Biomass-specific respiration rates of benthic meiofauna: Demonstrating a novel oxygen micro-respiration system Leon Moodley a, ⁎ , Maaike Steyaert b , Eric Epping c , Jack J. Middelburg a , Magda Vincx b , Pim van Avesaath a , Tom Moens b , Karline Soetaert a a Netherlands Institute of Ecology, (NIOO-KNAW), Korringaweg, 4401 NT Yerseke, The Netherlands b Marine Biology Section, Department of Biology, Ghent University, Krijgslaan 281/S8, 9000 Ghent, Belgium c Royal Netherlands Institute for Sea Research, P.O. Box 59, 1790 AB Den Burg, Texel, The Netherlands Received 11 October 2007; received in revised form 13 December 2007; accepted 17 December 2007 Abstract Meiofauna (small-sized Metazoa and Foraminifera) may constitute a significant part of seafloor biomass and potentially play an important role in benthic metabolism. However, respiration measurements are limited and the methods used are diverse together complicating comparison or upscaling. Here we describe a novel glass micro-respiration chamber used to perform non-invasive measurements (built-in oxygen-sensitive fluorogenic membrane and stirrer) and together with direct organic carbon measurements report initial biomass-specific respiration rates of common intertidal meiofauna. Results indicate large differences between respiration rates of different taxa (biomass 0.7–5.2 μg C per individual) but very similar organic carbon biomass-specific respiration rates (1.6–2.5 μl O 2 h - 1 mgC - 1 or on average 2.0 ± 0.3 μl O 2 h - 1 mgC - 1 ). This new, rapid and accurate method allows the study of metabolic allometry of the different small-sized taxa and determining their functional role in benthic metabolism. © 2007 Elsevier B.V. All rights reserved. Keywords: Benthic; Foraminifera; Meiofauna; Optodes; Respiration 1. Introduction Understanding the relationship between sediment community structure and seafloor ecological processes remains a central area of focus in current marine ecology and the growing concern of marine biodiversity loss has provided an extra impetus for experimental work aimed at ascertaining the ecological role of different benthic compartments in basic ecosystem functioning (e.g. Austen et al., 2002; Raffaelli et al., 2003; Solan et al., 2006). Seafloor ecosystem functioning can be defined as the ecological processes (such as nutrient or elemental cycling) or the flow of energy and material through the biotic and abiotic components of benthic ecosystems (Bremner et al., 2006). To this end, one approach has been to utilize 13 C-enriched algal material as a tracer (Blair et al., 1996; Levin et al., 1999) to follow organic carbon flow pathways and the fate (community respiration versus assimilation by microbes and fauna) of incoming organic carbon (e.g. Moodley et al., 2002; Witte et al., 2003; Buhring et al., 2006; Woulds et al., 2007). These studies have provided new insight into the relationship between seafloor community structure and functioning but have at the same time revealed gaps in our knowledge of some very fundamentals aspect of benthic ecology. For example, differential assimilation of tracer organic matter among the different benthic groups hints toward resource selectivity but knowledge on the trophodynamics of the different taxa is limited. Similarly, a fraction of organic carbon ingested by biota is respired but respiration rates are known only for a very limited number of taxa. Oxygen consumption is a measure of respiration rates and represents that part of food intake that provides energy to support life processes. Conse- quently, understanding organic carbon flow pathways or fluxes calls for documentation of both carbon assimilation and Journal of Experimental Marine Biology and Ecology 357 (2008) 41 – 47 www.elsevier.com/locate/jembe ⁎ Corresponding author. Korringaweg 7, 4401 NT Yerseke, The Netherlands. Tel.: +31 113 577418; fax: +31 113 573616. E-mail address: [email protected] (L. Moodley). 0022-0981/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.jembe.2007.12.025

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

y and Ecology 357 (2008) 41–47www.elsevier.com/locate/jembe
Journal of Experimental Marine Biolog
Biomass-specific respiration rates of benthic meiofauna:Demonstrating a novel oxygen micro-respiration system
Leon Moodley a,⁎, Maaike Steyaert b, Eric Epping c, Jack J. Middelburg a, Magda Vincx b,Pim van Avesaath a, Tom Moens b, Karline Soetaert a
a Netherlands Institute of Ecology, (NIOO-KNAW), Korringaweg, 4401 NT Yerseke, The Netherlandsb Marine Biology Section, Department of Biology, Ghent University, Krijgslaan 281/S8, 9000 Ghent, Belgium
c Royal Netherlands Institute for Sea Research, P.O. Box 59, 1790 AB Den Burg, Texel, The Netherlands
Received 11 October 2007; received in revised form 13 December 2007; accepted 17 December 2007
Abstract
Meiofauna (small-sized Metazoa and Foraminifera) may constitute a significant part of seafloor biomass and potentially play an important rolein benthic metabolism. However, respiration measurements are limited and the methods used are diverse together complicating comparison orupscaling. Here we describe a novel glass micro-respiration chamber used to perform non-invasive measurements (built-in oxygen-sensitivefluorogenic membrane and stirrer) and together with direct organic carbon measurements report initial biomass-specific respiration rates ofcommon intertidal meiofauna. Results indicate large differences between respiration rates of different taxa (biomass 0.7–5.2 µg C per individual)but very similar organic carbon biomass-specific respiration rates (1.6–2.5 µl O2 h
−1 mgC−1 or on average 2.0±0.3 µl O2 h−1 mgC−1). This new,
rapid and accurate method allows the study of metabolic allometry of the different small-sized taxa and determining their functional role in benthicmetabolism.© 2007 Elsevier B.V. All rights reserved.
Keywords: Benthic; Foraminifera; Meiofauna; Optodes; Respiration
1. Introduction
Understanding the relationship between sediment communitystructure and seafloor ecological processes remains a central areaof focus in current marine ecology and the growing concern ofmarine biodiversity loss has provided an extra impetus forexperimental work aimed at ascertaining the ecological role ofdifferent benthic compartments in basic ecosystem functioning(e.g. Austen et al., 2002; Raffaelli et al., 2003; Solan et al., 2006).Seafloor ecosystem functioning can be defined as the ecologicalprocesses (such as nutrient or elemental cycling) or the flow ofenergy and material through the biotic and abiotic components ofbenthic ecosystems (Bremner et al., 2006). To this end, oneapproach has been to utilize 13C-enriched algal material as a
⁎ Corresponding author. Korringaweg 7, 4401 NT Yerseke, The Netherlands.Tel.: +31 113 577418; fax: +31 113 573616.
E-mail address: [email protected] (L. Moodley).
0022-0981/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jembe.2007.12.025
tracer (Blair et al., 1996; Levin et al., 1999) to follow organiccarbon flow pathways and the fate (community respiration versusassimilation by microbes and fauna) of incoming organic carbon(e.g. Moodley et al., 2002; Witte et al., 2003; Buhring et al.,2006; Woulds et al., 2007). These studies have provided newinsight into the relationship between seafloor communitystructure and functioning but have at the same time revealedgaps in our knowledge of some very fundamentals aspect ofbenthic ecology. For example, differential assimilation of tracerorganic matter among the different benthic groups hints towardresource selectivity but knowledge on the trophodynamics of thedifferent taxa is limited. Similarly, a fraction of organic carboningested by biota is respired but respiration rates are known onlyfor a very limited number of taxa. Oxygen consumption is ameasure of respiration rates and represents that part of foodintake that provides energy to support life processes. Conse-quently, understanding organic carbon flow pathways or fluxescalls for documentation of both carbon assimilation and

42 L. Moodley et al. / Journal of Experimental Marine Biology and Ecology 357 (2008) 41–47
respiration. Meiofauna may constitute a significant part of sea-floor biomass and potentially play an important role in benthicmetabolism (e.g. Heip et al., 1985). However, respiration mea-surements are limited and the methods used are diverse togethercomplicating comparison or upscaling. In contrast to macro-fauna, the small size of meiofauna is troublesome for rapid androutine measurements of oxygen uptake. However, this is needednot only to determine basic allometric relationships, but also tostudy impact of different environmental factors such astemperature and salinity (e.g. Moens et al., 1996).
Meiofauna, the operationally defined group of fauna passingthrough a 1 mm mesh openings but retained on sieves with a32–63 µmmesh opening encompasses juvenile macrofauna and“permanent” taxa (taxa that remain of small size) includingsingle-celled foraminifera of meiofauna size. Of the metazoans,nematodes are by far the most dominant taxon in terms of bothdensities and biomass but are often equalled by foraminifera(e.g. Widbom, 1988; Moodley et al., 2000 and referencestherein). Inherent to the division of scientific disciplines andassociated target organisms; different meiofauna taxa are oftenstudied in isolation and with different tools and methods.Cartesian divers and related techniques are highly sensitive butare extremely time- and labour-consuming (e.g. Bradshaw,1961; Price and Warwick, 1980; Herman and Heip, 1982;Hannah et al., 1994). Polarographic oxygen electrodes (e.g.Atkinson, 1973; Atkinson and Smith, 1973; Moens et al., 1996;Moens and Vincx, 2000) require more specimens per measure-ment, as well as adequate stirring which is sometimes difficultto accommodate in small volume vessels; small incubationvessels are preferred in order to limit incubation time whileobtaining sufficient decrease in oxygen content. Alternatively,oxygen optodes (measurements based on dynamic fluorescencequenching) have similar accuracy as electrodes and are lesssensitive to stirring (Klimant et al., 1995). Optodes have beensuccessfully applied for measurements on small animals (e.g.Gatti et al., 2002; Irwin et al., 2007) but have yet to be utilizedfor meiofauna taxa in small volume chambers.
From the limited data available, there is a wide range inmeiofauna respiration rates measured at 20 °C; e.g. for marinenematodes respiration ~1.7–33 nl O2 h
−1 individual−1 (Moenset al., 1999; Moens and Vincx, 2000) and for foraminifera ~2–58 nl O2 h
−1 individual−1 (Hannah et al., 1994 and referencestherein). Although it may seem that foraminifera may havehigher respiration rates, cross-taxon comparison is not onlyhindered due to methodological differences but also compli-cated as a result of different measures of weight or biomass. Dryor wet weight is often used for metazoan meiofauna and volumeestimates for foraminifera with inherent complication due todifficulty in measuring test (shell) thickness; further extrapola-tion/conversion to organic carbon biomass can be an extrasource of error (Hannah et al., 1994). Given differences inweight/volume and weight/organic carbon ratios between thetaxa, determining or expressing respiration rates directly as afunction of organic carbon allows more accurate comparisons(e.g. Schneider, 1992).
Here we report on a micro-respiration chamber with anoxygen sensor spot immobilized on the inside wall of a glass
chamber with measurements conducted from the outsidethrough the glass wall (so called “non-invasive” measurements,see www.presens.de). At the end of incubations, the organiccarbon content of the organisms was measured directly on aCHN analyser to accurately determine organic carbon biomass-specific respiration rates.
2. Materials and methods
The main aim was to have a respiration chamber as small aspossible with the least amount of potential sources of oxygen.Additionally, given that foraminifera and nematodes are moreactive in the absence of sediment and tend to rapidly settle downonce surrounded by particles (personal observations), weincluded a substrate. However, inclusion of a substratenecessitates non-destructive stirring of the water to avoiddevelopment of oxygen gradients in the case of less mobiletaxon such as foraminifera. The smallest size of the micro-respiration chamber was in part determined by the size of thesmallest magnet-bar that was available (teflon coated 2×5 mm)but also by the physical limitations of working with glass.Perspex or Teflon both potentially hold air that may slowly bereleased into the surrounding water upon fauna consumption ofdissolved oxygen. Consequently all components were made ofglass. Similarly, a teflon-coated magnet may be a source ofoxygen, hence we replaced the teflon coating by a glass coating(Glassinstruments B.V., Arnemuiden, The Netherlands) with afinal dimension of 3×7 mm.
Decrease in oxygen concentration in seawater was measuredusing an oxygen sensor spot (Presens, Germany) where oxygenacts as a dynamic fluorescence quencher of a luminophore in apolymer matrix. In contrast to (micro) optodes, this polymermatrix (available as spots immobilized on a thin foil) was gluedonto the inner wall of the micro-respiration chamber (Presens,Germany) and this allowed non-invasive measurements ofoxygen content through the glass wall from the outside. Anoptic fibre connected to the OXY 4 oxygen meter (Presens,Germany, source and receiver of optical signal connected to aPC for continuous monitoring) was guided and positioned fromthe outside of the chamber on the spot. Theoretical details onluminophore quenching and on the use of phase modulation forthe calculation of oxygen saturation can be found in Klimant etal. (1995) and on line (www.presens.de). Furthermore, a salinitycorrection (Garcia and Gordon, 1992) was embedded by themanufacturer in the original software in order to convert %oxygen saturation to µmol O2/l. The validity and accuracy of thebasic principle of the optodes have been amply demonstrated(Holst et al., 1997; Gatti et al., 2002; Irwin et al., 2007). Here wedescribe and demonstrate a specific design that allows accurate,rapid measurements of oxygen consumption by meiofauna.
This micro-respiration chamber is illustrated in Fig. 1. Aglass vessel (1 cm high and an inner diameter of 0.9 cm with awall thickness of 1.5 mm) was the basic chamber into which isfitted a glass table upon which a glass-coated magnet resides.First the chamber is filled with a layer (ca. 1 mm thick) of glassbeads (100–110 µm diameter, Sartorius, Germany), whichconstitutes the substrate. Consistent thickness was obtained by

Fig. 1. Sketch of the glass micro-respiration chamber with an oxygen-sensitivespot immobilized on the inside glass (Presens, Germany). The different parts are:(1) assembled glass chamber with glass beads (diameter 100–110 µm, Sartorius,Germany) as substrate at the bottom of the chamber. (2) Glass chamber withoxygen sensor spot (red) on the inside right hand wall. (3) Part 1 of glass tablewith detachable table top, legs of table with opening to allow insertion oforganisms. (4) Part 2 of glass table: the top with a tiny hole to promote goodmixing fits into part 1 to support magnet. This allows stirring without destructionof the organisms. (5) Glass-coated magnet stirrer. (6) Butyl rubber septum thatcan be pierced to release pressure built up when bolted down with metal plate.(7) Metal plate with hole to reach the septum with a sterile needle. (8) Bolts toscrew down the retaining plate to produce an assembled chamber (1) where apolymer optical fibre is positioned directly against the outer glass wall of thechamber at the point of the sensor spot. This fibre delivers the optic signal fromthe OXY 4 meter and receives the signal from the sensor spot back to thephotodetector in the oxygen meter. (For interpretation of the references to colorin this figure legend, the reader is referred to the web version of this article.)
43L. Moodley et al. / Journal of Experimental Marine Biology and Ecology 357 (2008) 41–47
using a fixed weight of glass beads per chamber. Exact portionswere heat sterilized (4 h at 400 °C) in a glass bottle (2 mlvolume) and were transferred into the respiration chamber usingdouble 0.2 µm filtered seawater with a glass pipette. In thechambers then partially filled with incubation water, the 4-legged table without top is inserted with a sterile metal pincetand pushed down evenly. Then the fauna (following cleaningsteps, see below) is pipetted into the chamber through the tableopening after which the table top is inserted into place to holdthe glass-coated magnet in position. A tiny hole in the tabletopensures rapid thorough mixing of the water (tests with inkverified that water was pulled up the hole and mixing was rapid;total mixing of a drop of ink placed at the bottom was withinminutes). Finally, the chamber is filled to the rim with seawaterand an acid cleaned butyl rubber septum placed above withouttrapping air. This septum was held down on the chamber with aheavy metal plate guided along pins on each side (see Fig. 1)and secured by screw bolts. The pressure in the chamber isreleased by piercing the septum with a sterile needle through an
opening in the metal cover (Fig. 1). This assembled chamberwas then positioned with spot directly in front of the polymeroptical fibre (to transmit and receive optical signals, positionfixed and ensured through a plastic pipe as guide) in a waterbath maintained at 20 °C in a temperature-controlled laboratorymaintained at the same temperature. The temperature bath wasplaced on a magnetic stirring table and in total, 4 chambers weresimultaneously incubated with the polymer optical fibresconnected to a 4-channel oxygen meter (OXY 4). Details andworking of the different components of the planar oxygensensor spot and the oxygen meter are available online (www.presens.de).
Two-point calibration of the sensor spots was done atexperimental temperature as specified by the manufacturers. Allcomponents were cleaned by first thoroughly washing with hotwater followed by a rinse with MilliQ water, a soak in a weakacid (10 min in 3% HCl), rinsed again with MilliQ water andthen sterilized by soaking in ethanol for a few minutes afterwhich components were dried through evaporation acceleratedwith a flow of nitrogen. All components were stored dry anddark in the temperature-controlled laboratory until initiation ofexperiments (within a couple of days) and rinsed once with0.2 µm filtered MilliQ and 0.2 µm filtered incubation seawaterjust prior to usage.
Test organisms were handpicked from the intertidal sedimentfree of mud (fine particles removed through sieving) collectedduring low tide at different locations in the Westerscheldeestuary (southwest Netherlands). Organisms were carefullycleaned of adhering detritus (thoroughly with a brush in case ofshelled organisms) in ambient seawater before being transferredto incubation seawater (double 0.2 µm filtered Atlantic openocean seawater, chosen because of low nutrient and dissolvedorganic carbon content). Bacteria contamination was reducedthrough several changes of this sterile water. Finally, organismswere transferred to the micro-respiration chamber by pipetting(Foraminifera and Ostracoda) or by needle transfer (Nematodes)and chambers only containing similar traces of the final washwere used as a control against bacterial contamination. Thus,per run, two controls and two fauna incubations wereconducted. It took on average approximately 30 min to fill all4 chambers that were then transferred (after verifying theabsence of air bubbles) into the water bath that was maintainedat 20 °C by a continuous flow of 20 °C water from aThermoMetric thermostat. The incubations were initiated withonline measurements preset every 5 min for 3–6 h. Temperaturein the temperature bath was also continuously monitored andremained fairly constant (20±0.6 °C) throughout the entiremeasurement period. Upon significant decrease in oxygenconcentration, measurements were stopped and chambers werechecked for air bubbles and the position of animals was noted.The metal plate firmly holding down the septum was thenunscrewed to remove the septum. The septum was blotted withpre-weighed blotting paper under the microscope (for laterdetermination of incubation water volume through weight)simultaneously checking for fauna. The fauna activity andposition were noted and in all cases most of the animals werepositioned in the substrate. Then each part was carefully
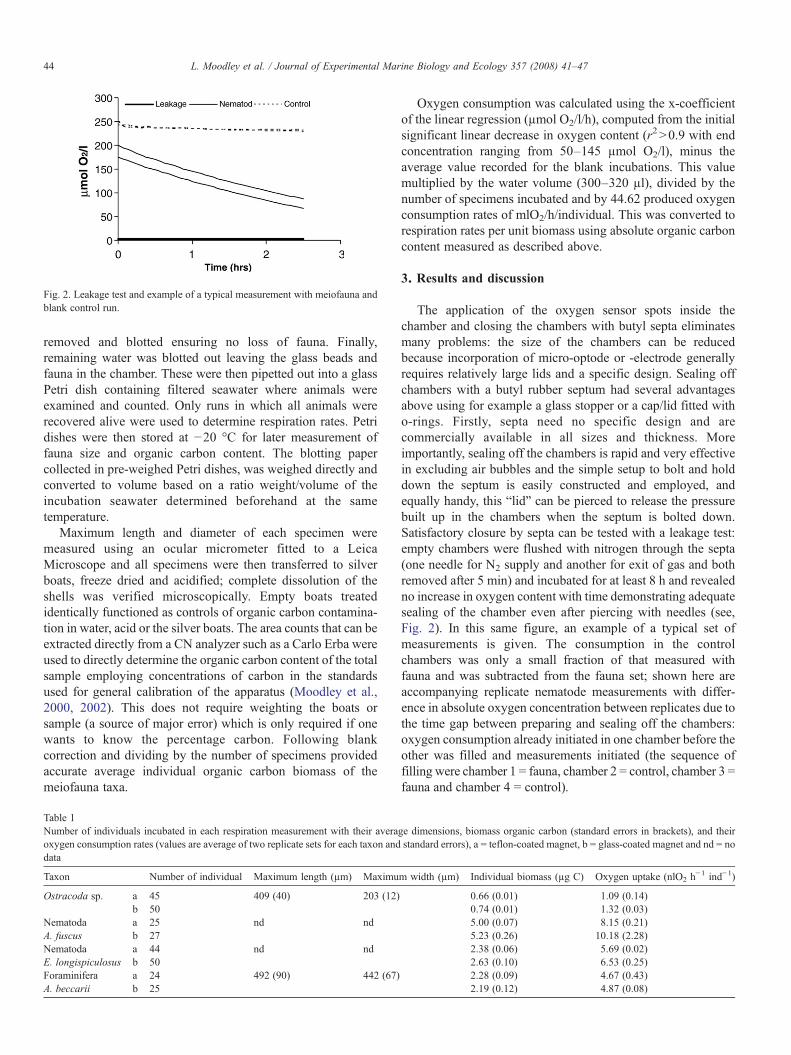
Fig. 2. Leakage test and example of a typical measurement with meiofauna andblank control run.
44 L. Moodley et al. / Journal of Experimental Marine Biology and Ecology 357 (2008) 41–47
removed and blotted ensuring no loss of fauna. Finally,remaining water was blotted out leaving the glass beads andfauna in the chamber. These were then pipetted out into a glassPetri dish containing filtered seawater where animals wereexamined and counted. Only runs in which all animals wererecovered alive were used to determine respiration rates. Petridishes were then stored at −20 °C for later measurement offauna size and organic carbon content. The blotting papercollected in pre-weighed Petri dishes, was weighed directly andconverted to volume based on a ratio weight/volume of theincubation seawater determined beforehand at the sametemperature.
Maximum length and diameter of each specimen weremeasured using an ocular micrometer fitted to a LeicaMicroscope and all specimens were then transferred to silverboats, freeze dried and acidified; complete dissolution of theshells was verified microscopically. Empty boats treatedidentically functioned as controls of organic carbon contamina-tion in water, acid or the silver boats. The area counts that can beextracted directly from a CN analyzer such as a Carlo Erba wereused to directly determine the organic carbon content of the totalsample employing concentrations of carbon in the standardsused for general calibration of the apparatus (Moodley et al.,2000, 2002). This does not require weighting the boats orsample (a source of major error) which is only required if onewants to know the percentage carbon. Following blankcorrection and dividing by the number of specimens providedaccurate average individual organic carbon biomass of themeiofauna taxa.
Table 1Number of individuals incubated in each respiration measurement with their averaoxygen consumption rates (values are average of two replicate sets for each taxon anddata
Taxon Number of individual Maximum length (µm) Maximu
Ostracoda sp. a 45 409 (40) 203 (12)b 50
Nematoda a 25 nd ndA. fuscus b 27Nematoda a 44 nd ndE. longispiculosus b 50Foraminifera a 24 492 (90) 442 (67)A. beccarii b 25
Oxygen consumption was calculated using the x-coefficientof the linear regression (µmol O2/l/h), computed from the initialsignificant linear decrease in oxygen content (r2N0.9 with endconcentration ranging from 50–145 µmol O2/l), minus theaverage value recorded for the blank incubations. This valuemultiplied by the water volume (300–320 µl), divided by thenumber of specimens incubated and by 44.62 produced oxygenconsumption rates of mlO2/h/individual. This was converted torespiration rates per unit biomass using absolute organic carboncontent measured as described above.
3. Results and discussion
The application of the oxygen sensor spots inside thechamber and closing the chambers with butyl septa eliminatesmany problems: the size of the chambers can be reducedbecause incorporation of micro-optode or -electrode generallyrequires relatively large lids and a specific design. Sealing offchambers with a butyl rubber septum had several advantagesabove using for example a glass stopper or a cap/lid fitted witho-rings. Firstly, septa need no specific design and arecommercially available in all sizes and thickness. Moreimportantly, sealing off the chambers is rapid and very effectivein excluding air bubbles and the simple setup to bolt and holddown the septum is easily constructed and employed, andequally handy, this “lid” can be pierced to release the pressurebuilt up in the chambers when the septum is bolted down.Satisfactory closure by septa can be tested with a leakage test:empty chambers were flushed with nitrogen through the septa(one needle for N2 supply and another for exit of gas and bothremoved after 5 min) and incubated for at least 8 h and revealedno increase in oxygen content with time demonstrating adequatesealing of the chamber even after piercing with needles (see,Fig. 2). In this same figure, an example of a typical set ofmeasurements is given. The consumption in the controlchambers was only a small fraction of that measured withfauna and was subtracted from the fauna set; shown here areaccompanying replicate nematode measurements with differ-ence in absolute oxygen concentration between replicates due tothe time gap between preparing and sealing off the chambers:oxygen consumption already initiated in one chamber before theother was filled and measurements initiated (the sequence offilling were chamber 1 = fauna, chamber 2 = control, chamber 3 =fauna and chamber 4 = control).
ge dimensions, biomass organic carbon (standard errors in brackets), and theirstandard errors), a = teflon-coated magnet, b = glass-coated magnet and nd = no
m width (µm) Individual biomass (µg C) Oxygen uptake (nlO2 h−1 ind−1)
0.66 (0.01) 1.09 (0.14)0.74 (0.01) 1.32 (0.03)5.00 (0.07) 8.15 (0.21)5.23 (0.26) 10.18 (2.28)2.38 (0.06) 5.69 (0.02)2.63 (0.10) 6.53 (0.25)2.28 (0.09) 4.67 (0.43)2.19 (0.12) 4.87 (0.08)
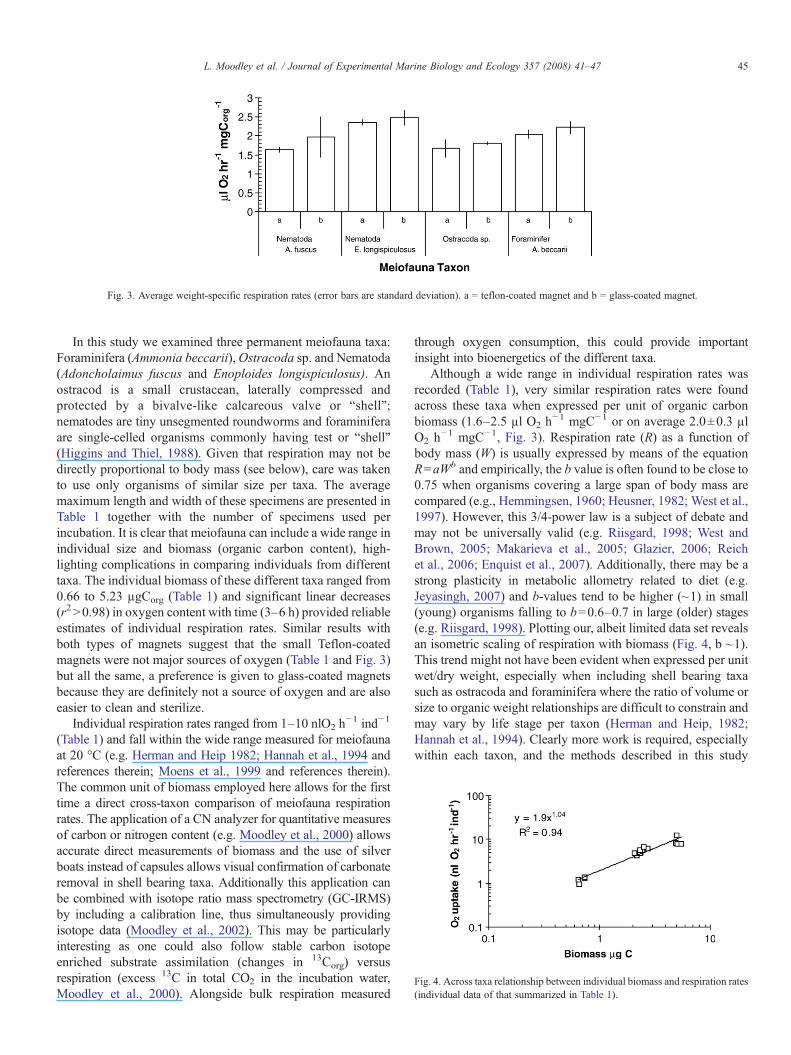
Fig. 3. Average weight-specific respiration rates (error bars are standard deviation). a = teflon-coated magnet and b = glass-coated magnet.
Fig. 4. Across taxa relationship between individual biomass and respiration rates(individual data of that summarized in Table 1).
45L. Moodley et al. / Journal of Experimental Marine Biology and Ecology 357 (2008) 41–47
In this study we examined three permanent meiofauna taxa:Foraminifera (Ammonia beccarii),Ostracoda sp. and Nematoda(Adoncholaimus fuscus and Enoploides longispiculosus). Anostracod is a small crustacean, laterally compressed andprotected by a bivalve-like calcareous valve or “shell”;nematodes are tiny unsegmented roundworms and foraminiferaare single-celled organisms commonly having test or “shell”(Higgins and Thiel, 1988). Given that respiration may not bedirectly proportional to body mass (see below), care was takento use only organisms of similar size per taxa. The averagemaximum length and width of these specimens are presented inTable 1 together with the number of specimens used perincubation. It is clear that meiofauna can include a wide range inindividual size and biomass (organic carbon content), high-lighting complications in comparing individuals from differenttaxa. The individual biomass of these different taxa ranged from0.66 to 5.23 µgCorg (Table 1) and significant linear decreases(r2N0.98) in oxygen content with time (3–6 h) provided reliableestimates of individual respiration rates. Similar results withboth types of magnets suggest that the small Teflon-coatedmagnets were not major sources of oxygen (Table 1 and Fig. 3)but all the same, a preference is given to glass-coated magnetsbecause they are definitely not a source of oxygen and are alsoeasier to clean and sterilize.
Individual respiration rates ranged from 1–10 nlO2 h−1 ind−1
(Table 1) and fall within the wide range measured for meiofaunaat 20 °C (e.g. Herman and Heip 1982; Hannah et al., 1994 andreferences therein; Moens et al., 1999 and references therein).The common unit of biomass employed here allows for the firsttime a direct cross-taxon comparison of meiofauna respirationrates. The application of a CN analyzer for quantitative measuresof carbon or nitrogen content (e.g. Moodley et al., 2000) allowsaccurate direct measurements of biomass and the use of silverboats instead of capsules allows visual confirmation of carbonateremoval in shell bearing taxa. Additionally this application canbe combined with isotope ratio mass spectrometry (GC-IRMS)by including a calibration line, thus simultaneously providingisotope data (Moodley et al., 2002). This may be particularlyinteresting as one could also follow stable carbon isotopeenriched substrate assimilation (changes in 13Corg) versusrespiration (excess 13C in total CO2 in the incubation water,Moodley et al., 2000). Alongside bulk respiration measured
through oxygen consumption, this could provide importantinsight into bioenergetics of the different taxa.
Although a wide range in individual respiration rates wasrecorded (Table 1), very similar respiration rates were foundacross these taxa when expressed per unit of organic carbonbiomass (1.6–2.5 µl O2 h
−1 mgC−1 or on average 2.0±0.3 µlO2 h−1 mgC−1, Fig. 3). Respiration rate (R) as a function ofbody mass (W) is usually expressed by means of the equationR=aWb and empirically, the b value is often found to be close to0.75 when organisms covering a large span of body mass arecompared (e.g., Hemmingsen, 1960; Heusner, 1982; West et al.,1997). However, this 3/4-power law is a subject of debate andmay not be universally valid (e.g. Riisgard, 1998; West andBrown, 2005; Makarieva et al., 2005; Glazier, 2006; Reichet al., 2006; Enquist et al., 2007). Additionally, there may be astrong plasticity in metabolic allometry related to diet (e.g.Jeyasingh, 2007) and b-values tend to be higher (~1) in small(young) organisms falling to b=0.6–0.7 in large (older) stages(e.g. Riisgard, 1998). Plotting our, albeit limited data set revealsan isometric scaling of respiration with biomass (Fig. 4, b ~1).This trend might not have been evident when expressed per unitwet/dry weight, especially when including shell bearing taxasuch as ostracoda and foraminifera where the ratio of volume orsize to organic weight relationships are difficult to constrain andmay vary by life stage per taxon (Herman and Heip, 1982;Hannah et al., 1994). Clearly more work is required, especiallywithin each taxon, and the methods described in this study

46 L. Moodley et al. / Journal of Experimental Marine Biology and Ecology 357 (2008) 41–47
allow accurate and rapid quantification of organic carbon-specific respiration rates of fauna including the tiny meiofauna.This novel method was demonstrated using relatively largemeiofauna specimens and a larger number of specimens will berequired to study the smaller species or juveniles. However, theisolation and cleaning of meiofauna is very time consuming andthe number of specimens required can be reduced drastically byusing smaller chambers. The size of the chambers can besignificantly reduced by excluding the glass substrate andtherefore also the need to stir the sample so that the table andmagnet can be omitted. A thinner layer of substrate may also beused without a stirrer, especially in a smaller volume of waterthat does not promote the development of oxygen gradients thatmay bias the measurements. In upscaling respiration rates, it isimportant to compare the allometry of the different taxa thatwould provide a better estimate of the functional role of thedifferent taxa especially the protozoan versus metazoanmeiofauna in benthic metabolism.
Acknowledgements
Special thanks to Ronald Rutgers and Lennart van Ijzerloofor excellent and accurate technical assistance during differentstages of this study. The brothers Peene, Bert Sinke and LouisHaazen are acknowledged for their patience passing throughseveral designs. We are grateful to Harm Simons (Glassinstru-ments B.V.) for intricate creations based on vague descriptions.T.M. is a postdoctoral fellow with the Flemish Fund forScientific Research FWO. This work was financially supportedby the Dutch and Flemish Funds for Scientific Research (NWOand FWO) in the framework of the joint ‘Flemish-Dutchprogramme on coastal marine research’. This is publication4229 of The Netherlands Institute of Ecology (NIOO-KNAW),Yerseke. [SS]
References
Atkinson, H.J., 1973. The respiratory physiology of the marine nematodesEnoplus brevis (Bastian) and E. communis (Bastian). I. The influence ofoxygen tension and body size. J. Exp. Biol. 59, 255–266.
Atkinson, H.J., Smith, L., 1973. An oxygen electrode microrespirometer. J. Exp.Biol. 59, 247–253.
Austen, M.C., Lambshead, P.J.D., Hutchings, P.A., Boucher, G., Snelgrove, P.V.R.,Heip, C., King, G., Koike, I., Smith, C., 2002. Biodiversity links above andbelow the marine sediment-water interface that may influence communitystability. Biodivers. Conserv. 11, 113–136.
Blair, N.E., Levin, L.A., DeMaster, D.J., Plaia, G., 1996. The short-term fate of freshalgal carbon in continental slope sediments. Limnol. Oceanogr. 41, 1208–1219.
Bradshaw, J.S., 1961. Laboratory experiments on the ecology of foraminifera.Contrib. Cushman Found. Foraminifer. Res. 12, 87–106.
Bremner, J., Rogers, S.I., Frid, C.L.J., 2006. Matching biological traits toenvironmental conditions in marine benthic ecosystems. J. Mar. Syst. 60,302–316.
Buhring, S.I., Lampadariou, N., Moodley, L., Tselepides, A., Witte, U., 2006.Benthic microbial and whole-community responses to different amounts ofC13-enriched algae: in situ experiments in the deep Cretan Sea (EasternMediterranean). Limnol. Oceanogr. 51, 157–165.
Enquist, B.J., Allen, A.P., Brown, J.H., Gilloolyl, J.F., Kerkhoff, A.J., Niklas,K.J., Price, C.A., West, G.B., 2007. Does the exception prove the rule?Nature 445, E9–E10.
Garcia, H.E., Gordon, L.I., 1992. Oxygen solubility in seawater: better fittingequations. Limnol. Oceanogr. 37, 1307–1312.
Gatti, S., Brey, T., Muller, W.E.G., Heilmayer, O., Holst, G., 2002. Oxygenmicrooptodes: a new tool for oxygen measurements in aquatic animalecology. Mar. Biol. 140, 1075–1085.
Glazier, D.S., 2006. The 3/4-power law is not universal: evolution of isometric,ontogenetic metabolic scaling in pelagic animals. Bioscience 56, 325–332.
Hannah, F., Rogerson, A., Laybourn-Parry, J., 1994. Respiration rates andbiovolumes of common benthic foraminifera (Protozoa). J. Mar. Biol.Assoc. U.K. 74, 301–312.
Heip, C., Vincx, M., Vranken, G., 1985. The ecology of marine nematodes.Oceanogr. Mar. Biol. Annu. Rev. 23, 399–489.
Hemmingsen, A.M., 1960. Energy metabolism as related to body size andrespiratory surfaces and its evolution. Rep. Steno Mem. Hosp. Nord.Insulinlab. 9 II (Copenhagen 1960).
Herman, P.M.J., Heip, C., 1982. Growth and respiration of Cyprides torosaJones 1850 (Crustacea Ostracoda). Oecologia 54, 300–303.
Heusner, A.A., 1982. Energy metabolism and body size. I. Is the 0.75 massexponent of Kleiber's equation a statistical artifact? Respir. Physiol. 48,1–12.
Higgins, R.P., Thiel, H., 1988. Introduction to the Study of Meiofauna.Smithsonian Institution Press, Washington D.C.
Holst, G., Glud, R.N., Kuhl, M., Klimant, I., 1997. A microoptode array for fine-scale measurement of oxygen distribution. Sensore and Actuators B 38–39,122–129.
Irwin, S., Wall, V., Davenport, J., 2007. Measurement of temperature andsalinity effects on oxygen consumption of Artemia franiscana K., measuredusing fibre-optic oxygen microsensors. Hydrobiologia 575, 109–115.
Jeyasingh, P.D., 2007. Plasticity in metabolic allometry: the role of dietarystoichiometry. Ecol. Lett. 10, 282–289.
Klimant, I., Meyer, V., Kuhl, M., 1995. Fibre-optic oxygen microsensors, a newtool in aquatic biology. Limnol. Oceanogr. 40, 1159–1165.
Levin, L.A., Blair, N.E., Martin, C.M., DeMaster, D.J., Plaia, G., Thomas, C.J.,1999. Macrofaunal processing of phytodetritus at two contrasting sites onthe Carolina margin: in situ experiments using 13C-labeled diatoms. Mar.Ecol. Prog. Ser. 182, 37–54.
Makarieva, A.M., Gorshkov, V.G., Li, B.-L., 2005. Energetics of the smallest:do bacteria breathe at the same rate as whales? Proc. R. Soc. B. 272,2219–2224.
Moens, T., Vincx, M., 2000. Temperature, salinity and food thresholds in twobrackish-water bacterivorous nematode species: assessing niches from foodabsorption and respiration experiments. J. Exp.Mar. Biol. Ecol. 243, 137–154.
Moens, T., Vierstraete, A., Vanhove, S., Verbeke, M., Vincx, M., 1996. A handymethod for measuring meiobenthic respiration. J. Exp. Mar. Biol. Ecol. 197,177–190.
Moens, T., Verbeeck, L., Vincx, M., 1999. Feeding biology of a predatory and afacultatively predatory nematode (Enoploides longispiculosus andAdoncholaimus fuscus). Mar. Biol. 134, 585–593.
Moodley, L., Boschker, H.T.S., Middelburg, J.J., Pel, R., Herman, P.M.J., deDeckere, E.M.G.T., Heip, C.H.R., 2000. Ecological significance of benthicforaminifera: C-13 labelling experiments. Mar. Ecol. Prog. Ser. 202, 289–295.
Moodley, L., Middelburg, J.J., Boschker, H.T.S., Duineveld, G.C.A., Pel, R.,Herman, P.M.J., Heip, C.H.R., 2002. Bacteria and Foraminifera: key playersin a short-term deep-sea benthic response to phytodetritus. Mar. Ecol. Prog.Ser. 236, 23–29.
Price, R., Warwick, R.M., 1980. The effect of temperature on the respiration ofmeiofauna. Oecologia 44, 145–148.
Raffaelli, D., Emmerson, M., Solan, M., Biles, C., Paterson, D., 2003.Biodiversity and ecosystem processes in shallow coastal waters: anexperimental approach. J. Sea Res. 49, 133–141.
Reich, P.B., Tjoelker, M.G., Machado, J.-L., Oleksyn, J., 2006. Universalscaling of respiratory metabolism, size and nitrogen in plants. Nature 439,457–461.
Riisgard, H.U., 1998. No foundation of a “3/4 power scaling law” for respirationin biology. Ecol. Lett. 1, 71–73.
Schneider, G., 1992. A comparison of carbon-specific respiration rates ingelatinous and non-gelatinous zooplankton: a search for general rules inzooplankton metabolism. Helgo. Meres. 46, 377–388.

47L. Moodley et al. / Journal of Experimental Marine Biology and Ecology 357 (2008) 41–47
Solan, M., Raffaelli, D.G., Paterson, D.M., White, P.C.L., Pierce, G.J., 2006.Marine biodiversity and ecosystem function: empirical approaches andfuture research needs. Mar. Ecol. Prog. Ser. 311, 175–178.
West, G.W., Brown, J.H., 2005. The origin of allometric scaling laws in biologyfrom genomes to ecosystems: towards a quantitative unifying theory ofbiological structure and organization. J. Exp. Biol. 208, 1575–1592.
West, G.W., Brown, J.H., Enquist, B.J., 1997. A general model for the origin ofthe allometric scaling laws in biology. Science. 276, 122–126.
Widbom, B., 1988. The benthic meiofauna of three coastal areas: structure,seasonal dynamics and response to environmental perturbations. DoctoralThesis. University of Stockholm.
Witte, U., Aberle, N., Sand, M., Wenzhofer, F., 2003. Rapid response of a deep-sea benthic community to POM enrichment: an in situ experimental study.Mar. Ecol. Prog. Ser. 251, 27–36.
Woulds, C., Cowie, G.L., Levin, L.A., Andersson, J.H., Middelburg, J.J.,Vandewiele, S., Lamont, P.A., Larkein, K.E., Gooday, A.J., Schumacher, S.,Whitcraft, C., Jeffreys, C., Schwartz, M., 2007. Oxygen as control onseafloor biological communities and their role in sedimentary carboncycling. Limnol. Oceanogr. 52, 1698–1709.
Related Documents











