Crop Protection 24 (2005) 197–206 Review Biological control of weeds and plant pathogens in paddy rice by exploiting plant allelopathy: an overview $ Tran Dang Xuan a , Tawata Shinkichi a , Tran Dang Khanh b , Chung Ill Min b, a Department of Bioscience & Biotechnology, Faculty of Agriculture, University of the Ryukyus, Okinawa 903-0213, Japan b Research Team of Friendly Environmental Low Input Natural Herbicide New Material Study, Konkuk University, Seoul 143-701, Korea Received 2 June 2004; received in revised form 11 August 2004; accepted 18 August 2004 Abstract The allelopathic properties of plants can be exploited successfully as a tool for pathogen and weed reduction. Over a decade, preliminary surveys of some hundred allelopathic plants in the Southeast Asia and Japan ecosystems were made, and more than 30 species including crops which exhibited the greatest allelopathic potential were selected and examined for their impacts on emergence of pathogens and weeds. Observations showed that application of these plant materials at 1–2 tonne ha 1 can reduce weed biomass by about 70%, and increase rice yield by about 20%. Some species showed strong inhibition on major plant pathogens and they might become effective tools in reducing plant pathogens and weeds. However, application of 1–2 tonne ha 1 of plant material to the field makes heavy fieldwork. Numerous growth inhibitors identified from these allelopathic plants are responsible for their allelopathic properties and may be a useful source for the future development of bio-herbicides and pesticides. r 2004 Elsevier Ltd. All rights reserved. Keywords: Allelopathy; Allelochemical; Inhibition; Pest; Rice; Weed ARTICLE IN PRESS www.elsevier.com/locate/cropro Contents 1. Introduction ............................................................................. 198 2. Effects on paddy weeds ..................................................................... 198 2.1. Screening for plants with strong allelopathic potential ........................................... 198 2.2. Dose of treatments .................................................................... 198 2.3. Frequency of treatment................................................................. 199 2.4. Effect of allelopathic plants on weed species is selective .......................................... 200 2.5. Varietal effects on weed control ........................................................... 201 3. Effects on plant fungi ....................................................................... 201 4. Allelochemicals ........................................................................... 203 0261-2194/$ - see front matter r 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.cropro.2004.08.004 $ This paper is dedicated to Professor Tsuzuki Eiji, an Allelopathy Pioneer in Japan. Corresponding author. Fax: +822-446-7856. E-mail address: [email protected] (I.M. Chung).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
Contents
1. Introduc
2. Effects o
2.1. Sc
2.2. Do
2.3. Fr
2.4. Ef
2.5. Va
3. Effects o
4. Alleloche
0261-2194/$ - se
doi:10.1016/j.cr
$This paper�CorrespondE-mail addr
Crop Protection 24 (2005) 197–206
www.elsevier.com/locate/cropro
Review
Biological control of weeds and plant pathogens in paddy rice byexploiting plant allelopathy: an overview$
Tran Dang Xuana, Tawata Shinkichia, Tran Dang Khanhb, Chung Ill Minb,�
aDepartment of Bioscience & Biotechnology, Faculty of Agriculture, University of the Ryukyus, Okinawa 903-0213, JapanbResearch Team of Friendly Environmental Low Input Natural Herbicide New Material Study, Konkuk University, Seoul 143-701, Korea
Received 2 June 2004; received in revised form 11 August 2004; accepted 18 August 2004
Abstract
The allelopathic properties of plants can be exploited successfully as a tool for pathogen and weed reduction. Over a decade,
preliminary surveys of some hundred allelopathic plants in the Southeast Asia and Japan ecosystems were made, and more than 30
species including crops which exhibited the greatest allelopathic potential were selected and examined for their impacts on
emergence of pathogens and weeds. Observations showed that application of these plant materials at 1–2 tonne ha�1 can reduce
weed biomass by about 70%, and increase rice yield by about 20%. Some species showed strong inhibition on major plant
pathogens and they might become effective tools in reducing plant pathogens and weeds. However, application of 1–2 tonne ha�1 of
plant material to the field makes heavy fieldwork. Numerous growth inhibitors identified from these allelopathic plants are
responsible for their allelopathic properties and may be a useful source for the future development of bio-herbicides and pesticides.
r 2004 Elsevier Ltd. All rights reserved.
Keywords: Allelopathy; Allelochemical; Inhibition; Pest; Rice; Weed
tion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 198
n paddy weeds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 198
reening for plants with strong allelopathic potential . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 198
se of treatments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 198
equency of treatment. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199
fect of allelopathic plants on weed species is selective . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 200
rietal effects on weed control. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 201
n plant fungi. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 201
micals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 203
e front matter r 2004 Elsevier Ltd. All rights reserved.
opro.2004.08.004
is dedicated to Professor Tsuzuki Eiji, an Allelopathy Pioneer in Japan.
ing author. Fax: +822-446-7856.
ess: [email protected] (I.M. Chung).

ARTICLE IN PRESS
5. Suggestions and future prospects. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 203
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 205
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 205
T.D. Xuan et al. / Crop Protection 24 (2005) 197–206198
1. Introduction
The current trend is to find a biological solution tominimize the perceived hazardous impacts from herbi-cides and insecticides in agriculture production. Allelo-pathy is defined as a beneficial or detrimental effect froma donor plant to the recipient by chemical pathway(Rice, 1984). The harmful impact of allelopathy can beexploited for pest and weed control (Narwal, 1994;Kohli et al., 1998). Much research has documented thepotential of allelopathic plants to reduce pathogens andaffect weed emergence. An explanation of the allelo-pathy mechanism has been attempted and the influencefrom a complex of phytochemicals synthesized in plantsas secondary metabolites has been proposed. Theallelochemicals can be broadly classified as plantphenolics and terpenoids, which show great chemicaldiversity and are involved in a number of metabolic andecological processes. The most common effects ofallelochemicals may occur through leaching, volatiliza-tion, root exudation, and the death and decay of thefallen plant parts are either via biotic or abiotic means(Anaya et al., 1990).The question of what allelopathic plants should be
selected, how they are applied, and their benefits shouldbe seen as a requisite before introducing them to thefarmers for field usage. This report reviews our researchcarried out in a period of 10 years (1993–2003) on theuse of allelopathic plants for pathogen and weed controlin paddy rice.
2. Effects on paddy weeds
2.1. Screening for plants with strong allelopathic
potential
This is a prerequisite before a programme of pathogenand weed control starts. The screening is aimed atvarious crops and plants in the ecosystems to search forspecies with strong allelopathic properties, beforeexamining their effects on pathogen and weeds. Thescreening should meet certain criteria: (a) to assessaccurately the allelopathic activity excluding the com-petitive resources (viz. nutrients, light, water, and otherfactors), (b) in a minimum space, a large number ofplant varieties can be screened in a relatively shortperiod of time, (c) need simple procedures, experimentalmaterials, implements and reagents, and (d) be relatively
inexpensive and could be carried out all year round.Furthermore, indicator plants are the crucial factors toaccurately determine allelopathic impact. Lactuca sativa
L. (lettuce), Raphanus sativus L. (radish), Lepidium
sativum (cress), or Amaranthus spp. have been used astested plants in the screenings. However, these plants aretoo sensitive to chemicals at low concentration, as theysometimes over-estimate the actual allelopathy (Olofs-dotter, 2001). These plants may be more useful in thetrials for evaluating the varietal difference of allelo-pathic potential, in which disparity among plantcultivars is small. The uses of common crop plants,vegetables, and major noxious weeds may bring otheragronomic impacts.The preliminary requirements to determine plants
collected for screening should include: (a) their inva-siveness and area in the plant ecosystems; (b) the plantshave less natural weed density in their canopy andsurroundings than other plants in their ecosystem; and(c) are traditionally used as green manure, weed or pestmanagement by local farmers (Hong et al., 2003).A preliminary survey on plants and crops in the
ecosystems of Southeast Asia and Japan was conductedbased upon these criteria. The potential crops and plantsare listed in Table 1. The allelopathic potential fromtheir different plant parts (leaves, stem, and root) wasevaluated, and the average magnitude of inhibition onindicator plants are shown. Plants with strong allelo-pathic potential were selected for applying to the paddyfields to control weed emergence, as well as a source forgrowth inhibitor identification.
2.2. Dose of treatments
Numerous greenhouse and field trials were conductedto examine the most appropriate concentration that canreduce maximum weed emergence. Species showingmore than 20% inhibition were selected for weedcontrol purposes. Although rice hull did not reach20% of suppression (15.9%), it was included as it isreadily available and has been regularly incorporatedinto paddy field by local farmers. The effect of theseallelopathic plants on paddy weeds and rice yield arelisted in Table 2.It was found that a dose lower than 1 tonne ha�1 can
inhibit weed growth, but did not significantly suppressemergence of major paddy weeds such as Echinochloa
crus-galli, Monochoria vaginalis, Rotala indica, Cyperus
difformis, Digitaria ciliaris, and Azadirachta indica
ARTICLE IN PRESS
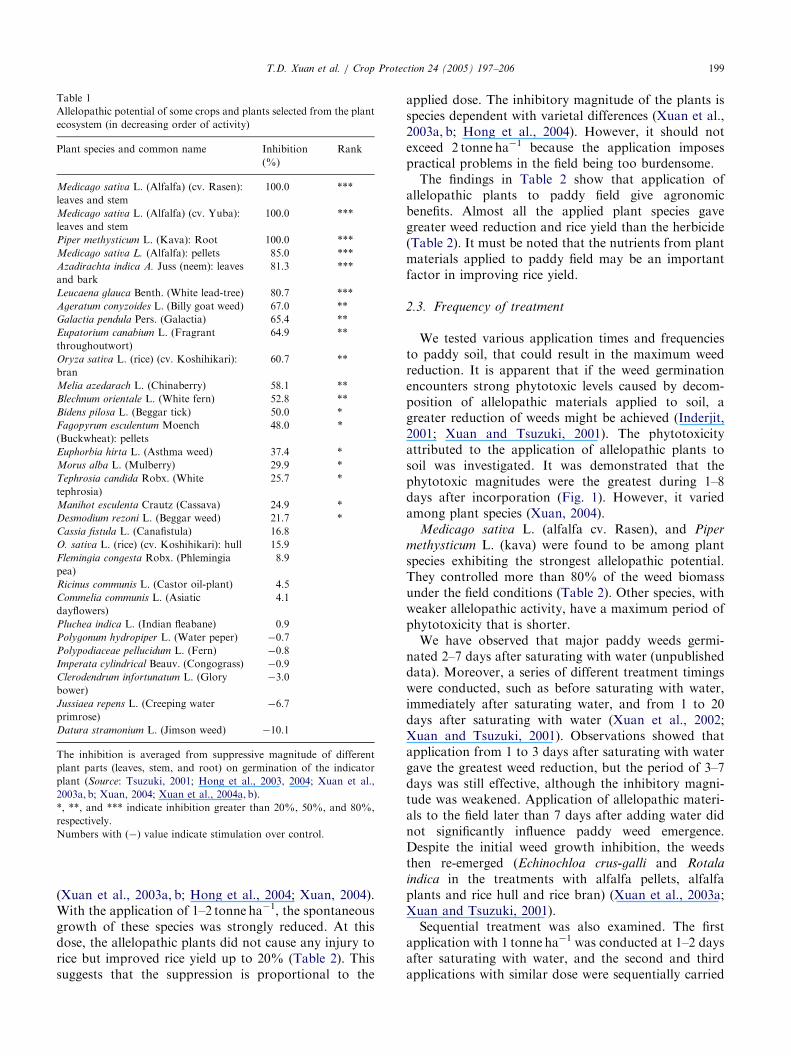
Table 1
Allelopathic potential of some crops and plants selected from the plant
ecosystem (in decreasing order of activity)
Plant species and common name Inhibition
(%)
Rank
Medicago sativa L. (Alfalfa) (cv. Rasen):
leaves and stem
100.0 ***
Medicago sativa L. (Alfalfa) (cv. Yuba):
leaves and stem
100.0 ***
Piper methysticum L. (Kava): Root 100.0 ***
Medicago sativa L. (Alfalfa): pellets 85.0 ***
Azadirachta indica A. Juss (neem): leaves
and bark
81.3 ***
Leucaena glauca Benth. (White lead-tree) 80.7 ***
Ageratum conyzoides L. (Billy goat weed) 67.0 **
Galactia pendula Pers. (Galactia) 65.4 **
Eupatorium canabium L. (Fragrant
throughoutwort)
64.9 **
Oryza sativa L. (rice) (cv. Koshihikari):
bran
60.7 **
Melia azedarach L. (Chinaberry) 58.1 **
Blechnum orientale L. (White fern) 52.8 **
Bidens pilosa L. (Beggar tick) 50.0 *
Fagopyrum esculentum Moench
(Buckwheat): pellets
48.0 *
Euphorbia hirta L. (Asthma weed) 37.4 *
Morus alba L. (Mulberry) 29.9 *
Tephrosia candida Robx. (White
tephrosia)
25.7 *
Manihot esculenta Crautz (Cassava) 24.9 *
Desmodium rezoni L. (Beggar weed) 21.7 *
Cassia fistula L. (Canafistula) 16.8
O. sativa L. (rice) (cv. Koshihikari): hull 15.9
Flemingia congesta Robx. (Phlemingia
pea)
8.9
Ricinus communis L. (Castor oil-plant) 4.5
Commelia communis L. (Asiatic
dayflowers)
4.1
Pluchea indica L. (Indian fleabane) 0.9
Polygonum hydropiper L. (Water peper) �0.7
Polypodiaceae pellucidum L. (Fern) �0.8
Imperata cylindrical Beauv. (Congograss) �0.9
Clerodendrum infortunatum L. (Glory
bower)
�3.0
Jussiaea repens L. (Creeping water
primrose)
�6.7
Datura stramonium L. (Jimson weed) �10.1
The inhibition is averaged from suppressive magnitude of different
plant parts (leaves, stem, and root) on germination of the indicator
plant (Source: Tsuzuki, 2001; Hong et al., 2003, 2004; Xuan et al.,
2003a, b; Xuan, 2004; Xuan et al., 2004a, b).
*, **, and *** indicate inhibition greater than 20%, 50%, and 80%,
respectively.
Numbers with (�) value indicate stimulation over control.
T.D. Xuan et al. / Crop Protection 24 (2005) 197–206 199
(Xuan et al., 2003a, b; Hong et al., 2004; Xuan, 2004).With the application of 1–2 tonne ha�1, the spontaneousgrowth of these species was strongly reduced. At thisdose, the allelopathic plants did not cause any injury torice but improved rice yield up to 20% (Table 2). Thissuggests that the suppression is proportional to the
applied dose. The inhibitory magnitude of the plants isspecies dependent with varietal differences (Xuan et al.,2003a, b; Hong et al., 2004). However, it should notexceed 2 tonne ha�1 because the application imposespractical problems in the field being too burdensome.The findings in Table 2 show that application of
allelopathic plants to paddy field give agronomicbenefits. Almost all the applied plant species gavegreater weed reduction and rice yield than the herbicide(Table 2). It must be noted that the nutrients from plantmaterials applied to paddy field may be an importantfactor in improving rice yield.
2.3. Frequency of treatment
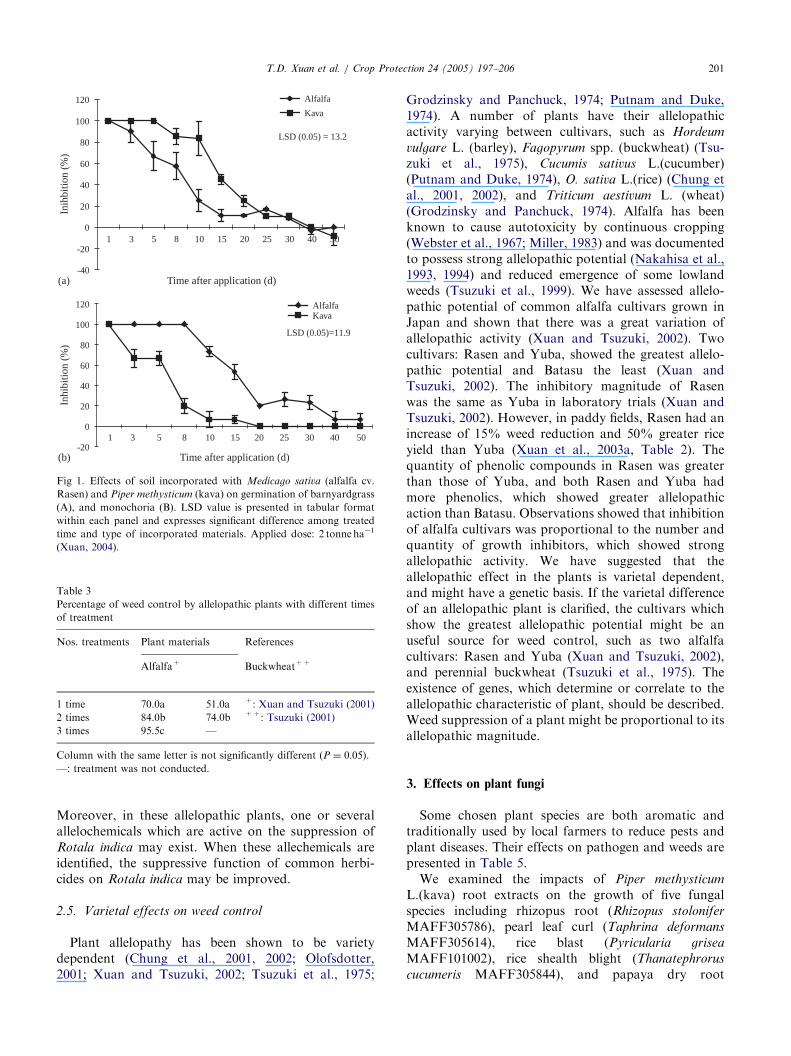
We tested various application times and frequenciesto paddy soil, that could result in the maximum weedreduction. It is apparent that if the weed germinationencounters strong phytotoxic levels caused by decom-position of allelopathic materials applied to soil, agreater reduction of weeds might be achieved (Inderjit,2001; Xuan and Tsuzuki, 2001). The phytotoxicityattributed to the application of allelopathic plants tosoil was investigated. It was demonstrated that thephytotoxic magnitudes were the greatest during 1–8days after incorporation (Fig. 1). However, it variedamong plant species (Xuan, 2004).
Medicago sativa L. (alfalfa cv. Rasen), and Piper
methysticum L. (kava) were found to be among plantspecies exhibiting the strongest allelopathic potential.They controlled more than 80% of the weed biomassunder the field conditions (Table 2). Other species, withweaker allelopathic activity, have a maximum period ofphytotoxicity that is shorter.We have observed that major paddy weeds germi-
nated 2–7 days after saturating with water (unpublisheddata). Moreover, a series of different treatment timingswere conducted, such as before saturating with water,immediately after saturating water, and from 1 to 20days after saturating with water (Xuan et al., 2002;Xuan and Tsuzuki, 2001). Observations showed thatapplication from 1 to 3 days after saturating with watergave the greatest weed reduction, but the period of 3–7days was still effective, although the inhibitory magni-tude was weakened. Application of allelopathic materi-als to the field later than 7 days after adding water didnot significantly influence paddy weed emergence.Despite the initial weed growth inhibition, the weedsthen re-emerged (Echinochloa crus-galli and Rotala
indica in the treatments with alfalfa pellets, alfalfaplants and rice hull and rice bran) (Xuan et al., 2003a;Xuan and Tsuzuki, 2001).Sequential treatment was also examined. The first
application with 1 tonne ha�1 was conducted at 1–2 daysafter saturating with water, and the second and thirdapplications with similar dose were sequentially carried
ARTICLE IN PRESS
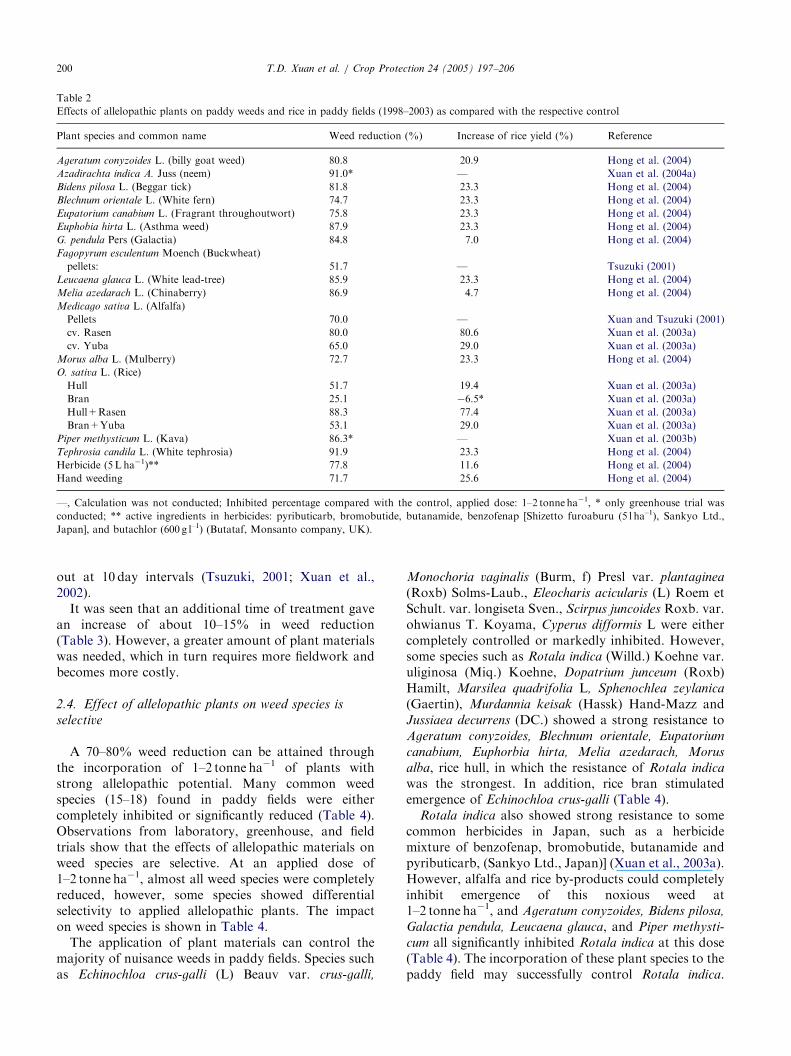
Table 2
Effects of allelopathic plants on paddy weeds and rice in paddy fields (1998–2003) as compared with the respective control
Plant species and common name Weed reduction (%) Increase of rice yield (%) Reference
Ageratum conyzoides L. (billy goat weed) 80.8 20.9 Hong et al. (2004)
Azadirachta indica A. Juss (neem) 91.0* — Xuan et al. (2004a)
Bidens pilosa L. (Beggar tick) 81.8 23.3 Hong et al. (2004)
Blechnum orientale L. (White fern) 74.7 23.3 Hong et al. (2004)
Eupatorium canabium L. (Fragrant throughoutwort) 75.8 23.3 Hong et al. (2004)
Euphobia hirta L. (Asthma weed) 87.9 23.3 Hong et al. (2004)
G. pendula Pers (Galactia) 84.8 7.0 Hong et al. (2004)
Fagopyrum esculentum Moench (Buckwheat)
pellets: 51.7 — Tsuzuki (2001)
Leucaena glauca L. (White lead-tree) 85.9 23.3 Hong et al. (2004)
Melia azedarach L. (Chinaberry) 86.9 4.7 Hong et al. (2004)
Medicago sativa L. (Alfalfa)
Pellets 70.0 — Xuan and Tsuzuki (2001)
cv. Rasen 80.0 80.6 Xuan et al. (2003a)
cv. Yuba 65.0 29.0 Xuan et al. (2003a)
Morus alba L. (Mulberry) 72.7 23.3 Hong et al. (2004)
O. sativa L. (Rice)
Hull 51.7 19.4 Xuan et al. (2003a)
Bran 25.1 �6.5* Xuan et al. (2003a)
Hull+Rasen 88.3 77.4 Xuan et al. (2003a)
Bran+Yuba 53.1 29.0 Xuan et al. (2003a)
Piper methysticum L. (Kava) 86.3* — Xuan et al. (2003b)
Tephrosia candila L. (White tephrosia) 91.9 23.3 Hong et al. (2004)
Herbicide (5L ha�1)** 77.8 11.6 Hong et al. (2004)
Hand weeding 71.7 25.6 Hong et al. (2004)
—, Calculation was not conducted; Inhibited percentage compared with the control, applied dose: 1–2 tonne ha�1, * only greenhouse trial was
conducted; ** active ingredients in herbicides: pyributicarb, bromobutide, butanamide, benzofenap [Shizetto furoaburu (5 l ha–1), Sankyo Ltd.,
Japan], and butachlor (600 g l–1) (Butataf, Monsanto company, UK).
T.D. Xuan et al. / Crop Protection 24 (2005) 197–206200
out at 10 day intervals (Tsuzuki, 2001; Xuan et al.,2002).It was seen that an additional time of treatment gave
an increase of about 10–15% in weed reduction(Table 3). However, a greater amount of plant materialswas needed, which in turn requires more fieldwork andbecomes more costly.
2.4. Effect of allelopathic plants on weed species is
selective
A 70–80% weed reduction can be attained throughthe incorporation of 1–2 tonne ha�1 of plants withstrong allelopathic potential. Many common weedspecies (15–18) found in paddy fields were eithercompletely inhibited or significantly reduced (Table 4).Observations from laboratory, greenhouse, and fieldtrials show that the effects of allelopathic materials onweed species are selective. At an applied dose of1–2 tonne ha�1, almost all weed species were completelyreduced, however, some species showed differentialselectivity to applied allelopathic plants. The impacton weed species is shown in Table 4.The application of plant materials can control the
majority of nuisance weeds in paddy fields. Species suchas Echinochloa crus-galli (L) Beauv var. crus-galli,
Monochoria vaginalis (Burm, f) Presl var. plantaginea
(Roxb) Solms-Laub., Eleocharis acicularis (L) Roem etSchult. var. longiseta Sven., Scirpus juncoides Roxb. var.ohwianus T. Koyama, Cyperus difformis L were eithercompletely controlled or markedly inhibited. However,some species such as Rotala indica (Willd.) Koehne var.uliginosa (Miq.) Koehne, Dopatrium junceum (Roxb)Hamilt, Marsilea quadrifolia L, Sphenochlea zeylanica
(Gaertin), Murdannia keisak (Hassk) Hand-Mazz andJussiaea decurrens (DC.) showed a strong resistance toAgeratum conyzoides, Blechnum orientale, Eupatorium
canabium, Euphorbia hirta, Melia azedarach, Morus
alba, rice hull, in which the resistance of Rotala indica
was the strongest. In addition, rice bran stimulatedemergence of Echinochloa crus-galli (Table 4).
Rotala indica also showed strong resistance to somecommon herbicides in Japan, such as a herbicidemixture of benzofenap, bromobutide, butanamide andpyributicarb, (Sankyo Ltd., Japan)] (Xuan et al., 2003a).However, alfalfa and rice by-products could completelyinhibit emergence of this noxious weed at1–2 tonne ha�1, and Ageratum conyzoides, Bidens pilosa,
Galactia pendula, Leucaena glauca, and Piper methysti-
cum all significantly inhibited Rotala indica at this dose(Table 4). The incorporation of these plant species to thepaddy field may successfully control Rotala indica.
ARTICLE IN PRESS
Table 3
Percentage of weed control by allelopathic plants with different times
of treatment
Nos. treatments Plant materials References
Alfalfa+ Buckwheat++
1 time 70.0a 51.0a +: Xuan and Tsuzuki (2001)
2 times 84.0b 74.0b ++: Tsuzuki (2001)
3 times 95.5c —
Column with the same letter is not significantly different (P ¼ 0:05).—: treatment was not conducted.
-40
-20
0
20
40
60
80
100
120
1 3 5 8 10 15 20 25 30 40 50
Inih
bitio
n (%
)
Alfalfa
Kava
Time after application (d)
LSD (0.05) = 13.2
-20
0
20
40
60
80
100
120
Time after application (d)
Inhi
bitio
n (%
)
AlfalfaKava
LSD (0.05)=11.9
1 3 5 8 10 15 20 25 30 40 50
(a)
(b)
Fig 1. Effects of soil incorporated with Medicago sativa (alfalfa cv.
Rasen) and Piper methysticum (kava) on germination of barnyardgrass
(A), and monochoria (B). LSD value is presented in tabular format
within each panel and expresses significant difference among treated
time and type of incorporated materials. Applied dose: 2 tonne ha�1
(Xuan, 2004).
T.D. Xuan et al. / Crop Protection 24 (2005) 197–206 201
Moreover, in these allelopathic plants, one or severalallelochemicals which are active on the suppression ofRotala indica may exist. When these allechemicals areidentified, the suppressive function of common herbi-cides on Rotala indica may be improved.
2.5. Varietal effects on weed control
Plant allelopathy has been shown to be varietydependent (Chung et al., 2001, 2002; Olofsdotter,2001; Xuan and Tsuzuki, 2002; Tsuzuki et al., 1975;
Grodzinsky and Panchuck, 1974; Putnam and Duke,1974). A number of plants have their allelopathicactivity varying between cultivars, such as Hordeum
vulgare L. (barley), Fagopyrum spp. (buckwheat) (Tsu-zuki et al., 1975), Cucumis sativus L.(cucumber)(Putnam and Duke, 1974), O. sativa L.(rice) (Chung etal., 2001, 2002), and Triticum aestivum L. (wheat)(Grodzinsky and Panchuck, 1974). Alfalfa has beenknown to cause autotoxicity by continuous cropping(Webster et al., 1967; Miller, 1983) and was documentedto possess strong allelopathic potential (Nakahisa et al.,1993, 1994) and reduced emergence of some lowlandweeds (Tsuzuki et al., 1999). We have assessed allelo-pathic potential of common alfalfa cultivars grown inJapan and shown that there was a great variation ofallelopathic activity (Xuan and Tsuzuki, 2002). Twocultivars: Rasen and Yuba, showed the greatest allelo-pathic potential and Batasu the least (Xuan andTsuzuki, 2002). The inhibitory magnitude of Rasenwas the same as Yuba in laboratory trials (Xuan andTsuzuki, 2002). However, in paddy fields, Rasen had anincrease of 15% weed reduction and 50% greater riceyield than Yuba (Xuan et al., 2003a, Table 2). Thequantity of phenolic compounds in Rasen was greaterthan those of Yuba, and both Rasen and Yuba hadmore phenolics, which showed greater allelopathicaction than Batasu. Observations showed that inhibitionof alfalfa cultivars was proportional to the number andquantity of growth inhibitors, which showed strongallelopathic activity. We have suggested that theallelopathic effect in the plants is varietal dependent,and might have a genetic basis. If the varietal differenceof an allelopathic plant is clarified, the cultivars whichshow the greatest allelopathic potential might be anuseful source for weed control, such as two alfalfacultivars: Rasen and Yuba (Xuan and Tsuzuki, 2002),and perennial buckwheat (Tsuzuki et al., 1975). Theexistence of genes, which determine or correlate to theallelopathic characteristic of plant, should be described.Weed suppression of a plant might be proportional to itsallelopathic magnitude.
3. Effects on plant fungi
Some chosen plant species are both aromatic andtraditionally used by local farmers to reduce pests andplant diseases. Their effects on pathogen and weeds arepresented in Table 5.We examined the impacts of Piper methysticum
L.(kava) root extracts on the growth of five fungalspecies including rhizopus root (Rhizopus stolonifer
MAFF305786), pearl leaf curl (Taphrina deformans
MAFF305614), rice blast (Pyricularia grisea
MAFF101002), rice shealth blight (Thanatephrorus
cucumeris MAFF305844), and papaya dry root

ARTICLE IN PRESS
Table 4
Effects of allelopathic plants at 1–2 tonne ha�1 applied dose on emergence of common paddy weeds
Plant materials Weed species References
A B C D E F G H I J K L M N O P Q R
Ageratum conyzoides L. � * + � � � � � + ! * + + * ! ! * + Hong et al. (2004)
Azadirachta indica A. Juss + * � � � � � � � � � � � � � � � � Xuan et al. (2004a, b)
Bidens pilosa L. � + + � � � � � + + + + * * + * + + Hong et al. (2004)
Blechnum orientale L. � * ! � � � � � + NS + + NS * ! ! + + Hong et al. (2004)
Eupatorium canabium L. � NS ! � � � � � + NS + + ! * * + ! * Hong et al. (2004)
Euphorbia hirta L. � * ! � � � � � + ! NS + + * + + + + Hong et al. (2004)
G. pendula Pers � + + � � � � � + + + + + * ! + + + Hong et al. (2004)
Fagopyrum esculentum Moench
Pellets: + + � + + * � � � � � * � � � � � � Tsuzuki (2001)
L. glauca L. � * + � � � � � + NS + + * * + + + + Hong et al. (2004)
Melia azedarach L. � + ! � � � � � * + * + + * + ! * + Hong et al. (2004)
Medicago sativa L.
Pellets + + * * + * � * � � � + � � � � � � Xuan and Tsuzuki (2001);
Xuan et al. (2002)
cv. Rasen * * * + � + + � � � � � � � � � � � Xuan et al. (2003a)
cv. Yuba * * + + � + + � � � � � � � � � � � Xuan et al. (2003a)
Morus alba L. � + ! � � � � � + ! + + + * ! + + + Hong et al. (2004)
O. sativa L.
Hull * * ! + � ! * � � � � � � � � � � � Xuan et al. (2003a)
Bran ! * + ! � * NS � � � � � � � � � � � Xuan et al. (2003a)
Hull+Rasen * * * NS � * * � � � � � � � � � � � Xuan et al. (2003a)
Bran+Yuba * * + NS � + * � � � � � � � � � � � Xuan et al. (2003a)
Piper methysticum L. + + + + � � � � � � � + � � � � � � Xuan et al. (2003b)
Tephrosia candila L. � + + � � � � � + + * + + * * NS + + Hong et al. (2004)
A: Echinochloa crus-galli; B: Monochoria vaginalis; C: Rotala indica; D: Eleocharis acicularis; E: Scirpus juncoides; F: Doparium juncencum; G:
Lindernia pyxidaria (L.); H: Elatine triandrae Schk var. pedicellate Krylov; I: Marsilea quadrifolia; J: Fimbristylis miliacea (L) Vahl; K: Leptochloa
chinensis (L); L: Cyperus difformis; M: Sphenochlea zeylanica Gaertin; N: Commelina diffusa Burm; O: Murdannia keisak; P: J. decurrens; Q:
Dactyloctenium aegyptium (L) Beauv.; R: Brachiaria mutica.
*: completely inhibited; +: significantly inhibited;�: Measurement was not conducted.; !: promoted ; NS: not significant or no effect.
Table 5
Effects of applied plants on pests and plant diseases
Plant species Effects on pests and plant diseases Reference
Ageratum conyzoides L. Inhibited growth and development of Staphylococus aureus, Bacillus subtilis,
Escherichia coli, Pseudomonas aeruginosa.
Almagboul et al. (1985)
Reduced larvae emergence of Meloidogyne incognita Shabana et al. (1991)
Inhibited predators of spidermite Panonychus citri, Pu et al. (1990);
Phyllocoptruta oleivora, Brevipalpus phoenicis Liang et al. (1994);
Gravena et al. (1993)
Azadirachta indica A.
Juss
Control domestic insects, stored grains pests, crop pests Sankaram (1987)
10% neem oil inhibited growth of Aspergilus nigher, Fusarium moniliforma,
Macrophormina phaseolina, Drechlea rostrata in vitro Tungo baciliform, spherical
virus
Sankaram (1987)
Fagopyrum esculentum Reduce wireworm larvae and wireworm population Valenzuela and Smith
(2002)
Melia azedarach L. Infection by pathogen such as Pyricularia grisea and Thanatephorus cucumeris is
reduced if applied with Melia azedarach into rice field
Hieu (2003)
T.D. Xuan et al. / Crop Protection 24 (2005) 197–206202
(Fusarium solani MAFF306358). The inhibition on thesenuisance plant fungi is shown in Table 6.Kava root had a strong allelopathic potential and
exhibited a great suppression on paddy weed emergence
(Tables 2 and 4). However, the extract of kava root alsosignificantly inhibited the five noxious fungi studied.Rhizopus stolonifer was the most inhibited, andother species were also strongly suppressed by kava.

ARTICLE IN PRESS
Table 6
Inhibitory effects on fungi treated with kava root extracts (as percent compared to the respective control)
Dose (g l�1) Fungal species
F. solani Pyricularia grisea R. stolonifer T. deformans Thanatephorus cucumeris
25.0 25.8a 28.6a 44.0a 30.5a 26.1a
50.0 39.9b 42.8b 85.5b 41.7a 36.6b
100.0 43.3b 64.4c 100.0c 66.9b 56.7c
200.0 61.2c 85.5d 100.0c 71.6b 87.7d
LSD (0.05) 11.1 7.8 7.3 11.5 8.6
Source: Xuan et al. (2003b).
T.D. Xuan et al. / Crop Protection 24 (2005) 197–206 203
Application of kava root at 200 g l�1 concentration gave70–100% control of these plant fungi (Table 5). Themagnitude of kava inhibition on individual fungiappeared to be species dependent and proportional tothe applied dose.Allelochemicals isolated from Alpinia speciosa K.
SCHUM, and Ananas comosus var. Cayenne (pineapple) were examined for their biological impact onfungi. Dihydro-5,6-dehydrokawain isolated from Alpi-
nia speciosa and its derivative dimethyl phosphorothio-nate exhibited the greatest antifungal activitity of 91%and 72% against Corticium rolfsii and Pythium spp.,respectively (Tawata et al., 1996a). Cinamic acid, p-coumaric acid, and ferulic acid were isolated fromAnanas comosus and twenty-four kinds of esters weremade from these acids, alcohols, and the components ofAlpinia (Tawata et al., 1996b). Among these com-pounds, isopropyl 4-hydroxycinnamate and butyl 4-hydroxy-cinnamate were observed to have similarantifungal activity on Pythium spp. at 10 ppm.The incorporation of plants with strong allelopathic
potential may give beneficial effects on both weeds andfungi. Our research examining the extracts of plant withstrong allelopathic properties on major pests and plantfungi is continuing.
4. Allelochemicals
The chemical exudates from allelopathic plants areproposed to play a major role in the allelopathy mode ofaction. Evidence showed that higher plants release adiversity of allelochemicals into the environment, whichincludes phenolics, alkaloids, long-chain fatty acids,terpenoids, and flavonoids (Rice, 1984; Chou, 1995).However, these compounds are available in lowconcentrations in most allelopathic plants. The inhibi-tion of the emergence of paddy weeds is the interactionof many allelochemicals, not a single compound. Whena single compound responsible for the allelopathicpotential of the allelopathic plants is isolated andidentified, it will be a breakthrough to develop bio-
herbicides. In fact, several compounds extracted fromhigher plants viz., cineole (Shell), benzoxazinones(BASF), quinolinic acid (BASF) and leptospermones(Zeneca), have been used in agriculture with promisingresults (Kohli et al., 1998). We have attempted to isolateand identify growth inhibitors in plants with strongallelopathic potential by the uses of high-pressure liquidchromatography (HPLC), gas chromatography massspectrometry (GC-MS), and nuclear magnetic resonance(NMR). Our identified allelochemicals are listed inTable 7.In bioassays, these identified compounds showed
strong suppression on indicator plants such as lettuce,radish, rice seedlings, and some paddy weeds. However,the inhibition varies among allelochemicals. Arachidicacid, behenic acid, palmitic acid, and stearic acidshowed significant reduction on emergence of riceseedlings at 250 ppm, whilst caffeic acid and ferulic acidsignificantly inhibited rice seedlings at 10 ppm (Xuanand Tsuzuki, 2004).These identified compounds are putative allelochem-
icals and commonly found in allelopathic plants in lowconcentration (Xuan and Tsuzuki, 2004) and shown tohave interactive impacts (Xuan et al., 2003c). In kava, wefound that there might be some water-soluble substances,which show marked inhibition on lettuce growth 30 daysafter incorporation in soil (Xuan, 2004). This compoundhas not been identified but was observed to be present ingreat quantity in kava root, and it may have a major rolein the allelopathy activity (Xuan et al., 2003d). We arecarrying out the isolation and identification of allelo-chemicals from other allelopathic plants shown in thisnote (Tables 1 and 2) and concentrating on novel growthinhibitors which can be exploited for the development ofbio-herbicides and pesticides.
5. Suggestions and future prospects
Amendment of plant materials with strong allelo-pathic properties brings important agronomic impacts:(i) a 70% reduction in weed biomass and a 20% increase

ARTICLE IN PRESS
Table 7
Allelochemicals identified in applied plant species
Plant species and common
name
Plant parts Identified allelochemicals References
Ageratum conyzoides (billy
goat weed)
Leaves, stem and root Benzoic acid, coumalic acid, gallic acid,
protocatechuic acid, p-hydroxybenzoic acid, p-
coumaric acid, sinapic acid
Xuan et al. (2004a)
Azadirachta indica (neem) Leaves and bark Benzoic acid, gallic acid, ferulic acid, p-coumaric
acid, p-hydroxybenzoic acid, vanillic acid, trans-
cinamic acid
Xuan (2004)
Fagopyrum spp.
(buckwheat)
Leaves and stem Arachidic acid, behenic acid, caffeic acid,
chrologenic acid, ferulic acid, palmitic acid,
stearic acid
Tsuzuki et al. (1987);
Tsuzuki and Yamamoto
(1987)
Medicago sativa L. (alfalfa) Leaves and stem Catechin, gallic acid, ferulic acid, protocatechuic
acid, p-coumaric acid, p-hydroxybenzoic acid,
syringic acid, sinapic acid, vanillin, vanillic acid
Xuan et al. (2003d)
Leaves and stem, and in
decomposed residue in soil
Ferulic acid, salicylic acid Nakahisa et al. (1994)
Piper methysticum L.
(kava)
Root Gallic acid, ferulic acid, protocatechuic acid, p-
hydroxybenzoic acid, p-coumaric acid, salicylic
acid, trans-o-coumaric acid, trans-cinamic acid
Xuan et al. (2003d)
T.D. Xuan et al. / Crop Protection 24 (2005) 197–206204
in rice yield, (ii) soil improvement: additional nutrientsfrom plant materials to soil may minimize the quantityof synthetic fertilizers needed for rice or crop growthand development, (iii) the application of allelopathicplants does not provide the same level of weed control assynthetic herbicides, but the remaining of 30% weedsleft after application of plant materials can be controlledsuccessfully by a lower dose of herbicides and by riceshading. This leads to a reduction of applied herbicides.Hence, the soil environment might be kept from possibledetrimental effects caused by synthetic agrochemicals,but this point needs further evaluation. In addition, thequality of crops between incorporation of allelopathicplants and agrochemicals also requires detailed analysis,(iv) the infestation of pests and pathogens might bereduced simultaneously with the reduction of weeds. Forinstance, Ageratum conyzoides L.(billy goat weed),Fagopyrum esculentum Moench (buckwheat), Melia
azedarach L.(Chinaberry), Piper methysticum L.(kava),and Azadirachta indica A. Juss (neem) gave growthreduction of pests and plant diseases (Tables 5 and 6).Future determination of whether allelochemicals or theother compounds available in the allelopathic plants areresponsible for pest and pathogenic suppression isneeded.Heavy fieldwork is required to apply an amount of
1–2 tonne ha�1 of plant materials to the fields, and thistask does not meet future agriculture practice, as thelabour force in the agricultural sector is declining. Theproduction of plant materials incorporated or formu-lated as ‘pellets’ may help with easier application. Whenallelochemicals with strong action on weed suppression
are synthesized for the development of bio-herbicides,the burden from plant residue application can beminimized.Another shortcoming of the incorporation of allelo-
pathic plant material is that water is necessary for theirdecomposition, therefore the control of weed in uplandrice faces more difficulties because of water availability.For the paddy field, the plant materials should beapplied to the fields within 5 days after saturating withwater, to achieve the greatest amount of weed reduction(Xuan et al., 2004a). However, for upland fields, theplant materials should be incorporated into soil beforethe cropping starts, to kill weeds during their germina-tion. Otherwise, a great amount of water is needed forthe decomposition of plant materials.Another challenge is that the suppression of plant
materials on weed species is selective. A plant speciesexhibits strong allelopathic properties but could notinhibit every weed species, and the impact of allelopathicplants on weeds is species dependent (Table 4). Hence,the combination of different plant materials appliedsimultaneously to the fields, which may suppress mostmajor weeds, might successfully overcome this problem.The recent trend of allelopathic researchers is to
determine and locate genes correlated with allelopathy.In fact, the allelopathic characteristic of plants [such asMedicago sativa L. (alfalfa) (Xuan and Tsuzuki, 2002),H. vulgare L. (barley), Fagopyrum spp. (buckwheat)(Tsuzuki et al., 1975), cucumber (Cucumis sativus L.)(Putnam and Duke, 1974), O. sativa L. (rice) (Chung etal., 2001, 2002), and Triticum aestivum L. (wheat)(Grodzinsky and Panchuck, 1974)] is documented to

ARTICLE IN PRESST.D. Xuan et al. / Crop Protection 24 (2005) 197–206 205
be varietal dependent and suggested to be geneticallybased. When allelopathic gene(s) in the plants arelocated, then plant breeding techniques might help toenhance the allelopathic potential of the target crops.
Acknowledgements
The authors wish to acknowledge the financialsupport of the Rural Development Administrationmade to the BioGreen 21 in 2003. Thanks are also dueto the research fund provided by Japan Showa ShellSekiyu (2003–2004).
References
Almagboul, A.Z., Farrog, A.A., Tyagi, B.R., 1985. Antimicrobial
activity of certain Sudanese plants used in folkloric medicine:
screening for antibacterial activity. Fitoterapia 56, 103–109.
Anaya, A.L., Calera, M.R., Mata, R., Miranda, R.P., 1990.
Allelopathic potential of compounds isolated from Ipomoea tricolor
Cav. (Convolvulaceae). J. Chem. Ecol. 16, 2145–2152.
Chou, C.H., 1995. Allelopathy and sustainable agriculture. In: Inderjit,
Dakshini, K.M.M., Einhellig, F.A. (Eds.), Allelopathy: Organisms,
Process and Applications. ACS Symposium Series 582. American
Chemical Society, Washington, DC, pp. 211–233.
Chung, I.M., Ahn, J.K., Jun, S.J., 2001. Assessment of allelopathic
potential of barnyardgrass (Echinochloa crus-galli) on rice (Oryza
sativa L.) cultivars. Crop Prot. 20, 921–928.
Chung, I.M., Kim, K.H., Ahn, J.K., Chun, S.C., Kim, C.S., Kim, J.T.,
Kim, S.H., 2002. Screening of allelochemicals on barnyardgrass
(Echinochloa crus-galli) and identification of potentially allelo-
pathic compounds from rice (Oryza sativa) variety hull extracts.
Crop Prot. 21, 913–920.
Gravena, S., Coletti, A., Yamamoto, P.T., 1993. Influence of green
cover with Ageratum conyzoides and Eupatorium pauciflorum on
predatory and phytophagous mites in citrus. Bul. OILB-SROP 16,
104–114.
Grodzinsky, M.M., Panchuck, M.A., 1974. Allelopathic properties of
crop residues of wheat–wheat grass hybrids. In: Grodzinsky, A.M.
(Ed.), Physiological–Biochemical Basis of Plant Interactions in
Phytocenosis. Naukova Dumka, Kiev, pp. 51–55 (in Russian with
English summary).
Hieu, H.H., 2003. Screening for allelopathic potential of higher plants
in Southeast Asia. Graduation Thesis, Thai Nguyen University,
Vietnam, 64pp.
Hong, N.H., Xuan, T.D., Tsuzuki, E., Terao, H., Matsuo, M., Khanh,
T.D., 2003. Screening for allelopathic potential of higher plants
from Southeast Asia. Crop Prot. 22 (6), 829–836 [International
Association for the Plant Protection Sciences (IAPPS)].
Hong, N.H., Xuan, T.D., Tsuzuki, E., Khanh, T.D., 2004. Paddy weed
control by higher plants from Southeast Asia. Crop Prot. 23,
255–261.
Inderjit, 2001. Soil: environment effects on allelochemical activity.
Agron. J. 93, 79–84.
Kohli, R.K., Batish, D., Singh, H.P., 1998. Allelopathy and its
implications in agroecosystems. J. Crop Prod. 1, 169–202.
Liang, W.G., Hui, W., Lee, W.K., 1994. Influence of citrus orchard
ground cover plants on arthropod communities in China: a review.
Agric. Ecosyst. Environ. 50, 29–37.
Miller, D.A., 1983. Allelopathic effects of alfalfa. J. Chem. Ecol. 9,
1059–1072.
Nakahisa, K., Tsuzuki, E., Mitsumizo, T., 1993. Study on the
allelopathy of alfalfa (Medicago sativa L.). 1. Observation of
allelopathy and survey for substances inducing growth inhibition.
Jpn. J. Crop Sci. 62, 294–299.
Nakahisa, K., Tsuzuki, E., Terao, H., Kosemura, S., 1994. Study on
the allelopathy of alfalfa (Medicago sativa L.). 2. Isolation and
identification of allelopathic substance in alfalfa. Jpn. J. Crop Sci.
63, 278–284.
Narwal, S.S., 1994. Allelopathy in Crop Production. Scientific
Publisher, Jodhpur, India 288pp.
Olofsdotter, M., 2001. Rice—a step toward use of allelopathy. Agron.
J. 93, 3–8.
Pu, T.S., Liao, K.Y., Chang, T., 1990. Investigations on predations
mite resources in citrus orchards in Guang Xi and their utilization.
Acta Phytophyarica Sin. 17, 355–358.
Putnam, A.R., Duke, W.B., 1974. Biological suppression of weeds:
evidence for allelopathy in accession of cucumber. Science 186,
370–372.
Rice, E.L., 1984. Allelopathy, Second ed. Academic Press Inc.,
Orlando, FL, p. 422.
Sankaram, A.V.B., 1987. Chemistry, biological activity and utilization
aspects of some promising neem extractives. In: Proceedings of the
Third International Neem Conference, Nairobi, Kenya, July 10,
1986, pp. 127–148.
Shabana, N., Husain, S.I., Nisar, S., 1991. Allelopathic effects of some
plants on the larval emergence of Meloidogyne incognita. J. Indian
Appl. Pure Biol. 5, 129–130.
Tawata, S., Taira, S., Kobamoto, S., Ishihara, M., Toyama, S.,
1996a. Syntheses and biological activities of dihydro-5,6-
dehydrokawain derivatives. Biosci. Biotechnol. Biochem. 60,
1643–1645.
Tawata, S., Taira, S., Kobamoto, N., Zhu, J., Ishihara, M., Toyama,
S., 1996b. Synthesis and antifungal activity of cinamic acid esters.
Biosci. Biotechnol. Biochem. 60, 909–910.
Tsuzuki, E., 2001. Application of buckwheat as a weed control. Agric.
Hortic. 76, 55–62 (in Japanese).
Tsuzuki, E., Yamamoto, Y., 1987. Isolation and identification of
phenolic substances from wild perennial buckwheat (F. cymosum
M.). Bulletin Faculty of Agriculture, Miyazaki University. 34, pp.
289–295.
Tsuzuki, E., Katsuki, A., Shida, S., Danjo, T., 1975. On the growth
inhibitors contained in buckwheat plants. I. The effects of water
soluble substances on germination of some crops and growth of
rice seedling. Report Kyushu Branch of Crop Sci. Soc. Japan. 42,
pp. 83–84.
Tsuzuki, E., Yamamoto, Y., Shimizu, T., 1987. Fatty acids
in buckwheat are growth inhibitor. Ann. Bot. (London) 60,
69–70.
Tsuzuki, E., Miura, M., Sakaki, N., Yoshino, T., 1999. Study on the
control of weeds by using higher plants. Bull. Kyushu Branch Crop
Sci. Soc. Japan 65, 39–40 (in Japanese).
Xuan, T.D., 2004. Utilization of allelopathy in higher plants
for control of paddy weeds. Ph.D. Thesis, School of
Agricultural Sciences, Kagoshima University, Kagoshima,
Japan, 167pp.
Xuan, T.D., Tsuzuki, E., 2001. Effects of application of alfalfa
pellet on germination and growth of weeds. J. Crop Prod. 4,
303–312.
Xuan, T.D., Tsuzuki, E., 2002. Varietal difference in allelopathic
potential of alfalfa (Medicago sativa L.). J. Agron. Crop Sci. 188,
2–7.
Xuan, T.D., Tsuzuki, E., 2004. Allelopathic plants: Buckwheat.
Allelopathy J. 13, 137–148.
Xuan, T.D., Tsuzuki, E., Uematsu, H., Terao, H., 2002. Effects of
alfalfa (Medicago sativa L.) on weed control in rice. Allelopathy J.
9, 195–203.

ARTICLE IN PRESST.D. Xuan et al. / Crop Protection 24 (2005) 197–206206
Xuan, T.D., Tsuzuki, E., Terao, H., Matsuo, M., Khanh, T.D., 2003a.
Alfalfa, rice by-products, and their incorporation for weed control
in rice. Weed Biol. Manage. 3, 137–144.
Xuan, T.D., Chikara, J., Ogushi, Y., Tsuzuki, E., Terao, H., Khanh,
T.D., Matsuo, M., 2003b. Application of kava (Piper methysticum
L.) root as potential herbicide and fungicide. Crop Prot. 22,
873–881.
Xuan, T.D., Tsuzuki, E., Matsuo, M., Khanh, T.D., 2003c. Correla-
tion between inhibitory exhibition and suspected allelochemicals in
alfalfa (Medicago sativa L.). Plant Prod. Sci. 6, 165–171.
Xuan, T.D., Tsuzuki, E., Terao, H., Matsuo, M., Khanh, T.D., 2003d.
Identification of potential allelochemicals in kava (Piper methys-
ticum. L) root. Allelopathy J. 12, 197–204.
Xuan, T.D., Tsuzuki, E., Tawata, S., Khanh, T.D., 2004a. Methods to
determine allelopathic potential of crop plants for weed control.
Allelopathy J. 13, 149–164.
Xuan, T.D., Tawata, S., Hong, N.H., Khanh, T.D., Chung, I.M.,
2004b. Assessment of phytotoxic action of Ageratum conyzoides L.
(billy goat weed) on weeds. Crop Prot. 23, 335–345.
Valenzuela, H., Smith, J., 2002. Sustainable agriculture green
manure crops: Buckwheat. Cooperative extension service.
College of Tropical Agriculture and Human Resources. Hawaii
University, p. 4.
Webster, G.R., Khan, S.U., Moore, A.W., 1967. Poor growth of
alfalfa (Medicago sativa L.) on some Alberta soils. Agron. J. 59,
37–42.
Related Documents











