http://informahealthcare.com/bty ISSN: 0738-8551 (print), 1549-7801 (electronic) Crit Rev Biotechnol, Early Online: 1–14 ! 2014 Informa Healthcare USA, Inc. DOI: 10.3109/07388551.2014.961003 REVIEW ARTICLE Biotechnological approaches to the production of shikonins: a critical review with recent updates Sonia Malik 1,2 , Shashi Bhushan 1 , Madhu Sharma 1 , and Paramvir Singh Ahuja 1 1 Division of Biotechnology, CSIR-Institute of Himalayan Bioresource Technology, Palampur, Himachal Pradesh, India and 2 Department of Chemical Biology and Genetics, Centre of the Region Hana ´ for Biotechnological and Agricultural Research, Palacky ´ University, Olomouc, Czech Republic Abstract Shikonins are commercially important secondary compounds, known for array of biological activities such as antimicrobial, insecticidal, antitumor, antioxidants, etc. These compounds are usually colored and therefore have application in food, textiles and cosmetics. Shikonin and its derivatives, which are commercially most important of the naphthoquinone pigments, are distributed among members of the family Boraginaceae. These include different species of Lithospermum, Arnebia, Alkanna, Anchusa, Echium and Onosma. The growing demand for plant- based natural products has made this group of compounds one of the enthralling targets for their in vitro production. The aim of this review is to highlight the recent progress in production of shikonins by various biotechnological means. Different methods of increasing the levels of shikonins in plant cells such as selection of cell lines, optimization of culture conditions, elicitation, in situ product removal, genetic transformation and metabolic engineering are discussed. The experience of different researchers working worldwide on this aspect is also considered. Further, to meet market demand, the needs for continuous and reliable production systems, as well as future prospects, are included. Keywords Alkannin, boraginaceae, cell culture, in vitro culture, naphthoquinones, natural products, pigments, secondary metabolites History Received 21 December 2013 Revised 16 July 2014 Accepted 23 July 2014 Published online 9 October 2014 Introduction Plants are capable of synthesizing a bewildering array of chemical compounds, which are used as pharmaceuticals, agrochemicals, flavors, fragrances, pigments, dyes, cosmetics and food additives (Lubbe & Verpoorte, 2011). These chemical compounds, commonly known as secondary metab- olites or natural products are heterogeneous compounds, generally not only required for normal growth and develop- ment of plants but increase the plant’s ability to survive and overcome local challenges by allowing them to interact with their environment (Harborne, 1993). These are produced during specific developmental stages of plant or under specific (seasonal, stress or nutritional) conditions and their production levels are often low (Malik et al., 2009, 2010a, 2013a; Verpoorte, 2000). Secondary metabolites of plant origin are in demand due to a resurgence of public interest in plant-based products. Every year, the market for plant-based products is increasing at a rate of 12–15% (Raskin et al., 2002). Table 1 summarizes the source of plant species for various secondary metabolites, their economic use and market value. Secondary metabolites are mainly classified into three main groups: polyphenols, alkaloids and terpenes. Naphthoquinones (belong to a class of polyphenols formed on a C6–C4 skeleton) have developed great interest due to their broad range of activities (Babula et al., 2009). These are widespread in nature as secondary metabolites of fungi, micro-organisms as well as plants. Naphthoquinones which include shikonin, alkannin and their derivatives, juglans and many more present a larger group of quinone pigments (Figure 1). The molecular formula of naphthoquinone is C 10 H 6 O 2 . These metabolites range from simple structures such as 5-hydroxy derivative juglone, to pigments with isoprenyl attachment such as alkannin. Shikonin, and its derivatives, are commercially the most important of the naphthoquinone pigments and this review is focused on the production of shikonin and their derivatives in plants. The main objective of this review is to highlight the reproducible methods for in vitro production of shikonins and to illustrate the different production systems for these important classes of compounds. Shikonins: their occurrence, distribution and economic value Shikonin and its derivatives are the commercially most important of the napthoquinone pigments, distributed among members of the family Boraginaceae. These include different species of Lithospermum, Arnebia, Alkanna, Anchusa, Address for correspondence: Sonia Malik, Department of Chemical Biology and Genetics, Centre of the Region Hana ´ for Biotechnological and Agricultural Research, Palacky ´ University, S ˇ lechtitelu ˚ 11, 783 71 Olomouc, Czech Republic. Tel:+420 585 632 173. Fax: +420 585 634 870. E-mail: [email protected] Critical Reviews in Biotechnology Downloaded from informahealthcare.com by UNICAMP on 10/30/14 For personal use only.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

http://informahealthcare.com/btyISSN: 0738-8551 (print), 1549-7801 (electronic)
Crit Rev Biotechnol, Early Online: 1–14! 2014 Informa Healthcare USA, Inc. DOI: 10.3109/07388551.2014.961003
REVIEW ARTICLE
Biotechnological approaches to the production of shikonins: a criticalreview with recent updates
Sonia Malik1,2, Shashi Bhushan1, Madhu Sharma1, and Paramvir Singh Ahuja1
1Division of Biotechnology, CSIR-Institute of Himalayan Bioresource Technology, Palampur, Himachal Pradesh, India and 2Department of Chemical
Biology and Genetics, Centre of the Region Hana for Biotechnological and Agricultural Research, Palacky University, Olomouc, Czech Republic
Abstract
Shikonins are commercially important secondary compounds, known for array of biologicalactivities such as antimicrobial, insecticidal, antitumor, antioxidants, etc. These compounds areusually colored and therefore have application in food, textiles and cosmetics. Shikonin and itsderivatives, which are commercially most important of the naphthoquinone pigments, aredistributed among members of the family Boraginaceae. These include different species ofLithospermum, Arnebia, Alkanna, Anchusa, Echium and Onosma. The growing demand for plant-based natural products has made this group of compounds one of the enthralling targets fortheir in vitro production. The aim of this review is to highlight the recent progress in productionof shikonins by various biotechnological means. Different methods of increasing the levels ofshikonins in plant cells such as selection of cell lines, optimization of culture conditions,elicitation, in situ product removal, genetic transformation and metabolic engineering arediscussed. The experience of different researchers working worldwide on this aspect is alsoconsidered. Further, to meet market demand, the needs for continuous and reliable productionsystems, as well as future prospects, are included.
Keywords
Alkannin, boraginaceae, cell culture,in vitro culture, naphthoquinones, naturalproducts, pigments, secondary metabolites
History
Received 21 December 2013Revised 16 July 2014Accepted 23 July 2014Published online 9 October 2014
Introduction
Plants are capable of synthesizing a bewildering array of
chemical compounds, which are used as pharmaceuticals,
agrochemicals, flavors, fragrances, pigments, dyes, cosmetics
and food additives (Lubbe & Verpoorte, 2011). These
chemical compounds, commonly known as secondary metab-
olites or natural products are heterogeneous compounds,
generally not only required for normal growth and develop-
ment of plants but increase the plant’s ability to survive and
overcome local challenges by allowing them to interact with
their environment (Harborne, 1993). These are produced
during specific developmental stages of plant or under
specific (seasonal, stress or nutritional) conditions and their
production levels are often low (Malik et al., 2009, 2010a,
2013a; Verpoorte, 2000).
Secondary metabolites of plant origin are in demand due to
a resurgence of public interest in plant-based products. Every
year, the market for plant-based products is increasing at a
rate of 12–15% (Raskin et al., 2002). Table 1 summarizes the
source of plant species for various secondary metabolites,
their economic use and market value.
Secondary metabolites are mainly classified into three
main groups: polyphenols, alkaloids and terpenes.
Naphthoquinones (belong to a class of polyphenols formed
on a C6–C4 skeleton) have developed great interest due to
their broad range of activities (Babula et al., 2009). These are
widespread in nature as secondary metabolites of fungi,
micro-organisms as well as plants. Naphthoquinones which
include shikonin, alkannin and their derivatives, juglans and
many more present a larger group of quinone pigments
(Figure 1). The molecular formula of naphthoquinone is
C10H6O2. These metabolites range from simple structures
such as 5-hydroxy derivative juglone, to pigments with
isoprenyl attachment such as alkannin. Shikonin, and its
derivatives, are commercially the most important of the
naphthoquinone pigments and this review is focused on
the production of shikonin and their derivatives in plants. The
main objective of this review is to highlight the reproducible
methods for in vitro production of shikonins and to illustrate
the different production systems for these important classes
of compounds.
Shikonins: their occurrence, distribution andeconomic value
Shikonin and its derivatives are the commercially most
important of the napthoquinone pigments, distributed among
members of the family Boraginaceae. These include different
species of Lithospermum, Arnebia, Alkanna, Anchusa,
Address for correspondence: Sonia Malik, Department of ChemicalBiology and Genetics, Centre of the Region Hana for Biotechnologicaland Agricultural Research, Palacky University, Slechtitelu 11, 783 71Olomouc, Czech Republic. Tel:+420 585 632 173. Fax: +420 585 634870. E-mail: [email protected]
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

Echium and Onosma. Shikonin, alkannin and their derivatives
are known to possess a wide range of pharmaceutical
properties and are also used as natural dyes for coloring
silk, cosmetics, as well as food additives (Andujar et al.,
2013; Babula et al., 2009; Lu et al., 2013).
The biolipstick, a lipstick with biological colorant
‘‘shikonin’’ was a big hit in the cosmetics market in 1985.
In spite of its high price (US$ 30 a stick), 2 million pieces
(sticks) were sold within a few days by Kanebo, Japanese
Cosmetics Company. The idea was that shikonin-containing
lipstick is not only biologically prepared but it also protects
against bacteria and has other anti-microbial activities
(Renneberg, 2008).
Shikonin was the first product produced from cell cultures
of Lithospermum erythrorhizon by Mitsui Petrochemical
Industries Ltd. (now Mitsui Chemicals Inc.), Tokyo, Japan
in 1984 (DiCosmo & Misawa, 1996). The production of these
secondary metabolites in its natural habitat is hampered due
to poor germination, non-availability of planting material,
over-exploitation and geographical limitation as well as lack
of support for R&D work by the industries. Also, the plant
requires three to four years for noticeable production of these
pigments. In this context, the in vitro production of commer-
cially important secondary metabolites by various biotechno-
logical means offers an attractive proposition (Malik et al.,
2013b; Rao & Ravishankar, 2002).
Biosynthesis of shikonins
In L. erythrorhizon, it is well documented that shikonin is
derived from two precursors originating from different
pathways (Figure 2). The aromatic precursor 4-hydroxyben-
zoic acid (4HB) formed via the shikimate and the phenyl-
propanoid pathway, while the isoprenoid precursor,
geranyldiphosphate (GPP) is derived from the mevalonate
pathway (Li et al., 1998). The 4HB geranyltransferase
reaction links the aromatic precursor 4HB to the isoprenoid
precursor GPP and yields 3-geranyl-4-hydroxybenzoate
(GBA), which is the first specific intermediate in shikonin
biosynthesis (Heide & Tabata, 1987b).
It has been reported that hydroxyl-3-methylglutaryl-
coenzymeA reductase (HMGR) and 4HB geranyltransferase
Table 1. Useful secondary metabolites from plants, their uses and market value.
Plant species Family Secondary product Economic use Price* (US$/Kg)
PharmaceuticalsRauwolfia serpentina Apocynaceae Ajmaline Sedative 75 000Artemisia annua Asteraceae Artemisinin Anti-malarial 400Atropa belladonna Solanaceae Atropine Anti-cholinergic n/aCatharanthus roseus Apocynaceae Ajmalicine Anticancer 37 000
Vinblastine Anticancer 1 000 000Vincristine Anti-leukemic 3 000 000
Coptis japonica Ranunculaceae Berberine Antiseptic 3250Papaver somniferum Papaveraceae Codeine Sedative 17 000
Morphine Analgesic 340 000Colchicum speciosum Colchicaceae Colchicine Antitumor 35 000Digitalis lanata Scrophulariaceae Digoxin Cardiatonic 3000Dioscorea deltoidea Dioscoreaceae Diosgenin Steroid, anti-fertility agent 1000Orchrosia elliptica Apocynaceae Ellipticine Antitumor 240 000Podophyllum sp., Linum sp. Berberidaceae Podophyllotoxin Viral diseases and skin cancer 800 000Cinchona ledgeriana Rubiaceae Quinine Anti-malarial 500Sanguinaria canadensis Papaveraceae Sanguinarine Antibiotic, expectorant, analgesic 4800Datura stramonium Solanaceae Scopolamine Anti-cholinergic n/a
PigmentsMorinda citrifolia Polygonaceae Anthraquinones Natural dye, purgative, cathartic n/aLithospermum erythrorhizon,
Arnebia sp.Boraginaceae Shikonin derivatives Cosmetics, natural dye,
anti-cancer, anti-bacterial4500
AgrochemicalsAzadirachta indica Meliaceae Azadirachtin Insecticide n/aJuniperus viriginiana Cupressaceae Cederene Repellant n/aNicotiana tabaccum Solanaceae Nicotine Insecticide n/aChrysanthemumcinerariaefolium
Asteraceae Pyrethrins, Pyretheroids Insecticide 413
Flavors and fragrancesCapsicum frutescens Solanaceae Capsicin Condiment n/aCymbopogon winterianus Gramineae Citronellal Food flavor n/aCrocus sativus Iridaceae Crocin Saffron n/aJasminum spp. Oleaceae Jasmine (oil) Perfume 350Citrus sp. Rutaceae Limonene Food flavor n/aMentha sp. Labiatae Menthol Food flavor n/aVanilla planifolia Orchidaceae Vanillin Flavor 1000
n/a, not available.
Figure 1. Chemical structure of Naphthoquinone.
2 S. Malik et al. Crit Rev Biotechnol, Early Online: 1–14
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

are the important enzymes for regulation of shikonin biosyn-
thesis (Gaisser & Heide, 1996; Heide et al., 1989; Lange
et al., 1998). Srinivasan & Ryu (1992) observed high
activities of the biosynthetic enzymes, phenylalanine ammo-
nia lyase (PAL) and 3-hydroxyl-3-methylglutaryl-CoA-reduc-
tase (HMGR) in shikonin producing cells of L. erythrorhizon.
Biotechnological approaches for the productionof shikonins
Biotechnological advancements provide us the opportunity to
make use of cells, tissue or organs of economically important
plants by growing them under in vitro conditions and to
genetically manipulate them to obtain the desired compounds.
Further, in vitro cultures could supersede the extraction of
valuable compounds from wild or cultivated plants (Kumar
et al., 2011; Malik et al., 2011a, 2014; Ravishankar & Rao,
2002).
In vitro production of secondary metabolites by cell and
tissue culture systems could be helpful in understanding the
biosynthesis of these compounds. Table 2 summarizes the
commercial or semi-industrial production of secondary com-
pounds from different plant species by in vitro means.
In vitro production of shikonin and its derivatives
Micropropagation and regeneration in shikoninbearing plants
Plant cells or tissues exhibit totipotency under appropriate
conditions and this potential can be exploited by the selection
of suitable media and culture conditions. Ji & Wang (2001)
obtained direct regeneration from excised leaves of Arnebia
euchroma on a modified MS (Murashige & Skoog, 1962)
medium with 0.4 mg/l Naphthalene Acetic Acid (NAA) and
1.5 mg/l Kinetin (Kn). In another report, Jiang et al. (2005)
used Thidiazuron (TDZ, 4.5 lM) for direct regeneration from
cotyledons and hypocotyls in same plant species. The
importance of pre-culture time and concentration of TDZ
for direct regeneration from in vitro leaves (attached to
shoots) of A. euchroma was studied by our group (Malik
et al., 2010b). Direct regeneration could be obtained from
shoots, which were initially pre-cultured for 40 days on
medium having higher TDZ concentration (20.0 lM) followed
by transfer to lower concentration (5.0 lM; Figure 3a–c). The
identity of shoot buds was confirmed by histological studies
(Figure 3d–f). Regenerated shoots were cultured on Kn
(5.0 lM) containing media for proliferation and rooted
plantlets were transferred to greenhouse (Figure 3g and h).
The mode of regeneration depends mainly on the species,
type of organ used and the level of growth regulating
substances in the medium. Manjkhola et al. (2005) achieved
shoot regeneration and somatic embryogenesis from leaf
derived callus on MS medium supplemented with 2.5 mM
Benzylaminopurine (BAP) + 2.5 mM Indole-3-butyric acid
(IBA) and 2.5 mM BAP + 1.0 mM IBA, respectively, while
root derived callus did not show any sign of organogenesis.
Later on, Ji & Wang (2001) reported regeneration from bud
derived callus from the same plant species after 3 months
culture on Linsmaier & Skoog (LS) (1965) medium contain-
ing NAA in combination with Kn or BAP.
Figure 2. Shikonin biosynthesis pathway(Singh et al., 2010).
Glucose Glyceraldehyde 3-phosphate
Chorismic acid
Phenylalanine
Coumaric acid
Dimethylallyl pyrophosphate
Cinnamic acid
p-hydroxybenzoic acid
SHIKIMATE PATHWAY
Diphosphomevalonate Decarboxylase
Deoxyshikonin Shikonin
m-geranyl-p-hydroxybenzoic acid
m-geranylhydroxy quinone
Mevalonate pyrophosphate
HMGCoA synthase
Pyruvate
Acetoacetyl CoA
3-Hydroxy 3-methylglutaryl CoA
Mevalonate
Mevalonate phosphate
Isopentenyl pyrophosphate
Geranylpyrophosphate
Mevalonate kinase
4-Coumaroyl Co A ligase IPP Isomerase
Acetoacetyl CoA
PHB geranyl transferase
Phenylalanine ammonia lyase
Phosphomevalonate kinase
Acetyl CoA
HMG CoA reductase
Geranyl diphosphate synthase
MEVALONATE PATHWAY
Cinnamic acid-4 hydroxylase
DOI: 10.3109/07388551.2014.961003 Biotechnological approaches to the production of shikonins 3
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

Tissue and cell culture
The first successful attempt for in vitro production of shikonin
derivatives was made from callus cultures of L. erythrorhizon
by Tabata et al. (1974). The induction, growth and color of
callus depend upon the type of explant used and presence of
plant growth regulators (PGRs). In A. euchroma, friable callus
was obtained from in vitro leaves within 20–25 days of culture
on MS medium fortified with 10.0 mM BAP and 5.0 mM IBA
(Figure 4).
Tabata et al. (1974) observed that callus culture derived
from seedlings of L. erythrorhizon accumulated shikonin
derivatives on LS medium supplemented with IAA (1.0 mM)
and Kn (10.0mM). The derivatives were similar to that of root
bark of the plant. Exposure to light and presence of 2,4-D in
the medium suppressed the synthesis of these compounds.
Callus cultures of L. erythrorhizon were able to produce
shikonin derivatives in agar gelled LS media but cell
suspension cultures in the liquid medium of same compos-
ition did not show the production of these derivatives (Fujita
et al., 1981a).
It has been reported that production of shikonin derivatives
from cells of L. erythrorhizon is affected by the supply of
nutrients, availability of oxygen and the presence of agar in
culture medium (Hara et al., 1987). Fujita et al. (1981b)
established the cell suspension cultures of L. erythrorhizon in
a modified WH medium with the possibility of large scale
production of these compounds under in vitro conditions. The
cell suspension cultures were shown to produce all the
shikonin derivatives as found in the roots of the intact plant
(Fujita et al., 1983). The annual consumption of shikonin
derivatives is 150 kg (Nosov, 2012). To meet the demand of
shikonin derivatives in Japan, almost 10 000 kg roots of L.
erythrorhizon are used annually. This plant is almost extinct
in Japan due to uninhibited collection from the wild (Fujita,
1988b). Even cultivation of these plants failed to provide
sufficient raw materials for commercial production of this
valuable product. Moreover, shikonin derivatives reached
only a maximum of 1–2% in their roots after 5–7 years growth
(Fujita, 1988a,b). Therefore, Japan had to import ten metric
tons of raw material (valued app. 4500 US$ per kg) annually
from South Korea and China. However, supply of plant
material from countries other than Japan could not meet the
increasing demand of this valuable product due to geographic
reasons as well as the high price associated with import of the
plants (Renneberg, 2008). Hence, Japanese researchers
focused on in vitro production of these compounds. Mitsui
Chemicals Inc., Japan became successful in producing
shikonin from cell culture and presented the first example
of commercial production of a secondary metabolite from cell
culture. Chemical synthesis of shikonin is not economically
Table 2. Commercial and emerging/semi-emerging large-scale in vitro production of plant secondary metabolites.
Bioreactor
Product Plant species Type Max. Volume (L) Producer(s) Country
Anthocyanins (c) Euphorbia milliiAralia cordata
Rotary culture system – Nippon Paint Company Japan
Arbutin (c) Catharanthus roseus – – Mitsui PetrochemicalIndustries
Japan
Berberine (c) Thalictrum minus,Coptis japonica
Batch and continuousflow; impeller driven
4000 Mitsui PetrochemicalIndustries Ltd.
Japan
Betacyanins (c) Beta vulgaris – – Nippon Shinyaku JapanCamptothecin (h) Camptotheca acuminate – – ROOTec bioactives GmbH SwitzerlandCarthamin (c) Carthamus tinctorius – – Kibun JapanDigoxin (c) Digitalis lanata Airlift system 300 Boehringer GermanyGinsenosides (c, r) Panax ginseng – 20 000 Nitto Denko Corp.
CBN BiotechJapanRepublic of Korea
ImmunostimulatingPolysaccharides (c)
Echinacea purpurea,E. auguslifolia
– – Diversa Germany
Podophyllotoxin (c,h) Podophyllum sp. – – Nippon OilROOTec bioactives GmbH
JapanSwitzerland
Purpurin (c) Rubia akane – – Mitsui PetrochemicalIndustries Ltd., NittoDenko Corp.
Japan
Rosmarinic acid (c) Coleus blumei – – Nattermann GermanySanguinarine (c) Papaver somniferum Airlift system 300 Vipont Research Labs USAScopolamine (c) Duboisia sp. – – Sumitomo Chemical
IndustriesJapan
Shikonin (c) Lithospermum erythrorhizon Batch 750 Mitsui PetrochemicalIndustries Ltd.
Japan
Taxol (Paclitaxel) (c) Taxus brevifolia,T. cuspidata
Impeller driven and air-lift reactor systems
75 000 ESCAgenetics Phyton Inc.Nippon Oil CompanySamyang, Genex Co. Ltd.Bristol-Myers Squibb Co.
USAJapanKoreaGermany
Vanillin(Phyto vanilla) (c)
Vanilla planifolia Impeller driven reactor 72 ESCAgenetics USA
Large-scale commercial production of secondary compounds from in vitro culture is recognized for shikonin, ginsenosides, berberine and taxol. c: cellculture; r: root culture; h: hairy root culture.
Data collected from Malik et al. (2011a) and Nosov (2012).
4 S. Malik et al. Crit Rev Biotechnol, Early Online: 1–14
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

feasible as it involves twelve step reactions and the final yield
is only 0.7% (Terada et al., 1983). In order to find an
alternative plant source to meet the still emerging demand
of this product, other members of the family Boraginaceae
were also exploited for their potential to produce shikonin
derivatives.
Pietrosiuk et al. (1999) isolated shikonin derivatives and
pyrrolizidine alkaloids from callus and cell suspension
cultures of A. euchroma. The presence of acetylshikonin
and b-acetoxyisovalerylshikonin in cell suspension cultures of
A. euchroma has been reported by our group (Sharma et al.,
2008). The cell culture of A. hispidissima was found to
produce arnebins (Jain et al., 1999). There are reports on the
production of shikonin derivatives in callus cultures of
Echium lycopsis (Fukui et al., 1983; Inouye et al., 1981).
Isolation procedures for isohexenylnaphthazarin compounds
from callus and cell suspension cultures of A. euchroma have
been demonstrated by Damianakos et al. (2012).
Methods for improving productivity
Although plant cells are capable of producing secondary
metabolites under in vitro conditions, until now only a few
plant metabolites have been commercialized or reached at
industrial scale through cell culture. In order to improve the
yield for commercial exploitation, efforts were focused on
Figure 3. Regeneration in A. euchroma. (a) Shoot cultures with vitrified leaves (arrow marked) on 20.0 lM TDZ supplemented medium. (b) Shoot budregeneration (arrow marked) from intact leaves (attached to shoots) on 5.0 lM TDZ. (c) Growth of shoot buds on 5.0 lM TDZ. (d) A part of swollenleaf with protuberances (arrow marked) and scattered vascular bundles. (e) Initiation of shoot bud (continuation of vascular strand with the motherexplant). (f) Shoot bud showing shoot apex (SA), subtending leaves (SL) and vascular strand (VS) in continuity with the mother explant (arrowmarked). (g) Rooted plantlet on 0.25mM IBA after 20 days. (h) Hardened plants under greenhouse conditions. Reprinted from Malik et al. (2010b)Copyright (2010) International Federation for Cell Biology, with permission from John Wiley and Sons.
Figure 4. A. euchroma callus (a) induction and (b) proliferation on MSmedium with BAP (10.0 mM) + IBA (5.0mM).
DOI: 10.3109/07388551.2014.961003 Biotechnological approaches to the production of shikonins 5
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.
isolating the biosynthetic activities of cultured cells, by
selecting high yielding cell lines, optimizing the nutrient
media and the culture conditions (Dornenburg & Knorr,
1995).
Selection of high yielding cell lines
Several techniques have been employed to isolate secondary
metabolite producing cells from non-producing ones (Dix,
1986). Colored cells are visually selected at subculture by
colorimeteric tests, fluorescence (Chaprin & Ellis, 1984) or
optical microspectrophotometry (Hall & Yeoman, 1986).
Such a strategy resulted in stable and high production of
shikonin derivatives in L. erythrorhizon (Tabata et al., 1974).
Using cell-aggregate cloning techniques, Mizukami et al.
(1978) obtained two stable cell lines in L. erythrorhizon with
improved growth and pigment production. In A. euchroma, we
screened eight cell lines showing either a higher growth rate
or pigmentation over a period of 2 years. Amongst colour
producing lines, the stable cell line was selected which was
used for further studies (Figure 5).
Bulgakov et al. (2001) used PFP for selection of high
pigment producing cell lines in A. euchroma and reported
two-fold higher shikonin content in the selected p-fluorophe-
nylalanine resistant callus lines. Zakhlenjuk et al. (1993) also
selected PFP-resistant cell lines in A. euchroma where the
biosynthesis of shikonins was found to be inhibited in this
selected cell line.
Optimization of nutrient media
The composition of the culture medium had a pronounced
effect on cell growth and shikonin derivatives production.
Fujita et al. (1981b) studied the effect of different basal
media and reported the optimum cell growth in Linsmaier &
Skoog (1965) (LS) medium. However, shikonin derivatives
were obtained only in the White (1954) (WH) medium. In cell
suspension cultures of A. euchroma, a dark red pigment was
observed when WH medium was modified by varying the
concentration of different macro nutrients (designated as
Arnebia Production Medium, APM; Figure 6, Malik et al.,
2008). The red pigment extracted from cells growing in APM
turned blue when dissolved in 2.5% KOH solution, indicating
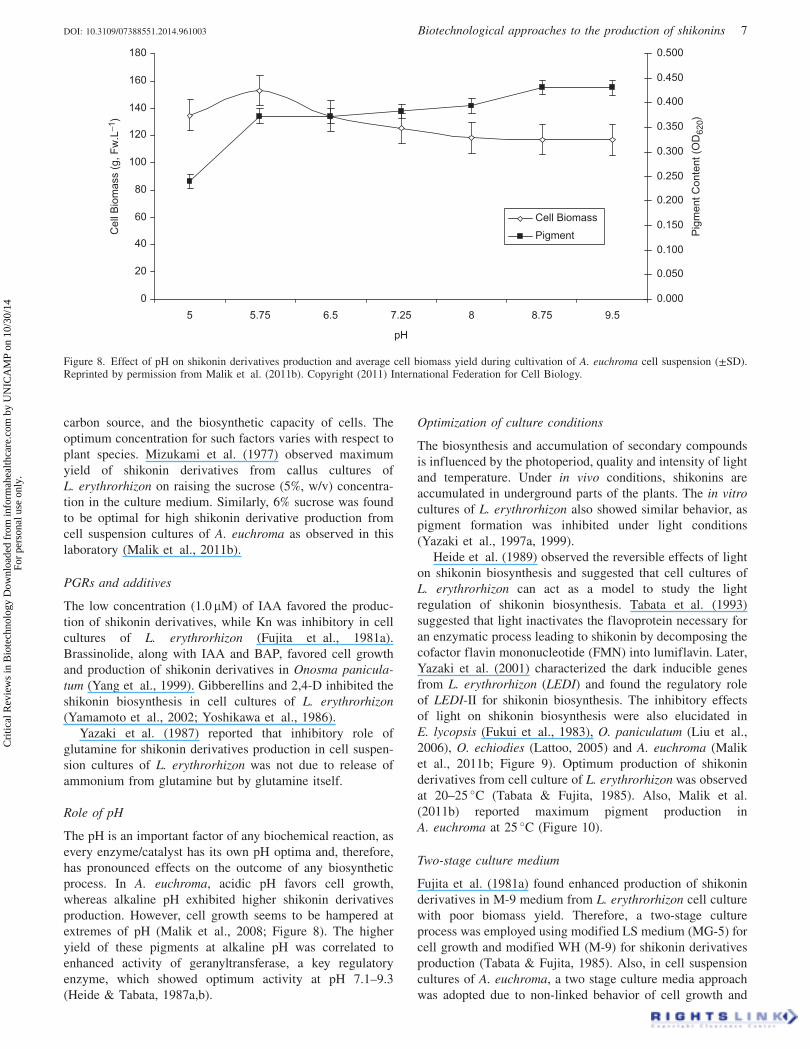
the presence of shikonins (Figure 7). Spectrophotometric
analysis showed maximum absorbance at 620 nm. The
pigment was confirmed to be a shikonin derivative as
compared with the standard (Malik, 2009).
In Echium italicum, the maximum induction of callus
producing shikonin acetate was reported in the WH medium
(Zare et al., 2011). On the basis of a requirement for each
component in the culture medium for the optimal production
of shikonin derivatives from cell culture of L. erythrorhizon,
M-9 medium has been devised and the yield of shikonin
derivatives from cells was enhanced from 120 mg/l (2% DW),
as found in WH medium, to 1400 mg/l (12% DW; Fujita
et al., 1981a).
Carbon source
The growth and production of secondary compounds in cell
culture depends greatly on the type and concentration of
Figure 7. (a) Cells of A. euchroma showing pigment in APM and(b) qualitative reaction of red pigment with 2.5% KOH solution.
Figure 6. Cell suspension cultures of A. euchroma on (a) MS and(b) APM.
Figure 5. (a) Non-pigmented and (b) pigmented callus line ofA. euchroma obtained after selection.
6 S. Malik et al. Crit Rev Biotechnol, Early Online: 1–14
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.
carbon source, and the biosynthetic capacity of cells. The
optimum concentration for such factors varies with respect to
plant species. Mizukami et al. (1977) observed maximum
yield of shikonin derivatives from callus cultures of
L. erythrorhizon on raising the sucrose (5%, w/v) concentra-
tion in the culture medium. Similarly, 6% sucrose was found
to be optimal for high shikonin derivative production from
cell suspension cultures of A. euchroma as observed in this
laboratory (Malik et al., 2011b).
PGRs and additives
The low concentration (1.0mM) of IAA favored the produc-
tion of shikonin derivatives, while Kn was inhibitory in cell
cultures of L. erythrorhizon (Fujita et al., 1981a).
Brassinolide, along with IAA and BAP, favored cell growth
and production of shikonin derivatives in Onosma panicula-
tum (Yang et al., 1999). Gibberellins and 2,4-D inhibited the
shikonin biosynthesis in cell cultures of L. erythrorhizon
(Yamamoto et al., 2002; Yoshikawa et al., 1986).
Yazaki et al. (1987) reported that inhibitory role of
glutamine for shikonin derivatives production in cell suspen-
sion cultures of L. erythrorhizon was not due to release of
ammonium from glutamine but by glutamine itself.
Role of pH
The pH is an important factor of any biochemical reaction, as
every enzyme/catalyst has its own pH optima and, therefore,
has pronounced effects on the outcome of any biosynthetic
process. In A. euchroma, acidic pH favors cell growth,
whereas alkaline pH exhibited higher shikonin derivatives
production. However, cell growth seems to be hampered at
extremes of pH (Malik et al., 2008; Figure 8). The higher
yield of these pigments at alkaline pH was correlated to
enhanced activity of geranyltransferase, a key regulatory
enzyme, which showed optimum activity at pH 7.1–9.3
(Heide & Tabata, 1987a,b).
Optimization of culture conditions
The biosynthesis and accumulation of secondary compounds
is influenced by the photoperiod, quality and intensity of light
and temperature. Under in vivo conditions, shikonins are
accumulated in underground parts of the plants. The in vitro
cultures of L. erythrorhizon also showed similar behavior, as
pigment formation was inhibited under light conditions
(Yazaki et al., 1997a, 1999).
Heide et al. (1989) observed the reversible effects of light
on shikonin biosynthesis and suggested that cell cultures of
L. erythrorhizon can act as a model to study the light
regulation of shikonin biosynthesis. Tabata et al. (1993)
suggested that light inactivates the flavoprotein necessary for
an enzymatic process leading to shikonin by decomposing the
cofactor flavin mononucleotide (FMN) into lumiflavin. Later,
Yazaki et al. (2001) characterized the dark inducible genes
from L. erythrorhizon (LEDI) and found the regulatory role
of LEDI-II for shikonin biosynthesis. The inhibitory effects
of light on shikonin biosynthesis were also elucidated in
E. lycopsis (Fukui et al., 1983), O. paniculatum (Liu et al.,
2006), O. echiodies (Lattoo, 2005) and A. euchroma (Malik
et al., 2011b; Figure 9). Optimum production of shikonin
derivatives from cell culture of L. erythrorhizon was observed
at 20–25 �C (Tabata & Fujita, 1985). Also, Malik et al.
(2011b) reported maximum pigment production in
A. euchroma at 25 �C (Figure 10).
Two-stage culture medium
Fujita et al. (1981a) found enhanced production of shikonin
derivatives in M-9 medium from L. erythrorhizon cell culture
with poor biomass yield. Therefore, a two-stage culture
process was employed using modified LS medium (MG-5) for
cell growth and modified WH (M-9) for shikonin derivatives
production (Tabata & Fujita, 1985). Also, in cell suspension
cultures of A. euchroma, a two stage culture media approach
was adopted due to non-linked behavior of cell growth and
0
20
40
60
80
100
120
140
160
180
5 5.75 6.5 7.25 8.758 9.5
pH
Cel
l Bio
mas
s (g
, Fw
.L−1
)
0.000
0.050
0.100
0.150
0.200
0.250
0.300
0.350
0.400
0.450
0.500
Pig
men
t Con
tent
(O
D62
0)
Cell Biomass
Pigment
Figure 8. Effect of pH on shikonin derivatives production and average cell biomass yield during cultivation of A. euchroma cell suspension (±SD).Reprinted by permission from Malik et al. (2011b). Copyright (2011) International Federation for Cell Biology.
DOI: 10.3109/07388551.2014.961003 Biotechnological approaches to the production of shikonins 7
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

shikonin derivatives production. Cells were first cultured
for 8 days in a liquid MS + BAP (10.0mM) + IBA (5.0 mM)
medium for cell growth (Growth Medium) and then
transferred to shikonin derivatives or pigment production
medium (APM; Malik et al., 2008).
Addition of precursors and adsorbents
The addition of a precursor, phenylalanine has been reported
to increase the accumulation of shikonins in a cell suspension
culture of L. erythrorhizon (Mizukami et al., 1977). Fukui
et al. (1984) observed that cells of L. erythrorhizon, cultured
in M-9 medium containing activated charcoal, started
producing an orange color benzoquinone derivative, echino-
furan B instead of shikonin. The presence of an adsorbent
PVP (1.0 g/l) in cell suspension culture of A. euchroma was
found to promote cell growth and shikonin derivative
production (Zakhlenjuk et al., 1992).
Use of elicitors/inducers
Elicitation is one of the strategies employed in plant cell
culture to improve the productivity of secondary metabolites
(Namdeo, 2007). Endogenous polysaccharides were found to
induce the biosynthesis of shikonin derivatives in cell cultures
of L. erythrorhizon (Fukui et al., 1990). Ge et al. (2006)
reported the use of rare earth elements in cell cultures of
A. euchroma for enhanced cell growth and shikonin derivative
production. In order to enhance the shikonin content in cell
culture of A. euchroma, fungal elicitors were reported by
Fu & Lu (1999). They obtained 2.24 times higher production
of shikonin derivatives from cells on addition of Rhizopus
oryzae in the medium. Ning et al. (1994) observed the higher
production of shikonin derivatives in cell suspension cultures
of Onosma paniculatum on addition of fungal elicitor. There
are reports of enhanced production of shikonin derivatives in
cell culture of boraginaceous plants by methyl jasmonate.
Enhanced production of shikonin was observed from cells of
L. erythrorhizon (Gaisser & Heide, 1996) and A. euchroma
(Bychkova et al., 1993) and there was an increase in alkannin
pigment, an enantiomer of shikonin from Alkanna tinctoria
cells (Urbanek et al., 1996) in response to methyl jasmonate.
Urmantseva et al. (1999) reported that methyl jasmonate
induced shikonin biosynthesis in cells of A. euchroma by
activation of phenylalanine ammonia-lyase. The elicitation
activity of methyl jasmonate was higher than that of yeast
extract (Mizukami et al., 1992, 1993). Touno et al. (2005)
studied the role of ethylene on shikonin biosynthesis in stem
cultures of L. erythrorhizon.
Permeabilization
Permeabilization of the cell membrane enables the passage of
various enzymes or molecules into and out of the cell. Chung
et al. (2006) studied the effect of gamma-irradiation on
shikonin derivatives production from callus cultures of
L. erythrorhizon. A brief exposure of cells to ultrasound
resulted in a 60–70% increase in the yield of shikonin
derivatives in L. erythrorhizon (Lin & Wu, 2002). It has been
reported that the addition of 1–2% (w/v) Na2EDTA and 1–5%
(v/v) Triton X-100 at the end of growth phase stimulated
the accumulation of shikonin derivatives in cell suspension
cultures of A. euchroma (Zakhlenjuk et al., 1992).
Immobilization
Immobilization can be an effective tool to improve the
effectiveness of a plant cell production process (Dornenburg,
2004). Kim & Chang (1990b) showed 2.5 times higher
production of shikonin from cells of L. erythrorhizon by
immobilization using calcium alginate.
In situ product removal and two-phase culture
Use of a two-phase culture system to improve secondary
metabolites production from different plant species, its types
as well as applications have been described in a recent review
by Malik et al. (2013b). In situ extraction of shikonin in cell
suspension culture of L. erythrorhizon using n-hexadecane as
lipophilic phase was reported by Deno et al. (1987).
Shimomura et al. (1991) and Sim & Chang (1993) success-
fully employed this method in hairy roots of L. erythrorhizon
(Bruce & Daugulis, 1991). The time period for addition
of organic solvent affects the cell growth and secondary
metabolite production. The addition of n-hexadecane to the
0 100 200 300 400 500 600 700
2
4
6
8
10
12
14
Day
s
Shikonin derivatives (µg/g FW)
20 °C 25 °C 30 °C
c
a
b
a
a
a
a
b
b
b
b
bb
cc
cc
cc
Figure 10. Effect of temperature on shikonin derivatives production incell suspension cultures of A. euchroma (± SD). Different letters column(a, b, c) on the bars showing the significant difference (Tukey test,p� 0.05). Reprinted from Malik et al. (2011b) Copyright (2011)International Federation for Cell Biology, with permission from JohnWiley and Sons.
Figure 9. Cell suspension cultures of A. euchroma under (a) light and(b) dark conditions after 8 days of culture. Reprinted from Malik et al.(2011b) Copyright (2011) International Federation for Cell Biology, withpermission from John Wiley and Sons.
8 S. Malik et al. Crit Rev Biotechnol, Early Online: 1–14
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

medium on 12th day in A. euchroma culture yielded 2.49
times higher shikonin derivatives as compared to normal
culture without solvents (Fu & Dewei, 1998). In
A. hispidissima, addition of adsorbents such as liquid paraffin
and charcoal in liquid culture medium stimulated alkannin
production by ca. 3-fold (Singh et al., 2002). Zare et al.
(2010) used liquid paraffin to extract shikonin and alkannin
from cell suspension cultures of Echium italicum. There are
reports on the combined effect of in situ extraction and
immobilization, elicitation or permeabilization for better
production of shikonin derivatives (Fu & Lu, 1999; Kim &
Chang, 1990a; Lin & Wu, 2002; Zhang et al., 2013).
Organ culture
Although plant cell cultures showed great potential to produce
an array of valuable products, limited success has been
achieved at the industrial scale. This is attributed to poor
genetic stability of in vitro cell cultures and reduced
metabolite yield with scalability (Shanks & Morgan, 1999).
The culture of differentiated cells or organs viz., shoot or root
culture offer an advantage as these are more stable and do
not undergo rapid genetic variation as observed in unorgan-
ized cells.
Shoot culture
Touno et al. (2000a) reported the production of shikonin
derivatives in shoot cultures of L. erythrorhizon. Shoot
cultures showed the highest shikonin production in liquid
B5 medium at 25 �C and under dark conditions. On the basis
of microscopic studies of shoot culture, Touno et al. (2000b)
showed the presence of a red pigment on stem surface
and hairs.
Scale up studies
Efforts of scientific groups for several years enabled the
Mitsui Petrochemical Company, Japan to become the first of
its kind in the production of shikonin derivatives on the
commercial scale using plant cell culture technology. The
initial experiments on optimization of nutrient media, culture
conditions and downstream parameters increased the yield of
shikonin derivatives up to 13.6% (DW) using L. erythrorhizon
cell cultures, as compared to 1–2% (DW) in roots of intact
plants (Fujita et al., 1981b; Tabata & Fujita, 1985). Later,
shikonin derivative production has been scaled up success-
fully by Mitsui Chemicals Inc. in stirred airlift type bioreac-
tors having 200 L working volume in the growth stage and
750 L in the production stage with residence times of 9 and 14
days, respectively (Fujita, 1988a, b; Tabata & Fujita, 1985).
The cells in bioreactor produced 23% of shikonin in only 23
days as compared to only 2% in 3–7 years from wild grown
plants. Approximately, 5 kg of shikonin was produced in each
bioreactor run and out of 150 kg of its annual demand, 65 kg
was produced by Mitsui Chemicals (Renneberg, 2008). The
production of shikonin derivatives from hairy roots of L.
erythrorhizon by employing a bubble phase bioreactor was
reported by Sim & Chang (1993).
Experiments on the up-scaling of shikonin derivative
production from cell suspension cultures in A. euchroma
using 7.5 L stirred tank bioreactor were carried out
(Figure 11). Cell suspension cultures in growth medium
were transferred from 250 ml flasks to 500, 1000, 2000, 3000,
5000 flasks and finally to 7.5 L BioFlo 110 Benchtop
Fermentor (New Brunswick Scientific Company Inc.,
Edison, NJ) containing APM for large scale production of
shikonin derivatives.
Dong et al. (1993) studied some characteristics of cell
suspension and fermentation culture in A. euchroma. Process
of progressive scale-up culture in A. euchroma from 250 mL
shake flask to 9 L and 25 L airlift bioreactors has been
established by Chen et al. (1994). The use of periodically
submerged airlift bioreactor systems was investigated in a
two-stage culture system of plant cell culture and enhanced
secondary metabolite production by Ge et al. (2006). The
advances in shikonin derivative production from different
plant species under in vitro conditions are highlighted in
Table 3.
Figure 11. (a) Shikonin derivatives production from cell suspension cultures of A. euchroma in 7.5 L bioreactor and (b) Close up of the same.
DOI: 10.3109/07388551.2014.961003 Biotechnological approaches to the production of shikonins 9
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

Table 3. Chronological studies on in vitro culture of shikonin bearing plant species.
Year References
1974 Shikonin derivatives obtained from callus cultures of Lithospermum erythrorhizon. Tabata et al. (1974)1978 Stable cell lines in L. erythrorhizon with improved growth and pigment production obtained using cell-
aggregate cloning technique .Mizukami et al. (1978)
1981 Effect of different basal media was studied on cell growth and shikonin derivatives production in cellsuspension culture of L. erythrorhizon.
Fujita et al. (1981b)
1981 A new medium developed for the production of shikonin derivatives from cell suspension culture ofL. erythrorhizon.
Fujita et al. (1981a)
1981 Production of shikonin derivatives in callus cultures of Echium lycopsis. Inouye et al. (1981);Fukui et al. (1983)
1983 Analysis of shikonin derivatives in cell suspension cultures of L. erythrorhizon. Fujita et al. (1983)1983 Commercial production of shikonin derivatives achieved from cell culture of L. erythrorhizon. Fujita (1988a)1987 Tissue culture of Arnebia euchroma initiated.1987 Factors affecting the production of shikonin derivatives in callus and cell suspension culture studied
in L. erythrorhizon.Hara et al. (1987)
1989 Reversible effect of light on shikonin biosynthesis in L. erythrorhizon. Heide et al. (1989)1991 Two phase culture system for the extraction of shikonin derivatives from hairy roots of L. erythrorhizon. Shimomura et al. (1991);
Sim & Chang (1993)1993 PFP-resistant cell lines selected in A. euchroma. Zakhlenjuk et al. (1993)1996 Cell lines of A. euchroma selected during different stages of culture by in vitro screening method. Sokha et al. (1996)1996 Important enzymes involved in regulation of shikonin biosynthesis were identified. Gaisser & Heide (1996);
Lange et al. (1998)1997 Effects of elicitor methyl jasmonate were investigated on shikonin biosynthesis in cell culture of
L. erythrorhizon.Yazaki et al. (1997b)
1997 Effect of various auxins and cytokinins on organogenesis and embryogenesis in selected line ofL. erythrorhizon.
Yu et al. (1997)
1999 Synergistic effect of in situ extraction and elicitation on shikonin derivatives production in cellsuspension cultures of A. euchroma.
Fu & Lu (1999)
1999 Isolation of naphthoquinones and pyrrolizidine alkaloids from callus and cell suspension culturesof A. euchroma.
Pietrosiuk et al. (1999)
1999 Arnebins showing antibacterial and antifungal activity were found in cell culture of A. hispidissima. Jain et al. (1999)1999 Studies were carried out to modify the biosynthetic pathway for enhanced production of shikonin
derivatives.Sommer et al. (1999)
2000 Cell suspension culture of L. erythrorhizon were found to produce different groups of compounds. Yamamoto et al. (2000)2001 Regulatory mechanism of light was studied on formation of shikonin derivatives in L. erythrorhizon. Yazaki et al. (2001)2001 Direct regeneration from leaves of A. euchroma. Ji & Wang (2001)2001 High shikonin content in the selected p-fluorophenylalanine resistant callus lines in A. euchroma. Bulgakov et al. (2001)2002 Production of alkannin from hairy roots of A. hispidissima. Singh et al. (2002)2002 Enhancement in production of shikonin derivatives in cell suspension culture of L. erythrorhizon by using
low energy ultrasound.Lin & Wu (2002)
2002 Different shikonin derivatives were observed in cell suspension cultures of L. erythrorhizon. Yamamoto et al. (2002)2005 Direct regeneration from cotyledons and hypocotyls in A. euchroma. Jiang et al. (2005)2005 Shoot regeneration and somatic embryogenesis from leaf derived callus in A. euchroma. Manjkhola et al. (2005)2008 Role of pH on shikonin derivatives production in cell suspension culture. Malik et al. (2008)2008 Presence of acetylshikonin and b-acetoxyisovaleryl shikonin in cell suspension cultures of A. euchroma. Sharma et al. (2008)2009 Shikonin formation in cell suspension culture of Onosma paniculatum was reported to be regulated by
Nitric oxide.Wu et al. (2009)
2010 In vitro induction of hairy root culture and shikonin production in A. hispidissima. Chaudhury & Pal (2010)2010 Direct plant regeneration, micropropagation and shikonin induction in A. hispidissima. Pal & Chaudhury (2010)2010 The importance of pre-culture time and concentration of TDZ for direct regeneration from in vitro leaves
of A. euchroma.Malik et al. (2010b)
2010 Pathway for shikonins biosynthesis was identified. Singh et al. (2010)2010 A two-phase liquid culture system using liquid paraffin was established to elicit shikonin and alkannin
derivatives in E. italicum.Zare et al. (2010)
2011 Different physico-chemical factors affecting the shikonin derivatives production in cell suspensioncultures of A. euchroma were studied.
Malik et al. (2011b)
2011 Micropropagation and establishment of callus and cell suspension culture of A. hispidissima foroptimization of alkannin production.
Shekhawat & Shekhawat(2011)
2011 Shikonin production potential of E. italicum callus. Zare et al. (2011)2011 LeERF-1, a novel AP2/ERF family gene within the B3 subcluster was down-regulated by different
light conditions in L. erythrorhizon.Zhang et al. (2011)
2012 Enhanced production of shikonin derivatives in hairy roots of L. canescens. Sykłowska-Baranek et al.(2012)
2013 In vitro propagation and genetic fidelity of micropropagated plants was assessed for A. hispidissima. Phulwaria et al. (2013)2013 The effects of fungal elicitor and macroporous adsorption resin were studied on shikonin accumulation in
hairy roots of A. euchroma.Zhang et al. (2013)
2013 Shikonin production from cell suspension culture of Arnebia sp. and it’s up-scaling through bioreactor. Gupta et al. (2013)
10 S. Malik et al. Crit Rev Biotechnol, Early Online: 1–14
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

Genetic transformation and transformed plants
A low yield of secondary compounds from plant tissues under
in vitro conditions is a major bottleneck in their production at
the commercial level (Charlwood & Pletsch, 2002). Through
genetic engineering, it is possible to manipulate the regulatory
steps of the biosynthetic pathway in order to increase the yield
of required compounds (Verpoorte et al., 1999). There are
reports on the transformation of shikonins bearing plants with
Agrobacterium rhizogenes to enhance the secondary metab-
olite content (Fukui et al., 1998; Shimomura et al., 1991;
Yazaki et al., 1998). Yazaki et al. (1998) obtained transfor-
mants in L. erythrorhizon using A. rhizogenes strain 15 834
with stable and higher production of shikonin derivatives.
Shimomura et al. (1991) transformed the shoot cultures of L.
erythrorhizon with A. rhizogenes strain 15 834 and found a red
pigment after 2–3 weeks culturing on agar gelled and liquid
root culture media. Similarly in L. canescens, hairy roots were
shown to produce ca. a 10% higher content of acetylshikonin
and isobutrylshikonin as compared to natural roots (Pietrosiuk
et al., 2006). Hairy root transformation for other genera of
the Boraginaceae family has also been reported. In another
report by Singh et al. (2002), alkannin was obtained on a
half-strength agar solidified MS medium from hairy roots
of A. hispidissima induced with A. rhizogenes strain 15 834.
Sommer et al. (1999) modified the biosynthetic pathway
to 4HB in hairy root cultures of L. erythrorhizon by
introducing bacterial gene ubiC but it did not show any
significant increase in yield of shikonin derivatives in
transformed plants. Boehm et al. (2000) made an attempt to
manipulate the biosynthetic pathway of secondary metabol-
ism by the introduction of the bacterial gene ubiA in hairy
root cultures of L. erythrorhizon. The transformed hairy roots
showed high ubiA activities, however, ubiA overexpression
was not found sufficient to increase the shikonin content. In A.
euchroma, callus cultures were transformed with
pCAMBIA1302 containing HMGR cDNA (unpublished
data). The cell suspension cultures, raised from transformed
callus, did not show significant alteration in the shikonin
derivative content as compared to the control. Despite high
activity of HMGR, the shikonin content remained unchanged.
A high activity of HMGR in L. erythrorhizon hairy root
cultures was reported by transformation with A. rhizogenes
containing HMGR and ubiC genes under the control of
(octopine synthase)3 mannopine synthase promoter (Kohle
et al., 2002). In spite of high activity using the soluble
cytosolic domain of HMGR in cultures, the shikonin content
remained unchanged. Based on this, it was suggested that the
overexpression of CPL or HMGR was insufficient to increase
shikonin formation. Therefore, it seems that either simultan-
eous overexpression or down regulation of different genes
may influence the biosynthesis of shikonin derivatives.
Conclusions and future prospects
Considering the consumer driven market demand for natural
products as well as commercial and societal importance of
and shikonin derivatives, noticeable efforts on in vitro
production R&D activities are evident from the recent
literature. Biotechnological interventions not only help in
the conservation of biodiversity of natural habitats by
reducing the demand for raw materials from the wild, but
will also lay a foundation towards development of alternative
strategies to fulfill industrial demand. Production of second-
ary compounds using various biotechnological means could
offer an important strategy, but so far have provided with only
limited commercial success. More detailed studies are
required to understand the complex biosynthetic pathways
in order to obtain the desired product. The technology could
be utilized for studies related to metabolic engineering in
shikonins bearing plants to suppress or overexpress the genes/
enzymes involved in the pathway for enhanced production of
secondary compounds.
Acknowledgements
The authors are grateful to the Council of Scientific and
Industrial Research (CSIR), India for providing financial
support. SM would also like to acknowledge the grant from
Centre of the Region Hana for Biotechnological and
Agricultural Research, Czech Republic (CZ.1.05/2.1.00/
01.0007 (SPP 813103061/15)).
Declaration of interest
All the authors have read the manuscript and there are no
conflicts of interest.
References
Andujar I, Rıos JL, Giner RM, Recio MC. (2013). Pharmacologicalproperties of shikonin – a review of literature since 2002. Planta Med,79, 1685–97.
Babula P, Adam V, Havel L, Kizek R. (2009). Noteworthy secondarymetabolites naphthoquinones – their occurrence, pharmacologicalproperties and analysis. Curr Pharm Anal, 5, 47–68.
Boehm R, Sommer S, Li SM, Heide L. (2000). Genetic engineering onshikonin biosynthesis: expression of the bacterial ubiA gene inLithospermum erythrorhizon. Plant Cell Physiol, 41, 911–19.
Bruce LJ, Daugulis AJ. (1991). Solvent selection strategies for extractivebiocatalysis. Biotechnol Prog, 7, 116–24.
Bulgakov VP, Kozyrenko MM, Fedoreyev SA, et al. (2001). Shikoninproduction by p-fluorophenylalanine resistant cells of Lithospermumerythrorhizon. Fitoterapia, 72, 394–401.
Bychkova TP, Nanenina EB, Berzin VB. Miroshnikov AI. (1993).Influence of jasmonic acid and 12-oxophytodienoic acid on thebiosynthesis of shikonin in a cell culture of Arnebia euchroma.Bioorganicheskaya Khimiya, 19, 1008–12.
Chaprin N, Ellis BE. (1984). Microspectrophotometric evaluation ofrosmarinic acid accumulation in single cultured plant cells. Can J Bot,62, 2278–82.
Charlwood BV, Pletsch M. (2002). Manipulation of natural productaccumulation in plants through genetic engineering. J Herbs SpicesMed Plants, 9, 139–51.
Chaudhury A, Pal M. (2010). Induction of Shikonin production in hairyroot cultures of Arnebia hispidissima via Agrobacterium rhizogenes-mediated genetic transformation. J Crop Sci Biotechnol, 13, 99–106.
Chen SY, Hou SS, Zhang J, et al. (1994). Progressive scale-up culture ofArnebia euchroma (Royle) Johnston cells. J Plant Resources Environ,3, 27–31.
Chung BY, Lee YB, Baek MH, et al. (2006). Effects of low-dose gamma-irradiation on production of shikonin derivatives in callus cultures ofLithospermum erythrorhizon S. Radiat Phys Chem, 75, 1018–23.
Damianakos H, Kretschmer N, Sykłowska-Baranek K, et al. (2012).Antimicrobial and cytotoxic isohexenylnaphthazarins from Arnebiaeuchroma (Royle) Jonst. (Boraginaceae) callus and cell suspensionculture. Molecules, 17, 14310–22.
Deno H, Suga C, Morimoto T, Fujita Y. (1987). Production of shikoninderivatives by cell suspension cultures of Lithospermum erythrorhi-zon: VI. Production of shikonin derivatives by a two-layer culturecontaining an organic solvent. Plant Cell Rep, 6, 197–9.
DOI: 10.3109/07388551.2014.961003 Biotechnological approaches to the production of shikonins 11
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

DiCosmo F, Misawa M. (1996). Plant cell culture secondary metabolism:toward industrial application. New York: CRC Press, 229 p.
Dix P. (1986). Cell line selection. In: Yeoman MM, ed. Plant cell culturetechnology. Oxford, London: Blackwell Scientific Publications,143–201.
Dong JW, Ye HC, Wu X, et al. (1993). Studies on cell suspensionculture and fermentation culture of Arnebia euchroma. Acta Bot Sin, 35,57–61.
Dornenburg H, Knorr D. (1995). Strategies for the improvement ofsecondary metabolite production in plant cell cultures. EnzymeMicrob Technol, 17, 674–84.
Dornenburg H. (2004). Evaluation of immobilisation effects on meta-bolic activities and productivity in plant cell processes. ProcessBiochem, 39, 1369–75.
Fu X, Dewei L. (1998). Enhancement of shikonin production in cellsuspension cultures of Arnebia euchroma employing two-liquid-phasesystems. Chin J Chem Eng, 6, 1–5.
Fu XQ, Lu DW. (1999). Stimulation of shikonin production by combinedfungal elicitation and in situ extraction in suspension cultures ofArnebia euchroma. Enzyme Microb Technol, 24, 243–6.
Fujita Y, Hara Y, Ogino T, Suga C. (1981a). Production of shikoninderivatives by cell suspension cultures of Lithospermum erythrorhi-zon. I. Effects of nitrogen source on the production of shikoninderivatives. Plant Cell Rep, 1, 59–60.
Fujita Y, Hara Y, Suga C, Morimoto T. (1981b). Production of shikoninderivatives by cell suspension cultures of Lithospermum erythrorhizonII. A new medium for the production of shikonin derivates. Plant CellRep, 1, 613.
Fujita Y, Maeda Y, Suga C, Morimoto T. (1983). Production of shikoninderivatives by cell suspension cultures of Lithospermum erythrorhizonIII. Comparison of shikonin derivatives of cultured cells andKo-shikon. Plant Cell Rep, 2, 192–3.
Fujita Y. (1988a). Industrial production of shikonin and berberine. In:Bock G, Marsh J, eds. Applications of plant cell and tissue culture.Chichester, UK: John Wiley & Sons Ltd., 228–35.
Fujita Y. (1988b). Shikonin production by plant (Lithospermumerythrorhizon) cell cultures. In: Bajaj YPS, ed. Biotechnology inagriculture and forestry, Vol 4. Medicinal and aromatic plants. Berlin,Heidelberg: Springer-Verlag, 225 p.
Fukui H, Tsukada M, Mizukami H, Tabata M. (1983). Formation ofstereoisomeric mixtures of naphthoquinone derivatives in Echiumlycopsis callus cultures. Phytochemistry, 22, 453–6.
Fukui H, Yazaki K, Tabata M. (1984). Two phenolic acids fromLithospermum erythrorhizon cell suspension cultures. Phytochemistry,23, 2398–9.
Fukui H, Tani M, Tabata M. (1990). Induction of shikonin biosynthesisby endogenous polysaccharides in Lithospermum erythrorhizon cellsuspension cultures. Plant Cell Rep, 9, 73–6.
Fukui H, Feroz Hasan AFM, Ueoka T, Kyo M. (1998). Formation andsecretion of a new brown benzoquinone by hairy root cultures ofLithospermum erythrorhizon. Phytochemistry, 47, 1037–9.
Gaisser S, Heide L. (1996). Inhibition and regulation of shikoninbiosynthesis in suspension cultures of Lithospermum. Phytochemistry,41, 1065–72.
Ge F, Yuan X, Wang X, et al. (2006). Cell growth and shikoninproduction of Arnebia euchroma in a periodically submerged airliftbioreactor. Biotechnol Lett, 28, 525–9.
Gupta K, Garg S, Singh J, Kumar M. (2013). Enhanced production ofnapthoquinone metabolite (shikonin) from cell suspension culture ofArnebia sp. and its up-scaling through bioreactor. 3 Biotech, 4, 263–73.
Hall RD, Yeoman MM. (1986). Temporal and spatial heterogeneity inthe accumulation of anthocyanins in cell culture of Catharanthusroseus (L.) G. Don. J Exp Bot, 37, 48–60.
Hara Y, Morimoto T, Fujita Y. (1987). Production of shikonin derivativesby cell suspension cultures of Lithospermum erythrorhizon V.Differences in the production between callus and suspension cultures.Plant Cell Rep, 6, 8–11.
Harborne JR. (1993). Introduction to ecological biochemistry. 4th ed.London: Elsevier.
Heide L, Tabata M. (1987a). Enzyme activities in cell-free extractsof shikonin-producing Lithospermum erythrorhizon cell suspensioncultures. Phytochemistry, 26, 1645–50.
Heide L, Tabata M. (1987b). Geranylpyrophosphate: p-hydroxybenzoategeranyltransferase activity in extracts of Lithospermum erythrorhizoncell cultures. Phytochemistry, 26, 1651–5.
Heide L, Nishioka N, Fukui H, Tabata M. (1989). Enzymatic regulationof shikonin biosynthesis in Lithospermum erythrorhizon cell cultures.Phytochemistry, 28, 1873–77.
Inouye H, Matsumura H, Kawasaki M, et al. (1981). Two quinines fromcallus cultures of Echium lycopsis. Phytochemistry, 20, 1701–5.
Jain SC, Singh B, Jain R. (1999). Arnebins and antimicrobial activitiesof Arnebia hispidissima (Lehm.) DC cell cultures. Phytomedicine,6, 474–6.
Ji QL, Wang WG. (2001). Asexual propagation of Arnebia euchromaand exploration of hereditary stability in regenerated plantlets. PlantPhysiol Commun, 37, 499–502.
Jiang B, Yang YG, Guo YM, Chen YZ. (2005). Thidiazuron-inducedin vitro shoot organogenesis of the medicinal plant Arnebia euchroma(Royle) Johnst. In Vitro Cell Dev Biol Plant, 41, 677–81.
Kim DJ, Chang HN. (1990a). Effect of growth hormone modifications onshikonin production from Lithospermum erythrorhizon cell cultureswith in situ extraction. Biotechnol Lett, 12, 289–94.
Kim DJ, Chang HN. (1990b). Enhanced shikonin production fromLithospermum erythrorhizon by in situ extraction and calcium alginateimmobilization. Biotechnol Bioeng, 36, 460–6.
Kohle A, Sommer S, Yazaki K, et al. (2002). High level expression ofchorismate pyruvate-lyase (ubiC) and HMG-CoA reductase in hairyroot cultures of Lithospermum erythrorhizon. Plant Cell Physiol, 43,894–902.
Kumar R, Sharma N, Malik S, et al. (2011). Cell suspension cultureof Arnebia euchroma (Royle) Johnston – a potential source ofnaphthoquinone pigments. J Med Plants Res, 5, 6048–54.
Lange BM, Severin K, Bechthold A, Heide L. (1998). Regulatory roleof microsomal 3-hydroxy-3-methylglutaryl-coenzymeA reductase forshikonin biosynthesis in Lithospermum erythrorhizon cell suspensioncultures. Planta, 204, 234–41.
Lattoo S. (2005). In vitro approaches for the production of shikonin fromOnosma echioides var. hispidum. PhD thesis. Jammu: University ofJammu.
Li SM, Henning S, Heide L. (1998). Shikonin: a geranyl diphosphate-derived plant hemiterpenoid formed via the mevalonate pathway.Tetrahedron Lett, 39, 2721–4.
Lin L, Wu J. (2002). Enhancement of shikonin production in single andtwo-phase suspension cultures of Lithospermum erythorhizon cellsusing low-energy ultrasound. Biotechnol Bioeng, 78, 81–8.
Linsmaier EM, Skoog F. (1965). Organic growth factor requirements oftobacco tissue cultures. Physiol Plant, 18, 100–27.
Liu CZ, Qi JL, Chen L, et al. (2006). Effect of light on gene expressionand shikonin formation in cultured Onosma paniculatum cells. PlantCell Tissue Organ Cult, 84, 39–46.
Lu JJ, Bao JL, Wu GS, et al. (2013). Quinones derived from plantsecondary metabolites as anti-cancer agents. Anticancer Agents MedChem, 13,456–63.
Lubbe A, Verpoorte R. (2011). Cultivation of medicinal and aro-matic plants for specialty industrial materials. Ind Crops Prod, 34,785–801.
Malik S, Bhushan S, Verma SC, et al. (2008). Production ofnaphthoquinone pigments in cell suspension cultures of Arnebiaeuchroma (Royle) Johnston: influence of pH on growth kinetics andacetylshikonin. Med Aromat Plant Sci Biotechnol, 2, 43–9.
Malik S. (2009). In vitro cultures of Arnebia euchroma (Royle) Johnst.and Rheum emodi Wall. for secondary metabolite production. PhDthesis. Amritsar, India: Guru Nanak Dev University.
Malik S, Kumar R, Vats SK, et al. (2009). Regeneration in Rheum emodiWall.: a step towards conservation of an endangered medicinal plantspecies. Eng Life Sci, 9, 130–4.
Malik S, Sharma N, Sharma UK, et al. (2010a). Qualitative andquantitative analysis of anthraquinone derivatives in rhizomes oftissue culture-raised Rheum emodi Wall. plants. J Plant Physiol, 167,749–56.
Malik S, Sharma S, Sharma M, Ahuja PS. (2010b). Direct shootregeneration from intact leaves of Arnebia euchroma (Royle) Johnstonusing thidiazuron. Cell Biol Int, 34, 537–42.
Malik S, Cusido RM, Mirjalili MH, et al. (2011a). Production of theanticancer drug taxol in Taxus baccata suspension cultures: a review.Process Biochem, 46, 23–34.
Malik S, Bhushan S, Sharma M, Ahuja PS. (2011b). Physico-chemicalfactors influencing the shikonin derivatives production in cellsuspension cultures of Arnebia euchroma (Royle) Johnston, amedicinally important plant species Cell Biol Int, 35, 152–7.
12 S. Malik et al. Crit Rev Biotechnol, Early Online: 1–14
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

Malik S, Andrade SAL, Sawaya ACHF, et al. (2013a). Root-zonetemperature alters alkaloid synthesis and accumulation inCatharanthus roseus and Nicotiana tabacum. Ind Crops Prod, 49,318–25.
Malik S, Mirjalili MH, Fett-Neto AG, et al. (2013b). Living between twoworlds: two-phase culture systems for producing plant secondarymetabolites. Crit Rev Biotechnol, 33, 1–22.
Malik S, Biba O, Gruz J, et al. (2014). Biotechnological approaches forproducing aryltetralin lignans from Linum species. Phytochem Rev.doi: 10.1007/s11101-014-9345-5.
Manjkhola S, Dhar U, Joshi M. (2005). Organogenesis, embryogenesis,and synthetic seed production in Arnebia euchroma – a criticallyendangered medicinal plant of the Himalaya. In Vitro Cell Dev BiolPlant, 41, 244–8.
Mizukami H, Konoshima M, Tabata M. (1977). Effect of nutritionalfactors on shikonin derivative formation in Lithospermum calluscultures. Phytochemistry, 16, 1183–6.
Mizukami H, Konoshima M, Tabata M. (1978). Variation in pigmentproduction in Lithospermum erythrorhizon callus cultures.Phytochemistry, 17, 95–7.
Mizukami H, Ogawa T, Ohashi H, Ellis BE. (1992). Induction ofrosmarinic acid biosynthesis in Lithospermum erythrorhizon cellsuspension cultures by yeast extract. Plant Cell Rep, 11, 480–3.
Mizukami H, Tabira Y, Ohashi H, Ellis BE. (1993). Methyl jasmonate-induced rosmarinic acid biosynthesis in Lithospermum erythrorhizoncell suspension cultures. Plant Cell Rep, 12, 706–9.
Murashige T, Skoog F. (1962). A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol Plant, 15,473–97.
Namdeo AG. (2007). Plant cell elicitation for production of secondarymetabolites: a review. Pharmacogn Rev, 1, 69–79.
Ning W, Zhao Q, Xia Z, Cao R. (1994). Effects of fungal elicitor onshikonin derivatives formation in Onosma paniculatum cell cultures.Acta Phytophysiol Sinica, 20, 325–31.
Nosov AM. (2012). Application of cell technologies for production ofplant derived bioactive substances of plant origin. Appl BiochemMicrobiol, 48, 609–24.
Pal M, Chaudhury A. (2010). High frequency direct plant regeneration,micropropagation and shikonin induction in Arnebia hispidissima.J Crop Sci Biotech, 13, 13–20.
Phulwaria M, Rai M, Shekhawat NS. (2013). An improved micropropa-gation of Arnebia hispidissima (Lehm.) DC and assessment of geneticfidelity of micropropagated plants using DNA-based molecularmarkers. Appl Biochem Biotechnol, 170, 1163–73.
Pietrosiuk A, Urmantseva V, Furmanowa M. (1999). Some naphthoqui-nones and pyrolizidine alkaloids in cell culture of Arnebia euchroma(Royle) Jonst. Herba Polonica, 65, 354–61.
Pietrosiuk A, Syklowska-Baranek K, Wiedenfeld H, et al. (2006). Theshikonin derivatives and pyrrolizidine alkaloids in hairy root culturesof Lithospermum canescens (Michx.) Lehm. Plant Cell Rep, 25,1052–8.
Rao RS, Ravishankar GA. (2002). Plant cell cultures: chemical factoriesof secondary metabolites. Biotechnol Adv, 20, 101–53.
Raskin I, Ribnicky DM, Komarnytsky S, et al. (2002). Plants andhuman health in the twenty-first century. Trends Biotechnol, 20,522–31.
Ravishankar GA, Rao RS. (2002). Biotechnological production of phyto-pharmaceuticals. J Biochem Mol Biol Biophys, 4, 73–102.
Renneberg R. (2008). Green biotechnology. In: Demain AL, ed.Biotechnology for beginners. New York: Academic Press, 210–3.
Shanks JV, Morgan J. (1999). Plant ‘hairy root’ culture. Curr OpinBiotechnol, 10, 151–5.
Sharma N, Sharma UK, Malik S, et al. (2008). Isolation and purificationof acetylshikonin and b-acetoxyisovalerylshikonin from cell suspen-sion cultures of Arnebia euchroma (Royle) Johnston using rapidpreparative HPLC. J Sep Sci, 31, 629–35.
Shekhawat MS, Shekhawat NS. (2011). Micropropagation of Arnebiahispidissima (Lehm).DC. and production of alkannin from callus andcell suspension culture. Acta Physiol Plant, 33, 1445–50.
Shimomura K, Sudo H, Saga H, Kamada H. (1991). Shikonin productionand secretion by hairy root cultures of Lithospermum erythrorhizon.Plant Cell Rep, 10, 282–5.
Sim SJ, Chang HN. (1993). Increased shikonin production by hairy rootsof Lithospermum erythrorhizon in two phase bubble column reactor.Biotechnol Lett, 15, 145–50.
Singh B, Sahu PM, Sharma MK, et al. (2002). Production and secretionof alkannin by hairy root cultures of Arnebia hispidissima (Lehm.)D.C. J Plant Biol, 29, 293–300.
Singh RS, Gara RK, Bhardwaj PK, et al. (2010). Expression of3-hydroxy-3-methylglutaryl-CoA reductase, p-hydroxybenzoate-m-geranyltransferase and genes of phenylpropanoid pathway exhibitspositive correlation with shikonins content in arnebia [Arnebiaeuchroma (Royle) Johnston]. BMC Mol Biol, 11, 88.
Sokha V, Nikolaeva L, Pank F. (1996). The shikonin production inplant tissue culture: selection of cell lines with high productivity.Proceedings of the International symposium on Breeding Research onMedicinal and Aromatic Plants, Germany, 2, 322–6.
Sommer S, Kohle A, Yazaki K, et al. (1999). Genetic engineering ofshikonin biosynthesis hairy root cultures of Lithospermum erythror-hizon transformed with the bacterial ubiC gene. Plant Mol Biol, 39,683–93.
Srinivasan V, Ryu DDY. (1992). Enzyme activity and shikoninproduction in Lithospermum erythrorhizon cell cultures. BiotechnolBioeng, 40, 69–74.
Sykłowska-Baranek K, Pietrosiuk A, Gawron A, et al. (2012). Enhancedproduction of antitumour naphthoquinones in transgenic hairy rootlines of Lithospermum canescens. Plant Cell Tissue Organ Cult, 108,213–9.
Tabata M, Mizukami H, Hiraoka N. (1974). Pigment formation in callusculture of Lithospermum erythrorhizon. Phytochemistry, 13, 927–32.
Tabata M, Fujita Y. (1985). Production of shikonin by plant cell cultures.In: Zaitlin M, Day P, Hollaender A, eds. Biotechnology in plantscience: relevance to agriculture in the eighties. New York: AcademicPress, Inc. Harcourt Brace Jovanovich Publishers, 207–18.
Tabata M, Yazaki K, Nishikawa Y, Yoneda F. (1993). Inhibitionof shikonin biosynthesis by photodegradation products of FMN.Phytochemistry, 32, 1439–42.
Terada A, Tanoue Y, Hatada A, Sakamoto H. (1983). Total synthesis ofshikalkin [(±)-shikonin]. J Chem Soc Chem Commun, 18, 987.
Touno K, Harada K, Yoshimatsu K, et al. (2000a). Shikonin deriva-tive formation on the stem of cultured shoots in Lithospermumerythrorhizon. Plant Cell Rep, 19, 1121–6.
Touno K, Harada K, Yoshimatsu K, et al. (2000b). Histologicalobservation of red pigment formed on shoot stem of Lithospermumerythrorhizon. Plant Biotechnol, 17, 127–30.
Touno K, Tamaoka J, Ohashi Y, Shimomura K. (2005). Ethylene inducedshikonin biosynthesis in shoot culture of Lithospermum erythrorhizon.Plant Physiol Biochem, 43, 101–5.
Urbanek H, Bergier K, Saniewski M, Patykowski J. (1996). Effect ofjasmonates and exogenous polysaccharides on production of alkanninpigments in suspension cultures of Alkanna tinctoria. Plant Cell Rep,15, 637–41.
Urmantseva VV, Karyagina TB, Chertkova RV, et al. (1999). Inductionof phenylalanine ammonia-lyase by methyl jasmonate in cultured cellsof Arnebia euchroma. Russ J Plant Physiol, 46, 749.
Verpoorte R, ten Hoopen HJG, Memelink J. (1999). Metabolicengineering of plant secondary metabolite pathways for the produc-tion of fine chemicals. Biotechnol Lett, 21, 467–79.
Verpoorte R. (2000). Secondary metabolism. In: Verpoorte R,Alfermann AW, eds. Metabolic engineering of plant secondarymetabolism. London: Kluwer Academic Publishers, 1–29.
White PR. (1954). The cultivation of animal and plant cells. New York:The Ronald press Co.
Wu SJ, Qi JL, Zhang WJ, et al. (2009). Nitric oxide regulates shikoninformation in suspension-cultured Onosma paniculatum cells. PlantCell Physiol, 50, 118–28.
Yamamoto H, Inouye K, Li SM, Heide L. (2000). Geranylhydroquinone 3-hydroxylase, a cytochrome P-450 monooxygenase from Lithospermumerythrorhizon cell suspension cultures. Planta, 210, 312–7.
Yamamoto H, Zhao P, Yazaki K, Inoue K. (2002). Regulation of litho-spermic acid B and shikonin production in Lithospermum erythrorhi-zon cell suspension cultures. Chem Pharm Bull, 50, 1086–90.
Yang YH, Zhang H, Cao RQ. (1999). Effect of brassinolide on growthand shikonin formation in cultured Onosma paniculatum cells. J PlantGrowth Regul, 18, 89–92.
Yazaki K, Fukui H, Kikuma M, Tabata M. (1987). Ruglation of shikoninproduction by glutamine in Lithospermum erythrorhizon cell cultures.Plant Cell Rep, 6, 131–4.
Yazaki K, Matsuoka H, Sato F. (1997a). cDNA cloning and functionalanalysis of LEDI-2, a gene preferentially expressed in the dark in
DOI: 10.3109/07388551.2014.961003 Biotechnological approaches to the production of shikonins 13
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.

Lithospermum erythrorhizon cell suspension cultures (abstractno.1348). Plant Physiol, 114, S–261.
Yazaki K, Takeda K, Tabata M. (1997b). Effects of methyl jasmonateon shikonin and dihydroechinofuran production in Lithospermumerythrorhizon cell cultures. Plant Cell Physiol, 38, 776–82.
Yazaki K, Tanaka S, Matsuoka H, Sato F. (1998). Stable transform-ation of Lithospermum erythrorhizon by Agrobacterium rhizogenesand shikonin production of the transformants. Plant Cell Rep, 18,214–9.
Yazaki K, Matsuoka H, Ujihara T, Sato F. (1999). Shikonin biosynthesisin Lithospermum erythrorhizon: light-induced negative regulation ofsecondary metabolism. Plant Biotechnol, 16, 335–42.
Yazaki K, Matsuoka H, Shimomura K, et al. (2001). A novel dark-inducible protein LeDI-2 and its involvement in root specificsecondary metabolism in Lithospermum erythrorhizon. PlantPhysiol, 125, 1831–41.
Yoshikawa N, Fukui H, Tabata M. (1986). Effect of gibberellin A onshikonin production in Lithospermum callus cultures. Phytochemistry,25, 621–2.
Yu HJ, Oh SK, Oh MH, et al. (1997). Plant regeneration from calluscultures of Lithospermum erythrorhizon. Plant Cell Rep, 16, 261–6.
Zakhlenjuk OV, Vidmachenko TV, Kunakh VA. (1992). Effect of someabiotic elicitors on the shikonin accumulation in macrotomia suspen-sion culture. Abstr 3rd Ukr Confr Med Bot Kiev, 62 p.
Zakhlenjuk OV, Vidmachenko TV, Kunakh VA, et al. (1993).Comparative characteristics of shikonin accumulating Arnebiaeuchroma suspension culture and p-fluorophenylalanine-resistantvariant. Acta Horticult, 330, 293–8.
Zare K, Nazemiyeh H, Movafeghi A, et al. (2010). Bioprocessengineering of Echium italicum L.: induction of shikonin and alkanninderivatives by two-liquid-phase suspension cultures. Plant Cell TissueOrgan Cult, 100, 157–64.
Zare K, Khosrowshahli M, Nazemiyeh H, et al. (2011). Callus cultureof Echium italicum L. towards production of a shikonin derivative. NatProd Res, 25, 1480–7.
Zhang W, Zou A, Miao J, et al. (2011). LeERF-1, a novel AP2/ERFfamily gene within the B3 subcluster, is down-regulated by lightsignals in Lithospermum erythrorhizon. Plant Biol, 13, 343–8.
Zhang P, Wang F, Zhu C. (2013). Influence of fungal elicitorand macroporous resin on shikonin accumulation in hairy rootsof Arnebia euchroma (Royle) Johnst. Chinese J Biotechnol, 29,214–23.
14 S. Malik et al. Crit Rev Biotechnol, Early Online: 1–14
Cri
tical
Rev
iew
s in
Bio
tech
nolo
gy D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
NIC
AM
P on
10/
30/1
4Fo
r pe
rson
al u
se o
nly.
Related Documents











