IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO. 1, JANUARY 1981 1113 Biophysics of Geomagnetic Field Detection A. J. KALMIJN Abstract-In biology, the studyof geomagnetic orientation has gained new momentum since the discovery of magnetic field detectors in aquatic organisms. Sharks and rays respond to dc and low frequency voltage gradients of 0.005 pV/cm. By moving through the earth’s magnetic field, they induce electric fields well within the sensitivity range of their keen electric sense. As these fields depend on the direc- tion in which the animal is heading, the induced voltage gradients may serve as the biophysical basis of an electromagnetic compass sense. The ability of sharks and rays to orient to the earth’s magnetic field has been demonstrated in behavioral experiments. Also, various marine and freshwater mud bacteria are endowed with permanent magnetic dipole moments, directed parallel to the axis of motility. When sepa- rated from the sediments, these bacteria return to the mud by migrating downward along the earth’s inclined magnetic field lines. Their orienta- tion is largely determined by the principles of statistical mechanics and may be expressed in terms of the directive magnetic force, the randomizing effect of thermal agitation, and the cells’ flagellar thrust. Observations on live bacteria yield individual dipole moments circa 15 X kT/G. A INTRODUCTION LTHOUGH the principles of motional-electric and ferro- magnetic orientation date back to the early days of physics, their biological relevance remained a matter of specu- lation until very recently. Acceptance of the electromagnetic principle hinged on the unusually high electrical sensitivity required. For ferromagnetic orientation, it was crucial to find the necessary biomagnetic material. This paper relates the discovery of an acute electric sense in elasmobranch fishes and the revelation of highly ordered permanent magnetic dipoles in mud bacteria. The findings pertain directly to the question of geomagnetic orientation, for both elasmobranchs and mag- netic bacteria show remarkable abilities in this regard. The discussion of whether the same principles apply to other or- ganisms is expected to arouse a renewed interdisciplinary interest in thestudyof animal orientation, navigation, and homing. Elasmobranchs Dijkgraaf s observations on the behavior of sharks in response to metal objects led theauthor to investigate the electrical sensitivity of elasmobranch fishes [l] . Both sharks and the kindred skates showed transient cardiac decelerations when submitted to uniform square-wave fields of 5 Hz, the skates at voltage gradients as low as 0.01 pV/cm [2] . The electric field Manuscript received March 18, 1980; revised September 15, 1980. This work was supported by the Office of Naval Research, Oceanic Biology Program, under Contract NOOO14-79-C-0071. Paper presented at the 1980 INTERMAG Conference, Boston, MA, April 21-24. The author is with the Woods Hole Oceanographic Institution, Woods Hole. MA 02543. receptors were identified as the ampullae of Lorenzini [3], [4]. However, the heartbeat-as well as observed eye-blink and ventilatory reflexes-conveyed littleinformation on the significance of the fishes’ electrical sensitivity. It was only after finding dc and low frequency bioelectric fields in the vicinity of aquatic animals, including the prey of elasmobranch fishes, that more meaningful responses were recorded [2] , [5]. The fields of prey animals appeared to be detectable to small sharks and skates up to a distance of 25 cm [6]. It was dem- onstrated that sharks and skates, when hungry or aroused by odor, sharply cue in on their prey electrically, even when the victim is hiding under sand. Apparent feeding responses were also obtained by passing electrical currents between two buried electrodes establishing prey-simulating fields with dipole mo- ments of $4 pA X 5 cm (dc to 8 Hz) at a seawater resistiv- ity of 19-26 L? cm. The electrical aspects of predation have recently been verified on free-roaming sharks in the shallow waters off Cape Cod, MA [7], [8] . Larger scale fields of purely physical origin are induced by ocean currents flowing through the earth‘s magnetic field. Their voltage gradients range from about 0.05-0.5 pV/cm when measured with towed electrodes [9] and may inform the animals of their passive drift in tidal and open-ocean currents [SI, [6]. In training experiments, marine stingrays have re- vealed their ability not only to detect these fields, but also to appreciate the direction and polarity, and to orient with respect to these parameters at field strengths of 0.01 pV/cm and less [8], [lo] . By the same principle, when swimming through the earth‘s magnetic field, the fishes induce electric fields of their own. These additional self-induced voltage gradients contain all physical cues marine elasmobranchs may need for anactive electromagnetic compass sense [6], [ll] . The stingrays’ability to direct themselves relative to the earth’s magnetic field has been ascertained in behavioral experiments PI , DO].. Mud Bacteria After Blakemore’s discovery of mud bacteria that respond to the approach of small bar magnets, he and the author jointly established the organisms’ directed migration in strictly uni- form fields of geomagnetic strength [ 121 , [ 131 . In Woods Hole, MA, bacteria collected from marine marshes and fresh- water swamps were observed to swim consistently north when separated from the sediments. Upon reversal of the ambient magnetic field, they executed U turns, several cell diameters wide, therebyturningintothe direction of the field again. Thus it was postulated that these bacteria are endowed with permanent magnetic dipole moments, pointing into the direc- tion of motility and passively aligning the organisms with the

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO. 1, JANUARY 1981 1113
Biophysics of Geomagnetic Field Detection A. J. KALMIJN
Abstract-In biology, the study of geomagnetic orientation has gained new momentum since the discovery of magnetic field detectors in aquatic organisms. Sharks and rays respond to dc and low frequency voltage gradients of 0.005 pV/cm. By moving through the earth’s magnetic field, they induce electric fields well within the sensitivity range of their keen electric sense. As these fields depend on the direc- tion in which the animal is heading, the induced voltage gradients may serve as the biophysical basis of an electromagnetic compass sense. The ability of sharks and rays to orient to the earth’s magnetic field has been demonstrated in behavioral experiments. Also, various marine and freshwater mud bacteria are endowed with permanent magnetic dipole moments, directed parallel to the axis of motility. When sepa- rated from the sediments, these bacteria return to the mud by migrating downward along the earth’s inclined magnetic field lines. Their orienta- tion is largely determined by the principles of statistical mechanics and may be expressed in terms of the directive magnetic force, the randomizing effect of thermal agitation, and the cells’ flagellar thrust. Observations on live bacteria yield individual dipole moments circa 15 X kT/G.
A INTRODUCTION
LTHOUGH the principles of motional-electric and ferro- magnetic orientation date back to the early days of
physics, their biological relevance remained a matter of specu- lation until very recently. Acceptance of the electromagnetic principle hinged on the unusually high electrical sensitivity required. For ferromagnetic orientation, it was crucial to find the necessary biomagnetic material. This paper relates the discovery of an acute electric sense in elasmobranch fishes and the revelation of highly ordered permanent magnetic dipoles in mud bacteria. The findings pertain directly to the question of geomagnetic orientation, for both elasmobranchs and mag- netic bacteria show remarkable abilities in this regard. The discussion of whether the same principles apply to other or- ganisms is expected to arouse a renewed interdisciplinary interest in the study of animal orientation, navigation, and homing.
Elasmobranchs Dijkgraaf s observations on the behavior of sharks in response
to metal objects led the author to investigate the electrical sensitivity of elasmobranch fishes [l] . Both sharks and the kindred skates showed transient cardiac decelerations when submitted to uniform square-wave fields of 5 H z , the skates at voltage gradients as low as 0.01 pV/cm [2] . The electric field
Manuscript received March 18, 1980; revised September 15, 1980. This work was supported by the Office of Naval Research, Oceanic Biology Program, under Contract NOOO14-79-C-0071. Paper presented at the 1980 INTERMAG Conference, Boston, MA, April 21-24.
The author is with the Woods Hole Oceanographic Institution, Woods Hole. MA 02543.
receptors were identified as the ampullae of Lorenzini [3], [4]. However, the heartbeat-as well as observed eye-blink and ventilatory reflexes-conveyed little information on the significance of the fishes’ electrical sensitivity. It was only after finding dc and low frequency bioelectric fields in the vicinity of aquatic animals, including the prey of elasmobranch fishes, that more meaningful responses were recorded [2] , [ 5 ] . The fields of prey animals appeared to be detectable to small sharks and skates up to a distance of 25 cm [ 6 ] . It was dem- onstrated that sharks and skates, when hungry or aroused by odor, sharply cue in on their prey electrically, even when the victim is hiding under sand. Apparent feeding responses were also obtained by passing electrical currents between two buried electrodes establishing prey-simulating fields with dipole mo- ments of $ 4 pA X 5 cm (dc to 8 Hz) at a seawater resistiv- ity of 19-26 L? cm. The electrical aspects of predation have recently been verified on free-roaming sharks in the shallow waters off Cape Cod, MA [7], [8] .
Larger scale fields of purely physical origin are induced by ocean currents flowing through the earth‘s magnetic field. Their voltage gradients range from about 0.05-0.5 pV/cm when measured with towed electrodes [9] and may inform the animals of their passive drift in tidal and open-ocean currents [SI , [ 6 ] . In training experiments, marine stingrays have re- vealed their ability not only to detect these fields, but also to appreciate the direction and polarity, and to orient with respect to these parameters at field strengths of 0.01 pV/cm and less [8], [lo] . By the same principle, when swimming through the earth‘s magnetic field, the fishes induce electric fields of their own. These additional self-induced voltage gradients contain all physical cues marine elasmobranchs may need for an active electromagnetic compass sense [ 6 ] , [l l] . The stingrays’ ability to direct themselves relative to the earth’s magnetic field has been ascertained in behavioral experiments P I , DO]..
Mud Bacteria After Blakemore’s discovery of mud bacteria that respond to
the approach of small bar magnets, he and the author jointly established the organisms’ directed migration in strictly uni- form fields of geomagnetic strength [ 121 , [ 131 . In Woods Hole, MA, bacteria collected from marine marshes and fresh- water swamps were observed to swim consistently north when separated from the sediments. Upon reversal of the ambient magnetic field, they executed U turns, several cell diameters wide, thereby turning into the direction of the field again. Thus it was postulated that these bacteria are endowed with permanent magnetic dipole moments, pointing into the direc- tion of motility and passively aligning the organisms with the

1114 IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO, 1 , JANUARY 1981
ambient field. The permanent magnetic nature of the response was revealed by subjecting the bacteria to strong magnetic pulses of short duration (200-800 G, 1-2 ps zero-to-peak), antiparallel to an ambient field of 0.5 G [I41 , as originally suggested by E. M . Purcell. Thus stimulated, the cells executed U turns similar to those described above, although now turning against the direction of the field. That is, the pulses lastingly changed the bacteria’s tendency from north-seeking to south- seeking, as one would expect from reversal of the intrinsic dipole moments. From the equal widths of the U turns elicited by reversal of either the ambient field or the intrinsic dipole moments, the bacteria appeared to behave magnetically as single-domain dipole particles. The dipole moments are due to endogenous chains of tightly coupled single-domain magnetite crystals, oriented nearly parallel to the cell axis as defined by flagellar insertion (Fig. 4) [12] , [I51 . The total dipole mo- ment, estimated from the number and size-range of the mag- netite grains, would-in dead cells at least-be sufficient to overcome thermal agitation, in conformity with the postulated orientation mechanism, As for the biological significance of the response, at Woods Hole the earth’s steeply inclined mag- netic field lines guide these microaerophylic organisms, when stirred up from the bottom sediments, back to the mud in which they thrive [ 121 , [ 141 .
PHYSICS OF ORIENTATION MECHANISMS The physics of geomagnetic field detection in elasmobranch
fishes and magnetic mud bacteria are rather different in nature. The motional-electric principle traces back to Faraday [16] , who not only gave the first coherent description of the various induction phenomena, but also mentioned their possible use for open-ocean navigation. The oceanographic aspects of electromagnetic induction have been addressed in particular by Longuet-Higgins et al. [ 171 . The biological implications of Faraday’s ideas for the study of animal orientation have been worked out by the author [ 6 ] , [I I ] as part of his research on the electric sense of elasmobranch fishes. Lodestone and magnetized compass needles were known for their directive properties more than a millennium ago. They were instrumen- tal to the discovery of the earth‘s magnetic field; cf., Gilbert [I81 . Through Amp6re’s imaginative reasoning, the compass needle became an integral part of the classical theory of elec- tricity and magnetism. Langevin’s [ 19 J statistical mechanics approach and Weiss’ domain theory refined AmpBre’s concept. The magnetic bacteria have manifested the biological relevance of these earlier studies [ 121 , [ 141 . Elasmobranchs
A shark cruising at a velocity v through the earth’s magnetic field % induces a voltage gradient v X fd as implied by Faraday’s law. Interaction with the horizontal component of the earth’s magnetic field %h yields a vertically directed voltage gradient v X $h, the magnitude of which depends on the speed of swimming, the strength of the magnetic field, and-most importantly-on the sine of the angle from v to gh; i.e., on the animal’s compass heading. With the shark swimming from west to east, the induced voltage gradient is in the positive ventrodorsal direction. That is, the animal acts as a source of
Fig. 1. Shark swimming through earth’s magnetic field induces electric field giving animal its compass heading. (After Kalmijn, 1974.)
electromotive force (EMF)
S=%(.X$,) .da,
the internal resistance of which is determined by the vertical extent of the animal and the average resistivity p of the skin and body tissues (Fig. 1).
The induced EMF gives rise to a current field of density 9, directed ventrodorsally within the animal and completing its circuit through the seawater environment relative to which the animal moves. Along current lines, a voltage gradient -pQ develops according to Ohm’s law. Thus the resulting potential difference across the animal is given by
With v rectilinear, the induced voltage gradient v X $h is in- herently uniform. The ohmic voltage gradient - p j would be uniform also if the skin and body tissues were homogeneous in resistivity, and the animal was of ellipsoid shape. For the present discussion one may picture it that way without adverse consequences (Fig. 2).
The potential difference f$ between the dorsal and ventral surfaces of the animal depends on the resistance of the body (internal part of the circuit) relative to the resistance of the surrounding seawater (external part of the circuit). Regarding the geometry of the internal and external parts, the animal’s resistance is expected to be 1-2 times that of its aquatic envi- rons. Furthermore, the average resistivity of skin and body tissues is 10-100 times the resistivity of seawater, say 20 C2 cm. That means that the animal is heavily loaded electrically, and within the body
dr dr ’ - p 9. da- -in (v X EBh) da.
vn (3)
Hence the potential difference $J between the dorsal and ven- tral surfaces of the animal tends to zero.
Sharks have several hundred electroreceptive organs, known as the ampullae of Lorenzini. Each sense organ consists of a small pore in the skin, giving access to a jelly-filled canal which ends in a blind sensory swelling: the ampulla proper. While the pores are distributed over most of the head, the ampullae proper are clustered in capsules, mainly located in the protrud- ing snout. From the capsules, the ampullary canals radiate in practically all directions. While the jelly in the canals is almost

KALMIJN: BIOPHYSICS O’F GEOMAGNETIC FIELD DETECTION 1115
Fig. 2. Cross section of shark heading east With velocity u in magnetic field of horizontal intensity $h. Two sense organs represent all those having ventrodorsal component. Field lines indicate current density 9. To right, potential distribution @ along ampullary axes o 1 anda2. (After Kalmijn, 1974).
as conductive as seawater, the wall of the ampullary canals has the highest mural resistance known of animal tissues [20]. Thus the organs form almost ideal electrically open-ended “submarine cables,” allowing virtually no dc and low frequency losses to occur along their jelly-filled canals. The receptor cells bridge the wall of the ampullae proper, sensing the poten- tial differences between the ampullary jelly and the capsule stroma in which the organs are embedded. In short, the am- pullae of Lorenzini offer high ohmic voltage-sensitive devices for the detection of dc and low frequency electric fields [6].
How may sharks apply these marvelous sense organs to the detection of their magnetic compass heading? Fig. 2 depicts two ampullary organs representing all the ones having a ventro- dorsal component. Since the animal as a source of motional electricity is heavily loaded, the potentials at the skin pores do not appreciably differ from each other and from those in the tissues around the ampullae proper, indicated by the circuit null. In the blind, well-insulated ampullary canals, however, the induced voltage gradients v X !i$.rernain uncom- pensated, as -pJ is vanishingly small. They lead to sizable potential differences between ampullae proper and skin pores, which measure by good approximation
amp 1 pore 2
-lorn 1 s,,, 2 (v X 53h)-& and - (v X $ h ) - d q .
(4) These then are the stimuli appearing across the sehsory epithe- lia, signaling the animal its compass direction (at given swim- ming speed I v i ) . With the fish heading east, the ampullary potentials are as indicated in Fig. 2; with the fish heading west, they are of opposite polarity. At all times, the signals are pro- portional to the sine of the angle from v to 53h. At a cruising speed of 1 m/s and ahorizontal magnetic component of 0.25 G, the amplitude of the stimulus gradient tends to 0.25 pV/cm. Given a threshold sensitivity of 0.01 pV/cm, this value demon- strates the physical feasibility of the proposed orientation mechanism.
One may also arrive at these results following a more rigor- ous, but perhaps conceptually less lucid, approach. Integration along a closed path from ventral to dorsal pore through the two ampullae (ha) and back from dorsal to ventral pore through
the tissues (+) yields
J^_dr I:‘ (v x $h - p y ) * dhU + (v x 9 h - p j) d&, = 0. (5)
For uniform v, the v X !Bh terms cancel. J - p J da,, the ohmic potential difference across the sensory epithelia (ne- glecting the losses in the capsule stroma and along the jelly- filled ampullary canals), equals J - p J * dht, the ohmic po- tential difference across the animal’s body. Fundamentally, v X !Bh is not accessible to the animal [6]. However, by mea- suring the integral of - p ~ across the body, elasmobranchs may estimate the integral of v X $h on the basis of (3).
On close examination, one may note an apparent ambiguity in the electromagnetic orientation mechanism, as swimming northeast and southeast, for example, leads to the same elec- trical stimuli. However, while swimming, sharks normally swerve alternately right and left, thereby “scanning” the in- duced field over a certain arc. As these scanning motions give rise to response patterns that for the northeast and southeast directions are mirror images of each other, the animals may discriminate between the two without further norlelectrical information (Fig. 3). The dynamics of magnetic field detec- tion are of great importance as the sense organs are not true dc but very low frequency ac receptors with a time constaat of several seconds. In prey detection and in the orientation to voltage gradients of inanimate origin, sharks also rely pre- dominantly on dc fields which they detect by moving in the appropriate way [ 6 ] .
Up to this point, only the interaction with the horizontal component of the earth’s magnetic field 91h has been taken into account. By the same token, interaction with the vertical component 53, stimulates the ampullae having their canals parallel to the fish‘s left-right axis. The total field induced is obtained by vector addition of the fields resulting from the individual magnetic components. The fields induced by inter- action with the vertical component may be used a) to monitor swimming speed I v I as part of the electromagnetic orientation process, and b) to detect geomagnetic field inclination, in- structing the animal about the latitude of its position on the globe.
In conclusion, through their keen electric sense, sharks,

1116 IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO. 1, JANUARY 1981
W-
s Fig. 3. Shark swimming northeast or southeast induces ventrodorsal
EMF E’; while swimming northwest pr southwest it induces dorso- ventral EMF E-. Changes 8’ and 6- in EMF, due to sweiving right and left, ake mirror images of each other about east-west axis. Thus swim- ming normally, shark may gain unambiguous information on its magnetic compass heading.
Fig. 4. Bacterium migrating in magnetic field 9. Rate of migratiori in direction of field equals swimmirig speed along axis of motility times cosine of angle of deviation. N and S: poles of magnetite string. (Bacterium not to scale.)
skates, A d rays may gain all the information necessary for geomagnetic field orientation. Whether they are able to, and indeed do, use the earth’s magnetic field as ari orientational cue are questions only the fishes can answer by expressing themselves behaviorally in response to magnetic fields of suit- able design.
Mud Bacteria As free-moving dipoles, magnetic bacteria are subject to
a) the directive force of geomagnetic field alignment and b) the randomizing effect .of thermal agitation. They thereby re- semble the inanimate dipole particles in Langevin’s theory of magnetization.
The bacteria under study propel themselves unidirectionally by flagellar action. Swimming at constant forward speed, their rate of migration in the direction of the field is a measure of how well they follow the field lines (Fig. 4). More precisely, the meari rate of migration ( V ) equals the average cosine of the angle of deviation ( cos 8 ) times the speed of swimming VO :
( v ) = ( COS e ) v,. (6)
This concept originally emerged from discussions between C. P. Bean and the author and has been further developed in
partial collaboration with B. D. Teague and M. K. Gilson [21] . The great merit of the approach is in its obvious ties with Langevin’s theory of magnetization, which gives (cos €0 as a function of the intrinsic magnetic dipole moment m, the am- bient magnetic field strength B, and the thermal energy kT. The Langevin concept is tested by timing the migration rates of individual bacteria as a function of ambient magnetic field strength. The experiments reveal the statistical mechanics nature of the bacteria’s orientation and yield excellent esti- mates of the cells’ magnetic dipole moments.
Under conditions of thermal equilibrium, the probability density function p(B) for the angle of deviation 8 of a magnetic moment m in magnetic field 53 is proportional to the Boltz- mann factor:
p ( e ) = Ae-@/kT = Aeffl .$/kT = AemB cos OIkT , (7)
with CP = - m 43 = -mB cos 8 the potential energy, and kT the thermal energy of the dipole particle. The constant A is deter- miried by the normalizing condition that the total probability of finding the dipole oriented in any direction equals unity.
For the average cosine of 0 it follows that
I” cos ,g p B cos 6lkT d a (COS 8) = COS e p(e ) da = ( 8 )
with dh the element of, the solid angle corresponding to all dipole orientations between 8 and 0 + dB. For d a one may write 2n sin 0 dB. The denominator is then readily evaluated by substitution of -d cos 0 for sin 6’ dB. The numerator is just the partial derivative of the denominator with respect to mBlkT. Thus after some calculation, one obtains
(9)
which is known as the Langevin function L. For a statistical analysis of the data, the variance or mean
squared deviation of the Ldgevin distribution Will also be needed, which is by definition
U’ = ((COS e - (COS e) )2) = (cos2 e) - (cos eY . (1 0)
In (lo), (cos 8)’ may be recognized as the Langevin function (9) squared. (cos2 0 ) equals (cf. (8))
in which the expression within braces is the Langevin function again. Substitution of (1 2) and the square of (9) into (10) gives

KALMIJN: BIOPHYSICS OF GEOMAGNETIC FIELD DETECTION 1 1 1 7
o2 = 1 - coth2 mBlkT + 1 (mBlkT)2 ’
which is the Langevin variance. Before the Langevin theory is applied, it will be necessary
first to review its underlying assumptions. a) In its original form, the Langevin function deals with an
ensemble of identical independent dipole particles in thermal equilibrium. The bacteria may be sufficiently independent, but their dipole moments actually differ from one another, often by as much as a factor of two [22] . Also, their forward thrust, although individually quite constant, varies appreciably from cell to cell. As the migration rate as a function of B is nonlinear in its parameters Vo and m, it would be incorrect to test the Langevin concept on ensembles of bacteria, not to mention the, practical difficulties in estimating the average migration rates of cell aggregates. This suggests that bacteria should be tracked singly.
b) In individual bacteria, the statistics are applied not to a distribution of simultaneous orientations of many specimens, but to the distribution of successive orientations of the one bacterium under study. By this approach, the requirement of identical particles is met, but the orientations are no longer independent of each other, unless they are sampled at time intervals long enough to ensure randomization (within the con- straints of the Langevin distribution). Thus the data obtained are analyzed on the assumption of a constant randomization time At characteristic of each individual bacterium. The valid- ity of this assumption may be judged by the magnitude and constancy of the experimentally determined values of At.
c) The motility of the bacteria is on the one hand used as a means of measuring the organisms’ average cosine of devia- tion; on the other hand, it adds biological noise to’the cell’s thermal-equilibrium agitation. Moreover, one cannot deny the remote possibility that the bacteria actively control their direction of swimming. Whether the modified Langevin con- cept will be adequate to explain the bacterial migrations de- pends largely on the randomizing and directive energy that the bacteria put in themselves relative to kT and mB. In the present research, it is determined to what extent the biological aspects make live bacteria deviate from the behavior of “self- propelled Langevin particles.”
BIOLOGICAL ORGANISMS AND EXPERIMENTAL PROCEDURES
All tests are conducted on healthy organisms under condi- tions matching those of the natural habitats as closely as pos- sible. The magnetic fields applied are highly uniform, well calibrated, and free from spurious artifacts. Thus utmost care is taken to guarantee the biological and physical validity of the data.
Elasmobranchs After some preliminary observations on leopard sharks [6],
the, warm-water stingray Urolophus hallen was selected for the present experiments. Urolophus is a hardy lively animal of proven electrical sensitivity. Its small adult size helps in reduc- ing the technical demands. The stingrays are tested, two or three at a time, in circular fiberglass tanks measuring 1.8 m in diameter and filled with natural seawater to a depth of.15 cm
/v s
Reversed
Fig. 5. Stingray orienting to earth’s magnetic field. To receive reward and avoid punishment, animal enters enclosure in magnetic east, dis- regarding similar enclosure in magnetic west. Field is reversed from trial to trial in random order.
(resistivity 23 i-2 * cm at 2OoC). To keep the animals interested in orientation, the setup has been implemented with an artifical daily sun shift. The applied magnetic field mimics that of the San Diego, CA area, where the fish were collected (horizontal intensity 0.26 G; vertical intensity 0.42 G).
The animal tanks are enclosed by a 12-sided hut, erected in the woods, far from any major buildings. Two orthogonal pairs of Helmholtz coils, mounted on the outside of the hut, control the magnetic field in the tanks. The power supplies, one for the coils and one for heating and illumination, are located in a remote utility box. Magnetic interference is minimized by the use of dc power and tightly twisted wires. The hut is kept virtually free from ferromagnetic materials. The tanks are electrically isolated from ground and no metal is allowed to come into contact with the water.
In the actual orientation experiments, the stingrays are trained to procure food from an enclosure in the magnetic east, but not to enter a similar enclosure in the magnetic west of their habitat. To preclude the use of alternative cues, the horizontal component of the magnetic field is reversed ran- domly from trial to trial. After the field polarity is selected, the two plastic enclosures are lowered into the tank with their entrances facing the wall (Fig. 5). As soon as a stingray enters one of the enclosures, the entrances of both are closed off by gates. After an incorrect choice, the animal is lightly punished with a Plexiglas rod; after a correct choice, it is rewarded with a small piece of herring. Subsequently, the enclosures are moved to the north and south of the tank and the animal is set free in preparation for the next trial. The number of trials per series depends on the amount of food the animals can take without becoming sated (approximately seven pieces per fish per day). The final tests are carried out in a double blind fashion. The two experimenters handle the enclosures unaware of the field direction, while a third person, having no view of the animals, hands out the feeding stick or punishing rod after a choice has been made.
Mud Bacteria The freshwater magnetic bacteria were collected from Cedar
Swamp, the marine species from Eel Pond, Woods Hole area. It was on bacteria from these localities that the organisms’ geomagnetic orientation was first established [I21 , [ 141 . The mud samples are taken from the top layer of the sedi- ments and are kept in glass jars under laboratory conditions. The bacteria chosen for this study have their flagellae at one end and propel themselves unidirectionally, swimming steadily when separated from the substrate (Fig. 4). Aged mud samples

1118 IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO. 1, JANUARY 1981
contain predominantly cells of only a few types. Although no taxonomic information is available as yet, one may repro- ducibly identify the types by size and performance with rea- sonable confidence.
The bacterium setup is located in the basement of the Insti- tution’s estate house, where the earth’s magnetic field is fairly undistorted. Two pairs of Helmholtz coils null the earth‘s magnetic field in the room; another two pairs produce a hori- zontal test field of either normal or reversed polarity. The field strength is selected from seven pre-calibrated settings, ranging from $ to 8 G. The bacteria are observed under a Zeiss OPMI 1 Operation Microscope at a magnification of 83.2X. This microscope lends itself particularly well to mag- netic studies because of its long working distance (10 cm) and its relative freedom from ferromagnetic materials. By oblique illumination the bacteria may be seen as bright dots against a dark background.
To measure their rate of migration, one to a few bacteria are transferred from the mud into a small observation chamber built upon a standard hemacytometer slide. In the chamber, anaerobic conditions are maintained to keep the bacteria from slowing down due to oxygen exposure. A graticule etched in the bottom of the chamber provides a calibrated grid with its major divisions 1 mm apart. The migration rate of single bacteria is determined by recording the time of travel in the direction of the field over the measured 1-mm distance. After each run the magnetic field is reversed to make the bacterium traverse the same distance in the opposite direction. Single cells may swim back and forth for up to 100 crossings without noticeably slowing down. The observer signals the beginning and the end of each run and reverses the field with a foot- switch. A second person records the time of travel in l/lOOths of a second and sets the field strength. Thus the rate of migra- tion is sampled over several 1-mm runs at each of the pre- selected gauss settings.
RESULTS AND STATISTICAL EVALUATION Except for the physical stimulus, the elasmobranch experi-
ments are entirely different from those on the bacteria. The elasmobranchs must be strongly motivated to seek food de- spite the known danger of punishment. The motionalelectric energy is extremely small, and the conditions have to be just right for the animals to reveal their orientational abilities. Also, the elasmobranchs may choose to bear in any direction they wish with respect to the field. The magnetic bacteria, although required to swim for the present study, orient equally well even when dead. The response is mainly passive; no signal processing on the part of the organism is needed. The mag- netic torque acts directly on the axis of motility. In fact, it remains to be determined whether free-swimming cells can escape geomagnetic alignment for any length of time.
Elasmobranchs Before the actual tests, the stingrays are first trained to
receive food in an enclosure. Then, they are conditioned only to enter the enclosure in the magnetic east of the tank and not to visit the one in the west. During the initial period, the experimenters do not work for a good score but concentrate
TABLE I ELASMOBRANCH GEOMAGNETIC ORIENTATION: MAGNETIC EAST
VERSUS MAGEETIC WEST’
Day it correct 0 incorrect # t o t a l 1 2 3 4 1 2 3 4 1 2 3 4
1 7 5 6 7 3 1 4 5 10 5 1 0 1 2 2 8 5 1 0 8 4 1 2 3 12 7 1 2 1 1
4 8 1 7 5 3 3 5 5 11 4 1 2 1 1 3 9 4 a 1 0 3 2 1 1 12 5 9 1 1
5 6 3 1 0 * 9 4 4 2 4 10 7 1 2 1 3 6 6 6 3 6 2 0 5 6 8 6 8 1 2 7 6 6 7 6 3 2 5 5 9 8 1 2 1 1 8 8 4 I 9 5 1 5 3 13 5 1 2 1 2 9 10 11 12 13
15 14
16 17
9 * 4 9 5 7 4 1 2 7 9 5 7 4 7 9* 10 9
1 1 . . 7 7 . . 4
7 . . 5 * 9 . . . 9** . . .
4 2 2 3 2 4
3 3 5 4 1 4 5 . . 4 . .
4 . . 3 . .
3 . .
12 6 13 7 11 5 14 11 12 8 12 10 11 10 14 12 1 2 . . 7
1 0 . . 6 1 5 . . 8
13 . . . 12 . . .
aFour series of trials-1-3: three out of four versus random; 4: seven
*Series turns significant at P < 0.001. **Series turns significant at P < 0.00001.
out of ten versus random.
on teaching the animals their task to give them the necessary confidence. For a successful completion of the behavioral tests, one must realize that it is the scientist who has to learn how to conduct the experiments. In the end, though, the laurels go to the animals. Table I summarizes the performances of two of the stingrays in four (3 + 1) independent series of trials,
To make the most economical use of the responses, the data are analyzed by the method of sequential analysis [23] . This procedure allows the experimenter to decide from trial to trial whether a) to accept the stingray’s orientation, b) to reject the hypothesis, or c) to collect more data. After each trial, one calculates the probability p o that the data thus far obtained derive from, say, a three-out-of-four-correct orientation (0.75), and the probability p r that the data thus far obtained are the result of random selection (0.50). When p o is much larger than p,., orientation is accepted; when p o is much smaller than p r , orientation is rejected; if the ratio of po to p r does not allow any conclusion yet, more data are collected. The criteria for acceptance and rejection depend on the risks (11 and p of making a wrong decision either way. Both CY and 0 are con- servatively set at 0.001 or less to attach to these studies a high level of confidence.
According to mathematical theory, if
orientation is accepted or rejected, respectively. With c, the number of correct choices thus far and in the number of incor- rect choices, a decision is made whenever the following critical values for e, are attained:
1 - 0.75 1 - 0.001 -in In ___ + l n
1 - 0.50 0.001 in In 2 f In 999 c, = - -
0.75 In 3/2 ln- 0.50
(1 5)
The critical values of e, and the actual number of correct choices may be plotted versus the number of incorrect choices
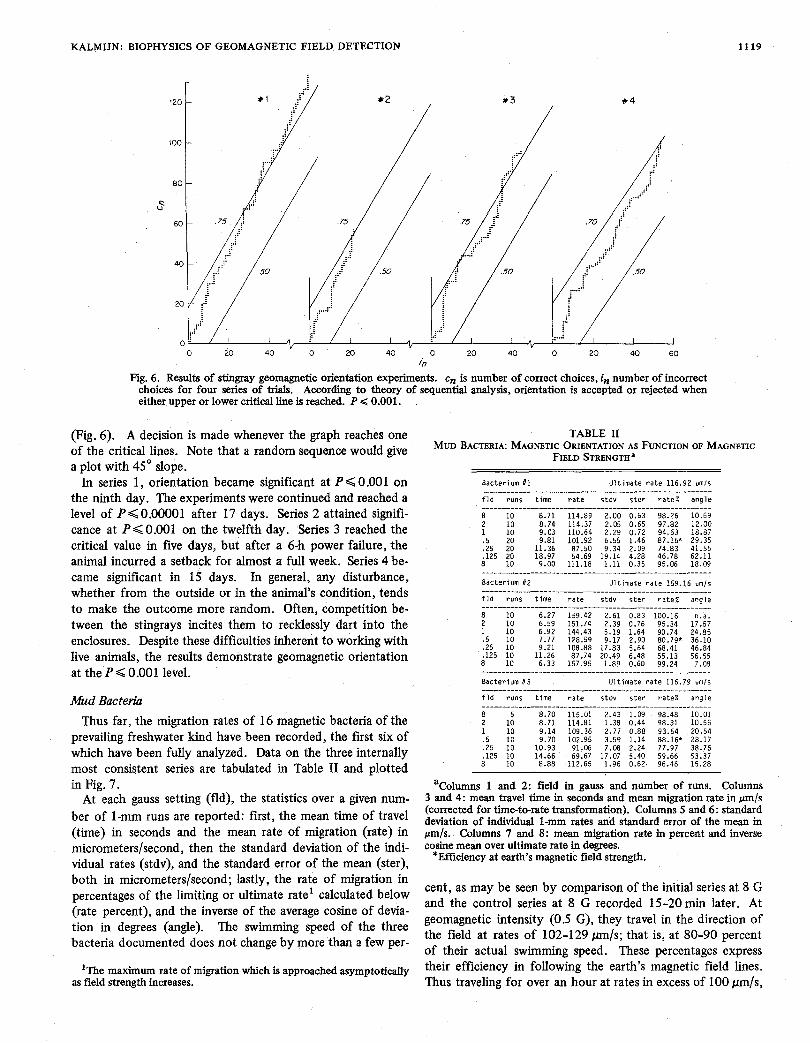
KALMIJN: BIOPHYSICS OF GEOMAGNETIC FIELD DETECTION 1119
either.upper or lower critical line is reached.-P < 0.001.
(Fig. 6) . A decision is made whenever the graph reaches one of the critical lines. Note that a random sequence would give a plot with 45" slope. In series 1 , orientation became significant at P < 0.001 on
the ninth day. The experiments were continued and reached a level of P< 0.00001 after 17 days. Series 2 attained signifi- cance at P < 0.001 on the twelfth day. Series 3 reached the critical value in five days, but after a 6-h power failure, the animal incurred a setback for almost a full week. Series 4 be- came significant in 15 days. In general, any disturbance, whether from the outside or in the animal's condition, tends to make the outcome more random. Often, competition be- tween the stingrays incites them to recklessly dart into the enclosures. Despite these difficulties inherent to working with live animals, the results demonstrate geomagnetic orientation at the P < 0.001 level.
Mud Bacteria Thus far, the migration rates of 16 magnetic bacteria of the
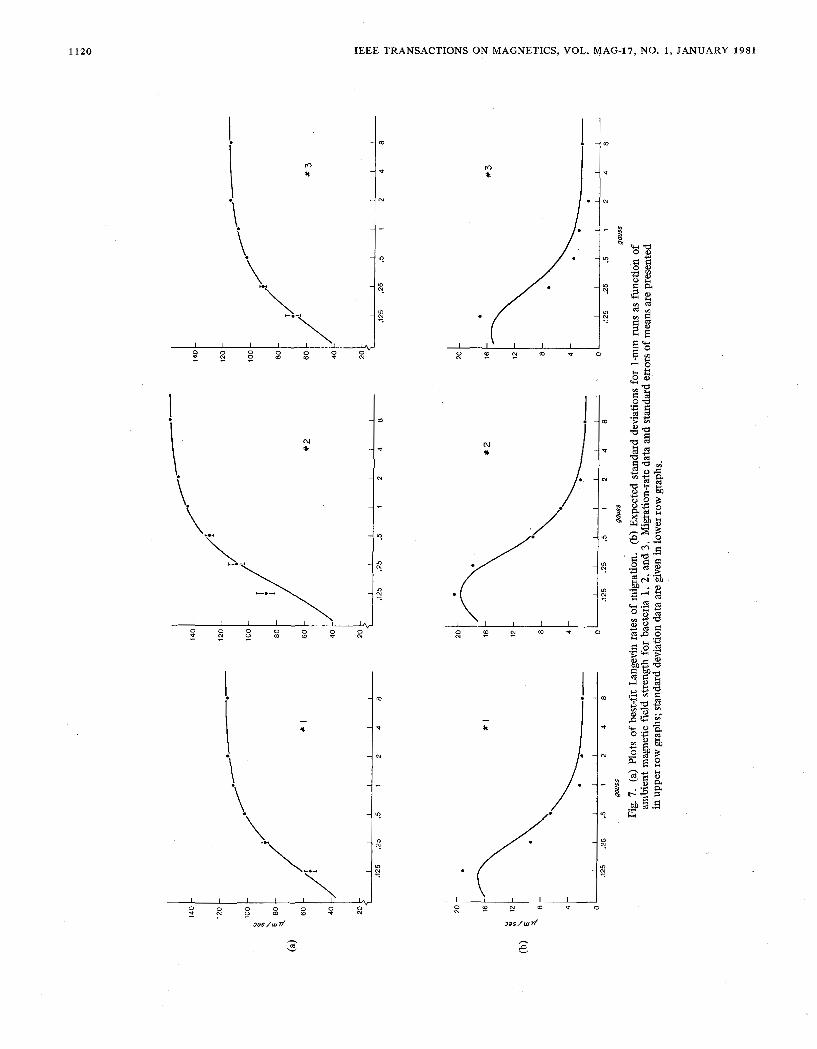
prevailing freshwater kind have been recorded, the first six of which have been fully analyzed. Data on the three internally most consistent series are tabulated in Table I1 and plotted in Fig. 7.
At each gauss setting (fld), the statistics over a given num- ber of 1-mm runs are reported: first, the mean time of travel (time) in seconds and the mean rate of migration (rate) in micrometers/second, then the standard deviation of the indi- vidual rates (stdv), and the standard error of the mean (ster), both in micrometers/second; lastly, the rate of migration in percentages of the limiting or ultimate rate' calculated below (rate percent), and the inverse of the average cosine of devia- tion in degrees (angle). The swimming speed of the three bacteria documented does not change by more than a few per-
'The maximum rate of migration which is approached asymptotically as field strength increases.
TABLE I1 MUD BACTERIA: MAGNETIC ORIENTATION AS FUNCTION OF MAGNETIC
FIELD STRENGTH^
Bacterium I1 Ultimate rate 116.92 umls
fld runs time rate stdv ster rate% angle ~ ~~~~~
8 10 8.71 114.89 2.00 0.63 98.26 10.69 2 10 8.74 114.37 2.05 0.65 97.82 12.00 1 10 9.03 iio.64 2.29 0.72 94.63 18.87
~ ~~
.25 20 11.36 87.50 9.34 2-09 74.83 41.55
.5 20 9.81 101.92 6.55 1.46 87.16* 29.35
8 10 9.00 111.18 1.11 0.35 95.06 18.09 .125 20 18.97 54.69 19.14 4.28 46.78 62.11
fld runs time rate stdv rter rate% angle ~~~~~
8 10 6.27 159.42 2.61 0.83 100.16 n.a. 2 10 6.59 151.74 2.39 0.76 95.34 17.57 1 10 6.92 144.43 5.19 1.64 90.74 24.85 .5 10 7.77 128.59 9.17 2.90 80.79* 36.10 .25 10 9.21 108.88 17.83 5.64 68.41 46.84 '.125 10 11.26 87.74 20.49 6.48 55.13 56.55 8 10 6.33 157.95 1.89 0.60 99.24 7.08
Bacterium 13 Ultimate rate 116.79 umls ........................................................
fld runs time rate stdv ster rate% angle
8 5 8.70 115.01 2.43 1.09 98.48 10.01 2 10 8.71 114.81 1.38 0.44 98.31 10.56 1 10 9.14 109.36 2.77 0.88 93.64 20.54 .5 10 9.70 102.95 3.59 1.14 88.16* 28.17
,125 10 14.66 69.67 17.07 5.40 59.66 53.37 .25 10 10.93 91.06 7.08 2.24 77.97 38.76
8 10 8.88 112.65 1.96 0.62. 96.46 15.28
aColumns 1 and 2: field in gauss and number of runs. Columns 3 and 4: mean travel time in seconds and mean migation rate in pm/s (corrected for time-to-rate transformation). Columns 5 and 6: standard deviation of individual 1-mm rates and standard error of the mean in pm/s. Columns 7 and 8: mean migration rate in percent and inverse cosine mean over ultimate rate in degrees.
*Efficiency at earth's magnetic field strength.
cent, as may be seen by comparison of the initial series at 8 G and the control series at 8 G recorded 15-20 min later. At geomagnetic intensity (0.5 G), they travel in the direction of the field at rates of 102-129 p r n / s ; that is, at 80-90 percent of their actual swimming speed. These percentages express their efficiency in following the earth's magnetic field lines. Thus traveling for over an hour at rates in excess of 100 pm/s,
1120 IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO. 1 , JANUARY 1981
nr #
yc)
#
N #

KALMIJN: BIOPHYSICS OF GEOMAGNETIC FIELD DETECTION 1121
magnetic bacteria cover distances of 36 cm and more. Note that at 0.5 G the bacteria deviate from a straight line at an inverse average cosine of 28-36”, in accordance with the ob- served 10-20 percent loss of speed due to swerving.
The bacterial performances are compared to those of the hypothetical self-propelled Langevin particles by nonlinear regression analysis. Curve fitting is accomplished by the least squares method [24], [25]. As the variance of the model depends on the field strength, and also fluctuations in swim- ming speed and errors in timing add variability to the data, the squared deviations are weighted by the expected total migration-rate variations. .The sum of the weighted squared deviations is
in which *vi is the experimentally determined mean migration rate; Li the Langevin function (9); Vo the ultimate migration rate; and LjVo the mean migration rate as predicted by the Langevin model at the selected field strengths. In the denomi- nator, *u: is the Langevin variance of the runs, taking into account the number of randomizations per run (the expected time of travel divided by the randomization time At). I J~ is the experimentally determined swimming and timing variance; ri is the number of runs at the particular gauss setting i, and n the number of gauss settings. In (16), the quantities marked with an asterisk (*)have been corrected to allow for the trans- formation of “travel time” into “rate of migration,” following a procedure derived from the diffusion approximation to the Langevin model. In [22] it will be shown that:
*vi = vi - u . i ” ~ , ” ~ t ( 1 6 4
*I$ = o.i“LiV:At t 2(u;V;At (1 6b)
with Vi the uncorrected mean migration rate and u; the Langevin variance given by (13).
To obtain the least squares, expression (16) is minimized by a suitable choice of its parameters Vo and m at an estimated At. To establish this minimum, the partial derivatives with
j respect to Vo and m are set equal to zero. The resulting non- linear “normal equations” are solved numerically for Vo and m by the two-dimensional Newton-Raphson method, which follows an iterative procedure, making use of two-dimensional Taylor series. Subsequently, the variances of the model are fitted to the variances of the bacteria with At as the parameter [22]. Thus a best fit is obtained in terms of the ultimate migration rate Vo , the magnetic moment m, and the randomi- zation time At. This method also gives the chi-square values for the migration rates, for the variances, and for the total fit with the model as null hypothesis, and yields the approximate confidence limits for the estimated parameters. Table I11 and the graphs of Fig. 7 summarize the results.
The limiting or ultimate migration rate estimates the bacte- ria’s actual swimming speed (forward thrust), which ranges in the recorded specimens from 117-159 pm/s at 23°C. The swimming speed usually remains remarkably constant for up to a hundred runs (and reversals), after which the bacteria gradually slow down before eventually coming to a grinding
TABLE 111 CURVE-FITTING DATA FOR BACTERIA^
Bacter ium ..._.... ii 1 112 113
U l t i m a t e r a t e , 1, 116.92 159.16 95% conf. l i m i t s 115.34-118.32 157.23-161.34 114.70-118.74
116.79
Magn. moment, m 6.23 4.88 7.33 95% conf . f l imi ts 5.31-7.70 2.97-6.77 5.73-9.69
Random. t ime, P C I .83 0.97 1.49 95% conf. l i m i t s 1.20-3.85 0.72-7.30 0.82-5.82
Chi -sauare ra te data 9.0415 11.1079 degr . f r . , r e j . 5% 4 no 4 yes 4 no
Chi-sauare vrnc data 4.8763 d e g r . f r . , r e j . 5% 5 no
0.9768 5 no 5 no
6.3033
Chi-square t o t a l f i t 13.9178 12.0847 10.3339 degr.fr.. r e j . 5% 9 no 9 no 9 no
4.0305
VO Ultimate migration rate, wm/s. m Magnetic dipole moment, ergs/G.
At Randomization time, s. ag5 percent confidence limits $ corresponding units; chi-square
values for migration-rate data, for variance data, and for total fit; de- grees of freedom, and rejection “yes” or “no” at five percent signifi- cance level. Bacteria 1 and 3 show acceptable fits; 2 is formally rejected for its lack of fit in migration-rate data.
halt. The best-fit dipole moments of bacteria 1 and 3 are, respectively, 6.23 and 7.33 X ergs/(;, the randomization times 1.83 and 1.49 s. The conservatively rated confidence limits indicate the precision of these estimates. The chi-square values for the migration rates, for the variances, and for the fit in its entirety imply that bacteria 1 and 3 do not signifi- cantly differ in their behavior from the proposed self-propelled Langevin particles at the ‘five percent significance level (at the degrees of freedom given). The migration rates of bacterium 2 do not agree with those of the model at the five percent level, though the variance data fit fhe model very well. This lack of fit formally invalidates the parameter estimates for this bacterium. It should be noted, however, that not only in bac- terium 2 but also in 1 the migration-rate curve deviates sys- tematically from that of the model. In bacterium 1 the curve has a steeper slope, while in 2 the slope is less steep than in the model as may be seen from Fig. 7(a). Bacterium 3 fits the model best. In brief, individual bacteria may noticeably deviate from the modified Langevin model, each in its own characteristic manner, although not often significantly so.
DISCUSSION The reported results leave no doubt that. elasmobranchs
sense the earth‘s magnetic field and may direct themselves accordingly, whereas magnetic bacteria are committed to swimming along field lines by passive dipole alignment. The stingrays exhibit great proficiency in geomagnetic orientation, and one may safely assume that the. animals had practiced their talents long before they were taught to do so in the de- scribed training experiments. The orientation of magnetic bacteria is largely determined by purely physical forces and seems virtually inescapable to free-moving cells. In the mud, interface forces complicate their behavior. Thus the elasmo- branchs’ compass orientation and the bacteria’s magnetically directed migration suggest an important role of their “mag- netic sense” in daily life. However, adequate field studies will be required to verify such a plausible, yet conjectural conclusion.

1122 IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO. 1 , JANUARY 1981
In elasmobranchs, the magnetic sense was anticipated from the physics of electromagnetic orientation and the animals’ astounding electrical sensitivity [6]. Hence the demonstration of their magnetic abilities has tacitly been interpreted as evidence in support of the proposed mode of operation. Although the results are perfectly consistent with this inter- pretation, formal identification of the mechanism(s) involved has yet to follow. Regarding the theory of electromagnetic orientation, it should be emphasized that elasmobranchs do not directly detect the EMF of induction but rather the result- ing ohmic voltage gradient integrated across the body ( 5 ) [6]. In marine species, the ohmic gradient reaches almost its full value because of the high resistivity of the animal’s skin and body tissues relative to the conductivity of the seawater envi- ronment. In freshwater fish the mechanism is less likely, in terrestrial and air-borne animals it is highly unlikely as far as circuits embracing the animal and its environment are con- cerned [lo] . The ohmic voltage gradients of internally circu- lating induction currents, resulting from changes in swimming direction or from relative motion of body parts, are less effec- tive in stimulating the elasmobranchs’ ampullary organs, the canals of which connect to the outside by small pores in the skin. The prerequisites for the detection of internally circulat- ing induction currents and their possible role in other animal species will be discussed elsewhere [26].
When orienting electromagnetically, the animals operate their electric sense in the active mode with the signal energy deriving from the fish’s own swimming movements (active electroorientation), whereas in the detection of fields from ocean currents and prey, they operate their electric sense in the passive mode with the signal energy deriving from external sources (passive electroorientation) [6] . In marine elasmo- branchs, the physical characteristics of the electroreceptor system show themselves fully compatible with this functional duality. Behaviorally, the stingrays respond to the distributed dipole fields of prey, the practically uniform electric fields of ocean currents, and the self-induced electromagnetic fields discriminately without apparent confusion. This biological ver- satility calls for highly developed signal-processing capabilities.
Elasmobranchs exhibit the highest dc and low frequency electrical sensitivity known in the animal kingdom. In the stingray Urolophus halleri, orientation to uniform dc fields has recently been established at 0.005 pV/cm (at 23 . cm), a value half that reported formerly [27]. In small sharks (Mustelus canis) equally low values were determined from the distance of response to electrically simulated prey ([7] , [8] , and unpublished data). This great sensitivity is dictated by the strengths of the bioelectric, geophysical, and electrochemi- cal fields available in the natural habitat [6] . Rough calcula- tion indicates that the electric sense operates down to about the physical noise level of the seawater-receptor system circuit. While electroreceptors detect submicrovolt stimuli, the regen- erative response of nerve fibers usually requires a depolariza- tion of several millivolts. In nervous conduction, however, the pertinent signals are generated by other parts of the transmis- sion system itself. Therefore, signal strength and electrical excitability may together be elevated well above noise levels in order to achieve reliable operation. On the other hand, the
graded integrative action of the central nervous system may depend on signals much weaker than hitherto expected. In this respect, the electric sense of elasmobranch fishes might be very instructive, as has been pointed out by Adey [28]. For the neurophysiology of elasmobranch electroreception, the reader may also refer to ] 291 -[33] .2
With the mud bacteria, nature has offered an example of the most straightforward biomagnetic orientation mechanism con- ceivable. Nevertheless, the same principles may have been implemented in higher organisms as well [34] . Thus the bac- terial findings are just the beginning of a whole new chapter in “sensory reception.” The aim of the present project has been to study the physics and physiology of this novel orienta- tion mechanism at work in living organisms. The strength of the approach is in its capacity to determine the magnetic characteristics of individual bacteria by a nondestructive method, applied under quite acceptable biological conditions. It is established that the orientation mechanism, though ex- ceedingly simple, is highly effective in directing the bacteria along the earth‘s magnetic field lines. At a geomagnetic inten- sity of 0.5 G, the rate of migration is 80-90 percent of the actual swimming speed of 100-160 ,um/s, which is in itself higher than recorded in bacteria before [22]. Preliminary results have been reported in [I41 and [21] .
To adapt the Langevin theory to bacterial migration, the concept of randomization time was introduced. Although treated as the duration of an all-or-none phenomenon, rather than the time constant of a graded process, the randomization time adequately explains the observed variance of the bacteria’s migration rate as a function of ambient magnetic field strength. It may be noted that at lower field strengths, and correspond- ingly longer times of travel, the increased number of randomi- zations reduce both the observed and the model variances, as is particularly obvious from the standard deviation graph of bacterium 2 (Fig. 7). In sum, the randomization time param- eter allows in most cases good least squares fits between bac- terium and Langevin model (Table 111). Moreover, the experi- mentally estimated randomization times of 1-2 s seem to be perfectly reasonable when checked against the loss of direc- tionality observed in clouds of migrating bacteria after sudden cancelation of the ambient magnetic field.
From the size range and number of magnetite crystals per cell (as measured by Blakemore), one calculates for the bac- teria studied in this paper an approximate dipole moment of 8.6 X ergs/G, which is the product of the average num- ber of crystals per cell (10.2), the estimated volume of each single-domain crystal (56 X 56 X 56-nm cube), the saturation magnetization of magnetite (92 G * cm3/g), and the density of magnetite (5.2 g/cm3). The values arrived at by the Langevin approach range from about 4-8 X ergs/G, in good agree- ment with the value calculated above, given that a) the flagellar thrust raises the effective temperature of the bacteria above that of the surrounding bog water, and b) any imperfection or misalignment of the single-domain magnetite crystals lowers the net dipole moment. Deviation of the net dipole moment
21n [32], the physics of electromagnetic orientation are at variance with [ 6 ] ; in [33], the results of [6] are used without proper reference.

KALMIJN: BIOPHYSICS OF GEOMAGNETIC FIELD DETECTION
from the axis of motility reduces the effective speed of swim- ming. In the earth‘s magnetic field, the directive magnetic energy mB (4-8 X X 0.5 ergs) overrides the randomizing thermal energy kT (1.38 X X ’300 ergs, not counting swimming noise) by a factor of about 5-10, corresponding to the observed efficiency of 80-90 percent in following the earth’s magnetic field lines (cf. [15]). Doubling the amount of magnetite would increase the migration rate efficiency only 5-10 percentage points; i.e., the bacterial dipole strength is ample, but not excessive from an orientational point of view. Actually, cells from regions of different geomagnetic intensity may exhibit proportionally stronger or weaker dipole moments.
In conclusion, the postulated Langevin model leads to a con- sistent overall description of the observed magnetic orientation of living bacteria, although systematic deviations characteristic of individual cells do occur. As the experimentally determined dipole moments are not higher than expected from the amount of magnetite present in the cells, there is no need to invoke an active control system. Thus except for the inevitable swim- ming noise, the flagellar thrust propels the cell without notice- ably affecting the otherwise passive orientation mechanism. However simple their guidance system may be, magnetic bac- teria from the northern and southern hemispheres, having opposite magnetic polarities, both migrate downward to the mud where they flourish in great numbers [35] .
ACKNOWLEDGMENT The author wishes to thank his former sponsor at Scripps
Institution of Oceanography, Dr. Theodore H. Bullock, who encouraged him to undertake the electromagnetic studies. His wife, Vera Kalmijn, conducted most of the stingray training experiments. The bacterium project was initiated together with Dr. R. P. Blakemore, presently at the University of New Hampshire. Barbara D. Teague and Michael K. Gilson collabo- rated with the author on the single-cell experiments. Drs. C . P. Bean (G.E.), E. hi. Purcell (Harvard), R. B. Frankel (M.I.T.), and R. Schiller (Hoboken) generously shared their original ideas and critical comments. Dr. W. K. Smith (W.H.O.I.) offered invaluable help with the statistical analysis. Ms. C. S. Proctor and the author’s son Jelger executed most of the computer programs. T. A. Dourdeville, E. W. Heineke, and R. K. Gibbs scrutinized the manuscript. Dr. R. C . Tipper and Dr. B. J. Zahuranec lent great moral support by their continual interest in the project.
REFERENCES [ 11 S . Dijkgraaf and A. J. Kalmijn, “Verhaltensversuche zur Funktion
der Lorenzinischen Ampullen,” Naturwhenschaften, vol. 49, p. 400,1962.
[2] A. J. Kalmijn, “Elecko-perception in sharks and rays,” Nature,
[ 31 R. W. Murray, “The response of the afftpullae of Lorenzini of elasmobranchs to electrical stimulation, J. Exp. Biol., vol. 39,
V O ~ . 212, pp. 1232-1233,1966.
pp. 119-128,1962. [4] S . Dijkgraaf and A. J. Kalmijn, “Untersuchungen iiber die Funk-
tion der Lorenzinischen Ampullen an Haifischen,” Z. Vergl. Physiol., vol. 47, pp. 438-456, 1963.
[5] A. I. Kalmijn, “The electric sense of sharks and rays,” J. Exp. Biol.,vol. 55,pp. 371-383,1971.
[6] -, “The detection of electric fields from inanimate and animate sources other than electric organs,” in Handbook of Sensory
1123
Physiology, vol. 111/3, A. Fessard, Ed. New York: Springer- Verlag, 1974, pp. 147-200.
[7] -, “The electric and magnetic sense of sharks, skates,and rays,’’ Oceanus, vol. 20(3), pp. 45-52, 1977.
[8] -, “Electric and magnetic sensory world of sharks, skates, and rays,” in Sensory Biology of Sharks, Skates, and Rays, E. S. Hodgson and R. F. Mathewson, Eds. Washington, DC: Govern-. ment Printing Office, 1978, pp. 507-528.
[9] w. s. Von &x, An Introduction to Physical oceanography. Reading, MA: Addison-Wesley, 1962.
[ 101 A. J. Kalmijn, “Experimental evidence of geomagnetic orientation in elasmobranch fishes,” in Animal Migration, Navigation, and Homing, K. Schmidt-Koenig and W. T. Keeton, Eds. New York: Springer-Verlag, 1978, pp. 347-353.
[ 111 -, “Electro-orientation in sharks and rays: theory and experi- mental evidence,” Scripps Inst. Oceanography Reference Ser., Contr. 73-39, pp. 1-22,1973.
[12] R. P. Blakemore, “Magnetotactic bacteria,” Science, vol. 190,
[ 131 A. J. Kalmijn and R. P. Blakemore, “Geomagnetic orientation in marine mud bacteria,” Proc. Int. Union Physiol. Sci., vol. 13, p. 364, 1977.
[ 141 -, “The magnetic behavior of mud bacteria,” in Animal Migra- tion, Navigation, and Homing, K. Schmidt-Koenig and W. T. Keeton, Eds. New York: Springer-Verlag, 1978, pp. 354-355.
[15] R. B. Frankel, R. P. Blakemore, and R. S. Wolfe, “Magnetite in freshwater magnetotactic bacteria,” Science, vol. 203; pp. 1355- 1356,1979.
[16] M. Faraday, “Experimental researches in electricity,” Philos. Trans. R. SOC. Lond., vol. 122(1),pp. 125-194,1832.
[17] M. s. Longuet-Higgins, M. E. Stern, and H. Stommel, “The elec- trical field induced by ocean currents and waves, with applications to the method of towed electrodes,” Pap. Phys. Oceanogu. Met.,
pp. 377-379,1975.
V O ~ . 13(1), pp. 1-37,1954. [ 181 W. Gilbert, De Magnete. London: Petrus Short, 1600. ,
[ 191 S . Chikazumi, Physics of Magnetism. New York: Wiley, 1964. [20] B. Waltman, “Electrical properties and fine structure of the am-
pullary canals of Lorenzini,” Acta Physiol. Scand., vol. 66, Sup- plement 264, pp. 1-60,1966.
[21] B. D. Teague, M. K. Gilson, and A. J. Kalmijn, “Migration rate of mud bacteria as a function of magnetic field strength,” Biol. Bull., vol. 157, p. 399,1979.
[22] A. J. Kalmijn, “Physics and physiology of geomagnetic orienta- tion in bacteria,” forthcoming. (Detailed account of bacterium studies reported in present paper.)
[23] W. J. Dixon and F. J. Massey, Jr., Introduction to Statistical Analysis. New York: McGraw-Ri, 1969.
1241 J. H. Pollard, Numerical and Statistical Techniques. London: Cambridge Univ. Press, 1977.
[25] N. R. Draper and H. Smith, Applied Regression Analysis. New York: Wiley, 1966.
[26] A. J. Kalmijn, “A relativistic approach to animal orientation,” in preparation (to include an extensive discussion of the relevant literature).
[27] -, “Geoelectric-field orientation in marine and freshwater fishes,” in preparation (orientation with respect to naturdly occurring fields of inanimate origin).
[28] W. Ross Adey, “Long-range electromagnetic field interactions at brain cell surfaces,” in Magnetic Field Ejfects on Biological sys- tems, T. S . Tenforde, Ed. New York: Plenum, 1979, pp. 57-80.
[29] R. W. Murray, “The ampullae of Lorenzini,” in Handbook of Sensory Physiology, vol. 111/3, A. Fessard, Ed. New York: Springer-Verlag, 1974, pp. 125-146.
[30] S . Obara and M. V. L. Bennett, “Mode of operation of ampullae of Lorenzini of the skate, Raja,” J. Gen. Physiol., vol. 60, pp.
M. V. L. Bennett and W. T. Clusin, “Physiology of the ampulla of Lorenzini, the electroreceptor of elasmobranchs,” in Sensory Biology of Sharks, Skates, and Rays, E. S. Hodgson and R. F. Mathewson, Eds. Washington, DC: Government Printing Office,
[31] T. H. Bullock, “Processing of ampullary input in the brain: Com- parison of sensitivity and evoked responses among elasmobranch and siluriform fishes,” J. Physiol. (Paris), vol. 75, pp. 397-407, 1979.
[32] G. N. Andrianov, H. R. Brown, and 0. B. Ilyinsky, “Responses of central neurons to electrical and magnetic stimuli of the am-
534457,1972.
1978, pp. 483-505.

1124 IEEE TRANSACTIONS ON MAGNETICS, VOL. MAG-17, NO. 1 , JANUARY 1981
pullae of Lorenzini in the Black Sea skate,” J. Comp. Physiol., [34] C. Walcott, J. L. Gould, and J. L. Kirschvink, “Pigeons have mag-
[33] H. R. Brown and 0. B. Ilyinsky, “The ampullae of Lorenzini in [35] R. P. Blakemore, R. B. Frankel, and A. J. Kalmijn, “South-seeking the magnetic field,” J. Comp. Physiol., vol. 126, pp. 333-341, magnetotactic bacteria in the southern hemisphere,” Nature, in 1978. press.
VOI. 93, pp. 287-299,1974. nets,” Science, vol. 205, pp. 1027-1028,1979.
Fundamentals of Modeling I-Bars and Bubbles Using Fourier Series
Abstract-A Fourier series method is described for the calculation of the response of periodic arrays of I-bars to applied fields and bubbles. The underlying assumptions are emphasized for the exact solution of Poisson’s equation with an approximate constitutive equation. A gen- eral formulation is given for bubble-bar magnetostatic energy, the components of which are tabulated systematically. Results are given as comparisons with other investigators for calculated and measured potential wells and the variation of well with spacing, with applied field, and with bubble radius. Bubble-bubble interaction is treated.
I. INTRODUCTION
R ECENTLY, two major approaches have been described for the calculation of the response of Permalloy overlay
elements to applied fields and to the effect of bubbles. George and Hughes have described a two-component two-variable numerical simulation [ l ] , whereas Almasi and Lin [ 2 ] have described a flux and reluctance analytical model. Ishak and Della Torre developed an iterative method based on rapid con- vergence to a solution for the magnetostatic potential [3]. This paper details an approach which is based on the use of Fourier series and transforms.
Fourier series methods have been applied previously to magnetostatic calculations. Dove [4] used a Fourier series method to calculate the response of an infinite thin film to a nonuniform hard-axis applied field. His early paper demon- strated the usefulness of the series method, which permitted the demagnetizing field to be expressed as a function of the magnetization. The magnetostatic problem, therefore, became deterministic. Lin [5] first used Fourier series to calculate the response of finite I-bars using shape function multipliers to define the finite bars in two dimensions.
Manuscript received July 20, 1979; revised August 11, 1980. This work was supported in part by the National Science Foundation.
E. Huijer was with the University of Florida, Gainesville, FL 32611. He is now with Hewlett-Packard, Palo Alto, CA 94304.
D. B. Dove was with the University of Florida, Gainesville, FL 3261 1. He is now with the IBM Thomas J. Watson Research Center, Yorktom Heights, NY 10598.
J. K. Watson is with the University of Florida, Gainesville, FL 32611.
In earlier reports from the Florida group, the effects of bubbles were included [6] , and the consequences were noted of finite permeability of the Permalloy [7] and of bar-bar inieractions [8] . This paper lays out the magnetic assumptions behind the present method, presents details of the Fourier formulation, and gives detailed comparisons of the calculated results. A companion paper [9] contrasts continuum modeling with domain observations. The present approach defines a simplified constitutive equation for the magnetization with one component and one variable, yet calculates the response in three dimensions.
11. THE MAGNETIZATION PROBLEM In order to arrive at a solution of the magnetization problem
of Permalloy I-bars, a necessary first step is the definition of the geometrical conditions and the physical assumptions. The equation that results is then transformed into a set of linear equations using Fourier series and transforms. Solving these by computer yields quantitative results of magnetization distributions. Responses of several cases, of a uniform applied field, and a nonuniform (bubble stray) field have been treated.
A. Geometry In order to permit the use of Fourier series, it is desirable to
deal with geometries of interest as periodic structures. For the present model, structures will be used that are periodic in x and y directions. To approach the situation in bubble de- vices, a permeable overlay structure is placed at some distance over a uniformly magnetized layer carrying cylindrical domains of opposite magnetic orientation. The overlay structure con- sists of an array of I-bars. Fig. 1 shows the geometrical con- figuration of the array and defines the parameters X,, Xy , L , w, t. The layer of bubble material is treated as a layer with uniform magnetization -MBZ and an array of circular cylinders with magnetization +2MB2. As Fig. 2 shows, superposition of these two components yields a magnetized layer with an array of reversed bubble domains. The array of cylinders is charac-
0018-9464/81/0100-1124$00.75 0 1981 IEEE
Related Documents











