Biologia 64/6: 1047—1052, 2009 Section Cellular and Molecular Biology DOI: 10.2478/s11756-009-0190-8 Biochemical characterization of a raw starch degrading α-amylase from the Indonesian marine bacterium Bacillus sp. ALSHL3 Keni Vidilaseris 1 , Karina Hidayat 1 , Debbie S. Retnoningrum 2,3 , Zeily Nurachman 1 , Achmad Saefuddin Noer 1 & Dessy Natalia 1,3 * 1 Biochemistry Division, Faculty of Mathematics and Natural Sciences, Bandung Institute of Technology, Jl. Ganesha 10, Bandung, 40132, Indonesia; e-mail: [email protected] 2 School of Pharmacy, Bandung Institute of Technology, Jl. Ganesha 10, Bandung, 40132, Indonesia 3 Centre for Life Sciences, Bandung Institute of Technology, Jl. Ganesha 10, Bandung, 40132, Indonesia Abstract: An Indonesian marine bacterial isolate, which belongs to genus of Bacillus sp. based on 16S rDNA analysis and was identified as Bacillus filicolonicus according to its morphology and physiology, produced a raw starch degrading α-amylase. The partially purified α-amylase using a maize starch affinity method exhibited an optimum pH and temperature of 6.0 and 60 ◦ C, respectively. The enzyme retained 72% of its activity in the presence of 1.5 M NaCl. Scanning electron micrographs showed that the α-amylase was capable of degrading starch granules of rice and maize. This α-amylase from Bacillus sp. ALSHL3 was classified as a saccharifying enzyme since its major final degradation product was glucose, maltose, and maltotriose. Key words: α-amylase; Bacillus filicolonicus; raw starch; maize; marine. Abbreviations: DNSA, dinitrosalicylic acid; MB, marine broth; SBD, starch-binding domain. Introduction A critical step in starch hydrolysis is its solubilization at high temperatures, followed by liquefaction using ther- mostable amylases. A major reduction in energy de- mand in starch hydrolysis process can be achieved by avoiding starch solubilization and liquefaction at high temperatures by hydrolysis of granular starch at lower temperature using raw starch degrading α-amylase. The availability of suitable raw starch binding and de- grading α-amylases is currently very limited and they are largely identified in silico as putative enzymes via genome sequence analysis (Machovic & Janecek 2006). Approximately only 10% of all amylolytic enzymes pos- sess a distinct domain that facilitates binding to raw starch (Janecek & Sevcik 1999). α-Amylase (EC 3.2.1.1) from the family 13 of glycoside hydrolases cleaves internal α-1,4-glycosidic linkages in polysaccharides producing oligosaccharides and α-limit dextrins (van der Maarel et al. 2002). α- Amylases are multidomain proteins which consist of three main domains, domain A, B, and C (MacGregor et al. 2001). The structure of domain A is a (β/α) 8 - or TIM-barrel which contains two aspartates and one glutamate as the catalytic residues. Domain B is a long loop located between the third β-strand and the third α-helix of the TIM-barrel and forms an irregular β-like structure. Domain C constitutes the C-terminal part of enzyme and is made up of β strands. A few α-amylases contain additional domain named as a starch-binding domain (SBD) which has function to bind starch granules. SBDs are found in bacterial and fungal α-amylases (Janecek & Sevcik 1999). This domain is usually localized at the C-terminal end of enzymes. However α-amylase from Thermoactinomyces vulgaris has SBD at the N-terminus (Abe et al. 2004). Bacillus sp. IMD 434 degrades raw starch without ad- sorbing onto raw starch (Hamilton et al. 1998). Further- more, it should be noted that there are few amylolytic enzymes capable of binding and degrading raw starch without a specialized functional domain (Hamilton et al. 1998). Properties of α-amylases vary and reflect the growth environment of the producing organism. Indone- sia largely consists of a marine biosphere (70% of total area) that is an extremely rich natural habitat. We have screened this marine habitat for bacteria producing α- amylase enzymes. Forty-four of 267 isolates gave large clear zone on a iodine-strach plate assay and their α- amylase activities were measured at 37 ◦ C for 66 hours in liquid culture (D. S. Retroningrum & D. Natalia, un- published data). Two high producers were identified by 16S rDNA-based approach and based on their partial 16S rDNA nucleotide sequence, they have been identi- * Corresponding author c 2009 Institute of Molecular Biology, Slovak Academy of Sciences

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biologia 64/6: 1047—1052, 2009Section Cellular and Molecular BiologyDOI: 10.2478/s11756-009-0190-8
Biochemical characterization of a raw starch degrading α-amylasefrom the Indonesian marine bacterium Bacillus sp. ALSHL3
Keni Vidilaseris1, Karina Hidayat1, Debbie S. Retnoningrum2,3, Zeily Nurachman1,Achmad Saefuddin Noer1 & Dessy Natalia1,3*1Biochemistry Division, Faculty of Mathematics and Natural Sciences, Bandung Institute of Technology, Jl. Ganesha 10,Bandung, 40132, Indonesia; e-mail: [email protected] of Pharmacy, Bandung Institute of Technology, Jl. Ganesha 10, Bandung, 40132, Indonesia3Centre for Life Sciences, Bandung Institute of Technology, Jl. Ganesha 10, Bandung, 40132, Indonesia
Abstract: An Indonesian marine bacterial isolate, which belongs to genus of Bacillus sp. based on 16S rDNA analysisand was identified as Bacillus filicolonicus according to its morphology and physiology, produced a raw starch degradingα-amylase. The partially purified α-amylase using a maize starch affinity method exhibited an optimum pH and temperatureof 6.0 and 60◦C, respectively. The enzyme retained 72% of its activity in the presence of 1.5 M NaCl. Scanning electronmicrographs showed that the α-amylase was capable of degrading starch granules of rice and maize. This α-amylase fromBacillus sp. ALSHL3 was classified as a saccharifying enzyme since its major final degradation product was glucose, maltose,and maltotriose.
Key words: α-amylase; Bacillus filicolonicus; raw starch; maize; marine.
Abbreviations: DNSA, dinitrosalicylic acid; MB, marine broth; SBD, starch-binding domain.
Introduction
A critical step in starch hydrolysis is its solubilization athigh temperatures, followed by liquefaction using ther-mostable amylases. A major reduction in energy de-mand in starch hydrolysis process can be achieved byavoiding starch solubilization and liquefaction at hightemperatures by hydrolysis of granular starch at lowertemperature using raw starch degrading α-amylase.The availability of suitable raw starch binding and de-grading α-amylases is currently very limited and theyare largely identified in silico as putative enzymes viagenome sequence analysis (Machovic & Janecek 2006).Approximately only 10% of all amylolytic enzymes pos-sess a distinct domain that facilitates binding to rawstarch (Janecek & Sevcik 1999).
α-Amylase (EC 3.2.1.1) from the family 13 ofglycoside hydrolases cleaves internal α-1,4-glycosidiclinkages in polysaccharides producing oligosaccharidesand α-limit dextrins (van der Maarel et al. 2002). α-Amylases are multidomain proteins which consist ofthree main domains, domain A, B, and C (MacGregoret al. 2001). The structure of domain A is a (β/α)8-or TIM-barrel which contains two aspartates and oneglutamate as the catalytic residues. Domain B is a longloop located between the third β-strand and the thirdα-helix of the TIM-barrel and forms an irregular β-like
structure. Domain C constitutes the C-terminal part ofenzyme and is made up of β strands.A few α-amylases contain additional domain named
as a starch-binding domain (SBD) which has functionto bind starch granules. SBDs are found in bacterialand fungal α-amylases (Janecek & Sevcik 1999). Thisdomain is usually localized at the C-terminal end ofenzymes. However α-amylase from Thermoactinomycesvulgaris has SBD at the N-terminus (Abe et al. 2004).Bacillus sp. IMD 434 degrades raw starch without ad-sorbing onto raw starch (Hamilton et al. 1998). Further-more, it should be noted that there are few amylolyticenzymes capable of binding and degrading raw starchwithout a specialized functional domain (Hamilton etal. 1998).Properties of α-amylases vary and reflect the
growth environment of the producing organism. Indone-sia largely consists of a marine biosphere (70% of totalarea) that is an extremely rich natural habitat. We havescreened this marine habitat for bacteria producing α-amylase enzymes. Forty-four of 267 isolates gave largeclear zone on a iodine-strach plate assay and their α-amylase activities were measured at 37◦C for 66 hoursin liquid culture (D. S. Retroningrum & D. Natalia, un-published data). Two high producers were identified by16S rDNA-based approach and based on their partial16S rDNA nucleotide sequence, they have been identi-
* Corresponding author
c©2009 Institute of Molecular Biology, Slovak Academy of Sciences

1048 K. Vidilaseris et al.
fied as Bacillus sp. This current work focuses on the iso-late ALSH3 that was obtained from Cilauteuren, WestJava, Indonesia. It was isolated from sea water aboutfive meters from the shoreline.
α-Amylases previously characterized from Bacillussp. strain differ with respect to their molecular weights,and their optimal pH and temperature values. In thispaper we demonstrate a further distinguishing feature,degradation of granular raw starch, by an α-amylasefrom the marine Bacillus sp. ALSHL3 isolate.
Material and methods
Bacterial identificationAn α-amylase producing bacterium was isolated in pureculture from sea water samples collected from Cilauteuren,Pameungpeuk, Garut in West Java, Indonesia. The isolatewas grown in marine broth (MB) medium containing yeastextract 0.3% (b/v), peptone 0.5% (b/v), bacto agar 2%(b/v) and sea water at 37◦C. Bacteriology determinationof isolated strain was determined according to the methodsdescribed in Bergey’s Manual of Determinative Bacteriol-ogy (Buchanan et al. 1974) at Laboratory of Microbiology,School of Life Science and Technology, Bandung Instituteof Technology, Bandung, Indonesia.
Chromosomal DNA was isolated using Wizard Ge-nomic DNA Purification (Promega). The 16S rDNA genewas amplified by PCR using universal primers UniB1 andBactF1. The resulting 1.4 kb fragment was sequenced usingdye-end terminator (Macrogen, Korea).
Time course for enzyme productionA 1% (v/v) Bacillus sp. ALSHL3 overnight culture wastransferred into 100 mL MB medium containing 0.1% sol-uble starch (Sigma) and incubated in a shaking incubator(150 rpm) at 37◦C for 20 h. In time, samples were takenfor measurement of cell density (at 600 nm) and α-amylaseactivity. The culture supernatant was collected by centrifu-gation (11,100 × g for 3 min) and the α-amylase activitywas determined.
Raw starch affinity purificationPurification of α-amylase was conducted according to Na-jafi & Kembhavi (2005) with slight modification. A 5%(w/v) maize starch granule (Maizena, Honig) suspensionin aquadest was incubated for 10 min at room tempera-ture to wash the starch and then centrifuged at 11,100 × gfor 10 min. A 200 mL of crude enzyme was mixed with thestarch suspension and shaken gently for 50 min at 10◦C. Themixture was centrifuged at 11,100 × g for 10 min at 4◦C.Unbound enzyme was washed with 20 mM Tris-Cl bufferpH 7.0. Bound α-amylase was released by incubating with20 mM Tris-Cl buffer pH 7.0 at 50◦C for 30 min. The sus-pension was sentrifuged at 11,100 × g for 10 min and thesupernatant was colected and used for the α-amylase char-acterization. The enzyme was concentarated by CentriconMWCO 30 kDa (GE) at 8,000 × g. The unbound and boundamylases were subjected to protein gel electrophoresis andgel activity staining.
Enzyme assayDNS assay. The reducing sugar of α-amylase was deter-mined by the dinitrosalicylic acid (DNSA) method (Miller1959). A 25 µL of enzyme (10,671 U/mg) was mixed with25 µL of 0.75% (w/v) soluble starch (Sigma) dissolved in
50 mM universal buffer pH 6.0 at 60◦C for 10 minutes. α-Amylase activity was stopped by addition of 50 µL DNSAreagent. The reaction mixture was then incubated in boilledwater for 10 min. Subsequently, the reaction mixture wascooled down until reach room temperature and the ab-sorbance at 500 nm was measured. One unit of the α-amylase activity is defined as the amount of enzyme thatreleases the amount of reducing sugar equivalent to 1 µmolof reducing sugar under the assay conditions. All assays wereconducted in triplicates. D-glucose was used as a standard.
Iodine method. Fuwa method (Fuwa 1954) was usedto determine the effect of CaCl2 concentration on the α-amylase activity. A 50 µL of enzyme (44,464 U/mg) wasmixed with 50 µL of 0.125% (w/v) soluble starch (Sigma)dissolved in 50 mM universal buffer pH 6.0 at 60 ◦C. Oneunit of the α-amylase activity was defined as the amount ofenzyme that produced 10% reduction in starch-iodine stain-ing after 10 min of incubation under the experimental con-ditions. All assays were conducted in triplicates.
Effect of pH and temperature on α-amylase activityThe optimal pH of α-amylase activity was determined usingDNSA method by evaluating the hydrolysis reaction in therange of pH 4.0–10.0 using 50 mM universal buffers (succinicacid, NaH2PO4, glycine) at 60◦C. The optimal temperatureof α-amylase activity was determined by evaluating the hy-drolysis reaction in the temperature range of 27–90◦C at pH6.0.
Effect of sodium and calcium ions on α-amylase activityInfluence of sodium and calcium ions on the α-amylase ac-tivity was determined by pre-incubation of the enzyme withdifferent ions concentration at room temperature for 10 min.The controls were reaction mixture without ions. The ac-tivity of α-amylase was then determined using DNSA assay(Miller 1959) and Fuwa assay (Fuwa 1954) for the effect ofsodium and calcium ions, respectively.
Protein concentration, SDS-PAGE and zymography analysisProtein concentration was determined by the Bradfordmethod (Bradford 1976) using bovine serum albumin asa standard (Bio-Rad protein assay). SDS-PAGE was car-ried out according to Laemmli method (Laemmli 1970).The proteins were stained using Blue stain (BioRad). Gelstaining activity was performed by incubating the gel fromnative-PAGE or SDS-PAGE in solution containing 1% solu-ble starch in 50 mM universal buffer pH 6.0 at temperature50◦C for 30 min. The gel was then dipped in iodine solutionfor 10 min. The amylase activity was shown with the ap-pearance of clear zone in the gel (Lo et al. 2002). Pre-stainedprotein ladder (PageRulerTM, Fermentas) was used for theα-amylase molecular determination using SDS-PAGE.
Scanning electron microscopyA mixture of 5% (b/v) granular raw starch suspension andα-amylase (31.4 U) was incubated at 37◦C for 4 days. Themixture was centrifuged at 11,100 × g and the pellet waswashed three times with 95% ethanol followed by dryingat 35◦C for 18 h. Treated starch granules were attached toSEM holder and then coated with Au-Pd (80–20) using IonSputter JFC-110 at 1.2 KV and 6 mA for 4 min. Sampleswere inserted into specimen chamber SEM (JSM-35C) andthe pictures were taken.
Granular starch degradation by an Indonesian marine Bacillus α-amylase 1049
Table 1. Morphology and physiology of Bacillus sp. ALSHL3.
Character Bacillus sp. ALSHL3
Colony shape Circular-irregular, filamentous,umbonate, opaque
Cell shape Rods, spore formingGram stain +Motility +Nitrate reduction +Catalase production +H2S production −Indol production −Urease production −
Methyl red assay −Voges-Proskauer assay −TSI assay +Simmon’s sitrat assay −
Hydrolysis of starch +Hydrolysis of lipid +Hydrolysis of casein +Hydrolysis of gelatin +
Glucose fermentation +Sucrose fermentation +Lactose fermentation −
Mode of actionTwenty µL of α-amylase enzyme (31.8 U/mL) was addedto 100 µL soluble starch 1% (w/v) and incubated at 60◦C.Samples were removed at fixed time intervals. Products inthe reaction mixture were analyzed by thin layer chromatog-raphy with a stationary phase of silica gel and a solventsystem containing buthanol:ethanol:water (5:5:3). Starchproduct spots were visualized by spraying plates with 50%(v/v) H2SO4 in methanol and drying at 110◦C for 10 min.
Kinetic determinationInitial reaction rates using soluble starch as substrate weredetermined at substrate concentration of 5–17.5 mg/mL in50 mM universal buffer pH 6.0 at 60◦C. The kinetic con-stants, Km and Vmax, were estimated using linear regressionplots of Lineweaver and Burk.
Results and discussions
Identification of ALSHL3 isolate was determined ac-cording Bergey’s Manual of Determinative Bacteriol-ogy (Buchanan et al. 1974). Colony of the strain hasfilamentous, umbonate, opaque, and cilcular-irregularshape. The cell has stem form, producing endospore,and include to Gram-positive bacteria. Based on itsmorphology and physiology (Table 1), the ALSHL3isolate was identified as Bacillus filicolonicus. Molec-ular identification using 16s rDNA sequence compar-ison shows that the isolate ALSHL3 has 99% identitywith B. subtilis and B. amyloliquefaciens. Since the iso-late ALSHL3 has different morphology and physiologythan B. subtilis and B. amyloliquefaciens, it can be con-sidered as a different Bacillus species. To confirm thephylogeny of ALSHL3 isolate, further chemotaxonomicstudies have to be performed.
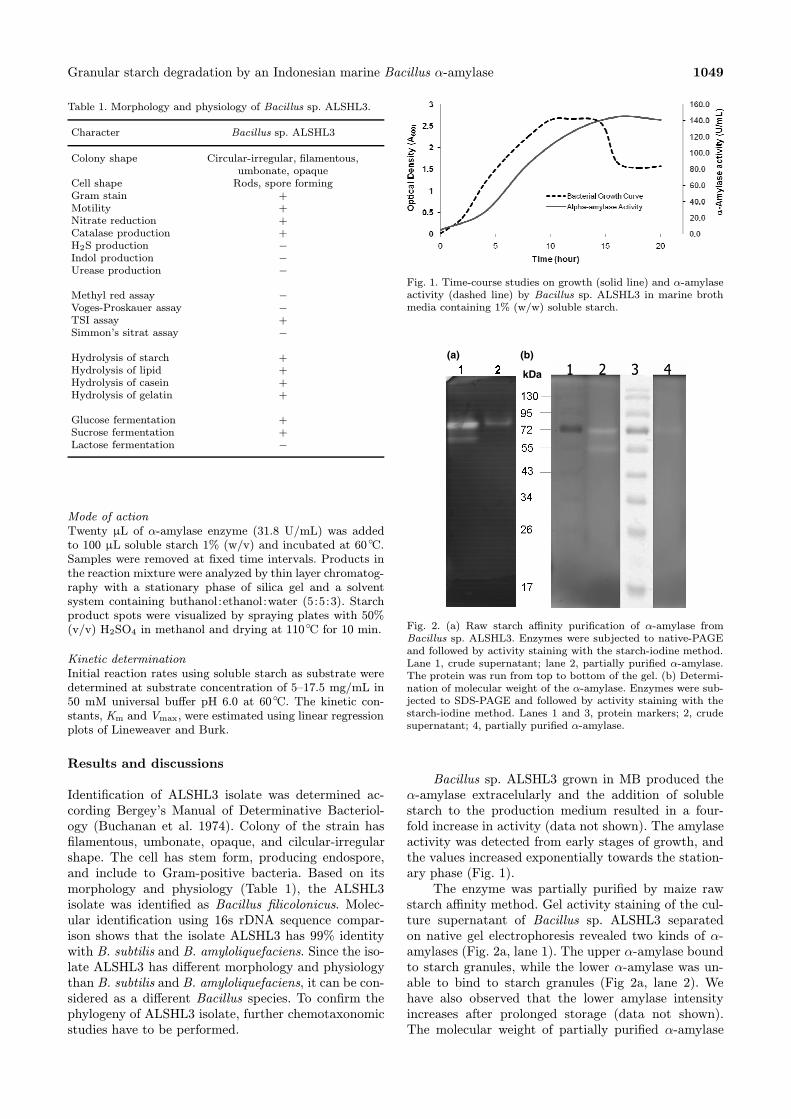
Fig. 1. Time-course studies on growth (solid line) and α-amylaseactivity (dashed line) by Bacillus sp. ALSHL3 in marine brothmedia containing 1% (w/w) soluble starch.
kDa
(a) (b)
Fig. 2. (a) Raw starch affinity purification of α-amylase fromBacillus sp. ALSHL3. Enzymes were subjected to native-PAGEand followed by activity staining with the starch-iodine method.Lane 1, crude supernatant; lane 2, partially purified α-amylase.The protein was run from top to bottom of the gel. (b) Determi-nation of molecular weight of the α-amylase. Enzymes were sub-jected to SDS-PAGE and followed by activity staining with thestarch-iodine method. Lanes 1 and 3, protein markers; 2, crudesupernatant; 4, partially purified α-amylase.
Bacillus sp. ALSHL3 grown in MB produced theα-amylase extracelularly and the addition of solublestarch to the production medium resulted in a four-fold increase in activity (data not shown). The amylaseactivity was detected from early stages of growth, andthe values increased exponentially towards the station-ary phase (Fig. 1).The enzyme was partially purified by maize raw
starch affinity method. Gel activity staining of the cul-ture supernatant of Bacillus sp. ALSHL3 separatedon native gel electrophoresis revealed two kinds of α-amylases (Fig. 2a, lane 1). The upper α-amylase boundto starch granules, while the lower α-amylase was un-able to bind to starch granules (Fig 2a, lane 2). Wehave also observed that the lower amylase intensityincreases after prolonged storage (data not shown).The molecular weight of partially purified α-amylase
1050 K. Vidilaseris et al.
(a) (b)
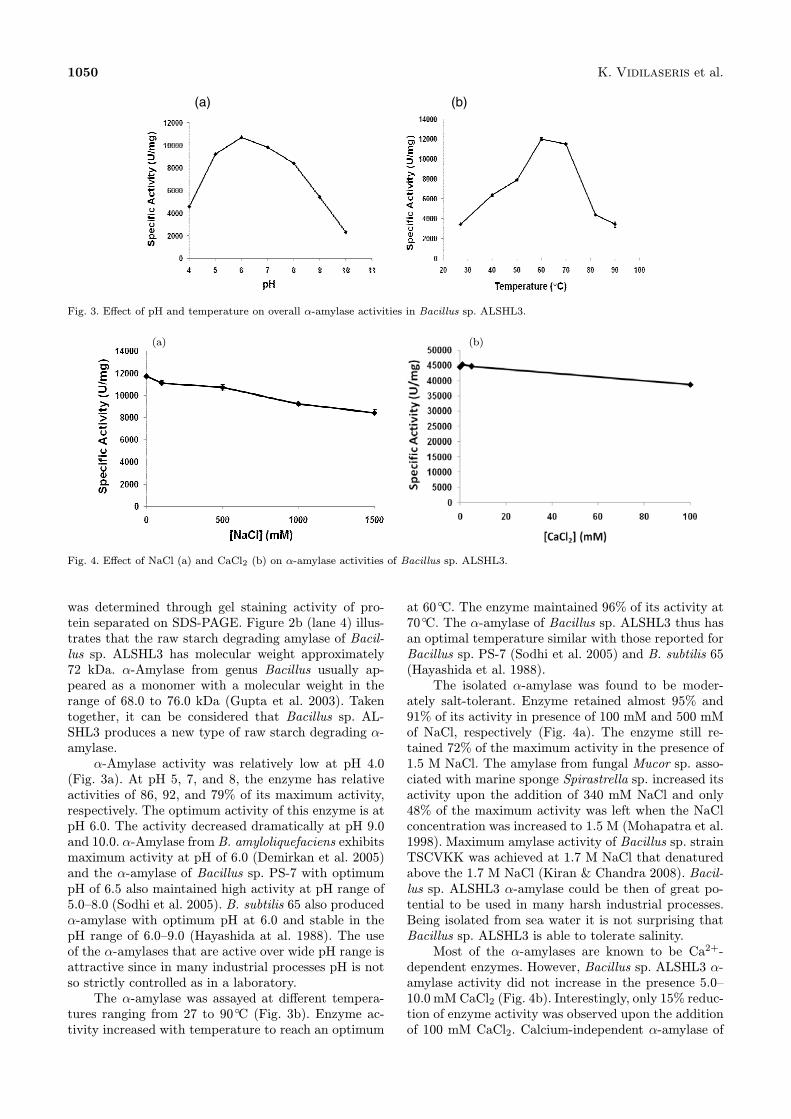
Fig. 3. Effect of pH and temperature on overall α-amylase activities in Bacillus sp. ALSHL3.
Fig. 4.
(a) (b)
Effect of NaCl (a) and CaCl2 (b) on α-amylase activities of Bacillus sp. ALSHL3.
was determined through gel staining activity of pro-tein separated on SDS-PAGE. Figure 2b (lane 4) illus-trates that the raw starch degrading amylase of Bacil-lus sp. ALSHL3 has molecular weight approximately72 kDa. α-Amylase from genus Bacillus usually ap-peared as a monomer with a molecular weight in therange of 68.0 to 76.0 kDa (Gupta et al. 2003). Takentogether, it can be considered that Bacillus sp. AL-SHL3 produces a new type of raw starch degrading α-amylase.
α-Amylase activity was relatively low at pH 4.0(Fig. 3a). At pH 5, 7, and 8, the enzyme has relativeactivities of 86, 92, and 79% of its maximum activity,respectively. The optimum activity of this enzyme is atpH 6.0. The activity decreased dramatically at pH 9.0and 10.0. α-Amylase from B. amyloliquefaciens exhibitsmaximum activity at pH of 6.0 (Demirkan et al. 2005)and the α-amylase of Bacillus sp. PS-7 with optimumpH of 6.5 also maintained high activity at pH range of5.0–8.0 (Sodhi et al. 2005). B. subtilis 65 also producedα-amylase with optimum pH at 6.0 and stable in thepH range of 6.0–9.0 (Hayashida at al. 1988). The useof the α-amylases that are active over wide pH range isattractive since in many industrial processes pH is notso strictly controlled as in a laboratory.The α-amylase was assayed at different tempera-
tures ranging from 27 to 90◦C (Fig. 3b). Enzyme ac-tivity increased with temperature to reach an optimum
at 60◦C. The enzyme maintained 96% of its activity at70◦C. The α-amylase of Bacillus sp. ALSHL3 thus hasan optimal temperature similar with those reported forBacillus sp. PS-7 (Sodhi et al. 2005) and B. subtilis 65(Hayashida et al. 1988).The isolated α-amylase was found to be moder-
ately salt-tolerant. Enzyme retained almost 95% and91% of its activity in presence of 100 mM and 500 mMof NaCl, respectively (Fig. 4a). The enzyme still re-tained 72% of the maximum activity in the presence of1.5 M NaCl. The amylase from fungal Mucor sp. asso-ciated with marine sponge Spirastrella sp. increased itsactivity upon the addition of 340 mM NaCl and only48% of the maximum activity was left when the NaClconcentration was increased to 1.5 M (Mohapatra et al.1998). Maximum amylase activity of Bacillus sp. strainTSCVKK was achieved at 1.7 M NaCl that denaturedabove the 1.7 M NaCl (Kiran & Chandra 2008). Bacil-lus sp. ALSHL3 α-amylase could be then of great po-tential to be used in many harsh industrial processes.Being isolated from sea water it is not surprising thatBacillus sp. ALSHL3 is able to tolerate salinity.Most of the α-amylases are known to be Ca2+-
dependent enzymes. However, Bacillus sp. ALSHL3 α-amylase activity did not increase in the presence 5.0–10.0 mM CaCl2 (Fig. 4b). Interestingly, only 15% reduc-tion of enzyme activity was observed upon the additionof 100 mM CaCl2. Calcium-independent α-amylase of
Granular starch degradation by an Indonesian marine Bacillus α-amylase 1051
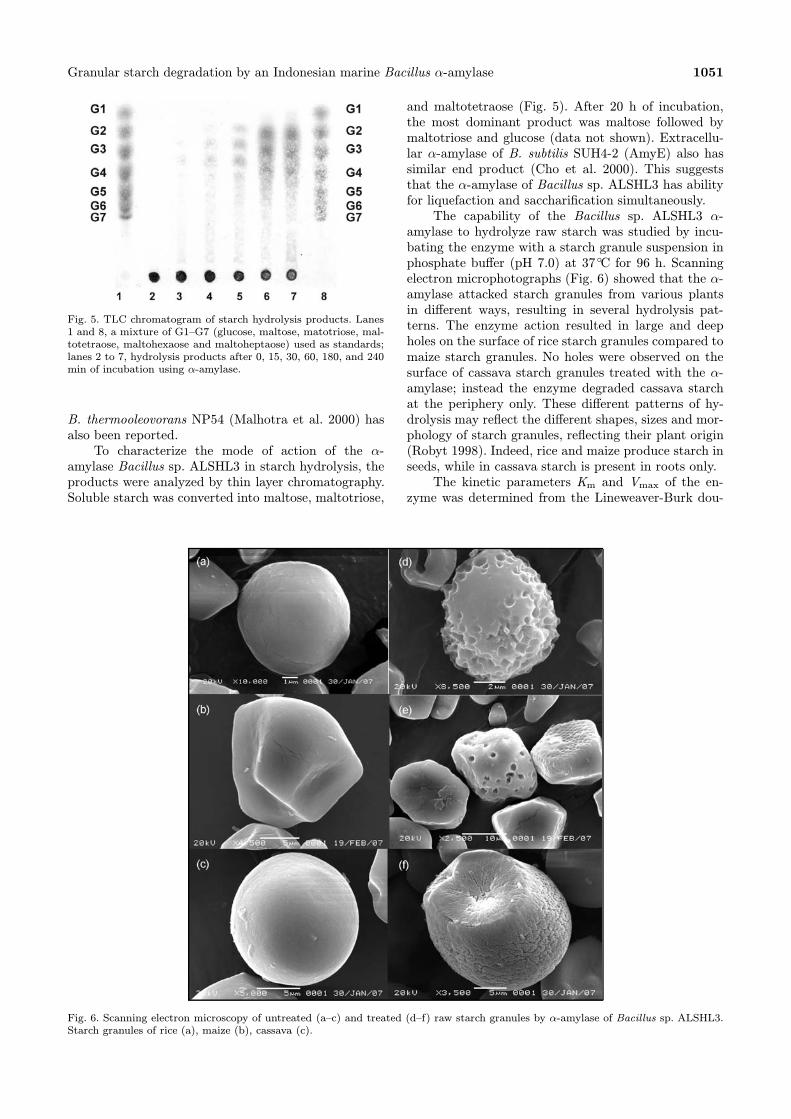
Fig. 5. TLC chromatogram of starch hydrolysis products. Lanes1 and 8, a mixture of G1–G7 (glucose, maltose, matotriose, mal-totetraose, maltohexaose and maltoheptaose) used as standards;lanes 2 to 7, hydrolysis products after 0, 15, 30, 60, 180, and 240min of incubation using α-amylase.
B. thermooleovorans NP54 (Malhotra et al. 2000) hasalso been reported.To characterize the mode of action of the α-
amylase Bacillus sp. ALSHL3 in starch hydrolysis, theproducts were analyzed by thin layer chromatography.Soluble starch was converted into maltose, maltotriose,
and maltotetraose (Fig. 5). After 20 h of incubation,the most dominant product was maltose followed bymaltotriose and glucose (data not shown). Extracellu-lar α-amylase of B. subtilis SUH4-2 (AmyE) also hassimilar end product (Cho et al. 2000). This suggeststhat the α-amylase of Bacillus sp. ALSHL3 has abilityfor liquefaction and saccharification simultaneously.The capability of the Bacillus sp. ALSHL3 α-
amylase to hydrolyze raw starch was studied by incu-bating the enzyme with a starch granule suspension inphosphate buffer (pH 7.0) at 37◦C for 96 h. Scanningelectron microphotographs (Fig. 6) showed that the α-amylase attacked starch granules from various plantsin different ways, resulting in several hydrolysis pat-terns. The enzyme action resulted in large and deepholes on the surface of rice starch granules compared tomaize starch granules. No holes were observed on thesurface of cassava starch granules treated with the α-amylase; instead the enzyme degraded cassava starchat the periphery only. These different patterns of hy-drolysis may reflect the different shapes, sizes and mor-phology of starch granules, reflecting their plant origin(Robyt 1998). Indeed, rice and maize produce starch inseeds, while in cassava starch is present in roots only.The kinetic parameters Km and Vmax of the en-
zyme was determined from the Lineweaver-Burk dou-
Fig. 6. Scanning electron microscopy of untreated (a–c) and treated (d–f) raw starch granules by α-amylase of Bacillus sp. ALSHL3.Starch granules of rice (a), maize (b), cassava (c).

1052 K. Vidilaseris et al.
Fig. 7. Double reciprocal plot for determining kinetic parametersof α-amylase of Bacillus. sp. ALSHL3.
ble reciprocal plots of the α-amylase activity at 60◦Cusing various concentration of soluble starch as sub-strate (Fig. 7). Apparent Km of the α-amylase fromBacillus sp. ALSHL3 was 4.79 mg/mL, Vmax was 0.322mM/min. The Km values obtained in the present studyare higher than those observed for B. amyloliquefaciens(1.92 mg/mL) (Demirkan et al. 2005), B. subtilis AX20(3.3 mg/mL) (Najafi et al. 2005) and B. licheniformisα-amylase (0.9 mg/mL) (Ivanova et al. 1993). There-fore, the α-amylase from Bacillus sp. ALSHL3 has lessaffinity for the soluble starch compared to other re-ported α-amylases.In conclusion, the novel raw starch degrading α-
amylase from marine Bacillus sp. ALSHL3 is a poten-tial candidate for application in the starch processingindustry and other biotechnological processes since itis thermotolerant, has the ability to act at a broad pHrange and tolerates high salinity. In the future it will beessential to perform further molecular characterizationin terms of both the gene and the protein to fully un-derstand the nature of the α-amylase from the marineBacillus sp. ALSHL3.
Acknowledgements
We thank Riset Unggulan ITB 2005 and International Re-search Grant ITB 2007 for funding this work.
References
Abe A., Tonozuka T., Sakano Y. & Kamitori S. 2004. Complexstructures of Thermoactinomyces vulgaris R-47 α-amylase 1with malto-oligosaccharides demonstrate the role of domainN acting as a starch-binding domain. J. Mol. Biol. 335: 811–822.
Asgher M., Asad M.J., Rahman S.U. & Legge R.L. 2007. A ther-mostable α-amylase from a moderately thermophilic Bacillussubtilis strain for starch processing. J. Food Eng. 79: 950–955.
Bradford M.M. 1976. A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the prin-ciple of protein-dye-binding. Anal. Biochem. 72: 248–254.
Buchanan R.E., Gibbons N.E., Cowan S.T., Holt J.G., Liston J.,Muray R.G.E., Niven C.F., Ravin A.W. & Stanier R.W. 1974.Bergey’s Manual of Determinative Bacteriology, 8th Edition.The Williams and Wilkins Company, Baltimore.
Cho H.Y., Kim Y.W., Kim T.J., Lee H.S., Kim D.Y., Kim J.W.,Lee Y.W., Lee S.B. & Park K.H. 2000. Molecular characteri-zation of a dimeric intracellular maltogenic amylase of Bacil-lus subtilis SUH4-2. Biochim. Biophys. Acta. 1478: 333–340.
Demirkan E.S., Mikami B., Adachi M., Higasa T., Utsumi S.2005. α-Amylase from B. amyloliquefaciens: purification,characterization, raw starch degradation and expression inE. coli. Process Biochem. 40: 2629–2636.
Fuwa H. 1954. A new method for microdetermination of amylaseactivity by the use of amylose as a substrate. J. Biochem. 21:219–230.
Gupta R., Gigras P., Mohapatra H., Goswami V.K. & ChauhanB. 2003. Microbial α-amylases: a biotechnological perspec-tive. Process Biochem. 38: 1599–1616.
Hamilton L.M., Kelly C.T. & Fogarty W.M. 1998. Raw starchdegradation by the non-raw starch-adsorbing bacterial α-amylase of Bacillus sp. IMD 434. Carbohydr. Res. 314: 251–257.
Hayashida S., Teramoto Y. & Inoue T. 1988. Production andcharacteristics of raw potato starch digesting amylase fromBacillus subtilis 65. Appl. Environ. Microbiol. 54: 1516–1522.
Ivanova V., Dobreva E. & Emanuilova E. 1993. Purificationand characterization of thermostable α-amylase from Bacilluslicheniformis. J. Biotechnol. 28: 277–289.
Janecek S. & Sevcik J. 1999. The evolution of starch-bindingdomain. FEBS Lett. 456: 119–125.
Kiran K.K. & Chandra T. S. 2008. Production of surfactant anddetergent-stable, halophilic, and alkalitolerant α-amylase bya moderately halophilic Bacillus sp. strain TSCVKK. Appl.Microbiol. Biotechnol. 77: 1023–1031.
Laemmli U.K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriphage T4. Nature227: 680–685.
Lo H.F., Lin L.L., Chiang W.Y., Chie M.C., Hsu W.H. & ChangC.T. 2002. Deletion analysis of the C-terminal region of theα-amylase of Bacillus sp. strain TS-23. Arch. Microbiol. 178:115–123.
MacGregor E.A., Janecek S. & Svensson B. 2001. Relationship ofsequence and structure to specificity in the α-amylase familyof enzymes. Biochim. Biophys. Acta 1546: 1–20.
Machovic M. & Janecek S. 2006. The evolution of putative starchbinding domains. FEBS Lett. 580: 6349–6356.
Malhotra R., Noorvez S.M. & Satyanarayana T. 2000. Produc-tion and partial characterization of thermostable and calciumindependent α-amylase of an extreme thermophile Bacillusthermooleovorans NP54. Lett. Appl. Microbiol. 31: 378–384.
Miller G.L. 1959. Use of dinitrosalicylic acid reagent for determi-nation of reducing sugar. Anal. Chem. 31: 426 428.
Mohapatra B.R., Banerjee U.C. & Bapuji M. 1998. Characteriza-tion of a fungal amylase from Mucor sp. associated with themarine sponge Spirastrella sp. J. Biotechnol. 60: 113–117.
Najafi M.F., Deobagkar D. & Deobagkar D. 2005. Purifica-tion and characterization of an extracellular α-amylase fromBacillus subtilis AX20. Protein Expr. Purif. 41: 349–354.
Najafi M.F. & Kembhavi A. 2005. One step purification and char-acterization of an extracellular amylase from marine Vibriosp. Enzyme Microb. Technol. 36: 535–539.
Robyt J.F. 1998. Essentials of Carbohydrate Chemistry. Springer,Boston, 399 pp.
Sodhi H.K., Sharma K., Gupta J.K. & Soni S.K. 2005. Productionof a thermostable amylase by solid-state fermentation and itssynergistic use in the hydrolysis of malt strarch for alcoholproduction. Process Biochem. 40: 525–534.
Van der Maarel M.J.E.C., van der Veen B., Uitdehaag J.C.M.,Leemhuis H. & Dijkhuizen L. 2002. Properties and applica-tion of starch converting enzymes of the amylase family. J.Biotechnol. 94: 137–155.
Received November 19, 2008Accepted July 18, 2009
Related Documents











