See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/262564975 Biodegradation of Azo Dyes a Review Article · October 2014 READS 550 2 authors: Waleed M. Sh. Alabdraba University of Tikrit 41 PUBLICATIONS 1 CITATION SEE PROFILE Mohamed Bayati University of Missouri 3 PUBLICATIONS 0 CITATIONS SEE PROFILE All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately. Available from: Waleed M. Sh. Alabdraba Retrieved on: 19 June 2016

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 1/16
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/262564975
Biodegradation of Azo Dyes a Review
Article · October 2014
READS
550
2 authors:
Waleed M. Sh. Alabdraba
University of Tikrit
41 PUBLICATIONS 1 CITATION
SEE PROFILE
Mohamed Bayati
University of Missouri
3 PUBLICATIONS 0 CITATIONS
SEE PROFILE
All in-text references underlined in blue are linked to publications on ResearchGate,
letting you access and read them immediately.
Available from: Waleed M. Sh. Alabdraba
Retrieved on: 19 June 2016

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 2/16
Biodegradation of Azo Dyes a Review
Waleed Mohammed Sheet Alabdraba Mohammed Burhan Ali Albayati
Assistant Prof. Assistant Lecturer
Environmental Engineering Department, College of Engineering, Tikrit University
Abstract
One of the main problems associated with the treatment of textile wastewater is the removalof dyes. Most (60-70%) of the more than 10,000 dyes applied in textile-processing industries are
Azo compounds, i.e. molecules with one or more Azo (N=N) bridges linking substituted
aromatic structures. Discharge of Azo dyes is undesirable, not only for aesthetic reasons, but also
because many Azo dyes and their breakdown products are toxic toward aquatic life, andmutagenic for humans. Reductive cleavage of Azo bond, leading to the formation of aromatic
amines, is the initial reaction during the biological metabolism of Azo dyes. Anaerobic/anoxic
Azo dye decolorization by several mixed and pure bacterial cultures have been reported. Under
these conditions, this reaction is nonspecific with respect to organisms as well as dyes. Variousmechanisms, which include direct enzymatic and indirect (mediated) and chemical Azo dye
reduction. Only few aerobic bacterial strains that can utilize Azo dyes as growth substrates have
been isolated. These organisms generally have a narrow substrate range. Degradation of aromaticamines depends on their chemical structure and the conditions. It is now known that simple
aromatic amines can be mineralized under methanogenic conditions. Sulfonated aromatic
amines, on the other hand, are resistant and require specialized aerobic microbial consortia fortheir mineralization. This review is focused on the biodegradation of Azo dyes and
mineralization of aromatic amines
Key Word: Azo dyes, Biodegradation, Aromatic amines, Anaerobic/aerobic treatment
1. IntroductionAzo dyes are one of the oldest man-made chemicals and they are still widely used in textile
printing and the food industries. The annual production worldwide is approximately 700,000 tons(Zollinger 1987). Of this amount, approximately 10 -15% of the dyes are released into the
environment during manufacturing and usage (Vaidya and Datye 1982). Some dyes and some of
their N-substituted aromatic biotransformation products are toxic and/or carcinogenic and,
therefore, the dyes are considered as important environmental pollutants (Chung and Cerniglia1992). During conventional aerobic wastewater treatment, Azo dyes are not degraded (Pagga and
Brown 1986; Shaul et al. 1991). However, Azo linkages are easily reduced under anaerobic
conditions (Brown and Laboureur 1983b), with digester sludge (Carliell et al. 1995; Carliell et al.
1994), anaerobic granular sludge (Donlon et al. 1997; RAzo-Flores et al. 1997) or sediments(Weber 1991). The Azo dyes act as electron acceptors for reduced flavin nucleotides and redox
mediators (Gingell and Walker 1971; Keck et al. 1997; Kudlich et al. 1997) enhance reduction.
The reduction of Azo dyes results in the formation of aromatic amines. Most aromatic amines,which accumulate after Azo cleavage, are not mineralized anaerobically (Brown and Hamburger
1987), with the exception of a few aromatic amines substituted with hydroxyl and carboxyl
groups which were fully degraded under methanogen conditions (RAzo-Flores et al. 1996).However, the aromatic amines are readily degraded aerobically (Brown and Laboureur 1983a;
Konopka 1993; Loidl et al. 1990). Generally, bacterial Azo dye biodegradation proceeds in two

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 3/16
stages. The first stage involves reductive cleavage of the dyes’ Azo linkages, resulting in the
formation of generally colorless but potentially hazardous aromatic amines. The second stage
involves degradation of the aromatic amines. Azo dye reduction usually requires anaerobicconditions, whereas bacterial biodegradation of aromatic amines is an almost exclusively aerobic
process (Fig. 1).
A combination of anaerobic and aerobic conditions is therefore proposed as a feasible
biological treatment strategy for Azo compounds (Field et al. 1995). Anaerobic and aerobicconditions can be applied in a sequential anaerobic/aerobic bioreactor system but also in a single
reactor in which anaerobic and aerobic micro niches occur side by side. Sequential
anaerobic/aerobic treatment for Azo dyes and textile industry wastewater has been evaluated inseveral studies (An et al. 1996; Brown and Hamburger 1987; FitzGerald and Bishop 1995;
Seshadri et al. 1994; Zaoyan et al. 1992). Also a simultaneous anaerobic/aerobic treatment
methodology for an Azo dye was recently described. (Esenceli and Tiyek 2014).
2. Decolorization of Azo dye by bacteria
Reductive cleavage of the – N=N- bond is the initial step of the bacterial degradation of Azodyes. Decolorization of Azo dyes occurs under anaerobic (methanogenic), anoxic and aerobic
condition by different groups of bacteria. Decolourization Azo dyes under these different is
briefly discussed in subsequent sections.
2.1. Azo dye decolorization under anaerobic conditions
Anaerobic Azo dye reduction is the reductive cleavage of Azo linkages, i.e. the transfer ofreducing equivalents resulting in the formation of aromatic amines. As aromatic amines are
Fig. 1 General overview of the fate of Azo dyes and aromatic amines during anaerobic/aerobic
treatment

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 4/16
generally colorless, Azo dye reduction is also referred to as Azo dye decolorization. The first
study on Azo dye reduction was published as early as 1937, when the decolorization of food Azo
dyes by lactic acid bacteria isolated from the human gut was reported (Brohm and Frohwein,1937). Hence, as the formation of toxic aromatic amines in humans is a matter of concern,
research on bacterial Azo dye reduction has traditionally mostly been focused on the activity of
(facultative) anaerobic bacteria from mammalian intestines (Brown, 1981; Chung et al., 1978; ;Chung et al., 1992; Rafii et al.1990; Walker, 1970). Later, when the removal of dyes fromwastewater became a topic, also bacteria from other origins were used to investigate anaerobic
Azo dye reduction, e.g. pure cultures (Wuhrmann et al., 1980), mixed cultures (Haug et al.,
1991), anaerobic sediments (Weber and Wolfe, 1987), digester sludge (Brown and Laboureur, 1983), (Carliell et al., 1994), anaerobic granular sludge (RAzo Flores, 1997) and activated sludge
under anaerobic conditions (Bromly-Challenor, 2000 ; Bell and Buckley, 2003; Albayati, 2010
and Aldoury et al., 2014 ).
2.2 Azo dye decolorization under anoxic conditions
Anoxic decolorization of various Azo dyes by mixed aerobic and facultative microbialconsortia has been reported ( Nigam et al., 1996; Kapdan et al., 2000; Padmavathy et al., 2003;
Khehra et al., 2005; Moosvi et al., 2005). Although many of these cultures were able to grow
aerobically, decolorization was achieved only under anaerobic conditions. Pure bacterial strains,
such as Pseudomonas sp. and Proteus mirabilis, decolorized Azo dyes under anoxic conditions(Chang et al., 2001; Chen et al., 1999; Yu et al., 2001). Azo dye decolorization by mixed, as well
as pure, cultures generally required complex organic sources, such as yeast extract, peptone, or
a combination of complex organic source and carbohydrate(Chen et al., 2003; Khehra et al., 2005). Glucose is the preferred substrate in anaerobic dye decolorization under methanogenic
conditions, but its suitability for anoxic dye decolorization by facultative anaerobic and
fermenting bacteria seems to vary, depending on the bacterial culture.
Decolorization of Mordant Yellow 3 by Sphingomonas xenophaga strain BN6 was greatlyenhanced by glucose, where as a significant decrease in Azo decolorization in its presence was
reported for P leuteola, Aeromonas sp. and few other mixed cultures (Haug et al., 1991; Kapdan
et al., 2000; Chang et al., 2001; Chen et al., 2003). The negative effect of glucose on anoxicdecolorization has been attributed either to a decrease in pH due to acid formation, or to
catabolic repression (Chen et al., 2003). HPLC and mass spectrometery data from culture
filtrates after the decolorization of reactive red 22 by P leuteola, showed the presence of twoaromatic amines, as well as a partially redused product (Chang et al., 2001). This is in
accordance with the two-step reduction mechanism of the Azo bond proposed by Gingell and
Walker (1971).
2.3. Azo dye decolorization under aerobic conditions
Conventional activated sludge treatment of wastes is often an effective and highly economic
system for reducing organic pollutants in wastewater. A fair amount of research has beenconducted assessing the viability of using activated sludge to treat textile effluents (Zissi et al.,
1997; Loyd, 1992; Shaul et al., 1991; Pagga and Brown, 1986). However, aerobic treatment of
Azo dye wastes has proven ineffective in most cases, but is often the typical method of treatmentused today (Edwards, 2000; Yang et al., 1998). Because aerobic microbes cannot reduce Azo

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 5/16
linkages, their ability to destroy dye chromogens is less than anaerobic bacterium. However,
aerobic sludges have been successfully used to stabilize dye metabolites (Brown and Laboureur,
1983). Several bacterial strains that can aerobically decolorize Azo dyes have been isolatedduring the past few years. Many of these strains require organic carbon sources, as they cannot
utilize dye as the growth substrate (Stolz, 2001). P. aeruginosa decolorized a commercial tannery
and textile dye, Navitan Fast blue S5R, in the presence of glucose under aerobic conditions. Thisorganism was also able to decolorize various other Azo dyes (Nachiyar and Rajkumar, 2003). There are only very few bacteria that are able to grow on Azo compounds as the sole carbon
source. These bacteria cleave – N=N – bonds reductively and utilize amines as the source of
carbon and energy for their growth.Such organisms are specific towards their substrate. Examples of bacterial strains with this
trait are Xenophilus Azovorans KF 46 (previously Pseudomonas sp. KF46) and Pigmentiphaga
kullae K24 (previously Pseudomonas sp. K24), which can grow aerobically on carboxy-orange I
and carboxy-orange II, respectively (Zimmermann et al., 1982; Kulla et al., 1983). Theseorganisms, however, could not grow on structurally analogous sulfonated dyes, acid orange 20
(Orange I) and AO7. Long adaptation of 4-aminobenzenesulfonate (4-ABS) degrading
Hydrogenophaga intermedia strain S1 for growth on 4-carboxy-40-sulfoAzobenzene (CSB) asthe sole organic carbon source led to the isolation of strain S5, which reduced CSB and utilized
the two amine metabolites (Blumel et al., 1998). Coughlin et al. (1999) have reported the
isolation of a Sphingomonas sp, strain 1CX, an obligate aerobe, which can grow on an Azo dye,
AO7, as sole carbon, energy and nitrogen source. This strain degraded only one of thecomponent amines (1-amino 2-naphthol) formed during AO7 decolorization. 4-
aminobenzesulfonate (4-ABS) degradation, however, required the additional presence of an
unidentified strain, SAD4i (Coughlin et al., 2003). Sphingomonas ICX could also decolorizeseveral Azo dyes consisting of either 1-amino-2-naphthol or 2-amino-1- naphthol coupled via the
Azo bond to a phenyl or naphthyl moiety (Coughlin et al., 1999). Similar Azo dyes, such as AO6
or AO20, which lack these structures, were not decolorized.
Three bacterial strains that could utilize Azo dye (AO 7 or acid red 88) as sole carbon sourcewere isolated from soil and sewage samples and were identified as Bacillus sp. OY1-2,
Xanthomonas sp. NR25-2 and Pseudomonas sp. PR41-1. (Sugiura et al., 1999). Recently, four
bacterial species have been isolated using methyl red as the sole carbon source. Two of thesestrains have been identified as Vibrio logei and P. nitroreducens. Amine products were not
detected in the culture medium, indicating their degradation (Adedayo et al., 2004). The
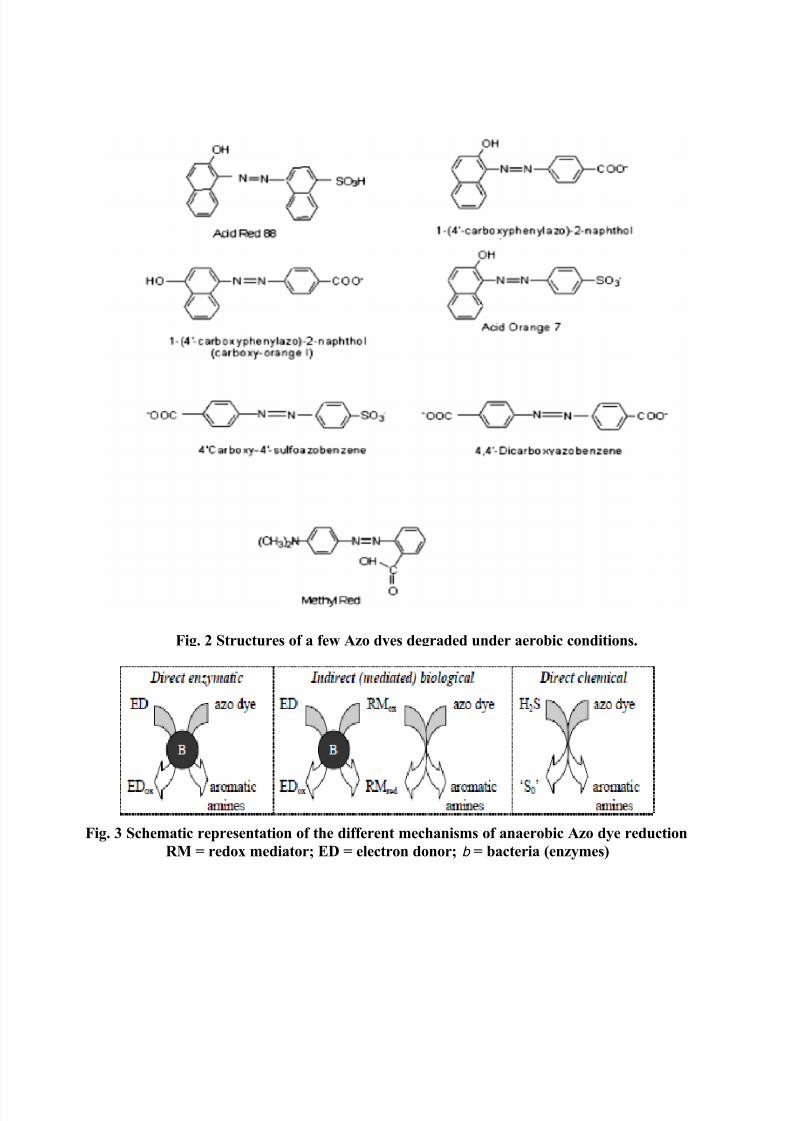
structures of few Azo dyes that are mineralized under aerobic conditions are presented in Fig. 2.
2.4. Mechanism of Azo dye reduction
The first step in the bacterial degradation of Azo dyes, in either anaerobic or aerobicconditions, is the reduction of the – N=N – bond. This reduction may involve different
mechanisms, such as enzymes, low molecular weight redox mediators, chemical reduction by
biogenic reductants like sulfide, or a combination of these (Fig. 3). Additionally, the location of
the reactions can be either intracellular or extracellular.
7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 6/16
Fig. 2 Structures of a few Azo dyes degraded under aerobic conditions.
Fig. 3 Schematic representation of the different mechanisms of anaerobic Azo dye reduction
RM = redox mediator; ED = electron donor; b = bacteria (enzymes)

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 7/16
2.4.1. Direct enzymatic Azo dye reduction
According to the first mechanism of biological Azo dye reduction, enzymes transfer the
reducing equivalents originating from the oxidation of organic substrates to the Azo dyes.
Enzymes that catalyze Azo dye reduction may either be specialized enzymes (catalysing only thereduction of Azo dyes) or non-specialized enzymes (non-specific enzymes that catalyze thereduction of a wide range of compounds, including Azo dyes). Evidence for the existence of
specialized Azo dye reducing enzymes, so-called .Azoreductases., has so far only been found in
studies with some aerobic and facultative aerobic bacteria that could grow with mostly simpleAzo compounds as sole source of carbon and energy. These strains grew under strict aerobic
conditions by using a metabolism that started with reductive cleavage of the Azo linkage (Ince
and Gonenc, 1997). The existence of enzymes catalysing Azo dye reduction in aerobic bacteria
was for the first time proven when two Azoreductases from obligate aerobic bacteria wereisolated and characterised (Zimmermann et al., 1984: Zimmermann et al., 1982). These
intracellular Azoreductases showed high specificity to dye structures. Aside from these specific
Azoreductases, also non-specific enzymes catalysing Azo dye reduction have been isolated fromaerobically grown cultures of Shigella dysenteriae (Ghosh et al., 1992) Escherichia coli (Ghosh
et al., 1993) and Bacillus sp. (Suzuki et al., 2001) . Where characterized enzymes were found to
be flavoproteins (Ghosh et al., 1993; Ghosh et al., 1992; Rafii and Cerniglia, 1995)
2.4.2. Indirect (mediated) biological Azo dye reduction.
According to the second mechanism of biological Azo dye reduction, Azo dyes are indirectlyreduced by enzymatically reduced electron carriers. Early research has hypothesised that reduced
flavins (FADH2, FMNH2, riboflavin) generated by flavindependent reductases can reduce Azo
dyes in a non-specific chemical reaction (Gingell and Walker, 1971; Roxon, 1967 ). Flavins were
indeed often found to stimulate Azo dye reduction and recent research has revealed that flavinreductases are indeed .anaerobic Azoreductases (Russ et al., 2000). Also other reduced enzyme
cofactors capable of direct Azo dye reduction have been reported, e.g. NADH ( Nam and
Renganathan, 2000), NADH and NADPH (Hernandez et al., 1967) and an NADPH-generatingsystem (Semdé et al., 1998). Aside from enzyme cofactors, also various artificial redox
mediating compounds are important stimulants of biological Azo dye reduction (methyl
viologen, benzyl viologen, Riboflavin, FAD, FMN, Phenosaphranin, Menadione, Neutral Red,2-hydroxy-1,4-naphthoquinone) (Chung et al., 1978).
2.4.3 Chemical Azo dye reduction
Textile manufacturers are aware that addition of reducing agents to dye containing effluents
leads to considerable decolorization (Glover, 1993). Hence, Azo dye can be reduced by chemical
reductants like dithionite (Davis et al., 1993; Zbaida and Levine, 1992) and zerovalent iron
( Nam and Renganathan, 2000). Moreover, chemical Azo dye reduction by biogenic reductantslike sulphide, is possible as well (Yoo et al., 2000). Dye-containing wastewaters usually contain
moderate to high sulphate concentrations. Sulphate is often an additive of dye baths or it is
formed by the oxidation of more reduced sulphur species used in dyeing processes, such assulphide, hydrosulphite and dithionite. Sulphate also results from neutralization of alkaline dye

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 8/16
effluents with sulphuric acid. Sulphide is therefore a relevant compound, as it will be generated
by sulphate reducing bacteria during treatment of these wastewaters in anaerobic bioreactors.
3. Degradation of aromatic amines
Aromatic compounds possess a large negative resonance energy, resulting in thermodynamicstability. Microorganisms, particularly bacteria, have evolved enzyme systems that degrade the benzene structure under aerobic and anoxic conditions (Gibson and Subramanian, 1984; Schink
et al., 2000). Common to both oxygen-dependent and anoxic metabolism of aromatic compounds
is a separation into peripheral and central pathways (Heider and Fuchs, 1997). Peripheral pathways convert the large variety of compounds into a few central intermediates. In aerobic
metabolism, the initial reactions involve the replacement of other functional groups of the
aromatic ring with hydroxyl groups, followed by cleavage by incorporating two oxygen atoms.
These reactions are catalysed by hydroxylases and oxygenases. Under anoxic conditions,dearomatization is achieved by ring reduction and also includes other unique reactions such as
carboxylation, reductive dehydroxylation and addition reactions, which are absent in the aerobic
metabolism (Heider and Fuchs, 1997).
3.1. Degradation of aromatic amines under anaerobic conditions
Decolorization of Azo dyes in anaerobic environments leads to the formation of aromaticamines, many of which were assumed to resist further degradation under these conditions (Stolz,
2001). Nevertheless, mineralization of few simple aromatic amines has been reported under
methanogenic conditions. They include the three isomers of aminobenzoate, 2- and 4-aminophenols, 2, 4 dihydroxyaniline and 5-aminosalicylic acid (5-ASA). (Connor and Young,
1993; RAzo-Flores et al., 1997b; Yemashova et al., 2004). Complete degradation of Azo
disalicylate and partial mineralization of the Azo dyes, Mordent Orange 1, AO6, AO7 and
AO52, under methanogenic conditions, has been reported (Donlon et al., 1997; RAzo-Flores etal., 1997b; Yemashova et al., 2004). Many reports have shown that sulfonated aromatic amines
(SAA) are nonbiodegradable under methanogenic conditions (Tan et al., 2005).
3.2. Aerobic fate of aromatic amines
The aerobic biodegradation of many aromatic amines has been extensively studied. Many ofthese compounds were found to be degraded under aerobic conditions (Baird et al. 1977; Brown
and Laboureur 1983a, Albayati, 2010 and Aldoury et al., 2014,), e.g. compounds like aniline
(Anson and MacKinnon 1984; Konopka 1993), carboxylated aromatic amines ( Stolz et al.
1992), chlorinated aromatic amines (Hwang et al. 1987; Loidl et al. 1990), and (substituted) benzidines (Baird et al. 1977). Several of the studies reveal evidence for partial or complete
removal of many aromatic amines in the aerobic stage. The decrease or disappearance of the,
sometimes unidentified, peaks in HPLC chromatograms (Harmer and Bishop, 1992; FitzGerald
and Bishop, 1995; Tan et al., 2000; Isik and Sponza, 2004a), the decrease or disappearance ofaromatic amines as detected with a diAzotization-based method ( Isik and Sponza, 2004c; Isik
and Sponza, 2004a; Sponza and Isik, 2005), as well as the decrease of UV absorbance (Cruz and
Buitro´ n, 2001; Shaw et al., 2002) all indicate removal of aromatic amines. Moreover, the largedecreases of toxicity (mostly suppression of bacterial luminescence or inhibition of respiration)

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 9/16
between the effluent of the anaerobic stage and the effluent of the anaerobic stage (O’Neill et al.,
2000b; Frijters et al., 2004) or between the effluent of a completely anaerobic reactor and the
effluent of a combined anaerobic – aerobic reactor (Gottlieb et al., 2003) provide indirect evidencefor the removal of aromatic amines. The results taken as a whole suggest that many of the
aromatic amines from anaerobic cleavage of Azo dyes were removed in the consequent aerobic
stage. However, some aromatic amines may not be removed. Especially cleavage products fromthe reactive Azo dyes Reactive Black 5 and Reactive Violet 5 were often reported not to beremoved aerobically (Lourenc-o et al., 2000; Lourenc-o et al., 2001; Shaw et al., 2002; Lourenc-
o et al., 2003). Also a relatively large fraction (50%) of the aromatic amines from the benzidine
based dye Direct Black 38 resisted removal in the aerobic stage (Sponza and Isik, 2005). Most ofthe studies reporting aromatic amine removal do not reveal the underlying mechanism.
As aerobic biodegradation of aromatic amines requires specific microorganisms, the type of
biomass may play a role. At least in one laboratory reactor study, the degradation of an aromatic
amine, sulfanilic acid, could only be achieved after bioaugmentation with a proper bacterialculture (Tan et al., 2000). Also the observation that introduction of biomass from a textile waste-
treating water works decreased the toxicity of the effluent from an Azo dye treating baffled
reactor (Gottlieb et al., 2003) suggests the involvement of specific bacteria. In many of thestudies reporting aromatic amine removal, it is not clear whether the removal is due to
biodegradation, adsorption or chemical reactions. Remarkably, although autoxidation of aromatic
amines during aerobic treatment, as suggested by an increase of color, has been observed in
some studies (Kalyuzhnyi and Sklyar, 2000; Cruz and Buitro´ n, 2001), a slight decrease of thecolor was much more often observed (An et al., 1996 ; Kuai et al., 1998; O’Neill et al., 2000b;
Shaw et al., 2002; Kapdan et al., 2003; Isik and Sponza, 2004c; Sponza and Isik, 2004; Kapdan
and Alparslan, 2005; Ong et al., 2005). Since many of the Azo dyes treated in these studies yieldaromatic amines that are expected to autoxidize, the latter observation suggests, in several cases,
removal of these compounds or their autoxidation products from the water phase.
4. Conclusion
Among the synthetic dyes, which are widely used for textile dyeing and other industrialapplications, those containing an Azo chromosphere constitute the largest class. Since dyes are
design to be resistant to microbial and physico-chemical attack, conventional processes of
wastewater treatment, including biological treatment, do not easily destroy most of them. Theavailable literature seems to indicate that anaerobic – aerobic biological methods may be
appropriate for the treatment of dye-containing wastewaters. However, there is a still a need to
assess the extent of mineralization of aromatic amines, as many amines can undergo
autoxidation, leading to the formation of soluble recalcitrant polymers, which may be toxic.Degradation of many amines, including SAA, requires the presence of specialized cultures. SAA
degraders have a very narrow substrate range. Hence, there is a requirement for developing
microbial consortia that harbor genes for the rapid degradation of mixtures of aromatic amines.
Such cultures may have to be used for the bioaugmentation of aerobic treatment units. Molecular biology techniques may also be used to improve the strains so that rapid mineralization of
aromatic amines can be achieved. Their use, however, requires caution. It may also be necessary
to combine AOP with biological processes to achieve the required degree of treatment of dye-containing wastewaters so that regulatory standards can be met

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 10/16
References
Adedayo, O., Javadpour, S., Taylor, C., Anderson, W.A., Moo-Young, M., (2004).Decolorization and detoxification of methyl red by aerobic bacteria from a wastewater
treatment plant. World Journal of Microbiology and Biotechnology 20, 545 – 550.
Albayati, M. B. (2010). Anaerobic/Aerobic Biological Treatment of Synthetic WastewaterContaining Two Types of Azo Dye. MSc Thesis College of Engineering, Tikrit University,Iraq
Aldoury, M. M. , Alabdraba, Waleed M. Sh. and Albayati , M. B. (2014). Performance of
Sequential Anaerobic/Aerobic Biological Treatment of Synthetic Wastewater Containing TwoTypes of Azo Dye. 2ed International Conference of Environmental Science and Technology
ICOEST2014 - Side, Turkey
An H., Yi Q., Xiasheng G. and Walter Z.T. (1996). Biological treatment of dye wastewaters
using an anaerobic oxic system. Chemosphere 33(12): 2533-2542. An, H., Qian, Y., Gu, X.S., Tang, W.Z., 1996. Biological treatment of dye wastewaters using an
anaerobic – oxic system. Chemosphere 33 (12), 2533 – 2542.
Anson J.G. and MacKinnon G. (1984). Novel Pseudomonas plasmid involved in aniline degradation. Applied and Environmental Microbiology 48(4): 868-869.
Baird, R., Camona, L. and Jenkins, R.L. (1977) Behavior of benzidine and other aromatic amines
in aerobic wastewater treatment. J. Water Pollut. Control Fed., 49: 1609-1615.
Bell J. and Buckley, C. A. (2003). Treatment of a textile dye in the anaerobic baffled reactor. Water SA Vol. 29 No. 2
Blumel, S., Contzen, M., Lutz, M., Stolz, M., Knackmuss, H.J., (1998). Isolation of a bacterial
strain with the ability to utilize the sulfonated Azo compound 4-carboxy-40- sulfoAzobenzene as sole source of carbon and energy. Applied and Environmental Microbiology 64, 2315 –
2317.
Brown, J.P. (1981). Reduction of polymeric Azo and nitro dyes by intestinal bacteria. Appl.
Environ. Microbiol., 41: 1283-1286. Brown, D. and Laboureur, P. (1983) The degradation of dyestuffs: Part I – Primary
biodegradation under anaerobic conditions. Chemosphere, 12: 397-404.
Brown D. and P. Laboureur. 1983a. The Degradation of Dyestuffs: Part I. PrimaryBiodegradation under Anaerobic Conditions. Chemosphere, 12(3): 397-404.
Brown D. and Laboureur P. (1983a). The aerobic biodegradability of primary aromatic amines.
Chemosphere 12(3): 405-414. Brown D. and Laboureur P. (1983b). The degradation of dyestuffs: Part I – Primary
biodegradation under anaerobic conditions. Chemosphere 12(3): 397-404.
Brown D. and Hamburger B. (1987). The degradation of dyestuffs: Part III - Investigations of
their ultimate degradability. Chemosphere 16(7): 1539-1553. Brohm, K. and Frohwein, E. (1937) Nachweis von durch Säueringentfarbten künstlichen
Eigelbfrabstoffen in Milchspeiseeis. Zbl. Lebensmitt. Forsch., 73: 30.
Bromly-Challenor, K.C.A., Knapp, J.S., Zhang, Z., Gray, N.C.C., Hetheridge, M.J. and Evans,
M.R. (2000) Decolorization of an Azo dye by unacclimated activated sludge under anaerobic conditions. Water Res., 34: 4410-4418.
Carliell C.M., Godefroy S.J., Naidoo N., Buckley C.A., Senior E., Mulholland D. and Martineigh
B.S. (1994). Anaerobic decolourisation of Azo dyes, Seventh International Symposium on

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 11/16
Anaerobic Digestion, RSA Litho (Pty) Ltd, Goodwood, South Africa, Cape Town, South
Africa, pp. 303-306
Carliell C.M., Barclay S.J., Naidoo N., Buckley C.A., Mulholland D.A. and Senior E. (1995).Microbial decolourisation of a reactive Azo dye under anaerobic conditions. Water SA 21(1):
61-69.
Carliell, C.M., Barclay, S.J., Naidoo, N., Buckley, C.A., Mulholland, D.A. and Senior, E. (1994) Anaerobic decolorisation of reactive dyes in conventional sewage treatment processes. Water SA, 20: 341-344
Chang, J.S., Chou, C., Lin, Y., Ho, J., Hu, T.L., (2001). Kinetic Characteristics of bacterial Azo-
dye decolorization by Pseudomonas luteola. Water Research 35, 2041 – 2850. Chen, K.C., Huang, W.T., Wu, J.Y., Houng, J.Y., 1999. Microbial decolorization of Azo dyes by
Proteus mirabilis. Journal of Industrial Microbiology and Biotechnology 23, 686 – 690
Chen, K.C., Wu, J.Y., Liou, D.J., Hwang, S.J., 2003. Decolorization of textile dyes by newly
isolated bacterial strains. Journal of Biotechnology 101, 57 – 68. Chung, K.T., Fulk, G.E. and Egan, M. (1978) Reduction of Azo dyes by intestinal anaerobes.
Appl. Environ. Microbiol., 35: 558-562
Chung, K.T., Fulk, G.E. and Egan, M. (1978) Reduction of Azo dyes by intestinal anaerobes. Appl. Environ. Microbiol., 35: 558-562.
Chung, K.T. and Cerniglia, C.E. (1992) Mutagenicity of Azo dyes: Structure-activity
relationships. Mutat. Res., 277: 201-220.
Chung K.T. and Cerniglia C.E. (1992). Mutagenicity of Azo dyes: Structure-activityrelationships. Mutation Research 277: 201-220.
Coughlin, M.F., Kinkle, B.K., Bishop, P.L., (1999). Degradation of Azo dyes containing amino
naphthol by Sphingomonas sp. strain ICX. Journal of Industrial Microbiology and Biotechnology 23, 341 – 346.
Coughlin, M.F., Kinkle, B.K., Bishop, P.L.,( 2003). High performance degradation of Azo dye
acid orange 7 and sulfanilic acid in a laboratory scale reactor after seeding with cultured
bacterial strains. Water Research 37, 2757 – 2763. Connor, O.A., Young, L.Y., (1993). Effect of nitrogen limitation on the biodegradability and
toxicity of nitro and aminophenol isomers to methanogenesis. Archives of Environment
Contamination and Toxicology 25, 285 – 291. Cruz, A., Buitro´ n, G., (2001). Biodegradation of Disperse Blue 79 using sequenced
anaerobic/aerobic biofilters. Water Sci. Technol. 44 (4), 159 – 166.
Davis, V.M. and Bailey, J.E.J. (1993) Chemical reduction of FD and C yellow No. 5 to determine combined benzidine. J. Chromatogr., 635: 160-164.
Donlon B.A., RAzo-Flores E., Luijten M., Swarts H., Lettinga G. and Field J.A. (1997).
Detoxification and partial mineralization of the Azo dye mordant orange 1 in a continuous
upflow anaerobic sludge-blanket reactor. Applied Microbiology and Biotechnology 47(1): 83-90.
Donlon, B., RAzo-Flores, E., Luijten, M., Swarts, H., Lettinga, G., Field, J.A., (1997).
Detoxification and partial mineralization of the Azo dye mordant orange 1 in a continuous
upflow anaerobic sludge — blanket reactor. Applied Microbiology and Biotechnology 47, 83 – 90.
Edwards, Jesscia C. (2000). Investigation of Color Removal by Chemical Oxidation for Three
Reactive Textile Dyes and Spent Textile Dye Wastewater. Masters Thesis, Virginia Polytechnic Institute and State University. 56 pp.

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 12/16
Esenceli, N. and Tiyek, I. (2014). Investigation of New Techniques Used in the Removal of Dyes
in Textile Wastewater. 2ed International Conference of Environmental Science and
Technology ICOEST2014 - Side, Turkey Field J.A., Stams A.J.M., Kato M. and Schraa G. (1995). Enhanced biodegradation of aromatic
pollutant in coculture of anaerobic and aerobic bacterial consortia. Antonie van Leeuwenhoek
67: 47-77. FitzGerald S.W. and Bishop P.L. (1995). Two stage anaerobic/aerobic treatment of sulfonated Azo dyes. Journal of Environmental Science and Health A30(6): 1251-1276.
Frijters, C.T.M.J., Vos, R.H., Scheffer, G., Mulder, R., 2004. Decolorizing and detoxifying
textile wastewater in a fullscale sequential anaerobic/aerobic system. In: Guiot, S.R. (Ed.),Proceedings of the 10th World Congress on Anaerobic Digestion. Anaerobic bioconversiony
Answer for Sustainability. 29 August – 2 September 2004, Montre´ al, Canada. IWA, Vol. 4,
pp. 2387 – 2390.
Ghosh, D.K., Mandal, A. and Chaudhuri, J. (1992) Purification and partial characterization of two Azoreductases from Shigella dysenteriae type 1. FEMS Microbiol. Lett., 98: 229-234.
Ghosh, D.K., Ghosh, S., Sadhukhan, P., Mandal, A. and Chaudhuri, J. (1993) Purification of two
Azoreductases from Escherichia coli K12. Ind. J. Exp. Biol., 31: 951-954. Glover, B. (1993) Getting rid of colour. J. Soc. Dyers Colour., 109: 273.
Gottlieb, A., Shaw, C., Smith, A., Wheatley, A., Forsythe, S., (2003). The toxicity of textile
reactive Azo dyes after hydrolysis and decolourisation. J. Biotechnol. 101 (1), 49 – 56.
Gingell R. and Walker R. (1971). Mechanisms of Azo reduction by Streptococcus faecalis II. The role of soluble flavins. Xenobiotica 1(3): 231-239.
Gibson, D.T., Subramanian, V., (1984). In: Gibson, D.T. (Ed.), Microbial Degradation of
Aromatic Hydrocarbons, in Microbial Degradation of Organic Compounds. Marcel Dekker, New York, pp. 181 – 252.
Harmer C. and Bishop P. (1992). Transformation of Azo dye AO-7 by wastewater biofilms.
Water Science Technology 26(3/4): 627-636
Haug, W., Schmidt, A., Nortemann, B., Hempel, D.C., Stolz, A. and Knackmuss, H.J. (1991)Mineralization of the sulfonated Azo dye Mordant Yellow 3 by a 6-aminonaphthalene-
2- sulfonate-degrading bacterial consortium. Appl. Environ. Microbiol., 57: 3144-3149.
Heider, J., Fuchs, G., (1997). Anaerobic metabolism of aroimatic compounds. European Journalof Biochemistry 243, 577 – 59
Hernandez, P.H., Gilette, R. and Mazel, P. (1967) Studies on the mechanism of action of
mammalian hepatic Azoreductase. I. Azoreductase activity of reduced nicotinamide adeninedinucleotide phosphate-cytochrome c reductase. Biochem. Pharmacol., 16: 1859-1875.
Hwang H.M., Hodson R.E. and Lee R.F. (1987). Degradation of aniline and chloroaniline by
sunlight and microbes in estuarine water. Water Research 21(3): 309-316.
Ince, N.H. and Gonenc, D.T. (1997) Treatability of a textile Azo dye by UV/H-2O-2. Environ. Technol., 18: 179-185.
Isik, M., Sponza, D.T., (2004a). Anaerobic/aerobic sequential treatment of a cotton textile mill
wastewater. J. Chem. Technol. Biotechnol. 79 (11), 1268 – 1274.
Isik, M., Sponza, D.T., (2004c). Monitoring of toxicity and intermediates of C.I. Direct Black 38Azo dye through decolorization in an anaerobic/aerobic sequential reactor system. J. Hazard.
Mater. 114 (1 – 3), 29 – 39.
Kalyuzhnyi, S., Sklyar, V., (2000). Biomineralisation of Azo dyes and their breakdown products in anaerobic – aerobic hybrid and UASB reactors. Water Sci. Technol. 41 (12), 23 – 30.

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 13/16
Kapdan, I.K., Kargi, F., McMullan, G., Marchant, R., (2000). Decolorization of textile dye stuffs
by a mixed bacterial consortium. Biotechnology Letters 22, 1179 – 1181.
Kapdan, I.K., Tekol, M., Sengul, F., (2003). Decolorization of simulated textile wastewater in an anaerobic – aerobic sequential treatment system. Process Biochem. 38 (7), 1031 – 1037.
Kapdan, I.K., Alparslan, S., (2005). Application of anaerobic – aerobic sequential treatment
system to real textile wastewater for color and COD removal. Enzyme Microb. Technol. 36 (2 – 3), 273 – 279. Keck A., Klein J., Kudlich M., Stolz A., Knackmuss H.J. and Mattes R. (1997). Reduction of
Azo dyes by redox mediators originating in the naphthalenesulfonic acid degradation pathway
of Sphingomonas sp. Strain BN6. Applied and Environmental Microbiology 63(9): 3684-3690.
Khehra, M.S., Saini, H.S., Sharma, D.K., Chadha, B.S., Chimni, S.S., (2005). Decolorization of
various Azo dyes by bacterial consortia. Dyes and Pigments 67, 55 – 61.
Konopka A. (1993). Isolation and characterization of a subsurface bacterium that degrades aniline and methylanilines. FEMS Microbiology Letters 111(1): 93-99.
Kuai, L., De Vreese, I., Vandevivere, P., Verstraete, W., (1998). GAC-amended UASB reactor
for the stable treatment of toxic textile wastewater. Environ. Technol. 19 (11), 1111 – 1117 Kudlich M., Keck A., Klein J. and Stolz A. (1997). Localization of the enzyme system involved
in anaerobic reduction of Azo dyes by Sphingomonas sp. Strain BN6 and effect of artificial
redox mediators on the rate of Azo dye reduction. Applied and Environmental Microbiology
63(9): 3691-3694. Kulla, H.G., Klausener, F., Meyer, O., Ludeke, B., Leisinger, T., (1983). Interference of aromatic
sulfo groups in the microbial degradation of the Azo dyes orange I and orange II. Archives of
Microbiology 135, 1 – 7. Lourenc-o, N.D., Novais, J.M., Pinheiro, H.M., (2000). Reactive textile dye colour removal in a
sequencing batch reactor. Water Sci. Technol. 42 (5 – 6), 321 – 328.
Lourenc-o, N.D., Novais, J.M., Pinheiro, H.M., (2001). Effect of some operational parameters on
textile dye biodegradation in a sequential batch reactor. J. Biotechnol. 89 (2 – 3), 163 – 174. Lourenc-o, N.D., Novais, J.M., Pinheiro, H.M., (2003). Analysis of secondary metabolite fate
during anaerobic – aerobic Azo dye biodegradation in a sequential batch reactor. Environ.
Technol. 24 (6), 679 – 686. Loidl M., Hinteregger C., Ditzelmuller G., Ferschl A. and Streichsbier F. (1990). Degradation of
aniline and monochlorinated anilines by soil-born Pseudomonas acidovorans strains.
Archives of Microbiology 155(1): 56-61. Loyd, Chapman K. (1992). Anaerobic/Aerobic Degradation of a Textile Dye Wastewater.
Masters Thesis, Virginia Polytechnic Institute and State University. 184 pp.
Moosvi, S., Keharia, H., Madamawar, D.,( 2005). Decolorization of textile dye reactive violet by
a newly isolated bacterial consortium. RVM 11.1. World Journal of Microbiology and Biotechnology 21, 667 – 672.
Nachiyar, C.V., Rajkumar, G.S., (2003). Degradation of tannery and textile dye, Navitan Fast
Blue S5R by Pseudomonas aeruginosa. World Journal of Microbiology and Biotechnology 19,
609 – 614. Nam, S. and Renganathan, V. (2000) Non-enzymatic reduction of Azo dyes by NADH.
Chemosphere, 40: 351-357.

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 14/16
Nigam, P., Banat, I.M., Singh, D., Marchant, R., (1996). Microbial process for the
decolonization of textile effluent containing Azo, diAzo and reactive dyes. Process
Biochemistry 31, 435 – 442. Ong, S.A., Toorisaka, E., Hirata, M., Hano, T., (2005). Decolorization of Azo dye (Orange II) in
a sequential UASB – SBR system. Sep. Purif. Technol. 42 (3), 297 – 302.
O’Neill, C., Lopez, A., Esteves, S., Hawkes, F.R., Hawkes, D.L., Wilcox, S., (2000b). Azo-dye degradation in an anaerobic – aerobic treatment system operating on simulated textile effluent. Appl. Microbiol. Biotechnol. 53 (2), 249 – 254.
Pagga U. and D. Brown. (1986). The Degradation of Dyestuffs: Part II. Behaviour of Dyestuffs
in Aerobic Biodegradation Tests. Chemosphere, 15(4): 479-491 Padmavathy, S., Sandhya, S., Swaminathan, K., Subrahmanyam, Y.V., Kaul, S.N., (2003).
Comparison of decolorization of reactive Azo dyes by microorganisms isolated from various
sources. Journal of Environmental Science (China) 15, 628 – 633, 4184.
Rafii, F., Franklin, W. and Cerniglia, C.E. (1990) Azoreductase activity of anaerobic bacteria isolated from human intestinal microflora. Appl. Environ. Microbiol., 56: 2146-2151.
Rafii, F. and Cerniglia, C.E. (1995) Reduction of Azo dyes and nitroaromatic compounds by
bacterial enzymes from the human intestinal tract. Environ. Health Persp., 103: 17-19. RAzo-Flores E., Donlon B.A., Field J.A. and Lettinga G. (1996). Biodegradability of
Nsubstituted aromatics and alkylphenols under methanogenic conditions using granular
sludge. Water Science and Technology 33(3): 47-57.
RAzo-Flores E., Luijten M., Donlon B.A., Lettinga G. and Field J.A. (1997). Complete biodegradation of the Azo dye Azodisalicylate under anaerobic conditions. Environmental
Science and Technology 31(7): 2098-2103.
RAzo-Flores, E., Luijten, M., Donlon, B., Lettinga, G., Field, J., (1997b). Complete biodegradation of the Azo dye Azosalicylate under anaerobic conditions. Environmental
Science and Technology 31, 2098 – 2103.
Roxon, J.J., Ryan, A.J. and Wright, S.E. (1967) Enzymatic reduction of tartrazine by Proteus
vulgaris from rats. Food Cosmet. Toxicol., 5: 645-656. Russ, R., Rau, J. and Stolz, A. (2000) The function of cytoplasmic flavin reductases in the
reduction of Azo dyes by bacteria. Appl. Environ. Microbiol., 66: 1429-1434.
Schink, B., Philipp, B., Muller, J., (2000). Anaerobic degradation of phenolic compounds. Review Naturwissenschaften 87, 12 – 23.
Seshadri S., Bishop P.L. and Agha A.M. (1994). Anaerobic/aerobic treatment of selected Azo
dyes in wastewater. Waste Management 14(2): 127-137. Semdé, R., Pierre, D., Geuskens, G., Devleeschouwer, M. and Moes, A.J. (1998) Study of some
important factors involved in Azo derivative reduction by Clostridium perfringens . Int. J.
Pharm., 161: 45-54.
Shaul G.M., Holdsworth T.J., Demmpsey C.R. and Dostal K.A. (1991). Fate of water soluble Azo dyes in the activated sludge process. Chemosphere 22: 107-119.
Shaw, C.B., Carliell, C.M., Wheatley, A.D., (2002). Anaerobic/ aerobic treatment of coloured
textile effluents using sequencing batch reactors. Water Res. 36 (8), 1993 – 2001.
Stolz, A., (2001). Basic and applied aspects in the microbial degradation of Azo dyes. Applied Microbiology and Biotechnology 56, 69 – 80.
Sugiura, W., Miyashita, T., Yokoyama, T., Arai, M., (1999). Isolation of Azo-dye degrading
microorganisms and their application to whitedischarge printing of fabric. Journal of Bioscience and Bioengineering 88, 577 – 581.

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 15/16
Suzuki, Y., Yoda, T., Ruhul, A. and Sugiura, W. (2001) Molecular cloning and characterization
of the gene coding for Azoreductase from Bacillus sp OY1-2 isolated from soil. J. Biol.
Chem., 276 (12): 9059-9065. Stolz A., Nortemann B. and Knackmuss H.J. (1992). Bacterial metabolism of 5- aminosalicylic
acid: Initial ring cleavage. Biochemical Journal 282(3): 675-680.
Stolz, A., (2001). Basic and applied aspects in the microbial degradation ofAzo dyes. AppliedMicrobiology and Biotechnology 56, 69 – 80.Sponza, D.T., Is-ik, M., (2005). Reactor performances and fate of aromatic amines through
decolorization of Direct Black 38 dye under anaerobic/aerobic sequentials. Process Biochem.
40 (1), 35 – 44.Sponza, D.T., Is-ik, M., (2004). Decolorization and inhibition kinetic of Direct Black 38 Azo dye
with granulated anaerobic sludge. Enzyme Microb. Technol. 34 (2), 147 – 158.
Tan N.C.G. and Field J.A. (2000). Biodegradation of sulfonated aromatic compounds. In:
Environmental technologies to treat sulfur pollution. Principles and engineering, Lens P. andHulshoff Pol L., IWA publishing, London, pp. 377-392.
Tan, N.C.G., van Leeuwen, A., van Voorthuizen, E.M., Slenders, P., Prenafeta-Boldu, F.X.,
Temmink, H., Lettinga, G., Field, J.A.,( 2005). Fate and biodegradability of sulfonated aromatic amines. Biodegradation 16, 527 – 537.
Vaidya A.A. and Datye K.V. (1982). Environmental pollution during chemical processing of
synthetic fibers. Colourage 14: 3-10.
Walker, R. (1970) The metabolism of Azo compounds: a review of the literature. Food Cosmet. Toxicol., 8: 659-676.
Weber, E.J. and Wolfe, L.N. (1987) Kinetic studies of the reduction of aromatic Azo compounds
in anaerobic sediment/water systems. Environ. Toxicol. Chem., 6: 911-919. Weber E.J. (1991). Studies of benzidine-based dyes in sediment-water systems. Environmental
Toxicology and Chemistry 10(5): 609-618.
Wuhrmann, K., Mechsner, K. and Kappeler, T. (1980) Investigation on rate-determining factors
in the microbial reduction of Azo dyes. Eur. J. Appl. Microbiol. Biotechnol., 9: 325-338. Yang Y., Wyatt D.T. II, Bahorshky M. (1998). Decolorization of Dyes Using UV/H2O2
Photochemical Oxidation. Textile Chemist and Colorist, 30:27-35.
Yemashova, N., Telegina, A., Kotova, I., Netrusova, A., Kalyuzhnyi, S., (2004). Decolorization and partial degradation of selected Azo dyes by methanogenic sludge. Applied Biochemistry
and Biotechnology 119, 31 – 40.
Yoo, E.S., Libra, J. and Wiesmann, U. (2000) Reduction of Azo dyes by Desulfovibrio desulfuricans. Water Sci. Technol., 41: 15-22.
Yu, J., Wang, X., Yue, P.L., (2001). Optimal decolorization and kinetic modeling of synthetic
dyes by Pseudomonas strains. Water Research 35, 3579 – 3586.
Zaoyan Y., Ke S., Guangliang S., Fan Y., Jinshan D. and Huanian M. (1992). Anaerobicaerobic treatment of dye waste-water by combination with activated sludge. Water Science and
Technology 26: 2093-2096.
Zbaida, S. and Levine, W.G. (1992) Role of electronic factors in binding and reduction of Azo
dyes by hepatic microsomes. J. Pharmacol. Exp. Ther ., 260: 554-561. Zimmermann, T., Kulla, H. and Leisinger, T. (1982) Purification and properties of orange
IIAzoreductase from Pseudomonas KF46. Experientia, 38: 1380.

7/25/2019 Biodegradation of Azo Dyes
http://slidepdf.com/reader/full/biodegradation-of-azo-dyes 16/16
Zimmermann, T., Kulla, H., Leisinger, T., (1982). Properties of purified orange II-Azoreductase,
the enzyme initiating Azo dye degradation by Pseudomonas KF46. European Journal of
Biochemistry 129, 197 – 203. Zimmermann, T., Gasser, F., Kulla, H. and Leisinger, T. (1984) Comparison of two bacterial
Azoreductases acquired during adaptation to growth on Azo dyes. Arch. Microbiol., 138: 37-
43. Zissi U., Lyberatos G. and Pavlou S. (1997). Biodegradation of p-aminoAzobenzene by Bacillus subtilis under aerobic conditions. Journal of Industrial Microbiology and Biotechnology
19(1): 49-55.
Zollinger H. (1987). Color chemistry - Syntheses, properties and applications of organic dyes pigments. VCH, New York, NY.
Related Documents











![Biotransformation and Detoxification of Xylidine Orange ... · The three most commonly used dyes are azo, anthraquinone and phthalocyanine [1]. Azo dyes, in particular, contain an](https://static.cupdf.com/doc/110x72/601cef69c50cac6a4b44f587/biotransformation-and-detoxification-of-xylidine-orange-the-three-most-commonly.jpg)