BIOE 109 Summer 2009 Lecture 10- part I Mating systems

BIOE 109 Summer 2009 Lecture 10- part I Mating systems.
Dec 15, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

BIOE 109Summer 2009
Lecture 10- part IMating systems

Types of Mating Systems
Mating Systems Mode Parental care
Monogamy One male and one female form bond
both
Polygyny Male mates with multiple females
female
Polyandry Female mates with multiple males
male
Promiscuity Both sexes mate with multiple partners
Both, either, or neither

Mating Systems In Nature

Polygyny Polygyny
MonogamyMonogamy
Polyandry
Monogamy
Mating Systems In Nature

?
Mating Systems In Nature
Promiscuous
Mating Systems In Nature

Promiscuous
Mating Systems In Nature

Hypothesis for the evolution of mating systems
• Based on parental care and ecological constraints

Hypothesis for the evolution of mating systems
• Based on parental care and ecological constraints
Who can ditch first?

Hypothesis for the evolution of mating systems
• Based on parental care and ecological constraints
Who can ditch first?
Is ditching worth it?

Sex allocation

Sex allocation• the allocation of resources to male versus
female production in sexual species (Charnov 1982).
• Sex Ratio?

What is sex ratio?
• sex ratio is defined as the proportion of males to females.

What is sex ratio?
• sex ratio is defined as the proportion of males to females.
• two distinct sex ratios exist:

What is sex ratio?
• sex ratio is defined as the proportion of males to females.
• two distinct sex ratios exist:
1. the population sex ratio
i.e., the proportion of males to females in the population

What is sex ratio?
• sex ratio is defined as the proportion of males to females.
• two distinct sex ratios exist:
1. the population sex ratio
i.e., the proportion of males to females in the population
2. the individual sex ratio

What is sex ratio?
• sex ratio is defined as the proportion of males to females.
• two distinct sex ratios exist:
1. the population sex ratio
i.e., the proportion of males to females in the population
2. the individual sex ratio
i.e., the sex ratio of progeny from a female

-In many species sex chromosomes cause 1:1 sex ratio
The evolution of sex ratio

Mammals: females are homogametic (XX)
males are heterogametic (XY) Birds: males are homogametic (ZZ)
females are heterogametic (WZ)
The evolution of sex ratio

Mammals: females are homogametic (XX)
males are heterogametic (XY) Birds: males are homogametic (ZZ)
females are heterogametic (WZ) Sex chromosomes do not guarantee a 1:1 sex ratio!

Why equal numbers of males and females?
• R.A. Fisher (1930) provided a genetic explanation for the evolution of a stable sex ratio of 1:1.

Why equal numbers of males and females?
• R.A. Fisher (1930) provided a genetic explanation for the evolution of a stable sex ratio of 1:1.
• since every individual has one mother and one father, each sex contributes equally, on average, to subsequent generations.

Why equal numbers of males and females?
• R.A. Fisher (1930) provided a genetic explanation for the evolution of a stable sex ratio of 1:1.
• since every individual has one mother and one father, each sex contributes equally, on average, to subsequent generations.
• therefore, males and females must have the same average fitness.

Suppose:25% male males will have high fitness75% female because they mate with
multiple females
Suppose:
75% male females will have high fitness 25% female because they mate with multiple
males
• Members of the rarer sex will experience increased reproductive success relative to common sex
• frequency-dependent selection results in stable equilibrium sex ratio of 1:1.

Exceptions to Fisher’s theory
NOT ALWAYS 1:1

Exceptions to Fisher’s theory
1. Local mate competition (Hamilton 1967)
2. Condition-dependent sex allocation (Trivers and Willard 1973)

Exceptions to Fisher’s theory
1. Local mate competition (Hamilton 1967)
• proposed to account for female-biased sex ratios (e.g., parasitoid wasps).

Exceptions to Fisher’s theory
1. Local mate competition (Hamilton 1967)
• proposed to account for female-biased sex ratios (e.g. parasitoid wasps).
• here, a single foundress produces a small group of closely related individuals that mate among themselves.

Exceptions to Fisher’s theory
1. Local mate competition (Hamilton 1967)
• proposed to account for female-biased sex ratios (e.g. parasitoid wasps).
• here, a single foundress produces a small group of closely related individuals that mate among themselves.
• females invest heavily in daughters and don’t “waste” effort in producing sons.
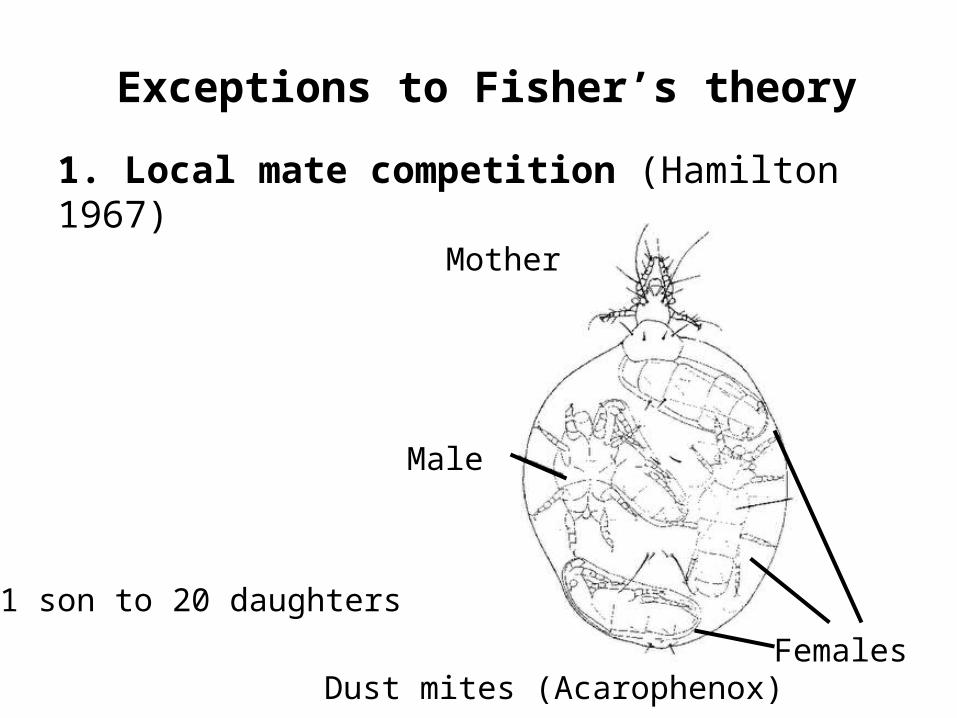
Exceptions to Fisher’s theory
1. Local mate competition (Hamilton 1967)
Male
Females
Mother
Dust mites (Acarophenox)
1 son to 20 daughters

Exceptions to Fisher’s theory
2. Condition-dependent sex allocation (Trivers and Willard 1973)
Red deer, Cervus elaphus

Exceptions to Fisher’s theory
2. Condition-dependent sex allocation (Trivers and Willard 1973)
• occurs in polygynous species when females invest heavily in producing and caring for their young.

Exceptions to Fisher’s theory
2. Condition-dependent sex allocation (Trivers and Willard 1973)
• occurs in polygynous species when females invest heavily in producing and caring for their young.
• a good mother can produce larger, or healthier, individuals when they mature.

Exceptions to Fisher’s theory
2. Condition-dependent sex allocation (Trivers and Willard 1973)
• occurs in polygynous species when females invest heavily in producing and caring for their young.
• a good mother can produce larger, or healthier, individuals when they mature.
• theory predicts that females in extremely good condition should produce males.

Exceptions to Fisher’s theory
2. Condition-dependent sex allocation (Trivers and Willard 1973)
• occurs in polygynous species when females invest heavily in producing and caring for their young.
• a good mother can produce larger, or healthier, individuals when they mature.
• theory predicts that females in extremely good condition should produce males.
• Why?

Exceptions to Fisher’s theory
2. Condition-dependent sex allocation (Trivers and Willard 1973)
• occurs in polygynous species when females invest heavily in producing and caring for their young.
• a good mother can produce larger, or healthier, individuals when they mature.
• theory predicts that females in extremely good condition should produce males.
• Why? Because sexual selection (usually) occurs more strongly in males and condition matters!

How is sex ratio adjusted by mother?
• Not known

Sex Allocation Recap
• Sex ratio• Why we see an unbiased sex ratio
– Sex chromosomes– Frequency dependent selection
• Exceptions to sex ratio:– Local mate competition– Condition-dependent sex allocation

Sex in Plants
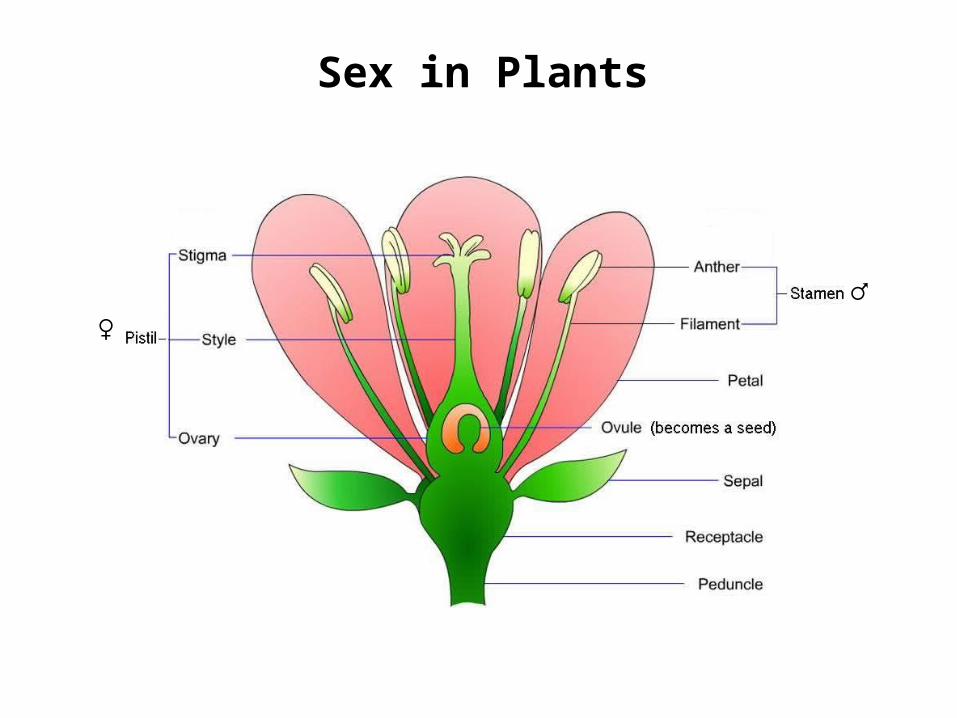
Sex in Plants
♂♀

Sex in Plants
• Why and how do they outbreed?
• Why do they inbreed?

The evolution of inbreeding and outbreeding
• many plant species have evolved traits to avoid inbreeding.

The evolution of inbreeding and outbreeding
• many plant species have evolved traits to avoid inbreeding. 1. Asynchronous male and female functions
• pollen shed after or before plant’s stigmas are receptive.

The evolution of inbreeding and outbreeding
• many plant species have evolved traits to avoid inbreeding. 1. Asynchronous male and female functions
• pollen shed after or before plant’s stigmas are receptive.

The evolution of inbreeding and outbreeding
• many plant species have evolved traits to avoid inbreeding. 1. Asynchronous male and female functions
• pollen shed after or before plant’s stigmas are receptive. 2. Monoecy
• male and female flowers separated on same plant.
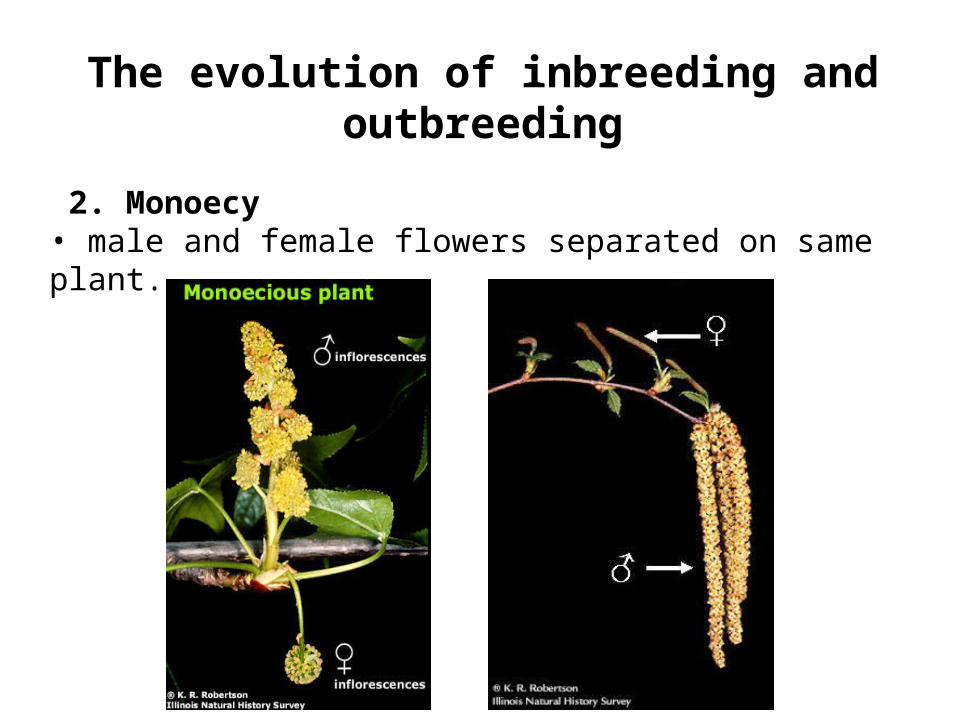
The evolution of inbreeding and outbreeding
2. Monoecy• male and female flowers separated on same plant.

The evolution of inbreeding and outbreeding
• many plant species have evolved traits to avoid inbreeding. 1. Asynchronous male and female functions
• pollen shed after or before plant’s stigmas are receptive.
2. Monoecious
• male and female flowers separated on same plant. 3. Dieocy
• sexes are separated in different individuals.

The evolution of inbreeding and outbreeding
3. Dieocy
• sexes are separated in different individuals.

The evolution of inbreeding and outbreeding
4. Self-incompatibility loci
• prevent selfing or breeding with close relatives.

The evolution of inbreeding and outbreeding
4. Self-incompatibility loci
• prevent selfing or breeding with close relatives. 5. Heterostyly
• two (distyly) or three (tristyly) forms of flowers exist in a species (on different plants).

The evolution of inbreeding and outbreeding
4. Self-incompatibility loci
• prevent selfing or breeding with close relatives. 5. Heterostyly
• two (distyly) or three (tristyly) forms of flowers exist in a species (on different plants).
• pollen is more effectively transferred between, rather than within, morphs.

The evolution of inbreeding and outbreeding
4. Self-incompatibility loci
• prevent selfing or breeding with close relatives. 5. Heterostyly
• two (distyly) or three (tristyly) forms of flowers exist in a species (on different plants).
• pollen is more effectively transferred between, rather than within, morphs.
• acts to maximize outcrossing.

Thrum flowered Primula polyneura Pin flowered Primula polyneura
Heterostyly in Primula polynera

Why inbreed?

Why inbreed?
• partial selfers have a fitness advantage over exclusive outcrossers because genes can be transmitted:

Why inbreed?
• partial selfers have a fitness advantage over exclusive outcrossers because genes can be transmitted: 1. through its ovules

Why inbreed?
• partial selfers have a fitness advantage over exclusive outcrossers because genes can be transmitted: 1. through its ovules2. through its pollen by selfing

Why inbreed?
• partial selfers have a fitness advantage over exclusive outcrossers because genes can be transmitted: 1. through its ovules2. through its pollen by selfing3. through its pollen by outcrossing.

Why inbreed?
• partial selfers have a fitness advantage over exclusive outcrossers because genes can be transmitted: 1. through its ovules2. through its pollen by selfing3. through its pollen by outcrossing.
• another advantage of selfing is reproductive assurance

Why inbreed?
• partial selfers have a fitness advantage over exclusive outcrossers because genes can be transmitted: 1. through its ovules2. through its pollen by selfing3. through its pollen by outcrossing.
• another advantage of selfing is reproductive assurance
• if pollinators are scarce, then a plant can produce at least some seeds by selfing.

OVERALL
• Mating Systems
• Sex allocation
• Sex ratios
• Exceptions to 1:1 sex ratio
• Mating in plants
Related Documents











