1 NOTICE: This is the author’s version of a work that was accepted for publication in CSIRO Publishing title: Biological Control of Weeds in Australia, eds Mic Julian, Rachel McFadyen, Jim Cullen. Any changes resulting from the publishing process, such as peer review, editing, corrections or structural formatting may not be reflected in this document. Changes may have been made to this work since it was submitted for publication. Link to Publisher’s version: <http://www.publish.csiro.au/pid/6509.htm>

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
NOTICE: This is the author’s version of a work that was accepted for publication in
CSIRO Publishing title: Biological Control of Weeds in Australia, eds Mic Julian, Rachel
McFadyen, Jim Cullen. Any changes resulting from the publishing process, such as peer
review, editing, corrections or structural formatting may not be reflected in this
document. Changes may have been made to this work since it was submitted for
publication. Link to Publisher’s version: <http://www.publish.csiro.au/pid/6509.htm>

2
Word count: 5,209
Heliotropium europaeum L. – common heliotrope
Andy W. Sheppard, Louise Morin and Jim Cullen
CSIRO Ecosystem Services, GPO Box 1700, Canberra, ACT 2601, Australia
ABSTRACT
Common heliotrope is a summer growing annual weed of crops and pasture, considered
native to Mediterranean Europe, Turkey and western North Africa, that is present in all states
and territories of Australia except Tasmania. It can be an important component of annual
pastures in southern Australia, where it often dominates fallow, disturbed or ploughed land
from December to April. When eaten by livestock it causes cumulative liver damage and
causes the highest fatality rate from primary pyrrolizidine alkaloid poisoning of any plant.
The biological control program targeting common heliotrope started in the 1950’s, with a
peak of activities from the early 1970’s to the early 1990’s, and consisted of natural enemy
surveys of more than 15 annual Heliotropium spp. in the western Mediterranean, eastern
Africa and Iran (the centre of origin of the genus). Nearly 40 arthropods and pathogens were
identified from common heliotrope out of the 132 species found across Heliotropium spp.
Host-specificity testing in Europe was undertaken for three arthropod candidate agents; the
root and leaf feeding flea beetle, Longitarsus albineus, the root weevil, Pachycerus segnis
and the flower cyme feeding moth Ethmia distigmatella, and two plant pathogens; the rust
Uromyces heliotropii and the leaf-blotch fungus, Cercospora heliotropiicola. Thousands of
flea beetles were released in Australia from 1979 until the late 1980’s following the
development of effective laboratory mass-rearing techniques, but the insects never persisted
more than a few years at a site and this failure was never adequately explained. The rust
fungus was released between 1991 and 1993, widely established and is still occasionally
found, but has had no impact on the abundance of the weed. None of the other candidate
agents were released. The inability of released agents to reduce weed populations was put
down to the difficulty of achieving biological control of an ephemeral summer annual
agricultural weed with highly variable between-year dynamics.

3
Key words: Boraginaceae, Longitarsus albineus, Pachycerus cordiger, Ethmia
distigmatella, Uromyces heliotropii, Cercospora heliotropiicola, Cercospora taurica.
INTRODUCTION
Heliotropium europaeum L. (Boraginaceae), common heliotrope, is a summer annual herb
considered native to the Mediterranean region and south-west Asia. It is a weed in North
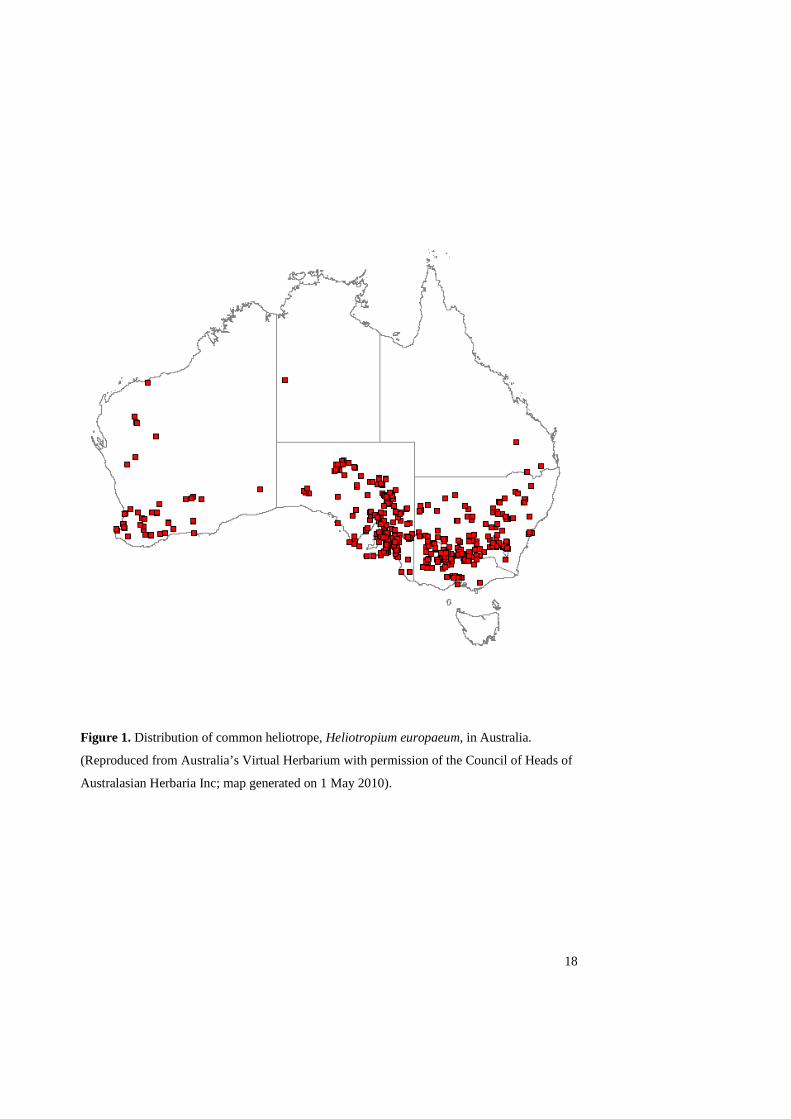
America (Parsons and Cuthbertson 2001), but its impacts are far worse in Australia. Its
distribution in Australia is given in Figure 1. It is most abundant north and west of the Great
Dividing Range in the south east of the continent, where it occurs in a broad arc from
southern Queensland, through NSW and northern Victoria to SA. It is also abundant in the
lower south west of Western Australia (Parsons and Cuthbertson 2001). This corresponds to a
zone of between 300-500mm annual rainfall (Moore 1956). In southern Australia it
germinates from spring to early summer on bare soil with soil moisture once soil
temperatures reach 24°C and grows from November to May, with abundance in any given
year driven by the seasonal distribution of summer rain and the degree of litter accumulation,
which provide favourably moist conditions for plant growth (Moore 1956, Hunt et al., 2008).
It is a very ephemeral summer annual that by its germination requirements rarely occurs in
the same place twice in consecutive years in the same season. It often dominates fallow,
disturbed or ploughed land and can be an important component of annual pastures. When
eaten by stock it causes cumulative liver damage and leads to the highest fatality rate from
primary pyrrolizidine alkaloid poisoning of any plant. Common heliotrope is six times more
toxic than Paterson’s curse, which also contains pyrrolizidine alkaloids (O'Dowd and Edgar
1989). Historically pyrrolizidine alkaloidisis accounts for more than 63% of sheep losses in
some years (Edgar et al. 1941), although the situation more recently is unclear. Outbreaks of
common heliotrope do not occur every year, but in years of high abundance (e.g., 1991-93)
farmers generally face economic losses due to the impact of the weed and costs of
implementing largely ineffective chemical or cultural control (Parsons and Cuthbertson
2001). It has been estimated that on average in the 1980s, common heliotrope cost the meat
and wool industry at least $46 million pa ($17.5 m from livestock mortality, $14 m from
decreased productivity, and $25 m in control costs) (Cullen and Delfosse 1990). In a

4
cropping context it is considered to use up valuable soil moisture over summer prior to
autumn/winter crop sowing (Delfosse and Cullen 1980).
Common heliotrope is a hairy, ephemeral summer-annual herb usually 10-30 cm high with a
well-developed tap root system and a somewhat offensive odour. It reproduces only by seed.
The grey-green erect or semi-prostrate shoot has alternate elliptical to ovate leaves. Flowers
are small, white with a yellow throat, in two rows on the flowering coiled cymes, that
straighten as seeds ripen. The fruit has four nutlets with brown to black small subglobular
seeds (Parsons and Cuthbertson 2001) (Figure 2). Plants produce flowers and seeds
concurrently with vegetative growth, soon after germination, thereby creating a persistent
seedbank of up to 3,000,000 seeds m-2. They senesce in autumn if they are not previously
killed by drought or rain-induced leaf-blight outbreaks over the summer.
In a comparative study of common heliotrope in both its native and exotic ranges, Sheppard
et al. (1996) found maximum plant densities and basic demography (e.g., seed production
versus density) to be similar in both regions although the average infestation size and density
were greater and their occurrences more frequent in Australia. The higher summer rainfall
and more extensive rotational agriculture in Australia appeared to be the dominant causes of
the plant’s abundance status. These conditions however, could not fully explain why the
seedbanks in Australia were ten times larger than in the native range.
Control options for common heliotrope are well documented in Parsons and Cuthbertson
(2001) and include cultivation, herbicides and crash grazing by sheep, but each of these
options has significant economic, practical and animal health constraints. The persistent
seedbank is the other major hindrance in rotational cropping systems. Classical biological
control was identified very early, in the 1950’s. When major activity in this biological
control program commenced there was no regulatory requirement to either nominate a weed
as a biological control target, or consider risks that potential arthropod agents might pose to
native Australian plants, though this was already an issue for plant pathogens.
BIOLOGICAL CONTROL HISTORY
Australia is the only country to have attempted a biological control program against common
heliotrope. The program started with survey work by Frank Wilson from about 1950 in

5
Europe and North Africa, but was deemed "not especially promising" (Delfosse and Cullen
1980) and was shelved until 1971 when exploration was renewed in Europe, the Middle East
and North Africa. The early surveys and the work from 1971 to the late 1980s was funded by
CSIRO and then jointly by CSIRO and the Meat Research Corporation from 1988 when the
program received encouragement from producers, politicians and state agencies with across
jurisdictional support through the Australian Weeds Committee. Adding biological control to
existing management practices was considered the quickest way towards a sustainable control
strategy. While the early focus of the program was almost entirely on arthropod agents, one
of which was eventually released, the success of the skeleton weed rust in Australia prompted
consideration of potential pathogen agents in the 1980’s (Hasan 1985; 1992).
PLANT TAXONOMY
Common heliotrope is a member of a complex of closely-related Heliotropium species in the
eastern part of its native range in Eurasia, which has caused confusion on whether certain
natural enemies collected during early surveys had indeed been found on the target. Only
common heliotrope, H. europaeum, is considered to be present in Australia, but there are
several Australian native Heliotropium spp.
EXPLORATION
Frank Wilson's initial exploration included southern France, eastern Spain, coastal Algeria
and Tunisia. He then conducted surveys in Kenya, Eritrea and the Sudan. Eventually he
concluded that the prospects for biological control of common heliotrope were low (Delfosse
and Cullen 1980). There was no further work until 1971 when increasing pressure in
Australia to find some solution to the problem, plus the presence of a long-term base in
Europe encouraged renewed exploration. This base allowed more extensive exploration to the
east including the Middle East known to have a high diversity of species in the genus.
Surveys were subsequently conducted in Iran in 1971 and 1974 and in Tunisia and Algeria in
1974 mainly by John Huber. Iran was chosen as a primary site because it has a large number
of annual Heliotropium spp., some of which are similar to H. europaeum, and because of the
climatic similarity to areas of Australia where H. europaeum is a problem. These surveys
yielded a greater diversity of insects on Heliotropium spp. in northern Iran than in

6
Mediterranean Europe or North Africa, though the most promising agent, the flea beetle
Longitarsus albineus (Foudras) (Coleoptera: Chrysomelidae) was present everywhere.
Surveys continued through the 1970s in Mediterranean Europe and Turkey with the
promising root feeding weevil Pachycerus segnis Germar (=Pachycerus cordiger Germar
,=Pachycerus scabrosus Brullé) (Coleoptera: Curculionidae) being found in Greece and
Turkey as well as in Iran.
A number of later surveys were made to Eastern Europe from CSIRO’s European Laboratory
in the 1980’s and 1990’s, either to collect live material or to try to understand host species
usage by agents under consideration.
CANDIDATES
Natural enemies
The surveys of Heliotropium spp. from 1950 until 1980 in the native range found 132 species
of natural enemy 37, of which were considered to have been found on common heliotrope
(list in Delfosse and Cullen 1980). Table 1 lists those species considered to have biological
control potential. Diversity of natural enemies on common heliotrope was higher towards the
eastern end of its distribution and closer to the presumed evolutionary centre of origin of the
genus. Five species were considered to be potentially specific to at least the genus from the
literature at the time (Delfosse and Cullen 1980). Longitarsus albineus (flea beetle),
Utethesia pulchella (L.) (Lepidoptera: Arctiideae), Cercospora heliotropiicola A.K. Kar &
M. Mandal (Moniliales: Dematiaceae) and the weevil P. segnis are found throughout the
native range of common heliotrope, Ethmia distigmatella Erschoff (Lepidoptera: Ethmiidae)
is only found in the eastern part of the range. Delfosse (1985) discussed strategies for release
stating that the original proposed order of release had been the flea beetle followed by the
weevil then E. distigmatella and finally the rust fungus (U. heliotropii ), but that this
changed to the rust followed by the moth and then the weevil and finally the flea beetle based
on the status of the host specificity testing and availability. As it turned out only the flea
beetle and the rust were ever released. Four species were found on common heliotrope in
Australia prior to the release of exotic agents; a native flea beetle Longitarsus sp. (nr L.
victoriensis Blackburn) (C Reid pers. comm. 1993), a native arctiid moth, Utethesia
pulchelloides (L.), a native gelechiid moth Stomopteryx isosceliscantha (Lower) (Delfosse

7
and Cullen 1980; CSIRO Entomology 1994) and a probably introduced leaf-blotch fungus
Cercospora taurica Tanzschel (J Walker pers. comm. 1991; CSIRO 1994).
Natural enemies tested but not released.
Curculionid weevil; Pachycerus segnis. This large univoltine weevil completes its lifecycle
on species in the Boraginaceae with larvae developing in an earthen cell on the roots of
common heliotrope. The taxonomy has been confusing (Huber and Vayssières 1990) being
considered as an eastern European species P. cordiger with a sister western European species
P. scabrosus by two weevil systematists or as one pan European species later accepted as P.
segnis by others. Huber and Vayssières (1990) found that adults from Eastern Europe
(Turkey and Greece) were larger than museum specimens of either species from Western
Europe and so tested weevil populations collected from common heliotrope in Greece and
Turkey against seven European Boraginaceae and 31 economically important crop species
and. They also found no Pachycerus species commonly on common heliotrope in western
European. They concluded that the tested material was specific to Boraginaceae in the
laboratory and to Heliotropium spp. in the field, and that therefore should be safe for
introduction to Australia. Later concerns that no Australian species in the Boraginaceae had
been included in the original tests led to further testing in Europe. Testing of eastern
European populations over four years in the laboratory in France and in the field in Greece
showed that no Australian species of Heliotropium tested (H. asperrimum R.Br., H.
bacciferum Forsk and H. crispatum F. Muell. Ex Benth.) was suitable for larval
development, but other Australian Boraginaceae; Cynoglossum australe R.Br. and Myosotis
australis R.Br. supported adult feeding, oviposition and larval development (MJ Cullen
unpublished). Whether or not the phenologies of these Australian species might reduce their
exposure to use in the field in Australia by P. segnis was hard to assume and meant the level
of risk to them was difficult to assess and work on this insect was suspended. A further study
discovered a morphologically identical Pachycerus species to that from Eastern Europe
restricted to H. europaeum populations in Western Europe, adding host range support that
there was one pan-European species (Brun et al. 1993), which has since been recognised as
P. segnis (Alonso-Zarazaga 2010). Given these results and the concerns over specificity, no
further application was made to release this species in Australia.
Ethmiid moth; Ethmia distigmatella. This flower-cyme feeding moth recorded from
common heliotrope in Crete, Turkey and south west Asia was only once successfully reared

8
in quarantine in Australia on common heliotrope in small numbers despite several attempts.
An initial host-specificity test carried out using freshly emerged adults found no oviposition
on common heliotrope or the Australian native Heliotropium ovalifolium Forsk. tested.
Using the Flora of Turkey (Davis 1984), a survey of the complex of closely-related species
including H. europaeum, and the insects on this complex in western Turkey in 1991 failed to
confirm whether or not the moth had indeed ever been recorded from common heliotrope in
the native range. Based on this it was considered that common heliotrope is not the preferred
host of this moth (CSIRO Entomology 1994).
Leaf-blotch fungus; Cercospora heliotropiicola. Following rainfall, this plant pathogen can
produce severe disease epidemics on common heliotrope in Europe that can prematurely kill
populations prior to much seed production (Brun et al. 1995; Hasan et al. 1995). However in
the 1980’s another Cercospora species, C. taurica, was observed causing epidemics on
common heliotrope in Australia (J Walker pers. comm. 1991). This species was also known
from eastern Europe, Russia and western Asia (Chupp 1953). The taxonomic status of the
two Cercospora spp. attacking common heliotrope was explored morphologically and using
isozyme electrophoresis (Brun et al. 1996). Results showed consistent differences between
isolates classed as either C. heliotropiicola (from France) or C. taurica (from Turkey and
Australia) used in the study.
Before host-specificity testing of C. heliotropiicola could be justified it was important to see
whether C. heliotropiicola (ex. France) was more pathogenic than C. taurica, which already
occurred in Australia. A comparative demographic study of common heliotrope in Australia
and Europe demonstrated that Cercospora epidemics were both more common and more
damaging in Europe than in Australia, potentially contributing to the observed differences in
the seed bank sizes between the native and exotic range (Brun et al. 1995; Sheppard et al.
1996). Brun et al. (1996) also showed that C. heliotropiicola (ex. France) was more
pathogenic on detached leaves of common heliotrope than C. taurica (ex. Australia) and that
all five Mediterranean Heliotropium species tested were susceptible to both species.
C. heliotropiicola was therefore imported from France into quarantine in Australia in 1996 to
undertake detailed comparative studies with C. taurica. In experiments using whole plants,
both Cercospora species produced a similar number of leaf lesions on plants exposed to the
same temperatures (14, 22, 30°C) and wetness periods (8, 16, 24, 48 h) during the initial

9
infection phase (Morin and Trueman 1997). Under optimum conditions, the pathogens had a
similar impact on plant growth and reduced plant dry weight by 63%. In the first 24 hours,
C. taurica produced more than five times the number of conidia produced by C.
heliotropiicola on infected necrotic tissue placed in high humidity conditions. For both
species, production of conidia increased as the period under high humidity increased,
although C. taurica always produced the highest number of conidia. These results suggested
that under the same (controlled) conditions, C. heliotropiicola did not appear to have greater
value than C. taurica for biological control of common heliotrope and was consequently not
tested further nor released in Australia.
Natural enemies released.
Chrysomelid flea beetle; Longitarsus albineus . This multi-voltine flea beetle feeds on the
exteriors of fine roots as larvae, pupating in the soil and feeding on foliage as adults. In the
field in the native range adult feeding was recorded on four annual Heliotropium species and
typical feeding damage was observed on another 12 species in the genus. A method for
observing and rearing the root-feeding larvae was developed in the late 1970’s and the beetle
was found to have three to four generations a year under field conditions (Huber 1979, 1981).
A parasitic nematode (13% of collections) and a hymenopteran parasitoid were recorded from
this insect in the native range (Huber 1981). Host-specificity testing was carried out against
41 non-target plants using no-choice adult oviposition and larval starvation tests. The test
plant list included only two Australian native species; a Eucalyptus sp. and an Acacia sp.
Adult feeding was observed on 20 of the test plant species, but severity of feeding was less
than a tenth of that seen on common heliotrope. Oviposition was observed across a wide
range of test plants, although the number of eggs laid was a third or less than that found on
common heliotrope. Larval development was recorded on the Heliotropium peruvianum L.
and Myosotis alpestris F.W.Schmidt, but adults emerged only from the former species. At
the time assessing risks to native Australian species was not a requirement for arthropod
agents and therefore Huber (1981) considered that this agent was specific enough for release,
despite the test results suggesting that some Australian native species in the Boraginaceae
were likely to be within its host range.
Common heliotrope seedlings were killed by only a few larvae in tests so significant impact
in the field was predicted. Huber (1981) also concluded that L. albineus was physiologically
ill suited to breeding on non-summer Heliotropium or other Boraginaceae, but that its broad

10
climatic adaptability, its host finding ability and its rapid rate of increase meant that it should
have no difficulty establishing in Australia and that survival of overwintering adults would be
the main determinant of its effectiveness under Australian conditions.
The flea beetle was imported into Australia and first released in 1979. Beetles were
recovered the following season at most sites, but establishment beyond this first season was
not confirmed. A mass-rearing program was successfully developed in Canberra that
produced over 31,000 beetles, which were then released at seven sites across four states
(NSW, SA, Vic and WA) from 1987 to 1989. Measures, such as irrigation, were taken to
ensure common heliotrope presence in subsequent summers. Successful recoveries of flea
beetles were made at four of these sites in lower numbers in the next year and individuals
were recovered at some sites until 1992, but not after. As no flea beetles persisted at any site
longer than three years and it was clear from the search effort that no spread had occurred
monitoring was terminated in 1994. As no further sightings have been made it has been
assumed that this agent failed to establish in Australia. Field experiments in the native range
suggested that unless the post-release densities of flea beetles were very high compared to
those observed in the native range, this agent would have little impact on common heliotrope
abundance, given the already highly variable between year dynamics of this weed (CSIRO
Entomology 1994).
Rust fungus; Uromyces heliotropii. This macrocyclic and autoecious rust was originally
discovered on common heliotrope in Turkey, but was subsequently observed in France, and is
generally found, though only sporadically, across its native range. Field experiments in the
native range suggested that this rust could significantly impact on and kill common heliotrope
seedlings and could survive winter conditions and reinfect plants the following season (Hasan
and Ayres 1990; Hasan and Aracil 1991). Host-specificity tests were carried out on 96 non-
target plant species including eight species of Heliotropium native to Australia and 16 other
native species in the Boraginaceae (Hasan et al. 1992). The Australian native Heliotropium
crispatum F. Muell. Ex Benth. and two exotic Heliotropium spp. present in Australia were
susceptible to the rust, albeit at a much lower level than common heliotrope (Hasan et al.
1992). Any potential threat posed by the rust to H. crispatum was discounted as this species
is both temporally (winter growing) and geographically (northern Australia) separated from
common heliotrope (Hasan and Delfosse 1995).

11
An isolate from Turkey was first released in 1991 at a site in NSW, followed by 19 further
releases at 13 sites across four states (NSW, SA, Vic and WA) over three years, using
sprayed spore suspensions in water, covered by plastic sheeting in the first 24 hours, at each
release point. Local spread of the rust up to 500 m from the release plot was observed at 11
sites within the season of release (Sheppard et al. 1993) and overwintering teliospores were
seen at many sites. Six sites were monitored in the subsequent season and natural re-
infection of common heliotrope by the rust was observed at three of them, but only on one or
a few plants per site and only at sites where good spread had been observed in the initial year.
Infection in the second year never spread nor multiplied the way it did in the initial year and
the rust could only be found at one of the monitored sites as spot infections two years after
the releases. While climate appeared limiting in WA (too dry), the main cause of poor
establishment of this agent is probably the inability of the overwintering teliospores to
synchronise germination with the reappearance of the target in spring (S Hasan pers. com.
1992). The annual life cycle and ephemeral nature of common heliotrope may also limit the
abundance and therefore the potential impact of the rust, which, following winter without its
host, must undergo a sexual cycle on re-infection each spring. While this agent has had no
apparent impact on the target in Australia, its persistence continued to be confirmed through
occasional sightings on common heliotrope at sites across south-eastern Australia unrelated
to those where original releases were made.
DISCUSSION
As one of the longest running of Australia’s biological control programs, with very extensive
native range research activities across Europe, North Africa and the Middle East, the program
against common heliotrope is most notable by its failure to deliver any kind of effective
control of the target. Frank Wilson, the earliest scientist to work on it, did not rate the
prospects of biological control very high. His 1950 report has since been lost so we can not
be sure of his reasoning, but it is likely he came to this conclusion after his initial efforts did
not find many apparently specific candidate agents in the 1950’s (listed in Delfosse and
Cullen1980), and it was only pressure from the agricultural sector for CSIRO to do
something, plus the opportunity to explore closer to the centre of origin of the species, that
stimulated more work. Also the limited chance of success of biological control against
summer annual cropping weeds is due to a number of factors, including their ephemeral
nature (Chaboudez and Sheppard 1995) was probably not as well appreciated at the time.

12
Annual plants are also likely to support a smaller community of natural enemies than longer
lived species because of this.
The only insect to be released, L. albineus, was as Huber (1981) rightly considered “a good
looking agent on anyone’s books”. That it was so hard to establish, despite a highly
successful mass-rearing program, is a significant enigma, unless Australian soil conditions or
summer temperatures or over-wintering conditions were just too unsuitable in some way for
successful beetle survival. As it was, there was every likelihood this agent would have had to
reach very high numbers very quickly within a growing season to have any impact on
common heliotrope populations and would have also attacked many native Heliotropium spp.
and possibly other Boraginaceae had it become widespread (CSIRO Entomology 1994).
This program also fell at a time when Australia was enthusiastic about plant pathogens as
potential agents following the successful use of a rust fungus for the biological control of
skeleton weed in Australia, another cropping weed. Expectations that the two promising
pathogens found on common heliotrope, including a rust fungus, could achieve similar levels
of success would have been quite high. With hindsight, could the failure of the common
heliotrope rust to impact on the weed have been predicted from its rarity in the native range?
Perhaps, but this program was the first to show that rust fungi do have an Achilles heal when
their host is so ephemeral. Most of the teliospores, the stage of the life-cycle of rusts that
enables survival over winter without a host, could have germinated with moisture in the
spring when environmental conditions were suitable and produced infective basidiospores,
but before temperatures were high enough for germination of the host plant. That is their may
be a problem of phenological synchrony. In comparison, the Cercospora spp. found on
common heliotrope cause a much more damaging disease, often resulting in severe epidemics
in both the native range and Australia (Brun et al. 1995; Sheppard et al. 1996). The impact
of these pathogens however, is driven by summer rainfall, which provides optimal conditions
for infection and associated rain splash necessary for local spread. Keeping the infected litter
in the paddocks may be a way of encouraging such epidemics, provided the period until new
host foliage is present at the sites is not too long.
This common heliotrope program provided a reality check on over optimism that successful
biological control of cropping weeds was possible. It also provided key lessons on the need
to focus on the target weed species rather than the target genus when there are con-generic

13
species native to Australia in order to not waste time investigating ineffective candidate
agents. Also since the early 1980’s agents to be released in Australia have had to be more
specific anyway to address the legislative requirements of the Wildlife and Countryside Act
of 1981.
ACKNOWLEDGEMENTS
We gratefully acknowledge the contribution of Laurent Brun, Ernest Delfosse, Siraj Hasan,
John Huber, Bobby Lewis, Carey Smith, Tony Wapshere, Frank Wilson, Tim Woodburn and
many technical staff to this biological control program.
REFERENCES Alonso-Zarazaga, MA (2010), Fauna Europaea: Pachycerus segnis (Germar 1824). Fauna
Europaea version 2.2, http://www.faunaeur.org
Brun LA, Sheppard AW and Carrara A (1993) Host range of Pachycerus cordiger Germar
(=P. scabrosus Brulle) (Col.: Curculionidae). Entomophaga 38, 537-539
Brun LA, Sheppard AW, Hasan S, Jourdan M, and Delmotte E (1995) Development of
Cercospora blight epidemics and effect on the summer annual weed Heliotropium
europaeum L. in the field. Annals of Applied Biology 127, 137-150.
Brun LA, Sheppard AW and Lopes V (1996) Variation in pathology of isolates of
Cercospora spp. attacking Heliotropium spp. Annals of Applied Biology 128, 565-570
Chaboudez P and Sheppard AW (1995) Are particular weeds more amenable to biological
control? - a reanalysis of mode of reproduction and life history. In: Proceedings of the VIIIth
International Symposium on Biological Control of Weeds. 2-7 February 1992, Canterbury,
New Zealand. (Eds ES Delfosse and RR Scott) pp. 95-102. DSIR/CSIRO, Melbourne,
Australia.
Chupp C (1953) A monograph of the fungus genus Cercospora. Ithaca, New York. pp. 667.
Comment [jul007 1]: Use en dashes rather than hyphens in the refs.

14
CSIRO Entomology (1994) Project CS.104 Biological control of common heliotrope,
Heliotropium europaeum. Contracted Research Report for Meat Research Corporation.
Cullen JM and Delfosse ES (1990) Progress and prospects in biological control of weeds. In:
Proceedings of the Ninth Australian Weeds Conference. 6-10 August, 1990, Adelaide. (Ed.
JW Heap) pp. 452-470. Crop Science Society of South Australia, Adelaide, Australia..
Davis P (1984) Flora of Turkey and the East Aegean Islands. Edinburgh University Press.
Delfosse ES (1985) Re-evaluation of the biological control program for Heliotropium
europaeum in Australia. In: Proceedings of the VI International Symposium on Biological
Control of Weeds. 19-25 August 1984, Vancouver, Canada. (Ed. ES Delfosse) pp 735-742.
Agriculture Canada, Ottawa, Canada.
Delfosse ES and Cullen JM (1980) New activities in biological control of weeds in Australia.
I. Common heliotrope, Heliotropium europaeum. In: Proceedings of the V International
Symposium on Biological Control of Weeds. 22-27 July 1980, Brisbane, Australia. (Ed. ES
Delfosse) pp. 545-561. CSIRO, Melbourne, Australia.
Edgar G, Hindmarsh WL, Keast JC and Rose AL (1941) The distribution and incidence of
enzootic or toxaemic jaundice in south-eastern Australia with special reference to New South
Wales. Australian Veterinary Journal 17, 120-130.
Hasan S (1985) Prospects for biological control of Heliotropium europaeum by fungal
pathogens. In: Proceedings of the VI International Symposium on Biological Control of
Weeds. 19-25 August 1984, Vancouver, Canada. (Ed. ES Delfosse) pp. 617-623. Agriculture
Canada, Ottawa, Canada.
Hasan S (1992) An update of the biological control of common heliotrope in Australia using
plant pathogens. In: Proceedings of the First International Weed Control Congress, Vol. 2.
17-21 February 1992, Melbourne, Australia. (Ed. RG Richardson) pp. 212-214. Weed
Science Society of Victoria Inc., Melbourne, Australia.

15
Hasan S and Aracil E (1991) Biology and effectiveness of Uromyces heliotropii Sred., a
potential biological control agent of Heliotropium europaeum L. New Phytologist 118, 559-
563
Hasan S and Ayres PG (1990) Tansley Review No. 23. The control of weeds through fungi :
principles and prospects. New Phytologist 115, 201-222.
Hasan S and Delfosse ES (1995) Susceptibility of the Australian native, Heliotropium
crispatum, to the rust fungus Uromyces heliotropii introduced to control common heliotrope,
Heliotropium europaeum. Biocontrol Science and Technology 5, 165-174
Hasan S, Delfosse ES, Aracil E and Lewis RC (1992) Host-specificity of Uromyces
heliotropii, a fungal agent for the biological control of common heliotrope (Heliotropium
europaeum) in Australia. Annals of Applied Biology 121, 697-705.
Hasan S, Jourdan M, Brun L and Sheppard AW (1995) Axenic culture and Influence of
wetness period and inoculum concentration on infection and development of cercospora
blight of Heliotropium europaeum. European Journal of Plant Pathology 101, 491-496
Huber JT (1979) A method for observing and rearing the root-feeding larvae of Longitarsus
albineus (Foudras) (Col. Chrysomelidae). Bulletin de la Société Entomologique Suisse 52,
431-433.
Huber JT (1981) Observations on the heliotrope flea beetle, Longitarsus albineus (Col.:
Chrysomelidae) with tests on its host specificity. Entomophaga 26, 265-73.
Huber JT and Vayssieres JF (1990) Life cycle and host specificity of the heliotrope weevil,
Pachycerus cordiger (= madidus Auct.) [Col.: Curculionidae]. Entomophaga 35, 475-484.
Hunt JR, Cousens RD and Knights SE (2008) The Biology of Australian Weeds 51.
Heliotropium europaeum L. Plant Protection Quarterly 23, 146-152
16
Moore CWE (1956) Observations on the autoecology of Heliotropium europaeum L. in New
South Wales and Victoria. CSIRO Division of Plant Industry Technical Paper No 7. CSIRO,
Melbourne.
Morin L and Trueman HE (1997) Pathogenicity and inoculum potential of two Cercospora
species on the weed Heliotropium europaeum. In: Programme and Summaries: 11th Biennial
Conference of the Australasian Plant Pathology Society. p. 49.
O’Dowd DJ and Edgar JA (1989) Seasonal dynamics in the pyrrolizidine alkaloids of
Heliotropium europaeum. Australian Journal of Ecology 14, 95-105.
Parsons WT and Cuthbertson EG (2001) Noxious Weeds of Australia. Second Edition.
CSIRO Publishing, Collingwood, Victoria.
Sheppard AW, Lewis RC and Delfosse ES (1993) The establishment of Uromyces heliotropii
Sred., a biological control agent of Heliotropium europaeum L. In: Proceedings of the 10th
Australian & 14th Asian -Pacific Weed Conference.6-10 September 1993, Brisbane,
Australia. p. 89-93. Queensland Weed Society, Brisbane, Australia.
Sheppard AW, Brun LA, Lewis RC. (1996) A demographic comparison of common
heliotrope, Heliotropium europaeum L.: southern Australia and southern France. Proceedings
of the 11th Australian Weeds Conference. 30 September-3 October 1996, Melbourne,
Australia. (Ed. RCH Shepherd) pp. 286-290. Weed Science Society of Victoria Inc.,
Melbourne, Australia.
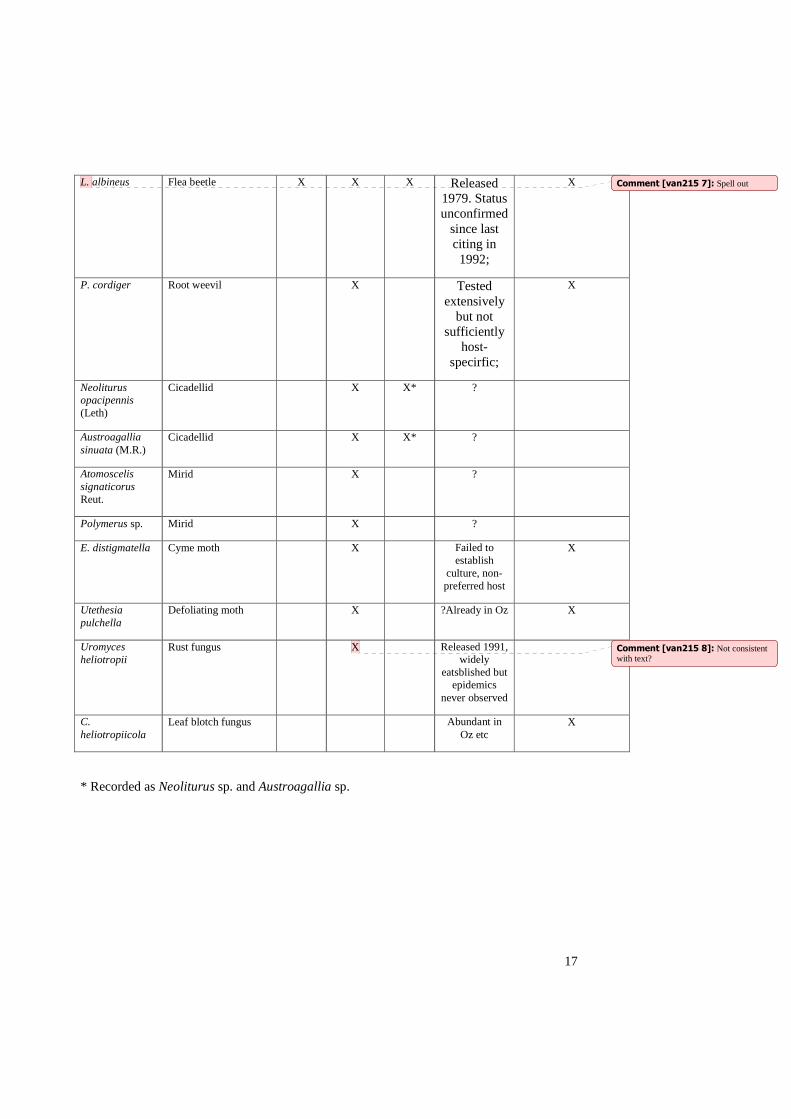
Table 1. Species considered of possible biological control potential discovered during surveys of H. europaeum.
Species Common name S. France,
E. Spain,
Algeria, Tunisia
1950
Iran 1971 1974
Tunisia, Algeria
1974
Med. Europe, Turkey. 1970s
Coleoptera
Comment [jul007 2]: More details?
Comment [van215 3]: These aren't really "common names" as such
Comment [van215 4]: Perhaps replace with "feeding damage/type" column
Comment [van215 5]: Not clear what dates refer to and what their significant is. Perhaps just have a column with "distribution" and a text field summarising it. (possibly with survey dates in brackets if that is important)
Comment [van215 6]: An additional column summarising state of play would be useful. E.g. Released 1979. Status unconfirmed since last citing in 1992; Tested extensively but not sufficiently host-specirfic;
17
L. albineus Flea beetle X X X Released 1979. Status unconfirmed
since last citing in 1992;
X
P. cordiger Root weevil X Tested extensively
but not sufficiently
host-specirfic;
X
Neoliturus opacipennis (Leth)
Cicadellid X X* ?
Austroagallia sinuata (M.R.)
Cicadellid X X* ?
Atomoscelis signaticorus Reut.
Mirid X ?
Polymerus sp. Mirid X ?
E. distigmatella Cyme moth X Failed to establish
culture, non-preferred host
X
Utethesia pulchella
Defoliating moth X ?Already in Oz X
Uromyces heliotropii
Rust fungus X Released 1991, widely
eatsblished but epidemics
never observed
C. heliotropiicola
Leaf blotch fungus Abundant in Oz etc
X
* Recorded as Neoliturus sp. and Austroagallia sp.
Comment [van215 7]: Spell out
Comment [van215 8]: Not consistent with text?
18
Figure 1. Distribution of common heliotrope, Heliotropium europaeum, in Australia.
(Reproduced from Australia’s Virtual Herbarium with permission of the Council of Heads of
Australasian Herbaria Inc; map generated on 1 May 2010).

19
A
B
Figure 2. Common heliotrope Heliotropium europaeum A. The plant. B. An infestation near
Gundagai, NSW.

20
A
B
Figure 3. A. The flea beetle Longitarsus albineus B. The rust fungus, Uromyces heliotropii.
Related Documents











