Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia Essam F. Sharaf 1, 3 , Marcelle K. BouDagher-Fadel 2 , J. A. (Toni) Simo 3, 4 , and A. R. Carroll 3 1 Department of Geology, Faculty of Science, Mansoura University, Mansoura 35516, Egypt email: [email protected] 2 Postgraduate Unit of Micropalaeontology of Geological sciences, University College London, Gower Street, London WC1E 6BT, U.K, email: [email protected] 3 Department of Geology and Geophysics, UW-Madison, 1215 W. Dayton Street, Madison, WI 53706, USA 4ICREA - Dept. Eng. Terreny, Cartografia y Geofisica, UPC, 08034 Barcelona, Spain INTRODUCTION The Oligocene and Miocene of the East Java Basin provide an excellent opportunity to examine the comparative responses of clastic and carbonate depositional systems to eustatic and tec- tonic controls (Bransden and Matthews 1992). In this area, large and isolated carbonate buildups surrounded by deep ma- rine carbonates or shallow- and deep-water siliciclastic rocks. The variation in facies distributions and faunal assemblages are among the major consequences of the eustatic and tectonic evo- lution of the basin, but their analysis requires improved geo- chronology. Larger benthic foraminifera (LBF) are most prolific in warm shallow water carbonate sediments. On modern reefs, they may constitute up to 15% of the skeletal debris associated with other reef builders (Maxwell 1968). Cenozoic LBF are very abundant in Indo-Pacific Ocean, having been described from many local- ities in Philippines (Carozzi 1976); Japan (Matsumaru 1977); Borneo (BouDagher-Fadel, and Lord 2001; BouDagher-Fadel and Wilson 2000) and Indonesia. Many LBF have biotopes closely associated with carbonate environments. Their geo- graphic distribution is controlled by environmental parameters such as light penetration, nutrient availability, turbidity, water depth, water temperature, and salinity (Cole 1957, 1963; Renema 2002). In general, most LBF taxa have long strati- graphic ranges, but well-established genera are morphologi- cally distinct and have different stratigraphic ranges (see BouDagher-Fadel and Banner 1999). The co-occurrence of planktonic foraminifera and LBF in the same section is a rare opportunity to refine the biostratigraphic ranges of some of these LBF and to refine the biostratigraphic frameworks based on these two groups. The first attempt to use the planktonic foraminiferal biostrati- graphy in the Indonesian region was made by Bolli (1966), when he applied his 1957 Trinidad and Tobago zonation to the Wonocolo Formation in well Bojonogoro-1 at the southwestern part of east Java Basin, Indonesia. The planktonic zones intro- duced by Blow (1969) and revised by Berggren et al., (1995) are used in the zonation scheme of this work and are correlated with the geologic time scale of Ogg and Smith, (2004). Addi- tional previous work on the Neogene planktonic foraminifera in EJB was published by Muhar (1957) and Brouwer (1966). The main purposes of this paper are to refine the biostrati- graphic framework of Oligocene-Miocene outcrops in East Java Basin and to calibrate the stratigraphic ranges obtained from the identified fossil assemblages with the geochronology of the ex- posures based on the strontium isotopic compositions of some larger benthic forminiferal content. GEOLOGIC AND STRATIGRAPHIC SETTING The study area occupies the northern part of the East Java Basin (text-fig. 1). It is located within the Rembang Zone and the northern part of Randublatung Zone (Van Bemmelen 1949). The Rembang Zone consists of series of E-W oriented hills with maximum elevation of about 500 m (text-fig. 1). Those hills gener- ally represent anticlines that may or may not be faulted. The Randublatung Zone is to the south (text-fig.1) and represents a phys- iographic depression that contains folds such as Pegat and Ngimbang anticlines (Duyfjes 1938). During the Cenozoic, the relative movement of the Indian, Eur- asian and Australian plates controlled the tectonic setting of the EJB. The basin exhibits a complex history of initial extension followed by differential basin subsidence and later tectonic in- version (Hamilton 1979). During the rifting, Paleogene exten- sion defined NE-SW trending horsts and lacustrine grabens (Cole and Crittenden 1997; Hamilton 1979). During the Eocene to early Oligocene, a marine transgression associated with mod- erate differential subsidence flooded the area and marine car- bonates were deposited across the entire region. Isolated carbonate mounds (red algae and coral dominated) grew above structural highs. Mound growth continued, with more aerial re- striction, into the Middle Miocene. The mounds were sur- rounded by off-mound facies mostly of marls and chalks with occasional shallow-water carbonate debris derived from the mounds. The Burdigalian-Langhian represents the initiation of clastic influx (Bransden and Matthews 1992); clastic deltas prograded from the north (Koesoemadinata and Pulunggono 1975). The clastic influx initially digitated with carbonate mounds but overwhelmed the entire region in the Serravallian. During the Late Miocene-Middle Pliocene (˜7 Ma- 3.6 Ma), northern East Java was affected by contraction forming south-verging faults and en-echelon east-west folds. A final re- organization of the basin occurred during the Late Pliocene un- til Recent (˜3.6 Ma- 0 Ma) where the southern area of EJB (Kendeng Zone) was affected by north-verging thrusts and up- stratigraphy, vol. 2, no. 3, pp. xxx-xxx, text-figures x-xx, plates x-x, tables x-x, 2005 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biostratigraphy and strontium isotope dating ofOligocene-Miocene strata, East Java, Indonesia
Essam F. Sharaf 1, 3, Marcelle K. BouDagher-Fadel 2, J. A. (Toni) Simo3, 4, and A. R. Carroll3
1Department of Geology, Faculty of Science, Mansoura University, Mansoura 35516, Egypt
email: [email protected] Unit of Micropalaeontology of Geological sciences, University College London,
Gower Street, London WC1E 6BT, U.K,
email: [email protected] of Geology and Geophysics, UW-Madison, 1215 W. Dayton Street, Madison, WI 53706, USA
4ICREA - Dept. Eng. Terreny, Cartografia y Geofisica, UPC, 08034 Barcelona, Spain
INTRODUCTION
The Oligocene and Miocene of the East Java Basin provide anexcellent opportunity to examine the comparative responses ofclastic and carbonate depositional systems to eustatic and tec-tonic controls (Bransden and Matthews 1992). In this area,large and isolated carbonate buildups surrounded by deep ma-rine carbonates or shallow- and deep-water siliciclastic rocks.The variation in facies distributions and faunal assemblages areamong the major consequences of the eustatic and tectonic evo-lution of the basin, but their analysis requires improved geo-chronology.
Larger benthic foraminifera (LBF) are most prolific in warmshallow water carbonate sediments. On modern reefs, they mayconstitute up to 15% of the skeletal debris associated with otherreef builders (Maxwell 1968). Cenozoic LBF are very abundantin Indo-Pacific Ocean, having been described from many local-ities in Philippines (Carozzi 1976); Japan (Matsumaru 1977);Borneo (BouDagher-Fadel, and Lord 2001; BouDagher-Fadeland Wilson 2000) and Indonesia. Many LBF have biotopesclosely associated with carbonate environments. Their geo-graphic distribution is controlled by environmental parameterssuch as light penetration, nutrient availability, turbidity, waterdepth, water temperature, and salinity (Cole 1957, 1963;Renema 2002). In general, most LBF taxa have long strati-graphic ranges, but well-established genera are morphologi-cally distinct and have different stratigraphic ranges (seeBouDagher-Fadel and Banner 1999). The co-occurrence ofplanktonic foraminifera and LBF in the same section is a rareopportunity to refine the biostratigraphic ranges of some ofthese LBF and to refine the biostratigraphic frameworks basedon these two groups.
The first attempt to use the planktonic foraminiferal biostrati-graphy in the Indonesian region was made by Bolli (1966),when he applied his 1957 Trinidad and Tobago zonation to theWonocolo Formation in well Bojonogoro-1 at the southwesternpart of east Java Basin, Indonesia. The planktonic zones intro-duced by Blow (1969) and revised by Berggren et al., (1995)are used in the zonation scheme of this work and are correlatedwith the geologic time scale of Ogg and Smith, (2004). Addi-tional previous work on the Neogene planktonic foraminifera inEJB was published by Muhar (1957) and Brouwer (1966).
The main purposes of this paper are to refine the biostrati-graphic framework of Oligocene-Miocene outcrops in East JavaBasin and to calibrate the stratigraphic ranges obtained from theidentified fossil assemblages with the geochronology of the ex-posures based on the strontium isotopic compositions of somelarger benthic forminiferal content.
GEOLOGIC AND STRATIGRAPHIC SETTING
The study area occupies the northern part of the East Java Basin(text-fig. 1). It is located within the Rembang Zone and thenorthern part of Randublatung Zone (Van Bemmelen 1949).The Rembang Zone consists of series of E-W oriented hills withmaximum elevation of about 500 m (text-fig. 1). Those hills gener-ally represent anticlines that may or may not be faulted. TheRandublatung Zone is to the south (text-fig.1) and represents a phys-iographic depression that contains folds such as Pegat andNgimbang anticlines (Duyfjes 1938).
During the Cenozoic, the relative movement of the Indian, Eur-asian and Australian plates controlled the tectonic setting of theEJB. The basin exhibits a complex history of initial extensionfollowed by differential basin subsidence and later tectonic in-version (Hamilton 1979). During the rifting, Paleogene exten-sion defined NE-SW trending horsts and lacustrine grabens(Cole and Crittenden 1997; Hamilton 1979). During the Eoceneto early Oligocene, a marine transgression associated with mod-erate differential subsidence flooded the area and marine car-bonates were deposited across the entire region. Isolatedcarbonate mounds (red algae and coral dominated) grew abovestructural highs. Mound growth continued, with more aerial re-striction, into the Middle Miocene. The mounds were sur-rounded by off-mound facies mostly of marls and chalks withoccasional shallow-water carbonate debris derived from themounds. The Burdigalian-Langhian represents the initiation ofclastic influx (Bransden and Matthews 1992); clastic deltasprograded from the north (Koesoemadinata and Pulunggono1975). The clastic influx initially digitated with carbonatemounds but overwhelmed the entire region in the Serravallian.During the Late Miocene-Middle Pliocene (˜7 Ma- 3.6 Ma),northern East Java was affected by contraction formingsouth-verging faults and en-echelon east-west folds. A final re-organization of the basin occurred during the Late Pliocene un-til Recent (˜3.6 Ma- 0 Ma) where the southern area of EJB(Kendeng Zone) was affected by north-verging thrusts and up-
stratigraphy, vol. 2, no. 3, pp. xxx-xxx, text-figures x-xx, plates x-x, tables x-x, 2005 1
geouser
Inserted Text
are
geouser
Cross-Out
geouser
Inserted Text
as well as

lifted. The uplift was accompanied by volcaniclastic influxfrom south to north (Soeparjadi et al. 1975).
Oligocene-Miocene outcrops in the EJB include carbonates, shalesand sandstones that are rich in coralline algae, corals, larger benthicand planktonic foraminifera. These lithologies and exposures arenot widely known; there is little previous stratigraphic, sediment-ologic and biostratigraphic work published of these rocks. Previouspaleontological studies in this area include Duyfjes (1936 and1938); Van Bemmelen (1949); Muhar (1957); Bolli (1966);Brouwer (1966); Van Vlerk and Postuma (1967); Pringgo-prawiro et al. (1977); van Vessem (1978); Ardhana et al. (1993)and Lunt et al. (2000). We follow the nomenclature establishedby Bataafsche Petroleum Maaschappiji, BPM, (1950) and JOBPertamina-Tuban (1990) as cited by Ardhana et al. (1993);within each formation we have recognized different lithologies(text-fig. 2). The Oligocene-Miocene stratigraphic units of in-terest of this work and exposed in East Java are the Kujung,Tuban and Ngrayong Formations and the Bulu Member ofWonocolo Formation (text-fig. 3). Briefly, these stratigraphic unitsare summarized below.
Kujung Formation
The Kujung Formation is the oldest formation exposed in thestudy area (text-figs. 1 and 2). The age of Kujung Formation
has been established as latest Early Oligocene (Rupelian, P20)to Early Miocene (Aquitanian N4) (Najoan 1972; Duyfjes 1941;cited in Lunt et al. 2000). It is subdivided into three, mappeable,sedimentary packages (Indonesian Map 1997). The exposed lowerKujung is represented by reefal carbonates (Darman and Sidi 2000).The exposed middle Kujung is represented by interbedded shaleand chalk lithologies rich in planktonic foraminifera. The ex-posed upper Kujung forms a resistant ridge (Prupuh Ridge, nearthe Prupuh village text-fig. 1) that exposes interbedded chalkycarbonates and graded-bedded grainstone with scour surfacesand load cast features. These lithologies are also known as thePrupuh Member carbonates (text-fig. 2).
Tuban Formation
The Tuban Formation is widely exposed along the EJB(text-fig.1) and has been dated as Burdigalian to Langhian(Ardhana et al. 1993). The Tuban lithologies are highly vari-able. Informally, we have distinguished three main lithologies(sandstone, carbonates and shale) that alternate in space andtime. The Tuban sandstone and carbonate lithologies are wellexposed in western Rembang area while the Tuban shale lithologiesare thicker and more abundant in the eastern Rembang area. TheTuban sandstone lithology consists of subtidal to intertidal,bioturbated interbedded sandstone, siltstone and shale beds with
2
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
TEXT-FIGURE 1Geologic map of East Java showing the main stratigraphic units, main areas and the localities of Sr isotope samples.
geouser
Inserted Text
and Sharaf et al the AAPG bull paper
geouser
Inserted Text
; Sharaf et al aapg bull paper

larger benthic foraminifera, thin planar corals and mollusk frag-ments, specially in the upper parts of the section. The Tuban carbon-ates are well exposed forming an E-W ridge that allowed measuringmultiple stratigraphic sections. The Tuban carbonates are highlyfossiliferous and characterized by massive coral-rich beds, LBF-rich bedded shelf strata, and red-algal thick-bedded carbonates. TheTuban shale consists of massive, featureless, green shale rich inplanktonic foraminifera. The Tuban Formation is interpreted asa mixed carbonate-siliciclastic shelf with prograding deltasintertonguing with shelfal carbonates and buildups.
Ngrayong Formation
The Ngrayong Formation is well exposed in quarries and riverbanks along the Lodan Anticline and Prantakan River (Text-fig.1).The age of Ngrayong stratigraphic unit is Middle Miocene(Ardhana et al. 1993; Lunt et al. 2000). The exposed Ngrayongsuccession is rarely fossiliferous, however, this formation consists atthe base of argillaceous fine sand and shale that grades upward intointerbedded fine to medium sand with thin mudstones layers andcoal seams. The top of the formation consists of coarse and friablesandstone. The Ngrayong Formation is interpreted as progradingtidal delta.
Bulu Member of Wonocolo Formation
The Wonocolo Formation in the study area consists of a basal car-bonate (Bulu Member) overlain by a thick succession of shale andmarl with thin sandstone intervals. The Bulu Member forms a mas-sive and resistant carbonate bench (10-20m thick). Two mainlithologies are identified: well bedded carbonates rich in large ben-thic foraminifera and planar corals, and sandy fossiliferous carbon-ates. The Bulu Member truncates the underlying NgrayongFormation; throughout the study area, meter-scale channels filledwith conglomerates separates the two formations. The Bulu Mem-ber is Late Serravallian-Early Tortonain in age based on the pres-ence of Cycloclypeus annulatus (Ardhana et al. 1993; Lunt et al.2000).
METHODS
Seventeen stratigraphic sections were measured throughout the area.A total of 650 samples were collected from these sections, and 250samples were collected from isolated sites.
The paleontological contribution to this study includes the identifi-cation of larger benthic foraminifera (LBF) and planktonicforaminifera. The study of LBF is based on the study of 250 thin
3
Stratigraphy, vol. 2, no. 3, 2005
TEXT-FIGURE 2Summary of the stratigraphy and the proposed ages for the different stratigraphic units in EJB.

sections of carbonate samples from the Prantakan, Mahindu, andPrupuh areas. As the specimens studied here are from randomthin sections of limestones, biometric measurements on iso-lated, solid specimens of larger foraminifera have rarely beenpossible. We attempt to combine the broad results gained byequatorial sections of the megalospheric nepiont of theMiogypsinidae (as published by Drooger 1993) with those ob-tained by vertical sections of the whole test (following Tan SinHok 1936, 1937). Similarly, we combine equatorial sections ofthe megalospheric nepiont of the lepidocyclinids (as publishedby Chaproniere 1984; Van Vessem 1978) with those obtainedby vertical sections of the whole test (as made by Cole 1957,1963).
The study of planktonic foraminifera was done either in thin sec-tions or from washed residues. The washed residues were obtainedby separating the foraminifera from 10 shale and soft chalk samples.The samples were soaked in a solution of 10% hydrogen peroxide(H2O2) and sodium hexametaphosphate (Na6(PO3)6, Calgon) for24 hours and then the insoluble residue wet sieved with water using63 micron sieve and checked for the fossil content. The planktonicforaminifera were examined under the binocular microscope.
Selected LBF were micro-drilled for strontium isotope analysis.Primary LBF shell materials were carefully selected. Extra stepswere performed to insure mineralogical homogeneity of eachsample including: elimination of the diagenetically deformedforms and matrix, staining of the thin sections of the selectedcarbonate samples with Alizarin Red S and potassium ferri-cyanide to check their carbonate mineralogy and running quali-tative x-ray analysis of picked foraminifera from a few shalesamples selected for the strontium dating. Approximately 10 mgof the powdered samples were dissolved in 1mL 1M ultra pureacetic acid. (See Asahara et al. 1995; Quade et al. 1995; Singh etal. 1998 for further discussion on acid leaching techniques.) Thesamples were then dried down in 1mL 8M hydrochloric acid inpreparation for ion-exchange chromatography. Mass analysiswas completed on a thermal-ionization mass spectrometer at theUniversity of Wisconsin – Madison, using a multi-collectoranalysis with exponential normalization to 86Sr/88Sr = 0.1194.The 87Sr/ 86Sr ratios obtained were dated numerically usingthe standard tables of McArthur et al. (2001) based on the seawater Sr-isotope curve of the Tertiary listed in McArthur et al.(2001) and Hodell et al. (1999) and the geologic time scale ofGradstein, Ogg and Smith (2004). During the data collection for
4
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
TEXT-FIGURE 3Idealized stratigraphic framework of the different rock units in East Java Basin, Indonesia.
this study, 4 analyses of NBS-987 yielded 87Sr/86Sr values of0.710269 +/-0.000015 (2-standard deviations) n=406 for NISTSRM-987 and 0.709192 +/-0.000013 (2-standard deviations)n=7 for EN- 1. The curves for age calculation assume 0.710248for NIST SRM-987 and 0.709175 for EN-1. To correct theinterlaboratory bias and make the strontium data agree withMcArthur et al. (2001), the 87Sr/86Sr ratios were multiplied by0.9999725 which yielded corrected ratios of 0.710249 for theNIST SRM-987 standard and 0.709172 for EN-1.
BIOSTRATIGRAPHY
The first biostratigraphic study of the Neogene of Indonesia was re-ported by Martin (1880; cited by Brouwer 1925) using the “Molluskstratigraphy” method. This method was based on the Linnaean ra-tios of extant to extinct mollusk fauna to estimate the relative ages.However, this method had numerous shortcomings (Brouwer 1925)and failed to correlate between mollusk species in East IndianOcean and the species in Europe. Since the 1920s, the larger benthicforaminifera have become the most important biostratigraphicmethod for age determination in the Indo-Pacific region for theirlarge diversity and regional distribution. The Tertiary Letter Stage
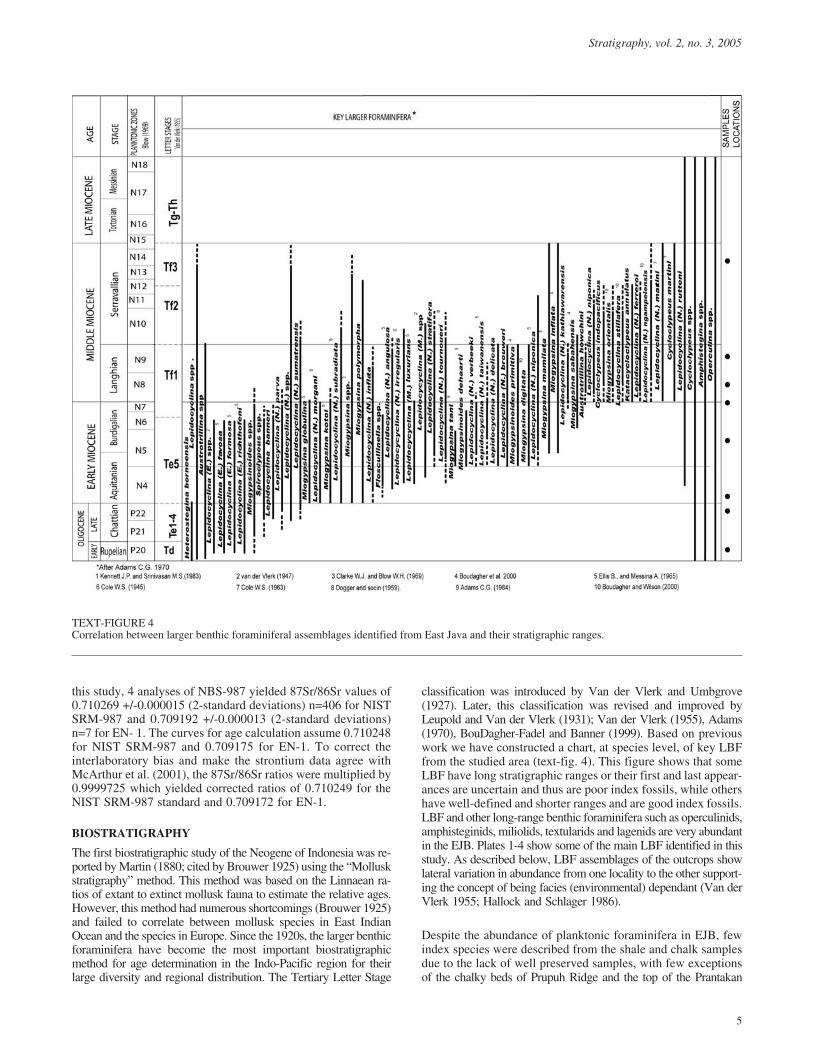
classification was introduced by Van der Vlerk and Umbgrove(1927). Later, this classification was revised and improved byLeupold and Van der Vlerk (1931); Van der Vlerk (1955), Adams(1970), BouDagher-Fadel and Banner (1999). Based on previouswork we have constructed a chart, at species level, of key LBFfrom the studied area (text-fig. 4). This figure shows that someLBF have long stratigraphic ranges or their first and last appear-ances are uncertain and thus are poor index fossils, while othershave well-defined and shorter ranges and are good index fossils.LBF and other long-range benthic foraminifera such as operculinids,amphisteginids, miliolids, textularids and lagenids are very abundantin the EJB. Plates 1-4 show some of the main LBF identified in thisstudy. As described below, LBF assemblages of the outcrops showlateral variation in abundance from one locality to the other support-ing the concept of being facies (environmental) dependant (Van derVlerk 1955; Hallock and Schlager 1986).
Despite the abundance of planktonic foraminifera in EJB, fewindex species were described from the shale and chalk samplesdue to the lack of well preserved samples, with few exceptionsof the chalky beds of Prupuh Ridge and the top of the Prantakan
5
Stratigraphy, vol. 2, no. 3, 2005
TEXT-FIGURE 4Correlation between larger benthic foraminiferal assemblages identified from East Java and their stratigraphic ranges.

River and Mahindu outcrop, where the beds are rich in planktonicassemblage of long geologic range such as Globigerina woodi,Globigerinoides sacculifer, Orbulina sp. and Orbulina suturalis.Plate 5 shows some of the main identified planktonic foraminiferafrom the studied area.
The Upper Oligocene, Lower Kujung, LBF assemblage is domi-nated by Spiroclypeus sp., Heterostegina borneensis, Lepido-cyclina (Eulepidina) favosa, L. (E.) richthofeni, L. formosa,Lepidocyclina banneri, Lepidocyclina sp., Miogypsinoides sp.,Miogypsinella boninensis (Plate 1) and the index planktonicforaminifera Globigerina ciperoensis described from a shale sam-ple near Dandu village, eastern Rembang area (text-fig. 1).
The Upper Oligocene-Lower Miocene (Chattian-Aquitanian,P22-N4, equivalent to Te1-4-Te5), Upper Kujung, is characterizedby an assemblage dominated by Lepidocyclina (Eulepidina)formosa, L. (E) favosa, L. (N.) parva, L. (N.) morgani, L. (N.)verbeeki, Lepidocyclina (Nephrolepidina) sumatrensis, Mio-gypsina sabahensis, Miogypsinoides dehaarti, Spiroclypeus sp.and Flosculinella sp. (pl. 1). The chalk beds are characterizedby a planktonic assemblage of Globoquadrina dehiscens (pl. 5,fig. 1), Globigerinoides quadrilobatus (pl. 5, fig. 2) andGlobigerinoides trilobus (pl. 5, fig. 3). Two shale samples col-lected along Prupuh Ridge are rich in Globorotalia kugleri andGlobigerinoides primordius.
The Burdigalian-Langhian (N5-N9, equivalent to upper Te5-Tf1),Tuban sandy carbonate beds are defined by Lepidocyclina(Eulepidina) formosa and Miogypsina (Miogypsina) tani. TheTuban carbonates are highly fossiliferous at the Prantakan area(text-fig. 1). The association at this area consists of Austro-trillina howchini, Lepidocyclina (Nephrolepidina) ferreroi, L. (N.)martini, Lepidocyclina (E.) sp., L. delicata, L. (N.) stratifera, L.(N.) inflata, L. (N.) angulosa, L. (N.) brouweri, L (N.) tournoueri, L.(N.) irregularis, L. (N.) kathiawarensis, Miogypsina digitata,Katacycloclypeus annulatus (pls. 2 and 3) . To the east, Mahinduarea (text-fig. 1), the dominant LBF’s are Lepidocyclina (N.)verbeeki, Miogypsina sp., Miogypsinoides sp., and Katacyclo-clypeus annulatus. And farther to the west (Dermawu village,text-fig.1) the association consists of Lepidocyclina (Nephro-lepidina) verrucosa, L. (N.) ferreroi, L. (N.) sumatrensis andKatacycloclypeus annulatus. The bed on top of the Tuban car-bonates (Mahindu area) contains the planktonic foraminifersGloborotalia praemenardii, Orbulina sp. and O. suturalis (pl.5, fig. 6)
The Late Serravallian assemblage (N12 and younger, equivalent tostage Tf2), Bulu, is characterized by dominance of Katacyclo-clypeus annulatus, Lepidocyclina (N.) ngampelensis (pl. 4),Orbulina sp. and Orbulina suturalis (pl. 5, fig. 6).
In this paper, we systematically describe significant benthiclarger foraminifera. We only listed the occurrences of theplanktonic foraminifera.
SYSTEMATIC DESCRIPTIONS
The usage of the subgeneric names Lepidocyclina (Lepidocyclina)and Lepidocyclina (Nephrolepidina), with the use of Lepido-cyclina s.l. for microspheric forms, follow the system proposed byBouDagher-Fadel and Banner (1997). The nomenclatural revi-sion of BouDagher-Fadel and Banner (1997) makes the genusLepidocyclina sensu lato available for the naming ofmicrospheric forms, but the megalospheric forms are divisibleinto the essentially Palaeogene Lepidocyclina (Lepidocyclina
L.), in which the protoconch and deuteroconch are of nearlyequal size and separated by a straight wall (this has been called“isolepidine”), and the essentially Miocene Lepidocyclina(Nephrolepidina) where the smaller proloculus is followed by amuch larger, reniform deuteroconch. The latter forms developquadrate proloculi in the later stages of many lineages.
Family LEPIDOCYCLINIDAE Scheffen 1932Subfamily LEPIDOCYCLININAE Scheffen 1932Subfamily Lepidocyclininae SCHEFFEN 1932Genus LEPIDOCYCLINA Gümbel 1870 emend. Boudagher-
Fadel and Banner 1997
Lepidocyclina banneri BouDagher-Fadel, Noad and Lord 2000Plate 1, figure 3
Lepidocyclina banneri BOUDAGHER-FADEL, NOAD, LORD 2000,p. 348, pl. 1, figs. 5-6.
Dimensions: Maximum measured length 4mm.
Remarks: This species is characterized by its possession of mas-sive pillars in the centrum of the strongly biconvex test. Much nar-rower hyaline pillars are scattered over the whole of the lateralsides of the test. L. banneri is broadly biconvex in form in compar-ison with L. delicata (Pl.3, fig.4).
Distribution: This species was originally described from theGomantong Limestone from the Upper Te, Aquitanian-Burdigalian of northwest Borneo. In this study it is found in theUpper Chattian of the Lower Kujung Formation in East Java,Kujung area Sample (6.3.3, 2001).
Lepidocyclina delicata Scheffen 1932Plate3, figure 4
Lepidocyclina delicata SCHEFFEN 1932, p. 18, pl. 1, fig. 4. –BOUDAGHER-FADEL and WILSON 2000, p. 153, pl. 1, fig. 5. –BOUDAGHER-FADEL et al. 2000b, p. 348, pl. 1, figs. 7-8.
Dimensions: Maximum measured length 20mm.
Remarks: Lepidocyclina delicata is characterised by the dark,very finely, microgranular pillars which are restricted to the in-ner lateral layers of the centrum only. Beyond these pillars,hyaline, glossy radial pillars are to be found radiating from theinner layers of the centrum to the outer surface.
Distribution: L. delicata Scheffen ranges from middle Tf1 toTf2 (Langhian to early Serravallian). It was first described fromJava, Indonesia. BouDagher-Fadel et al. (2000b) record the oc-currence of L. delicata in the Tf2 of the Darai Limestone in cen-tral south Papua-New Guinea. It has been found in Kalimantanwith Katacycloclypeus in the early Serravallian (Tf2) byBouDagher-Fadel and Wilson (2000) and from the Tf2(Serravallian) of Sadeng section (SAD) in the Gunung Sewu areaof South Central Java by (BouDagher-Fadel et al. 200B). In thisstudy, it was found in the Tf1 “letter stage” of Late Burdigalianto Langhian age of East Java, Gabalan outcrop, Rembang west,Sample # 1.
Lepidocyclina stratifera Tan Sin Hok 1935Plate 2, figure 1B
Lepidocyclina stratifera TAN SIN HOK 1935, p.9, pl. 1(4), figs. 1-3,pl.2(5), fig.11, pl.3(6), fig.9, pl. 4(7), fig.1, pl. 1, fig. 1.
Lepidocyclina (Nephrolepidina) stratifera Tan Sin Hok. – BARBERI etal. 1987, pl. 4, figs. 4,11. – BOUDAGHER-FADEL and WILSON
6
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
2000, p. 154, pl. 1, fig. 3, pl.2, fig. 6. – BOUDAGHER-FADEL 2002,p. 164, pl. 3, fig. 10.
Maximum measured length 4mm.
Remarks: L. stratifera has a biconvex test with many layers oflow cubiculae in which their platforms are as thick as or thickerthan the cubicular lumena. Club-shaped hyaline pillars are de-veloped from the outer periphery of the centrum to the surfaceof the test.
Distribution: L. stratifera was first described from Java, Indo-nesia being found together with Miogypsina, Katacycloclypeusand Trybliolepidina, an assemblage characteristic of middleSerravallian, Tf2 (see BouDagher-Fadel and Banner 1999).Barberi et al. (1987) reported L. stratifera from the Tf1(Burdigalian-Serravallian) of the carbonate sequence of the Is-land of Sumbawa, Indonesia. It was described by BouDagher-Fadel and Wilson (2000) in the Tf1 of late Burdigalian toLanghian age, of eastern Borneo, Kalimantan. BouDagher-Fadel (2002) recorded similar forms from the Tf1 (UpperBurdigalian) and Tf2 (Serravallian) of the Gunung Sewu area ofSouth Central Java. In this study it was found in the Tf1 “letterstage” of Late Burdigalian to Langhian age of East Java,Prantakan area, Sample (12.2.18, 2001).
Subgenus Eulepidina H. Douvillé 1911[EDITOR'S QUERY:
Please supply systematics (remarks, synonymy, etc.) for the fol-lowing three taxa.
Lepidocyclina (Eulepidina) favosa (Cushman)Plate 1, figure 8
Lepidocyclina (Nephrolepidina) tourneri Lemoine and DevillePlate 2, figure 4
Miogypsina spiralisPlate 2, figure 6
End Query, resume proof text]
Lepidocyclina (Eulepidina) formosa (Schlumberger)Plate 3, figure 2
Lepidocyclina formosa SCHLUMBERGER 1902, p. 251, pl. 7, figs. 1-3.
Dimensions: Maximum measured length 5mm.
Remarks: Lepidocyclina (Eulepidina) Formosa is characterizedby the four angles of the test prolonged into tapering rays.
Distribution: This species was originally described from theMiocene of Borneo. In this study it is found in the Lower Mio-cene deposits (Upper Te/Te5) of East Java. Sample 8.1.3.
Subgenus Nephrolepidina H. Douvillé 1911 emend. Bou-Dagher-Fadel and Banner 1997
Lepidocyclina (Nephrolepidina) ferreroi Provale 1909Plate 3, figure 1B, Plate 4, figure 2
Lepidocyclina ferreroi PROVALE 1909, p.70, pl.2, figs 7-13.Lepidocyclina (Nephrolepidina) ferreroi Provale. – BOUDAGHER-
FADEL and LORD 2000, pl.2, figs 2-9.
Dimensions: Maximum measured length 3.6mm.
Remarks: The species is characterized in having a quadrateprotoconch, a quadrilateral test, depressed medially but with amedian layer which has 4 high, pillared, lateral lobes and high,numerous cubiculae with relatively thin walls and.
Distribution: Lepidocyclina (Nephrolepidina) ferreroi with aquadrate protoconch were found in the Burdigalian-Langhian ofsouth east Kalimantan (pl.2, figs. 7, 8). In this study it is foundin the Tf1 “letter stage” of Late Burdigalian to Langhian age ofEast Java. West Rembang, Mahindu area, (GPS 115, 2002).
Lepidocyclina (Nephrolepidina) kathiawarensis Chatterji 1961Plate 2, figure 1A
Lepidocyclina (Nephrolepidina) kathiawarensis CHATTERJI 1961, p.429, pl. 2, fig. 9.
Dimensions: Maximum measured length 4mm.
Remarks: This species is characterized by having a bilocularembryonic apparatus typical of a tryliolepidine type.
Distribution: This species was first described from the UpperBurdigalian of western India. In this study it was found in the
7
Stratigraphy, vol. 2, no. 3, 2005
TABLE 1Strontium isotope analyses of selected samples in EJB, illustrated by decimal latitude and longitude coordinates, and their corresponding ages. The87Sr/86Sr ratios were normalized by multiplication by 0.9999725 which yielded corrected ratios of 0.710249 for the NIST SRM-987 standard and0.709172 for EN-1.

Tf1 “letter stage” of Late Burdigalian to Langhian age of EastJava. Sample 12.2.18.
Lepidocyclina (Nephrolepidina.) ngampelensis Gerth in Caudri1939Plate 6, figure 5
Lepidocyclina (Nephrolepidina) ngampelensis GERTH, in CAUDRI1939, p. 159, pl. 1, figs. 3-4.
Dimensions: Maximum measured length 5mm.
Remarks: This species is characterized by very low, thickfloored cubiculae, with strong pillars scattered along the centreof the test.
Distribution: This species was found in the “Tertiary” of Java.Similar forms occur in the Serravallian (Tf2) of the Darai Lime-stone in central south Papua-New Guinea (BouDagher-Fadel etal., in preparation). Our specimens were found in theSerravallian (Tf2) of East Java. Prantakan River Sample (2.3).
Lepidocyclina (Nephrolepidina) sumatrensis (Brady 1875)Plate 3, figure 3c, 6; plate 4, figure 4A
Orbitoides sumatrensis BRADY 1875 p. 536, pl. 14, fig. 3a-c.Lepidocyclina (Nephrolepidina) sumatrensis (Brady). – COLE 1957, p.
343, pl. 104, figs. 1-9, pl. 105, fig. 18, pl. 106, fig. 5, pl. 109, figs. 1-3.–CHAPRONIERE 1983, p. 41, pl. 3, figs. 11, 12, pl. 5, figs. 9-12, pl. 6,figs. 1-10. – CHAPRONIERE 1984, p. 66, pl. 10, figs. a-c, pl. 22, fig.14, pl. 23, figs. 1-7, pl. 26, figs. 15, 16, fig.21. – BARBERI et al. 1987,pl. 5, fig. 3, pl. 6, fig. 4. – BOUDAGHER-FADEL and WILSON2000, p. 156, pl. 2, fig. 4. – BOUDAGHER-FADEL et al. 2000b, p.352, pl.3, fig.2.
Dimensions: Maximum measured length 5mm.
Remarks: This species is characterized by a strongly biconvexspecies with a narrow equatorial flange and with many small,narrow pillars.
Distribution: Brady (1857) described L. (N.) sumatrensis fromthe “Early Tertiary” of Nias Island, west coast of Sumatra. Cole(1957) obtained his specimens from the Upper Te “LetterStage” of Saipan, Chaproniere (1983, 1984) from the Oligo-
cene-Miocene of Australia, while those of BouDagher-Fadeland Wilson (2000) were from the Tf1, upper Burdigalian toLanghian, of eastern Borneo, Kalimantan. The figured speci-mens of BouDagher-Fadel et al. (2000b) were from the Goman-tong Limestone, upper Upper Te, Burdigalian, of north Borneo.BouDagher-Fadel et al. (200B) recorded L. (N.) sumatrensisfrom the Tf1 (Upper Burdigalian) and Tf2 (Serravallian) ofRongkop section (SAD) in the Gunung Sewu area of South Cen-tral Java. In this study it is found in the Tf1 “letter stage” of LateBurdigalian to Langhian age of East Java. Sample (GPS 115,2002), East Dermawu, Sample 15, and Prantakan area, Sample(4.3.1, 2001).
Lepidocyclina (Nephrolepidina) verrucosa Scheffen 1932Plate 4, figure 4
Lepidocyclina verrucosa SCHEFFEN 1932, p. 33, pl. 7, figs. 2-4, p. 13,text-fig. 4. – BOUDAGHER-FADEL and WILSON 2000, p. 156, pl.2, figs. 7-8.
Dimensions: Maximum measured length 3.5mm.
Remarks: This species is characterized by possessing pairs ofpillars radiating from about the fifth layer of cubiculae to thesurface, the pillars diverging at an angle of about 60°.
Distribution: L. (N.) verrucosa was first described from Tf2 ofJava. BouDagher-Fadel and Wilson (2000) found similar formsin the Tf1 “letter stage”, upper Burdigalian to Langhian, of east-ern Borneo, Kalimantan and of the Tf1 of the Darai Limestonein Papua. In this study it is found in the Tf1 “letter stage” of LateBurdigalian to Langhian age of East Java. East Dermawu out-crop, (Sample #15).
Superfamily NUMMULITACEA de Blainville 1827Family CYCLOCLYPIDAE BouDagher-Fadel. 2002Genus KATACYCLOCLYPEUS Tan Sin Hok 1932
Katacycloclypeus annulatus (Martin 1880)Plate 1, figure 6; plate 2, figure 4; plate 4, figures 1, 6
Cycloclypeus annulatus MARTIN 1880, p. 157, pl. 28, figs. 1a-1i. –DOUVILLÉ 1916, p. 30, pl. 6, figs. 2,3 [not pl. 5, fig. 6, pl. 6, figs. 1-4).
8
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
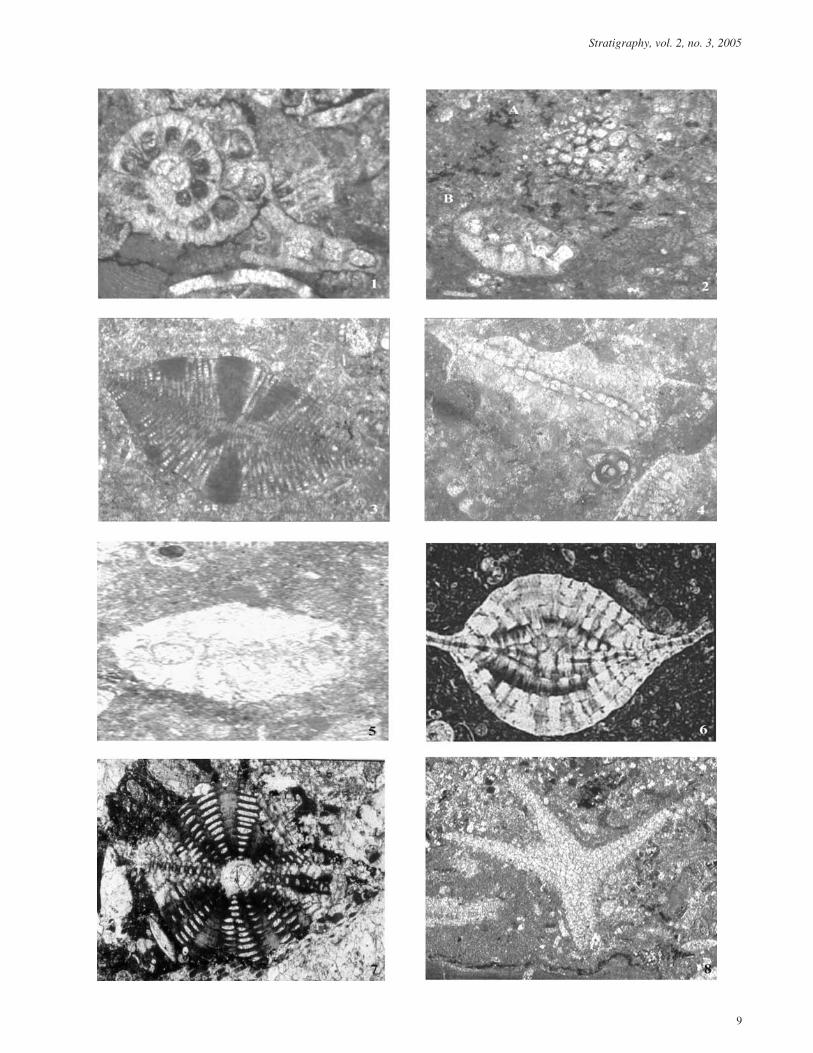
PLATE 1Thin section photomicrographs of Late Oligocene-Early Miocene larger benthic foraminifera
1 Miogypsinella boninensis Matsumaru 1996. Equato-rial slice of a megalospheric section. Sample 10.1.5.Dandu village, Kujung anticline. ×200.
2,5 Miogypsinoides dehaarti (Van Der Vlerk). A) verticalaxial section through the proloculus showing the solidlateral walls. B) equatorial section showing theproloculus surrounded by a whorl of embryonicchambers. 2. Sample 6.3.3; Dandu village, Kujunganticline, sample # 69, Prupuh section. ×50.
3 Lepidocyclina banneri BouDagher-Fadel, Noad andLord. Axial section showing massive pillars in thecenrum. Sample 6.3.3. ×100.
4 Miogypsina sabahensis BouDagher-Fadel, Lord andBanner. Axial section showing the stacks of cubiculaeseparated by massive pillars. Sample P1.69. ×100.
6 Spiroclypeus sp. Axial section. Sample # 42.8 Prupuhsection, ×50.
7 Lepidocyclina (Eulepidina) favosa (Cushman). Axialsection. Sample # 24.3, Prupuh section, ×50.
8 Lepidocyclina (Eulepidina) formosa (Schlumberger).Sample 8.1.3.
9
Stratigraphy, vol. 2, no. 3, 2005

Cycloclypeus (Katacycloclypeus) annulatus Martin - COLE 1963, p.E19,pl. 6, fig. 13, 14; pl. 7, fig. 7, pl. 8, figs. 4-6, 8-11, pl. 9, figs. 14, 17.
Katacycloclypeus annulatus (Martin). – BOUDAGHER-FADEL andWILSON 2000, p. 157, pl. 3, fig. 8, pl. 4, fig. 2. – BOUDAGHER-FADEL 2002, p. 168, pl. 3, fig. 1.
Dimensions: Maximum measured length up to 6mm (even on abroken specimen)
Remarks: This species is characterized by having a large, thintest with a central umbo surrounded by several widely spacedannular inflations of the solid lateral walls.
Distribution: K. annulatus was first described from the Early Mio-cene of West Java, Indonesia. Cole (1963) reported it from theTertiary of Guam and Fiji. Similar forms were found in theSerravallian, Tf2, of eastern Sabah and the Darai Limestone ofPapua New Guinea (BouDagher-Fadel et al., in preparation)while those of BouDagher-Fadel, and Wilson (2000) came fromthe lower Mid Miocene, Tf2 (early Serravallian) of Kalimantan.BouDagher-Fadel (2002) recorded this form the Lower Mio-cene of the Tacipi Formation, Indonesia. In this study similarforms occurred in the Tf2 (Lower Serravallian) of East Java,Prantakan outcrop (sample PR.2), Prantakan River, Sample(4.3.1, 2001) and Mahnidu area, Sample (9.1.2, 2001).
Family MIOGYPSINIDAE Vaughan 1929Genus MIOGYPSINELLA Hanzawa 1940
Miogypsinella boninensis Matsumaru 1996Plate1, figure 1
Miogypsinella boninensis MATSUMARU 1996, p.50, pl.5, figs 1-7; pl.6,figs 1-12; pl.7, figs 1-16; Fig.23-4. – BOUDAGHER-FADEL, LORDand BANNER 2000, p.144, pl.2, figs 1,2,4.
Dimensions: Maximum measured length 1mm.
Remarks: M. boninensis is characterised by having a biconvex testwith several umbilical plug-pillars. The embryonic chambers nearthe apex are followed by nepionic chambers disposed in a trochoidspire.
Distribution: This form was first described from the UpperOligocene of Japan. In this study, similar forms occur in the UpperOligocene, Lower Te, of North East Borneo (BouDagher-Fadel,Lord and Banner 2000). This species is here found in the UpperOligocene, Lower Te of East Java, Kujung area, Sample (10.1.5,2001).
Genus MIOGYPSINOIDES Yabe and Hanzawa 1928
Miogypsinoides dehaarti (Van Der Vlerk 1924)Plate1, figure 2, 5
Miogypsina dehaarti Van Der Vlerk 1924, p. 429-431, text-fig. 1-3.Miogypsinoides dehaarti (Van Der Vlerk). – COLE 1957, p. 339, pl. 111,
figs. 5-16. – VAN DER VLERK 1966, pl. 1, figs. 1-6, pl. 2, figs. 1-3.Miogypsina (Miogypsinoides) dehaarti Van Der Vlerk. – RAJU 1974, p.
80, pl. 1, figs. 19-25; pl. 3, fig. 8; pl. 4, figs. 2-4. – BOUDAGHER-FADEL, LORD and BANNER 2000c, p. 145, pl. 2, fig. 5.
Dimensions: Maximum measured length 1.5mm.
Remarks: M. dehaarti is characterized by having very thick lateralwalls and being smooth exteriorly, sometimes with slightly raiseddark pillars of finely granular structure.
Distribution: This species was first described from the LowerMiocene of Larat, Netherlands East Indies. An equatorial sec-tion of an ideotype was photographed by Van Der Vlerk (1966,pl.2, fig.1). RAJU (1974) registered the occurrence of this spe-cies in the Indopacific and Mediterranean regions.BouDagher-Fadel et al. (2000c) found similar forms in the Up-per Te, Aquitanian-Burdigalian, of North East Borneo, and isknown in Papua New Guinea to range up into Tf1 (EarlyLanghian). This species is here found in the Lower Miocene ofEast Java, Lower Aquitanian (Lower Te5). Prupuh outcropSample 1.69.
Genus MIOGYPSINA Sacco 1893
Miogypsina digitata Tan Sin Hok 1937Plate 3, figure 3A
Miogypsina (Miogypsina) kotôi Hanzawa forma digitata TAN SIN HOK1937, p. 101, pl. 2, figs. 1-5, fig. 1a.
10
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
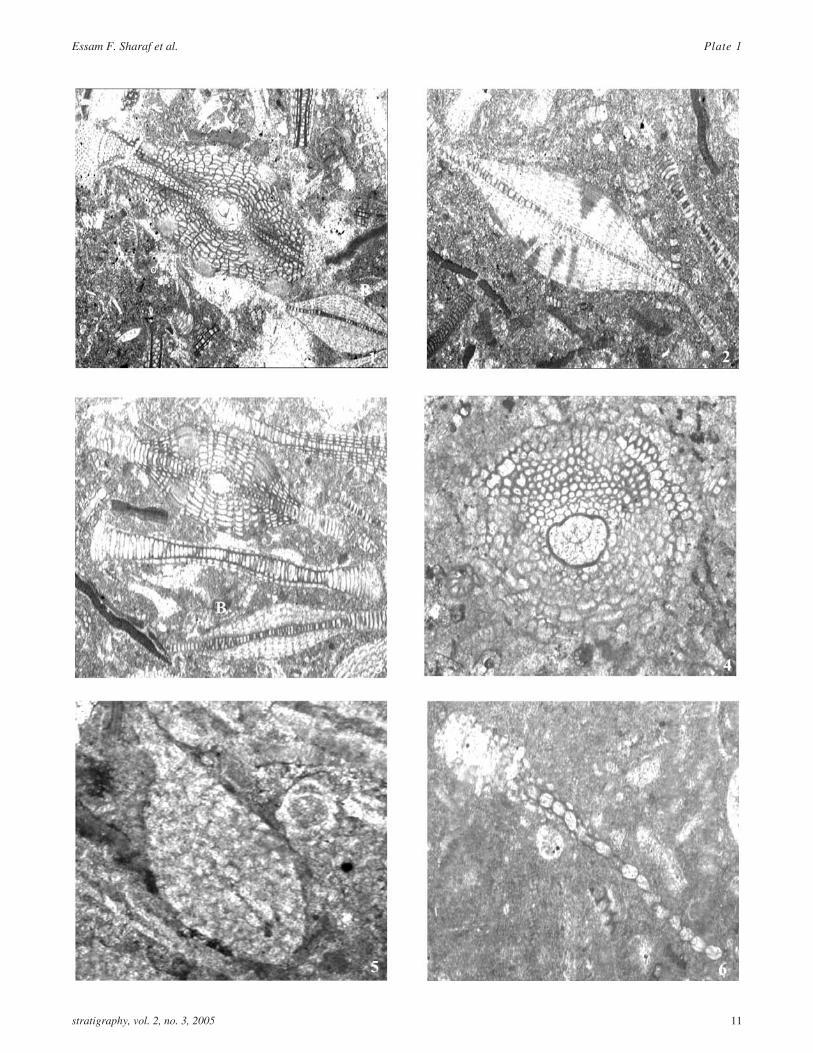
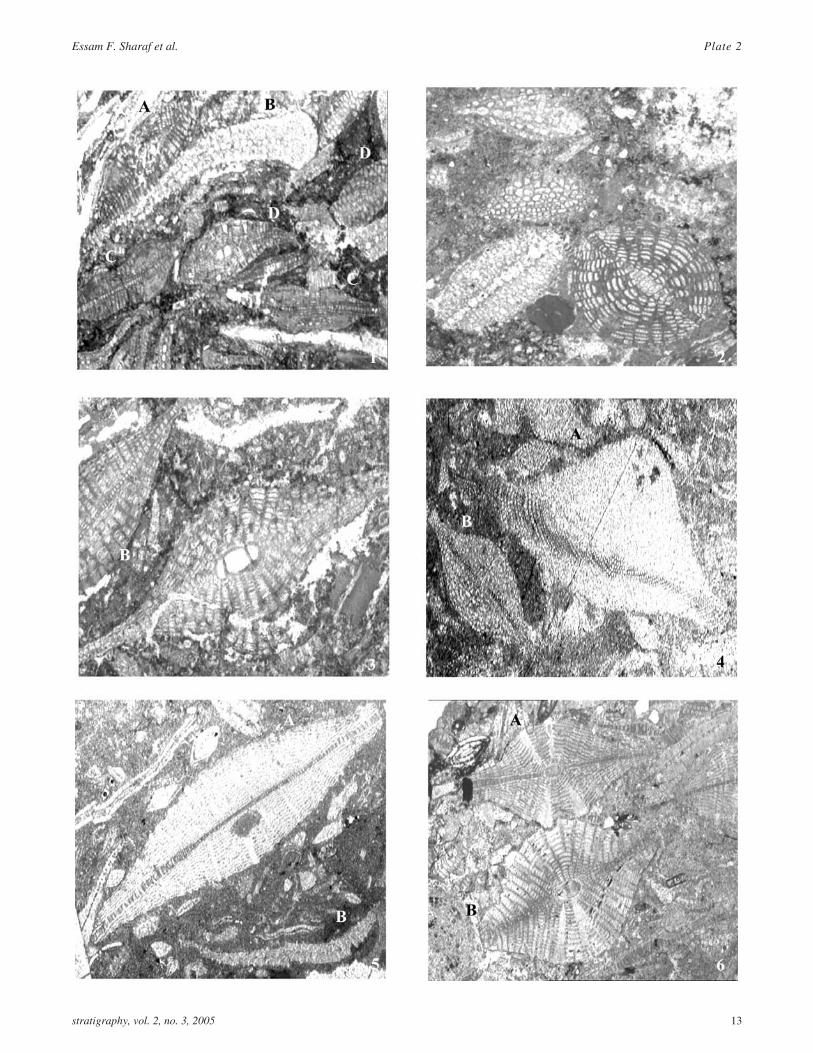
PLATE 2Thin section photomicrographs of Early-Middle Miocene (Burdigalian-Langhian) larger benthic foraminifera
1 A) Lepidocyclina (Nephrolepidina) kathiawarensisChatterji, oblique equatorial section showing aquadrate protoconch strongly embraced by adeuteroconch. Pillars are present only towards the pe-riphery in our specimens, however towards both endsof the test. B) Lepidocyclina stratifera Tan Sin Hok.Sample 12.3.18. Prantakan section. ×100.
2 Lepidocyclina stratifera Tan Sin Hok. Sample12.3.18. Prantakan section ×50.
3 A) Lepidocyclina (Nephrolepidina) verrucosa(Scheffen). A) An axial section showing pillars di-verging at an angle of about 60o. B) Lepidocyclina
stratifera Tan Sin Hok. Sample 12.3.18. Prantakansection. ×50.
4 Lepidocyclina (Nephrolepidina) tourneri (Lemoineand Douvillé). Sample # 2, Prantakan section. ×150.
5 Miogypsina kotoi Hanzawa. Vertical axial section.Sample 4.3.1. Prantakan River area. ×50
6 Miogypsina spiralis Rutten. Axial section showing alarge embryonic structure. Sample 11.2. 11. Mahinduarea. ×50.
stratigraphy, vol. 2, no. 3, 2005 11
Essam F. Sharaf et al. Plate 1

Miogypsina digitata Tan Sin Hok. – BOUDAGHER-FADEL, LORDand BANNER 2000c, p.146, pl.3, fig.7.
Dimensions: Maximum measured length 1mm.
Remarks: This form is characterized by having oval median cham-bers and numerous fine pillars.
Distribution: It was found by BouDagher-Fadel et al. (2000c) inthe early Burdigalian, upper Te, of North East Borneo. In thisstudy it is found in the Tf1 “letter stage” of Late Burdigalian ofEast Java. Sample 4.3.1.
Miogypsina kotoi Hanzawa 1931Plate 2, figure 5, plate 3, figure 3B
Miogypsina kotoi HANZAWA 1931, p.154, pl.25, figs 14-18. –BOUDAGHER-FADEL and WILSON 2000, p.157, pl.3, figs 1-2.
Dimensions: Maximum measured length 2.5 mm.
Remarks: This species is distinguished in having oval medianchambers, small thick-walled, but strongly convex cubicula; it hasa biserial nepiont which is strongly asymmetrical.
Distribution: Miogypsina kotoi was first described from theBurdigalian of Japan. BouDagher-Fadel and Wilson (2000) re-ported it from the Tf1 late Burdigalian to Langhian, of easternBorneo, Kalimantan. BouDagher-Fadel and Lokier (in press) re-corded similar forms from the Tf1 (Upper Burdigalian) and Tf2(Serravallian) of Djatirago (TAN) the Gunung Sewu area ofSouth Central Java. In this study it is found in the Tf1 “letterstage” of Late Burdigalian to Langhian age of East Java.Prantakan River, Sample (4.3.1, 2001).
Miogypsina sabahensis BouDagher-Fadel, Lord and Banner 2000Plate1, figure 4
Miogypsina sabahensis BOUDAGHER-FADEL, LORD and BANNER2000c, p. 147, pl.3, figs 4-6.
Dimensions: Maximum measured length 1.6mm.
Remarks: This species is characterized by having massive andheavy pillars when seen in vertical section.
Distribution: M. sabahensis was first described from the UpperTe of Burdigalian age of eastern Sabah and in the Tf1 (ofBurdigalian-Langhian age) of Kalimantan, both in North EastBorneo (BouDagher-Fadel et al. 2000c). This species is foundhere in the Lower Miocene of East Java, Te5. Prupuh outcrop,Sample 1.69.
Miogypsina tani Drooger,1952Plate 3, figure 1B
Miogypsina (Miogypsina) tani Drooger 1952, p. 26, 51, 52, pl. 2, figs.20-24. – RAJU 1974, p. 82, pl. 1, figs. 26-30; pl. 5, fig. 5. –WILDENBORG 1991, p. 113, pl. 4, figs. 1, 2, tabs. 19, I. –BOUDAGHER-FADEL, LORD and BANNER 2000c, p.147, pl.3,figs 4-6.
Dimensions: Maximum measured length 3mm.
Remarks: This species is distinguished by having a longmegalospheric nepionic coil of auxiliary chambers, low cubiculaewith inflated roofs and many scattered pillars.
Distribution: This species was first described from the so called“Middle Oligocene” of east-central Costa Rica. It was also foundin the Aquitanian stage, Upper Te, of Italy and southern Spain(Raju 1974). It has a wide distribution in the American, Mediterra-nean and Indopacific regions. Similar specimens were found byBouDagher-Fadel et al. (2000c) in the upper Upper Te,Burdigalian, of Borneo. In this study it is found in the Lower Tf1“letter stage” of Late Burdigalian of East Java. West Rembang,Mahindu area, sample (GPS 115, 2002).
STRONTIUM ISOTOPE CHRONOSTRATIGRAPHY
Strontium isotope chronostratigraphy was used to calibrate thenumerical ages of the exposed Oligocene-Miocene outcrops inEJB. Because the ranges of most LBF are relatively long andtheir abundance is a facies dependant, strontium isotope ratioswere used to help correlate the stratigraphic horizons. Table 1and Text-figures 2 and 3 summarize the proposed ages for eachstratigraphic unit throughout the studied area. The GPS loca-tions of the studied samples are converted to decimal latitudeand longitude co-ordinates and plotted on Table 1.
12
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
PLATE 3Thin section photomicrographs of Early-Middle Miocene (Burdigalian-Langhian) larger benthic foraminifera
1 Thin section photomicrograph of larger benthicforaminifera assemblage A). Lepidocyclina sp., B)Miogypsina tani Drooger. C) Lepidocyclina (Nephro-lepidina) ferreroi Provale. D) Axial and equatorialsections of Cycloclypeus sp. Sample GPS #115,Mahindu area. ×50.
2 Thin section photomicrograph of A) Miogypsinadigitata Drooger. B) Miogypsina kotoi Hanzawa. C)Lepidocyclina (Nephrolepidina) sumatrensis(Brady). Sample 4.3.1. Prantakan River. ×50.
3 Lepidocyclina (Nephrolepidina) sumatrensis(Brady). Axial section. GPS 115, Mahindu area. ×50.
4 Lepidocyclina delicata Scheffen, A) subequatorialsection showing a subcircular equatorial peripheryand broadly hexagonal median chamberlets. Gabalansection. Sample #1. ×100.
5 A) vertical axial section. Lepidocyclina (Nephro-
lepidina) subradiata (Douvillé). B) Miogypsina taniDrooger. ×50.
6 A) Lepidocyclina (Nephrolepidina) sumatrensis(Brady). B) Lepidocyclina (Nephrolepidina) verru-cosa (Scheffen). East Dermawu section. Sample #15.×50.
stratigraphy, vol. 2, no. 3, 2005 13
Essam F. Sharaf et al. Plate 2

Strontium isotope data from the lower Kujung Formation, the oldeststrata exposed in the EJB (Table 1), provide an age range of 28.78+0.74 to 28.20+ 0.74 Ma corresponding to Early to Late Oligocene,latest Rupellian- earliest Chattian respectively, (P21), and it is con-sistent with the presence of Heterostegina borneensis, Lepido-cyclina banneri, L. sp., L. formosa., Miogypsinoides sp., andMiogypsinella boninensis.
The index planktonic foraminifera Globigerina ciperoensis fromthe middle Kujung shale/chalk indicates correlation with Zone ofP22 for this interval. Strontium isotope dating of two samples fromthe upper Kujung (Sukowati village and west of Dandu village,Table 1) give ages of 23.44+ 0.74 and 24.31+ 0.74 (LateOligocene), correlating with Zone (P22-N4). For this interval, thebenthic foraminifera assemblage consists of Lepidocyclina (E)formosa, L. (E.) favosa, L (N.) sumatrensis, Miogypsinasabahensis, Miogypsinoides dehaarti corresponding to theAquitanian stage, and the planktonic foraminifera assemblage con-tains Globorotalia kugleri, Globigerinoides primordius, Globo-quadrina dehiscens, Globigerinoides quadrilobatus, Gdesprimordius and Globigerinoides trilobus.
The oldest sandy carbonate unit exposed of Tuban Formation has anage of 20.80 + 0.74 Ma (Table 1). Tuban carbonates yield an age of20.17+0.74 (base of Burdigalian, N5) to 15.25 +1.36 (base ofLanghian, N9). This is consistent with the association ofAustrotrillina howchini, Lepidocyclina (Nephrolepidina) ferreroi, L.(N.) martini, L. delicata, L. (N.) stratifera, L. (N.) inflata, L. (N.)angulosa, L. (N.) brouweri, L. (N.) tournoueri, L. (N.) irregularis, L.(N.) kathiawarensis, L. (N.) verbeeki, Miogypsina digitata andKatacycloclypeus annulatus. Two samples separating the Tubancarbonates from the Ngrayong Formation yield ages of 15.34 +1.36and 15.25+1.36 (Table 1). This is consistent with the appearance ofKatacycloclypeus annulatus, Orbulina sp. and Orbulina suturalistowards the very top of the Tuban carbonates. Although Orbulinasuturalis (pl. 5) and Orbulina sp. have long stratigraphic ranges,their first appearance datum is in upper Langhian (Zone N9 of Blow1969) indicating that the top of the exposed Tuban carbonates is asyoung as Late Langhian in age.
The Bulu Member from the base of the section at Prantakan River(Table 1) reveals an age of 12.98+1.36 Ma corresponding to theSerravallian age (N12, Table 1). This age is consistent with theage range of the Katacycloclypeus annulatus, Lepidocyclina
(Nephrolepidina) ngampelensis, Orbulina sp. and Orbulinasuturalis that are dominant at this stratigraphic unit.
DISCUSSION
Oligocene-Miocene sedimentary deposits of EJB are dominatedby warm water, shallow marine carbonates that are of crucialimportance as the product and record of climatic/oceanic condi-tions and interchange in tropical/subtropical inner-outer shelfenvironments. These deposits are dominantly biogenic in ori-gin, primarily of larger foraminfera and algae, with hermatypiccorals. Fluctuations in climate or oceanic currents have influ-enced the geographic distribution of the LBF, during this periodand abundance of LBF in the EJB appears to be facies dependant. InEast Java Basin LBF and planktonic foraminifera overlap in oc-currence in shelf and hemipelagic sediments allowing directcomparison of LBF zones with oceanic plankton scales. LBF arecommon to very abundant in facies dominated by corals in theLower Kujung, Tuban and Bulu carbonates. LBF are rare in skeletalgrainstones (Lower Kujung), red algae dominated facies (Tuban)and sandy carbonates (Tuban and Bulu). LBF are almost absent inthe Middle Kujung chalk, Tuban clastics, Ngrayong sand andWonocolo shale. LBF are abundant as reworked clastics in UpperKujung turbidites.
In these facies the occurrence of the LBF together with theplanktonic foraminifera, indicate the presence of strong currentstransporting the LBF towards deeper water marine waters in theopen shelf area (BouDagher-Fadel 2002). Accurate age datingof the studied sections is corroborated by Sr isotope datingwhere appropriate. This allowed us to revise the stratigraphicranges of some of theses LBF.
In correlating the LBF with the occurrence of the planktonicforaminifera and comparing the data with the strontium isotopedata, we were able to conclude that the LBF stratigraphic rangechart (text-fig. 4) is consistent with the ages obtained fromstrontium dating (text-fig. 2).
CONCLUSIONS
The age of exposed Lower Kujung Formation is late EarlyOligocene, Rupelian P20-Late Oligocene,Chattian, P21 respec-tively) based on identified foraminifera and strontium dating.The faunal assemblage in Prupuh Ridge area indicates that theexposed Upper Kujung (shale, chalk and turbidites) are rich in
14
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
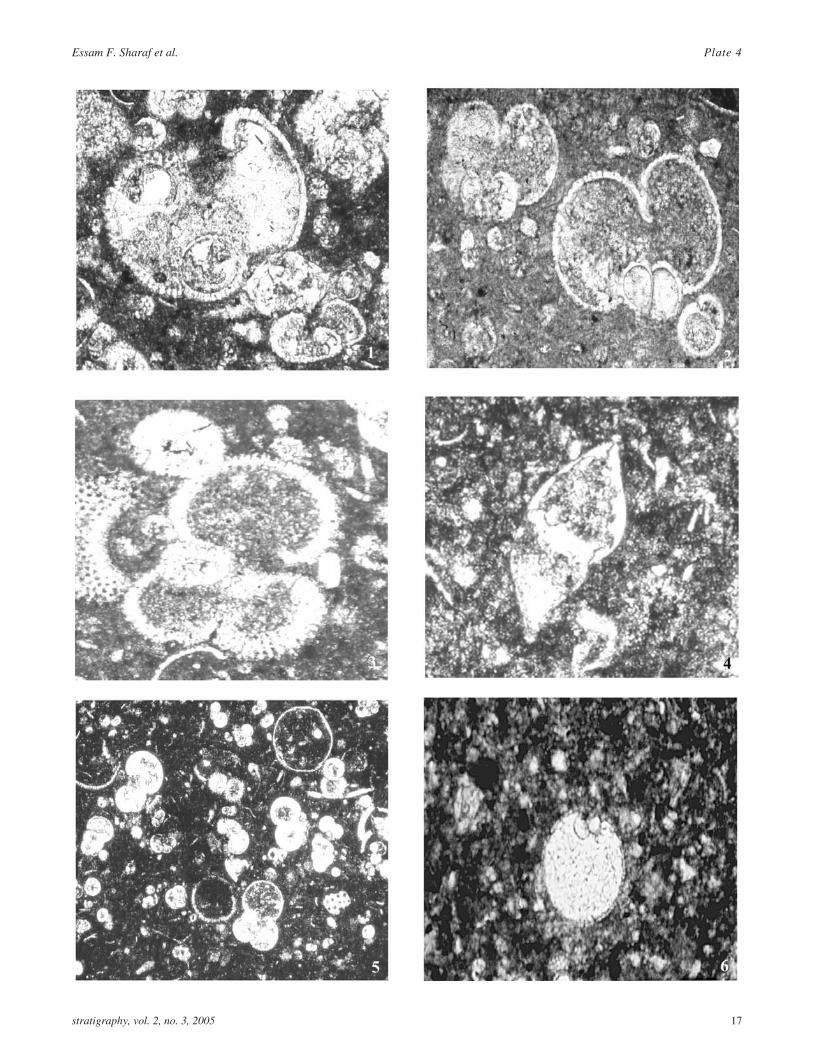
PLATE 4Thin section photomicrographs of Early-Middle Miocene (Burdigalian-Serravallian) larger benthic foraminifera
1,6 Thin section photomicrographs of Katacyclocypeusannulatus Martin. 1. Equatorial sections. Sample PR171. 6. Vertical section, Sample 9.1.2. Mahindu area.×200.
2 Thin section photomicrograph of Lepidocyclina(Nephrolepidina) ferreroi Provale. Prantakan area,GPS #150, 2001. ×50
3 Lepidocyclina (Nephrolepidina) ngampelensis Gerth.Kembang Baru section.Sample #10. ×150.
4 Lepidocyclina (N.) ferreroi Provale, Axial section,Sample GPS 150, 2001, Prantakan area, ×50.
4,6 Katacycloclypeus annulatus Martin. Axial section.Prantakan River. Sample #2. 4X50, 6 Sample 99.12×60.
5 Katacycloclypeus annulatus Martin. A) Fragments ofequatorial sections. B) Fragment of an axial section.Prantakan River section .Sample #3.1. ×150

stratigraphy, vol. 2, no. 3, 2005 15
Essam F. Sharaf et al. Plate 3

larger benthic and planktonic foraminifera with stratigraphicrange of Early Miocene Aquitanian age (Te5) which is equiva-lent to Zone N4 of Blow (1969). Strontium isotope dating of thePrupuh carbonates exposed at Sukowati village and along thewestern side of Prupuh Ridge gives an age of Late Oligocene(Chattian) to Early Miocene (Aquitanian) equivalent to ZonesP22 to N4 of Blow (1969). The Tuban outcrops have a longstratigraphic range from Upper Te5-Tf1 (Burdigalian-Langhian), which is equivalent to Zones N5-N9 of Blow(1969). The studied carbonate beds from Mahindu andPrantakan outcrops confirmed the presence of Orbulina sp. andOrbulina suturalis suggesting an age of late Langhian (N9) forthe top of the Tuban. This is consistent with the age constrainedfrom strontium dating of the shale unit at the top of Prantakanoutcrop.
The Bulu Member of the Wonocolo Formation is characterized byfaunal assemblage of late Mid –Late Miocene age (Serravallian–Tortonian). The strontium dating of the Bulu carbonates exposedat Prantakan River is consistent with the age range obtained fromthe LBF.
The co-occurrence of both coralgal benthic planktonicforaminifera rocks in the carbonate facies of the Oligocene andMiocene of EJB is a rare opportunity for correlating thebiostratigraphic framework of this region. These organismtypes although controlled by environment would have influ-enced production rates on the reef and platform.
ACKNOWLEDGMENTS
We would like to thank the Indonesian Institute of Technologyin Bandung (Prof. Lambok and Dardji), and specially NukiNugroho for providing field assistance. We are grateful to Dr.Clay Kelly, Department of Geology and Geophysics, Univer-sity of Wisconsin-Madison, Joseph Serra, University of Barce-lona, and Peter Lunt , Coparex Blora, Indonesia for their help inidentification of foraminifera and for their valuable recommen-dations. Martin Shield’s help in the field and was key to havinga better understanding of the EJB history. We want to thankProf. C.M. Johnson, the director of thermal-ionization massspectrometer lab at the University of Wisconsin-Madison, Dr.Brian Beard and in particular, Brooke Swanson for the stron-tium isotope analyses. We would like to thank the Micro-palaeontology Unit at UW-Madison for use of photographicfacilities. This work was funded by student grants from GSA,AAPG, and Department of Geology and Geophysics, Univer-
sity of Wisconsin-Madison. Exxon Mobil and Anadarko haveprovided partial support to this research. The strontium analyseswere funded by the donors of the Petroleum Research Fund ofthe American Chemical Society.
REFERENCES
ADAMS, C.G., 1970. A reconsideration of the Indian Letter classifica-tion of the Tertiary. Bulletin of the British Museum (Natural History)Geology, 19, 85-137
ADAMS, C.G., 1984. Neogene larger foraminifera, evolutionary andgeological events in the context of datum planes. In: Ikebe, I. andTuschi, R., Eds., Pacific Neogene Datum Planes, p. 47-68. Univer-sity of Tokyo Press.
ARDHANA, W., LUNT, P. and BURGON G.E., 1993. The deep marine
sand facies of the Ngrayong Formation in the Tuban Block, East Java
Basin, 118-175. Indonesian Petroleum Association, Clastic CoreWorkshop.
ASAHARA, Y., TANAKA, T., KAMIOKA, H., and NISHIMURA, A.,1995, Asian continental nature of 87Sr/86Sr ratios in north centralPacific sediments. Earth and Planetary Science Letters, 133:105-116.
BEMMELEN, R.w. van, 1949. The Geology of Indonesia. The Hague:Martinus Nijhoff, 732p.
BERGGREN, W.A., KENT, D.V., SWISHER, III, C.C., and AUBRY,M.P., 1995. A: revised Cenozoic geochronology and chronostrati-graphy. In: W. A. Berggren, D. V. Kent, M.P. Aubry and J.Hardenbol, Eds., Geochronology, time scales and global strati-
graphic correlation. SEPM Special Publication, 54: 129-212.
BOLLI, H.M., 1957. Planktonic foraminifera from Oligocene-MioceneCipero and Lengua Formations of Trinidad, B.W.I., U.S National
Museum Bulletin 215: 97-123.
———, 1966. The planktonic foraminifera in well Bojonegoro-1 ofJava. Eclogae Geologicae Helvetiae, 59 (1): 449-465.
BLOW W. H., 1969. Late Middle Eocene to recent planktonicforaminiferal biostratigraphy. Proceedings of the First International
Conference on Planktonic Microfossils Geneva, 1967, 1: 199-422.
BOUDAGHER-FADEL, M.K. and WILSON, M.E.J., 2000. A revisionof some larger foraminifera from the Miocene of East Kalimantan.Micropaleontology, 46: 153-165.
16
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia
PLATE 51 Globoquadrina dehiscens (Cushman, Parr and Col-
lins).Prupuh section. Sample # 35. ×150.
2 Oblique thin sections of two specimens of Globi-gerinoides quadrilobatus (d’Orbigny). Prupuh sec-tion. Sample # 35. ×250.
3 Globigerinoides trilobus (Reuss). Prupuh section.Sample # 35. ×150
4 Globorotalia praemenardii (Cushman and Stain-forth). Northern Gunung Manak area, Sample 3.9.1.×100.
5 Thin section photomicrographs of planktonicforaminifera: Globigerinoides spp., and Globigeri-noides quadrilobatus (d’Orbigny) . Prupuh section.Sample # 37. ×100.
6 Orbulina suturalis Brönnimann. Prantakan area, GPS# 150, 2001. ×200.
stratigraphy, vol. 2, no. 3, 2005 17
Essam F. Sharaf et al. Plate 4

BOUDAGHER-FADEL, M.K. 2002. The relationships between plank-tonic and larger benthic foraminifera, Middle Miocene to LowerPliocene facies of Sulawesi. Micropaleontology, 48:153-176.
BOUDAGHER-FADEL, M.K. and BANNER, F.T., 1997. The revisionof some genus-group names in Tethyan Lepidocyclininae. Paleo-
pelagos, 7: 3-16.
BOUDAGHER-FADEL, M.K., and BANNER, F.T 1999. Revision ofthe stratigraphic significance of the Oligocene –Miocene “ let-ter-Stages”. Revue de micropaléontologie, 42: 93-97.
BOUDAGHER-FADEL, M.K., and LORD A.R., 2000. The evolutionof Lepidocyclina (L.) isolepidinoides, L.(Nephrolepidina) nephro-
lepidinoides sp. Nov., L.(N.) brouweri and L. (N.) ferreroi in the LateOligocene-Miocene of the Far East. Journal of Foraminiferal Re-
search, 30(1): 71-76.
BOUDAGHER-FADEL, M.K., LORD, A.R., and BANNER, F.T.,2000. Some Miogypsindae (foraminifera) in the Miocene of Borneoand nearby countries. Revue paléobiologie, 19 (1): 137-156.
BRANSDEN, P.J.E. and MATTHEWS, S.J., 1992. Structural and strati-graphic evolution of the East Java Sea. Indonesia. Proceedings Indo-nesian Petroleum Association, 21st Annual Convention, I: 417-453.British Museum (Natural History), Geology Bulletin, 19: 85-137.
BROUWER, H.A., 1925. The geology of the Netherlands East Indies.
London: The Macmillan Company, 160p.
BROUWER, J. 1966. Stratigraphy of the younger Tertiary in
North-East Java and Madura. The Hague: Bataafse International Pe-troleum Maatschappij N.V.
CAROZZI, A., 1976. Microfacies and microfossils of the Miocene reefcarbonates of the Philippines. Philippine Oil Development Com-
pany, Inc., Special Publication, No.1, 80p.
CHAPRONIERE G.C.H., 1984. Oligocene and Miocene largerForaminiferida from Australia and New Zealand, Bureau of Mineral
Resources, Australia, Bulletin, v. 188, p.1-98.
COLE, W.S., 1957, Larger Foraminifera. U.S. Geological Survey Pro-
fessional Paper, 280-I, pt. 3, p. 321-360.
———, 1963. Tertiary larger foraminifera from Guam. U.S. Geological
Survey Professional Papers , 403-E: E1-E28.
COLE, J., M., and CRITTENDEN, S., 1997. Early Tertiary basin forma-tion and the development of lacustrine and quasi-lacustrine/marinesource rocks of the SundaShelf. In: A.J., Fraser, S.J., Matthews andR.W., Murphy, Eds., Petroleum Geology of Southeast Asia. Geolog-ical Society of London Special Publication, 5: 147-183.
DARMAN, H., and SIDI, H., 2000. An outline of the geology of Indone-
sia. Indonesian Association of Geologists (IGAI), 192 p.
DROOGER, C.W., 1993. Radial foraminifera; morphometrics and evolu-tion. Koninklijke Nederlandse Akademie van Wetenschappen,Am-
sterdam, 41: 1-241.
DUYFJES, J., 1936. Zur Geologie und Stratigraphie des Kendeg-
gebietes zwischen Trinil und Soerabaya (Java). Nederlandsch-Indiee, Dienst Mijnbouw, Section IV: p. 136 -149.
———, 1938. Geologishe Kaart van Java Toelichting bij Blad 109
(Geologic Map of Java, 1:100,000 explanatory notes to sheet 109),Lamongan.
———, 1938. Geologishe Kaart van Java Toelichting bij Blad 110
(Geologic Map of Java, 1:100,000 explanatory notes to sheet 110),Lamongan.
ELLIS, B. A., and MESSINA, A.R. 1965. Catalogue of index
foraminifera. The American Museum of Natural History. SpecialPublication, 1, Lepidocyclinids and Miogypsinids.
GRADSTEIN, F. M., OGG , J.G. and SMITH A.G., 2004. A Geologic
Time Scale. Edited by Felix M. Gradstein. New York: CambridgeUniversity Press, 610 p.
HALLOCK, P. and SCHLAGER, W., 1986. Nutrient excess and the de-mise of coral reefs and carbonate platforms. Palaios, 1: 389-398.
HAMILTON, W., 1979. Tectonics of the Indonesian region.US Geolog-ical Survey Professional Paper No.1078, 345 p.
HODELL, D.A., MUELLER P.A. and GARRIDO, J.R., 1991. Varia-tions in the strontium isotopic composition of seawater during theNeogene. Geology, 19: 24-27.
JOINT OPERATING BODY PERTAMINA- TREND TUBAN(JOB.P-TT), 1990. “East Java Field Work Report.” Unpublished re-port.
KOESOEMADINATA, R.P.K. and PULUNGGONO, A., 1975. Geol-ogy of the southern Sunda Shelf in reference to the tectonic frame-work of Tertiary sedimentary basins of Western Indonesia.Association of Indonesian Geologists, 2 (2): 1-11
LUNT, P. 2001. Introduction to the geology and biostratigraphy of the
Tuban Plateau. website: http://nummulites.net/Geo/NE_Java.pdf
LUNT, P., NORTHERWOOD, R. and BURGON, G., 2000. AAPG/IAPfield trip to Central Java, October, 2000. !!!!!PUBLISHER!!!!
LEUPOLD, W., and VLERK VAN DER, I.M. 1931. Stratigraphie vanNederlandsch Oost-Indie. Leidsche Geologische Mededelingen, 5:611-648.
MATSUMARU, K., 1977. Biostratigraphy and paleoecological transi-tion of larger foraminifera from the Minamizaki limestone, Chi-chi-Jima, Japan. Geological Research and Development Centre,
Special Publication, No.1: 63-88.
MATTHEWS, S.J. and BRANSDEN, P., J.E., 1995. Late Cretaceousand Cenozoic tectonostratigraphic development of East Java Sea Ba-sin, Indonesia. Marine and Petroleum Geology,12:499-510.
MAXWELL, W., G., H., 1968. Atlas of the Great Barrier Reef. Amster-dam: Elsevier, 258 p.
MCARTHUR, J. M., HOWARTH, R. J. and BAILEY, T. R., 2001.Strontium isotope stratigraphy: LOWESS Version 3: Best fit to themarine Sr-isotope curve for 0-509 Ma and accompanying look-up ta-ble for deriving numerical age. Journal of Geology, 109: 155-170.
MUHAR, A., 1956. Micropaleontoloisch onderzoek Van monstersofkomstig Van het geoloisch onderzoek Tuban (unpublished report).
NAJOAN, G.A.S. 1972. Correlation of the Tertiary lithostratigraphicunits in the Java Sea and adjacent areas. Proceedings of Indonesian
Petroleum Association, June: 11-30.
NOAD, J., 2001. The Gomantong Limestone of eastern Borneo; asedimentological comparison with the near-contemporaneousLuconia Province. Paleogeography, Palaeoclimatology, Palaeoec-
ology, 175; 1-4: 273-302
PRINGGOPRAWIRO, H., SOEHARSONO,N., and SUJANTO, F.X.,1977. Subsurface Neogene planktonic foraminifera biostratigraphy
of North-West Java Basin. Geological Research and DevelopmentCenter special publication.1:125-165.
PETROLEUM REPORT OF INDONESIA, 2002. Website:http://www.usembassyjakarta.org/econ/petro2002-toc.htm
18
Essam F. Sharaf et al.: Biostratigraphy and strontium isotope dating of Oligocene-Miocene strata, East Java, Indonesia

QUADE, J., CHIVAS, A.R., and MCCULLOCH, M.T., 1995. Stron-tium and carbon isotope tracers and the origins of soil carbonate inSouth Austral ia and Victoria . Palaeogeography,
Palaeoclimatology, Palaeoecology, 113: 103-117.
RENEMA , W., 2002. Larger foraminifera as marine environmental in-dicators. Scripta Geologica, 124: 263p.
SINGH, S.K., TRIVEDI, J.R., PANDE, K., RAMESH, R., and KRISH-NASWAMI, S., 1998, Chemical and strontium, oxygen, and carbonisotopic compositions of carbonates from Lesser Himalaya: Implica-tions to the strontium isotope composition of the source waters of theGanga, Ghagara, and the Indus rivers. Geochimica et Cosmochimica
Acta, 62: 743-755.
SOEPARJADI, R.A., NAYOAN, G.A., BEDDOES, L.R. and JAMES,W.V., 1975. Exploration play concepts in Indonesia. Proceedings of
the 9th World Petroleum Congress, London.3: 51-64.
SYSTEMATIC GEOLOGICAL MAP OF INDONESIA, 1997. Geo-
logical map of the Tuban Quadrangle, Jawa. 1:100,000, GeologicalResearch and development Centre, Surabaya.
TAN SIN HOK, 1936. Zur Kenntniss der Miogypsiniden. Ingenieur
Nederland.-Indië, Mijnbouw Geologie, 4(3): 45-61.
———, 1937. Weitere Untersuchungen über die Miogypsiniden; IIp.Ingenieur Nederland.-Indië, Mijnbouw Geologie, 4(6): 35-45.
VESSEM.E. J. VAN, 1978. Study of Lepidocyclinidae from SoutheastAsia, particularly from Java and Borneo. Utrecht Micropaleonto-
logical Bulletins, 19, 163 pp.
VLERK, I.M. VAN DER and UMBGROVE, J. H. F., 1927. Tertiairegidsforaminiferen van Ned.Oost-Indie. Wetenschappelijke Mede-
delingen Dienst van den Mijnbouw in Nederlandsch-Indië, 6: 1-31.
VLERK, I.M. VAN DER, 1955. Correlation of the Tertiary of the FarEast and Europe. Micropaleontology 1(1): 72-75.
———, 1963. Biometrical research on Lepidocyclina. Micropale-
ontology, 9: 425-426.
———, 1966. Miogypsinoides, Miogypsina, Lepidocyclina, andCycloclypeus of Larat, Moluccas. Eclogae Geologicae Helvetiae,
59(1): 421-429.
VLERK, I., M., VAN DER and POSTUMA, J.A., 1967. Oligo-MioceneLepidocyclinas and Planktonic foraminifera from East Java andMadura, Indonesia. Koninklijke Nederlandse Akademie van
Wetenschappen, Amsterdam, Proceedings, ser. B (76): 245-259.
Manuscript receivedManuscript accepted
19
Stratigraphy, vol. 2, no. 3, 2005
Related Documents











