Review Biogeochemistry of paddy soils Ingrid Kögel-Knabner a , Wulf Amelung b , Zhihong Cao c , Sabine Fiedler d , Peter Frenzel e , Reinhold Jahn f , Karsten Kalbitz g , Angelika Kölbl a, ⁎, Michael Schloter h a Lehrstuhl für Bodenkunde, TU München, 85350 Freising-Weihenstephan, Germany b Rheinische Friedrich-Wilhelms-Universität Bonn, Institut für Nutzpflanzenwissenschaften und Ressourcenschutz, Bereich Bodenwissenschaften, Nußallee 13, 53115 Bonn, Germany c The Institute of Soil Science, CAS Chinese Academy of Sciences, 71 East Beijing Rd., Nanjing 210008, PR China d Universität Hohenheim, Institut für Bodenkunde und Standortslehre, 70593 Stuttgart, Germany e MPI für terrestrische Mikrobiologie, Karl-von-Frisch-Str. 1, D-35043 Marburg, Germany f Martin-Luther-Universität Halle-Wittenberg, Institut für Agrar- und Ernährungswissenschaften, Weidenplan 14, 06108 Halle, Germany g Universiteit van Amsterdam, Institute for Biodiversity and Ecosystem Dynamics, Earth Surface Science, Nieuwe Achtergracht 166, NL-1018 WV Amsterdam, The Netherlands h Helmholtz Zentrum München, Institut für Bodenökologie, Ingolstädter Landstraße 1, D-85764 Neuherberg, Germany abstract article info Article history: Received 17 August 2009 Received in revised form 25 January 2010 Accepted 8 March 2010 Available online 10 April 2010 Keywords: Anthrosols Wetland Paddy management Mineral transformation Soil organic matter Soil N Soil solution chemistry Microbial community Paddy soils make up the largest anthropogenic wetlands on earth. They may originate from any type of soil in pedological terms, but are highly modified by anthropogenic activities. The formation of these Anthrosols is induced by tilling the wet soil (puddling), and the flooding and drainage regime associated with the development of a plough pan and specific redoximorphic features. Redox potential oscillations due to paddy management control microbial community structure and function and thus short-term biogeochemical processes. After flooding, microbial reduction processes sequentially use NO 3 − , Mn 4+ , Fe 3+ , SO 4 2− as electron acceptors, accompanied by the emission of the trace gases N 2 O, N 2 ,H 2 S, CH 4 and — due to reduction-induced increasing pH — NH 3 . This results in N losses and low N fertilizer use efficiency. However, transport of atmospheric O 2 to the roots via the rice plant's aerenchyma modifies conditions in the rhizosphere, leading to nitrification and methane oxidation, and precipitation of Mn and Fe oxides. High concentrations and fluxes of dissolved organic matter (DOM) in paddy soils from plant debris trigger microbial activity and thus the emission of greenhouse gases. Retention of DOM by soil minerals and its subsequent stabilisation against microbial decay depend on the redox state (e.g. DOM precipitation by Fe 2+ under anaerobic conditions). Oscillation in redox conditions may enhance retention and stabilisation of DOM by Fe oxyhydroxides. Induced by the periodic short-term redox cycles, paddy management over long periods has strong effects on long-term biogeochemical processes. Frequent irrigation intensifies mineral weathering and leaching processes. High concentrations of DOM during flooding seasons enhance the changes and the release of structural iron in clay minerals, and support the formation of ferrihydrite. Repeated redox alternations lead to a translocation of iron in various directions, and particularly increase the crystallinity of iron oxides. This results also in higher total iron oxide contents in paddy compared to non-paddy soils. The large accumulation of soil organic matter (SOM) observed in some, but not all paddy soils, is considered to be due to high input of plant residues and charred material associated with retarded decomposition under anaerobic conditions. There is also evidence of SOM stabilisation via occlusion into aggregates and phytoliths as well as interactions with clay minerals and iron oxides. SOM accumulation in paddy subsoils can be explained by downward movement of DOM and its stabilisation by interaction with iron oxides. A specific feature of paddy soils is the coupling of organic matter turnover with mineral transformations and fluxes, which seem to be intensified by the alternating redox conditions with increasing age of paddy soil development. Bioavailability of soil organic N is strongly coupled to SOM cycling and is a crucial parameter determining crop yield. Anaerobic conditions inhibit N mineralization, with a high risk of gaseous N losses. In paddy soils the management-induced, microbially mediated redox processes control the dynamics of soil minerals and soil organic matter, which are strongly related to the microbial accessibility of C and N, but also of Fe. © 2010 Elsevier B.V. All rights reserved. Geoderma 157 (2010) 1–14 ⁎ Corresponding author. Fax: + 49 8161 71 4466. E-mail address: [email protected] (A. Kölbl). 0016-7061/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.geoderma.2010.03.009 Contents lists available at ScienceDirect Geoderma journal homepage: www.elsevier.com/locate/geoderma

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Geoderma 157 (2010) 1–14
Contents lists available at ScienceDirect
Geoderma
j ourna l homepage: www.e lsev ie r.com/ locate /geoderma
Review
Biogeochemistry of paddy soils
Ingrid Kögel-Knabner a, Wulf Amelung b, Zhihong Cao c, Sabine Fiedler d, Peter Frenzel e, Reinhold Jahn f,Karsten Kalbitz g, Angelika Kölbl a,⁎, Michael Schloter h
a Lehrstuhl für Bodenkunde, TU München, 85350 Freising-Weihenstephan, Germanyb Rheinische Friedrich-Wilhelms-Universität Bonn, Institut für Nutzpflanzenwissenschaften und Ressourcenschutz, Bereich Bodenwissenschaften, Nußallee 13, 53115 Bonn, Germanyc The Institute of Soil Science, CAS Chinese Academy of Sciences, 71 East Beijing Rd., Nanjing 210008, PR Chinad Universität Hohenheim, Institut für Bodenkunde und Standortslehre, 70593 Stuttgart, Germanye MPI für terrestrische Mikrobiologie, Karl-von-Frisch-Str. 1, D-35043 Marburg, Germanyf Martin-Luther-Universität Halle-Wittenberg, Institut für Agrar- und Ernährungswissenschaften, Weidenplan 14, 06108 Halle, Germanyg Universiteit van Amsterdam, Institute for Biodiversity and Ecosystem Dynamics, Earth Surface Science, Nieuwe Achtergracht 166, NL-1018 WV Amsterdam, The Netherlandsh Helmholtz Zentrum München, Institut für Bodenökologie, Ingolstädter Landstraße 1, D-85764 Neuherberg, Germany
⁎ Corresponding author. Fax: +49 8161 71 4466.E-mail address: [email protected] (A. Kölbl).
0016-7061/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.geoderma.2010.03.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 August 2009Received in revised form 25 January 2010Accepted 8 March 2010Available online 10 April 2010
Keywords:AnthrosolsWetlandPaddy managementMineral transformationSoil organic matterSoil NSoil solution chemistryMicrobial community
Paddy soils make up the largest anthropogenic wetlands on earth. They may originate from any type of soil inpedological terms, but are highly modified by anthropogenic activities. The formation of these Anthrosols isinduced by tilling the wet soil (puddling), and the flooding and drainage regime associated with thedevelopment of a plough pan and specific redoximorphic features. Redox potential oscillations due to paddymanagement control microbial community structure and function and thus short-term biogeochemicalprocesses. After flooding, microbial reduction processes sequentially use NO3
−, Mn4+, Fe3+, SO42− as electron
acceptors, accompanied by the emission of the trace gases N2O, N2, H2S, CH4 and — due to reduction-inducedincreasing pH — NH3. This results in N losses and low N fertilizer use efficiency. However, transport ofatmospheric O2 to the roots via the rice plant's aerenchyma modifies conditions in the rhizosphere, leadingto nitrification and methane oxidation, and precipitation of Mn and Fe oxides. High concentrations and fluxesof dissolved organic matter (DOM) in paddy soils from plant debris trigger microbial activity and thus theemission of greenhouse gases. Retention of DOM by soil minerals and its subsequent stabilisation againstmicrobial decay depend on the redox state (e.g. DOM precipitation by Fe2+ under anaerobic conditions).Oscillation in redox conditions may enhance retention and stabilisation of DOM by Fe oxyhydroxides.Induced by the periodic short-term redox cycles, paddy management over long periods has strong effects onlong-term biogeochemical processes. Frequent irrigation intensifies mineral weathering and leachingprocesses. High concentrations of DOM during flooding seasons enhance the changes and the release ofstructural iron in clay minerals, and support the formation of ferrihydrite. Repeated redox alternations leadto a translocation of iron in various directions, and particularly increase the crystallinity of iron oxides. Thisresults also in higher total iron oxide contents in paddy compared to non-paddy soils.The large accumulation of soil organic matter (SOM) observed in some, but not all paddy soils, is consideredto be due to high input of plant residues and charred material associated with retarded decomposition underanaerobic conditions. There is also evidence of SOM stabilisation via occlusion into aggregates and phytolithsas well as interactions with clay minerals and iron oxides. SOM accumulation in paddy subsoils can beexplained by downward movement of DOM and its stabilisation by interaction with iron oxides. A specificfeature of paddy soils is the coupling of organic matter turnover with mineral transformations and fluxes,which seem to be intensified by the alternating redox conditions with increasing age of paddy soildevelopment. Bioavailability of soil organic N is strongly coupled to SOM cycling and is a crucial parameterdetermining crop yield. Anaerobic conditions inhibit N mineralization, with a high risk of gaseous N losses. Inpaddy soils the management-induced, microbially mediated redox processes control the dynamics of soilminerals and soil organic matter, which are strongly related to the microbial accessibility of C and N, but alsoof Fe.
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.

2 I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
Contents
1. Introduction — general characteristics of paddy soils . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22. Management and development of paddy soils . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23. Specific characteristics of short-term biogeochemical processes in paddy soils . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3.1. Redox cycling and pH conditions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33.2. Microbial metabolism and corresponding trace gas production . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43.3. Dynamics of dissolved organic matter (DOM) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
4. Specific characteristics of long-term biogeochemical processes in paddy soils . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74.1. Mineral weathering and (neo)transformation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74.2. Mineral transformation in the presence of organic matter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74.3. Accumulation and stabilization of soil organic matter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74.4. Soil organic N in paddy fields . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
5. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
1. Introduction — general characteristics of paddy soils
Rice is the most important food for more than 50% of the world'spopulation, and it is grown on almost 155 million ha of the world'ssurface (http://beta.irri.org/index.php/). World rice production in2008 was approximately 661 million tons, with more than 90%produced in Asia. At least 114 countries grow rice. China is the world'slargest rice producer, accounting for 30% of total world produc-tion, followed by India (22%), Indonesia (9%), and Bangladesh (7%)(http://beta.irri.org/statistics). Cultivated rice (Oryza sativa) isan annual grass that evolved from a semi-aquatic ancestor. Becauseof its semi-aquatic ancestry, rice is extremely sensitive to water shor-tage. Therefore, rice ismostly grownunderflooded lowland conditionsin bunded fields (paddies). The traditional way of growing paddyrice involves the tillage of flooded soil (puddling), followed by ricetransplantation and growing the crop in a submerged environmentfrom crop establishment to close to harvest (Gong, 1983; Sahrawat,2005; http://www.knowledgebank.irri.org/watermanagement).Nearly 80% of Asia's total rice growing area are managed aspaddy soils, corresponding to 139617000 ha in 2008 (http://www.knowledgebank.irri.org/oryza2000/Oryza_User_Manual; http://beta.irri.org/solutions). Most rice production is in the southern part ofChina in the Yangtze and Huaihe River Valley (or further south) whereample supplies of water are available. In south-eastern China, hightemperature and abundant rainfall provide an ideal environment forrice during a long growth period, and many areas grow two crops ofrice per year. In the Yangtze River Valley,much of the land is planted toa rice-wheat or other winter upland crop rotation (Cao and Zhang,2004; Lin et al., 2004).
Over the past few years, work on paddy soils has mostly beenconfined tomicrobiology and concerns about greenhouse gas emissions(Aulakh et al., 2001; Cai et al., 2003, 2005; Conrad and Frenzel, 2002;Conrad, 2007; Kimura et al., 2004; Neue and Roger, 2003; Wassmannet al., 2000a,b). When alternative electron acceptors are consumed,equimolar amounts of CO2 and CH4 are the end products of anaerobiccarbonmineralization (Kirk, 2004). However, only part of the producedCH4 is emitted, while a significant fraction may be oxidized at oxic-anoxic interfaces (Frenzel, 2000; Krüger et al., 2001, 2002), orpercolated to the subsoil layer (Kimura et al., 2004;Murase and Kimura,1994, 1996). Geochemical properties, such as the amount anddegradability of organic matter (OM) or iron minerals, affect microbialactivities. Conversely, microbes affect not only the turnover of theirprimary substrates, but also pH, redox potentials, complexation ofmetals, and solid phase chemistry bymodulating adsorption/desorptionand dissolution/precipitation.
Despite the large number of studies dealing with the chemistry,physics and microbiology of paddy soils, there is still a lack of
knowledge concerning the biogeochemistry of paddy soils. Theobjective of this review is to summarize the current knowledge onpaddy soil biogeochemistry with respect to short-term and long-term processes involved in the formation of these soils. Themanagement-induced redox cycles lead to specific changes in thesoil profile morphology. These are induced by short-term biogeo-chemical processes with specific characteristics of redox cycling andmicrobial metabolism that affect soil solution chemistry, trace gasproduction and dynamics of DOM. In the long term, the anthropo-genic soil management then controls the weathering and formationof minerals as well as accumulation of organic matter and organicnitrogen.
2. Management and development of paddy soils
Rice cultivation depends more on the moisture conditions underwhich the plant is grown than upon the former nature of the soils.Rice is not very sensitive to prior soil conditions with respect totexture and nutrient status, except for high sulphate contents(Barnes, 1990; Kawaguchi and Kyuma, 1977). Thus, paddy soildevelopment is driven by the specific soil management practices thatmask the soil's original character (Kirk, 2004). These are artificialsubmergence and drainage, ploughing and puddling (=ploughingand leveling the surface layer of a submerged soil), organic manuring(animal manure, rice straw and other crop residues, often fermentedwith sediments taken from the river or channel), liming andfertilization. The management-induced change of oxic and anoxicconditions results in temporal and spatial (vertical, horizontal)variations in reduction and oxidation (redox) reactions affecting thedynamics of organic and mineral soil constituents (Cheng et al.,2009).
Anoxic conditions prevail during most of the time of rice growth.However, when the fields are drained one to two weeks beforeharvest, the redox potential increases, i.e. reduced compounds suchas Fe2+ are oxidised (Jäckel et al., 2001; Krüger et al., 2001; Rateringand Conrad, 1998; Ratering and Schnell, 2000). Oxic conditions aresustained over a longer period of timewhen upland crops (e.g. wheator oil seeds) are grown after paddy rice cultivation.
This anthropogenic cycle might be accompanied by the progres-sive loss of clay during paddy cultivation, mainly by surface runoffduring heavy rain, as a result of the destruction of the soil structurethrough puddling (Eaqub and Blume, 1982; Zhang and Gong, 2003).The drop in redox potential leads to increasing flocculation anddispersion of clay particles, which enhances the migration of clay inpaddy soils (Li and Horikawa, 1997; Li et al., 1997). The migration ofclay into the bottom of the plough layer, in addition to mechanicalpressure exerted by buffalos or tractors, contributes to forming a

Fig. 1. Typical horizon sequence of a rice paddy soil (horizon designation according toFAO, 2006).
3I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
plough pan (Li, 1992). Repeated puddling over many years increasesthe plant available water capacity in the puddled layer, but reducesthe small coarse and meso pores (50 to 0.2 µm) in the plough pan(Janssen et al., 2006). The reduction of macropores and the increaseof micropores in the puddled horizon (Ap) correspond to thepuddling intensity which is, therefore, responsible for the waterretention characteristics of the puddled layers (Eickhorst andTippkötter, 2009).
The hydraulic properties of the plough pan depend mainly on thelength of time the soil is used for rice cultivation, with theconsolidation of the plough pan continuing for many decades(Lennartz et al, 2009). The increasing age of paddy fields isaccompanied by strongly decreasing infiltration rates, therebyconfirming the importance of maintaining an undisturbed, perma-nent plough pan in order to increase water use efficiency (Janssenand Lennartz, 2006, 2007). The differences in the properties (e.g.aggregation, accumulation of iron in the subsoil, number of biopores)of subsoil horizons apparently increase with the length of timeduring which paddy soil is formed and its diagnostic horizons. Theseare an anthraquic horizon (≥20 cm) and an underlying hydragrichorizon, according to the IUSS Working Group WRB (2006). Theanthraquic horizon is a human induced topsoil formed by wetcultivation and contains the puddled layer and the plough pan. Thediagnostic hydragric horizon is characterized by the abundance ofreduction features or the segregation of Fe/Mn in the matrix (IUSSWorking Group WRB, 2006). The anthraquic and an underlyinghydragric horizon are the diagnostic horizons of Hydragric Anthro-sols. Other paddy soils, in which substantial amounts of sedimentshave accumulated over time due to irrigation, show an irragrichorizon as diagnostic topsoil (Irragric Anthrosols).
Paddy management leads to the development of pedogenetichorizons that are specific to paddy soils (Fig. 1, horizon nomenclatureaccording to FAO, 2006):
W: thin layer of standing water that is the habitat of bacteria,phytoplankton, macrophytes and small fauna. This horizon is mainlyoxic.
Ap: oxic and partly oxic zone; the thickness of the oxic zone mayrange from several mm after flooding, to several cm when the riceplants are fully grown and start to release oxygen from their roots(Frenzel et al., 1992). Furthermore, pedoturbation by soil fauna,evapo-transpiration and percolation affect the extent of the oxic zone(Yu et al., 2007).
Arp: the upper part of an anthraquic horizon (IUSS WorkingGroupWRB, 2006) is the reduced puddled layer, characterized by theabsence of free oxygen in the soil solution and an rH value b20(rH=2pH+2Eh/59, at 25 °C), i.e. conditions which lead to theformation of Fe2+. Frequently, this layer has a thickness of about15 cm.
Ardp: the lower part of an anthraquic horizon or plough pan ortraffic pan. This horizon (N7 cm) is relatively compact, it has a platystructure, high mechanical strength and low hydraulic conductivity(0.34 to 0.83 mm day−1) (Chen and Liu, 2002). Consequently, stagnicand reducing conditions are maintained.
Hydraulic properties of the plough pan predominantly control thewater regime of the underlying B or C horizons which may haveeither oxic or reducing conditions. In flooded paddy soils, aquicmoisture regimes (i.e. gleyic conditions) prevail. The subsoil horizongenerally remains reduced, with mottling along cracks and widepores, and at least the interior of soil peds remains reduced duringmost years, unlike when the water table is perched.
Hydragric Anthrosols (IUSS Working Group WRB, 2006) mayoriginate from different reference soil groups, most often Fluvisols,Cambisols, Vertisols, Andosols, Ultisols, and Ferralsols. The specificsoil management and the continuing paddy soil use lead toalternating redox conditions and therefore to soil properties andmorphologies that are independent of the initial soil unit (Fig. 2).
3. Specific characteristics of short-term biogeochemical processesin paddy soils
3.1. Redox cycling and pH conditions
After flooding, water replaces the gaseous phase in the soil pores.Since oxygen diffusion times in water are four orders of magnitudelower than those in air, oxygen is depleted from the bulk topsoil within24 h (Boivin et al., 2002; Gao et al., 2002; Tanji et al., 2003). Besidesoxygen, other inorganic soil constituents contribute sequentially to theoxidationof organicmatter (Conradand Frenzel, 2002; Ponnamperuma,1972). Redox reactions in soils are mainly controlled by microbialactivity (Munch et al., 1978). Organisms use organic substances such ascarbon sources as electron donors during respiration.Molecular oxygenacts as the preferred electron acceptor as long as it is available. Floodinga field for subsequent rice cultivation cuts off the oxygen supply fromthe atmosphere, the microbial activities switch from aerobic (i.e. oxiccondition) to facultative (i.e. hypoxic condition) and to anaerobic (i.e.anoxic condition) fermentation of organic matter, where alternativeelectron acceptors are used. The sequence is determined by thermody-namics and includes, in terms of Eh, from high to low: aerobicrespiration, nitrification, denitrification, Mn4+-reduction, Fe3+-reduc-tion, SO4
2−-reduction and methanogenesis (Ponnamperuma, 1972;Patrick and Reddy, 1978; Patrick and Jugsujinda, 1992). In terms ofquantity, Fe3+ is by far themost important oxidant in rice field soils andthus determines the period during which organic matter is oxidized toCO2 (Yao et al., 1999). The critical Eh-value for Fe reduction and

Fig. 2. Typical paddy soil profiles after 100 y (Gleyic Cambisol (Eutric, Siltic)) and 1000 y (Endogleyic Stagnosol (Albic, Ruptic, Eutric, Siltic)) of paddy soil management in theZheijang Province, P.R. China (photos: R. Jahn).
4 I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
consequent dissolution is 100 mV at pH 7. However, different soils varyin the critical redox values for Fe transformation because the mineralphases of soils are mixed and the presence of dissolved organicsubstances can amplify reducing conditions (summarized by Fiedleret al., 2007). When microbially reducible iron is depleted, organiccarbon is disproportionated to CO2 plus CH4.
The magnitude of redox changes in paddy soils can vary from onesoil to another, and depends strongly on the ratio of oxidizing capacityto reducing capacity (Takai, 1961 cit. by Bouwman, 1991), which isthe result of the cultivation system and the length of cultivation (Xuet al., 2000). Particularly the incorporation of organic matter, i.e.electron donors, can be enhanced through the reducing conditionsoccurring shortly after flooding (Xu et al., 2000).
Beside macroscopic redox gradients between thin oxic layers (W,Apo) and anoxic A horizons, millimetre-scale gradients of Eh occuraround rice roots (Frenzel et al., 1992; Gilbert and Frenzel, 1998; Arthand Frenzel, 2000; Ratering and Schnell, 2000). Rice that is grown insubmerged paddy soils form aerenchymawhich enable the transport ofatmospheric O2 to the roots (Armstrong, 1971, Begg et al., 1994), wherea higher redox potential (Flessa and Fischer, 1992; Tyagi et al., 2004)mediates the detoxification of Mn4+, Fe2+ by oxidation and results inthe precipitation of Mn and Fe oxides in the root apoplast, which isvisible as so-called plaque at the root surface (Trolldenier, 1988). Theformation of these plaques can restrict the acquisition of phytotoxicelements such as Cu (Greipsson (1994), Ni (Greipsson, 1995), Cd (Luiet al., 2008) and As by rice (Bravin et al., 2008; Liu et al., 2005).
Soil reduction is accompanied by changes in the pH: An increase inpH of acid soils due to consumption of protons and a decrease in pH ofalkaline soils due to increasing PCO2 can be observed after flooding(Sahrawat, 2005). Changes in pH are reversible. The soil solutionreaction is neutral at the beginning of the cropping season, becomesalkaline at the end, but returns to neutral at the beginning of the nextcropping season (Kirk, 2004). According to Nernst's equation, redoxchanges of 59 mVare accompanied by pH changes of 1 unit, whereas Yuand Patrick (2003) suggested that the change of the redox potential perpH unit might even vary from 59 to 177 mV.
3.2. Microbial metabolism and corresponding trace gas production
The different anaerobic reduction processes are accompanied by anemission of trace gases, such as NH3, N2O, N2, H2S and CH4. The initialrise in pH at the beginning of reduction processes after flooding and theconsumption of CO2 by algae in thewater layer promotes gaseous lossesof ammonium as NH3. In Asia alone, more than 2 million tons of N arelost by this pathway, accounting for up to 23% of applied fertilizeramounts in certain regions (FAO, 2001). Furthermore, N losses vianitrate also play an important role, as nitrate is highly mobile and canleach easily to deeper soil layers or can be used by microbes as analternative terminal electron acceptor under anoxic conditions, whichleads to a stepwise reduction to nitrite, NO, N2O and N2. Therefore, notsurprisingly, N fertilizer use efficiency may frequently fall below 35%(DeDatta, 1981; Cao et al., 1984a,b), and may, hence, be less than onehalf of the efficiency typically found in other agricultural systems (Royand Misra, 2003). It might be speculated that the amount of nitritetransformed into ammonium depends on the amount of availablenitrate. If huge amounts of nitrate are available,microbeswill use this asthe alternative electron acceptor for energetic reasons and thus nitritecan be transformed into ammonia. If nitrate is limited, however,microbes will use nitrite, NO and N2O as terminal electron acceptors,resulting in theproductionof dinitrogen,whichwill be lost from the soil.Indeed it could be shown that a tight coupling between nitrification anddenitrification to dinitrogen exists (Arth et al., 1998). Furthermore,concentrations within a flooded paddy soil are highly variable at themillimetre scale, as shown by steep counter-gradients of ammoniumand nitrate measured around rice roots (Arth and Frenzel, 2000).
In most rice systems, methanogenesis is the most importantterminal process in carbon mineralization. Due to the radiative forcingcaused by methane, much work has been done on methane emissions.Emissions can be quite different depending on water management,mineralogy, rice cultivar, fertilization, and local climate (Cai et al., 2001;Denier van der Gon, 2000; Neue et al., 1996; Yan et al., 2005). Floodedrice paddies are one of the major biogenic sources of the atmosphericgreenhouse gasmethane (Scharpenseel et al., 1996; Liesack et al., 2000;

5I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
van der Gon et al., 2002) (Intergovernmental Panel on Climate Change,2007). Overall, wetland rice fields contribute about 10–25% to globalCH4 emission (Neue et al., 1997). Crop rotation with flooded rice insummeranduplandcrops suchaswheat or vegetables inwinter reducesmethane emission compared to repeated cropping with rice (Cai et al.,2003, 2005).
The rice plants supply the microbes in soil and rhizosphere withsubstrates: about 30 to 60% of net photosynthesized carbon is allocatedto roots, and asmuch as 40 to 90% of this fraction is translocated into thesoil (Lynch and Whipps, 1990). In paddies, this below-ground carbonflow provides a major carbon source for methane production (Minodaand Kimura, 1994; Minoda et al., 1996). Early in the season, however,rice straw and stubbles are assumed to provide substrates formethanogenesis, while exudates become more important during latetillering and ripening (Neue et al., 1996).Methane emission frompaddysoils planted to three Chinese cultivars has been found to differ up to afactor of ten (Jia et al., 2002). While methanogenic pathways are wellunderstood, we are just beginning to touch on microbial communitystructures. Similarly, our understanding of controls by abiotic and bioticfactors is far from complete. This is mainly related to the fact that evenkey microbial players in themost important soil processes have not yetbeen isolated. Therefore, a molecular toolbox, including genomics,transcriptomics and proteomics, has been developed over the pastdecade to characterize microbial community structure and functionwithout the need for cultivation.
Methanogenic archaea belonging to the Rice Cluster I (RC-I) havebeen identified as key players (Lu et al., 2005; Lu and Conrad, 2005).Based on metagenomic analysis, these organisms seem to beaerotolerant with an H2/CO2-dependent metabolism, and a uniqueset of antioxidant enzymes and DNA repair mechanisms. Altogether,RC-I seems to have a selective advantage over other methanogens inparticular in the rhizosphere (Erkel et al., 2006). However, methaneproduction not only occurs within the rhizosphere, driven by rootexudates, but also during litter degradation in the bulk soil. Peng et al.(2008) postulated that straw residues promote the growth ofMethanosarcinaceae, whereas the root residues favour Methanosaeta-ceae. The in situ concentrations of acetate and H2 also seem to be keyfactors controlling the structure of the methanogenic community(Krüger et al., 2005) and the anaerobic flow of carbon and reductants(Conrad and Frenzel, 2002).
Oxygen released from the roots enables oxidative processes likenitrification and methane oxidation. Therefore, methane emission notonly depends on production but also on oxidation according toemission=production–oxidation (Eller and Frenzel, 2001; Eller et al.,2005a; Eller and Frenzel, 2000; Krüger et al., 2002; Bilek et al., 1999;Bosse and Frenzel, 1998; Denier van der Gon and Neue, 1996; Jia et al.,2001). While some methanothrophs occur almost everywhere, somephylogenetic clusters are specific to wetland rice fields (Lüke et al.,2010). Within a rice field, methanothrophs show no spatial pattern(Krause et al., 2009). However, the seasonal patterns of methaneoxidation seem to be highly variable. At the very surface of the floodedpaddy, more than 90% of the potentially emitted methane may beoxidized (Rothfuss et al., 1996; Gilbert and Frenzel, 1998). However,overall mitigation of emission by methane oxidation accounts for onlyabout 10–50% (Bilek et al., 1999; Bosse and Frenzel, 1998; Denier vander Gon and Neue, 1996; Khalil et al., 1998; Krüger et al., 2001).Considering that most methane is emitted via the rice plant'saerenchyma (Aulakh et al., 2000, 2002; Denier van der Gon and vanBreemen, 1993;Wang et al., 1997), plant-associatedmethane oxidationis crucial inmitigatingmethane emission. However, the seasonality andcontrols are not fully understood. Recent work suggests that grazingprotists may be a factor controlling methanotrophs (Murase andFrenzel, 2007; Murase et al., 2006; Murase and Frenzel, 2008), but thishas not yet been explored in the field. Methane oxidation has beenreported to be inhibited by high concentrations of mineral nitrogen(Bosse et al., 1993; Conrad and Rothfuss, 1991). However, different
experiments have shown that ammonium-based fertilizationmay evenenhancemethaneoxidation in rice paddies, leading to reducedmethaneemissions (Bodelier et al., 2000a,b). Unfortunately, this may be only atransient effect (Krüger and Frenzel, 2003), suggesting that thecompetition for mineral nitrogen between rice roots and rhizosphericmicrobes is a key factor regulating microbial processes.
As the functional genes for methane oxidation (pmoA) andammonium oxidation (amoA) are very similar, ammonium oxidizingbacteria have been suggested to potentially contribute to methaneoxidation. Experimental work, however, has shown this to be ratherimprobable (Bodelier and Frenzel, 1999). Recent studies have demon-strated that ammonium-oxidizing archaea (AOA) predominate amongammonia-oxidizing prokaryotes in a paddy soil and in the rhizospherein particular (Chen et al., 2008). However, their contribution tonitrification or methane oxidation has not yet been verified. Anaerobicmethane oxidation has been assumed to be restricted to marineenvironments, but recently a freshwater enrichmenthas beendescribedas oxidizing methane anaerobically with nitrite as an electron acceptor(Ettwig et al., 2008, 2009). First evidence has been published thatanaerobic methane oxidation may be coupled to sulphate reductioneven in freshwater lakes (Eller et al., 2005b). However, it remains to beestablished whether these processes play a role in paddy soils.
In summary, N- and C-cycles are closely linked, as shown in asimplified sketch in Fig. 3. Anaerobic processes compete for fermenta-tion products,mainly acetate andhydrogen,while aerobic processes notonly compete for oxygen, but also for nutrients (as shown forammonium).
3.3. Dynamics of dissolved organic matter (DOM)
Typical values for seepage and percolation vary from 1–5 mm day−1
in heavy clay soils to 25–30mm day−1 in sandy and sandy loam soils(Bouman et al., 2006). Despite these large water fluxes, little is knownabout the dynamics of DOM in paddy soils. We found just one studypublished in English where fluxes of dissolved organic carbon (DOC)were determined in the field during a 98-day study (Katoh et al., 2004).Up to 320–630 kg DOC ha−1 a−1 percolated from the puddled horizoninto the subsoil, thus exceeding the large fluxes from Oa horizons offorest soils under temperate climates (100–400 kg C ha−1 a−1:Michalzik et al., 2001). In a lysimeter study, Maie et al. (2004) reported200 kg C ha−1 leaving the plough layer during the rice growth period ofapproximately 4 months. Li et al. (2005a,b) investigated a chronose-quence of paddy fields in subtropical China. In this study DOCconcentrations increased continuously after 10 years of rice cultivation.Theywere smaller in cultivated soils in comparison towastelands at thebeginning of cultivation.
These large DOC concentrations and fluxes in paddy soils might bethe result of longperiodswith anaerobic conditions in theAhorizon. It ispostulated that decomposition of organic matter is less efficient underanaerobic conditions, resulting also in an enrichment of water-solubleintermediate metabolites as organic anions like acetate, formate,propionate, and lactate (Ponnamperuma, 1972; Moore and Dalva2001; Fiedler and Kalbitz, 2003; Sahrawat, 2004). However, theconcentrations of low molecular organic anions are quite small; e.g.acetate concentrations are mostly smaller than 100 µM (Krüger et al.,2001, 2005). Therefore, such compounds should rarely contributemorethan 10% to total DOC. In contrast, plant derived C seems to play a largerrole for DOM in paddy soils than in upland soils (Lu et al., 2000, 2002,2004). The large proportion of degradableDOMderived from freshplantdebris (Kalbitz et al., 2003a) might be the reason for the close linkagebetweenDOCconcentrations,microbial biomass andproduction ratesofCO2 and CH4. (Lu et al., 2000, 2002; Lou et al., 2004; Cheng et al., 2005;Jiao et al., 2005),whichmay indicate thatmajor parts of the DOM add atleast potentially to an easily biodegradable C pool.
Lignin-derived compounds which are relatively stable againstmicrobial decay (Kalbitz et al., 2003a) might also make up a larger
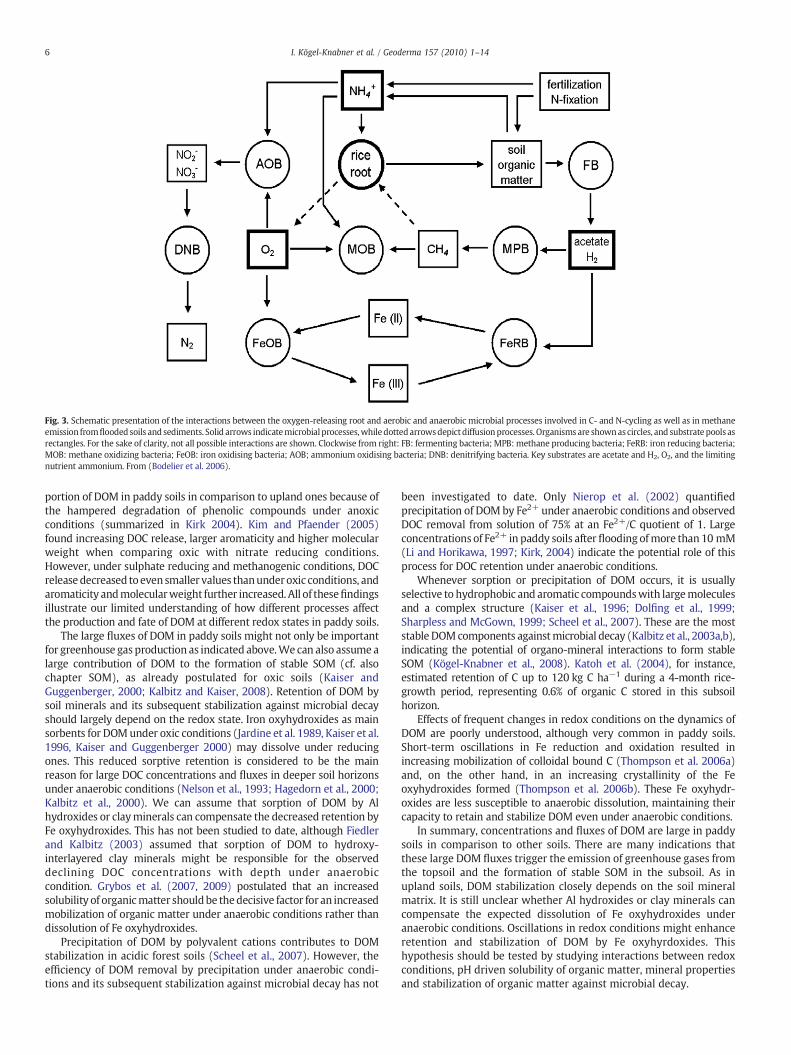
Fig. 3. Schematic presentation of the interactions between the oxygen-releasing root and aerobic and anaerobic microbial processes involved in C- and N-cycling as well as in methaneemission fromflooded soils and sediments. Solid arrows indicatemicrobial processes,whiledottedarrowsdepictdiffusionprocesses. Organismsare shownas circles, and substrate pools asrectangles. For the sake of clarity, not all possible interactions are shown. Clockwise from right: FB: fermenting bacteria; MPB: methane producing bacteria; FeRB: iron reducing bacteria;MOB: methane oxidizing bacteria; FeOB: iron oxidising bacteria; AOB; ammonium oxidising bacteria; DNB: denitrifying bacteria. Key substrates are acetate and H2, O2, and the limitingnutrient ammonium. From (Bodelier et al. 2006).
6 I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
portion of DOM in paddy soils in comparison to upland ones because ofthe hampered degradation of phenolic compounds under anoxicconditions (summarized in Kirk 2004). Kim and Pfaender (2005)found increasing DOC release, larger aromaticity and higher molecularweight when comparing oxic with nitrate reducing conditions.However, under sulphate reducing and methanogenic conditions, DOCrelease decreased to evensmaller values thanunderoxic conditions, andaromaticity andmolecularweight further increased. All of thesefindingsillustrate our limited understanding of how different processes affectthe production and fate of DOM at different redox states in paddy soils.
The large fluxes of DOM in paddy soils might not only be importantfor greenhouse gas production as indicated above.We can also assume alarge contribution of DOM to the formation of stable SOM (cf. alsochapter SOM), as already postulated for oxic soils (Kaiser andGuggenberger, 2000; Kalbitz and Kaiser, 2008). Retention of DOM bysoil minerals and its subsequent stabilization against microbial decayshould largely depend on the redox state. Iron oxyhydroxides as mainsorbents for DOMunder oxic conditions (Jardine et al. 1989, Kaiser et al.1996, Kaiser and Guggenberger 2000) may dissolve under reducingones. This reduced sorptive retention is considered to be the mainreason for large DOC concentrations and fluxes in deeper soil horizonsunder anaerobic conditions (Nelson et al., 1993; Hagedorn et al., 2000;Kalbitz et al., 2000). We can assume that sorption of DOM by Alhydroxides or clayminerals can compensate the decreased retention byFe oxyhydroxides. This has not been studied to date, although Fiedlerand Kalbitz (2003) assumed that sorption of DOM to hydroxy-interlayered clay minerals might be responsible for the observeddeclining DOC concentrations with depth under anaerobiccondition. Grybos et al. (2007, 2009) postulated that an increasedsolubility of organicmatter shouldbe thedecisive factor for an increasedmobilization of organic matter under anaerobic conditions rather thandissolution of Fe oxyhydroxides.
Precipitation of DOM by polyvalent cations contributes to DOMstabilization in acidic forest soils (Scheel et al., 2007). However, theefficiency of DOM removal by precipitation under anaerobic condi-tions and its subsequent stabilization against microbial decay has not
been investigated to date. Only Nierop et al. (2002) quantifiedprecipitation of DOM by Fe2+ under anaerobic conditions and observedDOC removal from solution of 75% at an Fe2+/C quotient of 1. Largeconcentrations of Fe2+ in paddy soils after flooding ofmore than 10 mM(Li and Horikawa, 1997; Kirk, 2004) indicate the potential role of thisprocess for DOC retention under anaerobic conditions.
Whenever sorption or precipitation of DOM occurs, it is usuallyselective to hydrophobic and aromatic compoundswith largemoleculesand a complex structure (Kaiser et al., 1996; Dolfing et al., 1999;Sharpless and McGown, 1999; Scheel et al., 2007). These are the moststable DOMcomponents againstmicrobial decay (Kalbitz et al., 2003a,b),indicating the potential of organo-mineral interactions to form stableSOM (Kögel-Knabner et al., 2008). Katoh et al. (2004), for instance,estimated retention of C up to 120 kg C ha−1 during a 4-month rice-growth period, representing 0.6% of organic C stored in this subsoilhorizon.
Effects of frequent changes in redox conditions on the dynamics ofDOM are poorly understood, although very common in paddy soils.Short-term oscillations in Fe reduction and oxidation resulted inincreasing mobilization of colloidal bound C (Thompson et al. 2006a)and, on the other hand, in an increasing crystallinity of the Feoxyhydroxides formed (Thompson et al. 2006b). These Fe oxyhydr-oxides are less susceptible to anaerobic dissolution, maintaining theircapacity to retain and stabilize DOM even under anaerobic conditions.
In summary, concentrations and fluxes of DOM are large in paddysoils in comparison to other soils. There are many indications thatthese large DOM fluxes trigger the emission of greenhouse gases fromthe topsoil and the formation of stable SOM in the subsoil. As inupland soils, DOM stabilization closely depends on the soil mineralmatrix. It is still unclear whether Al hydroxides or clay minerals cancompensate the expected dissolution of Fe oxyhydroxides underanaerobic conditions. Oscillations in redox conditions might enhanceretention and stabilization of DOM by Fe oxyhyrdoxides. Thishypothesis should be tested by studying interactions between redoxconditions, pH driven solubility of organic matter, mineral propertiesand stabilization of organic matter against microbial decay.

7I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
4. Specific characteristics of long-term biogeochemical processesin paddy soils
4.1. Mineral weathering and (neo)transformation
Since paddy soils can develop from different parent materials andsoil units, their chemical composition (Chandrajith et al., 2005; Egashiraet al., 1997), mineral assemblies (Mizota et al., 1982) and propertiessuch as texture (Prakongkep et al., 2008) can vary considerably.Hassannezhad et al. (2008) and Sano et al. (2006) postulated that claymineralogy is probablymore affected by parentmaterials than by redoxcycling. It is often observed that permanent strong reducing conditionseffectively slow down mineral weathering in iron-poor silicatic soils.However, there is some evidence that the alternating redox conditionsmay have an effect on mineral weathering and transformation. Oncemore, paddy soils contain numerous specific minerals such as siderite,jarosite and vivianite (Kyuma, 1978), phyto opal (Gong et al. 2007;Wickramasinghe and Rowell, 2006) and different types of green rust(Quantin et al., 2008).
Paddy soils originating from soils that are high in potassium-bearingminerals are characterized by strong changes in clay minerals due tomarked depotassification (Gong, 1983; Gong and Xu, 1990) and thepreferred formation of less siliceous, more ferro-magnesian chlorite (Liet al., 2003). In this case, the alternation between reductive andoxidative conditions in paddy soil induces the chloritisation ofexpanding 2:1 type minerals (Kyuma, 1978). Gong and Xu (1990)found that in paddy soils developed from calcareous purplish soils thecontent of hydromicas decreases and the content of vermiculiteincreases in contrast to the former soil.
Fe3+ and Fe2+ iron species occur either in pure or mixed form in awide range of primary and secondary soil minerals. A change in thestructural iron status leads to an imbalance in the overall electrostaticcharge of the crystal structure. Favre et al. (2002) investigated thechanges in structural iron in a flooded Vertisol containing smectiteand kaolinite as dominant clay minerals and managed with riceculture twice a year for 11 years. They found that structural smectiteFe2+ increased upon reduction but, nevertheless, the Fe contents ofthe rice field Ap horizon were less than those of soil in uncroppedneighbouring land, suggesting that inundation induced weatheringand eluviation of the minerals. The authors estimated that 0.4 cmolstructural iron kg−1 smectite clay is lost from the Ap horizons in eachcrop cycle. This figure equals amass loss of more than 10% if calculatedfor a period of one thousand years, which is, however, not reasonablefor soils from relatively iron-poormaterials. This suggests that the rateof iron loss decreases over time.
Upon the increasing availability of oxygen when the fields aredrained, the rapid oxidation of Fe2+ leads to a change in soil solutionchemistry (Ahmad and Nye, 1990; Kirk and Solivas, 1994; Kirk, 2004).Fast oxidation of Fe2+ is accompanied by a decrease in ionic strength.Frequently, the incipient oxidation product is expected to be a shortrange-ordered hydroxide such as ferrihydrite (Thompson et al., 2006a).Boivin et al. (2004) speculated that most iron (hydr)oxides in paddysoils are lepidocrocite. However, Thompson et al. (2006b) observed anincreasing crystallinity of iron oxides during soil redox alternation(200–700 mV) in short-termbatch experiments (56 days). The authors'findings fit verywell with the observation that (1) DCB-extractable iron(Fed) and its ratio with total iron (Fet) (i.e. Fed/Fet) increase with thelength of paddy cultivation (Zhang andGong, 2003), and (2) the total Fecontent is higher in paddy soils than in non-paddy soils (Cheng et al.,2009).Additionally, hydragric conditions seemto inhibit Al-substitutionin goethite, due to higher Fe-activity (Wang et al., 1992).
The translocation of iron in various directions and by differentprocesses (including eluviation and illuviation) is characteristic forpaddy soils, due to the high iron mobility during the flooding season(Gong, 1986). Eluviation of iron upon reduction is not obvious in youngpaddy soils but apparent in old ones (Gong and Xu, 1990), where
downward leaching and successive re-precipitation accounts for theformation of an Fe-rich illuvial horizon.
Specific to paddy soils is the accumulation of silica in phyto opal(phytoliths), depending on the amount of crop residues remaining inthe field (see below). Silica is also incorporated to a greater extent intoallophans in paddy soils formed from volcanic ash. This leads to anincreasing SiO2/Al2O3 molar ratio of the clay fraction (Kanno et al.,1957 cit. in Mizota et al., 1982) and ‘free’ oxides (Mitsuchi (1970), cit.in Mizota et al. (1982)). As a product of resilication resulting from thelong-term cultivation of rice, allophone can be transformed intohalloysite and metahalloysite. Aquic and anthraquic conditions inpaddy soils that develop on alluvial material enhance the formation ofsmectite (Hassannezhad et al., 2008).
Poor drainage and flooding during the rainy season might make apaddy soil prone to ferrolysis (Brinkman, 1978). This process assumesthat repeated redox and leaching cycles lead to strong acidicconditions and excessive weathering of clays, and consequently to avery small cation exchange capacity (CEC) of soil minerals in thetopsoils. However, many authors do not believe ferrolysis to be thedominant process in the genesis of paddy soils, because small CEC,high pH and low quantities of aluminium oxides can be found inmanypaddy soils, e.g. caused by addition of silty material with the irrigationwater, which makes ferrolysis very unlikely (Eaqub and Blume, 1982;Van Ranst and De Coninck, 2002). Due to the addition of materialswith the irrigation water and eluvial processes, it is in general difficultto trace back mineral changes of horizon sequences to weatheringprocesses.
4.2. Mineral transformation in the presence of organic matter
Suetsugu et al. (2005) observed a strong increase of both dissolvedand particulate organic matter during flooding seasons. High concen-trations of DOM favour the formation of aqueous organic Fe3+
complexes and support the formation of ferrihydrite (Ratering andSchnell, 2000; Rancourt et al., 2005). Many studies suggest thatoctahedral Fe3+ in the crystal structures of clayminerals can be reducedto clay by indigenous microbes (McKinley et al., 2000; Ueshima andTazaki, 2001; Li et al., 2004). The abundance of electron donors (DOM)should actually enhance the changes of structural iron in clay minerals.We can expect a direct link between the biodegradation of organicmatter associated with soil minerals and the status of iron in minerals.On the other hand, it can also be assumed that the close coupling oforganic matter dynamics to the biogeochemical interfaces in paddiessupportsdifferent stabilizationmechanisms involved inC sequestration,thereby also affecting the quality of soil OM.
4.3. Accumulation and stabilization of soil organic matter
Paddy soils are characterized by large carbon input via organicfertilizers and plant residues (Gong and Xu, 1990; Tanji et al., 2003). It iscommonly accepted that water logging associated with rice croppingenhances accumulation of SOC (Neue et al., 1997; Lal, 2002). Butaccording to Kirk (2004)wetland rice soils do not have particularly highorganic matter contents. The mean level of organic carbon in the topsoils of lowland rice soils ranges from 20 g kg−1 (tropical Asia,Greenland, 1997 rf. Kirk, 2004) to 29 g kg−1 (Japan; Shirato, 2005)and 27–41 g kg−1 in the Yangtze River Valley of China (Cao and Zhang,2004; Lin et al., 2004; Pan et al., 2008).
Rates of decomposition of added organic materials as well as ofsoil organic matter are considered to be slower under anaerobicconditions than under aerobic conditions, leading to a relativelygreater tendency to accumulate organic matter. High losses of soilorganic carbon (SOC) in rice–wheat systems contrast with resultsfrom continuous crops of wetland rice, due to differences in organicmatter decomposition pattern and products under more continuousmaintenance of anaerobic conditions (Sahrawat, 2004). Olk et al.

8 I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
(1996) report higher contents of OC in plots with higher intensity ofirrigated wetland rice cropping in a long-term experiment at theInternational Rice Research Institute in The Philippines. Li et al.(2005b) and Zhang and He (2004) reported that long-term ricecropping resulted in significantly increased OC and N contents in theplough layer. In contrast, Kirk (2004) points out that also the SOCcontents developing under rice-rice systems are not large.
Accumulation of OC and N in paddy subsoil is described by Li et al.(2005b) and Zhang and He (2004). The authors suggest that thesubsoil OC originates from downward movement of DOC or colloidalorgano-mineral associations, but do not provide any evidence forsuch processes. Results fromMaie et al. (2004) show that the subsoilOM of paddy soils originates at least partly from leached DOM,released from the plough layer. Part of this material seems to be lowmolecular weight compounds that are stabilized by interaction withiron oxides.
High bomb-14C inputs to larger depths and low C residence timesof C indicate rapid C cycling in paddy soils (Scharpenseel et al., 1996).The decomposition pattern of labelled rice straw was not different inpermanently submerged rice soils compared to aerobic soils in anexperiment conducted by Neue and Scharpenseel (1987) over 4 years.The authors pointed out that this is inconsistent with the hypothesisthat submergence generally retards decomposition because of lowerenergy yields of anaerobic fermentation, which is considered themajor factor controlling the rate of OM decomposition in anaerobicenvironments by Sahrawat (2004). Neue et al. (1997) conclude thatincreased SOC contents in tropical wetland soils are most probablycaused by increased OM inputs rather than retarded decomposition.This is supported by the four-year field study of Yang et al. (2005) inwhich continuous waterlogging increased SOC contents in paddysoils, especially under combined application of fertilizers andfarmyard manure or wheat straw. Cheng et al. (2009) found higherC contents in a paddy soil chronosequence established for severalhundred years under a rice/non-rice cropping system compared toupland soils in the same region. Zhang and He (2004) reported asignificant increase in organic carbon in the plough layer, up to 2%after 100 years of rice cropping, and a concomitant increase of OCcontents in the subsoil. The increasing C content after implementationof paddy soils could not be seen in the particulate organic C fraction(Zhang and He, 2004), i.e. different preservation mechanisms thanjust the mere accumulation of plant debris seem to be responsible forthis.
Sahrawat (2004) suggests that the accumulation of OM in paddysoils is due to the lack of oxygen underwaterlogging combinedwith thelack of other terminal electron acceptors, especially Fe3+ and SO4
2− thatdevelops with time in paddy soils. With these electron acceptorsmissing, no efficient OM oxidation can be performed at low Eh values,i.e. the destruction of organic materials slows down in submerged soilsand sediments. If this process is dominant, all plant-derived structuresaccumulate. As rice straw decomposes much faster than root residuesand especially the intact rice root system (Lu et al., 2003), theaccumulation of root-derived plant residues in particular is expected.And indeed, wetland rice cropping resulted in an increase in theproportion of less humified organic material in SOM, as revealed by theabundance of diester P, amide N and phenolic C in NMR spectra (Olket al., 1996, 1998, 1999, 2000; Mahieu et al., 2000b, 2002) in differenthumic acid extracts. Lignin is a peculiar tracer for plant residues in soils.The accumulation of lignin residues in paddy soil humic acids wasshown by pyrolysis-GC–MS and solid-state 13C NMR spectroscopicanalysis (Olk et al., 2002; Smernik et al., 2004). Therewas also a positivecorrelation with visible light absorbance. The authors suggest that slowlignin decomposition under restricted O2 supply in submerged soilsleads to incorporation of phenolic compounds into young SOM. Theaccumulation of lignin is promoted by crop intensification. In thesestudies a high intensity of rice cropping was generally associated withlarger proportions of less humified material (Olk et al., 1998).
It seems that the high inputs of organic matter in paddy soils favouraccumulationofOC. But it remains to be evaluated in detailwhether thisis due tohighorganicmatter inputs or to lowdecomposition rates underanoxic conditions.
Only few studies have investigated the mechanisms responsible forlong-term storage of OC (Neue et al., 1997; Kimura et al., 2004; Kirk,2004; Scharpenseel et al., 1996). The mechanisms suggested as beingresponsible for the accumulation of SOM in paddy soils are occlusion inaggregates, formation of organo-mineral associations, addition ofpyrogenic OM and phyto-opal associated stabilization of OC.
Although aggregationmight be considered to be of little importanceespecially in regularly puddled topsoil horizons (Kirk, 2004), severalreports have described the stabilization of SOM fractions associatedwith aggregation, i.e. occluded particulate organic matter. Indeed, it hasbeen found that paddy soils show a specific aggregation; aggregate sizere-effects redox potential and release of CH4 (e.g. Ramakrishnan et al.,2000, Tripathy and Singh, 2004). Zhang and He (2004) and Yang et al.(2005) found more water-stable aggregates in paddy soils withalternate wetting and drying water regime when supplied regularlywith OM amendments. Iron bridges add to the binding of particles toform aggregates. Lu et al. (1998) found a greater aggregate stability andhigher OC contents in Andosols after prolonged use as paddy comparedto soils under upland field or grassland management. They attributedthis specifically to high amounts of water-soluble carbohydrates.Saejiew et al. (2004) pointed out that flocculation of clay is supportedby the adsorption of Fe2+ under reduced conditions. These resultssuggest that occlusion into aggregates is a relevant process for OCstabilization in paddy soils. It remains to be investigated whether thehigh amounts of partly degraded plant residues found in many paddysoils are stabilized by occlusion in aggregates, and whether the conceptof aggregate hierarchy, commonly accepted for many aerobic soils(Oades and Waters, 1991), is valid for paddy soils.
Scharpenseel et al. (1996) considered adsorption to clayminerals tobe one of themajormechanisms of carbon storage in paddy soils. On theother hand, they mention a reduced formation of clay/oxide-humuscomplexes that is supposedly due to thepuddlingof the top soil. Nguyenet al. (2004) suggest that the susceptibility of OM to mineralization inpaddy soils is influenced by exchangeable cations, with Ca2+ respon-sible for a stronger binding of OM to soil surfaces than K+. Pan et al.(2003) suggest a stabilizing effect by iron cutans in paddy soils,indicated by good correlations between oxalate-extractable iron andSOC content. These differences are most probably due to different soilpedogenetic environments and need further investigation.
Post-harvest burning represents a common paddy soil manage-ment practice, needed to remove harvest residues and to controlweeds, insects and (plant) pathogenic microbes for the next ricecropping sequence. Studies in Japan have shown that this stubble androot material can amount to 600 to 2250 kg dry weight ha−1 (Kimuraet al., 2004). Because charring converts labile organic components ofplant remains, i.e. carbohydrates or amino acids, into highly aromaticand heteroaromatic forms (Almendros et al., 2003), these charredresidues (e.g. straw ash returned as a potassium source in China)exhibit an increased resistance against microbial degradation; andaromatic structures may prevail in the SOM of old paddies (Cao et al.,2006).
Rice belongs to a plant group known to accumulate silica in formingphytoliths (10 to 15% SiO2 of shoot dry weight; Marschner, 1995).Phytoliths are silicified structures formed from monosilicic acid (Si(OH)4) in soil solution taken up by plant roots and being subsequentlydeposited in the intra- and extra-cellular compartments throughouttheir leaf, stem, and root systems (Wilding, 1967; Parr and Sullivan,2005). The critical acid-extractable-Si concentration for plant growth inthe soil is 40 mg kg−1 (measured in1-Msodiumacetate buffered at pH4;Dobermann and Fairhurst, 2000). Depending on the amount of cropresidues remaining in the field, large amounts of phytoliths canaccumulate in soil (Fig. 4) (Wickramasinghe and Rowell, 2006). Wilding

Fig. 4. Rice phytoliths in a pedo complex, used as paddy soil in prehistoric, ancient and recent times (provided by Y.F. Zheng, Zhejiang Provincial Institute of Cultural Relics andArchaeology, Hangzhou 310014 (PR China)).
9I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
et al. (1967) reported phytoliths to hold up to 5% OC. Since it is highlyresistant to decomposition, phytolith OC can be a substantial componentof SOC, especially in subsoils (up to 80% of SOC; Parr and Sullivan 2005).The OC in different grass phytoliths, including Oryza sativa, is dominatedby O-alkyl C structures (55%), but also contained alkyl C (around 30%;Krull et al., 2003). The amount and type of OC associated with phytolithsin paddy soils has not yet been investigated in further detail.
The morphology of phytoliths is highly characteristic for plantspecies. Thus, phytoliths have beenused as indicators for rice cultivationin ancient paddy soils (e.g. Huang and Zhang, 2000; Gong et al., 2007).
Contrasting observations have been made on the solubility ofphyto opal. The solubility of biogenic opal in grasses is relative high,almost equivalent to that of amorphous silica (Wilding et al., 1979;Wickramasinghe and Rowell, 2006). However, opal in soils reactswith iron and aluminium complexes, which reduces its solubility to 10to 15 mg L−1 (Wilding et al., 1979). In addition, other factors such astemperature, pH, redox conditions, other inorganic anions and OMalso influence the dissolution of Si (Sommer et al., 2006). In contrastto the observations of Wilding et al. (1979), Wickramasinghe andRowell (2006) thus observed high dissolution rates of phyto opal fromrice straw in batch experiments. Therefore, it remains unclear to whatextent phytoliths either are preserved in soil over long periods(thousands of years) or serve as Si sources for plant growth. Anotheropen question is the biodegradability of organic matter trapped inphytoliths.
Overall, it is not clear whether paddy soils accumulate more SOMthan comparable upland soils. This is most probably due to the factthat paddy soils have a large variability in properties as they developfrom different parent materials and soil types that are converted topaddy management. The accumulation of SOM may be due to highinput of plant residues, probably associated with retarded decompo-sition of these residues under anaerobic conditions. Stabilizationprocesses of OM in paddy soils remain to be investigated in detail. Allaccumulation mechanisms, driven by retarded decomposition, chem-ical recalcitrance (e.g. likely for paddy-chars) and specific organo-mineral associations and inclusions are not well understood for paddysoils. Among the latter, phyto opal may play a special role in the long-term stabilization of C in paddy fields. The inconsistencies in ourunderstanding of C stabilization in paddy soils also lead to a lack ofsuitable models for C turnover in these soils (Sahrawat, 2004).
4.4. Soil organic N in paddy fields
Decomposition of the organic matter in paddy fields is not solelyimportant for the gross C balance and trace gas emissions. In naturalwetlands and minimally fertilized paddy fields, N release fromdecomposing OM is one of the crucial parameters determining biomassproduction and crop yield (Olk et al., 1996). Olk et al. (1996) reportedthat a small N utilization efficiency also leads to significant grain yielddeclines of more than 35% during 20–30 a of double and triple croppedrice soils. Despite declining yields, the total soil nitrogen contentremained more or less constant over the years. The authors concludedthat the bioavailability of the soil organic N (SON) had declined. Themechanisms behind this are just as unclear as those for bulk SOC.
Olk and Co-workers (e.g., Olk et al., 2006) published a number ofpapers on the fate of nitrogen in submerged rice soils. The majority ofthe SON detected by 15 N NMR spectroscopy was in amide forms(Mahieu et al., 2000a). This finding suggests that, similar to other soils,proteins make up the dominant SON fraction, i.e. changes in SONbioavailability might be reflected by changes within the soil proteinpool. Such proteins may survive in soils for prolonged periods of timeunder oxic conditions and thus certainly even longer undersubmerged ones (Amelung, 2003; Amelung et al., 2006).
Using two-dimensional nuclear magnetic resonance techniques,Schmidt-Rohr et al. (2004) found that significant amounts of theamide nitrogen were directly bound to aromatic rings in the paddysoils. As they also observed that the same soils were enriched inlignin, they concluded that N bound to lignin accounts for thereduced bioavailability of N. It was one of the first chemicalexplanations for the reduced N availability under waterloggedconditions. Little is known about other mechanisms that maypotentially contribute to reduced N supply but have not been studiedin paddy fields. Such additional mechanisms may include bothphysical encrustation of SON within other organic molecules (e.g.Knicker et al., 1996), iron-organo-mineral associations (microag-gregates; Ladd et al., 1993; Kögel-Knabner et al., 2008), andsequestration of N within microbial cell wall residues (e.g. Amelunget al., 2001). The latter are unfavourable for further decay,mainly dueto chemical recalcitrance from D-alanine in bacterial cell walls (Voetand Voet, 1995), encrustation of fungal chitin by melanin (Parsons,1981), or again by the close interaction of microbial residues with

10 I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
mineral matter (Ladd et al., 1993; Zhang et al., 1998; Amelung et al.,1999; see also discussion above for bulk SOM).
There is increasing evidence that all kinds of SON polymers mustbe broken down extracellularly to their monomeric units before beingcapable of microbial assimilation (Yang et al, 2006; Roberts et al.,2007). The concentration of amino acid-N strongly correlatedwith themineralizable N under waterlogged conditions (Mohapatra and Khan,1983), and concentrations of hydrolysable amino acids were higher inthe surface than in the subsurface soil and dominated by neutralstructures (Gotoh et al., 1986). Small proteins and free amino acidscan apparently even be taken up actively by (upland) rice plants(Yanamuro et al., 1999; Yamagata, 2000). The protein fraction may bebound to organic and inorganic fractions and, hence, be preserved.The underlying mechanisms are unclear, since both the acidic aminoacids (due to higher sorption strength of the carboxylic group) andbasic amino acids (due to higher intrinsic stability and possiblyadditional binding of cationic amide forms to clays) may be preservedin preference to other structures (e.g. Stevenson, 1982; Kaiser andZech, 2000). A significant proportion of the soil protein fraction mayalso not just be preserved, but taken up and sequestered or recycledby microorganisms. The high abundance of microbial biomass andmicrobial N containing fragments like DNA (e.g. Pande and Becker,2003; Cayhani et al., 2009; Gu et al., 2009) gives support to thehypothesis that microbes could well contribute to the storage of N inorganic forms.
To improve the N balance, organic amendments like wheat strawmay help to immobilize the liberated N in better developing microbialbiomass (Pande, 2005; Pande and Becker, 2003). If combined with Nfixation, e.g. by intercropping with green manure legumes, a positiveN balance has even been detected. Also organic fertilization withgreen manures like white clover and hairy vetch helped to improvethe N supply, with higher N use efficiency for the incorporatedfertilizers than for surface applied manures (Asagi and Ueno, 2009).However, to sustain crop growth it must also be assured that theadditional N is not sequestered in unavailable N forms bymechanismsoutlined above. As Olk et al. (2007) showed, nitrogen mineralizationwas inhibited in rice–rice rotation with anaerobic decompositioncompared with better-aerated treatments. The authors thus suggestincreased aeration of rice soils through aerobic decomposition of cropresidues or crop rotation as a promising management technique forimproving soil N supply in lowland rice cropping. Increased aeration,though, also induces the risk of water losses and crack formation. Forthis reason Lennartz et al. (2009) had suggested maintaining(saturated) equilibrium conditions over longer periods to reducesoil structural dynamics and the risk of water losses and chemicalleaching involved with preferential flow. This management in turn re-includes the risk of N losses as outlined above, and it may increase therisk of metal and As uptake under reduced conditions. Clearly, anytype of paddy management has beneficial and adverse effects onenvironment. Optimizing the first while minimizing the latter fordifferent soil properties and climates is still an urgent research goal forimproving the sustainability of the world's rice production.
5. Conclusions
Rice, mostly as paddy, grows on about 155 million ha of theworld's surface and paddy fields are therefore the largest man-madewetlands on earth. Paddy soils comprise diagnostic features of long-term anthropogenic flooding and puddling, with dense Arpd horizonscharacterized by microscaled Eh gradients in the rhizosphere above areduced or aerobic Bg or C horizon. Paddy soils develop in a uniquemanagement system that controls redox driven processes affectingmineral transformation and microbially mediated turnover of organicmatter. The vertical and horizontal redox gradients formed in paddysoil have a considerable effect on bacteria, fungi, and archaea in thosemicrohabitats.
In contrast to theoftenobserved fact thatpermanent strong reducingconditions effectively slow downmineral weathering, alternating redoxconditions occur in paddy soils and, depending onmineral assemblages,have different consequences for mineral weathering. Redox processestrigger the transformation of minerals and are responsible for themobility of elements. Potassiumand structural iron-containingmineralsalter faster under alternating redox conditions. Translocation of iron(and other redox sensitive elements) in various directions is character-istic for paddy soils and particularly increases the crystallinity of ironoxides. High concentrations of DOM during flooding seasons enhancethe release of structural iron in clayminerals, and support the formationof ferrihydrite. This also results in higher total iron oxide contents inpaddy compared to non-paddy soils. Since the existing studies aremostly from laboratory experiments underwell controlled conditions, itis rather difficult to derive figures for weathering and transformationrates over time. Differences in the mineral assemblage from parallelobservations on chronosequences (false time series) of paddy and non-paddy soils may partly result from differences of the parentmaterials orfrom the addition of materials with the irrigation water to paddy soils.
In paddy soils, concentrations and fluxes of DOM from the A intothe B horizon are larger than in other soils. The subsequent fate of DOMand associated nutrients depends on the Eh conditions of the subsoil.Since the abundance of electron donors (from SOM and DOM)enhances the changes of structural Fe3+ in clay minerals (e.g.nontronite) and iron oxyhydroxides, we expect a direct link ofbiodegradation of organic matter associated with soil minerals andthe status of iron in minerals. It is not known whether this change ofiron status in minerals is more intensive under alternating redox orunder permanent reducing conditions, because it is not clear to whatextent biodegradation of organic matter is affected by the dynamics ofredox processes. Many studies show that SOM decomposition and theformationof stable SOMproceed at a slower rate in hydromorphic soilsthan inwell-drained, aerated soils. However, there are also indicationsthat organicmatter decomposition is not retarded under the anaerobicconditions prevailing in paddy soils. OM accumulation in paddy soilsalso derives from high input of residues under intensivemanagement,partly also in the form of charred residues. The amount of organicmatter stabilized in paddy soils due to aggregation and associationwith minerals still has to be investigated. Due to high silica demand ofrice plants, the cycling of silicon is a special feature in paddy soils. Thusphytoliths may play an important role for carbon stabilization, but itremains to be investigated whether carbon trapped in phytoliths isavailable to microbial attack or not. The solubility of phytoliths is thecrucial property for the fate of OC trapped there.
Interactions between redox potential, pH, solubility of organicmatter and its biodegradation seem to play a decisive role for thebiogeochemistry of paddy soils. The availability of OC for microbialdegradation is of special interest, because in wetland soils such asflooded rice fields the anaerobic fermentation of SOM leads to therelease of methane and to denitrification losses of N. Tominimize suchlosses and to optimize paddy management in terms of environmentalsustainability, we need a quantitative and mechanistic understandingof the coupling of organic matter, N, and mineral transformations andfluxes and how they are regulated by redox potential, degree of soildevelopment and microbial community structure and function.
Acknowledgements
We thank two anonymous reviewers for their helpful commentsthat significantly improved the manuscript.
References
Ahmad, A.R., Nye, P.H., 1990. Coupled diffusion and oxidation of ferrous iron in soils. I.Kinetics of oxygenation of ferrous iron in soil suspension. J. Soil. Sci. 41, 395–409.

11I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
Almendros, G., Knicker, H., Gonzalez-Vila, F.J., 2003. Rearrangement of carbonandnitrogenforms in peat after progressive thermal oxidation as determined by solid-state C-13and N-15-NMR spectroscopy. Org. Geochem. 34, 1559–1568.
Amelung, W., 2003. Nitrogen biomarkers and their fate in soil. J. Plant Nutr. Soil Sci. 166,677–686.
Amelung, W., Flach, K.-W., Zech, W., 1999. Neutral and acidic sugars in particle-sizefractions as influenced by climate. Soil Sci. Soc. Am. J. 63, 865–873.
Amelung,W., Miltner, A., Zhang, X., Zech,W., 2001. Fate of microbial residues during litterdecomposition as affected by minerals. Soil Sci. 166, 598–606.
Amelung, W., Zhang, X., Flach, K.W., 2006. Amino acids in grassland soils: climatic effectson concentrations and chirality. Geoderma 130, 207–217.
Armstrong, W., 1971. Radial oxygen losses from intact rice roots as affected by distancefrom the apex, respiration and waterlogging. Physiol. Plant. 25, 192–197.
Arth, I., Frenzel, P., 2000. Nitrification and denitrification in the rhizosphere of rice: thedetection of processes by a newmultichannel electrode. Biol. Fert. Soils 31, 427–435.
Arth, I., Frenzel, P., Conrad, R., 1998. Denitrification coupled to nitrification in therhizosphere of rice. Soil Biol. Biochem. 30, 509–515.
Asagi, N., Ueno, H., 2009. Nitrogen dynamics in paddy soil applied with various N-15-labelled green manures. Plant Soil 332, 251–262.
Aulakh, M.S., Wassmann, R., Rennenberg, H., Fink, S., 2000. Pattern and amount ofaerenchyma relate to variable methane transport capacity of different rice cultivars.Plant Biol. 2, 182–194.
Aulakh, M.S., Wassmann, R., Rennenberg, H., 2001. Methane emissions from rice fields —quantification, mechanisms, role of management, and mitigation options, pp. 193–260.
Aulakh,M.S., Wassmann, R., Rennenberg, H., 2002. Methane transport capacity of twenty-two rice cultivars from fivemajor Asian rice-growing countries. Agr. Ecosyst. Environ.91, 59–71.
Barnes, G.L., 1990. Paddy soils now and then. World Archaeol. 22, 1–17.Begg, M.C.B., Kirk, G.J.D., MacKenzie, A.F., Neue, H.-U., 1994. Root-induced iron oxidation
and pH changes in the lowland rice rhizosphere. New Phytol. 128, 469–477.Bilek, R.S., Tyler, S.C., Sass, R.L., Fisher, F.M., 1999. Differences in CH4 oxidation and
pathways of production between rice cultivars deduced frommeasurements of CH4
flux and delta C-13 of CH4 and CO2. Global Biogeochem. Cy. 13, 1029–1044.Bodelier, P.L.E., Frenzel, P., 1999. Contribution ofmethanotrophic and nitrifying bacteria to
CH4 andNH4+ oxidation in the rice rhizosphere using newmethods for discrimination.
Appl. Environ. Microb. 65, 1826–1833.Bodelier, P.L.E., Hahn, A.P., Arth, I.R., Frenzel, P., 2000a. Effects of ammonium-based
fertilisation on microbial processes involved in methane emission from soils plantedwith rice. Biogeochem. 51, 255–257.
Bodelier, P.L.E., Roslev, P., Henckel, T., Frenzel, P., 2000b. Stimulation by ammonium-basedfertilizers of methane oxidation in soil around rice roots. Nature 403, 421–424.
Bodelier, P.L.E., Sorrell, B., Drake, H.L., Küsel, K., Hurek, T., Reinhold-Hurek, B., Lovell, C.,Megonigal, P., Frenzel, P., 2006. Ecological aspects of microbes and microbialcommunities inhabiting the rhizosphere of wetland plants. In: Bobbink, R., Beltman,B., Verhoeven, J.T.A., Whigham, D.F. (Eds.), Wetlands as a Natural Resource. Vol. 2.Ecological Studies, vol. 190. Springer-Verlag, New York, pp. 205–238.
Boivin, P., Favre, F., Hammecker, C., Maeght, J.L., Delarivère, J., Poussin, J.C., Wopeis, M.C.S.,2002. Processes driving soil solution chemistry in a flooded rice-cropped Vertisol:analysis of long-term monitoring data. Geoderma 110, 87–107.
Boivin, P., Saejiew, A., Grunberger, O., Arunin, S., 2004. Formation of soils with contrastingtextures by translocation of clays rather ferrolysis in flooded rice fields in NortheastThailand. Eur. J. Soil Sci. 55, 713–724.
Bosse, U., Frenzel, P., 1998. Methane emissions from rice microcosms: the balance ofproduction, accumulation and oxidation. Biogeochem. 41, 199–214.
Bosse, U., Frenzel, P., Conrad, R., 1993. Inhibition of methane oxidation by ammonium inthe surface layer of a littoral sediment. Fems Microbiol. Ecol. 13, 123–134.
Bouman, B.A.M., Humphreys, E., Tuong, T.P., Barker, R., 2006. Rice and Water. Adv. Agron.92, 187–237.
Bouwman, A.F., 1991. Agronomic aspects of wetland rice cultivation and associatedmethane emissions. Biogeochemistry 15, 65–88.
Bravin, M.N., Travassac, F., Floch, M.L., Hinsinger, P., Granier, J.-M., 2008. Oxygen inputcontrols the spatial and temporal dynamics of arsenic at the surface of afloodedpaddysoil and in the rhizosphere of lowland rice (Oryza sativa L.): a microcosm study. PlantSoil 312, 207–218.
Brinkman, R., 1978. Ferrolysis: chemical and mineralogical aspects of soil formation inseasonally wet acid soils, and some practical implications. Soil & Rice, I.R.R. Institute,ed., Los Banos, Philippines, 295-303.
Cai, Z.C., Tsuruta, H., Rong, X.M., Xu, H., Yuan, Z.P., 2001. CH4 emissions from rice paddiesmanaged according to farmer's practice in Hunan, China. Biogeochem. 56, 75–91.
Cai, Z.C., Tsuruta, H., Gao, M., Xu, H., Wei, C.F., 2003. Options for mitigating methaneemission from a permanently flooded rice field. Global Change Biol. 9, 37–45.
Cai, Z.C., Kang, G.D., Tsuruta, H., Mosier, A., 2005. Estimate of CH4 emissions from year-round flooded rice fields during rice growing season in China. Pedosphere 15,66–71.
Cao, Z.H., Zhang, H.C., 2004. Phosphorus losses to water from lowland rice fields underrice-wheat double cropping system in the Tai Lake region. Environ. Geochem. Hlth.26, 229–236.
Cao, Z.H., DeDatta, S.K., Fillery, I.R.P., 1984a. Effect of placement methods on floodwaterproperties and recovery of applied nitrogen 15 N labeled urea in wetland rice. Soil Sci.Soc. Am. J. 48, 196–203.
Cao, Z.H., DeDatta, S.K., Fillery, I.R.P., 1984b. 15Nitrogenbalance and residual effects of urea-N in wetland rice fields as affected by deep placement techniques. Soil Sci. Soc. Am. J.48, 203–208.
Cao, Z.H., Ding, J.L., Hu, Z.Y., Knicker, H., Kögel-Knabner, I., Yang, L.Z., Yin, R., Lin, X.G., Dong,Y.H., 2006. Ancient paddy soils from the Neolithic age in China's Yangtze River Delta.Naturwissenschaften 93, 232–236.
Cayhani, V.R.,Murase, J., Ishibashi, E., Asakawa, S., Kimura,M., 2009. Phylogenetic positionsof Mn2+-oxidizing bacteria and fungi isolated from Mn nodules in rice field subsoils.Biol. Fert. Soils 45, 337–346.
Chandrajith, R., Dissanayake, C.B., Tobscholl, H.J., 2005. The abundances of rarer traceelements in paddy (rice) soils of Sri Lanka. Chemosphere 58, 1415–1420.
Chen, S.-K., Liu, C.-W., 2002. Analysis of water movement in paddy rice fields (I)experimental studies. J. Hydrol. 260, 206–215.
Chen, X.P., Zhu, Y.G., Xia, Y., Shen, J.P., He, J.Z., 2008. Ammonia-oxidizing archaea:important players in paddy rhizosphere soil? Environ. Microbiol. 10, 1978–1987.
Cheng, W., Yagi, K., Sakai, H., Xu, H., Kobayashi, K., 2005. Changes in concentration and13C value of dissolved CH4, CO2 and organic carbon in rice paddies under ambientand elevated concentrations of atmospheric CO2. Org. Geochem. 36, 813–823.
Cheng, Y.-Q., Yang, L.-Z., Cao, Z.-H., Yin, S., 2009. Chronosequential changes of selectedpedogenic properties in paddy soils as compared with non-paddy soils. Geoderma151, 31–41.
Conrad, R., 2007. Microbial Ecology of Methanogens and Methanotrophs. In: Donald, L.S.(Ed.), Advances in Agronomy. Academic Press, pp. 1–63.
Conrad, R., Frenzel, P., 2002. Flooded soils. In: Britton, G. (Ed.), Encyclopedia of Environ.Microbiol. John Wiley & Sons, New York, pp. 1316–1333.
Conrad, R., Rothfuss, F., 1991. Methane oxidation in the soil surface layer of a flooded ridefield and the effect of ammonium. Biol. Fert. Soils 12, 28–32.
DeDatta, S.K., 1981. Principles and Practices of Rice Production. John Wiley, New York.Denier van der Gon, H., 2000. Changes in CH4 emission from rice fields from 1960 to
1990s–1. Impacts of modern rice technology. Global Biogeochem. Cycles 14, 61–72.Denier van der Gon, H., Neue, H.U., 1996. Oxidation of methane in the rhizosphere of
rice plants. Biol. Fert. Soils 22, 359–366.Denier van der Gon, H., van Breemen, N., 1993. Diffusion-controlled transport of methane
from soil to atmosphere as mediated by rice plants. Biogeochemistry 21, 177–190.Dobermann, A., Fairhurst, T., 2000. Rice: Nutrient disorders and management. Potash
and Phosphate, Singapore: Handbook series.Dolfing, J., Chardon, W.J., Japenga, J., 1999. Association between colloidal iron, aluminum,
phosphorus, and humic acids. Soil Sci. 164, 171–179.Eaqub, M., Blume, H.-P., 1982. Genesis of a so-called ferrolysed soil of Bangladesh. Z.
Pflanzenernähr. Bodenk. 145, 470–482.Egashira, K., Fujii, K., Yamasaki, S., Virakornphanich, P., 1997. Rare earth elements and clay
minerals of paddy soils from the central region of Mekong River, Laso. Geoderma 78,237–249.
Eickhorst, T., Tippkötter, R., 2009. Management-induced structural dynamics in paddysoils of south east China simulated in microcosms. Soil Till. Res. 102, 168–178.
Eller, G., Frenzel, P., 2000. Population dynamics and activities of rhizospheric CH4-oxidising bacteria. MPI für terrestrische Mikrobiologie, Marburg.
Eller, G., Frenzel, P., 2001. Changes in activity and community structure ofmethane oxidisingbacteria over the growth period of rice. Appl. Environ. Microb. 67, 2395–2403.
Eller, G., Kanel, L., Krüger, M., 2005a. Cooccurrence of aerobic and anaerobic methaneoxidation in thewater column of Lake Plußsee. Appl. Environ.Microb. 71, 8925–8928.
Eller, G., Krüger, M., Frenzel, P., 2005b. Comparing field and microcosm experiments: acase study onmethano- andmethylotrophic bacteria in paddy soil. Fems Microbiol.Ecol. 51, 279–291.
Erkel, C., Kube, M., Reinhardt, R., Liesack, W., 2006. Genome of Rice Cluster I archaea— thekey methane producers in the rice rhizosphere. Science 313, 370–372.
Ettwig, K.F., Shima, S., van de Pas-Schoonen, K., 2008. Denitrifying bacteria anaerobicallyoxidize methane in the absence of Archaea. Environ. Microbiol. 10, 3164–3173.
Ettwig, K.F., van Alen, T., van de Pas-Schoonen, K., Jetten, M.S.M., Strous, M., 2009.Enrichment andmolecular detection of denitrifying methanotrophic bacteria of theNC10 Phylum. Appl. Environ. Microb. 75, 3656–3662.
FAO, 2001. Global estimates of gaseous emissions of NH3, NO and N2O from agriculturalland. FAO and IFA, Rome. ftp://ftp.fao.org/agl/agll/docs/globest.pdf.
FAO, 2006. Guidelines for soil description. 4th edition. Publishing Management Service,Information Division, FAO, Rome.
Favre, F., Tessier, D., Abdelmoula, M., Génin, J.M., Gates, W.P., Boivin, P., 2002. Ironreduction and changes in cation exchange capacity in intermittentlywaterlogged soil.Eur. J. Soil Sci. 53, 175–183.
Fiedler, S., Kalbitz, K., 2003. Concentrations and properties of dissolved organic matterin forest soils as affected by the redox regime. Soil Sci. 168, 793–801.
Fiedler, S., Vepraskas, M.J., Richardson, J.L., 2007. Soil redox potential: importance, fieldmeasurements, and observations. Adv. Agron. 94, 1–57.
Flessa, H., Fischer, W.R., 1992. Plant-induced changes in the redox potentials of ricerhizospheres. Plant Soil 143, 55–60.
Frenzel, P., 2000. Plant-associated methane oxidation in ricefields and wetlands. Adv.Microb. Ecol. 16, 85–114.
Frenzel, P., Rothfuss, F., Conrad, R., 1992. Oxygen profiles and methane turnover in aflooded rice microcosm. Biol. Fertil. Soils 14, 84–89.
Gao, S., Tanji, K.K., Scardaci, S.C., Chow, A.T., 2002. Comparison of redox indicators in apaddy soil during rice-growing season. Soil Sci. Soc. Am. J. 66, 805–817.
Gilbert, B., Frenzel, P., 1998. Rice roots and CH4 oxidation: the activity of bacteria, theirdistribution and the microenvironment. Soil Biol. Biochem. 30, 1903–1916.
Gong, Z.-T., 1983. Pedogenesis of paddy soil and its significance in soil classification. SoilSci. 135, 5–10.
Gong, Z.T., 1986. Origin, evolution and classification of paddy soils in China. Adv. Soil Sci. 5,174–200.
Gong, Z.T., Xu, Q., 1990. Paddy soils. Soils of China. Science Press, Beijing, pp. 233–260.Gong, Z.T., Chen, H.Z., Yuan, D.G., Zhao, Y.G., Wu, Y.J., Zhang, G.L., 2007. The temporal
and spatial distribution of ancient rice in China and its implications. Chin. Sci. Bull.52, 1071–1079.
Gotoh, S., Araragi, M., Koga, H., Ono, S., 1986. Hydrolyzable organic forms of nitrogen insome rice profiles as affected by organic matter. Soil Sci. Plant Nutr. 32, 535–550.

12 I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
Greenland, D.J., 1997. The sustainability of rice farming. Wallingford, CAB International(cited from Kirk, 2004).
Greipsson, S., 1994. Effects of iron plaque on roots of rice on growth andmetal concentrationof seedsandplant tissueswhencultivated inexcess copper. Commun. Soil Sci. PlantAnal.25, 2761–2769.
Greipsson, S., 1995. Effect of iron plaque on roots of rice on growth of plants in excess zincand accumulation of phosphorus in plants in excess copper or nickel. J. Plant Nutr. 18,1659–1665.
Grybos, M., Davranche, M., Gruau, G., Petitjean, P., 2007. Is trace metal release in wetlandsoils controlled by organic matter mobility or Fe-oxyhydroxides reduction? J. ColloidInterface Sci. 314, 490–501.
Grybos, M., Davranche, M., Gruau, G., Petitjean, P., Pédrot, M., 2009. Increasing pH drivesorganic matter solubilization from wetland soils under reducing conditions. Geoderma154, 13–19.
Gu, Y.F., Zhang, X.P., Tu, S.H., Lindstrom, K., 2009. Soil microbial biomass, crop yields, andbacterial community structure as affected by long-term fertilizer treatments underwheat-rice cropping. Eur. J. Soil Biol. 45, 239–246.
Hagedorn, F., Kaiser, K., Feyen, H., Schleppi, P., 2000. Effects of redox conditions and flowprocesses on the mobility of dissolved organic carbon and nitrogen in a forest soil. J.Env. Qual. 29, 288–297.
Hassannezhad, H., Pashaee, A., Khormali, F., Mohammadian, M., 2008. Effect of soilmoisture regime and rice cultivation onmineralogical characteristics of paddy soils ofMazandarran Province, Northern Iran. Amol. Int. J. Soil Sci. 3, 138–148.
Huang, F., Zhang, M., 2000. Pollen and phytolith evidence for rice cultivation during theNeolithic at Longqiuzhuang, eastern Jianhuai, China. Veget. Hist. Archaeobot. 9, 161–168.
Intergovernmental Panel on Climate Change, 2007. Climate Change 2007: The PhysicalScience Basis. Summary for Policymakers. Contribution of Working Group I to theFourth Assessment Report of the Intergovernmental Panel on Climate Change.Summary for Policymakers formally approved at the 10th Session ofWorking Group Iof the IPCC, Paris.
IUSS Working Group WRB, 2006. World Reference Base for Soil Resources 2006. WorldSoil Resources Reports 103. FAO, Rome.
Jäckel, U., Schnell, S., Conrad, R., 2001. Effect of moisture, texture and aggregate size ofpaddy soil on production and consumption of CH4. Soil Biol. Biochem. 33, 965–971.
Janssen, M., Lennartz, B., 2006. Horizontal and vertical water fluxes in paddy rice fieldsof subtropical China. Adv. Geo. Ecol. 38, 344–354.
Janssen, M., Lennartz, B., 2007. Horizontal and vertical water and solute fluxes in paddyrice fields. Soil Till. Res. 94, 133–141.
Janssen, I., Peng, X., Horn, R., 2006. Physical soil properties of paddy fields as a functionof cultivation history and texture. Adv. Geo. Ecol. 38, 446–455.
Jardine, P.M., Weber, N.L., McCarthy, J.F., 1989. Mechanism of dissolved organic carbonadsorption on soil. Soil Sci. Soc. Am. J. 53, 1378–1385.
Jia, Z.J., Cai, Z.C., Xu, H., Li, X.P., 2001. Effect of rice plants on CH4 production, transport,oxidation and emission in rice paddy soil. Plant Soil 230, 211–221.
Jia, Z.J., Cai, Z.C., Xu, H., Tsuruta, H., 2002. Effects of rice cultivars on methane fluxes in apaddy soil. Nutr. Cycl. Agroecosys. 64, 87–94.
Jiao, Y., Huang, Y., Zong, L.G., Zheng, X.H., Sass, R.L., 2005. Effects of copper concentrationonmethane emission from rice soils. Chemosphere 58, 185–193.
Kaiser, K., Guggenberger, G., 2000. The role of DOM sorption to mineral surfaces in thepreservation of organic matter in soils. Org. Geochem. 31, 711–725.
Kaiser, K., Zech, W., 2000. Sorption of dissolved organic nitrogen by acid subsoil horizonsand individual mineral phases. Eur. J. Soil Sci. 51, 403–411.
Kaiser, K., Guggenberger, G., Zech, W., 1996. Sorption of DOM and DOM fractions toforest soils. Geoderma 74, 281–303.
Kalbitz, K., Kaiser, K., 2008. Contribution of dissolved organic matter to carbon storagein forest soils. J. Plant Nutr. Soil Sci. 171, 52–60.
Kalbitz, K., Solinger, S., Park, J.-H.,Michalzik, B.,Matzner, E., 2000. Controls on thedynamicsof dissolved organic matter in soils: a review. Soil Sci. 165, 277–304.
Kalbitz, K., Schmerwitz, J., Schwesig, D., Matzner, E., 2003a. Biodegradation of soil-deriveddissolved organic matter as related to its properties. Geoderma 113, 273–291.
Kalbitz, K., Schwesig, D., Schmerwitz, J., Kaiser, K., Haumaier, L., Glaser, B., Ellerbrock, R.,Leinweber, P., 2003b. Changes in properties of soil-derived dissolved organic matterinduced by biodegradation. Soil Biol. Biochem. 35, 1129–1142.
Kanno, I., Honjo, Y., Tokudome, S., 1957. Clay minerals of paddy soils in Kyushu, 1. OnKakehata humic clay loam derived from glassy volcanic ash in the Hitoyoshi Basin,Kumamoto. J. Sci. Soil Manure Jpn. 28, 173–175 (in Japanese, cited from Mizotaet al., 1892).
Katoh, M., Murase, J., Hayashi, M., Matsuya, K., Kimura, M., 2004. Nutrient leaching fromthe plow layer by water percolation and accumulation in the subsoil in an irrigatedpaddy field. Soil Sci. Plant Nutr. 50, 721–729.
Kawaguchi, K., Kyuma, K., 1977. Paddy soils in tropical Asia: their material nature andfertility. The University Press of Hawaii, Honolulu.
Khalil, M.A.K., Rasmussen, R.A., Shearer, M.J., 1998. Effects of production and oxidationprocesses on methane emissions from rice fields. J. Geophys. Res.-Atmos. 103,25233–25239.
Kim, H.S., Pfaender, F.K., 2005. Effects of microbially mediated redox conditions on PAH-soil interactions. Environ. Sci. Technol. 39, 9189–9196.
Kimura, M., Murase, J., Lu, Y.H., 2004. Carbon cycling in rice field ecosystems in thecontext of input, decomposition and translocation of organic materials and thefates of their end products CO2 and CH4. Soil Biol. Biochem. 36, 1399–1416.
Kirk, G., 2004. The biogeochemistry of submerged soils. Wiley, Chichester.Kirk, G.J.D., Solivas, J.L., 1994. Coupled diffusion and oxidation of ferrous iron in soils. III.
Further development of the model and experimental testing. Eur. J. Soil Sci. 45,369–378.
Knicker,H., Scaroni, A.W.,Hatcher, P.G., 1996. 13C and 15NNMRspectroscopic investigationon the formation of fossil algal residues. Org. Geochem. 23, 329–341.
Kögel-Knabner, I., Guggenberger, G., Kleber, M., Kandeler, E., Kalbitz, K., Scheu, S.,Eusterhues, K., Leinweber, P., 2008. Organo-mineral associations in temperate soils:integrating biology, mineralogy, and organic matter chemistry. J. Plant Nutr. Soil Sci.171, 61–82.
Krause, S., Lüke, C., Frenzel, P., 2009. Spatial heterogeneity ofmethanotrophs: a geostatisticalanalysis of pmoA-based T-RFLP patterns in a paddy soil. Environ. Microbiol. Rep. 1,393–397.
Krüger, M., Frenzel, P., 2003. Effects of N-fertilisation on CH4 oxidation and production,and consequences for CH4 emissions frommicrocosms and rice fields. Glob. ChangeBiol. 9, 773–784.
Krüger,M., Frenzel, P., Conrad, R., 2001.Microbial processes influencingmethane emissionfrom rice fields. Glob. Change Biol. 7, 49–63.
Krüger, M., Eller, G., Conrad, R., Frenzel, P., 2002. Seasonal variation in pathways of CH4
production and in CH4 oxidation in rice fields determined by stable carbon isotopesand specific inhibitors. Glob. Change Biol. 8, 265–280.
Krüger,M., Frenzel, P., Kemnitz,D., Conrad, R., 2005.Activity, structure anddynamics of themethanogenic archaeal community in aflooded Italian ricefield. FemsMicrobiol. Ecol.51, 323–331.
Krull, E.S., Skjemstad, J.O., Graetz, D., Grice, K., Dunning,W., Cook, G., Parr, J.F., 2003. 13C-depleted charcoal from C4 grasses and the role of occluded carbon in phytoliths.Org. Geochem. 34, 1337–1352.
Kyuma, K., 1978. Mineral composition of rice soils. Soils and Rice. International RiceResearch Institute, pp. 219–234. Los Banos, Philippines.
Ladd, J.N., Foster, R.C., Skjemstad, J.O., 1993. Soil structure: carbon and nitrogen metabolism.Geoderma 56, 401–434.
Lal, R., 2002. Soil carbon sequestration in China through agricultural intensification, andrestoration of degraded and desertified ecosystems. Land Degrad. Dev. 13, 469–478.
Lennartz, B., Horn, R., Duttmann, R., Gerke, H.H., Tippkötter, R., Eickhorst, T., Janssen, I.,Janssen, M., Rüth, B., Sander, T., Shi, X., Sumfleth, K., Taubner, H., Zhang, B., 2009.Ecological safe management of terraced rice paddy landscapes. Soil Till. Res. 102,179–192.
Li, C.K., 1992. Paddy soils of China Beijing China. Science Press, pp. 156–162.Li, Z.H., Horikawa, Y., 1997. Stability behavior of soil colloidal suspensions in relation to
sequential reduction of soils. II Turbidity changes by submergence of paddy soils atdifferent temperatures. Soil Sci. Plant Nutr. 43, 911–919.
Li, Z.H., Horikawa, Y., Tamagawa, S., 1997. Stability behavior of soil colloidalsuspensions in relation to sequential reduction of soils. I. Turbidity of soil colloidalsuspension and change induced by submergence of paddy soils. Soil Sci. Plant Nutr.43, 719–728.
Li, Z., Velde, B., Li, D., 2003. Loss of K-bearing clay minerals in flood-irrigation, rice-growing soils in Jiangxi Province, China. Clay. Clay. Miner. 51, 75–82.
Li, Y.L., Vali, H., Sear, S.K., Deng, B., Zhang, C.L., 2004. Iron reduction and alternation ofnontronite NAu-2 by a sulfate-reducing bacterium. Geochim. Cosmochim. Acta 68,3251–3260.
Li, Z.P., Zhang, T.L., Han, F.X., Felix-Henningsen, P., 2005a. Changes in soil C and N contentsandmineralization across a cultivation chronosequence of paddy fields in subtropicalChina. Pedosphere 15, 554–562.
Li, Z.P., Zhang, T.L., Li, D.C., Velde, B., Han, F.X., 2005b. Changes in soil properties ofpaddy fields across a cultivation chronosequence in subtropical China. Pedosphere15, 110–119.
Liesack, W., Schnell, S., Revsbech, N.P., 2000. Microbiology of flooded rice paddies. FEMSMicrobiol. Rev. 24, 625–645.
Lin, X.G., Yin, R., Zhang, H.Y., Huang, J.F., Chen, R.R., Cao, Z.H., 2004. Changes of soilmicrobiological properties caused by land use changing from rice-wheat rotation tovegetable cultivation. Environ. Geochem. Hlth. 26, 119–128.
Liu, W.J., Zhu, Y.G., Smith, F.A., 2005. Effects of iron and manganese plaques on arsenicuptake by rice seedings (Oryza sativa L.) grown in solution culture supplied witharsenate and arsenite. Plant Soil 277, 127–138.
Lou, Y.S., Li, Z.P., Zhang, T.L., Liang, Y.C., 2004. CO2 emissions fromsubtropical arable soils ofChina. Soil Biol. Biochem. 36, 1835–1842.
Lu, Y., Conrad, R., 2005. In situ stable isotope probing of methanogenic Archaea in therice rhizosphere. Sci. 309, 1088–1090.
Lu,G., Sakagami,K., Tanaka,H.,Hamada, R., 1998. Roleof soil organicmatter in stabilizationofwater-stable aggregates in soils under different types of landuse. Soil Sci. PlantNutr.44, 147–155.
Lu, Y.H., Wassmann, R., Neue, H.U., Huang, C.Y., 2000. Dynamics of dissolved organiccarbon and methane emissions in a flooded rice soil. Soil Sci. Soc. Am. J. 64,2011–2017.
Lu, Y.H., Watanabe, A., Kimura, M., 2002. Contribution of plant-derived carbon to soilmicrobial biomass dynamics in a paddy ricemicrocosm. Biol. Fertil. Soils 36, 136–142.
Lu, Y.H., Watanabe, A., Kimura, M., 2003. Carbon dynamics of rhizodeposits, root- andshoot-residues in a rice soil. Soil Biol. Biogeochem. 35, 1223–1230.
Lu, Y.H.,Watanabe, A., Kimura,M., 2004. Contribution of plant photosynthates to dissolvedorganic carbon in a flooded rice soil. Biogeochem. 71, 1–15.
Lu, Y.H., Lueders, T., Friedrich, M.W., Conrad, R., 2005. Detecting active methanogenicpopulations on rice roots using stable isotope probing. Environ. Microbiol. 7,326–336.
Lui, H., Zhang, J., Christie, P., Zhang, F., 2008. Influence of iron plaque on uptake andaccumulation of Cd by rice (Oryza sativa L.) seedling grown in soil. Sci. Total Environ.394, 361–368.
Lüke, C., Krause, S., Cavigiolo, S., Greppi, D., Lupotto, E., Frenzel, P., 2010. Biogeography ofwetland rice methanotrophs. Environmental Microbiology 12, 862–872.
Lynch, J.M., Whipps, J.M., 1990. Substrate flow in the rhizosphere. Plant Soil 129, 1–10.Mahieu, N., Olk, D.C., Randall, E.W., 2000a. Accumulation of heterocyclic nitrogen in
humified organic matter: a 15 N-NMR study of lowland rice soils. Eur. J. Soil Sci. 51,379–389.

13I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
Mahieu,N., Olk,D.C., Randall, E.W., 2000b.Analysis of phosphorus in twohumic acid fractionsof intensively cropped lowland rice soils by P-31-NMR. Eur. J. Soil Sci. 51, 391–402.
Mahieu, N., Olk, D.C., Randall, E.W., 2002. Multinuclear magnetic resonance analysis oftwo humic acid fractions from lowland rice soils. J. Environ. Qual. 31, 421–430.
Maie, N., Watanabe, A., Kimura, M., 2004. Chemical characteristics and potential source offulvic acids leached from the plow layer of paddy soil. Geoderma 120, 309–323.
Marschner, H., 1995. Mineral nutrition of higher plants, Second ed. Academic Press,London. pp. 889.
McKinley, J.P., Stevens, T.O., Westall, F., 2000. Microfossils and paleoenvironments indeep surface basalt samples. Geomicrobiol. J. 17, 43–54.
Michalzik, B., Kalbitz, K., Park, J.H., Solinger, S.,Matzner, E., 2001. Fluxes and concentrationsof dissolved organic carbon and nitrogen — a synthesis for temperate forests.Biogeochem. 52, 173–205.
Minoda, T., Kimura, M., 1994. Contribution of photosynthesized carbon to the methaneemitted from paddy fields. Geophys. Res. Lett. 21, 2007–2010.
Minoda, T., Kimura, M., Wada, E., 1996. Photosynthates as dominant source of CH4 andCO2 in soil water and CH4 emitted to the atmosphere from paddy fields. J. Geophys.Res.-Atmos. 101, 21091–21098.
Mitsuchi, M., 1970. Some characteristics of paddy soils developed on volcanogenousKuroboku soils (Andosols). J. Sci; Soil Manure Jpn. 41, 307–313 (in Japanese, citedfrom Mizota et al., 1982).
Mizota, C., Carrasco, M.A., Wada, K., 1982. Clay mineralogy and some chemical propertiesof Ap horizons of ando soils used paddy rice in Japan. Geoderma 27, 225–237.
Mohapatra, S.P., Khan, S.K., 1983. Mineralizable nitrogen in relation to soil nitrogenforms and soil properties. Oryza 20, 227–232.
Moore, T.R., Dalva, M., 2001. Some controls on the release of dissolved organic carbonby plant tissue and soils. Soil Sci. 166, 38–47.
Munch, J.C., Hillebrand, T., Ottow, J.C.G., 1978. Transformation in the Feo/Fed ratio ofpedogenetic ironoxides affectedby iron-reducingbacteria. Can. J. Soil Sci. 58, 475–486.
Murase, J., Frenzel, P., 2007. Amethane-drivenmicrobial foodweb in a wetland rice soil.Environ. Microbiol. 9, 3025–3034.
Murase, J., Frenzel, P., 2008. Selective grazing of methanotrophs by protozoa in a ricefield soil. FEMS Microbiol. Ecol. 65, 408–414.
Murase, J., Kimura, M., 1994. Methane production and its fate in paddy fields VI.Anaerobic oxidation ofmethane in plow layer soil. Soil. Sci. Plant Nutr. 40, 505–514.
Murase, J., Kimura, M., 1996. Methane production and its fate in paddy fields IX. Methaneflux distribution and decomposition of methane in the subsoil during the growthperiod of rice plants. Soil. Sci. Plant Nutr. 42, 187–190.
Murase, J., Noll, M., Frenzel, P., 2006. The impact of protists on activity and structure ofthe bacterial community in a rice field soil. Appl. Environ. Microb. 72, 5436–5444.
Nelson, P.N., Baldock, J.A., Oades, J.M., 1993. Concentration and composition of dissolvedorganic carbon in streams in relation to catchment soil properties. Biogechem. 19,27–50.
Neue, H.U., Roger, P.A., 2003. Rice agriculture: factors controlling emissions. In: Khalil,M.A.K. (Ed.), Atmospheric Methane.
Neue, H.U., Scharpenseel, H.W., 1987. Decomposition pattern of 14C-labeled rice strawin aerobic and submerged rice soilks of the Philippines. Sci. Total Environ. 62,431–434.
Neue, H.U., Wassmann, R., Lantin, R.S., Alberto, M.A.C.R., Aduna, J.B., Javellana, A.M., 1996.Factors affecting methane emission from rice fields. Atmos. Environ. 30, 1751–1754.
Neue, H.U., Gaunt, J.L., Wang, Z.P., Becker-Heidmann, P., Quijano, C., 1997. Carbon intropical wetlands. Geoderma 79, 163–185.
Nguyen, B.V., Olk, D.C., Cassman, K.G., 2004. Nitrogen mineralization from humic acidfractions in rice soils depends on degree of humifcation. Soil Sci. Soc. Am. J. 68,1278–1284.
Nierop, K.G.J., Jansen, B., Verstraten, J.A., 2002. Dissolved organicmatter, aluminium andiron interactions: precipitation induced bymetal/carbon ratio, pH and competition.Sci. Total Environ. 300, 201–211.
Oades, J.M., Waters, A.G., 1991. Aggregate hierarchy in soils. Aust. J. Soil. Res. 29, 815–828.Olk, D.C., Cassman, K.G., Randall, E.W., Kinchesh, P., Sanger, L.J., Anderson, J.M., 1996.
Changes in chemical properties of organic matter with intensified rice cropping intropical lowland soil. Eur. J. Soil Sci. 47, 293–303.
Olk, D.C., Cassman, K.G., Mahieu, N., Randall, E.W., 1998. Conserved chemical propertiesof young humic acid fractions in tropical lowland soil under intensive irrigated ricecropping. Eur. J. Soil Sci. 49, 337–349.
Olk, D.C., Brunetti, G., Senesi, N., 1999. Organic matter in double-cropped lowland ricesoils: chemical and spectroscopic properties. Soil Sci. 164, 633–649.
Olk, D.C., Brunetti, G., Senesi, N., 2000. Decrease in humification of organic matter withintensified lowland rice cropping: a wet chemical and spectroscopic investigation.Soil Sci. Soc. Am. J. 64, 1337–1347.
Olk, D.C., Dancel, M.C., Moscoso, E., Jimenez, R.R., Dayrit, F.M., 2002. Accumulation oflignin residues in organic matter fractions of lowland rice soils: a pyrolysis-GC-MSstudy. Soil Sci. 167, 590–606.
Olk, D.C., Cassman, K.G., Schmidt-Rohr, K., Anders, M.M., Mao, J.-D., Deenik, J.L., 2006.Chemical stabilization of soil organic nitrogen byphenolic lignin residues in anaerobicagroecosystems. Soil Biol. Biochem. 38, 3303–3312.
Olk, D.C., Samson, M.I., Gapas, P., 2007. Inhibition of nitrogen mineralization in younghumic fractions by anaerobic decomposition of rice crop residues. Eur. J. Soil Sci. 58,270–281.
Pan, G., Li, L., Wu, L., 2003. Storage and sequestration potential of topsoil organic carbonin China's paddy soils. Glob. Change Biol. 10, 79–92.
Pan, G., Wu, L., Li, L., Zhang, X., Gong, W., Wood, Y., 2008. Organic carbon stratificationand size distribution of three typical paddy soils from Taihu Lake region, China. J.Environ. Sci. 20, 456–463.
Pande, K.R., 2005. Managing seasonal soil N-dynamics in rice-wheat rotation systems ofNepal. PhD thesis at the University of Bonn.
Pande, K.R., Becker, M., 2003. Seasonal soil nitrogen dynamics in rice-wheat croppingsystems of Nepal. J. Plant Nutr. Soil Sci. 166, 499–506.
Parr, J.F., Sullivan, L.A., 2005. Soil carbon sequestration in phytoliths. Soil Biol. Biochem.37, 117–124.
Parsons, J.W., 1981. Chemistry and distribution of amino sugars in soils and soilorganisms. In: Paul, E.A., Ladd, J.N. (Eds.), Soil Biochemistry, Vol. 5. Marcel Dekker,New York, pp. 197–227.
Patrick, W.H., Jugsujinda, A., 1992. Sequential reduction and oxidation of inorganicnitrogen, manganese, and iron in flooded soil. Soil Sci. Soc. Am. J. 56, 1071–1073.
Patrick,W.H., Reddy, C.N., 1978. Chemical changes in rice soils. International Rice ResearchInstitute. International Rice Research Institute, Los Banos, pp. 361–379.
Peng, J.J., Lu, Z., Rui, J., Lu, Y.H., 2008. Dynamics of the methanogenic archaeal communityduring plant residue decomposition in an anoxic rice field soil. Appl. Environ. Microb.74, 2894–2901.
Ponnamperuma, F.N., 1972. The chemistry of submerged soils. Adv. Agron. 24, 29–96.Prakongkep, N., Suddhiprakam, A., Kheoruenromne, I., Smirk, M., Gilkes, R.J., 2008. The
geochemistry of Thai paddy soils. Geoderma 144, 310–324.Quantin, C., Grunberger, O., Suvannang, N., Bourdon, E., 2008. Landmanagement effects on
biogeochemical functioning of salt-affected paddy soils. Pedosphere 18, 183–194.Ramakrishnan, B., Lueders, T., Conrad, R., Friedrich, M., 2000. Effect of soil aggregate size
on methanogenesis and archaeal community structure in anoxic rice field soil.Fems Microbiol. Ecol. 32, 261–270.
Rancourt, D.G., Thibault, P.-J., Mavrocordatos, D., Lamarche, G., 2005. Hydrous ferricoxide precipitation in the presence of nonmetabolizing bacteria: constraints on themechanisms of a biotic effect. Geochim. Cosmochim. Acta 69, 553–577.
Ratering, S., Conrad, R., 1998. Effects of short-termdrainage andaerationon theproductionof methane in submerged rice soil. Glob. Change Biol. 4, 397–407.
Ratering, S., Schnell, S., 2000. Localization of iron-reducing activity in paddy soil by profilestudies. Biogeochem. 48, 341–365.
Roberts, P., Bol, R., Jones, D.L., 2007. Free amino sugar reactions in soil in relation to soilcarbon and nitrogen cycling. Soil Biol. Biochem. 39, 3081–3092.
Rothfuss, F., Bijnen, F.G.C., Conrad, R., Harren, F.J.M., Reuss, J., 1996. Combination ofphotoacoustic detector with gas diffusion probes for the measurement of methaneconcentration gradients in submerged paddy soil. Chemosphere 33, 2487–2504.
Roy, R.N., Misra, R.V., 2003. Economic and environmental impact of improved nitrogenmanagement in Asian rice-farming systems. Proceedings of the 20th Session of theInternational Rice Commission, Bangkok, Thailand, 23–26 July 2002, FAO, Rom.
Saejiew, A., Grunberger, O., Arunin, S., Favre, F., Tessier, D., Biovin, P., 2004. Criticalcoagulation of paddy soil clays in sodium-ferrous iron electrolyte. Soil Sci. Soc. Am. J.68, 789–794.
Sahrawat, K.L., 2004. Organic matter accumulation in submerged soils. Adv. Agron. 81,169–201.
Sahrawat, K.L., 2005. Fertility and organic matter in submerged rice soil. Curr. Sci. 88,735–739.
Sano, O., Ito, T., Ando, T., Nanzyo, M., Saito, G., Saito, K., Saigusa, M., 2006. Claymineralogical composition map of Paddy soils in Miyagi Prefecture, NortheasternJapan. 18th World Congress of Soil Science July 9-15, 2006, Philadelphia,Pennsylvania, USA, pp. 115–133.
Scharpenseel, H.W., Pfeiffer, E.-M., Becker-Heidmann, P., 1996. Organic carbon storagein tropical hydromorphic soils. In: Carter, M.R., Stewart, B.A. (Eds.), Structure andorganicmatter storage in agricultural soils. Adv. Soil Sci. Lewis Publishers, Boca Raton,pp. 361–392.
Scheel, T., Dörfler, C., Kalbitz, K., 2007. Precipitation of dissolved organic matter by Alstabilizes carbon in acidic forest soils. Soil Sci. Soc. Am. J. 71, 64–74.
Schmidt-Rohr, K., Mao, J.-D., Olk, D.C., 2004. Nitrogen-bonded aromatics in soil organicmatter and their implications for a yield decline in intensive rice cropping. Proc.Natl. Acad. Sci. USA 101, 6351–6354.
Sharpless, C.M., McGown, L.B., 1999. Effects of aluminum-induced aggregation on thefluorescence of humic substances. Environ. Sci. Technol. 33, 3264–3270.
Shirato, Y., 2005. Testing the suitability of the DNDC model for simulating long-termsoil organic dynamics in Japanese paddy oils. Soil Sci. Plant Nutr. 51, 183–192.
Smernik, R.J., Olk, D.C.,Mahieu, N., 2004. Quantitative solid-state 13CNMR spectroscopy oforganic matter fractions in lowland rice soils. Eur. J. Soil Sci. 55, 367–379.
Sommer, M., Kaczorek, D., Kuzyakov, Y., Breuer, J., 2006. Silicon pools and fluxes in soilsand landscapes — a review. J. Plant Nutr. Soil Sci. 169, 310–329.
Stevenson, F.J., 1982. Organic forms of soil nitrogen. In: Stevenson, F.J. (Ed.), Nitrogen inagricultural soils. American Society of Agronomy, Inc., Madison, WI, pp. 67–122.
Suetsugu, A., Sato, T., Kaneta, Y., Sato, A., 2005. Effects of organic-mineral complexes onflocculation, settlement and vertical distribution of bioelements in soil suspensions.Sci. Plant Nutr. 51, 323–331.
Takai, Y., 1961. Reduction and microbial metabolism in paddy soils. Nogyo Gijitsu(Agricultural Technology) 19, 122–126 (in Japanese, cited from Bouwman, 1991).
Tanji, K.K., Gao, S., Scardaci, S.C., Chow, A.T., 2003. Characterization redox status of paddysoils with incorporated rice straw. Geoderma 114, 333–353.
Thompson, A., Chadwick, O.A., Rancourt, D.G., Chorover, J., 2006a. Iron-oxide crystallinityincreases during redox oscillations. Geochim. Cosmochim. Acta 70, 1710–1727.
Thompson, A., Chadwick, O.A., Boman, S., Chorover, J., 2006b. Colloid mobilizationduring soil iron redox oscillations. Environ. Sci. Technol. 40, 5743–5749.
Tripathy, R., Singh, A.K., 2004. Effect of water and nitrogen management on aggregatesize and carbon enrichment in soil in rice–wheat cropping system. J. Plant Nutr. SoilSci. 167, 216–228.
Trolldenier, G., 1988. Visualisation of oxidising power of rice roots and of possibleparticipation of bacteria in iron deposition. Z. Pflanzenernaehr. Bodenkd. 151, 117–121.
Tyagi, L., Verma, A., Singh, S.N., 2004. Investigation on temporal variation in methaneemission from different rice cultivars under the influence of weeds. Environ. Monit.Assess. 93, 91–101.

14 I. Kögel-Knabner et al. / Geoderma 157 (2010) 1–14
Ueshima, M., Tazaki, K., 2001. Possible role of microbial polysaccharides in nontroniteformation. Clays Clay Min. 49, 292–299.
Van der Gon, H.A.C.D., Kropff, M.J., van Breemen, N., Wassmann, R., Lantin, R.S., Aduna,E., Corton, T.M., van Laar, H.H., 2002. Optimizing grain yields reduces CH4 emissionsfrom rice paddy fields. PNAS 99, 12021–12024.
Van Ranst, E., De Coninck, F., 2002. Evaluation of ferrolysis in soil formation. Eur. J. SoilSci. 53, 513–519.
Voet, D., Voet, J.G., 1995. Biochemistry. John Wiley & Sons Ltd., New York.Wang, Y., Zhou, X.,Wu, J., 1992.Mössbauer study on the iron oxideminerals of paddy soils
derived from red soil in Fujian, China. Hyperfine Interact. 70, 1037–1040.Wang, B., Neue, H.U., Samonte, H.P., 1997. Role of rice in mediating methane emission.
Plant Soil 189, 107–115.Wassmann, R., Neue, H.U., Lantin, R.S., Buendia, L.V., Rennenberg, H., 2000a. Character-
izationofmethaneemissions from ricefields inAsia, I. Comparison amongfield sites infive countries. Nutr. Cycl. Agroecosys. 58, 13–22.
Wassmann, R., Neue, H.U., Lantin, R.S., Makarim, K., Chareonsilp, N., Buendia, L.V.,Rennenberg, H., 2000b. Characterization of methane emissions from rice fields in Asia.II. Differences among irrigated, rainfed, and deepwater rice. Nutr. Cycl. Agroecosys. 58,13–22.
Wickramasinghe, D.B., Rowell, D.L., 2006. The release of silicon from amorphous silicaand rice straw in Sri Lankan soils. Biol. Fertil. Soils 42, 231–240.
Wilding, L.P., 1967. Radiocarbon dating of biogenetic opal. Sci. 156, 66–67.Wilding, L.P., Brown, R.E., Holowaychuk, N., 1967. Accessibility and properties of occluded
carbon in biogenic opal. Soil Sci. 103, 56–61.Wilding, L.P., Hallmark, C.T., Smeck, N.E., 1979. Dissolution and stability of biogenic opal.
Soil Sci. Soc. Am. Proc. 43, 800–802.Xu, H., Cai, Z.C., Li, X.P., Tsuruta, H., 2000. Effect of antecedent soil water regime and rice
strawapplication time onCH4 emission fromrice cultivation.Aust. J. Soil Res. 38, 1–12.
Yamagata, M., 2000. Preferential use of organic nitrogen by upland rice. Nogyo KankyxoGijutsu Kenkyosho Hokuku 18, 1–31 in Japanese, cited from abstract.
Yan, X., Yagi, K., Akiyama, H., Akimoto, H., 2005. Statistical analysis of the major variablescontrolling methane emission from rice fields. .
Yanamuro, S., Ueno,H., Takahashi, S., 1999. Fate of free amino acids inpaddy andupland soilsby using 13C and 15N tracer techniques. Nippon Dojo Hiryogaki Zasshi 70, 739–746 inJapanese, cited from abstract.
Yang, C., Yang, L., Ouyang, Z., 2005. Organic carbon and its fractions in paddy soils asaffected by different nutrient and water regimes. Geoderma 124, 133–142.
Yang, C., Rodionov, D.A., Li, X.Q., Laikova, O.N., Gelfand, M.S., Zagnitko, O.P., Romine, M.F.,Obraztsova, A.Y., Nealson, K.H., Osterman, A.L., 2006. Comparative genomics andexperimental characterization of N-acetylglucosamine utilization pathway of Shewa-nella oneidensis. J. Biol. Chem. 281, 29872–29885.
Yao, H., Conrad, R., Wassmann, R., Neue, H.U., 1999. Effect of soil characteristics onsequential reduction and methane production in sixteen rice paddy soils from China,the Philippines, and Italy. Biogeochem. 47, 269–295.
Yu, K., Patrick Jr., W.H., 2003. Redox range with minimum nitrous oxides and methaneproduction in rice soil under different pH. Soil Sci. Am. J. 67, 1952–1958.
Yu, K., Böhme, F., Rinklebe, J., Neue, K.-H., DeLaune, R.D., 2007. Major biogeochemicalprocesses in soils–microcosm incubation from reducing to oxidizing conditions.Soil Sci. Soc. Am. J. 71, 1406–1417.
Zhang, G.L., Gong, Z.T., 2003. Pedogenic evolution of paddy soils in different soil landscape.Geoderma 115, 15–29.
Zhang, M., He, Z., 2004. Long-term changes in organic carbon and nutrients of an Ultisolunder rice cropping in southeast China. Geoderma 118, 167–179.
Zhang, X., Amelung, W., Yuan, Y., Zech, W., 1998. Amino sugar signature of particle-sizefractions in soils of the native prairie as affected by climate. Soil Sci. 163, 220–229.
Related Documents











