Research Article Biotechnological conversions of bio-diesel- derived crude glycerol by Yarrowia lipolytica strains In the present report, crude glycerol, waste discharged from bio-diesel production, was used as carbon substrate for three natural Yarrowia lipolytica strains (LFMB 19, LFMB 20 and ACA-YC 5033) during growth in nitrogen-limited submerged shake-flask experiments. In media with initial glycerol concentration of 30 g/L, all strains presented satisfactory microbial growth and complete glycerol uptake. Although culture conditions favored the secretion of citric acid (and potentially the accumulation of storage lipid), for the strains LFMB 19 and LFMB 20, polyol mannitol was the principal metabolic product synthesized (maximum quantity 6.0 g/L, yield 0.20–0.26 g per g of glycerol consumed). The above strains produced small quantities of lipids and citric acid. In contrast, Y. lipolytica ACA-YC 5033 produced simultaneously higher quantities of lipid and citric acid and was further grown on crude glycerol in nitrogen-limited experiments, with constant nitrogen and increasing glycerol concentrations (70–120 g/L). Citric acid and lipid concentrations increased with increment of glycerol; maximum total citric acid 50.1 g/L was produced (yield 0.44 g per g of glycerol) while simultaneously 2.0 g/L of fat were accumulated inside the cells (0.31 g of lipid per g of dry weight). Cellular lipids were mainly composed of neutral fraction, the concentration of which substantially increased with time. Moreover, in any case, the phospholipid fraction was more unsaturated compared with total and neutral lipids, while at the early growth step, microbial lipid was more rich in saturated fatty acids (e.g. C16:0 and C18:0) compared with the stationary phase. Keywords: Citric acid / Crude glycerol / Mannitol / Single cell oil / Yarrowia lipolytica Received: July 31, 2009; revised: October 6, 2009; accepted: October 27, 2009 DOI: 10.1002/elsc.200900063 1 Introduction Bio-diesel fuels, defined as fatty acid methyl-esters deriving from trans-esterification of vegetable or animal fats, already constitute an alternative type of fuel for various types of diesel engines and heating systems [1–4]. Due to the potential exhaustion of the various conventional fuels currently used as well as the various environmental issues imposed (e.g. incre- ment of CO 2 emission, global planet warming phenomenon, etc.), the application of bio-diesel in a large commercial scale is strongly recommended by various authorities. This fact could very likely result in the generation of tremendous quantities of glycerol deposit in the market in the near future [1, 3–6]. Besides bio-diesel production plants, concentrated glycerol- containing waste-waters (crude glycerol) are also produced in significant quantities as the main by-product from fat sapo- nification and alcoholic beverage fabrication units [5, 7]. Therefore, conversion of (crude) glycerol to various high- added value products by the means of chemical and/or microbial technology attracts enormous interest [1, 3–5, 8, 9]. As far as microbial technology is concerned, in the last years a non-negligible number of scientific reports have appeared dealing with the fermentative valorization of (crude) glycerol. By far, the most classical fermentation of this resource that has been studied in significant details in biochemistry, microbial technology and down-stream processing levels refers to glycerol conversion into 1,3-propanediol by Clostridium butyricum and Klebsiella pneumoniae strains [1, 2, 6, 10–13]. Furthermore, glycerol has been recently used efficiently as Axel Andre ´ 1 Afroditi Chatzifragkou 1 Panagiota Diamantopoulou 2 Dimitris Sarris 1 Antonios Philippoussis 2 Maria Galiotou- Panayotou 1 Michael Komaitis 1 Seraphim Papanikolaou 1 1 Department of Food Science and Technology, Agricultural University of Athens, Athens, Greece 2 National Agricultural Research Foundation, Institute of Agricultural Machinery and Constructions, Laboratory of Edible Fungi, Athens, Greece Correspondence: Dr. Seraphim Papanikolaou ([email protected]), Assistant Professor in Food Bioprocesses, Department of Food Science and Technology, Laboratory of Food Microbiology and Biotechnology, Agricultural University of Athens, 75 Iera Odos, 11855 Athens, Greece. & 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com 468 Eng. Life Sci. 2009, 9, No. 6, 468–478

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Research Article
Biotechnological conversions of bio-diesel-derived crude glycerol by Yarrowia lipolyticastrains
In the present report, crude glycerol, waste discharged from bio-diesel production,was used as carbon substrate for three natural Yarrowia lipolytica strains (LFMB19, LFMB 20 and ACA-YC 5033) during growth in nitrogen-limited submergedshake-flask experiments. In media with initial glycerol concentration of 30 g/L, allstrains presented satisfactory microbial growth and complete glycerol uptake.Although culture conditions favored the secretion of citric acid (and potentiallythe accumulation of storage lipid), for the strains LFMB 19 and LFMB 20, polyolmannitol was the principal metabolic product synthesized (maximum quantity6.0 g/L, yield 0.20–0.26 g per g of glycerol consumed). The above strains producedsmall quantities of lipids and citric acid. In contrast, Y. lipolytica ACA-YC 5033produced simultaneously higher quantities of lipid and citric acid and was furthergrown on crude glycerol in nitrogen-limited experiments, with constant nitrogenand increasing glycerol concentrations (70–120 g/L). Citric acid and lipidconcentrations increased with increment of glycerol; maximum total citric acid50.1 g/L was produced (yield 0.44 g per g of glycerol) while simultaneously 2.0 g/Lof fat were accumulated inside the cells (0.31 g of lipid per g of dry weight).Cellular lipids were mainly composed of neutral fraction, the concentration ofwhich substantially increased with time. Moreover, in any case, the phospholipidfraction was more unsaturated compared with total and neutral lipids, while at theearly growth step, microbial lipid was more rich in saturated fatty acids (e.g. C16:0and C18:0) compared with the stationary phase.
Keywords: Citric acid / Crude glycerol / Mannitol / Single cell oil / Yarrowia lipolytica
Received: July 31, 2009; revised: October 6, 2009; accepted: October 27, 2009
DOI: 10.1002/elsc.200900063
1 Introduction
Bio-diesel fuels, defined as fatty acid methyl-esters derivingfrom trans-esterification of vegetable or animal fats, alreadyconstitute an alternative type of fuel for various types of dieselengines and heating systems [1–4]. Due to the potentialexhaustion of the various conventional fuels currently used aswell as the various environmental issues imposed (e.g. incre-ment of CO2 emission, global planet warming phenomenon,etc.), the application of bio-diesel in a large commercial scale isstrongly recommended by various authorities. This fact couldvery likely result in the generation of tremendous quantities of
glycerol deposit in the market in the near future [1, 3–6].Besides bio-diesel production plants, concentrated glycerol-containing waste-waters (crude glycerol) are also produced insignificant quantities as the main by-product from fat sapo-nification and alcoholic beverage fabrication units [5, 7].Therefore, conversion of (crude) glycerol to various high-added value products by the means of chemical and/ormicrobial technology attracts enormous interest [1, 3–5, 8, 9].As far as microbial technology is concerned, in the last years anon-negligible number of scientific reports have appeareddealing with the fermentative valorization of (crude) glycerol.By far, the most classical fermentation of this resource that hasbeen studied in significant details in biochemistry, microbialtechnology and down-stream processing levels refers toglycerol conversion into 1,3-propanediol by Clostridiumbutyricum and Klebsiella pneumoniae strains [1, 2, 6, 10–13].Furthermore, glycerol has been recently used efficiently as
Axel Andre1
Afroditi Chatzifragkou1
Panagiota
Diamantopoulou2
Dimitris Sarris1
Antonios Philippoussis2
Maria Galiotou-
Panayotou1
Michael Komaitis1
Seraphim Papanikolaou1
1Department of Food
Science and Technology,
Agricultural University of
Athens, Athens, Greece
2National Agricultural
Research Foundation,
Institute of Agricultural
Machinery and
Constructions, Laboratory
of Edible Fungi, Athens,
Greece
Correspondence: Dr. Seraphim Papanikolaou ([email protected]),
Assistant Professor in Food Bioprocesses, Department of Food Science
and Technology, Laboratory of Food Microbiology and Biotechnology,
Agricultural University of Athens, 75 Iera Odos, 11855 Athens, Greece.
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com
468 Eng. Life Sci. 2009, 9, No. 6, 468–478

starting material in fermentations other than that of 1,3-propanediol, and the production of microbial lipid (called alsosingle cell oil – SCO) [4, 14–17], food-grade pigments [18, 19],poly-hydroxy-alkanoates [20, 21], puryvic acid [22], microbialmass and enzymes [23] and erythritol [24] has been studied.Our research team has contributed to this effort with a numberof reports that dealt with the conversion of crude glycerol intovarious added-value compounds with the aid of yeast, bacterialor fungal species [7, 25–29].
The aim of the current investigation was to further assessthe potentialities of valorization of bio-diesel derived wasteglycerol, a residue used as a carbon source by newly isolatedYarrowia lipolytica strains in order for the production of citricacid, mannitol and lipids (SCO) to take place. It is known thatthe conversion of glycerol (or other similarly metabolizedcompounds like sugars) into citric acid or SCO occurs undernitrogen-limited conditions [3, 5]. While some reports haveappeared recently in relation with the transformation ofglycerol into SCO or citric acid by wild or mutant Y. lipolyticastrains [7, 24, 26, 29–34], in all cases not any simultaneousproduction of both metabolites occurred. Similarly, with theexemption of two recent reports [24, 34], there is lack ofinformation concerning the conversion of glycerol intomannitol by Y. lipolytica strains. Our investigation, thus, aimedat providing insights related with the conversion of crudeglycerol into citric acid, SCO and mannitol by newly isolatedY. lipolytica strains. Biochemical and kinetic considerationsrelated with these conversions were also assessed.
2 Materials and methods
2.1 Microorganisms and media
Y. lipolytica strain ACA-YC 5033 was isolated from varioustypes of sourdoughs [35]. Two other Y. lipolytica strains,isolated from edible products (fat-rich foods), were identifiedand characterized in the Laboratory of Food Microbiology andBiotechnology, Department of Food Science and Technology(Agricultural University of Athens, Greece) obtaining theculture numbers LFMB 19 and LFMB 20. All strains weremaintained on yeast peptone dextrose agar (YPDA) at 41C andsub-cultured every 4 months in order to maintain theirviability.
All experiments were performed in liquid-submerged flaskcultures. The culture medium used contained (in g/L):KH2PO4 7.0; Na2HPO4 2.5; MgSO4� 7H2O 1.5; CaCl2 0.15;FeCl3� 6H2O 0.15; ZnSO4� 7H2O 0.02; MnSO4�H2O 0.06.Ammonium sulfate [(NH4)2SO4] and yeast extract were usedas nitrogen sources in a concentration of 0.5 g/L each. Crudeglycerol provided from the ‘‘Hellenic Industry of Glycerin andFatty Acids SA’’ (purity770% w/w, impurities composed ofpotassium and sodium salts (12% w/w), non-glycerol organicmaterial (1% w/w), methanol (2% w/w) and water (14% w/w))was used as the sole carbon source in various concentrations.For all Y. lipolytica strains at a first trial, initial glycerol (Glol0)concentration into the medium was adjusted to 30 g/L. In anext approach, in order to investigate the impact of higherglycerol concentration and increased carbon excess upon the
biochemical behavior of Y. lipolytica ACA-YC 5033, this strainwas cultured in media containing Glol0 concentrations adjus-ted to 70, 90 and 120 g/L with all other culture parametersremaining constant. The initial pH for all media before andafter sterilization (1151C, 20 min) was 6.070.1. No acidaddition was required in any case, since although the pH of theglycerol feedstock was somehow elevated (around 10.0) thedilution of glycerol into the medium that presented a satis-factory buffer capacity resulted in a decrease of the pH of theculture medium to a value almost equal to 6.0.
2.2 Culture conditions
All experiments were performed in batch flask cultures in 250-mL flasks inoculated with 1 mL of exponential pre-culturecontaining 1–3� 107 [initial biomass concentration (X0)0.170.02 g/L] cells and incubated at T 5 281C with 180 rpmagitation in a rotary shaker (Lab-Line, IL, USA). The pre-culture was carried out in the above-mentioned syntheticmedium with initial glycerol concentration at 30 g/L. Pre-culture was incubated in the shaker under the same cultureconditions with the culture for 24 h.
In Y. lipolytica cultures, the initial pH of culture mediumwas 6.070.1, while during cultures, due to biosynthesis andaccumulation of organic acids (principally citric acid) into themedium, culture pH value presented a remarkable drop. Inorder to carry out cultures in a relatively controlled mode,the medium pH was maintained in a range between 4.5 and6.0 by adding (periodically and aseptically) small quantities(eg 500–600 mL) of 5 M KOH into the flasks (see Papanikolaouet al. [26]). The exact volume of KOH solution needed for pHcorrection was evaluated by measuring the volume of KOHsolution required for pH correction in one (at least) flask(collected daily). Then the appropriate volume of KOH solu-tion was aseptically added in the remaining flasks and the valueof pH reached was verified to be in a range of 4.5–6.0.
2.3 Analytical methods
Analyses carried out included determination of pH, drybiomass, glycerol, mannitol and organic acids quantity, resi-dual nitrogen concentration and intra-cellular lipids quantifi-cation and analysis. A Jenway 3020 pH-meter was used for pH-measurements. Cells were harvested by centrifugation(Heraeus Sepatech Suprafuge-22 apparatus) at 7000� g for20 min, were extensively washed with distilled water and werere-centrifuged in the same conditions. Cell concentration(X, g/L) was determined by dry weight (85751C/24 h).Glycerol (Glol, in g/L), mannitol (Ml, in g/L) and organic acidswere analyzed with the aid of a HPLC device. Filtered aliquotsof the culture medium were analyzed by a Waters Association600E apparatus at a 30.0 cm� 7.8 mm column Aminex HPX-87 (Bio-Rad, USA). The mobile phase used was H2SO4 at0.005 M with a flow rate 0.8 mL/min and the columntemperature was T 5 651C. Organic acids and polyols weredetected by an RI detector (differential refractometer 410-Waters). Under the above-mentioned analysis conditions, the
Eng. Life Sci. 2009, 9, No. 6, 468–478 Crude glycerol conversions by Yarrowia lipolytica 469
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

retention times of Cit, Ml, erythritol and Glol were 6.9, 8.7, 9.7and 11.0 min. respectively. Additionally, given that the HPLCanalysis carried out did not allow the satisfactory separation ofcitric acid from its isomer iso-citrate, in order to proceed witha more judicious determination of iso-citric acid in some ofthe fermentation points, an enzymatic method, based on themeasurement of the NADPH2 produced during conversion ofthe iso-citric to a-ketoglutaric acid, reaction catalyzed by iso-citrate dehydrogenase, was conducted [26]. As Cit (in g/L) willbe characterized the quantity of total citric acid (citric plus iso-citric acid) produced.
Dissolved oxygen (DO) concentration was determined withthe aid of a selective electrode (oxi200 Sensodirect, Lovinbod,UK). Before harvesting, the shaker was stopped and the probewas placed into the flask. Then, the shaker was switched onand the measurement was taken after DO equilibration(usually within 10 min). All trials were conducted in fullaerobic conditions (DO420% v/v for all growth steps of allfermentations). Ammonium ions determination was donewith the aid of an ammonium selective electrode (Hach 95-12,Germany).
Total cellular lipid (L, in g/L) was extracted from the drybiomass with a mixture of chloroform/methanol 2/1 (v/v) andwas determined gravimetrically. Furthermore, in some trialscellular lipids were fractionated into their lipid fractions. Inbrief, a known weight of extracted lipid (approximately300 mg) was dissolved in chloroform (3 mL) and was fractio-nated by using a column (25� 100 mm) of silicic acid, acti-vated by heating overnight at 1101C [36]. Successiveapplications of 1,1,1-trichloroethane, acetone and methanolproduced fractions containing neutral lipids (N), glycolipidsplus sphingolipids (G1S) and phospholipids (P), respectively[36]. The weight of each fraction was determined afterevaporation of the respective solvent. Cellular lipids or indi-vidual lipid fractions were converted to their methyl-esters in atwo-step reaction described in Papanikolaou et al. [26]. Fattyacid methyl-esters were analyzed according to Fakas et al. [36].
3 Results
3.1 Growth of Y. lipolytica strains in media withinitial glycerol at 30 g/L
In the first part of the present investigation, experiments wereperformed with all three natural Y. lipolytica strains used inmedia in which bio-diesel derived crude glycerol was utilizedas the sole carbon source in Glol0 5 30 g/L. It is known that insubmerged high carbon-excess cultures with glycerol (orsimilarly metabolized compounds) utilized as the solesubstrate, Y. lipolytica strains can potentially produce quan-tities of extra-cellular citric acid or intra-cellular reserve lipid[7, 26, 32]; therefore, kinetic studies were carried out in mediathat favor the production of the above-mentioned metabolites.The kinetic results of the experiments are presented in Table 1.In all experiments, microorganisms consumed with compar-able rates, the available extra-cellular nitrogen (initial NH4
1 at110715 ppm, exhaustion within 5075 h after inoculation).However, the strains LFMB 19 and LFMB 20 presented T
able
1.
Dat
ao
fY
arro
wia
lipol
ytic
ast
rain
so
rig
inat
edfr
om
kin
etic
so
nn
itro
gen
-lim
ited
med
iaw
ith
bio
-die
sel
der
ived
gly
cero
lu
tiliz
edas
the
sole
carb
on
sou
rce
atin
itia
lsu
bst
rate
con
cen
trat
ion
(Glo
l 0)
30
g/L
.a)
Stra
inT
ime
(h)
X(g
/L)
Glo
l co
ns
(g/L
)C
it(g
/L)
Ml
(g/L
)L
(g/L
)Y
L/X
(g/g
)Y
Cit
/Glo
l(g
/g)
YM
l/G
lol
(g/g
)
LF
MB
1969
i)6.
118
.64.
64.
80.
420.
070.
250.
26
117ii
)8.
125
.23.
95.
10.
810.
100.
150.
20
LF
MB
2090
i),
ii)
8.2
25.3
3.2
6.0
0.61
0.07
0.13
0.24
AC
A-Y
C50
3327
iii)
4.2
8.0
tr.b
)tr
.b)
0.82
0.20
––
187i)
5.4
29.8
11.7
tr.b
)0.
120.
020.
39–
a)R
epre
sen
tati
on
of
bio
mas
s(X
,g/L
),li
pid
(L,g
/L),
citr
icac
id(C
it,g
/L),
man
nit
ol
(Ml,
g/L
)an
dco
nsu
med
glyc
ero
l(G
lol c
on
s,g/
L)
con
cen
trat
ion
sat
dif
fere
nt
ferm
enta
tio
np
oin
tso
fea
chtr
ial:
(i)
wh
en
Cit
max
con
cen
trat
ion
was
ach
ieve
d;
(ii)
wh
enM
l max
con
cen
trat
ion
was
ach
ieve
d;
(iii
)w
hen
max
imu
mq
uan
tity
of
cell
ula
rli
pid
sin
dry
mat
ter
was
ach
ieve
d.
Fer
men
tati
on
tim
e,co
nve
rsio
nyi
eld
of
citr
icac
idp
rod
uce
dpe
rgl
ycer
ol
con
sum
ed(Y
Cit
/Glo
l,g/
g),c
on
vers
ion
yiel
do
fm
ann
ito
lp
rod
uce
dpe
rgl
ycer
ol
con
sum
ed(Y
Ml/
Glo
l,g/
g),l
ipid
ind
ryb
iom
ass
(YL
/X,g
/g)
and
con
sum
edgl
ycer
ol.
Val
ues
are
pre
sen
ted
for
all
po
ints
of
the
tria
ls.
Cu
ltu
reco
nd
itio
ns:
gro
wth
on
250-
mL
flas
ksat
185
rpm
,G
lol 0
530
g/L
,am
mo
niu
msu
lfat
ean
dye
ast
extr
act
at0.
5g/
Lo
fea
ch,
init
ial
pH
56.
070.
1,p
H
ran
gin
gb
etw
een
4.5
and
6.0,
DO4
40%
v/v,
incu
bat
ion
tem
per
atu
reT
5281C
.E
ach
po
int
isth
em
ean
valu
eo
ftw
oin
dep
end
ent
mea
sure
men
ts.
i)R
epre
sen
tati
on
sw
hen
Cit
max
con
cen
trat
ion
(in
g/L
)w
asac
hie
ved
.
ii)
Rep
rese
nta
tio
ns
wh
enM
l max
con
cen
trat
ion
(in
g/L
)w
asac
hie
ved
.
i),
ii)
Cit
max
and
Ml m
axw
ere
ob
tain
edsi
mu
ltan
eou
sly.
iii)
Rep
rese
nta
tio
ns
wh
enm
axim
um
YL
/X(i
ng/
g)w
asac
hie
ved
.
b)
tr.o
0.5
g/L
.
470 A. Andre et al. Eng. Life Sci. 2009, 9, No. 6, 468–478
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com
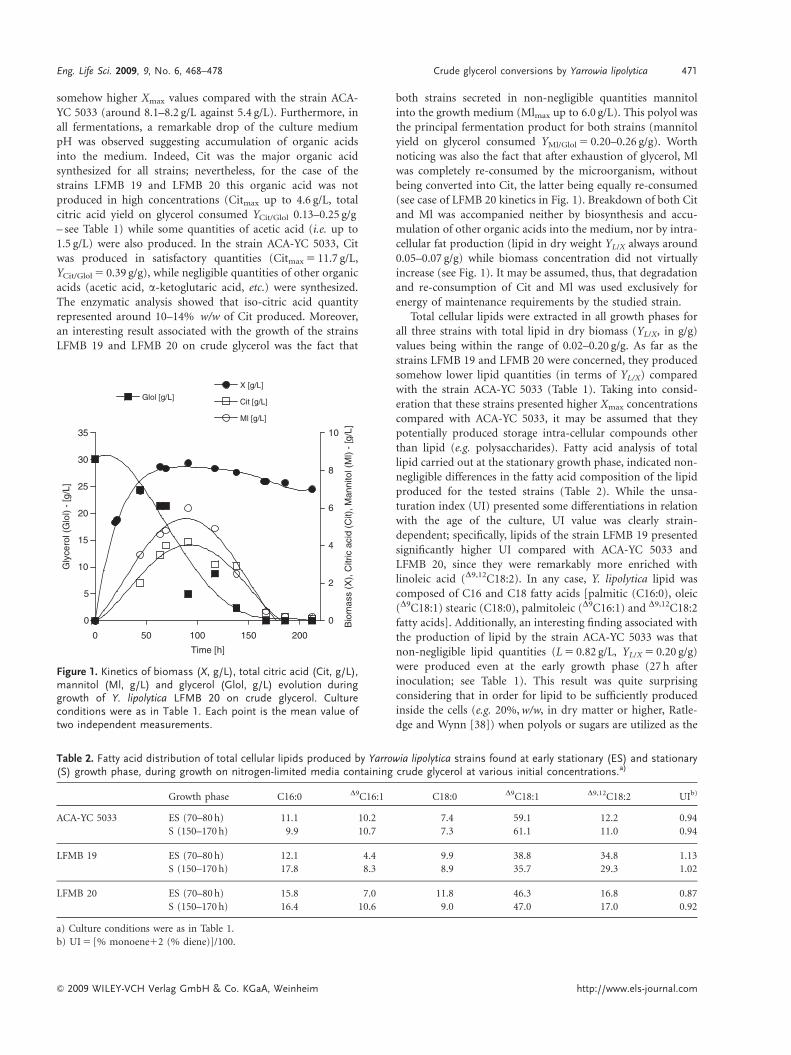
somehow higher Xmax values compared with the strain ACA-YC 5033 (around 8.1–8.2 g/L against 5.4 g/L). Furthermore, inall fermentations, a remarkable drop of the culture mediumpH was observed suggesting accumulation of organic acidsinto the medium. Indeed, Cit was the major organic acidsynthesized for all strains; nevertheless, for the case of thestrains LFMB 19 and LFMB 20 this organic acid was notproduced in high concentrations (Citmax up to 4.6 g/L, totalcitric acid yield on glycerol consumed YCit/Glol 0.13–0.25 g/g– see Table 1) while some quantities of acetic acid (i.e. up to1.5 g/L) were also produced. In the strain ACA-YC 5033, Citwas produced in satisfactory quantities (Citmax 5 11.7 g/L,YCit/Glol 5 0.39 g/g), while negligible quantities of other organicacids (acetic acid, a-ketoglutaric acid, etc.) were synthesized.The enzymatic analysis showed that iso-citric acid quantityrepresented around 10–14% w/w of Cit produced. Moreover,an interesting result associated with the growth of the strainsLFMB 19 and LFMB 20 on crude glycerol was the fact that
both strains secreted in non-negligible quantities mannitolinto the growth medium (Mlmax up to 6.0 g/L). This polyol wasthe principal fermentation product for both strains (mannitolyield on glycerol consumed YMl/Glol 5 0.20–0.26 g/g). Worthnoticing was also the fact that after exhaustion of glycerol, Mlwas completely re-consumed by the microorganism, withoutbeing converted into Cit, the latter being equally re-consumed(see case of LFMB 20 kinetics in Fig. 1). Breakdown of both Citand Ml was accompanied neither by biosynthesis and accu-mulation of other organic acids into the medium, nor by intra-cellular fat production (lipid in dry weight YL/X always around0.05–0.07 g/g) while biomass concentration did not virtuallyincrease (see Fig. 1). It may be assumed, thus, that degradationand re-consumption of Cit and Ml was used exclusively forenergy of maintenance requirements by the studied strain.
Total cellular lipids were extracted in all growth phases forall three strains with total lipid in dry biomass (YL/X, in g/g)values being within the range of 0.02–0.20 g/g. As far as thestrains LFMB 19 and LFMB 20 were concerned, they producedsomehow lower lipid quantities (in terms of YL/X) comparedwith the strain ACA-YC 5033 (Table 1). Taking into consid-eration that these strains presented higher Xmax concentrationscompared with ACA-YC 5033, it may be assumed that theypotentially produced storage intra-cellular compounds otherthan lipid (e.g. polysaccharides). Fatty acid analysis of totallipid carried out at the stationary growth phase, indicated non-negligible differences in the fatty acid composition of the lipidproduced for the tested strains (Table 2). While the unsa-turation index (UI) presented some differentiations in relationwith the age of the culture, UI value was clearly strain-dependent; specifically, lipids of the strain LFMB 19 presentedsignificantly higher UI compared with ACA-YC 5033 andLFMB 20, since they were remarkably more enriched withlinoleic acid (D9,12C18:2). In any case, Y. lipolytica lipid wascomposed of C16 and C18 fatty acids [palmitic (C16:0), oleic(D9C18:1) stearic (C18:0), palmitoleic (D9C16:1) and D9,12C18:2fatty acids]. Additionally, an interesting finding associated withthe production of lipid by the strain ACA-YC 5033 was thatnon-negligible lipid quantities (L 5 0.82 g/L, YL/X 5 0.20 g/g)were produced even at the early growth phase (27 h afterinoculation; see Table 1). This result was quite surprisingconsidering that in order for lipid to be sufficiently producedinside the cells (e.g. 20%, w/w, in dry matter or higher, Ratle-dge and Wynn [38]) when polyols or sugars are utilized as the
0
5
10
15
20
25
30
35
0
2
4
6
8
10
0 50 100 150 200
Glol [g/L]
X [g/L]
Cit [g/L]
Ml [g/L]
Gly
cero
l (G
lol)
- [g
/L]
Bio
mas
s (X
), C
itric
aci
d (C
it), M
anni
tol (
Ml)
- [g
/L]
Time [h]
Figure 1. Kinetics of biomass (X, g/L), total citric acid (Cit, g/L),mannitol (Ml, g/L) and glycerol (Glol, g/L) evolution duringgrowth of Y. lipolytica LFMB 20 on crude glycerol. Cultureconditions were as in Table 1. Each point is the mean value oftwo independent measurements.
Table 2. Fatty acid distribution of total cellular lipids produced by Yarrowia lipolytica strains found at early stationary (ES) and stationary(S) growth phase, during growth on nitrogen-limited media containing crude glycerol at various initial concentrations.a)
Growth phase C16:0 D9C16:1 C18:0 D9C18:1 D9,12C18:2 UIb)
ACA-YC 5033 ES (70–80 h) 11.1 10.2 7.4 59.1 12.2 0.94
S (150–170 h) 9.9 10.7 7.3 61.1 11.0 0.94
LFMB 19 ES (70–80 h) 12.1 4.4 9.9 38.8 34.8 1.13
S (150–170 h) 17.8 8.3 8.9 35.7 29.3 1.02
LFMB 20 ES (70–80 h) 15.8 7.0 11.8 46.3 16.8 0.87
S (150–170 h) 16.4 10.6 9.0 47.0 17.0 0.92
a) Culture conditions were as in Table 1.
b) UI 5 [% monoene12 (% diene)]/100.
Eng. Life Sci. 2009, 9, No. 6, 468–478 Crude glycerol conversions by Yarrowia lipolytica 471
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

sole carbon source, nitrogen-limited conditions are required;in the present culture conditions, 27 h after inoculation non-negligible NH4
1 quantities were always found into the medium.Extensive analysis of the lipid produced by strain ACA-YC5033 was performed at this fermentation step (t 5 27 h,exponential growth phase) and it was revealed that N consti-tuted the major fraction of cellular lipids produced (73.3% w/wof total lipid) (Table 3). Moreover, total fatty acid analysis ofcellular lipid produced showed that at the exponential growthphase, lipids were more saturated compared with the otherphases of the microbial growth (Tables 2 and 3). Theconcentrations of C16:0 and C18:0 decreased during growth,while the opposite trend was observed for D9C18:1 (Tables 2and 3). Analysis of G1S and P fractions showed that fatty acidcomposition of these fractions was somehow different withthat of total lipids produced, whereas in general the P fractionwas significantly more rich in poly-unsaturated fatty acids(principally D9,12C18:2) compared with total and neutral lipids(UI for P fraction was 1.07 while for the total lipids was 0.81).Moreover, kinetics of YL/X as function of the time for the strainACA-YC 5033 indicated that YL/X value was maintained withinthe range 0.10–0.16 g/g until Glol was found into the mediumin significant quantities (up to 70 h). Thereafter, cellular lipidswere re-consumed and final YL/X values were very low(0.02–0.04 g/g) (kinetics not presented).
3.2 Cultures of strain ACA-YC 5033 at elevatedconcentrations of crude glycerol
Taking into consideration that the strain ACA-YC 5033presented satisfactory microbial growth, while citric acid wasproduced in non-negligible quantities and simultaneous
potential accumulation of intra-cellular lipid occurred, thisstrain was selected for further trials with raw glycerol utilizedas the sole substrate. The above strain was cultured in mediapresenting a very high excess of carbon, in order to furtherfavor the production of citric acid and/or SCO. Therefore,cultures were done at increasing Glol0 quantities (70, 90 and120 g/L), with nitrogen concentration into the mediumremaining constant (see Table 4). Although the concentrationof glycerol did not significantly affect growth of Y. lipolytica,since in all cases almost equivalent final biomass concentra-tions were obtained, maximum specific growth rate (mmax) wassubstantially higher when the Glol0 concentration was lower,potentially due to some inhibition exerted at high Glol0concentration media. Moreover, at the relatively lower Glol0concentrations, the plateau of biomass production seemed toarrive earlier (kinetics not presented), while X concentrationslightly increased at high glycerol concentration media(Table 4). Similarly, nitrogen exhaustion that occurred around50 h after inoculation regardless of the initial glycerolconcentration, induced a remarkable Cit production, giventhat the above metabolite was the principal organic acidproduced. A Citmax value of 50.1 g/L was obtained, while theconversion yield YCit/Glol did not seem to remarkably increasewith Glol concentration increment into the culture medium(Table 4). However, Cit concentration (in absolute values, g/L)remarkably increased with Glol0 concentration increment.The representation of Cit produced per Glol consumed forall trials performed with the strain ACA-YC 5033 indicatedthat in the present investigation, the conversion threshold ofcitric acid produced per glycerol consumed could not be higherthan 0.44 g/g (Fig. 2). Moreover, significant quantities ofglycerol were consumed at the end of all trials, requiring,however, longer incubation time. The enzymatic analysis
Table 3. Fatty acid composition of total lipids, neutral lipids (N), glycolipids plus sphingolipids (G1S) and phospholipids (P) duringgrowth of Yarrowia lipolytica ACA-YC 5033 on raw glycerol-based media.a)
w/w% C16:0 D9C16:1 C18:0 D9C18:1 D9,12C18:2 UI
ACA-YC 5033 Total lipids 17.4 7.8 16.6 42.9 15.3 0.81
t 5 27 h N 73.3 19.4 7.1 16.4 40.3 16.8 0.81
G1S 20.1 22.4 2.7 7.4 53.3 14.2 0.84
P 6.6 13.8 8.1 4.1 49.4 24.6 1.07
a) Culture conditions as in Table 1, UI as in Table 2, sampling point in which lipids were analyzed was at the exponential growth phase (t 5 24 h after
inoculation).
Table 4. Data of Yarrowia lipolytica ACA-YC 5033 originated from kinetics on glycerol-based nitrogen-limited media at various initialglycerol concentrations (Glol0, g/L) and constant initial nitrogen concentration.a)
Glol0 (g/L) Time (h) Glolcons (g/L) X (g/L) Lmax (g/L) Citmax (g/L) mmax (h�1) YL/X (g/g) YCit/Glol (g/g)
70 187 66.7 5.6 0.9 28.0 0.14 0.16 0.42
90 220 73.0 5.9 1.4 31.2 0.12 0.24 0.43
120 375 114.0 6.5 2.0 50.1 0.06 0.31 0.44
a) Representation X (g/L) and Glolcons (g/L) when Citmax (g/L) and Lmax (g/L) values were achieved. YCit/Glol and YL/X (g/g) values are presented for the
above fermentation point. Maximum specific growth rate (mmax, h�1) was estimated by fitting the equation ln (X/X0) 5 f(t)within the early
exponential growth phase for each trial. Culture conditions and abbreviations were as in Table 1. Each point is the mean value of two independent
measurements.
472 A. Andre et al. Eng. Life Sci. 2009, 9, No. 6, 468–478
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

showed that iso-citric acid represented around 10% w/w, oftotal Cit produced, regardless of Glol0 used into the culturemedium.
Total intra-cellular lipids (L) were quantified at all growthsteps and all cultures. As shown in Table 4, the quantity oflipids produced (in absolute values, g/L, and relative valuesYL/X, g/g) substantially increased with Glol0 concentrationincrement. The higher quantity of lipid accumulated inside the
cells (YL/X) was 0.31 g/g in dry weight with respective absoluteLmax value of 2.0 g/L. The evolution of YL/X as function of thefermentation time for all trials shows that indeed SCO accu-mulation increased with Glol0 concentration increment(Fig. 3). The simultaneous non-negligible production of citricacid and SCO is a worth-noticing result, since in most of thecases strains belonging to Y. lipolytica species were revealedcapable to produce exclusively either citric acid or SCO [7, 26].Total fatty acid composition of cellular lipids was analyzed atthe stationary growth phase for all cultures. As previously, themain fatty acids were D9C18:1, C16:0, D9,12C18:2, D9C16:1 andC18:0. Not significant changes in the distribution of cellularfatty acids were observed regardless of the increment of Glol0concentration into the medium (Table 5). However, it seemedthat the UI of cellular lipids produced, slightly decreased withGlol0 concentration increment into the medium (see Tables 2and 5). Finally, analysis of the various lipid fractions (N, G1Sand P fractions) of SCO produced at the late exponential (LE)phase (50 h after inoculation) and late stationary (LS) phase(375 h after inoculation) demonstrated that progressivelyduring growth, N concentration significantly increased (from53.6% w/w to 87.1% w/w of total lipid produced), while, incontrast, polar fractions (G1S and P) were substantiallydecreased (Table 6). Moreover, as it was previously observed(Glol0 5 30 g/L), total lipids seemed to be somehow moresaturated and more rich in C16:0 and C18:0 fatty acids at theearly growth step (LE phase), compared with the onesproduced at the stationary phase (see Tables 5 and 6). Simi-larly, fatty acid composition of all lipid fractions was somehowaffected by the fermentation time, whereas, as previously, thefraction of P was more rich in poly-unsaturated fatty acids(principally D9,12C18:2) compared with total and neutral lipids(Table 6). As for the G1S fraction, it seemed to be slightlymore saturated compared with the total lipids and the Nfraction (Table 6).
4 Discussion
The potentiality of growth of three wild Y. lipolytica strains oncrude glycerol was assessed. In nitrogen-limited media withGlol0 5 30 g/L, although the limitation of nitrogen was high(C/N 5 100 moles/moles) glycerol was relatively rapidlyconsumed (substrate consumption within 4–6 days, dependenton the strain). Even when culture conditions favored theaccumulation of citric acid (e.g. pH value in medium 4.5–6.0,relatively increased DO saturation), this organic acid was notthe principal extra-cellular metabolite produced by the strainsLFMB 19 and LFMB 20; the principal one was the polyolmannitol. No other polyols (i.e. erythritol) were produced. Infew reports, secretion of polyols into the culture mediumtogether with citric acid, when glycerol was utilized as the solecarbon source in nitrogen-limited submerged experiments, hasbeen studied [24, 33, 34]. When mutant Y. lipolytica strains(e.g. Wratislavia AWG7 and K1 strains) were used in bioreactorexperiments, erythritol was the principal polyol produced, insome cases in significant concentrations (e.g. up to 47 g/L),while Ml was secreted in small amounts (e.g. up to 9 g/L)[24]. Nevertheless, Cit was the principal metabolite produced
0
10
20
30
40
50
60
0 20 40 60 80 100 120 140
Cit, Glol0=30 g/L
Cit, Glol0=70 g/L
Cit, Glol0=90 g/L
Cit, Glol0=120 g/L
Citr
ic a
cid
(Cit)
- [g
/L]
Glycerol (Glol) consumed - [g/L]
YCit/Glol
=0.44 g/g
Figure 2. Total citric acid produced (Cit, g/L) as function ofglycerol (Glol, g/L) consumed during growth of Y. lipolytica ACA-YC 5033 on glycerol-based nitrogen-limited media at variousinitial glycerol concentrations (Glol0, g/L) and constant initialnitrogen concentration in flask cultures. Culture conditions wereas in Table 1. Each point is the mean value of two independentmeasurements.
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0 50 100 150 200 250 300 350 400
Glol0 =70 g/L
Glol0 =90 g/L
Glol0 =120 g/L
Lipi
d in
dry
wei
ght,
YL/
X [g
/g]
Time (h)
Figure 3. Kinetics of evolution of lipid in dry weight (YL/X, g/g)during growth of Y. lipolytica ACA-YC 5033 on glycerol-basednitrogen-limited media at various initial glycerol concentrations(Glol0, g/L) and constant initial nitrogen concentration in flaskcultures. Culture conditions were as in Table 1. Each point is themean value of two independent measurements.
Eng. Life Sci. 2009, 9, No. 6, 468–478 Crude glycerol conversions by Yarrowia lipolytica 473
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

(in very high quantities, e.g. 87–139 g/L) [24]. Moreover, Ml inconcentrations 4–9 g/L together with erythritol (in concen-trations 4–15 g/L) have been produced by another mutantstrain (Wratislavia 1.31) in fed-batch bioreactor experimentswith glycerol utilized as the sole carbon source [34]. Aspreviously, very high Cit amounts have been produced(109–148 g/L) with polyols representing only a small portionof total metabolites produced [34]. Production of polyols fromglycerol as starting material is not common for Y. lipolyticastrains, and can present interest for Food Technology [34].Therefore, the fact that the natural LFMB 19 and LFMB 20strains can synthesize non-negligible Ml quantities as theprincipal extra-cellular metabolite is of interest and will bestudied in more details by our team. The biochemicalmechanism of Ml (and erythritol) biosynthesis by strainsgrowing on glycerol has not been completely elucidated. It hasbeen postulated that sugar alcohols, including Ml anderythritol, can protect various cells from osmotic stress [39],which likely occurs due to exposure of strains into relativelyhigh Glol concentrations into the medium. Moreover, in thecurrent investigation, after exhaustion of Glol from themedium, Ml was re-consumed without being transformed intocitric acid (in disagreement with Rywinska and Rymowicz[34]). Moreover, strains LFMB 19 and LFMB 20 completely re-consumed the previously produced Cit after Glol disap-pearance from the medium. Citric acid re-consumption hasbeen already reported for various Cit producing Y. lipolytica orCandida guilliermondii strains [26, 40].
In the cultures with Glol0 5 30 g/L, some quantities oflipids were produced for all strains. Strain ACA-YC 5033
produced higher fat quantities (lipid in dry weight at20% w/w) at the exponential growth phase, where ammoniumhad not yet been depleted from the medium. Y. lipolyticalipids were composed of C16 and C18 fatty acids, whereasstrain LFMB 19 presented higher concentration of the fattyacid D9,12C18:2 compared with the other strains tested.Therefore, cellular fatty acid composition can present strain-dependent differences, while cultures on glucose or glycerolcan result in differentiations in the fatty acid composition ofcellular lipids due to differences in initial substrate concen-tration, in the dilution rate imposed (for the continuouscultures) and addition of natural compounds into the medium[3, 7, 29, 41–43].
Strain ACA-YC 5033 was chosen for further trials withincreased Glol0 concentrations (70, 90 and 120 g/L) and allother parameters remaining constant. Although mmax valuedecreased with Glol0 increment in the medium (indicatingslight inhibitory effect due to the increase of substrateamount), biomass production was satisfactory within therange of Glol0 concentrations, presenting the tendency toincrease along with substrate increment. Equivocal results havebeen reported in relation with the potential inhibition exertedby the high initial glycerol or glucose concentration in themedia (e.g. quantities 70–200 g/L) upon growth of natural ormutant Y. lipolytica strains; in accordance with the resultsobtained by ACA-YC 5033, growth of other Y. lipolytica strainsseemed unaffected by the high substrate concentration, whileuninterrupted Cit production was observed [26, 29, 32, 42]. Incontrast, in different Y. lipolytica strains growing at increasinginitial glucose concentration media (e.g. from 50 to 200 g/L),
Table 6. Fatty acid composition of total lipids, neutral lipids (N), glycolipids plus sphingolipids (G1S) and phospholipids (P) duringgrowth of Yarrowia lipolytica ACA-YC 5033 on raw glycerol-based media.a)
w/w% C16:0 D9C16:1 C18:0 D9C18:1 D9,12C18:2 UI
ACA-YC 5033 Total lipids 16.9 9.2 16.4 44.3 13.2 0.80
t 5 50 h N 53.6 17.9 9.9 19.7 42.8 9.7 0.72
G1S 30.7 20.9 3.6 18.6 42.9 14.3 0.75
P 15.7 9.7 10.5 2.9 59.4 17.5 1.05
ACA-YC 5033 Total 11.6 10.1 8.7 60.4 9.2 0.88
t 5 375 h NL 87.1 13.2 8.6 9.3 57.8 11.1 0.89
G1S 4.2 19.3 7.1 14.3 48.4 10.9 0.77
P 8.7 9.6 10.2 7.4 59.1 13.7 0.97
a) Culture with Glol0 5 120 g/L, sampling point in which lipids were analyzed was at the late exponential (LE) growth phase (t 5 50 h after inoculation)
and the late stationary (LS) phase (t 5 375 h). Culture conditions were as in Table 1, UI as in Table 2.
Table 5. Fatty acid distribution of total cellular lipids produced by Yarrowia lipolytica ACA-YC 5033 found at early stationary (ES) andstationary (S) growth phase, during growth on nitrogen-limited media containing crude glycerol at various initial concentrations.a)
Glol0 (g/L) Growth phase C16:0 D9C16:1 C18:0 D9C18:1 D9,12C18:2 UI
70 ES (70–80 h) 13.2 11.2 7.3 57.1 11.2 0.91
S (150–170 h) 12.0 12.8 6.5 58.1 10.6 0.92
90 ES (70–80 h) 12.2 11.1 9.5 54.1 13.1 0.91
S (150–170 h) 13.0 10.2 7.9 54.9 14.0 0.93
120 ES (70–80 h) 14.2 10.0 9.6 55.1 11.1 0.87
S (150–170 h) 12.1 10.9 9.0 57.1 10.9 0.90
a) Culture conditions were as in Table 1, UI as in Table 2.
474 A. Andre et al. Eng. Life Sci. 2009, 9, No. 6, 468–478
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

a drastic depletion on values of mmax and Cit occurred dueto strong substrate inhibition [44]. Moreover, growth ofY. lipolytica ACA-YC 5033 was accompanied by satisfactoryCit production typically enhanced after nitrogen depletionfrom the medium. Elevation of citric acid production interms of absolute values (Citmax, g/L) was observed withGlol0 concentration increment (see Table 4). This canbe considered as an expected event since biosynthesis ofcitric acid from various carbon sources is a non-growthassociated process occurring after nitrogen depletion fromthe culture medium, and the more the carbon substrate isfound in excess, the more the microbial metabolism isdirected towards the biosynthesis of citric acid, due to‘‘metabolic over-flow’’ under carbon excess conditions [29, 37,43, 45, 46]. Additionally, kinetic analysis demonstratedsome ‘‘saturation’’ of glycerol assimilation at high Glol0concentrations, since under these conditions long incuba-tion period was required for the accomplishment of thefermentation (16 days). Similar observations have beendone for other natural Y. lipolytica strains growing onhigh glucose- or glycerol-amount media [29, 43]. In contrast,in cultures led by mutant Y. lipolytica strains (i.e. Wratislavia1.31 and K1) on highly agitated bioreactors, Glol assimilationrate was remarkably high, in spite of the fact the mediawere limited in nitrogen, resulting in excessive and rapidCit production (e.g. quantities 90–150 g/L within 5–7 days)[24, 32–34].
Strain ACA-YC 5033 showed similar performances for Citproduction in terms of YCit/Glol, regardless of the Glol0concentration; the threshold of Cit produced per Glolconsumed was 0.44 g/g, which is comparable with the valuesachieved in the international literature for glucose or polyols.The highest yield value reported in the literature was byWojtatowicz et al. [47], who revealed a yield of Cit equal to0.93 g/g in experiments with a strain of Y. lipolytica grown onhydrolyzed potato starch. Rane and Sims [48] have equallyreported high Cit yields for glucose-grown Y. lipolytica strains(0.77 g/g). In contrast, in most of the reports in which yeastshave been used in the conversion of sugars or polyols into Cit,similar values with the present study have been reported (e.g.0.30–0.62 g/g) [5, 26, 27, 31, 32, 37, 40, 43, 44]. YCit/S values aresignificantly dependent on the converted carbon source, sincesubstances with higher carbon and reductance degree can leadto higher conversion yields; for instance, growth of Y. lipolyticaon ethanol was accompanied by a yield of 0.87 g/g [46], whilegrowth on hydrophobic substrates (e.g. crude fats) can beaccompanied by conversion yields, which may be equal or even(notably) higher than 1.0 g/g [45, 49]. Moreover, Citmax
concentration achieved was 50.1 g/L, which can be consideredas satisfactory and comparable with the one reported by otherY. lipolytica or Candida oleophila strains; Klasson et al. [50]have reported a production of 51.5 g/L by C. lipolytica NRRL-Y-7576 during batch bioreactor cultures on glucose, while therespective values for C. lipolytica Y1095 grown on glucosewere 13.6–78.5 g/L [48]. Y. lipolytica ATCC 20346 produced50–69 g/L during growth on glucose-based fed-batch bior-eactor experiments [51]. C. oleophila ATCC 20177 produced50–79 g/L on glucose-based flask or bioreactor batch experi-ments [52] and 37.6–57.8 g/L on single-stage continuous
operations [37]. Strain ACA-DC 50109 produced 43 g/L duringgrowth on glucose [42] and 63 g/L on bio-diesel-derivedglycerol [29]. Imandi et al. [30] have reported concentrationsaround 78 g/L during growth of Y. lipolytica on bio-diesel-derived by-product containing glycerol, while Levinson et al.[31] have reported Citmax values of 24–31 g/L during growth ofvarious strains on pure glycerol. Finally, extremely highconcentrations (up to 150 g/L) have been achieved by mutantY. lipolytica strains cultivated on ethanol [46], vegetable oils[45] or raw glycerol [24, 32–34] in bioreactor operations. Inthe above-cited works, batch and fed-batch operations wereaccomplished rapidly resulting in high Cit concentrations (i.e.Cit 5 77–146 g/L) within 5–7 days [24, 32–34, 45, 49].
Interestingly, during growth of ACA-YC 5033 strain onglycerol-based nitrogen-limited cultures, the increment ofGlol0 concentration (and the ratio C/N) into the mediumnotably increased SCO accumulation. At high Glol0concentrations (120 g/L), 2.0 g/L of lipid with 0.31 g of lipidper g of dry weight were produced. SCO was mainly com-posed of neutral fractions, the concentration of whichsignificantly increased with the fermentation time, in fullagreement with the findings reported by other oleaginousfungi growing on glucose-based media [36]. Moreover,P fraction of SCO produced was richer in poly-unsaturatedfatty acids compared with total lipids and the N fraction, infull agreement with the results obtained by other oleaginousfungi like Cunninghamella echinulata [36]. Similarly, thedistribution of cellular fatty acids by Y. lipolytica ACA-YC 5033seemed to change with the age of the culture since at theexponential growth phase total lipids were more saturatedcompared with these on stationary phase (in agreement withthe results reported by the strain ACA-DC 50109 growing onglucose or glycerol [26]).
In general, there is confusion about the potentiality ofY. lipolytica to accumulate fat in flask experiments usingglucose or glycerol as substrate. While in continuous nitrogen-limited experiments with glycerol used as substrate and lowdilution rates imposed, noticeable quantities of lipid have beenproduced (Lmax 5 2.0–3.5 g/L, YL/X 0.25–0.43 g/g) along withnegligible quantities of Cit [7], in flask experiments smallquantities of lipids from this type of substrates were produced,since nitrogen limitation directed the metabolism towards Citsynthesis [26, 29]. This fact was attributed to the potentiallynot sufficient aeration occurring in the shake-flask experi-ments, since the not satisfactory O2 transfer could lead tochanges in gene expression resulting in down-regulation ofproteins with crucial importance for the process of lipidaccumulation (e.g. ATP-citrate lyase, NADPH1-malic enzyme)[38], with evident result of a decreased SCO production. Inany case, the current manuscript is the first report in whichnon-negligible quantities of SCO and citric acid are simulta-neously produced by Y. lipolytica during flask experiments.Apparently, the increment of Glol0 concentration and, hence,the increase of intra-cellular carbon flow, induced the activityof ATP-citrate lyase, while citrate synthase, enzyme-keyresponsible for the synthesis of citric acid [5], remained alwaysactivated. The finding presents importance since glycerolwaste could be recycled through SCO synthesis that couldbe used again for bio-diesel production. Of course, typical
Eng. Life Sci. 2009, 9, No. 6, 468–478 Crude glycerol conversions by Yarrowia lipolytica 475
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

lipid-producing yeasts (species Rhodosporidium toruloides,Lipomyces starkeyi, C. curvatus) can present significantly betterperformances concerning accumulation of fat during growthon sugars or polyols, since carbon flow is directed exclusivelytowards SCO production [14, 53, 54].
In conclusion, waste glycerol was an adequate substrate forY. lipolytica strains. Added value microbial products (citricacid, mannitol and SCO) were produced. The potentialsignificant increase of bio-diesel production in the near futurealready results in the necessity of discovery of variousintegrated (bio)-processes for valorization of this residue(‘‘bio-refinery approach’’). The current study aimed atproviding ways of valorizing this residue, by using it assubstrate of natural Y. lipolytica strains, in conversions rarelypresented in the literature.
Abbreviations : DO, dissolved oxygen; SCO, single cell oil; UI,
unsaturated index
Acknowledgements
The present work was financed by the PABET-2005 project(05PAB105) entitled ‘‘Valorization of agro-industrial residuesthrough the cultivation of the mushroom fungus Lentinulaedodes for the production of metabolic products of biotech-nological interest’’ (General Secretariat of Research andDevelopment, Greek Ministry of Development).
Conflict of interest
The authors have declared no conflict of interest.
5 References
[1] A. P. Zeng, H. Biebl, Bulk chemicals from biotechnology:
the case of 1,3-propanediol production and the new
trends. Adv. Biochem. Eng. Biotechnol. 2002, 74,
239–259.
[2] X. L. Xiu, A. P. Zeng, Present state and perspective of
downstream processing of biologically produced 1,3-propa-
nediol and 2,3-butanediol. Appl. Microbiol. Biotechnol. 2008,
78, 917–926.
[3] S. Papanikolaou, 1,3-Propanediol and citric acid produc-
tion from glycerol-containing waste discharged after bio-
diesel manufacturing process, in A. Koutinas, A. Pandey,
C. Larroche, (Eds.), Current Topics on Bioprocesses in Food
Industry, Vol. II, AsiaTech Publishers Inc., New Delhi 2008,
pp. 381–399.
[4] Z. Wen, D. J. Pyle, S. K. Athalye, Glycerol waste from
biodiesel manufacturing, in G. Aggelis, (Ed.) Microbial
Conversions of Raw Glycerol, Nova Science Publishers Inc.,
New York 2009, pp. 1–7.
[5] S. Papanikolaou, G. Aggelis, Biotechnological valorization of
biodiesel derived glycerol waste through production of single
cell oil and citric acid by Yarrowia lipolytica. Lipid Technol.
2009, 21, 83–87.
[6] Y. Mu, Z. L. Xiu, D. J. Zhang, A combined bioprocess of
biodiesel production by lipase with microbial production of
1,3-propanediol by Klebsiella pneumoniae. Biochem. Eng.
J. 2008, 40, 537–541.
[7] S. Papanikolaou, G. Aggelis, Lipid production by Yarrowia
lipolytica growing on industrial glycerol in a single-stage
continuous culture. Bioresour. Technol. 2002, 82, 43–49.
[8] D. T. Johnson, K. A. Taconi, The glycerin glut: Options for
the value-added conversion of crude glycerol resulting from
biodiesel production. Environ. Prog. 2007, 26, 338–348.
[9] A. Koutinas, R. H. Wang, C. Webb, The biochemurgist-
Bioconversion of agricultural raw materials for chemical
production. Biofuels Bioprod. Bioref. 2007, 1, 24–38.
[10] K. Menzel, A. P. Zeng, W. D. Deckwer, High concentration
and productivity of 1,3-propanediol from continuous
fermentation of glycerol by Klebsiella pneumoniae. Enzyme
Microb. Technol. 1997, 20, 82–86.
[11] K. Menzel, A. P. Zeng, W. D. Deckwer, Enzymatic evidence
for an involvement of pyruvate dehydrogenase in the
anaerobic glycerol metabolism of Klebsiella pneumoniae.
J. Biotechnol. 1997, 56, 135–142.
[12] Y. Mu, D. Zhang, H. Teng, W. Wang, Z. L. Xiu, Microbial
production of 1,3-propanediol by Klebsiella pneumoniae
using crude glycerol from bio-diesel preparation. Biotechnol.
Lett. 2006, 28, 1755–1759.
[13] Z. L. Xiu, B. H. Song, Z. T. Wang, L. H. Sun, E. M. Feng, A. P.
Zeng, Optimization of dissimilation of glycerol to 1,3-
propanediol by Klebsiella pneumoniae in one- and two-stage
anaerobic cultures. Biochem. Eng. J. 2004, 19, 189–197.
[14] P. A. E. P. Meesters, G. N. M. Huijberts, G. Eggink, High-cell-
density cultivation of the lipid accumulating yeast Crypto-
coccus curvatus using glycerol as a carbon source. Appl.
Microbiol. Biotechnol. 1996, 45, 575–579.
[15] Z. Chi, D. J. Pyle, Z. Wen, C. Frear, S. Chen, A laboratory
study of producing docosahexaenoic acid from biodiesel-
waste glycerol by microalgal fermentation. Proc. Biochem.
2007, 42, 1537–1545.
[16] D. J. Pyle, R. A. Garcia, Z. Wen, Producing docosahexaenoic
acid (DHA)-rich algae from biodiesel-derived crude
glycerol: effects of impurities on DHA production and
algal biomass composition. J. Agric. Food Chem. 2008, 56,
3933–3939.
[17] Z. Wen, D. J. Pyle, S. K. Athalye, Production of omega-3
polyunsaturated fatty acids from biodiesel-derived crude
glycerol by microalgal and fungal fermentation, in G. Aggelis,
(Ed.), Microbial Conversions of Raw Glycerol, Nova Science
Publishers Inc., New York 2009, pp. 41–63.
[18] F. Mantzouridou, E. Naziri, M. Z. Tsimidou, Industrial
glycerol as a supplementary carbon source in the production
of b-carotene by Blakeslea trispora. J. Agric. Food Chem. 2008,
56, 2668–2675.
[19] F. Mantzouridou, The potential of raw glycerol in the
production of food grade carotenoids by fungi, in G. Aggelis,
(Ed.), Microbial Conversions of Raw Glycerol, Nova Science
Publishers Inc., New York 2009, pp. 101–124.
[20] G. Mothes, C. Schnorpfeil, J. U. Ackermann, Production of
PHB from crude glycerol. Eng. Life Sci. 2007, 7, 475–479.
[21] G. Mothes, O. Otto, Biotechnological production of
bioplastics from raw glycerol, in G. Aggelis (Ed.), Microbial
476 A. Andre et al. Eng. Life Sci. 2009, 9, No. 6, 468–478
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

Conversions of Raw Glycerol, Nova Science Publishers Inc.,
New York 2009, pp. 65–84.
[22] I. Morgunov, S. Kamzolova, O. Perevoznikova, N. Shishka-
nova, T. Finogenova, Pyruvic acid production by a thiamine
auxotroph of Yarrowia lipolytica. Proc. Biochem. 2004, 39,
1469–1474.
[23] G. Volpato, R. C. Rodrigues, J. X. Heck, M. A. Z. Ayub,
Production of organic solvent tolerant lipase by Staphylo-
coccus caseolyticus EX17 using raw glycerol as substrate.
J. Chem. Technol. Biotechnol. 2008, 83, 821–828.
[24] A. Rywinska, W. Rymowicz, B. arowska, M. Wojtatowicz,
Biosynthesis of citric acid from glycerol by acetate mutants of
Yarrowia lipolytica in fed-batch fermentation. Food Technol.
Biotechnol. 2009, 47, 1–6.
[25] S. Papanikolaou, P. Ruiz-Sanchez, B. Pariset, F. Blanchard,
M. Fick, High production of 1,3-propanediol from industrial
glycerol by a newly isolated Clostridium butyricum strain.
J. Biotechnol. 2000, 77, 191–208.
[26] S. Papanikolaou, L. Muniglia, I. Chevalot, G. Aggelis, I. Marc,
Yarrowia lipolytica as a potential producer of citric acid from
raw glycerol. J. Appl. Microbiol. 2002, 92, 737–744.
[27] S. Papanikolaou, G. Aggelis, Modelling aspects of the
biotechnological valorization of raw glycerol: production of
citric acid by Yarrowia lipolytica and 1,3-propanediol by
Clostridium butyricum. J. Chem. Technol. Biotechnol. 2003,
78, 542–547.
[28] S. Papanikolaou, M. Fick, G. Aggelis, The effect of raw
glycerol concentration on the production of 1,3-propanediol
by Clostridium butyricum. J. Chem. Technol. Biotechnol. 2004,
79, 1189–1196.
[29] S. Papanikolaou, S. Fakas, M. Fick, I. Chevalot, M. Galiotou-
Panayotou, M. Komaitis, I. Marc, G. Aggelis, Biotechnolo-
gical valorisation of raw glycerol discharged after bio-diesel
(fatty acid methyl-esters) manufacturing process: production
of 1,3-propanediol, citric acid and single cell oil. Biomass
Bioenerg. 2008, 32, 60–71.
[30] S. B. Imandi, V. R. Bandaru, S. R. Somalanka, H. R. Garapati,
Optimization of medium constituents for the production of
citric acid from byproduct glycerol using Doehlert experi-
mental design. Enzyme Microb. Technol. 2007, 40, 1367–1372.
[31] W. E. Levinson, C. P. Kurtzman, T. M. Kuo, Characterization
of Yarrowia lipolytica and related species for citric acid
production from glycerol. Enzyme Microb. Technol. 2007, 41,
292–295.
[32] W. Rymowicz, A. Rywinska, B. Zarowska, P. Juszczyk, Citric
acid production from raw glycerol by acetate mutants of
Yarrowia lipolytica. Chem. Pap. 2006, 60, 391–394.
[33] W. Rymowicz, A. Rywinska, W. G"adkowski, Simultaneous
production of citric acid and erythritol from crude glycerol
by Yarrowia lipolytica Wratislavia K1. Chem. Pap. 2008, 62,
239–246.
[34] A. Rywinska, W. Rymowicz, Citric acid production from raw
glycerol by Yarrowia lipolytica Wratislavia 1.31, in G. Aggelis,
(Ed.), Microbial Conversions of Raw Glycerol, Nova Science
Publishers Inc., New York 2009, pp. 19–30.
[35] S. Paramithiotis, M. R. A. Mueller, M. A. Ehrmann,
E. Tsakalidou, H. Seiler, R. Vogel, G. Kalantzopoulos, Poly-
phasic identification of wild yeast strains isolated from Greek
sourdoughs. Syst. Appl. Microbiol. 2000, 23, 156–164.
[36] S. Fakas, S. Papanikolaou, M. Galiotou-Panayotou,
M. Komaitis, G. Aggelis, Lipids of Cunninghamella echinulata
with emphasis to g-linolenic acid distribution among lipid
classes. Appl. Microbiol. Biotechnol. 2006, 73, 676–683.
[37] S. Anastassiadis, H. J. Rehm, Continuous citric acid secretion
by a high specific pH dependent active transport system in
yeast Candida oleophila ATCC 20177. Electron J. Biotechnol.
2005, 8, 146–161.
[38] C. Ratledge, J. Wynn, The biochemistry and molecular
biology of lipid accumulation in oleaginous microorganisms.
Adv. Appl. Microbiol. 2002, 51, 1–51.
[39] E. P. W. Kets, E. A. Galinski, M. De Wit, J. A. M. De Bont,
H. J. Heipieper, Mannitol, a novel bacterial compatible
solute in Pseudomonas putida. J. Bacteriol. 1996, 178,
6665–6670.
[40] N. Gutierrez, I. McKay, C. French, J. Brooks, I. Maddox,
Repression of galactose utilization by glucose in a citrate-
producing yeast Candida guilliermondii. J. Ind. Microbiol.
Biotechnol. 1993, 11, 143–146.
[41] G. Aggelis, N. Athanassopoulos, A. Paliogianni, M. Komaitis,
Effect of Teucrium polium L. extract on the growth and the
fatty acid composition of Saccharomyces cerevisiae and
Yarrowia lipolytica. Antonie van Leeuwenhoek 1998, 73,
195–198.
[42] S. Papanikolaou, M. Galiotou-Panayotou, I. Chevalot,
M. Komaitis, I. Marc, G. Aggelis, Influence of glucose and
saturated free-fatty acid mixtures on citric acid and lipid
production by Yarrowia lipolytica. Curr. Microbiol. 2006, 52,
134–142.
[43] S. Papanikolaou, O. Gortzi, E. Margeli, I. Chinou,
M. Galiotou-Panayotou, S. Lalas, Effect of Citrus essential
oil addition upon growth and cellular lipids of Yarrowia
lipolytica yeast. Eur. J. Lipid Sci. Technol. 2008, 110,
997–1006.
[44] L. Moeller, B. Strehlitz, A. Aurich, A. Zehnsdorf, T. Bley,
Optimization of citric acid production from glucose by
Yarrowia lipolytica. Eng. Life Sci. 2007, 7, 504–511.
[45] S. Kamzolova, I. Morgunov, A. Aurich, S. Perevoznikova,
N. Shiskanova, U. Stottmeister, T. Finogenova, Lipase
secretion and citric acid production in Yarrowia lipolytica
yeast grown on animal and vegetable fat. Food Technol.
Biotechnol. 2005, 43, 113–122.
[46] S. Kamzolova, N. Shishkanova, I. Morgunov, T. Finogenova,
Oxygen requirements for growth and citric acid produc-
tion of Yarrowia lipolytica. FEMS Yeast Res. 2003, 3,
217–222.
[47] M. Wojtatowicz, W. Rymowicz, H. Kautola, Comparison of
different strains of the yeast Yarrowia lipolytica for citric acid
production from glucose hydrol. Appl. Biochem. Biotechnol.
1991, 31, 165–174.
[48] K. D. Rane, K. A. Sims, Production of citric acid by
Candida lipolytica Y 1095: Effect of glucose concentration on
yield and productivity. Enzyme Microb. Technol. 1993, 15,
646–651.
[49] S. Kamzolova, T. Finogenova, Y. Luniva, S. Perevoznikova,
L. Minachova, I. Morgunov, Characteristics of the growth on
rapeseed oil and synthesis of citric and isocitric acids by
Yarrowia lipolytica yeasts. Microbiology (Moscow) 2007, 76,
20–24.
Eng. Life Sci. 2009, 9, No. 6, 468–478 Crude glycerol conversions by Yarrowia lipolytica 477
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

[50] K. T. Klasson, E. C. Clausen, J. L. Gaddy, Continuous
fermentation for the production of citric acid from
glucose. Appl. Biochem. Biotechnol. 1989, 20/21,
491–509.
[51] C. Moresi, Effect of glucose concentration on citric acid
production by Yarrowia lipolytica–kinetics of the tropho-
phase, citrate lag phase and idiophase. J. Chem. Technol.
Biotechnol. 1994, 60, 387–395.
[52] S. Anastassiadis, A. Aivasidis, C. Wandrey, Citric
acid production by Candida strains under intracellular
nitrogen limitation. Appl. Microbiol. Biotechnol. 2002, 60,
81–87.
[53] Y. H. Li, Z. B. Zhao, F. W. Bai, High density cultivation of
oleaginous yeast Rhodosporidium toruloides Y4 in fed-batch
culture. Enzyme Microb. Technol. 2007, 41, 312–317.
[54] X. Zhao, X. Kong, Y. Hua, B. Feng, Z. B. Zhao,
Medium optimization for lipid production through co-
fermentation of glucose and xylose by the oleaginous yeast
Lipomyces starkeyi. Eur. J. Lipid Sci. Technol. 2008, 110,
405–412.
478 A. Andre et al. Eng. Life Sci. 2009, 9, No. 6, 468–478
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com
Related Documents










![Engineering the oleaginous yeast Yarrowia lipolytica for ... · Liu et al. Biotechnol Biofuels Page 2 of 11 compounds.Forexample,artemisininisasesquiterpene endoperoxidewitheectiveantimalarialproperties[12],](https://static.cupdf.com/doc/110x72/5f7b487302e1e8790c2f553c/engineering-the-oleaginous-yeast-yarrowia-lipolytica-for-liu-et-al-biotechnol.jpg)
