This article was downloaded by: [Hamed Ameri] On: 29 June 2015, At: 23:48 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Click for updates Historical Biology: An International Journal of Paleobiology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/ghbi20 Biostratigraphy of the Peri-Gondwana Cambrian trilobite fauna (northern Kerman, Iran) and correlation with other countries Hamed Ameri a & Fatane Zamani b a Department of Ecology, Institute of Science, High Technology and Environmental Sciences, Graduate University of Advanced Technology , Kerman, Iran b Department of Geology, Faculty of Sciences, University of Isfahan , Isfahan, Iran Published online: 29 Jun 2015. To cite this article: Hamed Ameri & Fatane Zamani (2015): Biostratigraphy of the Peri-Gondwana Cambrian trilobite fauna (northern Kerman, Iran) and correlation with other countries, Historical Biology: An International Journal of Paleobiology, DOI: 10.1080/08912963.2014.927870 To link to this article: http://dx.doi.org/10.1080/08912963.2014.927870 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by: [Hamed Ameri]On: 29 June 2015, At: 23:48Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Click for updates
Historical Biology: An International Journal ofPaleobiologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ghbi20
Biostratigraphy of the Peri-Gondwana Cambriantrilobite fauna (northern Kerman, Iran) and correlationwith other countriesHamed Ameria & Fatane Zamaniba Department of Ecology, Institute of Science, High Technology and Environmental Sciences,Graduate University of Advanced Technology , Kerman, Iranb Department of Geology, Faculty of Sciences, University of Isfahan , Isfahan, IranPublished online: 29 Jun 2015.
To cite this article: Hamed Ameri & Fatane Zamani (2015): Biostratigraphy of the Peri-Gondwana Cambrian trilobite fauna(northern Kerman, Iran) and correlation with other countries, Historical Biology: An International Journal of Paleobiology,DOI: 10.1080/08912963.2014.927870
To link to this article: http://dx.doi.org/10.1080/08912963.2014.927870
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Biostratigraphy of the Peri-Gondwana Cambrian trilobite fauna (northern Kerman, Iran) andcorrelation with other countries
Hamed Ameria* and Fatane Zamanib
aDepartment of Ecology, Institute of Science, High Technology and Environmental Sciences, Graduate University of AdvancedTechnology, Kerman, Iran; bDepartment of Geology, Faculty of Sciences, University of Isfahan, Isfahan, Iran
(Received 3 February 2014; accepted 21 May 2014)
Polymeroid trilobite fauna from two stratigraphic sections (Godbondar and Kuhbanan) of the Kuhbanan Formation innorthern Kerman (central Iran) were studied and subjected to biostratigraphic analysis. Eleven genera and species arerecognised from the latest Early Cambrian and Middle Cambrian Peri-Gondwanian successions of the study sections. Therecognised fauna includes Afghanocare lategenatum, Blountia blountia, Iranoleesia sp., Iranoleesia pisiformis, Kermanellakuhbananensis, Kermanella lata lata, Kermanella lata minuta, Kermanella sp., Redlichia chinensis, Redlichia noetlingi andRedlichia sp. Based on trilobite distribution, three trilobite biozones were recognised in the study sections, namely Redlichianoetlingi biozone, Kermanella kuhbananensis biozone and Iranoleesia pisiformis biozone. The age of the study sections islate Early Cambrian to late Middle Cambrian based on the recognised trilobite biozones. The recognised late EarlyCambrian trilobite assemblages (especially Redlichia and Kermanella) from northern Kerman are similar to those foundfrom some other parts of Gondwana or Peri-Gondwana terrains (north India, Pakistan, northwestern Kashmir, Tajikistan,South Australia, South China and Afghanistan) and show affinities with fauna found in some other parts of Iran (Alborz,northern Iran; Tabas, eastern Iran; southeast Karman).
Keywords: polymeroid trilobite; Cambrian; Kuhbanan Formation; Peri-Gondwana; central Iran
1. Introduction
Cambrian deposits are distributedwidely in different basins
of Iran. Each basin has distinct lithostratigraphic and
biostratigraphic characteristics and fossil contents. The
CambrianSystem in theAlborz andZagros basins (northern
and southeastern–northwestern Iran, respectively) consists
of upper parts of the Soltanieh Formation and Barut,
Zaigun, Lalun andMila Formations. The Cambrian System
in central Iran cropped out in two different sub-basins:
Kerman (southeastern Iran) and Tabas (eastern Iran). The
Cambrian deposits of northern Kerman sub-basin comprise
two series and three formations from base to top; they
include the Rizu and Dezu Series and Dahu, Kuhbanan and
Unnamed Formations. The above-mentioned deposits have
a thickness of 1000–2700m and consist of evaporite,
igneous and siliciclastic–carbonate rocks. (Almost all of
the deposits outcropped along the Kuhbanan fault.) The
Cambrian deposits of the Tabas sub-basin, Mila Group,
have a thickness of 1200–3100m and consist of evaporite
and siliciclastic–carbonate rocks. They are divided into the
Kalshaneh, Derenjal and Shirgesht Formations. Study on
the Cambrian trilobites of Iran started in the early twentieth
century. Iranian Cambrian trilobites have been studied by
many authors (King 1930, 1937; Wadia 1934; Balashova
1966; Wolfart 1974; Fortey and Rushton 1976; Kobayashi
1976; Kushan 1978; Wrona and Hamdi 2000; Ameri and
Dastanpour 2010; Ameri 2015). However, the biostrati-
graphy of the Iranian Cambrian trilobites remains poorly
documented.
Unfortunately, Iranian Cambrian trilobites are gen-
erally neglected in global assessments of Cambrian
stratigraphy and palaeontology. The Cambrian deposits
are widely distributed in Iran, but contain very few
fossils, such as the remains of brachiopods, gastropods,
stromatolites and trilobites (only in the Kuhbanan and
Mila Formations). Among these, trilobites are the most
abundant and diverse. Therefore, the study of Cambrian
trilobite fauna in the Cambrian successions is essential
for biostratigraphic and paleobiogeographic models in
Iran.
Based on the trilobite fauna, Iranian Cambrian
successions are deposited in two basins: the Alborz basin
(northern Iran) and central Iran (eastern and southeastern
Iran). Trilobite fauna in the Alborz basin include Lorasian
trilobites, whereas in the central basin (Kerman and Tabas
area) they include pre-Gondwana trilobites. Therefore,
these facies are comparable with Parahio-Spiti (northern
India) (Hayden 1904), Salt Range (Pakistan) (Kobayashi
1976), Hand Ward (northwest Kashmir) (Wadia 1934),
Rabut-Pamir (Tajikistan) (Balashova 1966), South Aus-
tralia (Paterson et al. 2007), South China (Kobayashi
1976) and Surkh Bum (Afghanistan) (Wolfart 1974).
This study aims to provide systematic descriptions of
the Cambrian trilobite taxa of Iran. It also attempts to
q 2015 Taylor & Francis
*Corresponding author. Email: [email protected]
Historical Biology, 2015
http://dx.doi.org/10.1080/08912963.2014.927870
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

construct a biostratigraphic scheme for Series nos 1 and 2
of the Cambrian System in the Kerman sub-basin.
2. Regional stratigraphy
2.1. Godbondar section
Godbondar section (with coordinates 3084801700N,5684404000E) is located to the east of Bondar village (east
Zarand) (Figure 1). TheEasternZarandMountains, ofwhich
Godbondar is a part, are bounded by the Kuhbanan fault to
the west and the Bidou syncline to the east. In the Eastern
Zarand Mountains, the Precambrian and Lower Palaeozoic
successions cropped out, and can be divided frombase to top
into the Rizu and Dezu Series, and the Dahu, Kuhbanan,
Unnamed, Katkoyeh and Niur Formations.
This study is focused mainly on the Kuhbanan
Formation. However, it presents a brief description of all
of the pre-Devonian successions cropping out in the study
area. Rizu Series is composed of siliciclastic and igneous
rocks. Dezu Series is lithologically similar to Rizu Series,
but gypsum and dolomitic rocks are present in its upper
parts. The component rocks of both of Rizu and Dezu
Series show a chaotic pattern. Dezu Series consists of grey
and black dolomites with interbedded gypsum, sometimes
developed in sequences up to 40m thick. The Series also
contains black limestones, violet–red thin-bedded micac-
eous sandstones and siltstones. No fossils have been
reported from the Dezu Series (Zhang et al. 1991).
According to Ruttner et al. (1968), the Dezu Series is
comparable with the Soltanieh and Barut Formations of
northern Iran. If this point of view is correct, it implies that
the lower part of the Dezu Series is Late Precambrian in
age. The Dahu Formation is a red sequence that typically
consists of red sandstones and red sandy shales. Cross-
bedding and ripple marks are present in its sandstones.
There are interbedded fine-grained breccias, and also
coarse sandstones in the Dahu Formation; the latter often
contain well-rounded grains of cherts. In the Godbondar
section the Dahu Formation is up to 45m thick. The Dahu
Formation is coeval with the Lalun Formation (in Zagros
and Alborz). The light-coloured (sandstones and quart-
zites) top zone of the Dahu Formation can be correlated
with the top quartzite of the Lalun Formation. The
Kuhbanan Formation consists of 110–150m of fine-
grained, red–brown sandstones with dolomite lenses and
thick- to thin-bedded limestones. The lithological nature of
the Kuhbanan Formation is very variable; it can be divided
into three informal members, as follows:
. Member 1 consists of thick-bedded, white quartzite
with ripple marks, and chertic pebbles (up to 8 cm in
diameter).. Member 2 includes grey to dark, partly dolomitic,
thick- to thin-bedded limestone (80–120m) with
interbeds of marly limestone. The limestone beds, in
particular, bear numerous remains of trilobites,
brachiopods, gastropods and Orthotheca.. Member 3 consists of unfossiliferous brown-bedded
dolomite with dolomitic lenses.
According to the occurrence of Redlichia, the
Kuhbanan Formation is late Early Cambrian to Middle
Cambrian in age. The Kuhbanan Formation in this area is
disconformably overlain with Unnamed Formation car-
bonate deposits (Cam-Ordovician?) and disconformably
underlain by the ‘top quartzite’ of the Early Cambrian
Dahu Formation (Figure 2).
The Katkoyeh Formation comprises 80m of buff and
grey–green siltstones and silty calcareous mudstones and
limestones with an interbedded red siltstone horizon. The
Katkoyeh Formation is the counterpart of the Shirgesht
Formation of the Tabas sub-basin and upper part of the
Mila Formation (in Alborz). The Katkoyeh Formation in
the study area gradually changes into the Niur Formation
(Silurian clastic deposits). Likewise, the dolomite succes-
sions of the Unnamed Formation disconformably underlie
the Katkoyeh Formation.
In the study area, clastic rocks of theNiur Formation are
partly developed and comprise white quartzitic sandstones,
siltstones, green sandy limestones and pure limestones. The
limestones are fossiliferous and contain different fauna of
brachiopods, trilobites, corals and conodonts.
2.2. Kuhbanan section
Kuhbanan section (with coordinates 3183201700N,
5681700400E), the most fossiliferous and the most complete
section of the Middle Cambrian, with 205m thickness, is
located near Kuhbanan town (about 170 km northwest of
Kerman) (Figure 3). The lower boundary of the Kuhbanan
Formation with the Dahu Formation is distinguished by
sharp lithological changes, so that the red siliciclastic
rocks of the Dahu are covered by the basal white quartzite
of the Kuhbanan. This Formation is conformably covered
by the Bahram deposits, with a sharp faulted contact
(Figure 4).
The Kuhbanan Formation can be subdivided into four
informal lithological members, as follows:
. Member 1 consists of thick-bedded, white to light
grey quartzite with oblique bedding, ripple marks
and chertic pebbles. The quartzite passes laterally
into medium- to coarse-grained, red–brown
sandstone.. Member 2 includes a 75-m thick sequence of
whitish to grey–brown, sandy, weakly lithified
dolomite and/or sandstones. Regionally, the dolo-
mite is partly replaced by fine-grained, often friable,
massive and thin-bedded, red sandstones. The lower
part of the member contains unfossiliferous small
2 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

chertic grains. Lateral changes are common in this
facies.. Member 3 is composed of grey to dark grey, partly
dolomitic or pseudo-oolitic, thick- to thin-bedded
limestonewith yellowish-brown to reddishweathering
interbeddedwith marly limestones. Themiddle part of
the limestones (from 30 to 65m) bears numerous
remains of trilobites, partly completed trilobites,
carapaces, brachiopods and gastropods. Orthotheca
are particularly numerous in coarse crystalline lime-
stone. The upper unfossiliferous part shows an
increasing content of dolomite and chertic layers.
. Member 4 includes the uppermost successions of the
uppermost Middle Cambrian Kuhbanan Formation.
This member with 35m thickness consists mainly of
fine-grained, red–brown, slightly solidified sandstone.
However, dolomitic lenses, grey to yellowish-brown,
bedded dolomite are present subordinately.
3. Material and methods
Rocks of theKuhbananFormationwere investigated and their
fossil contents sampled systematically at two stratigraphic
sections (Godbondar and Kuhbanan). All in situ and
Figure 1. (Colour online) Location of Godbondar section in northern Kerman, central Iran, with dark areas representing mountains inthe Kerman to Saghand region, and geological map of the study area.
Historical Biology 3
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

identifiable fossils were collected. In total, 300 specimens
were collected from the study sections. All materials were
deposited at the Institute of Science, High Technology and
Environmental Sciences, in Kerman, Iran (ISHE).
The examined materials were prepared from marly
limestones using a vibro tool. The specimens were coated
with ammonium chloride to heighten the contrast before
being photographed using ring-light illumination. In most
cases, additional highlight from the northwest quadrant
was used. Photographs were taken using a Nikon D90
digital camera with a 105-mm objective. Apart from
digital sharpening, deepening of contrast and blackening
of the background, the photographs have not generally
been retouched.
4. Biostratigraphy
Thebiostratigraphic zonationproposedhere for theCambrian
System of the Kerman sub-basin is based on the local
occurrence ofwell-known trilobite taxa. The term ‘zone’ was
appliedwhen the eponymous species occurred inmore than a
single bed. Intervals of ‘no zonation’ are indicated in thick,
apparently barren, strata between the recognised zones,
where further work is required to clarify the biostratigraphy.
Biostratigraphic study of Kuhbanan and Godbondar
sections enabled the authors to develop a biostratigraphic
scheme with three trilobite zones for the Cambrian System
of the study sections (Figures 2 and 4). Recognised
trilobite zones spanned from stage 4 to the middle of
Guzhangian Stage. In ascending order, they are the
Figure 2. Stratigraphic section and taxon range chart of the Kuhbanan Formation, Godbondar section.
4 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

Redlichia noetlingi zone, the Kermanella kuhbananensis
zone and the Iranoleesia pisiformis zone. The main
characteristics of the recognised biozones are as follows.
4.1. Redlichia noetlingi zone
The base of this biozone is characterised by the first
appearance datum (FAD) of Redlichia noetlingi and the
upper part of biozone is consistent with the last appearance
datum (LAD) of Redlichia noetlingi. Redlichia noetlingi
ranges from the uppermost Lower Cambrian to the
lowermost Middle Cambrian (Wolfart 1974; Ameri and
Dastanpour 2010), which is comparable with Shihchiao-
Sanshih stages in China. According to the global
chronostratigraphic criteria, Redlichia noetlingi ranges
from the late Early Cambrian to the lowermost Middle
Cambrian, which is comparablewith the Cambrian Series 2
(stage 4). The first sample of Redlichia noetlingi occurs
72m from the base of Godbondar section and 120m from
the base of Kuhbanan section. This zone includes Redlichia
noetlingi, Redlichia chinensis and Redlichia sp. The
Redlichia noetlingi zone is characteristic of member no. 3
of both study sections. Redlichia noetlingi has been
reported from the late Early Cambrian in some sections
from Iran (Wolfart 1974; Ameri 2015), Afghanistan
(Wolfart 1974), China (Kobayashi 1976), South Australia
Figure 3. (Colour online) Location of the Kuhbanan section in northern Kerman, central Iran, with dark areas representing mountains inthe Kerman to Saghand region, and geological map of study area.
Historical Biology 5
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

(Paterson et al. 2007) and the Pakistan Salt Range
(Kobayashi 1976). This biozone includes up to 20m of the
study sections. Trilobite taxa were commonly collected
from several horizons (sometimes in different lithology).
4.2. Kermanella kuhbananensis zone
This zone begins with the FAD of Kermanella kuhbana-
nensis. The species of Kermanella kuhbananensis ranges
from the lowermost Middle Cambrian (Cambrian stage 5)
to the Early Drumian stage, which is comparable with
Tangshih (Chinese stage). The first samples of Kermanella
kuhbananensis and Kermanella lata lata occur 96m from
the base of the Godbondar section and 140m from the base
of the Kuhbanan section, which indicates the Cambrian
stages 4 and 5 boundary. This zone contains Kermanella
kuhbananensis, Kermanella lata lata, Kermanella lata
minuta, and Kermanella sp. The upper limit of the interval
is defined by the LAD of Kermanella kuhbananensis.
Kermanella kuhbananensis has been reported from the
Middle Cambrian in the same sections in Iran (Wolfart
1974; Ameri and Dastanpour 2010; Ameri 2015).
4.3. Iranoleesia pisiformis zone
The base of this biozone is characterised by the FAD of
Iranoleesia pisiformis and the upper part of the biozone is
consistent with the LAD of Iranoleesia pisiformis.
Iranoleesia pisiformis is indicative of the uppermost Middle
Cambrian (Wolfart 1974; Ameri and Dastanpour 2010),
Figure 4. Stratigraphic section and taxon range chart of the Kuhbanan Formation, Kuhbanan section.
6 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

which is comparable with Kushanian stages in China.
According to the global chronostratigraphic criteria,
Iranoleesia pisiformis ranges from the Late Drumian stage
to the Early Guzhangian stage. The first sample of
Iranoleesia pisiformis occurs 180m from the base of the
Godbondar section and 190m from the base of theKuhbanan
section. This zone includes Blountia blountia, Afghanocera
lategenatum, Iranoleesia pisiformis and Iranoleesia sp.
5. Biogeographic considerations
The late Early Cambrian trilobite assemblages recognised in
central Iran show affinities with fauna found in some other
parts of Gondwana or Peri-Gondwana terrains (Figure 5)
The Redlichia noetlingi zone is found in six sections in
northern Kerman, namely Charmis, Dar Abe, Dahu,
Katkuh, Akbarabad and Banestan. Redlichia noetlingi,
which is characteristic of this assemblage, is known only
from central Iran.
Redlichia nobilis, which is recorded from South China
and the Pakistan Salt Range, is very similar to some of the
redlichiids of the central Iran that are assigned to Redlichia
noetlingi.
The Kermanella kuhbananensis zone is reported only
from Kerman region. It shows weak affinities with Pakistan
Salt Range andSouthChina by the occurrence ofAnomocare,
a Salt Range genus that is closely related to the Kermanian
genus Kermanella and the Chinese genus Shantungarpis.
The Iranoleesia pisiformis zone is found at several
localities: southern and northern Kerman (Wolfart 1974;
Ameri 2015), northern Iran (Mila Formation) (Kushan 1978),
Surkh Bum, Afghanistan (Wolfart 1974) and the Pakistan Salt
Range (Kobayashi 1976). It shows medium affinities with the
southern China platform and Australia by the occurrence of
Lioparia, a Chinese genus that is closely related to the
Australian genus Leiopyg olaevigala. This assemblage is
similar to some species of the eastern IranianLioparellagenus.
The biogeographic message of these assemblages seems
clear. The Antarctic faunas do not show any close similarity
with faunas from other geographic regions of the rest of
Gondwana. Thus, althoughGondwanawas a huge continent,
itmust have been located atmedian latitudes in the Early and
Middle Cambrian so that trilobite faunas could spread
widely across its marginal seas (Palmer and Rowell 1995).
The apparent faunal differences may be mainly due to
local habitat conditions. They could also be affected by the
availability of fossiliferous horizons in Peri-Gondwana
province. However, in the Cambrian, Gondwana must
have become isolated from Laurentia, another Cambrian
continent that occupied lower latitudes.
6. Systematic palaeontology
Some of the taxa that aremore abundant andmost important
for biozonation are described below. The measurements
used are illustrated in Figure 6.Measurements, terminology
and abbreviations are derived from Shaw (1957), Kaesler
et al. (1997), Sundberg and McCollum (1997, 2000, 2003)
and Ameri and Dastanpour (2010). Sagittal (sag.) and
exsagittal (exsag.) measurements refer to lengths, whereas
transverse (trans.) measurements refer to widths.
Class Trilobita (Walch, 1771)
Order REDLICHIIDA (Richter, 1932)
Suborder REDLICHIINA (Harrington, Henningsmoen,
Howell, Jaanusson and Lochman-Balk, 1959)
Superfamily REDLICHIACEA (Poulsen, 1927)
Family REDLICHIIDAE (Poulsen, 1927)
Genus Redlichia (Redlichia) (Cossmann, 1902)
Redlichia noetlingi (Redlich, 1899)
Type species Hoeferia noetlingi (Redlich, 1899)
(Figures 7(a),(b),(e) and 8(a),(b),(e))
1899. Hoeferia noetlingi ver. Late et angusta n. g. et n.
sp. – Redlich, Cambrian fauna Salt Range, p. 3–
4, Pl. 1, Fig. 1–8.
1910. Redlichia noetlingi (Redlich) – Reed, Cambrian
fossils Spite, p. 7–9, Pl. 1, Fig. 14.
1930. Redlichia chinensis Walcott – King, Cambrian
fauna Persia, p. 316–318, Pl. 17, Fig. 1.
1937. Redlichia chinensis Walcott. – King, Cambrian
trilobites Iran, p. 4–5, Pl. 1, Fig. 2(a)–(c).
1941. Redlichia noetlingi (Redlich) – King, Cambrian
fauna Salt Range, p. 3–10, Pl. 1, Fig. 1–3, Pl. 3,
Figs 1–9, Pl. 4, Fig. 1–6.
1955. Redlichia noetlingi (Redlich) Schindewolf and
Seilacher, Kambriun Salt Range p. 293–299,
Abb. 1–5, Pl. 6, Fig. 1–15, Pl. 7, Fig. 1–6.
1962. Redlichia cf. chinensis Walcott. Huckriede,
Kursten and Venzlaff, Geological Kerman, p.
35–39.
1962. Redlichia noetlingi (Redlich)/nobilisWalcott/Group-
Huckriede, Kursten and Venzlaff, Geological Ker-
man, p. 35.
1962. Redlichia chinensis Walcott and Redlichia nobilis
Walcott – Kobayashi, Cambrian Eastern Asia,
p. 492.
Materials: Four fragments were collected, mostly
cephala and genalspain.
Description: This species is a characteristic micro-
pygous, with a large cephalon, having opistoparian facial
sutures. The cranidium is large to medium size, length 37–
17mm, width high convexity (sag. and trans.). The
cranidium has a parallel-sided glabella, S1 (trans.)
glabellar furrow, eye ridges directed moderately poster-
olaterally, and moderately wide fixigena (71 ^ 8%
glabellar width). The cranidium area covers about 66–
Historical Biology 7
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

70% of the cephalon, and the anterior border furrow angle
is 120 ^ 58. Anterior width (J2) in relation to cranidial
width (J1) is 70 ^ 2%. Cranidial width (J1) in relation to
cranidial length (A1) is about 53–65%. The preglabellar
field is flattened with tropidium relief in the anterior border
of the preglabellar area. The occipital ring is in relief and
wide, and the occipital furrow is deep and protracted. The
eye ring starts from the anterior branch, with a trend of 308.The glabella is pear shaped and in relief. The preocular area
is absent. The palpebral area is small and limited. The
ocular lobe shows very sharp relief, with a narrow terocular
area. The glabella has three lobes: S1 is completed, S2 has
a deeper furrow and S3 has two branch-lines at the end.
Distribution: Redlichia biozone, Kuhbanan Formation
(north Kerman), Hormoz Formation (southwest Iran), Salt
Range (Pakistan), Himalayan region, southern Siberia,
Australia and south of China.
Redlichia noetlingi has been regarded as late Early
Cambrian in age.
Order PTYCHOPARIDA (Swinnerton, 1915)
Suborder PTYCHOPARIINA (Richter, 1933)
Superfamily ASAPHISCACEA (Raymond, 1924)
Family ASAPHISCIDAE (Raymond, 1924)
Genus Anomocarella (Walcott, 1905)
Kermanella kuhbananensis (Wolfart, 1974)
Type species Anomocarella chinensis
(Figures 9(h), 10(a)–(c), 11(b) and 12(d))
1930. Anomocare megalurus (Dames) – King,
Cambrian fauna Persia, p. 318, Pl. 17, Fig 4.
1962. Anomocare megalurus (Dames) – Huckriede
et al., Geology Kerman, p. 35.
1962. Alokistocariden? – Huckriede et al., Geology
Kerman, p. 38 (Chabdjereh), p. 40 (Charmis).
1967. Protolenoid (Anomocare megalurus) Kobayashi,
Cambrian Eastern Asia, p. 492.
Materials: In total, 102 samples were collected, mostly
cephalon and pygidium.
Description: The glabella has a cylindrical shape, with
basal convexity towards the thorax. The occipital ring is
convex and covers about 16–20% of the glabellar area.
The occipital furrow is deep and protracted; this species is
characterised by three lateral lobes, and S1–S3 have two
branch-lines with 508 trend at the end. The frontal lobes
Figure 5. Correlation of the Cambrian System between northern Kerman and other continents.
Figure 6. Measurement lobe and furrow labels used in thetrilobite descriptions.
8 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

are proximity flattened and show a relief from theirs upper
said to the preglabellar field. S1 has the deepest and widest
furrow. The glabellar field occupies about 41–45% of the
total cranidium area. The preglabellar field is wide and
flattened, and occupies about 14–16% of the cranidium
area. The eye ridge is in relief and parallel to the margin of
the preglabellar field. The preglabellar furrow is deep and
V-shaped. The posterior area is wide and proximately
flattened. The cranidium is wide (J1) in relation to its
length (A1), 50–57%, and the length of the occipital ring
(C) in relation to cranidium length (A1) is 25–28%.
Kermanella kuhbananensis is attributed to the early
Middle Cambrian (Drumian stage) in age.
Occurrence: The taxa were found 96m from the base
of Godbondar section.
Sample numbers: 1252 ISHE and 1253 ISHE.
Order PTYCHOPARIIDA (Swinnerton, 1915)
Suborder PTYCHOPARIINA (Richter, 1932)
Superfamily ASAPHISCACEA (Matthew, 1887)
Family ASAPHISCACEA (Raymond, 1924)
Genus Kermanella lata minuta (Wolfart, 1974)
(Figures 5(e) and 12(c))
Diagnosis: A sub-type of Kermanella lata with a broad
cranidium (average length) at 63% of the base width, wide
interocular fixed cheeks (on average 85% glabellar width),
small eyes (big eye index, on average 21%) and very
slightly divergent branches in the front seam.
Description: The outer contour of the cephalon is wide
and U-shaped, with short legs, slightly separate, posterior
margin in the area of the neck ring to the back, sides almost
Figure 7. All figures are dorsal view: a, b, e, Redlichia noetlingi, cranidium, £ 3; d, Redlichia chinensis, cranidium, £ 3; c, Redlichiasp., cranidium, £ 2.
Historical Biology 9
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015
straight, slightly angled to the rear running outside. The
front median border is slightly wider than the sides, and
much narrower (sag.) than the preglabellar field. The
frontal border furrow has a weak side, becoming shallower
backward. Behind border of the eyes is wider and larger
than their inside border. The neck ring represents a rather
broad (sag.) laughing-shape, with a narrow side at the
posterior margin. The neck furrow is shallow, transversely.
Back furrows are shallow, curved, very slightly convex,
slightly convergent forward and united from the front to
back furrow.
The glabellar outline has a dented rear edge and is
similar to that of Kermanella kuhbananensis, but its
anterior margin is slightly convex. The measured
cranidium length in 10 samples involves 57%–68% of
the cephalon. Preglabellar cranidial sections are less than
half of the neck rings lengths. The interocular fixed cheeks
are very wide; so that our investigations on the 10 samples
show that they involve 77–93% of the glabella. The
palpebral lobes are very small and slightly curved, with a
long eye. The eye strips are poorly developed, only slightly
curved inwards and extending obliquely forward, as the
front glabellar corners plunge back into the furrows. The
front seam branches diverge forward, curving slightly to
10 ^ 58, the seam extending rear branches are slightly
curved convexly forward, obliquely rearward outwardly,
and bend only in the region of behind it, but backwards.
Owing to the median position of the eyes and the course of
the seam behind the branches, the fixed cheek rear sections
are comparatively long (sag.) and wide (trans.). In side
view, the apex line is extended to the apex plane or it is
inclined very slightly to the front on the neck ring.
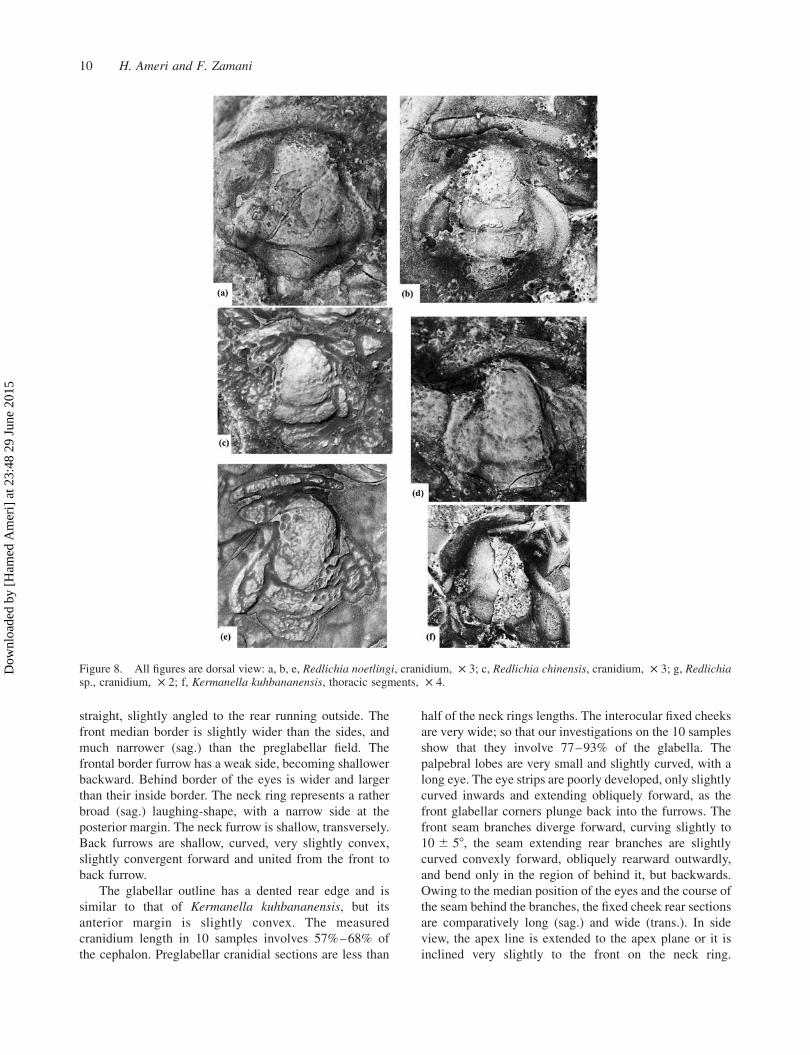
Figure 8. All figures are dorsal view: a, b, e, Redlichia noetlingi, cranidium, £ 3; c, Redlichia chinensis, cranidium, £ 3; g, Redlichiasp., cranidium, £ 2; f, Kermanella kuhbananensis, thoracic segments, £ 4.
10 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015
Gradually, the apex line then tends somewhat more
forward, until approximately from S3 to the entire front to
the leading edge the cranidial part slopes quite evenly. The
preglabellar field and front hemline rut are cause very
slight undulations over the crest line. The front view is
dominated by the relatively small total curvature of the
cephalon. The strongest cross bow is the neck ring, the
glabella rises only slightly above the general surface, fixed
interocular cheeks arch across very small free cheeks, and
the fixed cheek rear sections weakly slope outwards below.
The hypostome and pygidium are unknown.
Relations: Kermanella lata lata. The eyes are bigger
than minuta. The cephalon is positioned behind the centre.
It has a long eye index, on average 31% of minuta, a
deviating course of the back seam branches, and
differently shaped fixed cheek rear sections.
Time: Because of the association with Redlichia
noetlingi, it is only average to younger Lower Cambrian
in age.
Superfamily ASAPHISCACEA (Matthew, 1887)
Family ASAPHISCACEA (Raymond, 1924)
Genus Kermanella lata lata (Wolfart, 1974)
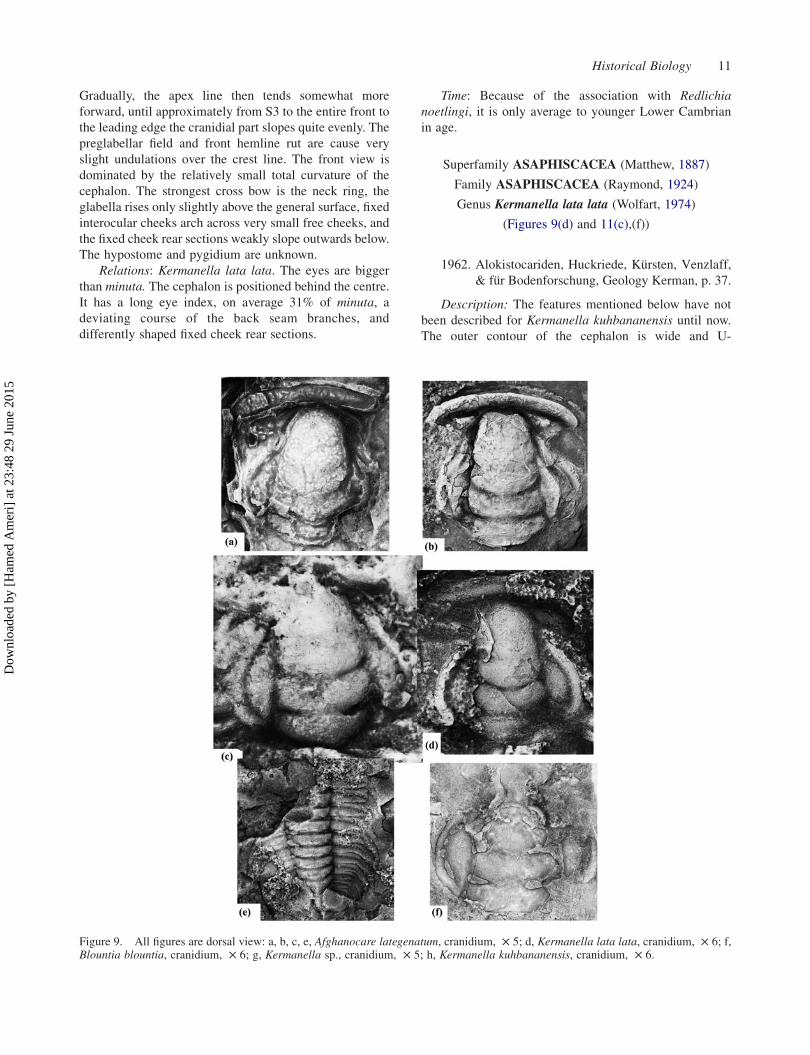
(Figures 9(d) and 11(c),(f))
1962. Alokistocariden, Huckriede, Kursten, Venzlaff,
& fur Bodenforschung, Geology Kerman, p. 37.
Description: The features mentioned below have not
been described for Kermanella kuhbananensis until now.
The outer contour of the cephalon is wide and U-
Figure 9. All figures are dorsal view: a, b, c, e, Afghanocare lategenatum, cranidium, £ 5; d, Kermanella lata lata, cranidium, £ 6; f,Blountia blountia, cranidium, £ 6; g, Kermanella sp., cranidium, £ 5; h, Kermanella kuhbananensis, cranidium, £ 6.
Historical Biology 11
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015
shaped, with short, slightly curved backward inside legs,
the rear edge of the neck ring portion bulges rearward
laterally, slightly curved rearwards and outwards. The
front hemline median is slightly wider (sag.) than the
side, the front hemline rut weak side seam furrow is
only vaguely developed. It is difficult to say that the
neck ring nodules are developed in K. lata lata, because
of their unfavourable conservation. The neck furrow is
weakly pronounced, as are the back furrows, which are
curved slightly concave, and unite before the glabella
to the end union, the back furrow is shallow, and this
is about the same width as the back longitudinal
furrows.
The glabella in outline is like Kermanella kuhbana-
nensis, except that the leading edge is sometimes slightly
curved convexly.
The measured cranidium length in 10 samples involves
55% and 70% of the cephalon. The preglabellar cranidial
section is less than half as long as the glabella (with neck
ring), before the glabella is weaker than the convex side
(sag.). The intraocular hard-cheeks are arched transver-
sely; so that our investigations on the 10 samples show that
they involve 75–91% of the glabella. The palpebral lobes
are slightly curved, and the eye plate index is between 25%
and 38%, somewhat higher than in Kermanella kuhbana-
nensis. Eye strips on the inside of the clear cranidium are
formed in a convex arc, the front end to the front of the
connecting through the back palpebral lobe furrow through
the glabella (range between S3 and S4). The intraocular
transfixed cheeks are hardly curved, slightly inclined
outwards. The fixed cheeks rear sections are comparatively
long (trans.) and narrow.
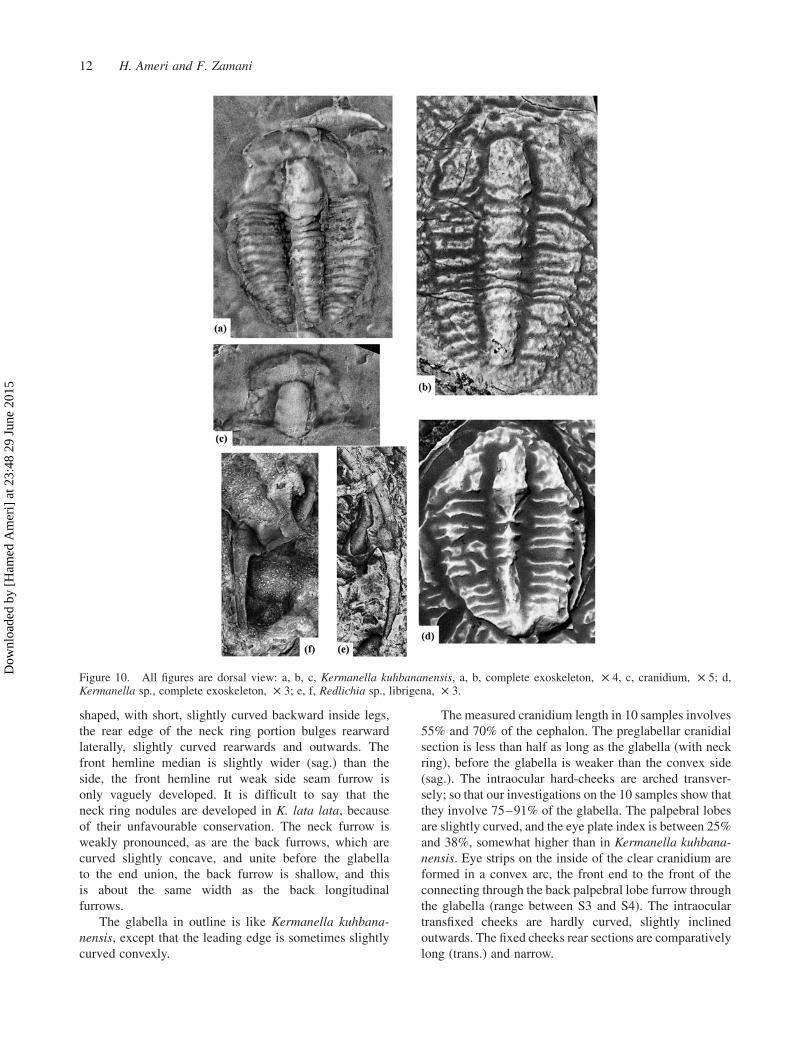
Figure 10. All figures are dorsal view: a, b, c, Kermanella kuhbananensis, a, b, complete exoskeleton, £ 4, c, cranidium, £ 5; d,Kermanella sp., complete exoskeleton, £ 3; e, f, Redlichia sp., librigena, £ 3.
12 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

The front seam diverges at an angle of about 10–158 tothe front hemline and turns closely bowing forwards
inside. Behind the seam, the first branches extend
obliquely rearwards and outwards, then turn at a fairly
large distance from the rear edge to the rear, and meet a
strong curve approximately at a right angle with the rear
edge.
In the side view, the apex line extends most pieces
from the neck ring to the rear edge S2, by the slightly
recessed neck furrow, broken below, almost at the apex
level, and sometimes even rising slightly. S2 is situated
before a gentle and gradual decline begins from the front
hemline rut. The flat to slightly domed front hemline fits
with a slight bend at the cranidium. Only a few pieces of
the end of the glabella are slightly more pronounced, so
that the apex line appears kinked. In front view, the entire
cephalon including the free cheeks as a very weakly
curved shield: the glabella is slightly convex, interocular
free cheeks are barely sprung, but the back grooves on the
outside are tilted weakly.
The pygidium in outline is transversely oval, and has a
curved outer edge which is slightly higher than the front
edge. Length of the pygidium is 1/3 of the cephalon length,
to the rear tapers only slightly, the rear border is blunt,
with a rounded front spindle ring curved sharply forward.
The furrow between the first and second rings is as wide as
the rings themselves. The second ring is hardly separated
from the rear undifferentiated stem section. Curved edges
are weakly curved with two or three low segments. Only
between the front two segments is a seam groove obtained.
A hem is thereby formed so that the grooves end in front of
the outer edge.
Figure 11. All figures are dorsal view: a, Kermanella sp., incomplete exoskeleton, £ 5; b, Kermanella kuhbananensis, completeexoskeleton, £ 4; c, f, Kermanella lata lata, complete exoskeleton, £ 4; d, e, Kermanella lata minuta, cranidium, £ 6; g, h, l, Iranoleesiapisiformis, cranidium, £ 4.
Historical Biology 13
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

Relations: Kermanella lata minuta. Distinguishing
features are known only from the head. Eyes small (sag.)
in median position, large eye index, average 21%. Fixed
cheeks rear sections rather long (sag.) and wide (trans.).
Time: Medium to younger Lower Cambrian, bearing
the Redlichia noetlingi (Sanshih Shihchiao-up stage?).
Family ORDOSIIDAE (Lu, 1954)
Subfamily CHELIDONOCEPHALINAE (Wittke, 1984)
Genus Iranoleesia (King, 1955)
Iranoleesia pisiformis (King, 1937)
(Figure 11(g),(h),(l))
1937. Irania pisiformis King, p. 12–13, Pl. 11, Fig. 6
(a)–(c).
1955. Iranoleesi apisiformis King, p. 86.
1967. Hundwarella pisiformis King Kobayashi, p. 493.
1976. Iranoleesia pisiformis King p. 330, Pl. 9, Fig. 6,
8–10 and 12.
1984. Iranoleesia pisiformis spinosa n., p. 106, Pl. 1,
Figs 12, 16 and 17.
Material: More than 40 cranidia, seven librigena, a
number of disarticulated fragments of thoracic segments,
and sex pygidium.
Occurrence: Kuhbanan Formation, Godbondar section
(ISHE 1997, ISHE 1996, ISHE 1992, ISHE 1993, ISHE
1994, ISHE 1998, ISHE 1980, ISHE 1981, ISHE 1972,
ISHE 1973, ISHE 1974, ISHE 1975, ISHE 1962 and ISHE
1952).
The glabella is forward tapering, with weakly curved
lateral margins. The front glabella is sub-truncate, with
Figure 12. All figures are dorsal view: a, d, Kermanella kuhbananensis, cranidium, £ 6; b, Kermanella lata lata, complete exoskeleton,£ 4; c, Kermanella lata minuta, cranidium, £ 4; e, Iranoleesia sp., cranidium, £ 5; f, Redlichia sp., librigena, £ 3; g, Kermanella sp.,cranidium, £ 5.
14 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

shallow indention medially. The lateral glabellar furrows
are relatively faint on the exterior, but marked by an
absence of surface ornament; they are clearly visible
directly anterior to S3 in most specimens. The occipital
furrow consists of well-incised, slightly backward
directed lateral sections and a shallow median section.
Frontal indention of glabella created by fused parafrontal
sections at anterolateral corners, which extend over axial
furrows into the eye ridges. The eye ridges are nearly
straight, slightly backwards directed from the dorsal
furrows, moderately well defined on internal moulds,
faint on the external cuticle, clearly separated from the
palpebral lobes by narrow grooves. Palpebral lobes of
26–33% cephalic length, typically protolenoid, sausage-
shaped, convex in trans. section, straight in exsag.
direction. The preocular area has a moderately steep
ventral slope, anteriorly defined by a gently curved, deep
anterior border furrow. The anterior border furrow has a
clearly shallower and slightly posteriorly deviated median
section in front of the glabella created by a plectrum
developed as a slightly swollen, weakly defined ridge in
front of the glabella. The lateral borders of the plectrum
can be understood as shallow posteriorly curved sections
originating from the anterior border furrow to meet the
preglabellar furrow immediately in front of the anterior-
most projections of the anterior corners. Well-preserved
specimens show creases that cross the anterior border
furrow in front of the glabella. The cranidium area covers
about 50–61% of the cranidium, and the anterior border
furrow angle is 132 ^ 58. The anterior width (J2) relation
to cranidial width (J1) is 73 ^ 2%. The cranidium width
(J1) relation to cranidial length (A1) is about 73–75%.
The preglabellar field is flattened with tropidium relief in
the anterior border of the preglabellar area. The occipital
ring is in relief and wide, and the occipital furrow is deep
and protracted.
7. Conclusions
Trilobite fauna from two stratigraphic sections (Godbondar
and Kuhbanan) of the Kuhbanan Formation in northern
Kerman (central Iran) were studied and subjected to
biostratigraphic analysis.
Eleven genera and species are recognised from the
latest Early Cambrian and Middle Cambrian Peri-
Gondwana successions of the study sections. Recognised
fauna include Afghanocare lategenatum, Blountia bloun-
tia, Iranoleesia sp., Iranoleesia pisiformis, Kermanella
kuhbananensis, Kermanella lata lata, Kermanella lata
minuta, Kermanella sp., Redlichia chinensis, Redlichia
noetlingi and Redlichia sp.
The age of the study sections is late Early Cambrian to
late Middle Cambrian, based on recognised trilobite
biozones.
Based on the trilobite distribution, three trilobite
biozones were recognised in the study sections, namely
Redlichia noetlingi biozone, Kermanella kuhbananensis
biozone and Iranoleesia pisiformis biozone.
The late Early Cambrian trilobite assemblages (Red-
lichia and Kermanella) recognised from northern Kerman
are similar to those found from some other parts of
Gondwana or Peri-Gondwana terrains, Parahio-Spiti
(northern India) (Hayden 1904), Salt Range (Pakistan)
(Kobayashi 1976), Hand Ward (northwest Kashmir)
(Wadia 1934), Rabut-Pamir (Tajikistan) (Balashova
1966), South Australia (Paterson et al. 2007), South
China (Kobayashi 1976) and Surkh Bum (Afghanistan)
(Wolfart 1974), and show affinities with fauna found in the
Mila Formation (Alborz, northern Iran) (Kushan 1978),
Derenjal Formation (Tabas, eastern Iran) (Ruttner et al.
1968) and South East Karman (Ameri and Dastanpour
2010; Ameri 2015).
Acknowledgements
We extend warm thanks to Dr. Ebrahim Mohammadi for readinga draft version of the manuscript and making helpful suggestions.
Funding
This work was supported by the Institute of Science, HighTechnology and Environmental Sciences, Graduate University ofAdvanced Technology, Kerman, Iran [number 1.1869].
Disclosure statement
No potential conflict of interest was reported by the authors.
References
Ameri H. 2015. Peri-Gondwana late Early-Middle Cambrian trilobites fromtheKuhbanan Formation inDahu section,Kerman Province, Iran. ArabJ Geosci. 8(3):1467–1478. doi:10.1007/s12517-013-1207-8.
Ameri H, Dastanpour M. 2010. Stratigraphy and palaeontology oftrilobites in Kuhbanan Formation in the Akbar-Abad section (northof Kerman). J Sci Islam Repub Iran. 21:37–42.
Balashova E. 1966. Trilobites from the Ordovician and Silurian beds ofPamir. Trudy Upravlenia Geologii. Sovieta Ministrov Tadzhiskoi. 2:191–262.
CossmannM. 1902. Rectification de la nomenclature. Rev Crit Paleozool.16:52.
Fortey R, Rushton A. 1976. Chelidonocephalus trilobite fauna from theCambrian of Iran. Bull Br Mus (Nat Hist) Geol. 27:321–340.
Harrington H, Henningsmoen G, Howell B, Jaanusson V, Lochman-BalkC, Moore, R, Poulsen, C, Rasetti, F, Richter, E and Richter, R. 1959.Arthropoda 1, Trilobitomorpha. In: Moore RC, editor. Treatise oninvertebrate paleontology. Part O. New York, NY: GeologicalSociety of America and University of Kansas; p. 560.
Hayden SHH. 1904. The geology of Spiti: with parts of Bashahr andRupshu. Mem Geol Surv India. 36:1–129.
Kaesler RL, Moore RC, Whittington HB. 1997. Treatise on invertebratepaleontology: Arthropoda 1: Trilobita, revised. Introduction, OrderAgnostida, Order Redlichiida. New York, NY: Geological Society ofAmerica.
Historical Biology 15
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015

King W. 1930. Notes on the Cambrian fauna of Persia. Geol Mag. 67(7):316–327. doi:10.1017/S0016756800099416.
King WBR. 1937. Cambrian trilobites from Iran (Persia). Calcutta:Geological Survey of India.
King, W.B.R. 1955. Iranoleesia, new trilobite name. GeologicalMagazine. 92(01):86–86.
Kobayashi T. 1976. Distribution of Cambrian trilobites in the Peri-Gondwana seas. Proc Jpn Acad. 52:187–190.
Kushan B. 1978. Stratigraphy and trilobite fauna of the Mila Formation(Middle Cambrian–Tremadocian) of the Alborz Range, North Iran.Tehran: Ministry of Information Press.
Lu, Y.-H. 1954. Two new trilobite genera of the Kushan Formation. ActaPalaeontologica Sinica. 2(4):409–438.
Matthew, G. 1887. Illustrations of the fauna of the St. John Group,Number 4, Pt. 2, The smaller trilobites with eyes (Ptychoparidae andEllipsocephalidae). Transactions and Proceedings of the RoyalSociety of Canada, series, 2(5):39–66.
Palmer AR, Rowel AJ. 1995. Early Cambrian trilobites from theShackleton limestone of the central Transantarctic Mountains.Paleontol Soc Mem. 45:1–28.
Paterson JR, Jago JB, Brock GA, Gehling JG. 2007. Taphonomy andpalaeoecology of the emuellid trilobite Balcoracania dailyi (earlyCambrian, South Australia). Palaeogeogr Palaeoclimatol Palaeoecol.249(3–4):302–321. doi:10.1016/j.palaeo.2007.02.004.
Poulsen C. 1927. The Cambrian, Ozarkian and Canadian faunas ofnorthwest Greenland. Medd Grønland. 70:233–343.
Raymond PE. 1924. New Upper Cambrian and Lower Ordoviciantrilobites from Vermont. Boston Soc Nat Hist Proc. 27:386–466.
Redlich K. 1899. The Cambrian fauna of the Eastern Salt Range.Calcutta: Geological Survey of India.
Richter R. 1932. In: Dittler R, Joos G, Korschelt E, Linek G, Oltmanns F,Schaum K, editors. Handworterbuch der Naturwissenschaften. 2nded. Jena: Gustav Fisher; p. 840–864.
Richter, R. 1933. Crustacea. Handworterbuch d. Naturwissenschaften.2nd ed. Jena: 840–863.
Ruttner AW,NabaviM,Hajtan J. 1968. Geology of the Shirgesht area (Tabasarea, east Iran).Tehran:MinistryofEconomy,GeologicalSurveyof Iran.
Shaw AB. 1957. Quantitative trilobite studies II. Measurement of thedorsal shell of non-agnostidean trilobites. J Paleontol. 31:193–207.
Sundberg FA, McCollum LB. 1997. Oryctocephalids (Corynexochida:Trilobita) of the Lower-Middle Cambrian boundary interval fromCalifornia and Nevada. J Paleontol. 71:1065–1090.
Sundberg FA, McCollum LB. 2000. Ptychopariid trilobites of the Lower-Middle Cambrian boundary interval, Pioche Shale, southeasternNevada. J Paleontol. 74(4):604–630. doi:10.1666/0022-3360(2000)074,0604:PTOTLM.2.0.CO;2.
Sundberg FA, McCollum LB. 2003. Early and Mid Cambrian trilobitesfrom the outer-shelf deposits of Nevada and California, USA.Palaeontology. 46(5):945–986. doi:10.1111/1475-4983.00328.
Swinnerton, H. 1915. II.—Suggestions for a Revised Classification ofTrilobites. Geological Magazine (Decade VI). 2(11): 487–496.
Wadia D. 1934. The Cambrian-Trias sequence of north-western Kashmir(parts of Muzaffarabad and Baramula districts). Rec Geol Surv India.68:121–176.
WalchJEI. 1771.DieNaturggeschichte derVersteinerungen.ZurErlauterungder Knorrischen Sammlung von Merkwurdigkeiten der Natur. [Thenatural historyof the fossils. To explain theKnorrischen collection of theMerkwurdigkeiten]. 4:184.
Walcott, C.D. 1905. Cambrian faunas of China. US Government PrintingOffice.
Wolfart R. 1974.Die Fauna (Brachiopoda,Mollusca, Trilobita) des alterenOber-Kambriums (Ober-Kushanian) von Dorah Shah Dad, Sudost-Iran, und Surkh Bum, Zentral-Afganistan. [The fauna (brachiopods,mollusca, Trilobita) of older Upper Cambrian (Upper- Kushanian)from Dorah Shah Dad, Southeast Iran, and Surkh Bum, centralAfghanistan]. Geol Jahrb. 8:71–184.
Wittke, H.W. 1984. Middle and Upper Cambrian trilobites from Iran:their taxonomy, stratigraphy and significance for provincialism.Palaeontographica Abteilung A: 91–161.
Wrona R, Hamdi B. 2000. Middle Cambrian Hadimopanella from MilaFormation in the Alborz Mountains, northern Iran. In: Acenolaza GF,Peralta S, editors. Cambrian from the southern edge. 6. Miscelana:Instituto Superior de Correlacion Geologica; p. 143–146.
Zhang S, Zhen Y, China MM, Nairn A. 1991. The Phanerozoic geology ofthe world: I. The Paleozoic A. Amsterdam: Elsevier.
16 H. Ameri and F. Zamani
Dow
nloa
ded
by [
Ham
ed A
mer
i] a
t 23:
48 2
9 Ju
ne 2
015
Related Documents











