Biomonitoring genotoxicity and cytotoxicity of Microcystis aeruginosa (Chroococcales, Cyanobacteria) using the Allium cepa test Haywood Dail Laughinghouse IV a, ⁎ , 1 , Daniel Prá a, b, f , Maria Estela Silva-Stenico c , Alexandre Rieger a , Viviane Dal-Souto Frescura d , Marli Fátima Fiore c , Solange Bosio Tedesco d, e a Laboratory of Biotechnology and Genetics, Department of Biology and Pharmacy, University of Santa Cruz do Sul, Santa Cruz do Sul, RS, Brazil b Laboratory of Genetics and Neurosciences, Center of Life and Health Sciences, Catholic University of Pelotas, Pelotas, RS, Brazil c Laboratory of Molecular Ecology of Cyanobacteria, Center for Nuclear Energy in Agriculture, University of São Paulo, Piracicaba, SP, Brazil d Agrobiology Graduate Program, Federal University of Santa Maria, RS, Brazil e Laboratory of Plant Cytogenetics and Genotoxicity, Department of Biology, Federal University of Santa Maria, RS, Brazil f Graduate Course in Health Promotion, University of Santa Cruz do Sul, Santa Cruz do Sul, RS, Brazil abstract article info Article history: Received 15 November 2011 Received in revised form 30 May 2012 Accepted 30 May 2012 Available online xxxx Keywords: Genotoxicity Cytotoxicity Cyanotoxins Microcystis aeruginosa Allium cepa test Water pollution caused by toxic cyanobacteria is a problem worldwide, increasing with eutrophication. Due to its biological significance, genotoxicity should be a focus for biomonitoring pollution owing to the increasing com- plexity of the toxicological environment in which organisms are exposed. Cyanobacteria produce a large number of bioactive compounds, most of which lack toxicological data. Microcystins comprise a class of potent cyclic hep- tapeptide toxins produced mainly by Microcystis aeruginosa. Other natural products can also be synthesized by cyanobacteria, such as the protease inhibitor, aeruginosin. The hepatotoxicity of microcystins has been well doc- umented, but information on the genotoxic effects of aeruginosins is relatively scarce. In this study, the gen- otoxicity and ecotoxicity of methanolic extracts from two strains of M. aeruginosa NPLJ-4, containing high levels of microcystin, and M. aeruginosa NPCD-1, with high levels of aeruginosin, were evaluated. Four endpoints, using plant assays in Allium cepa were applied: rootlet growth inhibition, chromosomal aberrations, mitotic di- visions, and micronucleus assays. The microcystin content of M. aeruginosa NPLJ-4 was confirmed through ELISA, while M. aeruginosa NPCD-1 did not produce microcystins. The extracts of M. aeruginosa NPLJ-4 were di- luted at 0.01, 0.1, 1 and 10 ppb of microcystins; the same procedure was used to dilute M. aeruginosa NPCD-1 used as a parameter for comparison, and water was used as the control. The results demonstrated that both strains inhibited root growth and induced rootlet abnormalities. The strain rich in aeruginosin was more geno- toxic, altering the cell cycle, while microcystins were more mitogenic. These findings indicate the need for future research on non-microcystin producing cyanobacterial strains. Understanding the genotoxicity of M. aeruginosa extracts can help determine a possible link between contamination by aquatic cyanobacteria and high risk of pri- mary liver cancer found in some areas as well as establish water level limits for compounds not yet studied. © 2012 Elsevier B.V. All rights reserved. 1. Introduction The decrease in water quality, especially in environments used for public water supply, irrigation, and recreation, is of increasing concern. Eutrophication in these environments has favored the abundance and/ or dominance of toxigenic cyanobacteria, such as the predominance of Microcystis aeruginosa (Kütz.) Kütz. This threatens human and animal health, as well as elevates water treatment costs. One of the worst problems resulting from cyanobacterial blooms is the production of toxins. As a principle, all cyanobacteria are considered potentially toxic, and there have been bioactive blooms associated to acute intoxications in humans and animals, reported worldwide (Hunter, 1998; Carmichael et al., 2001; Azevedo et al., 2002; Azevedo, 2006). Cyanotoxins can be characterized by their chemical structure as peptides, alkaloids, or lipopolysaccharides. However, due to their pharmacological action, the two principal classes of cyanotoxins are neu- rotoxins (anatoxin-a, anatoxin-a(s), and saxitoxins) and hepatotoxins (microcystins, nodularin, and cylindrospermopsin) (Azevedo, 2006). The most common intoxications by cyanobacteria, according to Carmichael (1991) and Roset et al. (2001) are provoked by hepatotoxins. Microcystis is known to produce microcystins, which inhibit pro- tein phosphatase (PP1 and PP2A), causing an increase in protein phosphorylation, related to its cytogenetic and tumorigenic effects Science of the Total Environment 432 (2012) 180–188 ⁎ Corresponding author at: Department of Botany, MRC-166, National Museum of Natural History, Smithsonian Institution, PO Box 37012, Washington, DC 20013‐7012, USA. Tel.: +1 202 431 4228. E-mail address: [email protected] (H.D. Laughinghouse). 1 Present address: College of Computer, Mathematical and Natural Sciences, Univer- sity of Maryland, College Park, MD, USA. 0048-9697/$ – see front matter © 2012 Elsevier B.V. All rights reserved. doi:10.1016/j.scitotenv.2012.05.093 Contents lists available at SciVerse ScienceDirect Science of the Total Environment journal homepage: www.elsevier.com/locate/scitotenv

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Science of the Total Environment 432 (2012) 180–188
Contents lists available at SciVerse ScienceDirect
Science of the Total Environment
j ourna l homepage: www.e lsev ie r .com/ locate /sc i totenv
Biomonitoring genotoxicity and cytotoxicity ofMicrocystis aeruginosa (Chroococcales,Cyanobacteria) using the Allium cepa test
Haywood Dail Laughinghouse IV a,⁎,1, Daniel Prá a,b,f, Maria Estela Silva-Stenico c, Alexandre Rieger a,Viviane Dal-Souto Frescura d, Marli Fátima Fiore c, Solange Bosio Tedesco d,e
a Laboratory of Biotechnology and Genetics, Department of Biology and Pharmacy, University of Santa Cruz do Sul, Santa Cruz do Sul, RS, Brazilb Laboratory of Genetics and Neurosciences, Center of Life and Health Sciences, Catholic University of Pelotas, Pelotas, RS, Brazilc Laboratory of Molecular Ecology of Cyanobacteria, Center for Nuclear Energy in Agriculture, University of São Paulo, Piracicaba, SP, Brazild Agrobiology Graduate Program, Federal University of Santa Maria, RS, Brazile Laboratory of Plant Cytogenetics and Genotoxicity, Department of Biology, Federal University of Santa Maria, RS, Brazilf Graduate Course in Health Promotion, University of Santa Cruz do Sul, Santa Cruz do Sul, RS, Brazil
⁎ Corresponding author at: Department of Botany, MNatural History, Smithsonian Institution, PO Box 37012USA. Tel.: +1 202 431 4228.
E-mail address: [email protected] (H.D. Laughingh1 Present address: College of Computer, Mathematica
sity of Maryland, College Park, MD, USA.
0048-9697/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.scitotenv.2012.05.093
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 November 2011Received in revised form 30 May 2012Accepted 30 May 2012Available online xxxx
Keywords:GenotoxicityCytotoxicityCyanotoxinsMicrocystis aeruginosaAllium cepa test
Water pollution caused by toxic cyanobacteria is a problemworldwide, increasingwith eutrophication. Due to itsbiological significance, genotoxicity should be a focus for biomonitoring pollution owing to the increasing com-plexity of the toxicological environment inwhich organisms are exposed. Cyanobacteria produce a large numberof bioactive compounds,most ofwhich lack toxicological data.Microcystins comprise a class of potent cyclic hep-tapeptide toxins produced mainly by Microcystis aeruginosa. Other natural products can also be synthesized bycyanobacteria, such as the protease inhibitor, aeruginosin. The hepatotoxicity of microcystins has beenwell doc-umented, but information on the genotoxic effects of aeruginosins is relatively scarce. In this study, the gen-otoxicity and ecotoxicity of methanolic extracts from two strains of M. aeruginosa NPLJ-4, containing highlevels ofmicrocystin, andM. aeruginosaNPCD-1,with high levels of aeruginosin, were evaluated. Four endpoints,using plant assays in Allium cepawere applied: rootlet growth inhibition, chromosomal aberrations, mitotic di-visions, and micronucleus assays. The microcystin content of M. aeruginosa NPLJ-4 was confirmed throughELISA, whileM. aeruginosa NPCD-1 did not produce microcystins. The extracts of M. aeruginosa NPLJ-4 were di-luted at 0.01, 0.1, 1 and 10 ppb of microcystins; the same procedure was used to dilute M. aeruginosa NPCD-1used as a parameter for comparison, and water was used as the control. The results demonstrated that bothstrains inhibited root growth and induced rootlet abnormalities. The strain rich in aeruginosin was more geno-toxic, altering the cell cycle, while microcystinsweremoremitogenic. These findings indicate the need for futureresearch on non-microcystin producing cyanobacterial strains. Understanding the genotoxicity of M. aeruginosaextracts can help determine a possible link between contamination by aquatic cyanobacteria and high risk of pri-mary liver cancer found in some areas as well as establish water level limits for compounds not yet studied.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
The decrease in water quality, especially in environments used forpublic water supply, irrigation, and recreation, is of increasing concern.Eutrophication in these environments has favored the abundance and/or dominance of toxigenic cyanobacteria, such as the predominance ofMicrocystis aeruginosa (Kütz.) Kütz. This threatens human and animalhealth, as well as elevates water treatment costs.
RC-166, National Museum of, Washington, DC 20013‐7012,
ouse).l and Natural Sciences, Univer-
rights reserved.
One of the worst problems resulting from cyanobacterial blooms isthe production of toxins. As a principle, all cyanobacteria are consideredpotentially toxic, and there have been bioactive blooms associatedto acute intoxications in humans and animals, reported worldwide(Hunter, 1998; Carmichael et al., 2001; Azevedo et al., 2002; Azevedo,2006). Cyanotoxins can be characterized by their chemical structureas peptides, alkaloids, or lipopolysaccharides. However, due to theirpharmacological action, the two principal classes of cyanotoxins are neu-rotoxins (anatoxin-a, anatoxin-a(s), and saxitoxins) and hepatotoxins(microcystins, nodularin, and cylindrospermopsin) (Azevedo, 2006).The most common intoxications by cyanobacteria, according toCarmichael (1991) and Roset et al. (2001) are provoked byhepatotoxins.
Microcystis is known to produce microcystins, which inhibit pro-tein phosphatase (PP1 and PP2A), causing an increase in proteinphosphorylation, related to its cytogenetic and tumorigenic effects

181H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
(Yoshizawa et al., 1990; Humpage and Falconer, 1999) or promotingliver cell necrosis (Takenaka, 2000). In addition, the mechanism ofmicrocystin genotoxicity can also be due to the induction of reactiveoxygen species (ROS). In non-lethal doses, these toxins have been re-lated with the development of tumors (Humpage and Falconer,1999). Aeruginosins are natural linear peptides with protease inhibitionactivity, characterized by the presence of four subunits: N-terminalamino acid group (3–4 hydroxyphenyl lactic acid—Hpla), a hydrophobicamino acid, a central residue 2-carboxy-6-hydroxyoctahydroindol (Choi)and a group with a guanidine-C terminal (Ishida et al., 2009). Structuralchanges may occur in Hpla subunits (mono-, di-chlorination andsulphation) and Choi (sulphation, glycosylation and chlorination).Due to these biological activities, these substances deserve more studiesso they can be placed in legislation applying to publicwater supply safety.
The hepatotoxicity induced by cyanotoxins is well documented, andinformation on their genotoxicity and potential carcinogenicity can befound in the review paper by Zegura et al. (2011). The main mechanismassociatedwith potential carcinogenic activity of microcystins is the inhi-bition of protein phosphatases, leading to a cellular hyperphosphorylationstate (Amado andMonserrat, 2010). The in vitro studieswithmammaliancells and in vivo rodent showed that microcystins induce ROS formation(Zegura et al., 2004, 2008), DNA damage (Lankoff et al., 2004; Palus etal., 2007; Zegura et al., 2008; Gaudin et al., 2008) andmicronuclei forma-tion (Zhan et al., 2004; Zegura et al., 2009). These toxins also interferewith the DNA damage repair pathway and double strand breaks contrib-uting to genetic instability of the exposed cells (Zegura et al., 2011). In ad-dition, microcystins induce the expression of tumor necrosis factor α (atumor promoter) and early-response genes (proto-oncogenes),suggesting a possible mechanism for their tumor-promoting activity(Sueoka et al., 1997).
To evaluate genotoxic substances, assays with vascular plants arewidely recommended by the United Nations Environment Programme-UNEP. Cytotoxicity tests using plant systems in vivo, such as Alliumcepa L., are validated by various researchers that have carried out animaltests in vitro together with the plant tests, obtaining similar results(Vicentini et al., 2001; Teixeira et al., 2003; Lubini et al., 2008). Lemeand Marin-Morales (2009) reviewed the Allium cepa test and its usefor testing environmental contamination, finding that vascular plantsare recognized as excellent genetic models for detecting environmentalmutagens; frequently used in monitoring studies. Tedesco andLaughinghouse (2012) showed that the A. cepa test is among the plantspecies used to evaluate DNA damages, chromosomal alterations, anddisturbances in the mitotic cycle. El-Shahaby et al. (2003) reinforcethe importance of using the Allium cepa test for detecting toxicity/genotoxicity, and the importance as an indicator for human health. Astudy conducted by McElhiney et al. (2001) showed that the differ-ence in the toxicities of microcystin-LR, -RR, and -LF on the growthof mustard seeds is significant. Wang et al. (2011) investigated thetoxic effects of microcystin-LR on lettuce (Lactuca sativa L.) andshowed that seedling growth was more sensitive to toxicant expo-sure than seed germination.
The aim of this study was to evaluate the possible ecotoxicological,cytotoxic, and genotoxic effects provoked by methanolic extracts oftwo strains of Microcystis aeruginosa (one positive for microcystin[NPLJ-4] and another negative for microcystin [NPCD-1]), using theAllium cepa test system.
2. Materials and methods
2.1. Cultivation of cyanobacterial strains
The Brazilian strain Microcystis aeruginosa NPCD-1 (non microcystin-producer) was isolated from a sewage treatment plant, Cidade deDeus, RJ, Brazil and NPLJ-4 (microcystin-producer) was isolated fromJacarepaguá lagoon, RJ, Brazil. Both strains were supplied by Dr. SandraM.F.O. Azevedo from the Instituto de Biofísica Carlos Chagas Filho,
Centro de Ciências da Saúde, Universidade Federal do Rio de Janeiro.The monospecific cultures were cultivated in 8 L of ASM-1 liquidmedium. Cultures were sparged with sterile air and maintained at23 °C under constant illumination by white fluorescent light(40 μmol m−2 s−1) for 20 days.
2.2. Enzyme-linked immunosorbent assays (ELISA)
Lyophilized cells were mixed with distilled water, microwaved for1 min, and then centrifuged for 15 min at 10,000 ×g. The supernatantwas collected and analyzed by an ELISA assay, using microplate kitsfor microcystins (Beacon Analytical Systems Inc., Portland, ME, USA)following the manufacturer's recommendations, with at least threereplicates. The detection limit of this method was 0.1 μg L−1.
2.3. Metabolite extraction
For metabolite extraction, M. aeruginosa NPCD-1 and NPLJ-4 cul-tures were harvested by centrifugation at 9000 ×g at 4 °C. Cellswere lyophilized (0.0135 g for NPCD-1 and 0.04 g for NPLJ-4) andextracted in methanol, using glass beads (ϕ 3 mm). The suspensionwas agitated for 1 min and afterwards, centrifuged at 9,000 ×g at4 °C. The supernatant was collected and evaporated. The dried extractwas kept at −20 °C for further analysis.
2.4. High performance liquid chromatography – mass spectrometry(HPLC-MS)
The dried methanolic extracts of M. aeruginosa NPLJ-4 and NPCD-1were resuspended each in 1 ml of methanol. Then the mixture was ho-mogenized for 30 s and filtered (0.22 μ). The samples were subjected toLC-MS and analysis of the extract composition was achieved using anAgilent 6410 Triple Quadrupole LC/MS equipped with Agilent 1200 Se-ries Binary Pump SL pumping system and Agilent 1200 Autosampler(Agilent Technologies Inc. Santa Clara, CA, USA). Data acquisition andanalysis were performed using Agilent MassHunterWorkstation soft-ware. The electrospray ionization (ESI) sourcewas employed in positivemode, and the source parameters for the MS detection were optimizedto maximize sensitivity for cyanopeptides, set as follows: drying gaswas nitrogen heated at 350 °C, gas flow was 7 L min−1, capillary volt-age was held at 4000 V. The MS/MS parameters were similarly opti-mized for cyanopeptides sensitivity and set as follows: dwell time was200 ms, fragmentor voltage was set at 200 V, collision gas was nitrogenand collision energy was set at 30 eV. The MS system was operated infull scan and multiple reaction mode (MRM). The mobile phase A con-sisted of ultrapure water (Milli-Q) and mobile phase B consisted ofmethanol, both containing 0.1% (v/v) of formic acid. The flow rate was0.3 ml min−1. The MS optimization was performed via direct infusionof the cyanobacterial extract into the ESI source of the mass spectrom-eter. Molecular ions were scanned in the range of 100 to 1200 Da.
2.5. Exposure of Allium cepa to cyanobacterial extracts
The dried methanolic extracts from NPLJ-4 and NPCD-1 strains weredissolved in distilledwater to afinal concentration of 6.58 mg L−1. Thesestock solutions for both strains were further diluted in distilled waterinto three test concentrations (for exposing the onions): 6.58 mg L−1
(Dilution 1—D1), 0.658 mg L−1 (Dilution 2—D2), and 0.0658 (Dilution3—D3). Seven Allium cepa bulbs were used (previously cultivated with-out the use of agrochemicals) for each treatment plus the negative con-trol. The external protective layers and the old rootlet tissue wereremoved, followed by the washing of each bulb before the test. The A.cepa bulbswere germinated in distilledwater for 24 h in a dark germina-tion chamber at 25 °C/22.5 °C 12/12 h. Afterwards, each bulb was placedin their respective test (D1, D2 or D3)/negative control for another 48 h.

182 H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
Bulbs that did not grow and/or outliers were removed, thus 4 bulbs foreach treatment were used in the study.
2.6. Determination of rootlet number and length / Fixation of the rootlets
At the end of the treatment, the A. cepa bulbs were taken out of theliquid treatment and the number of rootlets per bulb was counted(3–5 rootlets per bulb to determine the mean root length); rootletswere then removed. These were fixed in ethanol-acetic acid (3:1)and maintained refrigerated (4 °C) until slide preparation.
2.7. Slide preparation
To analyze cell division, micronuclei, and chromosomal aberra-tions, the rootlets were removed from the ethanol-acetic acid andplaced in 1 N HCl in a water bath at 60 °C for 10 min for hydrolysis.After hydrolysis, the rootlets were placed on ice for 10 min, and after-wards treated, in the dark, with the Feulgen and Fast-Green stains for10 min each according to Ma et al. (1995) and the squashing tech-nique was applied on the meristematic region following Guerra andSouza (2002). After squashing, the slide was dried using dry ice to re-move the cover slip. With cover slips removed, the slides were passedthrough a sequence of dehydration/staining made up of ethanol at in-creasing concentrations of 70, 90, and 100% (with Fast-Green at10 mg ml−1). Next, the slides were quickly immersed in ethanol100% and, finally, in xylol 100% to remove the rest of the stains andtotal dehydration, respectively. Entellan® (Merck Chemicals) wasadded, together with a cover slip to make the final permanent slide.
2.8. Microscope analysis
Slideswere scored (blind)with aminimumof 500 cells per bulb, usinga compound light microscope at 400×magnification for determining cellcycle phases, mitotic index, micronuclei, and chromosomal aberrations.
2.9. Cell cycle and mitotic index
The number of cells in each of the cell cycle phases [interphaseand division (prophase, metaphase, anaphase, and telophase)] wastallied. The mitotic index was determined by dividing the numberof cells in mitosis (prophase, metaphase, anaphase, and telophase)by the total number of cells observed and multiplying the result by100 (Pires et al., 2001; Tedesco and Laughinghouse, 2012).
2.10. Micronuclei and chromosomal aberrations
The number of micronuclei (MN) and chromosomal aberrations(CA) (sticky chromosomes, mitosis-C, anaphasic bridges, chromosomalfragments, and laggard chromosomes) were reported as the total num-ber of cells observed and multiplying the result by 1000. The meannumber of micronuclei and chromosomal aberrations were used inthe statistical analyses (Fiskejö, 1993, 1994). Bi-nucleated cells andbuds were also accounted for, but not included in the statistics.
2.11. Statistics
The non-parametric Mann–Whitney test followed by the Dunn'spost test was used to compare the values in relation to the negativecontrol. The significance level was pb0.05. The program GraphPadPrism (GraphPad Software, Inc., San Diego, California) was used toperform statistical analyses and create graphs.
3. Results
The M. aeruginosa NPLJ-4 cell extract elicited a positive immu-noreaction for microcystins in the ELISA test (171.71 μg ml−1), while
for the extract of strain NPCD-1 the result was negative. LC-MS was ap-plied to obtain, in high purity, the major microcystin of M. aeruginosaNPLJ-4, which was identified as MCYST-XR based on the study of Silva-Stenico et al. (2009). The presence of microcystin-XR (m/z 1037) and-LR (m/z 1023) were confirmed after fragmentation of individual peaksthat showed the Adda residue [C9H11O]+ atm/z 135 (Fig. 1A).
For aeruginosin analysis, the MS/MS spectrum of the protonatedmolecule at m/z 742 showed the characteristic ions at m/z 86, 100 and140, which corresponded to the leucine immonium, argal fragmentand the amino acid 2-carboxy-6-hydroxyoctahydroindole (Choi), re-spectively, all of which are found in aeruginosin molecules (Fig. 1B).
The results obtained from the analysis of the cell cycle and occur-rence of MN and CA, from the root-tip cells treatedwithmethanolic ex-tracts from the strainsM. aeruginosa NPLJ-4 and NPCD-1 populations indifferent concentrations are presented in Table 1 and Figs. 2–4.
The results presented in Table 1 show that there were cells in allphases of cell division. The highest number of cells in prophase and ana-phase was observed in bulbs treated withM. aeruginosa NPCD-1 D3. Theleast amount of cells in division was observed in bulbs treated withM. aeruginosa NPCD-1 D2. Furthermore, structural CA occurred in thecells observed from the rootlets treated with extracts of Microcystisaeruginosa. The mitotic index values ranged from 1.4 to 2.8% for NPCD-1; 3.2 to 4.2% for NPLJ-4, and in the controls (C1 and C2) it varied from1.5 to 2%, respectively. M. aeruginosa NPCD-1 D1 had the highest meanof MN (8.24), while M. aeruginosa NPCD-1 D3 had the lowest mean(2.14). M. aeruginosa NPLJ-4 D3 presented the highest MI mean (4.23),whileM. aeruginosaNPCD-1 D1 presented the lowest (1.36). The highestpercentage of CA was found inM. aeruginosa NPLJ-4 D3 (1.73), while M.aeruginosaNPLJ-4D1presented the lowest (0.42) (Table 1). Bulbs treatedwithM. aeruginosa NPLJ-4 D3 contained the highest number of MN.
Fig. 2 presents the MI (A), the MN (B), and the CA (C) induced bythe tested aqueous extracts. The treatment with the extracts of bothstrains increased, although not significantly, the MI at the lowest con-centration. As the extract concentrations increased, the MI forM. aeruginosa NPLJ-4-treated cell and the MI for NPCD-1-treatedcells decreased. In regards to the frequency of MN, a concentration-dependent increase for M. aeruginosa NPCD-1 was observed, with asignificant increase compared to the control in the highest concentra-tion (pb0.01). For M. aeruginosa NPLJ-4, a slight rise in MN frequencywas observed only in the lowest dosage, although not significant. Fur-thermore, the CA frequency had a significant increase in both strainsat the lowest concentration (pb0.05 for M. aeruginosa NPCD-1 andpb0.01 for M. aeruginosa NPJL-4), followed by a decrease in thehigher concentrations.
Fig. 3 presents the frequencies of sticky chromosomes/mitosis-C(A) and anaphasic bridges/laggard chromosomes (B) induced by thetreatments with the tested aqueous extracts. The treatments with ei-ther cyanobacterial strain significantly increased the index of stickychromosomes/mitosis-C in the lowest concentration (pb0.05 forM. aeruginosa NPCD-1 and pb0.01 for M. aeruginosa NPJL-4). However,as the extract concentrations increased, a decline for M. aeruginosaNPLJ-4-treated cells and a stabilization for NPCD-1-treated cells wereobserved. Concerning anaphasic bridges/laggard chromosomes, therewas a significant increase for M. aeruginosa NPLJ-4 (pb0.05) and asmall rise for M. aeruginosa NPCD-1 in the lowest concentrations,followed by a significant decrease in the higher concentrations.
Fig. 4 shows the rootlet growth (A) and the number of rootlets (B)with regards to the tested aqueous extracts. The treatment amplifiedthe rootlet growth in the lowest concentration of M. aeruginosa NPCD-1, followed by a decline at higher dosages, which was common withthe methanolic extract treatment usingM. aeruginosa NPLJ-4, in the sig-nificance limit for a response-concentration correlation (r=−0.9163and p=0.0837 for M. aeruginosa NPLJ-4).Pertaining to the number ofrootlets, there was a small increase in number with the M. aeruginosaNPCD-1 treatments in the lowest concentrations, followed by a decline,then a slight increase in the highest concentration. Relating to M.

Fig. 1. Fragment mass spectra of selected microcystin-XR (m/z 1037) produced by M. aeruginosa NPLJ-4 (A) and aeruginosin (m/z 742) produced by NPCD-1 (B).
183H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
aeruginosa NPCD-1, in the lowest concentrations, there was a decreasein rootlet number, followed by a concentration-dependent increase.
In this study, the following effects were observed: chromosomaldisruptions during metaphase (Figs. 5A, B and 6C, D, E, F), anaphasicbridge formation (Fig. 6B), chromosomal disruptions during anaphase
Table 1Cell cycle profile (interphase, prophase, metaphase, anaphase, and telophase), micronucleuswith aqueous extracts of Microcystis aeruginosa. Data were generated as averages of 4 repli
Treatment NPCD-1 NPCD-1 NPCD-1
Dillution D1 D2 D3Total cell number 2277 2238 2601Prophase 12 13 33Metaphase 6 4 14Anaphase 4 4 9Telophase 9 12 22Micronucleus (MN) 18 11 6Bud 1 0 0Sticky-chromosome/mitosis-C
16 14 22
Anaphasic-bridge/laggard
6 5 15
Binucleate cell 5 7 6Apoptotic body 3 0 0Necrosis 4 0 0MN/1000 8.24 4.73 2.14Mitotic index % (MI) 1.36 1.55 2.79Chromosomeaberration % (CA)
0.90 0.83 1.36
NPCD-1=extract containing aeruginosin.NPLJ-4=extract containing microcystin.D: concentrations of cells in mg L−1: Dilutions D3: 0.0658; D2: 0.658; D1: 6.580.The treatments C1 and C2 are controls, using (distilled) water.
(Fig. 5F), micronuclei (Fig. 5C, D, E), morphological alterations (Figs. 5and 6), and apoptosis (Fig. 6A). In addition, apoptotic bodies and cellsin necrosis were observed in the treatment with NPCD-1 (Table 1).The detected alterations undoubtedly indicate the presence of toxicand/or genotoxic substances in both extracts.
, chromosome aberrations, and cytotoxicity (necrosis and apoptosis) in rootlets treatedcations of each treatment.
NPLJ-4 NPLJ-4 NPLJ-4 C1 C2
D1 D2 D3 0 03887 2374 2221 4738 220799 28 37 32 1832 17 16 18 315 11 19 6 245 18 23 14 179 6 9 1 05 1 0 0 16 8 12 0 1
15 8 28 0 0
4 7 7 1 00 0 0 0 00 0 0 0 02.37 2.50 4.52 0.20 03.97 3.16 4.23 1.53 2.030.42 0.68 1.73 0 0.05

Fig. 2.Mitotic index (A), frequency of micronuclei (B), and chromosome aberration (C)induced by the tested methanolic cyanobacterial extracts.
Fig. 3. Indices of sticky-chromosomes/mitosis-C (A) and anaphasic bridges/laggardchromosomes (B) induced by the tested methanolic cyanobacterial extracts.
Fig. 4. Root length (A) and root number (B) according to the tested methanoliccyanobacterial extracts.
184 H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
4. Discussion
The Allium cepa test is important since it is an excellentmodel in vivo,where the roots grow directly in contact with the substance of interest(i.e. effluent, toxin, chemical mix, etc.) which is being tested, enablingputative damage to the eukaryotic DNA to be predicted. Therefore,these data can be extrapolated to other animal and plant diversity(Tedesco and Laughinghouse, 2012). In this manner, the Allium cepatest has been shown to be efficient as a pre-screening technique for stud-ies concerning and assessing (geno)toxicity (Lubini et al., 2008; Bagatiniet al., 2009).
Plants are considered to be similar tomammals for detecting cytoge-netic alterations induced by chemical substances (Chauhan et al., 1999).Fiskejö (1994) commented that even though plantmetabolism is differ-ent than animal metabolism, the A. cepa test is an excellent parameterfor cytotoxicity analysis,where other researchers report on its efficiencyto analyze genotoxicity (Tedesco and Laughinghouse, 2012).
The cyanobacterial extracts of strains NPLJ-4 and NPCD-1, containingmicrocystin and aeruginosin, respectively, naturally present in aquaticenvironments, were chemically characterized and evaluated for their cy-totoxicity and genotoxicity. The intracelullar extract of M. aeruginosaNPLJ-4 contained twomicrocystin variants (‐XR and -LR), where the var-iant -XR is the major component. This strain is also an aeruginosin pro-ducer (Silva-Stenico et al., 2011), but according to the purificationsystem used for microcystin in this study, the presence of aeruginosinwas not detected using LC-MS. The acute toxicity tests using miceshowed that microcystin produced by this strain corresponds to the de-gree of high toxicity (Kuroda et al., 2007). The strain NPCD-1 was con-firmed as a non-microcystin producer, but the intracellular extract ofM. aeruginosaNPCD-1 contained a high concentration of the protease in-hibitor aeruginosin (Silva-Stenico et al., 2011). These molecules arefound in aquatic environments, in particular in cyanobacterial blooms.Aeruginosins were already tested against thrombin, trypsin and plasminproteases. It is known that thrombin is the main factor in blood clotting,activating platelets and acting as an inducer of mitosis for various celltypes. For these reasons, this protein has become a special target forthe isolation of natural antithrombotic agents (Toyota et al., 2005; Ella-Menye et al., 2008). Despite its importance, there is still a lack of studiesregarding the potential genotoxicity of these molecules. Therefore, the

Fig. 5. Microphotographs of the morphological alterations of Allium cepa cells treated with methanolic extracts ofM. aeruginosaNPCD-1: chromosomal disruptions in metaphase (A and B);micronuclei (C, D, and E); chromosomal disruptions in anaphase (F).
185H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
meristematic root-tip cells of Allium cepa (onion) were used as a planttest system, through the analysis of cell cycle disturbances, MN, CA,apoptosis and necrosis, to determine the ecotoxicological, genotoxic,and cytotoxic effects of these strains.
From the results obtained and presented in Table 1, it was observedthat the extract of M. aeruginosa NPCD-1 formed apoptopic bodies,which did not occur in the other treatments. However, this was also pro-portional to the cells that suffered necrosis. This demonstrates the rela-tionship of both processes, apoptosis and necrosis, although distinctcellular processes. Somehow, in D1 of theM. aeruginosa extract NPCD-1the apoptopic mechanism was induced. This aspect was observed inthe treatment with extract NPCD-1, which also had a greater decreasein mitotic index when compared to the control with water. This hasalso been observed by Sousa et al. (2010) and Lubini et al. (2008)
when studying Cymbopogon and Psychotria, respectively. Sousa et al.(2010) observed apoptopic bodies in only one of the studied extracts ofCymbogon citratus (lemon grass), associated to programmed cell death.Interestingly, cyanobacterial extracts have been frequently shown tohave apoptogenic activity, even in strains cultured for decades. This activ-ity has been shown to occur frequently in the Microcystis genus (Oftedalet al., 2011).
Analyzing the results of the MI and inhibition of rootlet growth,we observed that the M. aeruginosa NPLJ-4 methanolic extract had atendency to increase the number of cell divisions as the concen-tration increased. However, the extract of M. aeruginosa NPCD-1,showed a tendency to reduce the number of cell divisions, with asignificant increase in the number of MN (pb0.01 for the mostconcentrated).
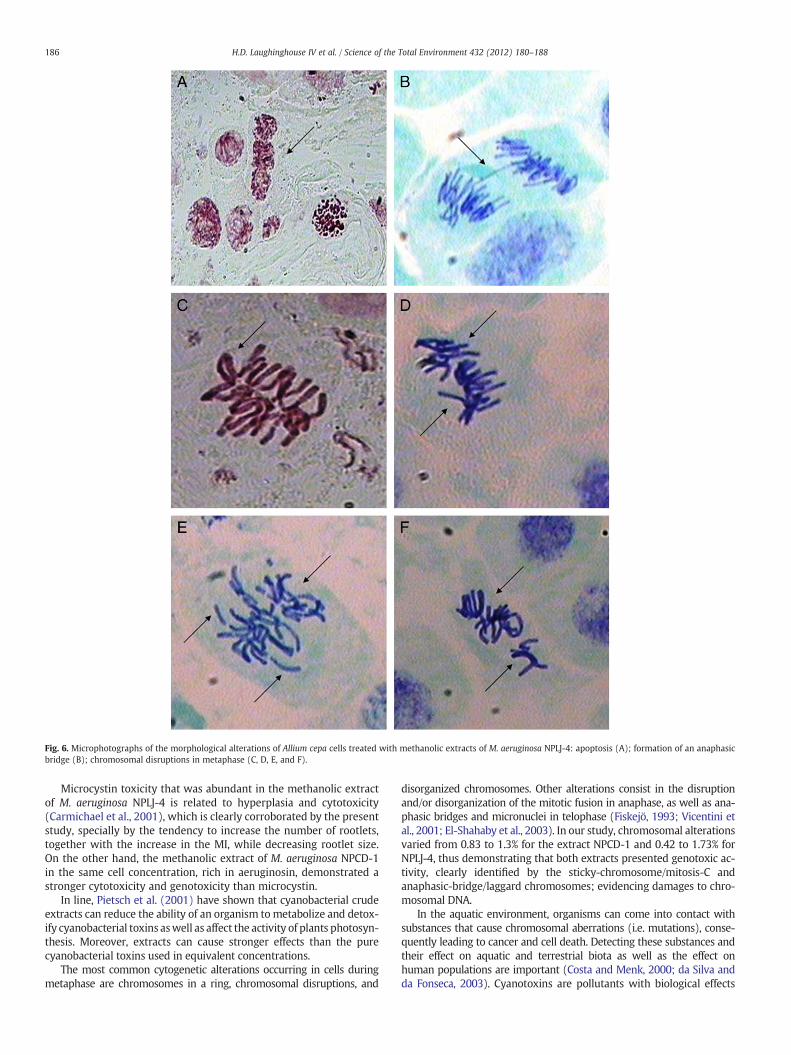
Fig. 6. Microphotographs of the morphological alterations of Allium cepa cells treated with methanolic extracts of M. aeruginosa NPLJ-4: apoptosis (A); formation of an anaphasicbridge (B); chromosomal disruptions in metaphase (C, D, E, and F).
186 H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
Microcystin toxicity that was abundant in the methanolic extractof M. aeruginosa NPLJ-4 is related to hyperplasia and cytotoxicity(Carmichael et al., 2001), which is clearly corroborated by the presentstudy, specially by the tendency to increase the number of rootlets,together with the increase in the MI, while decreasing rootlet size.On the other hand, the methanolic extract of M. aeruginosa NPCD-1in the same cell concentration, rich in aeruginosin, demonstrated astronger cytotoxicity and genotoxicity than microcystin.
In line, Pietsch et al. (2001) have shown that cyanobacterial crudeextracts can reduce the ability of an organism to metabolize and detox-ify cyanobacterial toxins aswell as affect the activity of plants photosyn-thesis. Moreover, extracts can cause stronger effects than the purecyanobacterial toxins used in equivalent concentrations.
The most common cytogenetic alterations occurring in cells duringmetaphase are chromosomes in a ring, chromosomal disruptions, and
disorganized chromosomes. Other alterations consist in the disruptionand/or disorganization of the mitotic fusion in anaphase, as well as ana-phasic bridges and micronuclei in telophase (Fiskejö, 1993; Vicentini etal., 2001; El-Shahaby et al., 2003). In our study, chromosomal alterationsvaried from 0.83 to 1.3% for the extract NPCD-1 and 0.42 to 1.73% forNPLJ-4, thus demonstrating that both extracts presented genotoxic ac-tivity, clearly identified by the sticky-chromosome/mitosis-C andanaphasic-bridge/laggard chromosomes; evidencing damages to chro-mosomal DNA.
In the aquatic environment, organisms can come into contact withsubstances that cause chromosomal aberrations (i.e. mutations), conse-quently leading to cancer and cell death. Detecting these substances andtheir effect on aquatic and terrestrial biota as well as the effect onhuman populations are important (Costa and Menk, 2000; da Silva andda Fonseca, 2003). Cyanotoxins are pollutants with biological effects

187H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
that are not totally understood. Although there is legislation limiting theconcentration of microcystins in water, the numerous other bio-compounds produced by cyanobacteria are still not legislated.
Our results demonstrated the existence of genotoxic activity of thecyanobacterial extracts on the Allium cepa cells in division by the inhibi-tion or increase of the observed MI or MN, thus demonstrating the ca-pability of using this method as a pre-screening technique duringcyanobacterial blooms, assessing the genotoxicity and cytotoxicity ofthese blooms. Moreover, significant differences between the controland the treatments among the studied strains and dilutions were ob-served, illustrating different actions of microcystin and aeruginosin, in-clusively indicating genotoxicity for aeruginosin, which previously hadnot been reported in literature.
In line with the observation the present results, Oftedal et al. (2011)screened the apoptosis activity of 83 cyanobacterial extracts against im-mune and hematopoietic human cancerous cells and have shown thatcyanobacteria produce a diversity of still uncharacterized bioactive com-pounds that have unpredictable effect over biological systems.
As evidenced by the positive result in the micronucleus test in thisstudy, strains producing aeruginosin should be further studied to un-derstand their full genotoxicity. As time goes by, human populationsare more prone to ingesting large quantities of these toxins, especiallyduring blooms. Therefore, carrying out studies directed to the under-standing of cyanobacterial genotoxicity and carcinogenicity is extreme-ly pertinent.
5. Conclusions
Inmany cases, in developing nations, such as Brazil, funds for impor-tant scientific research dealing with water quality are scarce. Further-more, the use of animals in research is becoming an ethical problem.Thus finding techniques, which can answer our hypothesis in a moreeconomical and bioethical manner, is very important. This study findsthat the use of the Allium cepa test is an excellent bioindicator of chro-mosomal alterations caused by two different bioactive compounds pro-duced by Microcystis aeruginosa. With correct interpretation, this testcan be used to assess the environmental impact that a cyanobacterialbioactive compound could cause on the environment. Moreover, resultsof the cyanobacterial extracts tested indicate the presence of a diversityof bioactive compounds that are likely to exert toxicity by differentmechanisms, many still uncharacterized.
Acknowledgements
This study was supported by grants from The State of São PauloResearch Foundation (FAPESP 2009/05474-5). M.F. Fiore would liketo thank CNPq for a research fellowship (308299/2009-4). M.E.Silva-Stenico was the recipient og a postdoctoral fellowship fromFAPESP (2010/09867-9) and CNPq (151931/2010-0). We thank Prof.Valdemar Tornisielo for LC-MS analysis support in the Laboratory ofEcotoxicology, CENA-USP, Brazil. We are also indebted to three anon-ymous reviewers that revised an earlier version of this manuscript.
References
Amado LL, Monserrat JM. Oxidative stress generation by microcystins in aquatic ani-mals: why and how. Environ Int 2010;36:226–35.
Azevedo SMFO. Cianobactérias tóxicas: o que precisamos fazer. In: de A. Mariath JE, dosSantos RP, editors. Os avanços da Botânica no início do século XXI: morfologia,fisiologia, taxonomia, ecologia e genética. Porto Alegre, Brazil: Sociedade Botânicado Brasil; 2006. p. 265–8.
Azevedo SMFO, Carmichael WW, Jochimsen EM, Rinehart KL, Lau S, Shaw GR, et al.Human intoxication by microcystins during renal dialysis treatment in Caruaru-Brazil. Toxicology 2002;181(182):441–6.
Bagatini MD, Vasconcelos TG, Laughinghouse IV HD, Martins AF, Tedesco SB. Bio-monitoring hospital effluents by the Allium cepa L. test. Bull Environ Contam Toxicol2009;82:590–2.
Carmichael WW. Blue-Green Algae: An Overlooked Health Threat. Health & Environ-ment Digest., 5(4). Freshwater Foundation; 1991. p. 1–4.
Carmichael WW, Azevedo SMFO, An JS, Molica RJR, Jochimsen EM, Lau S, et al. Humanfatalities from cyanobacteria: chemical and biological evidence for cyanotoxins.Environ Health Perspect 2001;109(7):663–8.
Chauhan LKS, Saxena PN, Gupta SK. Cytogenetic effects of cypermethrin and fenvalerateon the root meristem cells of Allium cepa. Environ Exp Bot 1999;42:181–9.
Costa RMA, Menk CFM. Biomonitoramento de mutagênese ambiental. BiotecnologiaCienc Desenvolv 2000;2(12):24–6.
da Silva J, da Fonseca MB. Estudos Toxicológicos no Ambiente e na Saúde Humana. In:da Silva J, Erdtmann B, Henriques JAP, editors. Genética Toxicológica. Porto Alegre:Alcance; 2003. p. 69–84. Ed.
Ella-Menye J-R, Nie X, Wang G. Synthesis of octahydropyrano[3,2-b]pyrrole-2-carboxylicacid derivatives from D-mannose. Carbohydr Res 2008;343:1743–53.
El-Shahaby AO, Abdel Migid HM, Soliman MI, Mashaly IA. Genotoxicity screening of in-dustrial wastewater using the Allium cepa chromosome aberration assay. Pak J BiolSci 2003;6:23–8.
Fiskejö G. The Allium test in wastewater monitoring. Environ Toxicol Water Qual1993;8:291–8.
Fiskejö G. The Allium Test II: assessment of chemical's genotoxic potential by recordingaberrations in chromosomes and cell divisions in root tips of Allium cepa L. EnvironToxicol Water Qual 1994;9:235–41.
Gaudin J, Huet S, Jarry G, Fessard V. In vivo DNA damage induced by the cyanotoxinmicrocystin-LR: comparison of intra-peritoneal and oral administrations by useof the comet assay. Mutat Res Genet Toxicol Environ Mutagen 2008;652:65–71.
Guerra M, Souza MJ. Como observar cromossomos—um guia de técnicas em citoge-nética vegetal, animal e humana. Ribeirão Preto: FUNPEC; 2002. 131 pp.
Humpage AR, Falconer IR. Microcystin-LR and liver tumor promotion: effects on cyto-kinesis, ploidy, and apoptosis in cultured hepatocytes. Environ Toxicol 1999;14:61–75.
Hunter PR. Cyanobacterial toxins and human health. J Appl Microbiol Symp Suppl1998;84:35S–40S.
Ishida K, Welker M, Christiansen G, Cadel-Six S, Bouchier C, Dittmann E, et al. Plasticityand evolution of aeruginosin biosynthesis in cyanobacteria. Appl EnvironMicrobiol 2009;75:2017-20126.
Kuroda EK, Minillo A, Rocha O, Rodrigues Filho E, Bernardo LD. Avaliação da toxicidadeaguda de uma cepa de Microcystis spp. por meio de testes com camundongos. EngSanit Ambient 2007;12:24–31.
Lankoff A, Krzowski L, Glab J, Banasik A, Lisowska H, Kuszewski T, et al. DNA damageand repair in human peripheral blood lymphocytes following treatment withmicrocystin-LR. Mutat Res Genet Toxicol Environ Mutagen 2004;559:131–42.
Leme DM, Marin-Morales MA. Allium cepa test in environmental monitoring: a reviewon its application. Mutat Res 2009;682:71–81.
Lubini G, Fachinetto JM, Laughinghouse IV HD, Paranhos JT, Silva ACF, Tedesco SB. Ex-tracts affecting mitotic division in root-tip meristematic cells. Biologia 2008;63:647–51.
Ma T-H, Xu Z, Xu C, McConnell H, Rabago EV, Arreola GA, et al. The improvedAllium/Vicia root tip micronucleus assay for clastogenicity of environmental pollut-ants. Mutat Res 1995;334:185–95.
McElhiney J, Lawton LA, Leifert C. Investigations into the inhibitory effects ofmicorcystins on plant growth, and the toxicity of plant tissue following exposure.Toxicon 2001;39:1411–20.
Oftedal L, Skjærven KH, Coyne RT, Edvardsen B, Rohrlack T, Skulberg OM, et al. Theapoptosis-inducing activity towards leukemia and lymphoma cells in a cyanobacterialculture collection is not associated with mouse bioassay toxicity. J Ind MicrobiolBiotechnol 2011;38:489–501.
Palus J, Dziubaltowska E, Stanczyk M, Lewinska D, Mankiewicz-Boczek J, Izydorczyk K,et al. Biomonitoring of cyanobacterial blooms in Polish water reservoir and thecytotoxicity and genotoxicity of selected cyanobacterial extracts. Int J Occup MedEnviron Health 2007;20:48–65.
Pietsch C, Wiegand C, Amé MV, Nicklisch A, Wunderlin D, Pflugmacher S. The Effects ofa cyanobacterial crude extract on different aquatic organisms: evidence forcyanobacterial toxin modulating factors. Environ Toxicol 2001;16:535–42.
Pires NM, Souza IRP, Prates HT, Faria TCL, Filho IAP, Magalhães PC. Efeito do extratoaquoso de leucena sobre o desenvolvimento, índice mitótico e atividade da perox-idase em plântulas de milho. Rev Bras Fisiol Veg 2001;13:55–65.
Roset J, Aguayo S, Muñoz MJ. Detección de cianobacterias y toxinas. Una revisión. RevToxicol 2001;18:65–71.
Silva-Stenico ME, Neto RS, Alves IR, Moraes LAB, Shishido TK, Fiore MF. Hepatotoxinmicrocystin—LR extraction optimization. J Braz Chem Soc 2009;20:535–42.
Silva-Stenico ME, Silva CSP, Lorenzi AS, Shishido TK, Etchegaray A, Lira SP, et al. Non-ribosomal peptides produced by Brazilian cyanobacterial isolates with antimicrobialactivity. Microbiol Res 2011;166:161–75.
Sousa SM, Silva PS, Viccini LF. Cytogenotoxicity of Cymbopogon citratus (DC) Strapf.(lemon grass) aqueous extracts in vegetal test systems. An Acad Bras Cienc2010;82:305–11.
Sueoka E, Sueoka N, Okabe S, Kozu T, Komori A, Ohta T, et al. Expression of the tumornecrosis factor a gene and early response genes by nodularin, a liver tumor promoter,in primary cultured rat hepatocytes. J Cancer Res Clin Oncol 1997;123:413–9.
Takenaka S. Effects of L-cysteine and reduced glutathione on the toxicities ofmicrocystin-LR: the effect for acute liver failure and inhibition of phosphatase 2Aactivity. Aquat Toxicol 2000;48:65–8.
Tedesco SB, Laughinghouse IV HD. Bioindicator of genotoxicity: the Allium cepa test. In:Srivastava JK, editor. Environmental Contamination. Rijeka: InTech Publisher;2012. p. 137–56.
Teixeira RO, Camparoto ML, Mantovani MS, Vicentini VEP. Assessment of two medici-nal plants, Psidium guajava L. and Achillea millefolium L. in in vivo assays. GenetMol Biol 2003;26:551–5.

188 H.D. Laughinghouse IV et al. / Science of the Total Environment 432 (2012) 180–188
Toyota E, Sekizaki H, Takahashi Y, Itoh K, Tanizawa K. Amidino-containing schiff basecopper (II) and iron (III) chelates as a thrombin inhibitor. Chem Pharm Bull2005;53:22–6.
Vicentini VEP, Camparoto ML, Teixeira RO, Mantovani MS. Averrhoa carambola L.,Syzygium cumini (L.) Skeels and Cissus sicyoides L.: medicinal herbal tea effectson vegetal and test systems. Acta Sci 2001;23:593–8.
Wang Z, Xiao B, Song L, Wu X, Zhang J, Wang C. Effects of microcystin-LR, linearalkylbenzene sulfonate and their mixture on lettuce (Lactuca sativa L.) seeds andseedlings. Ecotoxicology 2011;20:803–14.
Yoshizawa S, Matsushi R, Watanabe MF, Harada KI, Ichihara A, Carmichael WW, et al.Inhibition of protein phosphatases by microcystins and nodularin associatedwith hepatotoxicity. J Cancer Res Clin Oncol 1990;116:609–14.
Zegura B, Lah TT, Filipic M. The role of reactive oxygen species in microcystin-LR-induced DNA damage. Toxicology 2004;200:59–68.
Zegura B, Volcic M, Lah TT, Filipic M. Different sensitivities of human colon adenocarci-noma (CaCo-2), astrocytoma (IPDDC-A2) and lymphoblastoid (NCNC) cell lines tomicrocystin-LR induced reactive oxygen species and DNA damage. Toxicon2008;52:518–25.
Zegura B, Sever N, Koloŝa K, Filipic M. The influence of microcystin-LR on HepG2 cellproliferation and micronuclei formation. Proceedings of Tenth International Con-ference on Environmental Mutagenesis: The Renaissance of Environmental Muta-genesis. Firenze, Italy 20–25 August; 2009. p. 136.
Zegura B, Straser A, FilipicM. Genotoxicity and potential carcinogenicity of cyanobacterialtoxins —a review. Mutat Res 2011;727:16–41.
Zhan L, Sakamoto H, Sakuraba M, Wu DS, Zhang LS, Suzuki T, et al. Genotoxicity ofmicrocystin-LR in human lymphoblastoid TK6 cells. Mutat Res 2004;557:1–6.
Related Documents











