Hobbie SE Reich PB Oleksyn J Ogdahl M Zytkowiak R Hale C Karolewski P Tree species effects on decompositionand forest floor dynamics in a common garden Ecology 2006 87 2288ndash2297 [CrossRef]
113 Klimek B Niklinska M Jazwa M Tarasek A Tekielak I Musielok Ł Covariation of soil bacteria functional diversity andvegetation diversity along an altitudinal climatic gradient in the Western Carpathians Pedobiologia 2015 58 105ndash112 [CrossRef]
114 Niklinska M Klimek B Dynamics and stratification of soil biota activity along an altitudinal climatic gradient in WestCarpathians J Biol Res 2011 16 177ndash187
115 Taten R Takeda H Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soilnitrogen and light at the forest floor Ecol Res 2003 18 559ndash571 [CrossRef]
116 Espleta JF Clark DA Multi-scale variation in fine-root biomass in a tropical rain forest A seven-year study Ecol Monogr 200777 377ndash404 [CrossRef]
117 Leifeld J Bassin S Conen F Hajdas I Egli M Fuhrer J Control of soil pH on turnover of belowground organic matter insubalpine grassland Biogeochemistry 2013 112 59ndash69 [CrossRef]
118 Tan Z Lal R Smeck NE Calhoun FG Slater BK Parkinson B Gehring RM Taxonomic and geographic distribution ofsoil organic carbon pools in Ohio Soil Sci Soc Am J 2004 68 1896ndash1904 [CrossRef]
- Introduction
- Materials and Methods
-
- Study Area and Study Plot Selection
- Field Survey
- Sample Pretreatment Laboratory Analysis and Soil Organic Matter Stock Estimation
- Stand Characteristics and Coarse Root Biomass Estimation
- Data Analysis
-
- Results
-
- Predictor Variances Among Beech- Spruce- and Fir-Dominated Forests
- Determinants of Soil Organic Matter Stock and Fine Root Biomass in Beech- Spruce- and Fir-Dominated Forests
-
- Discussion
-
- Relationships Between Selected Forest Characteristics and Fine Root Biomass
- Relationships Between Selected Forest Characteristics and Soil Organic Matter Stock
- Effect of Abiotic Factors on Fine Root Biomass and Soil Organic Matter Stock
-
- Conclusions
- References
-
Related Documents











