This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Biological treatment of the effluent from a bleached kraft pulp mill usingbasidiomycete and zygomycete fungi
A.C. Freitas a,⁎, F. Ferreira a, A.M. Costa a, R. Pereira b, S.C. Antunes b, F. Gonçalves b, T.A.P. Rocha-Santos a,M.S. Diniz d, L. Castro d, I. Peres d, A.C. Duarte c
a ISEIT/Viseu, Instituto Piaget, Estrada do Alto do Gaio, Galifonge, 3515-776 Lordosa, Viseu, Portugalb CESAM & Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugalc CESAM & Department of Chemistry, University of Aveiro, 3810-193 Aveiro, Portugald IMAR-Instituto do Mar, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa - Departamento de Ciências e Engenharia do Ambiente,Quinta da Torre, Eliminate-2829-516 Caparica, Portugal
a b s t r a c ta r t i c l e i n f o
Article history:Received 3 November 2008Received in revised form 20 January 2009Accepted 26 January 2009Available online 6 March 2009
Keywords:FungiKraft effluentBiological treatment
Threewhite-rot fungi (Pleurotus sajor caju, Trametes versicolor and Phanerochaete chrysosporium) and one soft-rot fungi (Rhizopus oryzae) species confirmed their potential for future applications in the biological treatmentof effluents derived from the secondary treatment of a bleached kraft pulp mill processing Eucalyptus globulus.Among the four species P. sajor caju and R. oryzae were the most effective in the biodegradation of organiccompounds present in the effluent, being responsible for the reduction of relative absorbance (25–46% at250 nm and 72–74% at 465 nm) and of chemical oxygen demand levels (74 to 81%) after 10 days of incubation.Laccase (Lac), lignin (Lip) and manganese peroxidases (MnP) expression varied among fungal species, whereLac and LiP activities were correlated with the degradation of organic compounds in the effluent treated withP. sajor caju. The first two axes of a principal component analysis explained 88.9% of the total variation amongsub-samples treatedwith the four fungus species, after different incubationperiods. All the variablesmeasuredcontributed positively to the first component except for the MnP enzyme activity which was the only variablecontributing negatively to the first component. Absorbances at 465 nm, LiP and Lac enzyme activities were thevariables with more weight on the second component. P. sajor caju revealed to be the only species able toperform the biological treatment without promoting an increment in the toxicity of the effluent to the Vibriofischeri, as it was assessed by theMicrotox® assay. The oppositewas recorded for the treatments with the otherthree species of fungus. EC50–5 min values ranging between 28 and 57% (effluent concentrations)were recordedeven after 10 to 13 days of treatment with P. chrysosporium, R. oryzae or with T. versicolor.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Among other impacts, industrial effluents are responsible for slimegrowth, thermal changes, scum formation, colour problems and loss ofenvironmental aesthetic beauty on receiving freshwater systems(Pokhrel and Viraraghavan, 2004). The pulp and paper industry, inparticular, produces a strongly coloured effluent with high levels ofbiochemical oxygen demand (BOD) and chemical oxygen demand(COD). These effluents contain many organic compounds, derivedfrom lignin, which are responsible for their brown colour and also forincreasing water temperature and decreasing photosynthesis rate ofthe phytoplanktonic community. The content in lowmolecular weightchloro-organics of residues, generated by pulp and paper industry, isthe major contributors to mutagenicity and bioaccumulation, due to
their hydrophobicity and ability to penetrate cell membranes(Pedroza et al., 2007). All these organic compounds are toxic toaquatic organisms and resistant to microbial degradation, resulting ina decrease of the ecological value of natural systems surrounding thepulp mill (Esposito et al., 1991).
Biological treatments are particularly attractive for wastewatersrecovery since, in addition to colour removal, they also reduce the BODand COD of the effluent (Bajpai and Bajpai, 1994). Several authorsreported that different fungal species are able to remove colour fromkraft mill effluent (Durant et al., 1994; Sakurai et al., 2001). White-rotfungi have proved their potential for lignin/phenolic wastewatertreatment since they produce enzymes including lignin peroxidases,MnP-dependent peroxidases and laccases (Conesa et al., 2002) whichare able to degrade lignin as well as chlorinated lignins found in pulpbleaching effluents (Lankinen et al., 1990). Lignin-degrading perox-idases have been found in a wide number of basidiomycetes funginamely in species such as Phanerochaete chrysosporium, Trametes
Science of the Total Environment 407 (2009) 3282–3289
⁎ Corresponding author. Tel.: +351 232910017; fax: +351 232910123.E-mail address: [email protected] (A.C. Freitas).
0048-9697/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.scitotenv.2009.01.054
Contents lists available at ScienceDirect
Science of the Total Environment
j ourna l homepage: www.e lsev ie r.com/ locate /sc i totenv

Author's personal copy
versicolor and Pleurotus spp. (Kimura et al., 1990; Pelaez et al., 1995;Varela et al., 2000). In addition, the enzymatic systemof fungi includesseveral other non-specific extracellular enzymeswhich catalyze ligninand chlorolignins degradation and also oxidize aromatic and haloge-nated compounds (Bergbauer et al., 1991). The enzymes from white-rot fungi that catalyze the initial depolymerization are thought to playan important role in the initial depolymerization of lignin (Cullen andKersten, 2004). The resulting fragmentswith lowmolecularweight areultimately metabolized in intracellular environment to water andcarbon dioxide. These enzymes are responsible for generating highlyreactive free radicals that undergo a complex series of spontaneouscleavage reactions which have attracted considerable interest forseveral industrial applications such as biological pulping of paper andalso for effluent treatment (Valls and Roncero, 2009; Witayakran andRagauskas, 2009; Camarero et al., 2007;Wingate et al., 2005; Ünal andKolankaya, 2001).
Although species differ in their gross morphological patterns ofdecay, up to now, only white-rot basidiomycetes have convincinglyshown to efficientlymineralize lignin (Eriksson et al.,1990; Blanchette,1991; Daniel, 1994). However, other species belonging to zygomyceteare also of interest (Nagarathnamma and Bajpai, 1999) on thedecolourization and detoxification of kraft pulp effluents. Hence themain aim of this study was to compare the efficiency of the biologicaltreatment of an effluent, derived from the secondary treatment of ableached kraft pulp mill processing Eucalyptus globulus, performedwith basidiomycete and zygomycete fungi. To meet this purpose theeffluent was treated with four species of fungi (P. sajor caju, R. oryzae,T. versicolor and P. chrysosporium) through different periods of time.
2. Materials and methods
2.1. Inoculum: source and culture conditions
Three species of basidiomycete fungi and one species ofzygomycete fungi were obtained from UNESP (São Paulo) and fromBCCMTM/MUCL Culture Collection (Belgium): Pleurotus sajor caju(UNESP), P. chrysosporium (Burdsall, 38388), T. versicolor [(Linnaeus:Fries) Pilát, 38412)] and Rhizopus oryzae (Went & Prinsen Geerlings,31002). All species were cultured at 30 °C, except P. sajor caju, whichwas cultured at 25 °C, in media containing 20 g/L of malt extract, 1 g/L of peptone and 16 g/L of agar. After growth all species were main-tained at 4 °C on culture medium. For biological treatment, themycelia of P. sajor cajuwere grown in a liquid mediumwith the samecomposition during 7 days and, incubated at 25 °C and 120±10 rpm.Mycelia of P. chrysosporium, T. versicolor and of R. oryzae wereobtained promoting growth at the same conditions but at 30 °C. Afterthis period of growth, the mycelia of different species were collectedby filtration with a sterilized gaze and kept in sterilized plasticcontainers at 4 °C, for a maximum of 24 h, before being applied to thebiological treatment of the final effluent from the kraft pulp mill.
2.2. Effluent source
Afinal effluent, from a bleached kraft pulpmill processing E. globulusafter secondary treatment, was collected in glass bottles, acidified at pH2 and kept at room temperature before analysis. Table 1 describes thechemical characterization of the final effluent.
2.3. Effluent treatment in batch reactor
The ligninolytic fungi P. sajor caju, P. chrysosporium, T. versicolor andR. oryzae were used to treat 220 mL of final effluent in batch reactors.According to previous work, the efficiency of biological treatmentdepends on the composition of the final effluent. Hence, the biologicaltreatment with P. sajor caju was performed on the final effluentenriched with 2 g/L of glucose and with pH adjusted to 5.5. For the
biological treatment with P. chrysosporium, T. versicolor or R. oryzae,the final effluent medium composition contained (per litter) 1 g ofglucose, 1.5 g of calcium chloride, 0.2 g of magnesium sulphate, 1.0 g ofpotassium dihydrogen phosphate and 0.15 g of ammonium chlorideand the pH was adjusted to 3.9–4.0. This composition was based onrecommendations made by Nagarathnamma and Bajpai (1999).
A batch reactor with 220 mL of final effluent was inoculated witheach fungus species mycelia filtrated from the culture medium. Thebiological treatment was performed incubating at 25 °C (P. sajor caju)or at 30 °C (P. chrysosporium, T. versicolor and R. oryzae) with stirring at120±10 rpm, for a maximum of 17 days. Two-replicated sub-samplesof effluent were withdrawn throughout the incubation period, atdifferent moments, in order to monitor the biological treatment of theeffluent performed by each of the species. According to the degrada-tion rate of each species, different periods of time were considered forsampling: 0, 3, 6, 10, 13 and 17 days for P. sajor caju; 0, 1, 2, 3, 6 and10 days for P. chrysosporium and R. oryzae; and 0, 2, 3 6, 8 and 13 daysfor T. versicolor. The replicated sub-samples of the treated final effluentwere analysed for the relative absorbance ratio, pH, biomass, COD,ligninolytic enzymes and toxicity. Average values and their corre-sponding standard deviation values are reported.
2.4. Analytical procedures
Absorbance scan (200–600 nm) of sub-samples of the finaleffluent treated by each species of fungus and collected at differentincubation periods was obtained by spectrophotometry (GBC/Cintra10e). Before absorbance measurements, the pH of sub-samples wasadjusted to 5.5 by addition of sodium hydroxide (0.1–1 M) or hydro-chloric acid (0.1–1 M), and the sub-samples were filtered throughGF/F (47 Ø mm, Whatman) glass-fibber filters and diluted 5 timeswith deionised water (pH=5.5). The pH measurements wereperformed with a pH meter (Crison) throughout the biologicaltreatment experiments. Fungi biomass (wet weight) was recordedafter filtration. CODwas determined by the standardmethod ASTMD1252-88 (1994).
Production of ligninolytic enzymes during batch biological treat-ment of final effluent by each fungus species was determined bymonitoring manganese dependent peroxidase (MnP), laccase (Lac)and lignin peroxidase (LiP) activities by spectrophotometry. MnPactivity was determined by oxidation of phenol red at 431 nm(ε431=27,250 M−1 cm−1) according to the method described byRoy andArchibald (1993). Assaymixtures contained 0.2mMofMnSO4,50 mM of sodium malonate (pH 4.5), 0.1 mM of H2O2, 0.067 mM ofphenol red and 500 µL aliquots of treated effluent. One unit of MnPactivitywas defined as 1 µmol phenol red oxidized perminute, at 30 °C.Lac activity was determined bymeasuring the oxidation of ABTS [2,2′-azinobis (3ethylbenzothiazoline-6-sulfonic acid) diammonium salt)]at 420nm(ε420=36,000M−1 cm−1) (Eggert et al.,1996).Assaymixturescontained 500 µM ABTS, 50 mM sodium tartrate buffer (pH 4.5) and500 µL aliquots of treated effluent. One unit of Lac activity was definedas 1 µmol ABTS+ formed per minute, at 25 °C. LiP activity was measuredaccording to the method of Heinfling et al. (1998). The oxidation ofveratryl alcohol to veratryl aldehyde was recorded at 310 nm(ε310=9300 M−1 cm−1). Assay mixtures contained 2.0 mM veratrylalcohol, 0.1 mM H2O2, 10 mM sodium tartrate (pH 3.0) and 500 µLaliquots of treated effluent. One unit of LiP activity was defined as
Table 1pH, COD (mg/L) and absorbance values at 250, 325, 400 and 465 nm of final effluent.
Sample pH COD(mg/L)
Abs250nm
Abs325nm
Abs400nm
Abs465nm
Final effluenta 7.0±0.2 392±2 5.071±0.008 1.373±0.015 0.407±0.003 0.204±0.001
a For biological treatment the final effluent was enriched with glucose or additiveswhich increased the initial COD values as well the respective absorbance values.
3283A.C. Freitas et al. / Science of the Total Environment 407 (2009) 3282–3289
Author's personal copy
1 µmol of veratryl alcohol oxidized to veratryl aldehyde perminute, at25 °C. Aliquots of treated effluent with different fungus werepreviously filtered through GF/F glass-fibber filters, treated with 2%(w/v) of polyvinylpolypyrrolidone (PVPP) for colour removal andcentrifuged at 5000 rpm.
The toxicity of the effluent sub-samples was assessed following theBasic Test protocol (Microbics Corporation, 1992) and using aMicrotox® Model 500 Analyser (Azur Environmental). The endpointmeasured by Microtox® assay is the decrease in the intensity of lightemitted by the luminescent marine bacteria — Vibrio fischeri — after5 min of exposure to several dilutions of the treated effluent withprevious osmotic adjustment of the samples. Hence, EC50–5 min averagevalues, corresponding to the effluent dilution causing a 50% reductionin the luminescence of the bacteria, were calculated for each effluentsub-samples collected. The calculations following the proceduresdescribed on the Basic Test protocols were performed using an Excelworksheet and included corrections to mitigate the differences incolour of the different treated effluents. The sensitivity of each freezedried bacteria samplewas periodically checked in our laboratory usingthe zinc sulphate (ZnSO4.7H2O) as reference substance.
2.5. Statistical analysis
Pearson's correlation coefficients (r) were calculated to investigateassociations between the different parameters monitored during thetreatment of the final effluent by the four species of fungus.Additionally, Principal Component Analysis (PCA) was performedwith SPSS 15.0 for Windows to find relationships among the differenteffluent sub-samples biologically treated and, their correspondingincubation periods, based on the chemical and biochemical para-meters evaluated to assess the effectiveness of each treatment. Formultivariate analysis, average values from the chemical, biochemicaland ecotoxicological parameters assessed on each sub-sample oftreated effluents were used to build a standardized data matrix.
Cluster analysis is a method for combining similar objects (treatedeffluents) into groups or clusters, which can be displayed in adendrogram helping to find structure in data (Quinn and Keough,2002). The normalized Euclidean distance was applied to standar-dized data, in order to calculate the matrix of dissimilarities betweenall pairs of sampling sites. In dendrograms sub-samples of treatedeffluents joining each other at the bottom of the diagram are mostsimilar, whereas those joining at the top are the most dissimilar(Gauch, 1982).
3. Results and discussion
Table 2 displays the pH variation occurred throughout thebiological treatment by each fungus species, the initial biomassinoculated, and the respective growth rate. Higher growth rates wereachieved by P. sajor caju and R. oryzae, in spite of the differences in the
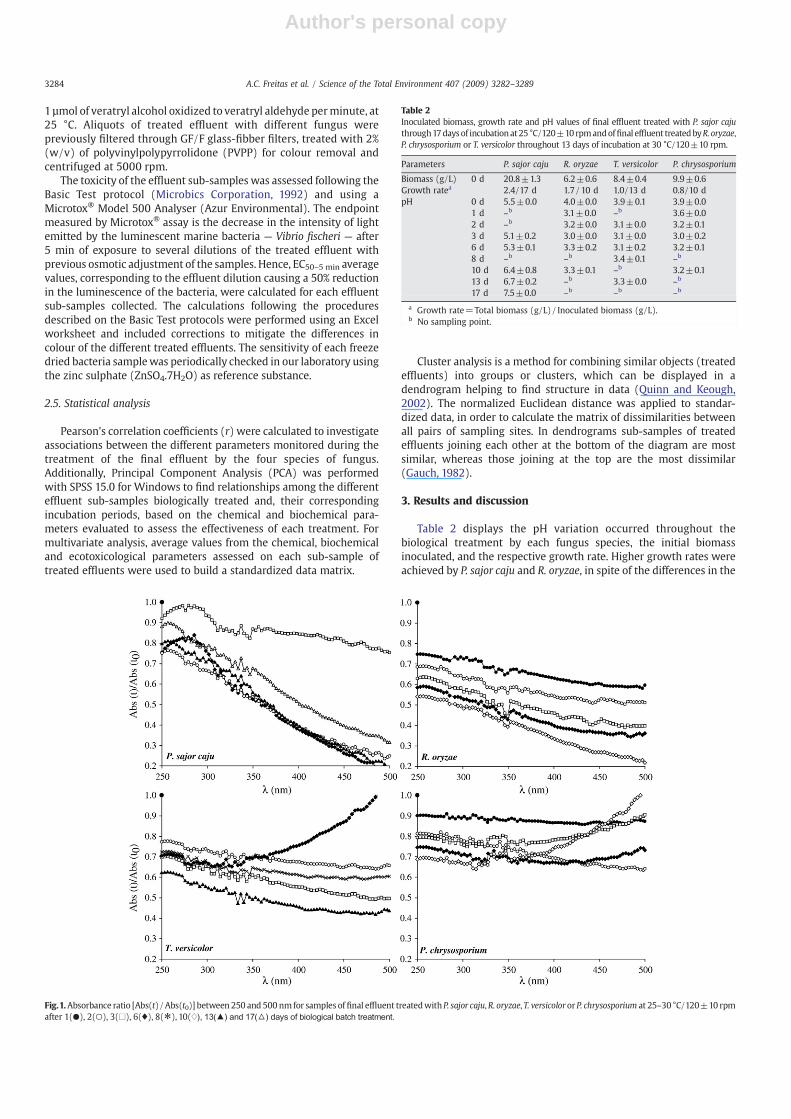
Fig.1.Absorbance ratio [Abs(t)/Abs(t0)] between 250 and 500 nmfor samples of final effluent treatedwithP. sajor caju,R. oryzae, T. versicolororP. chrysosporium at 25–30 °C/120±10 rpmafter 1(●), 2(○), 3(□), 6(♦), 8(*), 10(♢), 13(▲) and 17(△) days of biological batch treatment.
Table 2Inoculated biomass, growth rate and pH values of final effluent treated with P. sajor cajuthrough17daysof incubationat25 °C/120±10 rpmandoffinal effluent treatedbyR. oryzae,P. chrysosporium or T. versicolor throughout 13 days of incubation at 30 °C/120±10 rpm.
Parameters P. sajor caju R. oryzae T. versicolor P. chrysosporium
Biomass (g/L) 0 d 20.8±1.3 6.2±0.6 8.4±0.4 9.9±0.6Growth ratea 2.4/17 d 1.7/10 d 1.0/13 d 0.8/10 dpH 0 d 5.5±0.0 4.0±0.0 3.9±0.1 3.9±0.0
1 d –b 3.1±0.0 –b 3.6±0.02 d –b 3.2±0.0 3.1±0.0 3.2±0.13 d 5.1±0.2 3.0±0.0 3.1±0.0 3.0±0.26 d 5.3±0.1 3.3±0.2 3.1±0.2 3.2±0.18 d –b –b 3.4±0.1 –b
10 d 6.4±0.8 3.3±0.1 –b 3.2±0.113 d 6.7±0.2 –b 3.3±0.0 –b
17 d 7.5±0.0 –b –b –b
a Growth rate=Total biomass (g/L)/ Inoculated biomass (g/L).b No sampling point.
3284 A.C. Freitas et al. / Science of the Total Environment 407 (2009) 3282–3289
Author's personal copy
biomass inoculated. The initial biomass inoculated was based onpreliminaryexperiments performed to evaluate theminimumbiomassneeded, for each species, to perform successfully the biologicaltreatment. The pH profile throughout the period of incubation wasdifferent among the biological treatment with P. sajor caju and withother species under study. Increasing values of pHwere observed after10 days of treatment with P. sajor caju, reaching values higher than 7,which were well above its original value of 5. In opposition a decreasein pH was recorded throughout the incubation period for the otherspecies reaching values of 3 near their original values around 3.9 to 4.These pH variations throughout time of incubation point out for thepossible existence of different metabolic compounds produced byP. sajor caju which were responsible for increasing the pH of theeffluent and it was probably relatedwith addition of glucose instead ofthe additives suggested by Nagarathnamma and Bajpai (1999). Inprevious studies, performed by Belém et al. (2008), a final effluentfrom a bleached kraft pulp mill processing E. globulus, collected aftersecondary treatment, showed different pH profiles throughout theincubation period (14 days), at 25 °C and 120±10 rpm, with P. sajorcaju. Higher values of pH (4.9 to 7.6) were recorded for the effluentwith glucosewhereasmuch lower valueswere observed (3.0 to 5.4) forthe effluent with additives. Fungi are recognized for their great abilityto produce a large variety of extra cellular proteins, organic acids andother metabolites (Palma et al., 1999), being this process highlydependent from the substrates used by the fungi, which in turninfluences the pH of extracellular environment.
Fig. 1 shows the relative absorbance ratio [(Abs(t)/Abs(t0)]between 250 and 500 nm obtained for the final effluent treatedwith the four fungus species throughout different incubation periods.This procedure of scanning the absorbance for various wavelengthswas performed to complement the measurement of absorbanceat 465 nm. This single measure of decolourization, usually taken at
465 nm only, was thought to be rather limited when compared to thespectrum that is obtained, from the absorbance measurements oftreated final effluent at a range of wavelengths. Such scanningapproach is more innovative than classical procedures such as the oneused byWingate et al. (2005) in their study about colour remediationof pulp mill effluent by fungal enzymes. This author analysed colourreduction by a standard method, for colour determination in pulp andpaper wastewater (NCASI, 1971), in which colour is determined aschloroplatinate equivalent, by spectrometric measurement only at465 nm.
A decrease in the relative absorbance for all wavelengths and,throughout the incubation period, was observed in the final effluenteither treated by P. sajor caju or by R. oryzae. Lower values of relativeabsorbance were observed when using R. oryzae; after 1 day ofincubation at 30 °C, a reduction was recorded at 250 and 465 nm (25and 41%, respectively); these percentage values increased to 46 and74% after 10 days. Such trend at 465 nm was in agreement with datareported by Nagarathnamma and Bajpai (1999).
The final effluent sub-samples treated with P. sajor caju, after 6 to10 days of incubation, were, in general, associatedwith lower values ofrelative absorbance, i.e., reductions of 25 and 72%were attained at 250and 465 nm, respectively. Similar data was reported by Ragunathanand Swaminathan (2004) in kraft effluent treated with P. sajor caju.After 10 days of incubation the decreasing trend in absorbance wasreversed which could be related to secondary metabolic compoundsproduced by these species. This observation suggested that greatertime of incubation was not a positive factor for higher degradationrates of organic compounds present in the final effluent.
As also shown in Fig. 1, higher values of relative absorbance,with oscillations throughout time of incubation, were observed inthe biological treatment of the final effluent with T. versicolor or withP. chrysosporium, when compared with treatments made with theother two fungus species. Theminimumvalues of relative absorbancewere observed after 13 days of incubation with T. versicolor (38 and57% reduction at 250 and 465 nm, respectively). Comparable valueswere reported by Selvam et al. (2002) in the biological treatment ofkraft effluent with T. versicolor, after 4 days of incubation at 39 °C.Bergbauer et al. (1991) reported aswell about 90% of colour reductionin a bleach plant effluent, after 3 days of treatment. However theseresults were obtained for a kraft effluent collected from bleach plantwithout any chemical or biological treatment whereas in our studythe effluent was collected after secondary treatment. After 6 daysof incubation, the final effluent treated with T. versicolor becameturbid and this was responsible for the huge absorbance increase athigher wave lengths (N350 nm). This phenomenon could have beenrelated, at least in part, with the production of complex organiccompounds, or by the aggregation of more simple compounds.Polysaccharides production by T. versicolor through submersefermentation is a well documented fact (Kim et al., 2002; Cui andChisti, 2003).
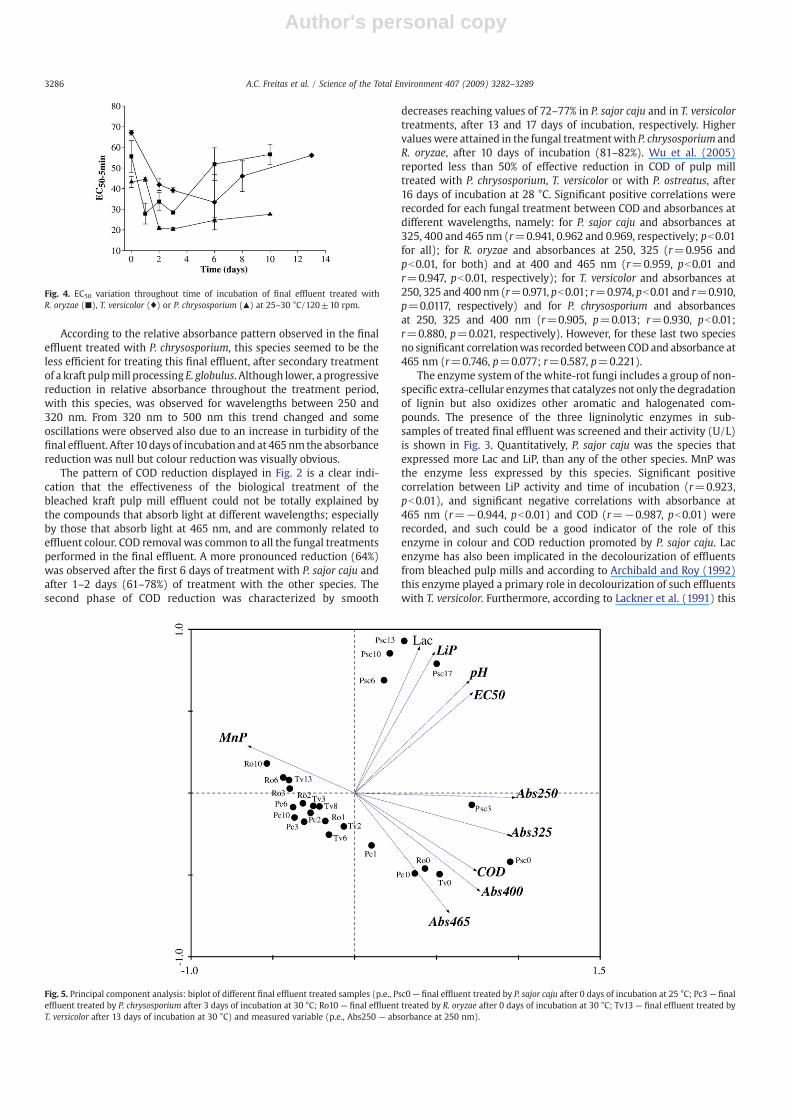
Fig. 3. Laccase,MnP and LiP activity throughout time of incubation offinal effluent treatedwith P. sajor caju (●),R. oryzae (■), T. versicolor (♦) or P. chrysosporium (▲) at 25–30 °C/120±10 rpm.
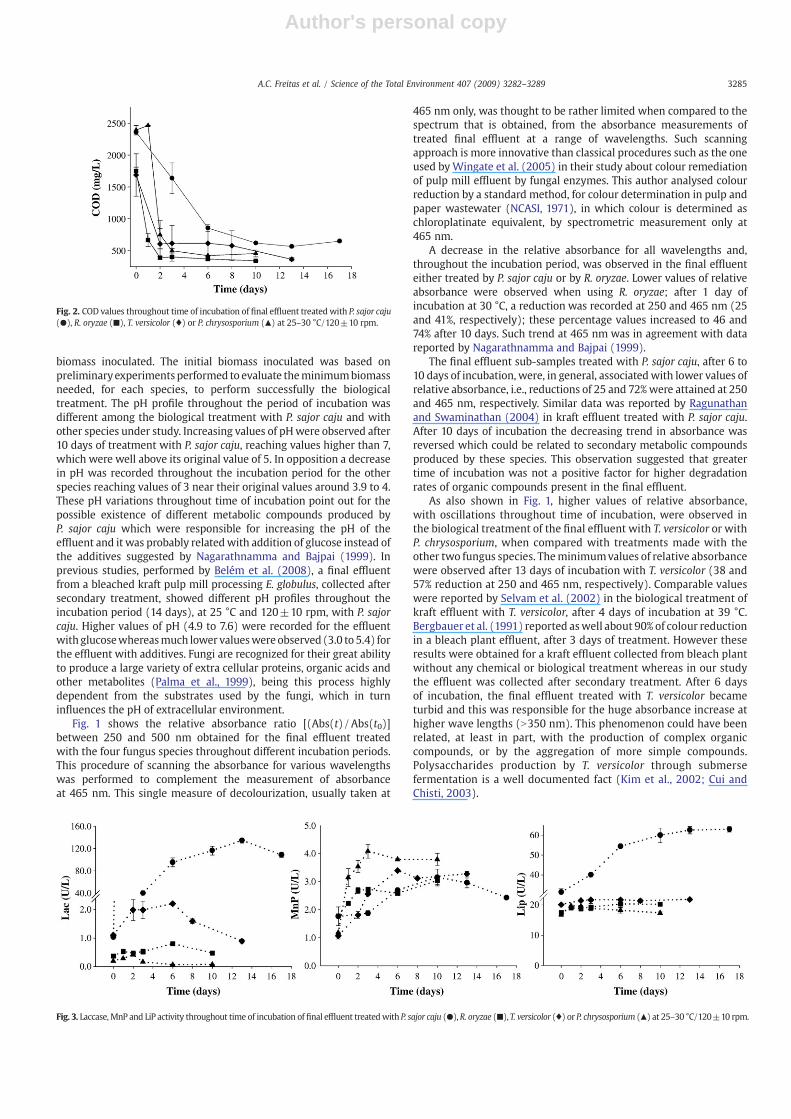
Fig. 2. COD values throughout time of incubation of final effluent treated with P. sajor caju(●), R. oryzae (■), T. versicolor (♦) or P. chrysosporium (▲) at 25–30 °C/120±10 rpm.
3285A.C. Freitas et al. / Science of the Total Environment 407 (2009) 3282–3289
Author's personal copy
According to the relative absorbance pattern observed in the finaleffluent treated with P. chrysosporium, this species seemed to be theless efficient for treating this final effluent, after secondary treatmentof a kraft pulpmill processing E. globulus. Although lower, a progressivereduction in relative absorbance throughout the treatment period,with this species, was observed for wavelengths between 250 and320 nm. From 320 nm to 500 nm this trend changed and someoscillations were observed also due to an increase in turbidity of thefinal effluent. After 10 days of incubation and at 465 nm the absorbancereduction was null but colour reduction was visually obvious.
The pattern of COD reduction displayed in Fig. 2 is a clear indi-cation that the effectiveness of the biological treatment of thebleached kraft pulp mill effluent could not be totally explained bythe compounds that absorb light at different wavelengths; especiallyby those that absorb light at 465 nm, and are commonly related toeffluent colour. COD removal was common to all the fungal treatmentsperformed in the final effluent. A more pronounced reduction (64%)was observed after the first 6 days of treatment with P. sajor caju andafter 1–2 days (61–78%) of treatment with the other species. Thesecond phase of COD reduction was characterized by smooth
decreases reaching values of 72–77% in P. sajor caju and in T. versicolortreatments, after 13 and 17 days of incubation, respectively. Highervalueswere attained in the fungal treatmentwith P. chrysosporium andR. oryzae, after 10 days of incubation (81–82%). Wu et al. (2005)reported less than 50% of effective reduction in COD of pulp milltreated with P. chrysosporium, T. versicolor or with P. ostreatus, after16 days of incubation at 28 °C. Significant positive correlations wererecorded for each fungal treatment between COD and absorbances atdifferent wavelengths, namely: for P. sajor caju and absorbances at325, 400 and 465 nm (r=0.941, 0.962 and 0.969, respectively; pb0.01for all); for R. oryzae and absorbances at 250, 325 (r=0.956 andpb0.01, for both) and at 400 and 465 nm (r=0.959, pb0.01 andr=0.947, pb0.01, respectively); for T. versicolor and absorbances at250, 325 and 400 nm (r=0.971, pb0.01; r=0.974, pb0.01 and r=0.910,p=0.0117, respectively) and for P. chrysosporium and absorbancesat 250, 325 and 400 nm (r=0.905, p=0.013; r=0.930, pb0.01;r=0.880, p=0.021, respectively). However, for these last two speciesno significant correlationwas recorded between COD and absorbance at465 nm (r=0.746, p=0.077; r=0.587, p=0.221).
The enzyme system of the white-rot fungi includes a group of non-specific extra-cellular enzymes that catalyzes not only the degradationof lignin but also oxidizes other aromatic and halogenated com-pounds. The presence of the three ligninolytic enzymes in sub-samples of treated final effluent was screened and their activity (U/L)is shown in Fig. 3. Quantitatively, P. sajor caju was the species thatexpressed more Lac and LiP, than any of the other species. MnP wasthe enzyme less expressed by this species. Significant positivecorrelation between LiP activity and time of incubation (r=0.923,pb0.01), and significant negative correlations with absorbance at465 nm (r=−0.944, pb0.01) and COD (r=−0.987, pb0.01) wererecorded, and such could be a good indicator of the role of thisenzyme in colour and COD reduction promoted by P. sajor caju. Lacenzyme has also been implicated in the decolourization of effluentsfrom bleached pulp mills and according to Archibald and Roy (1992)this enzyme played a primary role in decolourization of such effluentswith T. versicolor. Furthermore, according to Lackner et al. (1991) this
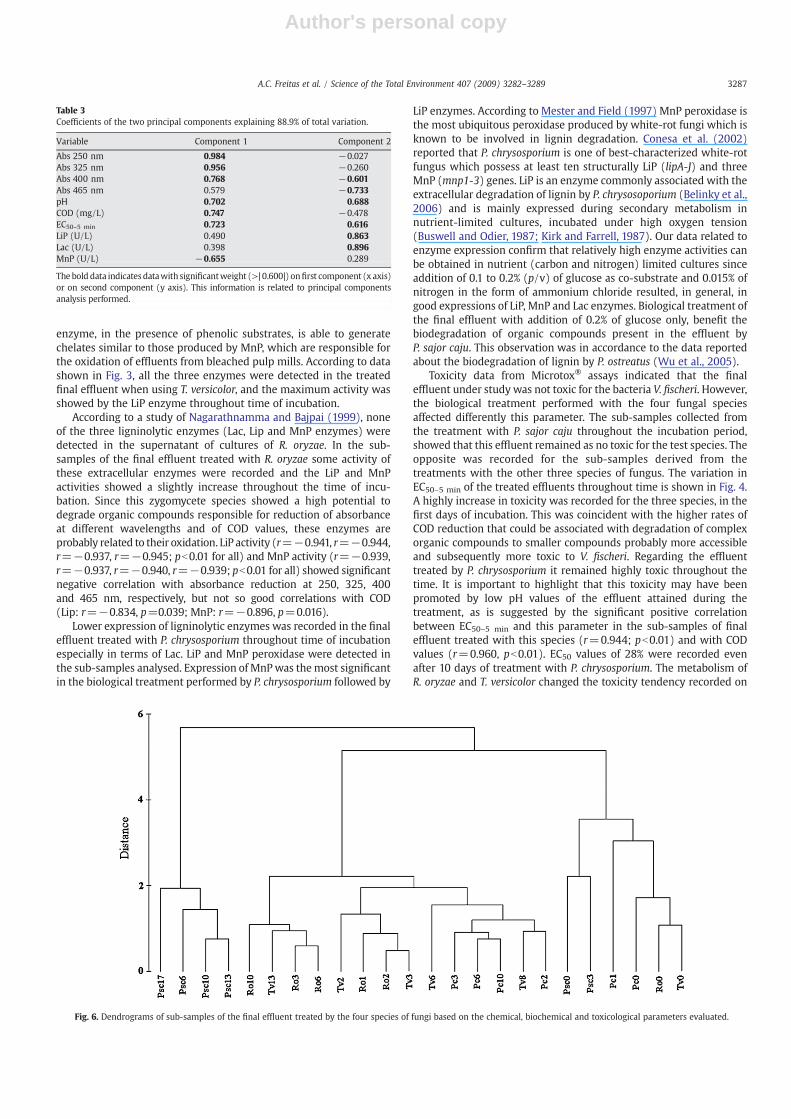
Fig. 4. EC50 variation throughout time of incubation of final effluent treated withR. oryzae (■), T. versicolor (♦) or P. chrysosporium (▲) at 25–30 °C/120±10 rpm.
Fig. 5. Principal component analysis: biplot of different final effluent treated samples (p.e., Psc0 — final effluent treated by P. sajor caju after 0 days of incubation at 25 °C; Pc3 — finaleffluent treated by P. chrysosporium after 3 days of incubation at 30 °C; Ro10 — final effluent treated by R. oryzae after 0 days of incubation at 30 °C; Tv13 — final effluent treated byT. versicolor after 13 days of incubation at 30 °C) and measured variable (p.e., Abs250 — absorbance at 250 nm).
3286 A.C. Freitas et al. / Science of the Total Environment 407 (2009) 3282–3289
Author's personal copy
enzyme, in the presence of phenolic substrates, is able to generatechelates similar to those produced by MnP, which are responsible forthe oxidation of effluents from bleached pulp mills. According to datashown in Fig. 3, all the three enzymes were detected in the treatedfinal effluent when using T. versicolor, and the maximum activity wasshowed by the LiP enzyme throughout time of incubation.
According to a study of Nagarathnamma and Bajpai (1999), noneof the three ligninolytic enzymes (Lac, Lip and MnP enzymes) weredetected in the supernatant of cultures of R. oryzae. In the sub-samples of the final effluent treated with R. oryzae some activity ofthese extracellular enzymes were recorded and the LiP and MnPactivities showed a slightly increase throughout the time of incu-bation. Since this zygomycete species showed a high potential todegrade organic compounds responsible for reduction of absorbanceat different wavelengths and of COD values, these enzymes areprobably related to their oxidation. LiP activity (r=−0.941, r=−0.944,r=−0.937, r=−0.945; pb0.01 for all) and MnP activity (r=−0.939,r=−0.937, r=−0.940, r=−0.939; pb0.01 for all) showed significantnegative correlation with absorbance reduction at 250, 325, 400and 465 nm, respectively, but not so good correlations with COD(Lip: r=−0.834, p=0.039; MnP: r=−0.896, p=0.016).
Lower expression of ligninolytic enzymes was recorded in the finaleffluent treated with P. chrysosporium throughout time of incubationespecially in terms of Lac. LiP and MnP peroxidase were detected inthe sub-samples analysed. Expression of MnPwas themost significantin the biological treatment performed by P. chrysosporium followed by
LiP enzymes. According to Mester and Field (1997) MnP peroxidase isthe most ubiquitous peroxidase produced by white-rot fungi which isknown to be involved in lignin degradation. Conesa et al. (2002)reported that P. chrysosporium is one of best-characterized white-rotfungus which possess at least ten structurally LiP (lipA-J) and threeMnP (mnp1-3) genes. LiP is an enzyme commonly associated with theextracellular degradation of lignin by P. chrysosoporium (Belinky et al.,2006) and is mainly expressed during secondary metabolism innutrient-limited cultures, incubated under high oxygen tension(Buswell and Odier, 1987; Kirk and Farrell, 1987). Our data related toenzyme expression confirm that relatively high enzyme activities canbe obtained in nutrient (carbon and nitrogen) limited cultures sinceaddition of 0.1 to 0.2% (p/v) of glucose as co-substrate and 0.015% ofnitrogen in the form of ammonium chloride resulted, in general, ingood expressions of LiP, MnP and Lac enzymes. Biological treatment ofthe final effluent with addition of 0.2% of glucose only, benefit thebiodegradation of organic compounds present in the effluent byP. sajor caju. This observation was in accordance to the data reportedabout the biodegradation of lignin by P. ostreatus (Wu et al., 2005).
Toxicity data from Microtox® assays indicated that the finaleffluent under study was not toxic for the bacteria V. fischeri. However,the biological treatment performed with the four fungal speciesaffected differently this parameter. The sub-samples collected fromthe treatment with P. sajor caju throughout the incubation period,showed that this effluent remained as no toxic for the test species. Theopposite was recorded for the sub-samples derived from thetreatments with the other three species of fungus. The variation inEC50–5 min of the treated effluents throughout time is shown in Fig. 4.A highly increase in toxicity was recorded for the three species, in thefirst days of incubation. This was coincident with the higher rates ofCOD reduction that could be associated with degradation of complexorganic compounds to smaller compounds probably more accessibleand subsequently more toxic to V. fischeri. Regarding the effluenttreated by P. chrysosporium it remained highly toxic throughout thetime. It is important to highlight that this toxicity may have beenpromoted by low pH values of the effluent attained during thetreatment, as is suggested by the significant positive correlationbetween EC50–5 min and this parameter in the sub-samples of finaleffluent treated with this species (r=0.944; pb0.01) and with CODvalues (r=0.960, pb0.01). EC50 values of 28% were recorded evenafter 10 days of treatment with P. chrysosporium. The metabolism ofR. oryzae and T. versicolor changed the toxicity tendency recorded on
Table 3Coefficients of the two principal components explaining 88.9% of total variation.
Variable Component 1 Component 2
Abs 250 nm 0.984 −0.027Abs 325 nm 0.956 −0.260Abs 400 nm 0.768 −0.601Abs 465 nm 0.579 −0.733pH 0.702 0.688COD (mg/L) 0.747 −0.478EC50–5 min 0.723 0.616LiP (U/L) 0.490 0.863Lac (U/L) 0.398 0.896MnP (U/L) −0.655 0.289
The bold data indicates datawith significantweight (>| 0.600|) onfirst component (x axis)or on second component (y axis). This information is related to principal componentsanalysis performed.
Fig. 6. Dendrograms of sub-samples of the final effluent treated by the four species of fungi based on the chemical, biochemical and toxicological parameters evaluated.
3287A.C. Freitas et al. / Science of the Total Environment 407 (2009) 3282–3289

Author's personal copy
the first days but, after 10 and 13 days, values of EC50–5 min becomesimilar to those recorded in the effluent immediately after theaddition of the fungus (0d). Surprisingly, the effluent that was nottoxic even after the addition of chemical compounds and pH adjustedto 3.9–4.0, becomes slightly toxic after the addition of fungus (exceptP. sajor caju). Some fungi are able to produce antimicrobial substancesthat are toxic to bacteria, and this may be a possible explanation forthis fact. For example polysaccharopeptides produced by Coriolus(Trametes) versicolor are beneficial in the therapy of opportunisticmicrobial infections that suppress the immune response (Cui andChisti, 2003).
In the PCA biplot (Fig. 5), three main groups can be discernedsetting apart along the first two components. One group (I) waslocated in the upper right quadrant and it included only the effluentsub-samples inoculated with P. sajor caju, for higher periods ofincubation (above 6 days). These sub-samples were correlated withhigher ligninolytic enzyme activity (Lac and LiP enzymes), higher pHvalues and higher EC50–5 min values (lower toxicity to V. fischeri) and allof these parameters were positively correlated with each other. Thegroup II, in lower right quadrant, included the sub-samples of non-treated final effluent (0d) inoculatedwith the four species of fungi andalso the final effluent treated with P. chrysosporium, after 1 day ofincubation at 30 °C, as well treated with P. sajor caju after 3 days ofincubation at 25 °C. All of these sub-samples were correlated withhigher values of COD and higher relative absorbance ratios. The thirdgroup III was set apart in the upper and lower left quadrants, and itincluded the final effluent sub-samples treated (2 to 13 days) withR. oryzae, P. chrysosporium or T. versicolor. In this third group, thesampleswith less incubation timewere, in general, joined in the lowerleft quadrant, in association to lower degrees of biological degradationwith high values of relative absorbance ratios and COD. In oppositionsamples with longer incubation timewere positioned in the upper leftquadrant, i.e., Ro6 and Ro10 (final effluent treated by R. oryzae after 6and 10 days of incubation at 30 °C, respectively) and Tv13 (final effluenttreated by T. versicolor after 13 days of incubation at 30 °C) which werecorrelated with higher activities of the MnP enzyme, lower values ofCODaswell as lower values of absorbance ratios. Thefirst twoaxesof thePCAexplained 88.9% of the total variation among the sub-samples of thefinal effluent treated by the four species of fungus, in terms of theparameters evaluated [absorbance at 250 325, 400 and at 465 nm, pH,COD, lignolytic enzymes (Lac, LiP andMnP) andEC50–5 min]. According tothe data shown in Table 3 and Fig. 5, all variables, contributed positivelyto the first component (x axis) except for MnP enzyme which was theonly variable with a negative contribution for the first component.Absorbances at 250, 325 and 400 nm as well COD and EC50–5 min werethe variables with more weight on this component. In the secondcomponent (y axis) several variables contributed negatively namely allabsorbances (250 to 465 nm) and COD. Absorbances at 465 nm, LiP andLac enzymes were the variables with more weight on the secondcomponent (y axis).
The dendogram derived from the cluster analysis (Fig. 6), helped todiscriminate among the sub-samples included in III group of the PCAbiplot, supporting the analysis previously performed and indicatingthree sub-groups in one of three major clusters. The sub-samplespresenting lower degree of biological degradation were groupedtogether and included those treated with P. chrysosporium (2 to10 days of incubation). Sub-samples treated with T. versicolor, during 6or 8 days, were also included in this subgroup probably due to thegreat increase of absorbance recorded in these sub-samples (Fig. 1).On the other hand, sub-samples with high levels of biologicaldegradation, some of them associated with longer incubation periodswere grouped together and included sub-samples treated withR. oryzae and T. versicolor (Ro3, Ro6, Ro10 and Tv 13). The third sub-group that could be visualized in the cluster was composed by theother sub-samples included in the group III of the PCA biplot, to whichintermediate levels of biological biodegradation were associated. A
high level of agreement was achieved between the composition ofgroups I and II from the PCA and the other two major clusters in thedendogram.
4. Conclusions
The four fungi species tested in this work confirmed their potentialfor future applications in the biological treatment of final effluentsderived from the secondary treatment of a bleached kraft pulp mill. Itis important to enhance that besides the secondary treatment of thiseffluent, its characteristics (ex. remaining colour, COD values, etc)indicate that additional treatment is important to decrease theirpotential to pollute the environment. The performance of the fourfungi species in the biological treatment resulted in remarkabledifferences in the ratios of relative absorbance but not so greatdifferences in COD effective reduction. The expression of ligninolyticenzymes varied among fungal species. While Lac and LiP activitiesseem to be correlated with the degradation of organic compounds, inthe effluent treated with P. sajor caju, MnP activity seemed to explainbetter the biological treatment performed by the other species, as itwas demonstrated by the PCA biplot. Among the four fungi speciesP. sajor caju and R. oryzae were the most effective in the biodegrada-tion of compounds present in the effluent, which are usually respon-sible for pollution, due to their great potential to reduce absorbance atdifferent wavelengths and to increase COD levels. However, sincetoxicity of effluents, in natural receptor systems, is also of concern,P. sajor caju revealed to be the only species able to perform thebiological treatment without promoting toxicity of the final effluentderived from the secondary treatment of a bleached kraft pulp mill.
Acknowledgments
This work has been developed under the scope of the FCT(Portugal) funded research project, POCT/CTA/45030/2002: ‘Deco-lourisation of effluents from pulp and paper mills: removal of organiccompounds and toxicity (DECORTOX)’.
References
Archibald F, Roy B. Production ofmanganic chelates by laccase from the lignin degradingfungus Trametes (Coriolus) versicolor. Appl Environ Microbiol 1992;l58:1496–9.
ASTM. Standard test methods for chemical oxygen demand of water. Report D 1252-88.Annual Book of ASTM Standards Philadelphia, USA: American Society for Testingand Material; 1994.
Bajpai P, Bajpai PK. Biological colour removal of pulp and paper mill wastewaters.J Biotechnol 1994;33:211–20.
Belém A, Panteleitchouk AV, Duarte AC, Rocha-Santos TAP, Freitas AC. Treatment of theeffluent from a kraft bleach plant with white-rot fungi: Pleurotus sajor caju andPleurotus ostreatus. Global NEST J 2008;10:426–31.
Belinky PA, Flikshtein N, Dosoretz C. Induction of lignin peroxidase via reactive oxygenspecies in manganese-deficient cultures of Phanerochaete chrysosporium. EnzMicrobiol Technol 2006;39:222–8.
Bergbauer M, Eggert C, Kraepelin G. Degradation of chlorinated lignin compounds in ableach plant effluent by the white-rot fungus Trametes versicolor. Appl MicrobiolBiotechnol 1991;35:105–9.
BlanchetteR.Delignificationbywood-decay fungi. AnnualRevPhytopathol 1991;29:381–98.Buswell JA, Odier E. Lignin biodegradation. Crit Rev Biotechnol 1987;6:1–60.Camarero S, Ibarra D, Martínez AT, Romero J, Gutiérrez A, del Río JC. Paper pulp
delignification using laccase and natural mediators. Enz Microbiol Technol2007;40: 1264–71.
Conesa A, Punt PJ, van den Hondel CAMJJ. Fungal peroxidases: molecular aspects andapplications. J Biotechnol 2002;93:143–58.
Cui J, Chisti Y. Polysaccharopeptides of Coriolus versicolor: physiological activity, usesand production. Biotechnol Adv 2003;21:109–22.
Cullen D, Kersten PJ. Enzymology and molecular biology of lignin degradation. In:Bramble R, Marzulf GA, editors. The Micota III, Biochemistry and Molecular Biology.2nd Edition. Berlin-Heidelberg, Germany: Springer-Verlag; 2004.
Daniel G. Use of electron microscopy for aiding our understanding of woodbiodegradation. FEMS Microbiol Rev 1994;13:199–233.
Durant N, Esposito E, Innicentini-Mei LH, Canhos PV. A new alternative process for krafteffluent treatment. Biodegradation 1994;5:13–9.
Eggert C, Temp U, Jeffrey DFD, Karl-Erik L, Eriksson L. A fungal metabolite mediatesdegradation of non-phenolic lignin structures and synthetic lignin by laccase. FEBSLetters 1996;391:144–8.
3288 A.C. Freitas et al. / Science of the Total Environment 407 (2009) 3282–3289

Author's personal copy
Eriksson KEl, Blanchette RA, Ander P. Microbial and enzymatic degradation of wood andwood components. Berlin Heidelberg New York, USA: Springer; 1990.
Esposito E, Canhos VP, DuranN. Screening of lignin degrading fungi for removal of colourfrom kraft mill wastewater with no additional extra carbon source. BiotechnolLetters 1991;13:571–6.
Gauch HGJ. Multivariate analysis in community ecology. New York, USA: CambridgeUniversity Press; 1982.
Heinfling A, Martínez MJ, Martínez AT, Bergbauer M, Szewzyk U. Transformation onindustrial dyes by manganese peroxidases from Bjerkandera adusta and Pleurotuseryngii in a manganese-independent reaction. Appl Env Microbiol 1998;64:2788–93.
Kim SW, Hwang HJ, Park JP, Cho YJ, Song CH, Yun JW. Mycelia growth and exo-biopolymer production by submerged culture of various edible mushrooms underdifferent media. Letters Appl Microbiol 2002;34:56–61.
Kimura Y, Asada Y, Kuwahara M. Screening of basidiomycetes for lignin peroxidasegenes using a DNA probe. Appl Microbiol Biotechnol 1990;32:436–42.
Kirk TK, Farrell RL. Enzymatic “combustion”: themicrobial degradation of lignin. AnnualRev Microbiol 1987;41:465–506.
Lackner R, Srebotnik E, Messner K. Oxidative degradation of high molecular weightchlorolignin by manganese peroxidase of Phanerochaete chrysosporium. BiochemBiophys Res Commun 1991;178:1092–8.
Lankinen VP, Inkeroninen MM, Pellinen J, Hatakka AI. The cost of lignin modifyingenzyme, decreases of AOX and colour removal bywhite-rot fungi: growth on bleachplant effluent. Water Sci Technol 1990;24:189–98.
Mester T, Field JA. Optimization of manganese peroxidase production by white rotfungus Bjerkandera sp. strain BOS55. FEMS Microbiol Lett 1997;155:161–8.
Microbics Corporation. Basic test detailed protocol. Microtox® Manual DetailedProtocols, vol. II. ; 1992. p. 107–27. Carlsbad, CA.
Nagarathnamma R, Bajpai P. Decolourization and detoxification of extraction-stageeffluent from chlorine bleaching of kraft pulp by Rhizopus oryzae. Applied EnvMicrobiol 1999;65:1078–82.
NCASI (National Council of the Paper Industry for Air and Stream Improvement). TechBull No. 253. USA: New York; 1971.
Palma C, Moreira MT, Mielgo I, Feijoo G, Lema JM. Use of fungal bioreactor as apretreatment or post-treatment step for continuous decolourisation of dyes. WatSci Technol 1999;40:131–6.
Pedroza AM, Rodolfo M, Alonso-Vante N, Rodríguez-Vásquez R. Sequential treatmentvia Trametes versicolor and UV/TiO2/RuxSey to reduce contaminants in wastewater resulting from the bleaching process during paper production. Chemosphere2007;67:793–801.
Pelaez F, Martinez MJ, Martinez AT. Screening of 68 species of basidiomycetes forenzymes involved in lignin degradation. Mycol Res 1995;99:37–42.
Pokhrel D, Viraraghavan T. Treatment of pulp and paper mill wastewater— a review. SciTotal Environ 2004;333:37–58.
Quinn GP, Keough MJ. Experimental design and data analysis for biologists. Cambridge,UK: Cambridge University Press; 2002.
Ragunathan R, Swaminathan K. Biological treatment of a pulp and paper industryeffluent by Pleurotus spp. World J Microbiol Biotechnol 2004;20:389–93.
Roy BP, Archibald F. Effects of kraft pulp and lignin on Trametes versicolor carbonmetabolism. Appl Env Microbiol 1993;59:1855–63.
Sakurai A, Yamamoto T, Makabe A, Kinoshita S, Sakakibara M. Removal of lignin in aliquid system by na isolated fungus. J Chem Technol Biotechnol 2001;77:9–14.
Selvam K, Swaminathan K, Song MH, Chae K. Biological treatment of a pulp and paperindustryeffluent by Fomes lividus and Trametes versicolor.World JMicrobiol Biotechnol2002;18:523–6.
Ünal A, Kolankaya N. Dechlorination of bleached Kraft pulp by laccase enzyme producedfrom some white-rot fungi. Turk J Biol 2001;25:67–72.
Valls C, Roncero MB. Using both xylanase and laccase enzymes for pulp bleaching.Bioresour Technol 2009;100:2032–9.
Varela E, Martinez AT, Martinez MJ. Southern blot screening for lignin peroxidase andaryl-alcohol oxidase genes in 30 fungal species. J Biotechnol 2000;83:245–51.
Witayakran S, Ragauskas AJ. Modification of high-lignin softwood Kraft pulp withlaccase and amino acids. Enz Microbiol Technol 2009;44:176–81.
Wingate KG, Stuthridge T, Mansfield SD. Colour remediation of pulp mill effluent usingpurified fungal cellobiose dehydrogenase: reaction optimisation and mechanism ofdegradation. Biotechnol Bioeng 2005;90:95–106.
Wu J, Xiao Y, Yu H. Degradation of lignin in pulp mill wastewaters by white-rot fungi onbiofilm. Bioresour Technol 2005;96:1357–63.
3289A.C. Freitas et al. / Science of the Total Environment 407 (2009) 3282–3289
Related Documents











