ORIGINAL ARTICLE Bioclimatology of beech (Fagus sylvatica L.) in the Eastern Alps: spatial and altitudinal climatic signals identified through a tree-ring network Alfredo Di Filippo 1 *, Franco Biondi 2 , Katarina Cufar 3 , Martı´n de Luis 4 , Michael Grabner 5 , Maurizio Maugeri 6 , Emanuele Presutti Saba 1 , Bartolomeo Schirone 1 and Gianluca Piovesan 1 1 DAF, Universita ` degli studi della Tuscia, Viterbo, Italy, 2 DendroLab, Department of Geography, University of Nevada, Reno, NV, USA, 3 Department of Wood Science and Technology, Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia, 4 Departamento de Geografı ´a y Ordenacio ´n del Territorio, Universidad de Zaragoza, Zaragoza, Spain, 5 University of Natural Resources and Applied Life Sciences, Vienna, Austria, 6 Istituto di Fisica Generale Applicata, Milan, Italy * Correspondence: Alfredo Di Filippo, DAF, Facolta ` di Agraria, Universita ` degli Studi della Tuscia, Via SC de’ Lellis snc, 01100, Viterbo, Italy. E-mail: difi[email protected]. SUMMARY Aim To 1 1 identify the dominant spatial patterns of Fagus sylvatica radial growth in the Eastern Alps, and to understand their relationships to climate variation and bioclimatic gradients. Location Fourteen beech stands in the Eastern Alps 1 2 , growing between 200 and 1500 m a.s.l. in Italy, Slovenia and Austria. Methods At each site, trees were sampled using increment borers or by taking discs from felled trees. Cores and discs were processed by measuring and crossdating ring width. Ring width series were standardized, averaged, and prewhitened to obtain site chronologies. Hierarchical Cluster Analysis (HCA) and Principal Components Analysis of prewhitened site chronologies were used to identify spatial and altitudinal growth patterns, related to the bioclimatic position of each stand. Bootstrap correlation and response functions were computed between monthly climatic variables and either principal component scores or composite chronologies from stands associated by HCA. The stability of dendroclimatic signals was analyzed by moving correlation functions (MCF). Correlation analysis (teleconnections) based on a data base of 37 Italian and Slovenian beech tree-ring chronologies revealed the spatial extent of Principal Component scores. Results Sampled trees were 200–400 years old, representing the oldest beech trees that have been crossdated for the Alps to date. Maximum age was directly related to altitude and to the presence of historical forms of conservation. Tree- ring parameters varied according to geographic patterns and the age of sampled trees. Stands were bioclimatically organized according to their location, and with reference to their elevation and distance from the Adriatic Sea. A direct response to winter temperature was found at all elevations. The altitudinal gradient ranged from low elevation stands, characterized by a Mediterranean-type, late spring– summer drought signal, to mountain and high elevation stands, characterized by a direct response to growing season temperature plus an inverse response to the previous year’s July temperature. The mountain and high elevation signal was evident in Austria, the Central Alps and Slovenia, while the low elevation signal was confined to mountains adjacent to the Adriatic Sea. MCF revealed trends in the response to climatic factors affecting tree-ring formation in mountain and high mountain stands linked to climatic warming. Main conclusions Dendroclimatic networks can be used for bioclimatic studies of tree populations. A biogeographical separation emerged between the Alps and the Apennines at the upper elevations, while different degrees of mediterraneity distinguished sites at lower elevations. This information will be useful in assessing Journal of Biogeography (J. Biogeogr.) (2007) ª 2007 The Authors www.blackwellpublishing.com/jbi 1 Journal compilation ª 2007 Blackwell Publishing Ltd doi:10.1111/j.1365-2699.2007.01747.x The definitive version is available at www.blackwell-synergy.com

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINALARTICLE
Bioclimatology of beech (Fagus sylvatica
L.) in the Eastern Alps: spatial and
altitudinal climatic signals identified
through a tree-ring network
Alfredo Di Filippo1*, Franco Biondi2, Katarina Cufar3, Martın de Luis4,
Michael Grabner5, Maurizio Maugeri6, Emanuele Presutti Saba1, Bartolomeo
Schirone1 and Gianluca Piovesan1
1DAF, Universita degli studi della Tuscia,
Viterbo, Italy,2DendroLab, Department of
Geography, University of Nevada, Reno, NV,
USA,3Department of Wood Science and
Technology, Biotechnical Faculty, University of
Ljubljana, Ljubljana, Slovenia,4Departamento
de Geografıa y Ordenacion del Territorio,
Universidad de Zaragoza, Zaragoza,
Spain,5University of Natural Resources and
Applied Life Sciences, Vienna, Austria,6Istituto
di Fisica Generale Applicata, Milan, Italy
* Correspondence: Alfredo Di Filippo, DAF,
Facolta di Agraria, Universita degli Studi della
Tuscia, Via SC de’ Lellis snc, 01100, Viterbo,
Italy.
E-mail: [email protected].
SUMMARY
Aim To11 identify the dominant spatial patterns of Fagus sylvatica radial growth in
the Eastern Alps, and to understand their relationships to climate variation and
bioclimatic gradients.
Location Fourteen beech stands in the Eastern Alps12 , growing between 200 and
1500 m a.s.l. in Italy, Slovenia and Austria.
Methods At each site, trees were sampled using increment borers or by taking
discs from felled trees. Cores and discs were processed by measuring and
crossdating ring width. Ring width series were standardized, averaged,
and prewhitened to obtain site chronologies. Hierarchical Cluster Analysis
(HCA) and Principal Components Analysis of prewhitened site chronologies were
used to identify spatial and altitudinal growth patterns, related to the bioclimatic
position of each stand. Bootstrap correlation and response functions were
computed between monthly climatic variables and either principal component
scores or composite chronologies from stands associated by HCA. The stability of
dendroclimatic signals was analyzed by moving correlation functions (MCF).
Correlation analysis (teleconnections) based on a data base of 37 Italian and
Slovenian beech tree-ring chronologies revealed the spatial extent of Principal
Component scores.
Results Sampled trees were 200–400 years old, representing the oldest beech
trees that have been crossdated for the Alps to date. Maximum age was directly
related to altitude and to the presence of historical forms of conservation. Tree-
ring parameters varied according to geographic patterns and the age of sampled
trees. Stands were bioclimatically organized according to their location, and with
reference to their elevation and distance from the Adriatic Sea. A direct response
to winter temperature was found at all elevations. The altitudinal gradient ranged
from low elevation stands, characterized by a Mediterranean-type, late spring–
summer drought signal, to mountain and high elevation stands, characterized by
a direct response to growing season temperature plus an inverse response to the
previous year’s July temperature. The mountain and high elevation signal was
evident in Austria, the Central Alps and Slovenia, while the low elevation signal
was confined to mountains adjacent to the Adriatic Sea. MCF revealed trends in
the response to climatic factors affecting tree-ring formation in mountain and
high mountain stands linked to climatic warming.
Main conclusions Dendroclimatic networks can be used for bioclimatic studies
of tree populations. A biogeographical separation emerged between the Alps and
the Apennines at the upper elevations, while different degrees of mediterraneity
distinguished sites at lower elevations. This information will be useful in assessing
Journal of Biogeography (J. Biogeogr.) (2007)
ª 2007 The Authors www.blackwellpublishing.com/jbi 1Journal compilation ª 2007 Blackwell Publishing Ltd doi:10.1111/j.1365-2699.2007.01747.x
The definitive version is available at www.blackwell-synergy.com

INTRODUCTION
The biogeographical study of plant–climate relationships has
been an important field of research since the 19th century, and
aims to explain vegetation patterns based on climate variation
(Woodward, 1987). In particular, the effect of climate on
species physiology and geographic range has provided the
causal mechanism linking climate with vegetation and biome
distribution (Walter, 1985). With respect to forest ecosystems,
dendroecology can contribute to bioclimatic studies by
improving the analysis of tree growth response to environ-
mental gradients, thereby refining the classifications that are
based on climate–vegetation interactions. This approach
considers tree-ring parameters as bioindicators that integrate
the environmental factors controlling forest growth. Tree-ring
records can show growth–climate relationships over space
(from a forest stand to a whole hemisphere) and time (from
seasons to centuries) (Fritts, 1976). The effect of climate
forcing on tree growth has been studied at local (Douglass,
1920), regional (Meko et al., 1993) and hemispheric (Briffa
et al., 2002) spatial scales. In this context, sampling stands of
the same species (e.g. beech) at different locations and altitudes
can provide an objective bioclimatic classification of tree
populations (Piovesan et al., 2005a).
A number of bioclimatic classification systems are currently
available (e.g. Walter, 1985; Bailey, 1996), with each system
defining bioclimatic units (e.g. ecoregions and zonobiomes)
based on different methods and types of data (e.g. Thompson
et al., 2005). Recently, the use of statistical techniques, digital
data bases and advanced spatial analytical approaches have
allowed the numerical classification of environmental variables
to clarify their role in affecting plant distribution (Laurent
et al., 2004; Metzger et al., 2005). However, especially at
regional scales, there is a need to integrate patterns of species
distribution with ecological processes, paying special attention
to the effects of climate variability (Whittaker et al., 2005).
Horizontal and vertical gradients in tree–climate relationships
provide the basis for defining bioclimatic units in terms of the
leading dendroclimatic signals. Such studies generate the basic
information necessary to perform climatic reconstructions
from tree rings (Frank & Esper, 2005), and can be the starting
point to define simulation models of plant community
response to a changing climate (Cook et al., 2001). In addition,
this way of classifying woody vegetation into bioclimatic units
offers a valuable tool for a science based management of forest
ecosystem, linking climate fluctuations to forest productivity
(Biondi, 1999). Finally, if long instrumental records33 are
available, it is possible to explore the temporal stability of
the observed climate–growth relationships (Biondi, 2000), and
formulate hypotheses about the potential effects of climatic
change on plant functioning and community dynamics (Jump
& Penuelas, 2005; Whittaker et al., 2005).
Dendroecologists have used European beech (Fagus sylvat-
ica L.) extensively during the last two decades (Eckstein &
Frisse, 1982; Gutierrez, 1988; Rozas, 2001; Dittmar et al.,
2003; Lebourgeois et al., 2005), taking advantage of the
widespread distribution, sensitivity to climate and longevity
of beech (Bourquin-Mignot & Girardclos, 2001; Piovesan
et al., 2005b). In Italy, investigations were carried out at local
(e.g. Biondi, 1993; Bernabei et al., 1996; Piutti & Cescatti,
1997; Piovesan et al., 2003) and regional (Biondi, 1992;
Biondi & Visani, 1996; Piovesan et al., 2005a) scales.
European beech approaches the southern edge of its
geographic range in Italy, where its altitudinal range extends
more than 1500 m, from about 300–400 to 2000–2100 m
a.s.l. in central and southern Italy, and from 200–300 to
1500–1600 m in the Alps. Our main hypothesis is that,
because beech is present in both the Alps and the Apennines,
its tree-ring records can be used to detect ecological
differences between these two mountain ranges. If a large
enough number of chronologies are available, it should also
be possible to distinguish dendroclimatic responses along
ecological gradients, such as elevation. To date, dendrocli-
matic networks in the Alps have focused on conifer species
growing at high elevation or at the tree line (e.g. Kienast
et al., 1987; Urbinati et al., 1997; Rolland, 2002; Frank &
Esper, 2005). In this study we tested our hypotheses using a
transnational network of 14 European beech stands sampled
at different elevations in the Eastern Alps (Italy, Slovenia and
Austria), which is a focus region for bioclimatic studies (e.g.
Walter, 1985; Ellenberg, 1988). Our main objectives were (1)
to identify the dominant spatial patterns of radial growth in
Fagus sylvatica, (2) to investigate how such patterns relate to
climatic variability and geographic location, and (3) to
improve the definition of bioclimatic gradients, and refine the
classification systems that rely on them.
any future climate-related bioclimatic shifts, especially for forests at ecotones and
along altitudinal gradients.
Keywords
Alps, altitudinal gradient, bioclimatology, dendroclimatology, ecological gradi-
ent, ecotone, Fagus, old-growth forests, tree growth, tree-ring analysis.
A. Di Filippo et al.
2 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

METHODS
Study areas
A total of 14 European beech (Fagus sylvatica L.) forests were
sampled in the following ranges: Julian Alps in Italy and
Slovenia, Carnic Alps in Italy, and northern Alps in Austria
(Fig. 1). Sampled sites were located from 46.19� to 48.33� N
latitude, and from 12.75� to 15.43� E longitude, covering an
altitudinal range of 1300 m, from 200 to 1500 m a.s.l.
(Table 1). This region occupies the central-southern portion
of the geographic range of beech distribution, where this
species has a noticeable spread in altitude (Ellenberg, 1988).
According to the classification of Koppen–Trewartha (Tre-
wartha, 1968), the Carnic and the Austrian ranges pertain to
temperate climates (D), while the Julian mountains are at the
boundary between temperate and subtropical dry summer (Cs)
climates. A recent environmental classification of Europe
(Metzger et al., 2005) defined the central bulk44 of our network
as the Alpine Environmental Zone, the southernmost Julian
Alps as the boundary with the Mediterranean Mountains Zone,
and the northernmost Dunkelsteinerwald as within the Con-
tinental Zone.
Almost all stands were managed as high forests where beech
was the dominant species; only the lower elevation sites
(MOT, NIM, PEC, TOA and TOB; see Table 1 for acronyms)
(b)
(a)
Figure 1 (a) Geographic distribution of
Fagus sylvatica (von Wuehlisch., 2006), with
outline (rectangle) of the study area. (b)
Enlargement of the study area showing the
location of sampled sites.
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 3ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

included species (e.g. Acer spp., Fraxinus spp. And Ostrya
carpinifolia Scop.) that were managed as coppice-with-stand-
ards, and even at those sites beech maintained a monocormic
stem. These low-elevation beech stands, located at the
altitudinal limit of the species, cover a few hectares each,
and are found within mixed deciduous forests. Low elevation
forests on the Italian side of the Julian Alps and in Slovenia
(MOT, NIM, PEC, TOA and TOB) were privately owned. The
Slovenian sites were strongly affected by the battles of World
War I, and later on they were over-exploited by the local
population. Therefore well-preserved forests are only found in
remote areas that are difficult to access. All stands sampled in
the Carnic Alps were publicly owned, being situated above
towns for protection against avalanches and rock falls
(‘protection forests’). Some of these forests (TIM, CLE, LAT
and GRA), called ‘boschi banditi’ (Fig. 2), were maintained by
the Republic of Venice during the 16th century to produce
timber (especially masts) for ships, or set aside by local
inhabitants to provide shade and food (beech nuts and
understorey species) for livestock (Paiero et al., 1975). As a
final note, at several sites included in this network specifically
because of their relatively undisturbed conditions and old-
growth characteristics, land managers stipulated that only one
core per tree could be taken so as to minimize damage. The
Table 1 Geographical and structural features of the sampled beech sites.
Site
Alpine
range Country Code
Latitude
(N)
Longitude
(E)
Elevation*
(m a.s.l.) Aspect
Slope
(%)
d.b.h.�
(cm)
Ht.�
(m)
La Motta J I MOT 46.1886 13.3153 250 (200-300) SW 10–40 39–65 20
Tolmin A J S TOA 46.2000 13.7333 355 (290-420) SW 40–80 39–50 24
Nimis J I NIM 46.2006 13.2769 560 SE 45 34–65 27
Pechinie J I PEC 46.2807 13.2027 670 NE 10–30 36–62 20
Tolmin B J S TOB 46.2167 13.7500 821 (797-845) SW 40–80 39–53 27
Tolmin C J S TOL 46.2333 13.7666 1328 (1240-1415) S 40–80 33–55 22
Gracco C I GRA 46.5514 12.8519 825 (750-900) S 70–90 42–72 21
Cleulis C I CLE 46.5584 13.0006 930 NE 20–60 35–70 24
Tre Confini C I TRE 46.5039 13.5831 1100 – 20–60 35–55 30
Timau C I TIM 46.5817 13.0050 1160 (825-1500) S 70–90 40–95 25
Paularo C I PAU 46.5297 13.1172 1385 (1275-1500) W 70–90 52–100 27
Lateis C I LAT 46.4594 12.7489 1450 (1370-1530) S 75–110 33–70 28
Dunkelsteinerwald N A DSW 48.3333 15.4333 650 E 40–60 30–50 22
Hallstatt N A HSA 47.8333 13.7500 1400 SE 100–120 15–36 12
C, Carnic Alps; J, Julian Alps; N, Northern Austrian Alps. I, Italy; S, Slovenia; A, Austria.
*Mean with range in parentheses.
�Diameter at breast height range of sampled trees.
�Mean height of the three to four tallest trees in the stand.
Figure 2 View of the Gracco ‘protection
forest’ in Carnia, where beech age exceeds
300 years (Photograph by G. Piovesan).
COLOUR
A. Di Filippo et al.
4 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

same constraint was imposed in privately owned forests,
including most of the low-elevation beech stands.
Sampling and chronology building
In each stand, tree selection focused on dominant or
co-dominant trees, either isolated or grouped, with the best
combination of old age and trunk health. Trees were cored at
breast height (1.3 m from the ground) using an increment
borer, taking one or two cores per tree. At the Slovenian sites,
all samples were cross-sections taken from trees already felled
for other purposes. Tree-ring chronologies were developed
from wood samples using standard dendrochronological
procedures (Stokes & Smiley, 1996). After surfacing and
preliminary visual crossdating, tree-ring widths were measured
to the nearest 0.01 mm using the system CCTRMD (Aniol,
1987) and the program CATRAS (Aniol, 1983) or the LINTAB
measuring table and TSAP/X programme (Rinn, 1996). Tree-
ring series were visually and statistically compared with each
other to ensure accuracy of crossdating and measurement
(Holmes, 1983; Grissino-Mayer, 2001). Locally absent rings
(LAR), when detected, were given ring width equal to zero.
The percentage of locally absent rings was calculated on the
entire length of the tree-ring series. Dendrochronological
parameters (mean ring width, standard deviation, mean
sensitivity and first-order autocorrelation) were computed
for all measured ring-width series. The same parameters were
then computed on raw site chronologies, obtained by arith-
metically averaging the ring-width series by site. The years
1942–2001 were a common period that included at least three
ring-width series at each site. Site location and tree age were
then used to explain the variability of dendrochronological
parameters, computed for the 1942–2001 period using the raw
site chronologies.
Standardized tree-ring chronologies were produced for each
site using the following formula:
�It ¼
Pi¼nti¼1 ðw0:5 � yÞit
ntþ cit
with �It = chronology value at year t; nt = number of samples
for year t, with nt ‡ 3; w = crossdated ring width of sample i
in year t; y = value of sample i in year t computed by fitting
a modified negative exponential with asymptote ‡ 0 or a
straight line with slope £ 0 to the ith ring-width series;
cit = constant added to sample i in year t so that the
standardized chronology has mean equal to 1. Although
‘standardized indices’ used in dendrochronology are often
calculated as ratios between the measurement and the fitted
curve value, there is evidence that variance-stabilized
residuals should be preferred (Cook & Peters, 1997; Biondi,
1999; Helama et al., 2004). Each standardized chronology
was prewhitened by fitting autoregressive (AR) models
(Biondi & Swetnam, 1987) to remove any biological trend
and enhance the climatic signal (Cook et al., 2001). The
linear correlation between prewhitened site chronologies was
computed for the common period 1942–2001. Finally,
composite chronologies were obtained by pooling together
all ring-width series from sites whose prewhitened chronol-
ogies were associated by multivariate analysis (see next
paragraph). The same procedure used to compute prewhit-
ened site chronologies was used to compute prewhitened
composite chronologies. Chronology confidence was evalu-
ated by computing the expressed population signal (EPS)
(Wigley et al., 1984). EPS values were computed for the
period 1942–2001, 1952–2001 and 1962–2001 on the pre-
whitened site chronologies. These three periods were chosen
to show how the increase in the number of available samples
affects the EPS statistic. The limited extension of the low
elevation sites, together with the restrictions imposed on the
number of samples per tree, were responsible for a reduced
sample depth in the early part of these site chronologies. For
prewhitened composite chronologies (see Table S4 in Sup-
plementary Material), EPS values were calculated over the
entire length of the chronology using 50-year moving
windows with a 40-year overlap.
Multivariate analysis
The period 1942–2001, common to the 14 prewhitened beech
chronologies, was considered for multivariate analysis. Hier-
archical Cluster Analysis (HCA) and Principal Components
Analysis (PCA) were based on the correlation matrix between
chronologies. HCA was used for the first detection of
groupings among the 14 chronologies (Ludwig & Reynolds,
1988). Distance between variables was based on (1 – r), with
r = Pearson product–moment correlation coefficient, while
clusters were identified by means of the average distance
between all pairs of variables contained in two groups
(Stenson & Wilkinson, 2004). Because clusters were generated
according to the degree of growth affinity between chronol-
ogies, the HCA dendrogram, when interpreted with consid-
eration to the spatial and altitudinal location of each forest,
could reveal the bioclimatic organization of the network
(Piovesan et al., 2005a).
The main modes of common growth variability among
stands were represented by Principal Component (PC) scores,
or amplitudes (Piovesan et al., 2005a). Component loadings
(eigenvectors), which display the pattern of association of
chronologies with each component, were employed to detect
groupings in the tree-ring network. Selection of PCs was
guided by Kaiser’s Rule (Kaiser, 1992). The combination of
HCA and PCA is essential because HCA produces a clear-cut
bioclimatic classification of the network, and PCA allows the
break down (thus the description) of the dominant climatic
signals responsible for the observed classification. The spatial
extent of the common signals was investigated by telecon-
nection analysis (Fritts, 1976), performed by correlating PC
scores for the period 1942–88 with beech chronologies
developed for Austria, Italy and Slovenia, standardized and
prewhitened as in our site chronologies55 . Graphical represen-
tations of correlation maps were produced with the software
GMT (Wessel & Smith, 1998).
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 5ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd
The climate–growth relationship
Dendroclimatic correlation and response functions (Biondi &
Waikul, 2004) were calculated between monthly climate
variables and two types of tree-ring bioindicators. One type
was the first and second principal component scores of
prewhitened site chronologies; the other type was the
prewhitened composite chronologies. The bootstrap method
(Efron & Tibshirani, 1986; Guiot, 1991) was used for
significance testing. Explanatory climate variables spanned a
17-month window, from October of the current growth year
to June of the previous year. Climatic data for the Italian and
Slovenian sites were developed under the research project
CLIMAGRI (Brunetti et al., 2006), and were organized in a
grid of 1� · 1� cells, with each cell containing the monthly
values of minimum and maximum temperature anomalies
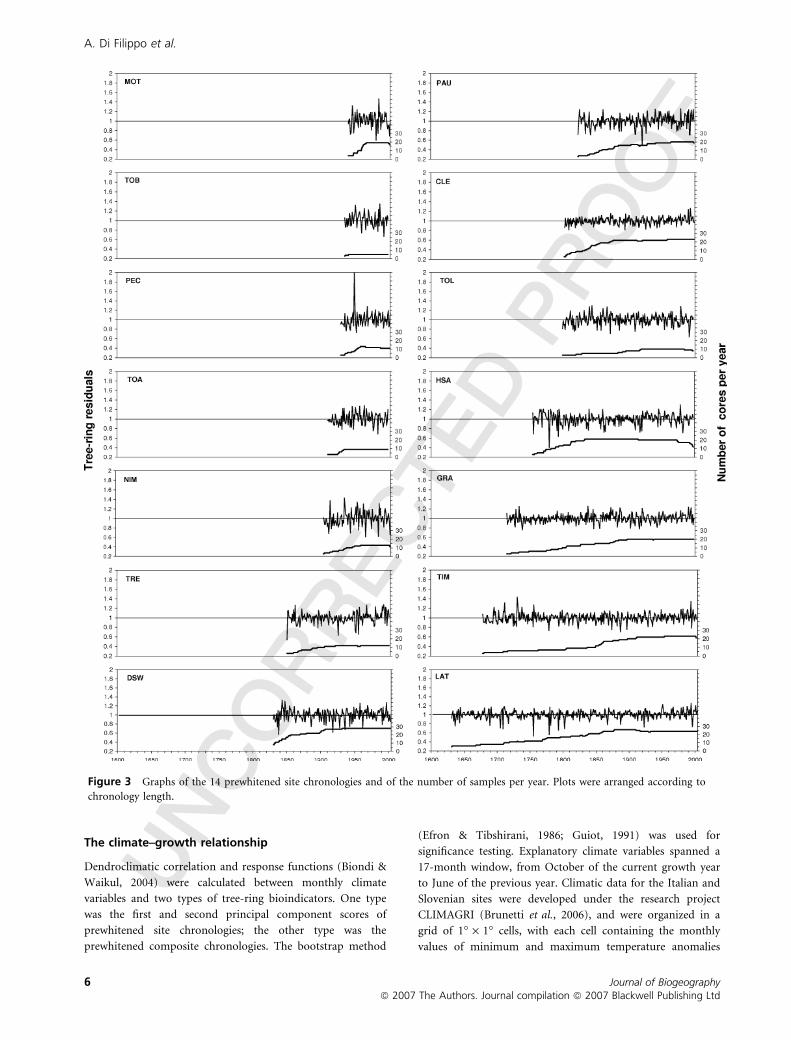
Figure 3 Graphs of the 14 prewhitened site chronologies and of the number of samples per year. Plots were arranged according to
chronology length.
A. Di Filippo et al.
6 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

and of precipitation ratios. Climate data for Italy and
Slovenia were obtained by averaging the two grid cells
(46� N, 13� E and 46� N, 14� E) that included our network
sites. In Austria, monthly total precipitation and mean air
temperature were available for the period 1942–2001 from
the meteorological station of Kremsmunster (48.052� N,
14.127� E, 403 m a.s.l.) for the Hallstatt site, and from the
station of Hohe Warte (48.299� N, 16.356� E, 203 m a.s.l.)
for the Dunkelsteinerwald site.
As climatic series for Italy and Slovenia reached the
beginning of the 19th century, we calculated moving correla-
tion functions (MCF) (Biondi, 1997) with the longest and best
replicated composite chronologies, i.e. those for mountain and
high mountain sites (see Table S4 in Supplementary Material),
using a 70-year window. By doing so, we investigated the
temporal stability of climatic signals identified for those
bioclimatic units.
RESULTS
Characteristics of the tree-ring network
A total of 248 cores from 188 trees were used in dendro-
chronological analyses (Tables 1 and 2a). Several old trees,
often exceeding 200–300 years of age, were identified (Fig. 3).
Mean ring width (MW) was negatively correlated with
elevation, with annual increments ranging from >4 mm in
the lowlands to c. 1 mm at the highest elevations (Table 2a).
The maximum age (Nmax) at each stand had a positive
correlation with altitude (r = 0.75, P < 0.001; this linear
relationship is graphically represented in Fig. 4). The main
features of raw and prewhitened site chronologies (plotted in
Fig. 3) are summarized in Table 2b.
Latitude was also negatively correlated with MW (Table 3).
Latitude may affect MW because the southern stands, which
are closer to the Adriatic Sea, experience66 a milder climate than
the northern, more continental sites. Finally, MW was
negatively correlated with tree age, as old-growth beech forests
are characterized by trees showing long periods of reduced
growth, occurring when they occupy the suppressed layers of
the canopy (Piovesan et al., 2005b). MW values were also
negatively correlated with LAR percentage (r = )0.50,
P < 0.05), possibly because LARs were frequent in old-growth
stands, especially during the suppressed growth periods. As
most LARs were present in the younger portion of measured
cores during periods of slow growth, it is likely that their
positive correlation with altitude can be related to the presence
of older trees at higher elevations.
The standard deviation (SD) had mean values varying
between 0.29 and 1.75 (Table 2a), and was negatively corre-
lated with altitude as well as to age of a site (Length) and of
individual trees (Nmax) (Table 3). This may be due to the
positive correlation between MW and SD (r = 0.82,
P < 0.001). The mean sensitivity (MS), a measure of year-to-
year variability, was comprised between 0.19 and 0.36, with
minimum values at CLE and at low elevation sites on the
Julian Alps (Table 2a). MS was correlated positively with
latitude (Table 3), most likely because both Austrian chronol-
ogies had MS values above 0.3 (Table 2a). The first-order
autocorrelation coefficient (A1), a measure of the persistence in
time series, varied between 0.61 and 0.79 (Table 2a), and
decreased with latitude, longitude and altitude (Table 3), but
correlations with longitude and altitude were immediately
below common significance thresholds. It is interesting to note
that A1 was inversely correlated with MS (r = )0.62, P < 0.05).
The order (p) of the autoregressive model used for prewhit-
Table 2(a) Summary of the ring width series for each site.
Site Code Trees Cores
MW*
(mm year)1)
SD*
(mm year)1) MS* A1*
Nmax
(years) Period
LAR
(%)
La Motta MOT 19 19 4.05 (1.70–5.44) 1.75 (1.10–2.49) 0.26 (0.18–0.44) 0.70 (0.44–0.94) 77 1928–2004 0
Tolmin A TOA 9 9 2.60 (1.71–3.50) 1.00 (0.59–1.41) 0.22 (0.15–0.29) 0.76 (0.57–0.86) 122 1880–2001 0
Nimis NIM 13 13 3.09 (1.46–4.34) 1.44 (0.75–1.89) 0.26 (0.20–0.34) 0.76 (0.53–0.88) 120 1883–2003 0
Pechinie PEC 12 13 3.11 (2.11–5.77) 1.40 (0.62–2.47) 0.22 (0.13–0.40) 0.72 (0.39–0.91) 79 1926–2004 0
Tolmin B TOB 5 5 3.28 (2.34–4.78) 1.18 (0.88–1.50) 0.20 (0.14–0.24) 0.77 (0.66–0.84) 83 1919–2001 0
Tolmin C TOL 10 10 1.61 (0.74–2.63) 0.69 (0.45–1.00) 0.25 (0.20–0.33) 0.74 (0.56–0.87) 271 1731–2001 0
Gracco GRA 20 20 1.43 (0.74–2.59) 0.70 (0.41–1.29) 0.24 (0.19–0.29) 0.78 (0.52–0.87) 318 1685–2002 0.099
Cleulis CLE 23 23 1.37 (0.40–3.19) 0.50 (0.19–0.90) 0.19 (0.13–0.26) 0.79 (0.60–0.91) 260 1744–2003 0
Tre Confini TRE 12 12 1.64 (0.85–2.81) 0.71 (0.41–1.00) 0.24 (0.16–0.30) 0.74 (0.58–0.86) 172 1831–2002 0.195
Timau TIM 21 27 1.28 (0.70–3.79) 0.72 (0.36–1.33) 0.26 (0.16–0.33) 0.77 (0.23–0.96) 348 1655–2002 0.162
Paularo PAU 20 20 2.11 (0.88–4.08) 0.80 (0.47–1.44) 0.27 (0.16–0.35) 0.61 (0.14–0.85) 261 1742–2002 0.110
Lateis LAT 24 28 1.05 (0.50–2.08) 0.49 (0.23–1.00) 0.25 (0.16–0.36) 0.77 (0.48–0.89) 380 1625–2004 0.187
Dunkelsteinerwald DSW 14 28 1.27 (0.79–2.06) 0.59 (0.36–0.86) 0.32 (0.24–0.37) 0.65 (0.25–0.86) 177 1827–2003 0
Hallstatt HSA 11 21 0.54 (0.25–0.73) 0.29 (0.15–0.42) 0.36 (0.29–0.43) 0.67 (0.40–0.85) 252 1751–2002 –
MW, mean ring-width; SD, standard deviation; MS, mean sensitivity; A1, first-order autocorrelation; Nmax, maximum number of rings counted on a
single core; Period, period covered by at least one sample; LAR, percentage of locally absent rings.
*Mean values with range in parentheses.
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 7ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

ening varied between 1 and 5 (Table 2b), and was positively
correlated with chronology length (Table 3).
Mountain (three sites) and high mountain (five sites)
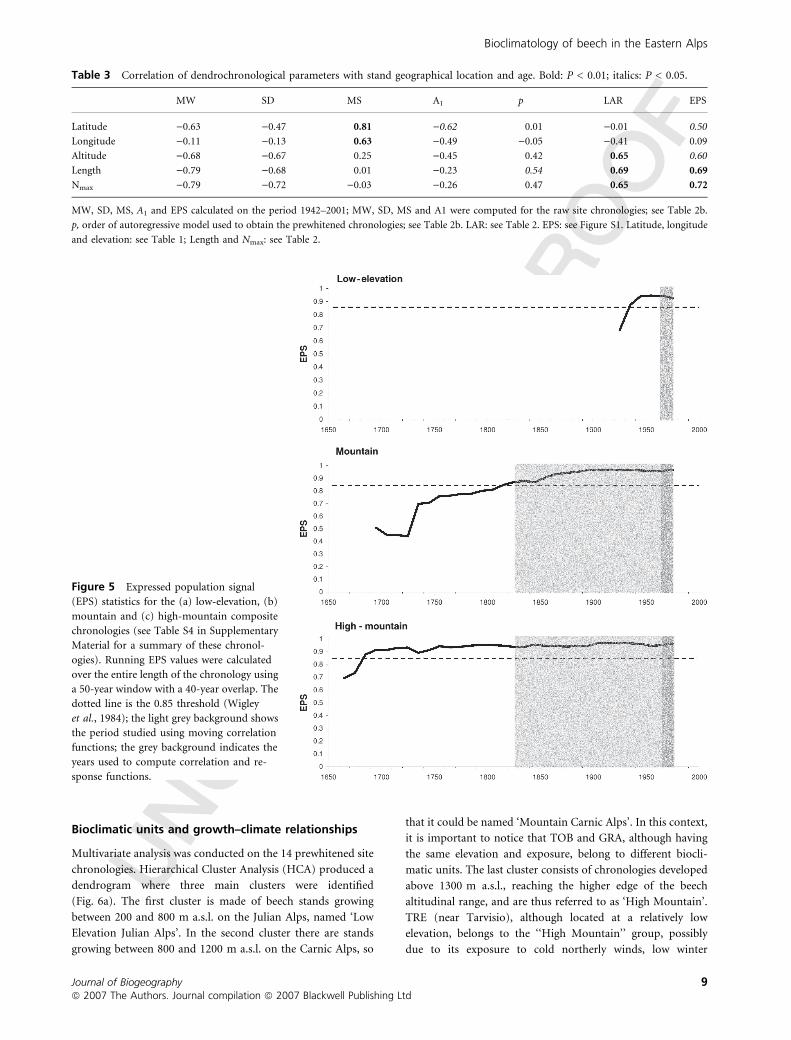
chronologies had EPS values >0.85. Of the six low elevation
sites, two had EPS > 0.85, and the other four chronologies
approached the EPS value in the most recent decades (see
Fig. S1). Given the overall high correlation between all tree-ring
chronologies (see Table S1), and the importance of our tree-ring
network for its ‘rear edge’ location (Hampe & Petit, 2005)
between Eurosiberian and Mediterranean regions, no chronol-
ogy was discarded based on its EPS value. Latitude and altitude
were directly correlated with EPS (Table 3), suggesting a
stronger climatic control on tree growth moving upward and
northward along the network. The highest correlationwas found
between EPS and stand age (Table 3), suggesting that old-
growth forests are characterized by greater synchronicity of
radial increment, perhaps because they have also escaped human
impacts for decades. For composite chronologies (see Table S4),
EPS values exceeded the 0.85 threshold during periods used to
quantify climate–tree growth relationships (Fig. 5).
Table 2(b) Summary of the raw and of the prewhitened site chronologies.
Site Code Trees Cores Period Length
Raw chronology Prewhitened
chronology
Entire length Common period (1942–
2001)
Entire length
MW
(mm
year)1)
SD
(mm
year)1) MS A1
MW
(mm
year)1)
SD
(mm
year)1) MS A1 p MW SD MS A1
La Motta MOT 19 19 1940–2004 65 2.99 1.66 0.21 0.93 3.50 1.47 0.16 0.88 2 1.00 0.16 0.19 )0.08
Tolmin A TOA 9 9 1910–2001 92 2.00 1.01 0.17 0.92 2.84 0.60 0.14 0.65 2 1.00 0.13 0.15 )0.03
Nimis NIM 13 13 1903–2003 101 2.64 1.08 0.18 0.86 3.47 0.69 0.13 0.62 2 1.00 0.16 0.18 )0.02
Pechinie PEC 12 13 1930–2004 75 2.64 1.34 0.16 0.92 3.08 1.20 0.14 0.83 1 1.00 0.17 0.16 )0.04
Tolmin B TOB 5 5 1934–2001 68 2.93 1.06 0.19 0.85 3.45 0.66 0.14 0.58 3 1.00 0.14 0.16 )0.03
Tolmin C TOL 10 10 1797–2001 205 1.25 0.57 0.22 0.85 1.53 0.36 0.16 0.64 4 1.00 0.11 0.13 0.00
Gracco GRA 20 20 1714–2002 289 1.15 0.39 0.16 0.83 1.25 0.29 0.13 0.70 2 1.00 0.09 0.10 )0.01
Cleulis CLE 23 23 1804–2003 200 1.08 0.41 0.16 0.90 1.41 0.28 0.12 0.66 1 1.00 0.07 0.09 )0.07
Tre Confini TRE 12 12 1850–2002 153 1.47 0.42 0.17 0.70 1.72 0.42 0.16 0.60 1 1.00 0.12 0.12 0.00
Timau TIM 21 27 1676–2002 327 1.16 0.65 0.19 0.83 1.58 0.46 0.13 0.75 3 1.00 0.10 0.11 )0.01
Paularo PAU 20 20 1824–2002 179 1.57 0.78 0.23 0.83 2.08 0.41 0.19 0.32 1 1.00 0.13 0.14 0.03
Lateis LAT 24 28 1626–2004 379 0.85 0.36 0.18 0.87 1.42 0.28 0.14 0.51 5 1.00 0.09 0.10 )0.01
Dunkelsteinerwald DSW 14 28 1828–2003 176 1.23 0.34 0.21 0.58 1.23 0.28 0.22 0.29 2 1.00 0.13 0.15 )0.02
Hallstatt HSA 11 21 1753–2002 250 0.57 0.22 0.28 0.58 0.57 0.17 0.25 0.49 3 1.00 0.12 0.13 0.00
Period, period covered by three or more samples; Length, number of years included in Period; MW, mean ring width; SD, standard deviation; MS,
mean sensitivity; A1, first-order autocorrelation; p, autoregressive model order.
Figure 4 Relationship between maximum age at each sampled forest and average elevation. ‘B’ indicates ‘boschi banditi’, forest stands
historically protected from logging.
A. Di Filippo et al.
8 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd
Bioclimatic units and growth–climate relationships
Multivariate analysis was conducted on the 14 prewhitened site
chronologies. Hierarchical Cluster Analysis (HCA) produced a
dendrogram where three main clusters were identified
(Fig. 6a). The first cluster is made of beech stands growing
between 200 and 800 m a.s.l. on the Julian Alps, named ‘Low
Elevation Julian Alps’. In the second cluster there are stands
growing between 800 and 1200 m a.s.l. on the Carnic Alps, so
that it could be named ‘Mountain Carnic Alps’. In this context,
it is important to notice that TOB and GRA, although having
the same elevation and exposure, belong to different biocli-
matic units. The last cluster consists of chronologies developed
above 1300 m a.s.l., reaching the higher edge of the beech
altitudinal range, and are thus referred to as ‘High Mountain’.
TRE (near Tarvisio), although located at a relatively low
elevation, belongs to the ‘‘High Mountain’’ group, possibly
due to its exposure to cold northerly winds, low winter
Table 3 Correlation of dendrochronological parameters with stand geographical location and age. Bold: P < 0.01; italics: P < 0.05.
MW SD MS A1 p LAR EPS
Latitude )0.63 )0.47 0.81 )0.62 0.01 )0.01 0.50
Longitude )0.11 )0.13 0.63 )0.49 )0.05 )0.41 0.09
Altitude )0.68 )0.67 0.25 )0.45 0.42 0.65 0.60
Length )0.79 )0.68 0.01 )0.23 0.54 0.69 0.69
Nmax )0.79 )0.72 )0.03 )0.26 0.47 0.65 0.72
MW, SD, MS, A1 and EPS calculated on the period 1942–2001; MW, SD, MS and A1 were computed for the raw site chronologies; see Table 2b.
p, order of autoregressive model used to obtain the prewhitened chronologies; see Table 2b. LAR: see Table 2. EPS: see Figure S1. Latitude, longitude
and elevation: see Table 1; Length and Nmax: see Table 2.
Figure 5 Expressed population signal
(EPS) statistics for the (a) low-elevation, (b)
mountain and (c) high-mountain composite
chronologies (see Table S4 in Supplementary
Material for a summary of these chronol-
ogies). Running EPS values were calculated
over the entire length of the chronology using
a 50-year window with a 40-year overlap. The
dotted line is the 0.85 threshold (Wigley
et al., 1984); the light grey background shows
the period studied using moving correlation
functions; the grey background indicates the
years used to compute correlation and re-
sponse functions.
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 9ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

temperatures and generally cool summers (Mennella, 1967;
Trewartha, 1968). Mountain and high mountain stands could
be further combined into a single group. The northernmost
site, DSW (Fig. 1 and Table 1), located at 650 m a.s.l., had low
affinity with the rest of the network and remained isolated
from the previous clusters. This site is located at the beginning
of the Continental Environmental Zone (Metzger et al., 2005),
and this may explain its uniqueness. Based on these results,
three sites were selected from each elevation range, so that the
high-mountain composite chronology incorporated all sam-
ples from LAT, PAU, TOL; the mountain composite chronol-
ogy was developed using TIM, CLE and GRA; and the low-
elevation composite chronology consisted of MOT, NIM and
TOA.
The first two principal components of the 14 prewhitened
chronologies explained 50% of the total variance, and were
retained for describing the beech network (see Fig. S2).
Mountain and high mountain chronologies have the highest
loadings on the first principal component, which explains 33%
of the total variance. The second principal component, which
accounts for 17% of the total variance, is mostly related to the
low elevation chronologies (see Fig. S2). Considering the
uniqueness of the Austrian site DSW, and the fact that gridded
climatic data covered the location of the Italian and Slovenian
sites, climate–tree growth relationships were analyzed sepa-
rately for the Austrian sites (see Tables S2 and S3). The first
two principal components of the 12 Italian and Slovenian
chronologies explained 55% of the total variance, and were
retained for describing the beech network (Fig. 6b,c). The first
component (PC1, 35.4% of the total variance) was mainly
related to mountain and high mountain chronologies; low
elevation stands were more related to the second component
(PC2, 19.3% of the total variance). Groups similar to those
identified by HCA were recognized by plotting the first and
second principal component loadings as a function of eleva-
tion (Fig. 6b,c).
Correlation and response function analysis of principal
component scores (Table 4) showed that radial growth in both
mountain and high mountain stands (PC1) was related to May
precipitation (negatively), September temperature (positively),
January minimum temperature (positively) and previous July
temperature (negatively). Radial growth in low-elevation beech
stands (PC2) was positively related to precipitation and
negatively to temperature in the months of May, July and
August; wood formation therefore appears to be drought-
limited during the growing season. Previous September and
October minimum temperatures were also negatively correla-
ted with low-elevation beech stands, while previous July and
November maximum temperatures appeared positively corre-
lated.
Correlation and response functions calculated for the period
1942–2001 using composite chronologies (see Table S4)
showed that growth in the high mountain forests was
positively correlated with May temperature and negatively
correlated with precipitation (Table 5), most likely because the
growing season at this elevation begins at the end of May
(Dittmar & Elling, 2005). July and August temperatures were
positively correlated with the high elevation composite,
whereas previous July temperature was inversely correlated.
No climatic variable for the growing season was significantly
correlated with the growth of mountain Carnic Alp stands, so
that the most important relationship was that with previous
summer temperatures. Thus, mountain and high mountain
chronologies, which were similar in terms of principal
component loadings, could be separated based on an eleva-
tion-related climatic signal. The low elevation composite was
R2
34.0=
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
0.1
004100210001008006004002
).l.s.am(edutitlA
Lo
ad
ing
(b)
(c)
PC1
PC2
R2
28.0=
0.1-
8.0-
6.0-
4.0-
2.0-
0.0
2.0
4.0
6.0
8.0
0.1
004100210001008006004002
).l.s.am(edutitlA
Lo
ad
ing
(a)
Figure 6 (a) Dendrogram showing results from the hierarchical cluster analysis made on the 14 prewhitened site chronologies (see
Table 2b). In parenthesis are reported the elevation and the alpine range of each site (see Table 1). Triangle = high-mountain forest;
circle = mountain forest; square = low elevation forest. Loadings of Italian and Slovenian prewhitened site chronologies on (b) the first and
(c) the second principal component are plotted as a function of site elevation.
A. Di Filippo et al.
10 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

correlated positively with June–July precipitation and negat-
ively with June–July temperature, pointing to summer drought
as the main climate signal. A positive correlation with
precipitation and a negative one with temperature also
appeared for the previous September. A positive correlation
with January minimum temperatures, which was reported for
PC1 (Table 4), was also found for low elevation and mountain
composites (Table 5). Climate–growth relationships for the
Austrian sites, DSW and HSA (see Tables S2 and S3),
reaffirmed their somewhat separate bioclimatic classification.
This separation could be the cause for the low correlations
between monthly precipitation recorded at the two meteoro-
logical stations in Austria and averaged over the two grid cells
for Italy and Slovenia (see Fig. S3).
Moving correlation function analysis was carried out on
mountain and high-mountain composite chronologies to
investigate the temporal stability of their main climatic
signals (Fig. 7). For mountain stands, the correlation in
growth with July maximum temperature during the previous
year has a progressive negative trend during the last
150 years (Fig. 7a), while the correlation with January
minimum temperature appears to increase in recent years
(Fig. 7b). With respect to high mountain sites, the inverse
correlations with May (Fig. 7c) and August precipitation
(data not shown) have emerged only in recent times. The
correlation of growth with August temperature has fluctu-
ated through time (Fig. 7d), with a tendency to higher
values in the most recent years, when correlation with July
and September temperature also becomes significant (data
not shown).
Teleconnection analysis
Teleconnection analysis of PCA scores served to evaluate the
geographic extent of climatic signals (Fig. 8). Correlations
between PC1 scores and other beech chronologies spread
mainly westward up to the Central Alps. It was less strong but
still significant eastward up to the Dinaric Mountains and the
hills of south-eastern Slovenia, and southward up to the
Northern Apennines (Fig. 8a). There was no significant
correlation with any beech chronologies further south, with
the only exception of Monte Cimino in Latium (r = 0.50,
P < 0.001). This might be due to the singularity of this site,
characterized by fertile volcanic soils, frequent fog and direct
exposure to northerly cold winds. Of the northernmost sites
DSW and HSA, which were excluded from the PC-based
dendroclimatic analysis, the former showed a less strong
correlation with PC1 scores (r = 0.32, P < 0.05) than the latter
(r = 0.47, P < 0.001). PC2 scores correlated eastward to the
Dinaric Mountains and south-eastern Slovenia, and southward
to low elevation stands in Central Italy (Latium) (Fig. 8b). In
that region, teleconnections could extend upward to 1050 m
a.s.l., where beech can grow in association with holm oak
(Quercus ilex L.) (Bernabei et al., 1996). No other Italian
chronologies from stands growing at higher elevations in the
Alpine region, the peninsula, or Sicily (Castorina et al., 2005),
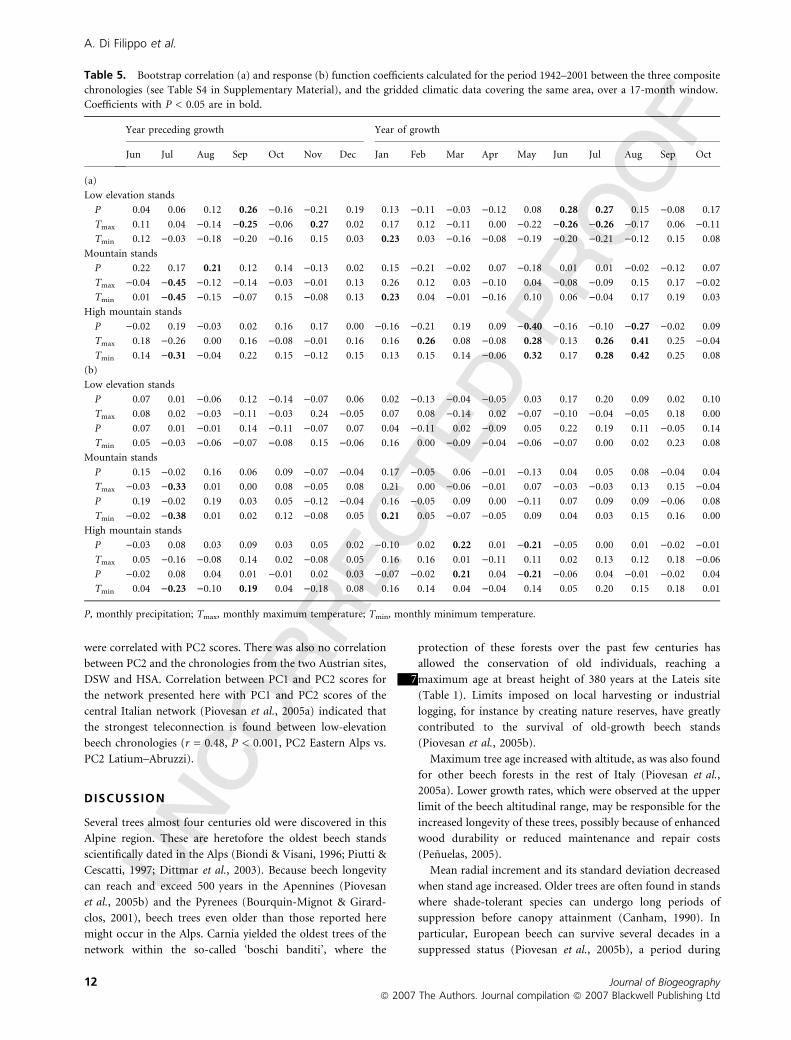
Table 4 Bootstrap (a) correlation and (b) response function coefficients calculated for the period 1942–2001 between the scores of the first
two PCs of the 12 prewhitened chronologies from Italy and Slovenia, and the gridded climatic data covering the same area, over a 17-month
window. Coefficients with P < 0.05 are in bold.
Year preceding growth Year of growth
Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct
(a)
PC1
P 0.14 0.18 0.12 0.14 0.06 0.00 0.06 0.00 )0.23 0.04 0.06 )0.27 )0.06 0.06 )0.11 )0.06 0.11
Tmax 0.10 )0.33 )0.11 )0.12 )0.08 0.06 0.12 0.22 0.20 0.03 )0.14 0.12 )0.02 0.00 0.21 0.27 )0.01
Tmin 0.09 )0.39 )0.16 )0.04 0.10 )0.03 0.11 0.23 0.12 0.03 )0.17 0.15 0.08 0.04 0.22 0.30 0.10
PC2
P 0.01 0.07 )0.02 )0.19 0.25 0.15 )0.19 )0.17 )0.06 0.11 0.09 )0.22 )0.16 )0.40 )0.25 0.04 )0.12
Tmax )0.07 )0.24 0.09 0.25 0.01 )0.23 0.11 )0.07 )0.03 0.14 )0.07 0.34 0.18 0.33 0.37 0.13 0.11
Tmin )0.06 )0.18 0.09 0.26 0.24 )0.17 0.07 )0.14 )0.02 0.23 )0.05 0.35 0.21 0.31 0.36 0.04 )0.05
(b)
PC1
P 0.12 0.00 0.05 0.12 )0.02 0.01 0.03 0.00 )0.05 0.09 )0.02 )0.16 0.01 0.09 0.04 0.02 0.01
Tmax 0.05 )0.20 )0.04 )0.03 0.04 0.04 0.04 0.19 0.09 )0.05 )0.10 0.07 )0.01 0.02 0.09 0.26 )0.01
P 0.14 0.01 0.11 0.08 )0.04 )0.03 0.03 0.03 )0.07 0.12 0.00 )0.15 0.03 0.13 0.04 )0.02 0.06
Tmin 0.03 )0.28 )0.06 0.03 0.08 )0.03 0.03 0.23 0.08 )0.04 )0.09 0.11 0.04 0.09 0.13 0.26 0.03
PC2
P )0.02 0.02 0.15 )0.08 0.16 0.03 )0.09 )0.02 0.10 0.12 )0.05 )0.12 )0.08 )0.28 )0.10 0.02 )0.06
Tmax )0.07 )0.14 )0.02 0.13 0.02 )0.25 0.10 0.01 )0.05 0.16 )0.09 0.17 0.06 0.06 0.16 )0.07 0.01
P )0.02 0.05 0.11 )0.12 0.12 0.01 )0.09 )0.04 0.06 0.07 0.02 )0.11 )0.09 )0.24 )0.11 0.08 )0.08
Tmin )0.05 )0.11 0.00 0.13 0.10 )0.19 0.09 )0.07 0.04 0.13 )0.04 0.18 0.07 0.06 0.10 )0.11 )0.05
P, monthly precipitation; Tmax, monthly maximum temperature; Tmin, monthly minimum temperature.
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 11ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd
were correlated with PC2 scores. There was also no correlation
between PC2 and the chronologies from the two Austrian sites,
DSW and HSA. Correlation between PC1 and PC2 scores for
the network presented here with PC1 and PC2 scores of the
central Italian network (Piovesan et al., 2005a) indicated that
the strongest teleconnection is found between low-elevation
beech chronologies (r = 0.48, P < 0.001, PC2 Eastern Alps vs.
PC2 Latium–Abruzzi).
DISCUSSION
Several trees almost four centuries old were discovered in this
Alpine region. These are heretofore the oldest beech stands
scientifically dated in the Alps (Biondi & Visani, 1996; Piutti &
Cescatti, 1997; Dittmar et al., 2003). Because beech longevity
can reach and exceed 500 years in the Apennines (Piovesan
et al., 2005b) and the Pyrenees (Bourquin-Mignot & Girard-
clos, 2001), beech trees even older than those reported here
might occur in the Alps. Carnia yielded the oldest trees of the
network within the so-called ‘boschi banditi’, where the
protection of these forests over the past few centuries has
allowed the conservation of old individuals, reaching a
maximum age at breast height of 380 years at77 the Lateis site
(Table 1). Limits imposed on local harvesting or industrial
logging, for instance by creating nature reserves, have greatly
contributed to the survival of old-growth beech stands
(Piovesan et al., 2005b).
Maximum tree age increased with altitude, as was also found
for other beech forests in the rest of Italy (Piovesan et al.,
2005a). Lower growth rates, which were observed at the upper
limit of the beech altitudinal range, may be responsible for the
increased longevity of these trees, possibly because of enhanced
wood durability or reduced maintenance and repair costs
(Penuelas, 2005).
Mean radial increment and its standard deviation decreased
when stand age increased. Older trees are often found in stands
where shade-tolerant species can undergo long periods of
suppression before canopy attainment (Canham, 1990). In
particular, European beech can survive several decades in a
suppressed status (Piovesan et al., 2005b), a period during
Table 5. Bootstrap correlation (a) and response (b) function coefficients calculated for the period 1942–2001 between the three composite
chronologies (see Table S4 in Supplementary Material), and the gridded climatic data covering the same area, over a 17-month window.
Coefficients with P < 0.05 are in bold.
Year preceding growth Year of growth
Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct
(a)
Low elevation stands
P 0.04 0.06 0.12 0.26 )0.16 )0.21 0.19 0.13 )0.11 )0.03 )0.12 0.08 0.28 0.27 0.15 )0.08 0.17
Tmax 0.11 0.04 )0.14 )0.25 )0.06 0.27 0.02 0.17 0.12 )0.11 0.00 )0.22 )0.26 )0.26 )0.17 0.06 )0.11
Tmin 0.12 )0.03 )0.18 )0.20 )0.16 0.15 0.03 0.23 0.03 )0.16 )0.08 )0.19 )0.20 )0.21 )0.12 0.15 0.08
Mountain stands
P 0.22 0.17 0.21 0.12 0.14 )0.13 0.02 0.15 )0.21 )0.02 0.07 )0.18 0.01 0.01 )0.02 )0.12 0.07
Tmax )0.04 )0.45 )0.12 )0.14 )0.03 )0.01 0.13 0.26 0.12 0.03 )0.10 0.04 )0.08 )0.09 0.15 0.17 )0.02
Tmin 0.01 )0.45 )0.15 )0.07 0.15 )0.08 0.13 0.23 0.04 )0.01 )0.16 0.10 0.06 )0.04 0.17 0.19 0.03
High mountain stands
P )0.02 0.19 )0.03 0.02 0.16 0.17 0.00 )0.16 )0.21 0.19 0.09 )0.40 )0.16 )0.10 )0.27 )0.02 0.09
Tmax 0.18 )0.26 0.00 0.16 )0.08 )0.01 0.16 0.16 0.26 0.08 )0.08 0.28 0.13 0.26 0.41 0.25 )0.04
Tmin 0.14 )0.31 )0.04 0.22 0.15 )0.12 0.15 0.13 0.15 0.14 )0.06 0.32 0.17 0.28 0.42 0.25 0.08
(b)
Low elevation stands
P 0.07 0.01 )0.06 0.12 )0.14 )0.07 0.06 0.02 )0.13 )0.04 )0.05 0.03 0.17 0.20 0.09 0.02 0.10
Tmax 0.08 0.02 )0.03 )0.11 )0.03 0.24 )0.05 0.07 0.08 )0.14 0.02 )0.07 )0.10 )0.04 )0.05 0.18 0.00
P 0.07 0.01 )0.01 0.14 )0.11 )0.07 0.07 0.04 )0.11 0.02 )0.09 0.05 0.22 0.19 0.11 )0.05 0.14
Tmin 0.05 )0.03 )0.06 )0.07 )0.08 0.15 )0.06 0.16 0.00 )0.09 )0.04 )0.06 )0.07 0.00 0.02 0.23 0.08
Mountain stands
P 0.15 )0.02 0.16 0.06 0.09 )0.07 )0.04 0.17 )0.05 0.06 )0.01 )0.13 0.04 0.05 0.08 )0.04 0.04
Tmax )0.03 )0.33 0.01 0.00 0.08 )0.05 0.08 0.21 0.00 )0.06 )0.01 0.07 )0.03 )0.03 0.13 0.15 )0.04
P 0.19 )0.02 0.19 0.03 0.05 )0.12 )0.04 0.16 )0.05 0.09 0.00 )0.11 0.07 0.09 0.09 )0.06 0.08
Tmin )0.02 )0.38 0.01 0.02 0.12 )0.08 0.05 0.21 0.05 )0.07 )0.05 0.09 0.04 0.03 0.15 0.16 0.00
High mountain stands
P )0.03 0.08 0.03 0.09 0.03 0.05 0.02 )0.10 0.02 0.22 0.01 )0.21 )0.05 0.00 0.01 )0.02 )0.01
Tmax 0.05 )0.16 )0.08 0.14 0.02 )0.08 0.05 0.16 0.16 0.01 )0.11 0.11 0.02 0.13 0.12 0.18 )0.06
P )0.02 0.08 0.04 0.01 )0.01 0.02 0.03 )0.07 )0.02 0.21 0.04 )0.21 )0.06 0.04 )0.01 )0.02 0.04
Tmin 0.04 )0.23 )0.10 0.19 0.04 )0.18 0.08 0.16 0.14 0.04 )0.04 0.14 0.05 0.20 0.15 0.18 0.01
P, monthly precipitation; Tmax, monthly maximum temperature; Tmin, monthly minimum temperature.
A. Di Filippo et al.
12 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

which growth is reduced to a minimum and locally absent
rings are more frequent in the lower stem of the tree (Lorimer
et al., 1999).
Geographic patterns characterized beech radial growth in
the Eastern Alps, as was previously reported for Apennine
forests (Piovesan et al., 2005a). Latitude was negatively
Figure 7 Moving correlation function coefficients calculated between selected climatic variables and composite mountain (a, b) and high-
mountain (c, d) beech chronologies. Period: 1803–2001; moving window: 70 years. See Table S4 in Supplementary Material for a summary
of the composite chronologies.
(a) (b)
Figure 8 Correlation map (teleconnections) between first (a) and second (b) principal component scores of 12 prewhitened chronologies
with Italian, Austrian and Slovenian beech site chronologies. Symbols relate to bioclimatic position as in Fig. 6, and were based on HCA of
different data sets (see Fig. 6a in this study, Fig. 2 in Piovesan et al., 2005a; and Fig. 3 in Di Filippo, 2006). Dimension of each symbol is
proportional to correlation value; in solid grey are correlations with P-value < 0.01. Calculations were made on the period 1942–1988.
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 13ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

correlated with mean ring-width, and was positively correlated
with mean sensitivity. Proximity to the Adriatic Sea most likely
generates a milder climate in the Julian Alps, and these
influences seemed to reach the south-facing Carnic valleys as
well. Relatively warmer temperatures, together with the
exceptional high levels of precipitation in this southern range
of the Eastern Alps (>2000–3000 mm year)1, Desiato et al.,
2005), create favourable conditions for beech growth, as shown
by the low mean sensitivity of these chronologies. The higher
mean sensitivity of the Austrian chronologies can be explained
by a progressive increase in continentality with latitude. In
particular, a shift from a Mediterranean to a Continental
Environmental Zone, passing through the Alpine one, can be
observed in our network going from south to north (Metzger
et al., 2005).
Altitude exerts an even stronger control on mean ring width,
with values increasing three to four times from high to low
elevation sites. This pattern is frequently observed along
altitudinal gradients (Monserud & Sterba, 1996; Piovesan
et al., 2005a). It can be explained considering that growing
season length and ecosystem productivity are closely linked
(e.g., White et al., 1999). Dittmar & Elling (2005), observed
that in Bavaria, beech growing season decreases by about 2–
3 days for every 100-m increase in elevation. Moreover, beech
growth may be negatively impacted by frequent late-frost
damage at higher elevations (Dittmar et al., 2006).
Multivariate analysis applied to tree-ring series has been
demonstrated to detect the effects of environmental gradients
on the growth of forest species (e.g. for Europe: Biondi &
Visani, 1996; Makinen et al., 2002; Dittmar et al., 2003;
Linderholm et al., 2003; Tardif et al., 2003; Frank & Esper,
2005; Piovesan et al., 2005a). Despite the elevational range of
this study, most beech forests in the network showed a
common climatic signal, characterized by the importance of
conditions at the beginning and the end of the growing period,
the risk of winter cold damage and the one-year lagged effect88 of
floral induction. Statistical relationships pointed to a negative
effect of May precipitation, which could be due to beech
sensitivity to soil saturation (Nielsen & Jørgensen, 2003),
especially considering the high precipitation levels at the study
areas (Desiato et al., 2005) and the fact that snow melt
typically occurs in late spring (Mennella, 1967). In addition,
cloudiness during May is another factor that could reduce
growth through light limitation (e.g. Graham et al., 2003).
During September, warmer temperatures can contribute to
latewood cell wall thickening (Lebourgeois et al., 2005). Before
the beginning of the growing season, minimum temperature in
January could be related to tree growth because extremely low
winter temperature can cause cold damage, such as freezing
embolism (Lemoine et al., 1999).
The highest correlations were found with previous July
temperature, which may be related to floral induction,
stimulated by a hot and dry summer in the year preceding
masting (Piovesan & Adams, 2001; Schmidt, 2006). This signal
was more pronounced in mountain and high mountain stands,
and less so at lower elevations. Previous summer conditions
were most important for mountain chronologies, while the
climate of the growing season gains significance at higher
elevations (Dittmar & Elling, 1999). This beech response to
previous summer conditions has been found throughout
Europe, e.g. in the Pyrenees (Gutierrez, 1988; Dittmar et al.,
2003), Cantabria (Rozas, 2001), Apennines (Piovesan &
Schirone, 2000), French hills (Lebourgeois et al., 2005) and
central Europe (Dittmar et al., 2003). Its widespread occur-
rence suggests a connection with climate controls on physio-
logical processes involved in resource accumulation (e.g. starch
reserves) and bud development (with particular reference to
leaf primordia) and differentiation of flower buds (see
Nakawatase & Peterson, 2006). Thus, the number of differen-
tiated flower buds should influence the amount of photosynt-
hates assigned to reproduction, rather than to growth, during
the following season. Beech mast years are known to have a
negative effect on wood production, and are sometime
responsible for the formation of ‘pointer years’ (see Figs
19.112 and 19.114 in Schweingruber, 1996; Piovesan &
Bernabei, 1997). Studies that have employed a long-term data
set have usually found trade-offs between radial growth,
masting and climate (Woodward et al., 1994; Piovesan &
Bernabei, 1997; Selas et al., 2002; Monks & Kelly, 2006).
Relationships between reproduction and growth in plants are
not easily detected (Banuelos & Obeso, 2004). Recently, Monks
& Kelly (2006) identified such relationships in Nothofagus,
which has a reproductive behaviour very similar to Fagus
(Richardson et al., 2005). Other authors had already reported a
negative relationship between growth and previous summer
temperature in Nothofagus menziesii without providing an
explanation for it (Cullen et al., 2001). On the other hand,
Fagus crenata does not always show a tree-ring response to
mast years (Yasumura et al., 2006). Mechanistic explanations
will most likely need to focus on the partitioning of carbon
allocation between reproductive and vegetative pathways (see
Hoch, 2005; and Yasumura et al., 2006). In our study, moving
correlations suggested an increased importance of this inter-
action during recent times, possibly due to the combination of
climatic warming (Schmidt, 2006) and the progressive aging
and development of sampled trees.
Close to the upper edge of its altitudinal range, beech
benefits from higher temperature and suffers from excessive
precipitation in late spring (May) and during summer. The
same behaviour was observed in central European high-
elevation beech populations (830–1240 m a.s.l.) (Dittmar
et al., 2003), and can be related to a thermal limit imposed
by altitude, plus the associated negative effect of cloudiness
and soil saturation. A similar temperature response during the
growing season is typical of conifer species growing at high
elevations in the Alps (Frank & Esper, 2005) and Pyrenees
(Tardif et al., 2003), or at high latitudes in Europe (e.g.,
Makinen et al., 2002;99 Linderholm et al., 2003). Moving
correlations showed a transient response of the main climatic
signals, and similar behaviour was recently reported in a
dendroclimatic study of Larix decidua in the same region
(Carrer & Urbinati, 2006).
A. Di Filippo et al.
14 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

A word of caution is needed on the interpretation of MCF.
In fact, it is simplistic to consider these changes in moving
correlations as evidence of deviation from the ‘principle of
uniformitarianism’ (Camardi, 1999), which assumes that
modern natural processes have acted similarly in the past,
and is equivalent to the statistical assumption of ‘stationarity’.
First, because results are based on empirical relationships,
changes in data quality (both dendrochronological and
instrumental) can be responsible for different correlation
values over time. Second, the ‘uniformity principle’ commonly
refers to the fact that limiting factors controlled tree-ring
parameters in the past just as they do today, but the role of
different factors at a single location or over an entire region
could change over time. This possibility has been raised, for
example, to explain the ‘divergence’ between temperature and
ring parameters (width and maximum latewood density)
during the late 20th century (Jacoby & D’Arrigo, 1995; Briffa
et al., 1998). In Alaska, recent increases in air temperature are
not reflected in tree-ring thickness because water (that is,
drought stress) has become the limiting factor (Barber et al.,
2000; Lloyd & Fastie, 2002; Wilmking & Juday, 2005). In
Siberia, on the other hand, reduced correlation of growth rates
with summer temperature has been attributed to increasing
winter precipitation, which leads to delayed snowmelt in
permafrost environments, thus shortening the tree growing
season (Vaganov et al., 1999).
The second PC of the 12 site chronologies in the Eastern
Alps showed contrasting growth patterns for low and high
mountain sites. Dendroclimatic results pointed to an environ-
mental gradient of increasing summer drought with low
elevation sites negatively influenced and high elevation ones
impacted positively (Table 5). Reversing tree-ring responses to
a certain climatic factor along an altitudinal gradient were
previously reported for European beech in the central Apen-
nines (Piovesan et al., 2005a), for Douglas fir and mountain
hemlock in western North America (Fagre et al., 2003; Zhang
& Hebda, 2004), and for birch in Japan (Takahashi et al.,
2005). Environmental differences associated with elevation
gradient, such as a decrease in temperature and in the length of
the growing season with increase in elevation, may lead to such
spatial variation in radial growth. With future warming, we
hypothesize a reduction of growth at the lower elevations,
whereas high-mountain beech productivity could increase due
to a milder and longer vegetative period (see also Nakawatase
& Peterson, 2006). At the lower elevations, reduced temper-
atures in previous summer–early autumn can favour harden-
ing-related processes that reduce the risk of early frost damage,
whereas mildness in November could enhance the storage of
reserves for the next growing season (Barbaroux & Breda,
2002)1010 .
Teleconnection analysis revealed that the Eastern Alps
climatic signal is still found in the Central Alps, spreads
northward in Austria, eastward to Slovenia, and southward to
the northern Apennines. The northern Apennines occupy an
intermediate position, as beech chronologies from that area are
correlated with both Alpine and central-southern Apennine
chronologies (Piovesan et al., 2005a). This mountain range is
placed in a transition zone between the Mediterranean and
Temperate climate (e.g. Walter, 1985; Bailey, 1996). From a
climatic point of view, the distinction between the Apennines
and the Alps is mainly a difference in precipitation regime
(Trewartha, 1968), as was confirmed by a recent climatic
zonation of Italy (Brunetti et al., 2006). As a possible
consequence, beech growth in central-southern Italy is char-
acterized by a dominant response to summer drought (Piov-
esan et al., 2005a), while in the Alps the response shifts
towards thermal factors (Di Filippo, 2006). Considering the
autoecology and biogeography of European beech, it is
reasonable to expect higher exposure to drought stress toward
the southern limit of the geographic range of the beech
(Becker, 1981), and an increased importance of temperature
moving northward and upward (Dittmar & Elling, 2005;1111 Fang
& Lechowicz, 2006). Evidence for the spatial separation
between these two bioclimatic zones emerged even in a study
of the climatic patterns influencing conifer tree rings across the
Northern Hemisphere (Fig. 7 in Briffa et al., 2002). Further
confirmation can be found in the bioclimatic classification of
Italy proposed by Pignatti (1979) according to vegetation
analysis.
Low elevation chronologies were correlated through central
and northern Italy and in Slovenia, suggesting the existence
of common climatic factors controlling growth at these
altitudes. Late spring–summer drought emerged as important
for tree-ring formation in both central Italy (Piovesan et al.,
2005a) and low elevation stands in the Julian Alps. From the
teleconnection analysis, a similar influence extended to the
Dinaric Mountains and south-east Slovenian hills. In partic-
ular, May climate seems to be important at hilly sites,
possibly because higher temperature at the beginning of the
growing season allows for early crown development, exposing
beech to water stress during the summer (Piovesan et al.,
2005a). This expansion of the Mediterranean region up to the
Pre-Alps is consistent with placing the Julian Alp sites at the
boundary between the Mediterranean Mountains and the
Alpine Environmental Zones (Metzger et al., 2005; see also
Zohary, 1973). Thus, TOB (Julian Alps) and GRA (Carnic
Alps), even though located at the same elevation on a
southern exposure, belong to, respectively, the low elevation
and the mountain range. Additional evidence of Mediterra-
nean influences at these sites comes from the presence upon
rocky cliffs of the evergreen holm oak (Quercus ilex L.), and
from the cultivation of the olive tree (Olea europea L.). These
two species reach the northernmost limit of distribution (the
former) and cultivation (the latter) in north-eastern Italy
(Pignatti, 1998). In low-elevation Julian stands, the response
to drought appears combined with one to early-frost damage.
A mixed response to summer drought stress and previous
autumn–winter temperature was found in hilly beech tree-
ring chronologies of central and eastern Europe (Dittmar
et al., 2003). In French beech sites, such responses are
generally observed below 600 m a.s.l. (Lebourgeois et al.,
2005).
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 15ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Genetic research has showed a different origin of the
Apennine and Alpine beech populations (Vettori et al., 2004;
Magri et al., 2006). The Apennines and the southern part of
the Balkan Peninsula were Mediterranean refuge areas for
European beech during the ice ages. These refugia were
separated from Central European populations, from which the
Alpine Fagus sylvatica stands originated (Magri et al., 2006).
Even though beech radial growth is sensitive to summer
drought both in the Apennines and at low elevation in the
Alps, the Mediterranean populations seem more drought
resistant (Nahm et al., 2006). Because summer drought is
becoming increasingly important for beech forest dynamics
(and eventually for natural selection) in central Europe
(Czajkowski et al., 2005) and in the Mediterranean Basin
(Jump et al., 2006), it is vital to understand how beech
populations with varying genetic material have responded to
drought in the past.
CONCLUSION
The Alps and the Apennines belong to two different biocli-
matic zones, both possessing a clear vertical dendroclimatic
zonation in altitudinal ranges, each with its own distinctive
climatic signal (Dittmar et al., 2003; Piovesan et al., 2005a). In
both zones, response to low temperature increases with
altitude. However, this phenomenon is more evident in the
Alps than the Apennines, probably because at high elevations
low temperatures can be limiting even in the middle of the
growing season (Dittmar et al., 2003). In the high-mountain
Mediterranean environment, temperature response is concen-
trated in early spring (late-frost damage and/or temperature
requirements for growth reactivation). Alpine high-elevation
beechwoods have dendroclimatic signals opposite those for
low elevation ones, which correlate with precipitation in May
and coolness in summer (Dittmar & Elling, 1999; Dittmar
et al., 2003). The cause may be the altitudinal difference in
temperature regime and, in particular, an altitude-mediated
phenological shift in growing season onset (Dittmar & Elling,
2005). While drought in the Apennines is a limiting factor
across all Mediterranean settings, from hilly to tree-line beech
stands (Piovesan et al., 2005a), its influence in the Alps
remains limited to the low elevation environments. Thus,
despite similar elevational response, summer influence on tree-
ring chronologies marks the bioclimatic separation between
the Apennines and the Alps.
Is this bioclimatic difference stable through time? Moving
response functions suggest a recent expansion of the
growing season for Alpine high-elevation beechwoods. As
for plant phenological changes (e.g. Parmesan & Yohe,
2003), the observed changes can be explained by the positive
temperature trend reported for Italy over the last 130 years
(Brunetti et al., 2006). This phenomenon is emphasized
when considering that the first decades used in moving
correlations fall within the Little Ice Age, which ended in
central Europe during the second half of the 19th century
(Xoplaki et al., 2005). Under a changing climate, bioclimatic
shifts could characterize vegetation arranged along altitudinal
gradients or at ecotonal boundaries (e.g. Penuelas & Boada,
2003). The beech old-growth network presented here will
therefore be useful to assess any future changes in forest
growth related to climate. As dendroecological data sets
provide ground-truth information on spatial and temporal
variability of climate–forest interactions, they can also
benefit the successful implementation of adaptive manage-
ment strategies.
ACKNOWLEDGEMENTS
We are grateful to Livio Silverio and to the Forest Service of
Friuli–Venezia Giulia for their support during the sampling
excursions, and to Anze Rutar for his contribution to beech
research at the Tolmin site. FB thanks Stanford University for
sabbatical support. KC thanks the Ministry of Education,
Science, and Sport of the Republic of Slovenia, Research
Program ‘Lesarstvo’, for financial support. The comments of
the Handling Editor and two anonymous referees helped in
improving on an earlier version of this manuscript.
REFERENCES
Aniol, R.W. (1983) Tree-ring analysis using CATRAS. Den-
drochronologia, 1, 45–53.
Aniol, R.W. (1987) A new device for Computer Assisted
Measurement of Tree-Ring Widths. Dendrochronologia, 5,
135–141.
Bailey, R.G. (1996) Ecosystem geography. Springer Verlag, New
York, NY.
Banuelos, M.-J. & Obeso, J.-R. (2004) Resource allocation in
the dioecious shrub Rhamnus alpinus: the hidden costs of
reproduction. Evolutionary Ecology Research, 6, 397–413.
Barbaroux, C. & Breda, N. (2002) Contrasting distribution and
seasonal dynamics of carbohydrate reserves in stem wood of
adult ring-porous sessile oak and diffuse porous beech trees.
Tree Physiology, 22, 1201–1210.
Barber, V.A., Juday, G.P. & Finney, B.P. (2000) Reduced growth
of Alaskan white spruce in the twentieth century from tem-
perature-induced drought stress. Nature, 405, 668–673.
Becker, M. (1981) Ecologie du hetre et de la hetraie – Ca-
racterisation climatique. Le hetre (ed. by E. Teissier du Cros,
F. Le Tacon, G. Nepveu, J. Parde, R. Perrin and J. Timbal),
pp. 71–77, INRA, Paris.
Bernabei, M., Lo Monaco, A., Piovesan, G. & Romagnoli, M.
(1996) Dendrocronologia del faggio (Fagus sylvatica L.) sui
Monti Sabini (Rieti). Dendrochronologia, 14, 59–70.
Biondi, F. (1992) Development of a tree-ring network for the
Italian Peninsula. Tree-Ring Bulletin, 52, 15–29.
Biondi, F. (1993) Climatic signals in tree-rings of Fagus sylv-
atica L. from the central Apennines, Italy. Acta Oecologica,
14, 57–71.
Biondi, F. (1997) Evolutionary and moving response functions
in dendroclimatology. Dendrochronologia, 15, 139–150.
A. Di Filippo et al.
16 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Biondi, F. (1999) Comparing tree-ring chronologies and
repeated timber inventories as forest monitoring tools.
Ecological Applications, 9, 216–227.
Biondi, F. (2000) Are climate–tree growth relationships
changing in north-central Idaho? Arctic, Antarctic and
Alpine Research, 32, 111–116.
Biondi, F. & Swetnam, T.W. (1987) Box–Jenkins models of
forest interior tree-ring chronologies. Tree-Ring Bulletin, 47,
71–95.
Biondi, F. & Visani, S. (1996) Recent developments in the
analysis of an Italian tree-ring network with emphasis on
European beech (Fagus sylvatica L.). Tree rings, Environ-
ment and Humanity (ed. by J.S. Dean, D.M. Meko and
T.W. Swetnam), pp. 713–725. Radiocarbon XXXXXX,
XXXXXX1212 .
Biondi, F. & Waikul, K. (2004) DENDROCLIM2002: a C++ pro-
gram for statistical calibration of climate signals in tree-ring
chronologies. Computers and Geosciences, 30, 303–311.
Bourquin-Mignot, C. & Girardclos, O. (2001) Construction
d’une longue chronologie de hetres au pays basque: la foret
d’Iraty et le Petit Age Glaciaire. Revue de Geographie des
Pyrenees et du Sud-Ouest Europeen, 11, 59–71.
Briffa, K.R. & Jones, P.D. (1990) Basic chronology statistics
and assessment. Methods of dendrochronology: applications in
the environmental sciences(ed. by E.R. Cook and L.A. Kai-
riukstis (ed by *), pp. 137–152. International Institute for
Applied Systems Analysis (IIASA). Kluwer Academic Pub-
lishers, Dodrecht.1313
Briffa, K.R., Schweingruber, F.H., Jones, P.D., Osborn, T.J.,
Shiyatov, S.G. & Vaganov, E.A. (1998) Reduced sensitivity of
recent tree-growth to temperature at high northern lati-
tudes. Nature, 391, 678–682.
Briffa, K.R., Osborn, T.J., Schweingruber, F.H., Jones, P.D.,
Shiyatov, S.G. & Vaganov, E.A. (2002) Tree-ring width and
density data around the Northern Hemisphere: Part 1, local
and regional climate signals. The Holocene, 12, 737–757.
Brunetti, M., Maugeri, M., Monti, F. & Nanni, T. (2006)
Temperature and precipitation variability in Italy in the last
two centuries from homogenised instrumental time series.
International Journal of Climatology, 26, 345–381.
Camardi, G. (1999) Charles Lyell and the Uniformity Princi-
ple. Biology and Philosophy, 14, 537–560.
Canham, C.D. (1990) Suppression and release during canopy
recruitment in Fagus grandifolia. Bulletin of the Torrey
Botanical Club, 117, 1–7.
Carrer, M. & Urbinati, C. (2006) Long-term change in the
sensitivity of tree-ring growth to climate forcing in Larix
decidua. New Phitologist, 170, 861–872.
Castorina, R., Di Filippo, A. & Piovesan, G. (2005) Dendro-
ecology of beech (Fagus sylvatica L.) on Mount Etna Natural
Park (Sicily, Italy). Abstract book of Eurodendro2005 (ed. by
M. Sarlatto, A. Di Filippo, G. Piovesan and M. Romagnoli),
pp. 44, Editore Sette Citta, Viterbo.
Cook, E.R. & Peters, K. (1997) Calculating unbiased tree-ring
indices for the study of climatic and environmental change.
The Holocene, 7, 361–370.
Cook, E.R., Glitzenstein, J.S., Krusic, P.J. & Harcombe, P.A.
(2001) Identifying functional groups of trees in West Gulf
Coast forests (USA): a tree-ring approach. Ecological
Applications, 11, 883–903.
Cullen, L.E., Palmer, J.G., Duncan, R.P. & Stewart, G.H. (2001)
Climate change and tree-ring relationships of Nothofagus
menziesii tree-line forests. Canadian Journal of Forest Re-
search, 31, 1981–1991.
Czajkowski, T., Kuhling, M. & Bolte, A. (2005) Impact of the
2003 summer drought on growth of beech sapling natural
regeneration (Fagus sylvatica L.) in north-eastern Central
Europe. Allgemeine Forst- und Jagdzeitung, 176, 133–143 [In
German].
Desiato, F., Lena, F., Baffo, F., Suatoni, B. & Toreti, A. (2005)
Indicatori del clima in Italia elaborati attraverso il sistema
SCIA. APAT, Rome.
Di Filippo, A. (2006) Study for a bioclimatic classification of
Italian beech forests on a dendroclimatic basis. PhD thesis,
Universita degli Studi della Tuscia, Viterbo (In italian).
Dittmar, C. & Elling, W. (1999) Radial growth of Norway
spruce and European beech in relation to weather and
altitude. Forstwissenschaftliches Centralblatt, 118, 251–270.
Dittmar, C. & Elling, W. (2005) Phenological phases of com-
mon beech (Fagus sylvatica L.) and their dependence on
region and altitude in Southern Germany. European Journal
of Forest Research, 125, 181–188.
Dittmar, C., Zech, W. & Elling, W. (2003) Growth variations
of common beech (Fagus sylvatica L.) under different cli-
matic and environmental conditions in Europe: a dendro-
ecological study. Forest Ecology and Management, 173,
63–78.
Dittmar, C., Fricke, W. & Elling, W. (2006) Impact of late frost
events on radial growth of common beech (Fagus sylvatica
L.) in Southern Germany. European Journal of Forest
Research, 125, 249–259.
Douglass, A.E. (1920) Evidence of climatic effects in the annual
rings of trees. Ecology, 1, 24–32.
Eckstein, D. & Frisse, E. (1982) The influence of temperature
and precipitation on vessel area and ring width of oak and
beech. Climate from tree rings (ed. by M.K. Hughes, P.M.
Kelly, J.R. Pilcher and V.C. LaMarche Jr), p. 12. Cambridge
University Press, Cambridge.
Efron, B. & Tibshirani, R. (1986) Bootstrap methods for
standard errors, confidence intervals, and other measures of
statistical accuracy. Statistical Science, 1, 54–75.
Ellenberg, H. (1988) Vegetation ecology of Central Europe.
Cambridge University Press, Cambridge.
Fagre, D.B., Peterson, D.L. & Hessl, A.E. (2003) Taking the
pulse of the mountains: ecosystem responses to climatic
variability. Climatic Change, 59, 263–282.
Fang, J. & Lechowicz, M.J. (2006) Climatic limits for the
present distribution of beech (Fagus L.) species in the world.
Journal of Biogeography, 33, 1804–1819.
Frank, D. & Esper, J. (2005) Characterization and climate
response patterns of a high elevation, multi-species tree-ring
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 17ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

network in the European Alps. Dendrochronologia, 22,
107–121.
Fritts, H.C. (1976) Tree rings and climate. Academic Press,
London.
Graham, E.A., Mulkey, S.S., Kitajima, K., Phillips, N.G. &
Wright, S.J. (2003) Cloud cover limits net CO2 uptake and
growth of a rainforest tree during tropical rainy seasons.
Proceedings of the National Academy of Sciences USA, 100,
572–576.
Grissino-Mayer, H.D. (2001) Evaluating crossdating accuracy:
a manual and tutorial for the computer program cofecha.
Tree-Ring Research, 57, 205–221.
Guiot, J. (1991) The bootstrapped response function. Tree-
Ring Bulletin, 51, 39–41.
Gutierrez, E. (1988) Dendroecological study of Fagus sylvatica
L. in the Montseny mountains (Spain). Acta Oecologica, 9,
301–309.
Hampe, A. & Petit, R.J. (2005) Conserving biodiversity under
climate change: the rear edge matters. Ecology Letters, 8,
461–467.
Helama, S., Lindholm, M., Timonen, M. & Eronen, M. (2004)
Detection of climate signal in dendrochronological data
analysis: a comparison of tree-ring standardization methods.
Theoretical and Applied Climatology, 79, 239–254.
Hoch, G. (2005) Fruit-bearing branchlets are carbon auton-
omous in mature broad-leaved temperate forest trees. Plant,
Cell and Environment, 28, 651–659.
Holmes, R.L. (1983) Computer-assisted quality control in tree-
ring dating and measurement. Tree-Ring Bulletin, 43, 69–78.
Jacoby, G.C. & D’Arrigo, R.D. (1995) Tree-ring width and
density evidence of climatic and potential forest change in
Alaska. Global Biogeochemical Cycles, 9, 227–234.
Jump, A.S. & Penuelas, J. (2005) Running to stand still:
adaptation and the response of plants to rapid climate
change. Ecology Letters, 8, 1010–1020.
Jump, A.S., Hunt, J.M. & Penuelas, J. (2006) Rapid climate
change-related growth decline at the southern range-edge of
Fagus sylvatica. Global Change Biology, Xxx, xxx–xxx1414 .
Kaiser, H.F. (1992) On Cliff’s formula, the Kaiser–Guttman
rule, and the number of factors. Perceptual and Motor Skills,
74, 595–598.
Kienast, F., Schweingruber, F.H., Braker, O.L. & Schar, E.
(1987) Tree-ring studies on conifers along ecological gra-
dients and the potential of single-year analyses. Canadian
Journal of Forest Research, 17, 683–96.
Laurent, J.M., Bar-Hen, A., Francois, L., Ghislain, M. &
Cheddadi, R. (2004) Refining vegetation simulation models:
from plant functional types to bioclimatic affinity groups of
plants. Journal of Vegetation Science, 15, 739–746.
Lebourgeois, F., Breda, N., Ulrich, E. & Granier, A. (2005)
Climate–tree-growth relationships of European beech
(Fagus sylvatica L.) in the French Permanent Plot Network
(RENECOFOR). Trees, 19, 385–401.
Lemoine, D., Granier, A. & Cochard, H. (1999) Mechanism of
freeze induced embolism in Fagus sylvatica L. Trees, 13,
206–210.
Linderholm, H.W., Solberg, B.Ø. & Lindholm, M. (2003) Tree-
ring records from central Fennoscandia: the relationship
between tree growth and climate a west–east transect. The
Holocene, 13, 889–897.
Lloyd, A.H. & Fastie, C.L. (2002) Spatial and temporal vari-
ability in the growth and climate response of treeline trees in
Alaska. Climatic Change, 52, 481–509.
Lorimer, C.G., Dahir, S.E. & Singer, M.T. (1999) Frequency of
partial and missing rings in Acer saccharum in relation to
canopy position and growth rate. Plant Ecology, 143, 189–202.
Ludwig, J.A. & Reynolds, J.F. (1988) Statistical ecology. John
Wiley and Sons, New York.
Magri, D., Vendramin, G.G., Comps, B., Dupanloup, I., Geb-
urek, T., Gomory, D., Latalowa, M., Litt, T., Paule, L., Roure,
J.M., Tantau, I., van der Knaap, W.O., Petit, R.J. & de Beau-
lieu, J.L. (2006) A new scenario for the Quaternary history of
European beech populations: palaeobotanical evidence and
genetic consequences. New Phytologist, 171, 199–221.
Makinen, H., Nojd, P., Kahle, H.-P., Neumann, U. & Tveite, B.
(2002) Radial growth variation of Norway spruce (Picea
abies (L.) Karst.) across latitudinal and altitudinal gradients
in central and northern Europe. Forest Ecology and Man-
agement, 171, 243–259.
Meko, D.M., Cook, E.R., Stahle, D.W., Stockton, C.W. &
Hughes, M. K. (1993) Spatial patterns of tree-growth
anomalies in the United States and southeastern Canada.
Journal of Climate, 6, 1773–1786.
Mencuccini, M. & Piussi, P. (1995) Production of seeds and
cones and consequences for wood radial increment in
Norway spruce (Picea abies (L.) Karst.). Giornale Botanico
Italiano, 129, 797–812.1515
Mennella, C. (1967) Il clima d’Italia. Fratelli Conti Editori,
Naples.
Metzger,M.J., Bunce, R.G.H., Jongman, R.H.G., Mucher, C.A. &
Watkins, J.W. (2005) A climatic stratification of the environ-
ment of Europe.Global Ecology andBiogeography, 14, 549–563.
Monks, A. & Kelly, D. (2006) Testing the resource-matching
hypothesis in the mast seeding tree Nothofagus truncata.
Austral Ecology, 31, 366–375.
Monserud, R.A. & Sterba, H. (1996) A basal area increment
model for individual trees growing in even- and uneven-
aged forest stands in Austria. Forest Ecology and Manage-
ment, 80, 57–80.
Nahm, M., Radoglou, K., Halyvopoulos, G., Geßler, A., Ren-
nenberg, H. & Fotelli, M.N. (2006) Physiological perform-
ance of beech (Fagus sylvatica L.) at its Southeastern
distribution limit in Europe: seasonal changes in nitrogen,
carbon and water balance. Plant Biology, 8, 52–63.
Nakawatase, J.M. & Peterson, D.L. (2006) Spatial variability in
forest growth–climate relationships in the Olympic Moun-
tains, Washington. Canadian Journal of Forest Research, 36,
77–91.
Nielsen, C.N. & Jørgensen, F.V. (2003) Phenology and diam-
eter increment in seedlings of European beech (Fagus sylv-
atica L.) as affected by different soil water contents: variation
A. Di Filippo et al.
18 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

between and within provenances. Forest Ecology and Man-
agement, 174, 233–249.
Paiero, P., Candidi-Tommasi, R. & Caniglia, G. (1975) Il bosco
‘bandito’ di Cleulis (Paluzza): fustaia di faggio derivata
dall’invecchiamento naturale di un ceduo matricinati. Monti
e Boschi, 4, 1–13.
Parmesan, C & Yohe, G. (2003) A globally coherent fingerprint
of climate change impacts across natural systems. Nature,
421, 37–42.
Penuelas, J. (2005) A big issue for trees. Nature, 437, 965–966.
Penuelas, J. & Boada, M. (2003) A global change-induced
biome shift in the Montseny Mountains (NE Spain). Global
Change Biology, 9, 131–140.
Pignatti, S. (1979) I piani di vegetazione in Italia. Giornale
Botanico Italiano, 113, 411–428.
Pignatti, S. (1998) I boschi d’Italia. UTET, Turin.
Piovesan, G. & Adams, J.M. (2001) Masting behaviour in
beech: linking reproduction and climatic variation. Cana-
dian Journal of Botany, 79, 1039–1047.
Piovesan, G. & Bernabei, M. (1997) L’influenza delle preci-
pitazioni estive sulla crescita e la riproduzione del faggio
(Fagus sylvatica L.) in una stazione meridionale dell’areale.
Italia Forestale e Montana, 6, 444–459.
Piovesan, G. & Schirone, B. (2000) Winter North Atlantic Oscil-
lation effects on the tree rings of the Italian beech (Fagus sylv-
atica L.). International Journal of Biometeorology, 44, 121–127.
Piovesan, G., Bernabei, M., Di Filippo, A., Romagnoli, M. &
Schirone, B. (2003) A long-term tree ring beech chronology
from a high-elevation old-growth forest of Central Italy.
Dendrochronologia, 21, 1–10.
Piovesan, G., Biondi, F., Bernabei, M., Di Filippo, A. & Schi-
rone, B. (2005a) Spatial and altitudinal bioclimatic zones of
the Italian Peninsula identified from a beech (Fagus sylvatica
L.) tree-ring network. Acta Oecologica, 27, 197–210.
Piovesan, G., Di Filippo, A., Alessandrini, A., Biondi, F. &
Schirone, B. (2005b) Structure, dynamics and dendroecol-
ogy of an old-growth Fagus forest in the Apennines. Journal
of Vegetation Science, 16, 13–28.
Piutti, E. & Cescatti, A. (1997) A quantitative analysis of the
interactions between climatic response and intraspecific
competition in European beech. Canadian Journal of Forest
Research, 27, 277–284.
Richardson, S.J., Allen, R.B., Whitehead, D., Carswell, F.E.,
Ruscoe, W.A. & Platt, K.H. (2005) Climate and net carbon
availability determine temporal patterns of seed production
by Nothofagus. Ecology, 86, 972–981.
Rinn, F. (1996) TSAP: Reference Manual (Version 3.0). Xxxxxx,
Heidelberg1616 .
Rolland, C. (2002) Decreasing teleconnections with inter-site
distance in monthly climatic data and tree-ring width net-
works in a mountainous Alpine area. Theoretical and Applied
Climatology, 71, 63–75.
Rozas, V. (2001) Detecting the impact of climate and distur-
bances on tree-rings of Fagus sylvatica L. and Quercus robur
L. in a lowland forest in Cantabria, Northern Spain. Annals
of Forest Science, 58, 237–251.
Schmidt,W. (2006) Temporal variation in beechmasting (Fagus
sylvatica L.) in a limestone beech forest (1981–2004). Allg-
emeine Forst- und Jagdzeitung, 177, 9–19 (In German).
Schweingruber, F.H. (1996) Tree rings and environment den-
droecology. Swiss Federal Institute of Forest, Snow and
Landscape Researches, WSL/FNP, Birmensdorf.
Selas, V., Piovesan, G., Adams, J.M. & Bernabei, M. (2002)
Climatic factors controlling reproduction and growth of
Norway spruce in southern Norway. Canadian Journal of
Forest Research, 32, 217–225.
Stenson, H. & Wilkinson, L. (2004) Factor analysis. systat 11
Statistics I (ed. by systat Software, Inc.), pp. 359–377,
Richmond, CA.
Stokes, M.A. & Smiley, T.L. (1996) An introduction to tree-ring
dating. University of Arizona Press, Tucson, AZ.
Takahashi, K., Tokumitsu, Y. & Yasue, K. (2005) Climatic
factors affecting the tree-ring width of Betula ermanii at the
timberline on Mount Norikura, central Japan. Ecological
Research, 20, 445–451.
Tardif, J., Camarero, J.J., Ribas, M. & Gutierrez, E. (2003)
Spatiotemporal variability in tree growth in the Central
Pyrenees: climatic and site influences. Ecological Mono-
graphs, 73, 241–257.
Thompson, R.S., Shafer, S.L., Anderson, K.H., Strickland, L.E.,
Pelltier, R.T., Bartlein, P.J. & Kerwin, M.W. (2005) Topo-
graphic, bioclimatic, and vegetation characteristics of three
ecoregion classification systems in North America: com-
parisons along continent-wide transects. Environmental
Management, 34, S125–S148.
Trewartha, G.T. (1968) An introduction to climate. McGraw-
Hill, New York, NY.
Urbinati, C., Carrer, M. & Sodiro, S. (1997) Dendroclimatic
response variability of Pinus cembra L. in upper timberline
forests of Italian eastern Alps. Dendrochronologia, 15,
101–17.
Vaganov, E.A., Hughes, M.K., Kirdyanov, A.V., Schweingru-
ber, F.H. & Silkin, P.P. (1999) Influence of snowfall and
melt timing on tree growth in subarctic Eurasia. Nature,
400, 149–151.
Vettori, C., Vendramin, G.G., Anzidei, M., Pastorelli, R., Paf-
fetti, D. & Giannini, R. (2004) Geographic distribution of
chloroplast of beech (Fagus sylvatica L.). Theoretical and
Applied Genetics, 109, 1–9.
von Wuehlisch., G. (2006) EUFORGEN Technical Guidelines
for genetic conservation and use for European beech (Fagus
sylvatica). International Plant Genetic Resources Institute,
Rome, Italy. http://www.ipgri.cgiar.org/networks/euforgen/
Euf_Distribution_Maps.asp
Walter, H. (1985) Vegetation of the Earth and ecological systems
of the geobiosphere. Springer-Verlag, Berlin.
Wessel, P. & Smith, W.H.F. (1998) New, improved version of
Generic Mapping Tools released. EOS Transactions of the
American Geophysical Union, 79, 579.
White, M.A., Running, S.W. & Thornton, P.E. (1999) The
impact of growing-season length variability on carbon
assimilation and evapotranspiration over 88 years in the
Bioclimatology of beech in the Eastern Alps
Journal of Biogeography 19ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

eastern US deciduous forest. International Journal of Bio-
meteorology, 42, 139–145.
Whittaker, R.J., Araujo, M.B., Jepson, P., Ladle, R.J., Watson,
J.E.M. & Willis, K.J. (2005) Conservation biogeography:
assessment andprospect.Diversity andDistributions, 11, 3–23.
Wigley, T.M.L., Briffa, K.R. & Jones, P.D. (1984) On the
average value of correlated time series, with applications in
dendroclimatology and hydrometeorology. Journal of Ap-
plied Meteorology, 23, 201–213.
Wilmking, M. & Juday, G.P. (2005) Longitudinal variation of
radial growth at Alaska’s northern treeline: recent changes
and possible scenarios for the 21st century. Global and
Planetary Change, 47, 282–300.
Woodward, F.I. (1987) Climate and plant distribution. Cam-
bridge University Press, Cambridge.
Woodward, F.I. & Cramer, W. (1996) Plant functional types
and climatic changes: introduction. Journal of Vegetation
Science, 7, 306–308.1717
Woodward, A., Silsbee, D.G., Schreiner, E.G. & Means, J.E.
(1994) Influence of climate on radial growth and cone
production in subalpine fir (Abies lasiocarpa) and mountain
hemlock (Tsuga mertensiana). Canadian Journal of Forest
Research, 24, 1133–1143.
Xoplaki, E., Luterbacher, J., Paeth, H., Dietrich, D., Steiner, N.,
Grosjean, M. & Wanner, H. (2005) European spring and
autumn temperature variability and change of extremes over
the last half millennium. Geophysical Research Letters, 32,
000–000.
Yasumura, Y., Hikosaka, K. & Hirose, T. (2006) Resource
allocation to vegetative and reproductive growth in relation
to mast seeding in Fagus crenata. Forest Ecology and Man-
agement, 229, 228–233.
Zhang, Q.-B. & Hebda, R.J. (2004) Variation in radial growth
patterns of Pseudotsuga menziesii on the central coast of
British Columbia, Canada. Canadian Journal of Forest
Research, 34, 1946–1954.
Zohary, M. (1973) Geobotanical foundations of the Middle East.
Fischer Verlag, Stuttgart, DE.
SUPPLEMENTARY MATERIAL
The following supplementary material is available for this
article online:
Table S1 Correlation matrix for the 14 prewhitened beech site
chronologies (period 1942–2001).
Table S2 Bootstrap correlation (a) and response (b) function
coefficients calculated between the Dunkelsteinerwald pre-
whitened site chronology and the 17-month climatic window
for the period 1942–2001.
Table S3 Bootstrap correlation (a) and response (b) function
coefficients calculated between the Hallstatt prewhitened site
chronology and the 17-month climatic window for the period
1942–2001.
Table S4 Summary of composite tree-ring chronologies,
which were developed using three sites each.
Figure S1 Expressed population signal (EPS) statistics for the
14 prewhitened site chronologies arranged according to their
length.
Figure S2 Eigenvectors (loadings) of the first two components
for the 14 prewhitened site chronologies (a). Eigenvalues
(percentage of explained variance) of the first two components
(b, total; c, PC1; d, PC2).
Figure S3 Correlation between climatic data used in dendro-
climatic analyses.
This material is available as part of the online article
from: http://www.blackwell-synergy.com/doi/abs/10.1111/j.
1365-2699.2007.01747.x
Please note: Blackwell Publishing are not responsible for the
content or functionality of any supplementary materials
supplied by the authors. Any queries (other than missing
material) should be directed to the corresponding author for
the article.
BIOSKETCHES
Alfredo Di Filippo conducted this research as a part of his
PhD thesis. He is currently working at the Universita della
Tuscia (Italy) as a research assistant, conducting investigations
in the fields of dendroclimatic networks and dendroecology
applied to the study of old-growth forests and disturbance
regimes.
Franco Biondi directs the DendroLab at the University of
Nevada, Reno (USA). His experience and interests are in
dendrochronology, climate change at multiple spatial and
temporal scales, numerical analysis of proxy climate data, and
late-Holocene dynamics in forests, woodlands and mountains.
Gianluca Piovesan is professor of Dendrology and Ecolog-
ical Forest-Landscape Management at the Universita della
Tuscia (Italy). His research interests include old-growth forest,
masting, dendroecology, forest response to climatic variability
and restoration of forest ecosystems.
Editor: Mark Bush
A. Di Filippo et al.
20 Journal of Biogeography
ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd
Related Documents











