Biochemical Characterization of a Novel Whey Protein from Murine Milk* (Received for publication, January 19, 1981, and in revised form, July 24, 1981) John E. PiletzS, Michael Heinlen, and Roger E. Ganschowg From the Children's Hospital Research Foundation, Cincinnati, Ohio 45229 and the Department of Pediatrics of the University of Cincinnati College of Medicine, Cincinnati, Ohio 45229 An electrophoretic variant of the major whey protein of murine milk has been uncovered in the YBR strain of mice. Both normal and variant forms of this protein, designated whey acidic protein (WAP), constitute a minimum of 2.4% of total mouse milk protein, display an acidic isoelectric point, exhibit a molecular weight of 14,000 after both denaturing and nondenaturing gel electrophoresis, and lack tyrosine and histidineas de- termined by amino acid analysis. Combined isotope incorporation and immunochemical studies show that WAP is synthesized by the mammary gland and not by the liver. WAP appears to be unique to mouse milk since no milk protein of similar properties has been described in other species and since goat anti-mouse- WAP antiserum does not cross-react with components of bovine, human, or rat milk. However, WAP does exhibit a structural similarity to certain apo-C lipopro- teins of human serum very light density lipoproteins. Amino acid and tryptic peptide analyses suggest that the variant form of WAP (WAP-B) contains one more arginine and one less cysteine than normal WAP (WAP- A). Genetic studies to be reported elsewhere show this difference to be under the control of a single Mendelian autosomal gene with alleles expressed in a co-dominant manner. The designation Wap is proposed for this gene with WapA and WapB as thewildtypeandmutant alleles, respectively. In contrast to the vast literature concerning the structure and function of milk proteins (1, 2), relatively little is known about the genetic control of these proteins because species in which milk protein variants have been identified, namely bovine and human (31, are not easily manipulated for genetic studies. In order to develop a system more suitable for genetic analysis, we have surveyed 58 inbred strains of mice for electrophoretic variations among milk proteins. Normal and variant forms of two milk proteins in inbred strains of mice have been identified and will be reported elsewhere. The biochemical characterization of one of these proteins, the major whey protein of murine milk, is reported here. We have investigated the composition, immunological character, and tissue site of synthesis of this previously undescribed protein. The results indicate no recognizablesimilarities to anymajor * This work was supported by Grants AM 14770 and HD 05221 from the National Institues of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accord- ance with 18 U.S.C. Section 1734 solely to indicate this fact. f Present address, Department of Nuclear Medicine and Radiation Biology, University of California at Los Angeles, Los Angeles, CA 90024. 8 To whom correspondence should be addressed. milk protein described in other species. We propose the name whey acidic protein for this novel protein, based on the fraction of milk from which it was derived and upon its acidic isoelectric point. Comparisons of wild type (WAP-A) and variant (WAP-B) forms of WAP' suggest that the electropho- retic mobility difference is controlled by the WAP structural gene which we designate as Wap, with the alleles WapAand WapBspecifying the two phenotypes, respectively. MATERIALS AND METHODS Animals YBR mice were obtained from Roswell Park Memorial Institute, Buffalo, NY and from the Kirschbaum Memorial Mouse Colony, North Eastern Ohio University, Kent, OH. Mice of the C57B1/6J strain were obtained from The Jackson Laboratory, Bar Harbor, ME. AU animals were maintained on a dietof Purina laboratorychow and water a d libitum. Milkwas collected according to the method of McBurney et al. (4) between 5 and 20 days postpartum. Purification of the Variant WheySrotein(WAP) Step 1: Fractionation of Milk into Curds and Whey-Fresh milk was pooled from several mice, diluted 50-fold with double-distilled water, and skimmed at 0 "C. Casein curds were precipitated by adjusting the pH to 4.6 with 0.1 M HCl and chilling for 10 rnin a t 0 "C. Precipitated casein was collected by centrifugation at 1,600 X g for 15 min at 4 "C. The whey was decanted and saved. Two additional washes of the casein pellet were performed with 30 original milk volumes of 0.1 M sodium acetate buffer, pH 4.6. The supernatants from these washes were discarded. The casein pellet was dissolvedin double-distilled water andadjusted to pH 6.2 with 0.1 M sodium hydroxide. The caseins were not purified further. Step 2: Ammonium Sulfate Fractionation-Residual casein in the whey fraction of milk was removed by sedimentation at 13,000 X g for 15 min at 4 "C. Solid ammonium sulfate was added to the resulting supernatant to a r i a l concentration of 33%; the mixture was incubated at 4 "C for 30 min and centrifuged at 2,000 X g for 15 min. The pellet was redissolved in water and used for additional purification of WAP. The ammonium sulfate concentration of the supernatant was adjusted to 80% and the resulting precipitate was collected at 13,000 X g for 30 min. The whey proteins precipitated at 80% ammonium sulfate were redissolved in water. Step 3: Gel Filtration Chromatography-The 33% ammonium sulfate precipitate from Step 2 was redissolved in water and further fractionated by gel filtration at 4 "C on a column (45 X 1.0 cm) of Bio-Gel P-60, using 75 mM ammonium bicarbonate for equilibration of the matrix and elution of the sample. Fractions (0.5 ml) were collected and monitored for absorbance at 280 nm on a Gilford Model 2400 spectrophotometer. The low molecular weight peak was pooled and lyophilized. Step 4: DEAE-cellulose Chromatography-The low molecular weight whey proteins were dialyzed against 500 volumes of 5 mM Tris-HC1, pH 8.6, at 4 "C for 8 h and applied to a DEAE-cellulose column (Whatman DE52) (8.0 X 0.8 cm) previously equilibrated with ' The abbreviations used are: WAP,whey acidic protein; SDS, sodium dodecyl sulfate. 11509

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochemical Characterization of a Novel Whey Protein from Murine Milk*
(Received for publication, January 19, 1981, and in revised form, July 24, 1981)
John E. PiletzS, Michael Heinlen, and Roger E. Ganschowg From the Children's Hospital Research Foundation, Cincinnati, Ohio 45229 and the Department of Pediatrics of the University of Cincinnati College of Medicine, Cincinnati, Ohio 45229
An electrophoretic variant of the major whey protein of murine milk has been uncovered in the YBR strain of mice. Both normal and variant forms of this protein, designated whey acidic protein (WAP), constitute a minimum of 2.4% of total mouse milk protein, display an acidic isoelectric point, exhibit a molecular weight of 14,000 after both denaturing and nondenaturing gel electrophoresis, and lack tyrosine and histidine as de- termined by amino acid analysis. Combined isotope incorporation and immunochemical studies show that WAP i s synthesized by the mammary gland and not by the liver. WAP appears to be unique to mouse milk since no milk protein of similar properties has been described in other species and since goat anti-mouse- WAP antiserum does not cross-react with components of bovine, human, or rat milk. However, WAP does exhibit a structural similarity to certain apo-C lipopro- teins of human serum very light density lipoproteins.
Amino acid and tryptic peptide analyses suggest that the variant form of WAP (WAP-B) contains one more arginine and one less cysteine than normal WAP (WAP- A). Genetic studies to be reported elsewhere show this difference to be under the control of a single Mendelian autosomal gene with alleles expressed in a co-dominant manner. The designation Wap is proposed for this gene with WapA and WapB as the wild type and mutant alleles, respectively.
In contrast to the vast literature concerning the structure and function of milk proteins (1, 2), relatively little is known about the genetic control of these proteins because species in which milk protein variants have been identified, namely bovine and human (31, are not easily manipulated for genetic studies. In order to develop a system more suitable for genetic analysis, we have surveyed 58 inbred strains of mice for electrophoretic variations among milk proteins. Normal and variant forms of two milk proteins in inbred strains of mice have been identified and will be reported elsewhere. The biochemical characterization of one of these proteins, the major whey protein of murine milk, is reported here. We have investigated the composition, immunological character, and tissue site of synthesis of this previously undescribed protein. The results indicate no recognizable similarities to any major
* This work was supported by Grants AM 14770 and HD 05221 from the National Institues of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accord- ance with 18 U.S.C. Section 1734 solely to indicate this fact.
f Present address, Department of Nuclear Medicine and Radiation Biology, University of California at Los Angeles, Los Angeles, CA 90024.
8 To whom correspondence should be addressed.
milk protein described in other species. We propose the name whey acidic protein for this novel protein, based on the fraction of milk from which it was derived and upon its acidic isoelectric point. Comparisons of wild type (WAP-A) and variant (WAP-B) forms of WAP' suggest that the electropho- retic mobility difference is controlled by the WAP structural gene which we designate as Wap, with the alleles WapA and WapB specifying the two phenotypes, respectively.
MATERIALS AND METHODS
Animals YBR mice were obtained from Roswell Park Memorial Institute,
Buffalo, NY and from the Kirschbaum Memorial Mouse Colony, North Eastern Ohio University, Kent, OH. Mice of the C57B1/6J strain were obtained from The Jackson Laboratory, Bar Harbor, ME. AU animals were maintained on a diet of Purina laboratory chow and water a d libitum. Milk was collected according to the method of McBurney et al. (4) between 5 and 20 days postpartum.
Purification of the Variant WheySrotein (WAP) Step 1: Fractionation of Milk into Curds and Whey-Fresh milk
was pooled from several mice, diluted 50-fold with double-distilled water, and skimmed at 0 "C. Casein curds were precipitated by adjusting the pH to 4.6 with 0.1 M HCl and chilling for 10 rnin at 0 "C. Precipitated casein was collected by centrifugation at 1,600 X g for 15 min at 4 "C. The whey was decanted and saved. Two additional washes of the casein pellet were performed with 30 original milk volumes of 0.1 M sodium acetate buffer, pH 4.6. The supernatants from these washes were discarded. The casein pellet was dissolved in double-distilled water and adjusted to pH 6.2 with 0.1 M sodium hydroxide. The caseins were not purified further.
Step 2: Ammonium Sulfate Fractionation-Residual casein in the whey fraction of milk was removed by sedimentation at 13,000 X g for 15 min at 4 "C. Solid ammonium sulfate was added to the resulting supernatant to a r i a l concentration of 33%; the mixture was incubated at 4 "C for 30 min and centrifuged at 2,000 X g for 15 min. The pellet was redissolved in water and used for additional purification of WAP. The ammonium sulfate concentration of the supernatant was adjusted to 80% and the resulting precipitate was collected at 13,000 X g for 30 min. The whey proteins precipitated at 80% ammonium sulfate were redissolved in water.
Step 3: Gel Filtration Chromatography-The 33% ammonium sulfate precipitate from Step 2 was redissolved in water and further fractionated by gel filtration at 4 "C on a column (45 X 1.0 cm) of Bio-Gel P-60, using 75 mM ammonium bicarbonate for equilibration of the matrix and elution of the sample. Fractions (0.5 ml) were collected and monitored for absorbance at 280 nm on a Gilford Model 2400 spectrophotometer. The low molecular weight peak was pooled and lyophilized.
Step 4: DEAE-cellulose Chromatography-The low molecular weight whey proteins were dialyzed against 500 volumes of 5 mM Tris-HC1, pH 8.6, at 4 "C for 8 h and applied to a DEAE-cellulose column (Whatman DE52) (8.0 X 0.8 cm) previously equilibrated with
' The abbreviations used are: WAP, whey acidic protein; SDS, sodium dodecyl sulfate.
11509

11510 A Novel Whey Prote
dialysis buffer. Two bed volumes each of 0.1 M, 0.2 M, and 0.3 M and 0.4 M and 0.5 M Tris buffer, pH 8.6, were passed through the column and 1-ml fractions were collected. Fractions were monitored for absorbance at 280 nm.
Isoelectric Focusing Electrofocusing was carried out in a column electrophoresis appa-
ratus (LKB Instrument Co.) using a 40% (v/v) Ampholine solution, pH 2.5-10 (LKB). Samples containing approximately 100 pg of protein were delipidated with glass-distilled tetramethylurea (5) and applied to the top of the prefocused gels according to the method of Albers and Scanu (6). Electrofocusing was performed at 400 V for 4 h. After focusing, the pH gradient in the gel was measured with a micro pH electrode (Microelectrodes Co.) before staining the focused gel with Coomassie blue R.
Molecular Weight Estimation The gel electrophoresis method of Laemmli (7) was used for
determining the molecular weight of denatured WAP. The slab gel contained 12.5% acrylamide and 0.32% bisacrylamide. Standard pro- teins were cytochrome c from horse heart (Sigma), soybean trypsin inhibitor (Sigma), carbonic anhydrase from bovine erythrocytes (Cal- biochem), DNase I from bovine pancreas (Worthington), beef liver alcohol dehydrogenase (Worthington), egg albumin grade I11 (Sigma), mouse serum albumin fraction V (Miles), and bovine a-lactalbumin (Sigma).
The molecular weight of undenatured WAP was determined in two ways. First, samples were chromatographed on a Bio-Gel P-60 gel filtration column (100 X 1.5 cm) at 4 "C as described above in Step 3 of the purification procedure. Fractions were monitored for absor- bance at 280 nm and examined electrophoretically on nondenaturing gels (8). The second method employed polyacrylamide gels of varying acrylamide concentrations according to the method of Hedrick and Smith (9). The electrophoretic conditions were those of Davis (10) with the following modifications. Slab gels of varying monomer con- centrations were made by diluting a stock solution of 40% (w/v) acrylamide and 1% (w/v) methylenebisacrylamide. After staining with Coomassie blue, mobilities of proteins were measured relative to the dye marker bromphenol blue.
Amino Acid Analysis Samples of purified WAP were extensively dialyzed against five 2-
liter washes of double-distilled water at 4 "C for 48 h prior to amino acid analysis. The postdialysis purity of each sample was determined electrophoretically. Samples were acid-hydrolyzed, evaporated to dry- ness, diluted, and analyzed for amino acids in a Durmm D-500 analyzer.
Carbohydrate Analysis The presence of carbohydrates associated with WAP was analyzed
by postelectrophoretic staining with periodic acid-Schiff stain (11) and with the cationic carbocyanine dye l-ethyl-2-[3-(l-ethylnaptho- [1,2d]-thiazolin-2-ylidene)-2-methylpropenyl]-naptho[l,3d]thi~o- lium bromide, known as "Stains-all" (Eastman) (12). Bovine casein and a-lactalbumin (Sigma), calf fetuin (gift of Dr. E. F. Zimmerman, Children's Hospital Research Foundation, Cincinnati), and murine serum albumin (Miles) were used as controls. The casein sample was dissolved in 7 M urea prior to electrophoresis.
Neuraminidase treatment of WAP was carried out in 200 pl of 0.1 M sodium acetate buffer (pH 5.0) with Clostridium perfringens Type VI neuraminidase (Sigma) for 120 min at 37 "C. Calf fetuin and a- fetoprotein served as controls. After treatment, 100-pg samples were electrophoresed at pH 9.5 on polyacrylamide slab gels containing 7 M urea (8) and stained with Coomassie blue.
Phosphate Analysis The presence of phosphate in WAP was analyzed after electropho-
resis by staining with either methyl green (13) or with "Stains-all'' (12). Bovine casein, a-lactalbumin, and murine serum albumin served as controls for the methyl green stain experiment.
Phosphatase digestion was performed on 200 pg of WAP incubated in 200 pl of 50 mM Tris buffer, pH 5.5, using 7 units of highly purified potato acid phosphatase (Sigma) for 12 h at 37 " c (14). similar amounts of murine curd protein, bovine a-lactalbumin, and bovine casein were digested as controls. After enzyme treatment, 150-pg samples were subjected to electrophoresis at pH 3.7 in 8 M urea (15).
i n from Murine Milk
Similar experiments were performed with calf intestinal alkaline phosphatase (Sigma).
Lipid Analysis The presence of lipids associated with WAP was determined by
staining on agarose gels with Sudan Black B (Sigma) (16). Approxi- mately 100 pg of WAP and 5-20 pg of human serum low density lipoprotein (courtesy of Dr. E. Gruenstein, University of Cincinnati) were electrophoresed. The agarose gel electrophoresis system of Nobel (16) was modified for use on the vertical slab gel apparatus (Model 220, Bio-Rad). A 3% acrylamide stacking gel (10) was layered atop the agarose gel and used to form the sample wells. Electropho- resis was performed using 50 mM barbital running buffer, pH 8.6, for 90 min at 100 mA with tap water cooling. The gels were stained with Sudan Black B as described by Noble (16).
Lipid extraction was performed on 200-pg samples of WAP, bovine a-lactalbumin, and human serum low density lipoprotein by the method of Folch et al. (17) or that of Kane et al. (5). After electro- phoresis on 10% nondenaturing polyacrylamide gels (lo), the delipi- dated proteins were stained with Coomassie brilliant blue and im- mediately photographed.
a-Lactalbumin Activity A modification of the method of Brew et al. (18) was used to
measure a-lactalbumin specifier protein activity for lactose synthetase in fractions of mouse milk. Reaction tubes contained 30 p1 of 0.5 M Tris-HC1, pH 7.4, 60 pl of 0.15 M glucose, 30 p1 of 0.4 M MnClz, 94 pl of 2 mM UDP-galactose, 30 pl of ['4C]UDP-galactose ( 5 0 nCi), 30 pl ( 8 4 microunits) of bovine galactosyltransferase (UDP-galactose:D-gh- cose-4-P-galactosyltransferase; EC 2.4.1.22) (Sigma), and varying amounts of milk sample to a final volume of 0.3 ml. Samples were passed over AG 1-X8 anion exchange columns (Bio-Rad) (4 X 0.7 cm) in the chloride form and eluted with water. Fractions were collected from each assay and dissolved in Triton-based scintillation fluor and ['4C]lactose radioactivity was determined by liquid scintillation count- ing.
For gel elution assays, gel slices were transferred to assay tubes containing reaction buffer minus MnClZ, ['4C]UDP-galactose, and galactosyltransferase. In order to prevent the manganese salt from precipitating in the presence of 7 M urea, the pH was lowered to 7.5 by addition of 25 pl of 0.5 M HCl before adding MnC12. Bovine a- lactalbumin activity was shown to be unchanged in 7 M urea. Protein was eluted from the gels by mincing the slices and allowing them to stand overnight a t room temperature. Lactose synthetase activity in each minced gel slice was then assayed. Bovine a-lactalbumin was co- electrophoresed (8) in the same gels and assayed.
WAP Antiserum Purified WAP from C57B1/6J mice was emulsified in complete
Freund's adjuvant and injected subcutaneously into a goat. One week later, an equal amount of emulsion was administered. Serum was collected between the 2nd and 3rd weeks and decomplemented. The immunospecificity of the goat serum to WAP was determined by Ouchterlony double diffusion analysis against various fractions of milk.
Synthesis of WAP in Explant Culture Mammary glands and liver were removed from a C57B1/6J female
mouse at midlactation and sliced with scissors. Four 200-mg slices of each tissue were mixed with 0.8 ml of incubation buffer containing 50 pCi of ~-[4,5-~H]leucine (Amersham) and allowed to incubate for 1 h at 37 "C according to the method of Brew and Campbell (19). In other experiments, ~-[3,4-~H]proline, sodium dihydrogen [32P]orth~ph~s- phate (Amersham), or ~-[U-'~C]tyrosin (New England Nuclear) re- placed ~-[4,5-~H]leucine. Tissues were homogenized using a Polytron tissue homogenizer (Brinkmann) at medium speed for 90 s/sample. A 5-pl aliquot of each homogenate was precipitated with 30% trichlo- roacetic acid, the precipitates were collected on glass fiber filters (Gelman), and the radioactivity was determined. The remainder of each homogenate was then mixed with 200 pg of C57B1/6J whey protein, adjusted to pH 4.6 with 1.0 N HCl, and clarified by filtration. The pH of the filtrate was readjusted to pH 8.0 with 1.0 N NaOH and the resulting precipitate was removed by centrifugation at 1600 X g for 15 min at 4 "C. Each supernatant was refiltered and mixed with 1.5 ml of anti-WAP antiserum. Duplicate samples were also mixed with 0.5 ml of rabbit anti-human serum albumin antiserum. Such

A Novel Whey Protein from Murine Milk
treatment resulted in complete precipitation of both WAP and serum albumin from mouse milk whey after incubation at room temperature for 1 h and then at 4 “C overnight. Samples were made 8% in polyethylene glycol (6,000 daltons, General Biochemical) and precip- itation was allowed to continue at 4 “C for 20 additional h. Immune complexes were collected by centrifugation for 5 min in a Beckman Microfuge B and washed twice with a buffer containing 2% Triton X- 100, 10 mM leucine, 10 mM NaZHPOd, and 0.14 M NaCl, pH 7.5. The immune precipitates and supernatants were analyzed for the presence of radioactivity in WAP by SDS-gel electrophoresis (7). Radioactivity in gel slices was determined by liquid scintillation spectrometry.
Tryptic Peptide Mapping Samples were delipidated and denatured prior to trypsin digestion.
Lipids were extracted as described above. The methanol phase was dried under nitrogen and the apoprotein was dissolved in 1 ml of a solution containing 0.3 M Tris, 6.0 M guanidine HCl (Sigma), and 0.2% EDTA (Schwarz/Mann), pH 8.5. Each sample was then reduced with 10 pl of P-mercaptoethanol (Eastman) under nitrogen ( 9 0 min at 56 “C) and then alkylated by addition of 0.16 ml of 1 M iodoacetic acid (Fisher) in 1 M NaOH. Alkylation proceeded for 30 min at 37 “C under nitrogen and the reaction was stopped by addition of 5 pl of /3-mercaptoethanol. The alkylated WAP samples were dialyzed against two 1-liter washes of deionized water for 10 h at 4 “C. Lowry protein analysis (20) was used to determine the amount of sample recovered from these steps. The dialyzed samples were lyophilized and reconstituted with water. Such samples were then heated for 5 min at 90 “C, cooled, and made 0.2 M in ammonium acetate, pH 8.5. Trypsin-tosylphenylalanyl chloromethyl ketone-treated (Worthing- ton) was added to the prepared samples in two portions of 2 pg of trypsin per 100 pg of WAP. Samples were incubated at 37 “C for 4 h, mixed with the second portion, and incubated for another 16 h. Ammonium acetate was removed by repeated lyophilization and the digests were reconstituted to 5 m g / d in deionized water. The trypsin digests were analyzed by two-dimensional thin layer chromatography- electrophoresis on cellulose Chromogram sheets (No. 13255, East- man) as described by Stephens (21). Fluorescamine (Roche) or nin- hydrin (Sigma) were used as stains.
RESULTS
In a survey of 58 inbred strains of mice for milk protein variants, milk of the YBR strain was found to contain a fast mobility form of a major protein when its protein components were separated by polyacrylamide gel electrophoresis at pH 3.7 (Fig. lA ). This protein was observed as a slow mobility form when the same milk proteins were electrophoretically resolved on polyacrylamide gels at pH 8.5 (as in Fig. 1B). Based upon its properties which we describe in the following studies, we call this component of mouse milk “whey acidic protein” with the acronym WAP. Only mice of the YBR strain exhibit the variant form of WAP (WAP-B). We have utilized milk from the C57BL/6J strain as the source of wild type WAP (WAP-A) in the following studies of the biochemical properties of this milk protein and its variant form.
Purification of WAP-Table I summarizes the results of purification of whey acidic protein. Both WAP-A and WAP-B purified identically. In each step of the purification, fractions were monitored electrophoretically for the presence and pu- rity of WAP. On non-SDS gels, both the variant and normal forms of WAP are easily distinguished from other proteins by their unique staining property with Coomassie blue, appearing royal blue by transmitted light and pinkish when viewed by reflected light. Extensive purification of whey acidic protein is achieved through a 33% ammonium sulfate fractionation of whey in which WAP is found to precipitate while most of the other whey proteins remain soluble. The major contaminant of WAP after Step 2 (Fig. lA) co-migrates on gels with authentic mouse serum albumin and is found in the 33-808 ammonium sulfate cut of whey.
WAP was further purified from the 33% ammonium sulfate fraction of whey by gel filtration chromatography and then by anion exchange chromatography. WAP-A and WAP-B
A 1 2 3
t”
C- A- B-
4
11511
B 1 2
B- A.
FIG. 1. Protein staining patterns of milk and milk fractions from mice of the CS7B1/6J and YBR strains after gel electro- phoresis. A, 80 pg of skimmed milk protein from C57B1/6J mice (Column I ) and from YBR mice (Column 2 ) , and 40 pg of protein from the 3040% ammonium sulfate cut of C57B1/6J (Column 3) and YBR milks (Column 4 ) were subjected to gel electrophoresis a t pH 3.7 and then stained as described under “Materials and Methods.” B, 35 pg of purified whey acidic protein after the DEAE-cellulose chro- matography step was subjected to gel electrophoresis a t pH 9.5 and stained as described under “Materials and Methods.” Column I represents WAP from C57B1/6J milk (WAP-A) and Column 2 that of YBR milk (WAP-B). The protein bands designated A represent WAP-A, those designated B represent WAP-B, and those designated C represent murine serum albumin.
TABLE I purification of whey acidic protein Fractions Volume Total protein
ml w I. skim milk 1.6 159.7
11. Whey separation 64.1 46.3 111.33% (NH4)804 precipitation 14.0 9.8 IV. Gel filtration 8.4 8.9 V. DEAE-cellulose chromatography 14.0 6.1
were found in a major low molecular weight peak. Following gel filtration on Bio-Gel P-60, the sem albumin-like contam- inant of the 33% ammonium sulfate fraction was excluded by the gel. The WAP-containing fraction was further purified by passage over DEAE-cellulose. The majority of WAP eluted from DEAE-cellulose with 0.1 M NaC1. WAP-B was distin- guished from WAP-A on DEAE-cellulose by a slightly re- tarded elution during the 0.1 M NaCl wash. Following elution from DEAE-cellulose, WAP-A and WAP-B appeared to be free of contaminating proteins (Fig. 1B).
Based on Lowry analysis (20) of the amount of whey acidic protein obtained after DEAE-cellulose chromatography (3.4 mg/ml of milk) and knowing that whole milk contains ap- proximately 140 mg of protein/ml, WAP represents a mini- mum of 2.4% of total milk protein (Table I). The actual percentage is somewhat higher since WAP was observed in fractions routinely discarded during the purification, espe- cially in the curd fraction due to trapping during curdling. No assessment of the yield of WAP was made due to the inability to precisely quantitate WAP in whole milk.
Isoelectric Focusing-Isoelectric focusing was performed on the 33% ammonium sulfate cut of whey. As in electropho- retic analyses, WAP is distinguished from other proteins by its unique pink color when stained with Coomassie blue. WAP-A and WAP-B exhibit a small, but reproducible, Mer- ence in isoelectric point; 4.7 for WAP-A and 4.8 for WAP-B.

11512 A Novel Whey Protein from Murine Milk
Molecular Weight Estimations-Molecular weight meas- urements of WAP were made with preparations partially purified through the gel filtration chromatography step of Table I. SDS-gel electrophoresis was used to estimate the molecular weight of denatured WAP-A and WAP-B (Fig. 2), while gel filtration chromatography (Fig. 3) and gel electro- phoresis (Fig. 4) techniques were used to estimate the molec- ular weight of undenatured WAP. The molecular weights of WAP-A and WAP-B are judged to be similar by all three
8
6 ? ? 4 % 3 $ 2 1
I ( ’ I I
0.2 0.3 0.4 0.5 R f
FIG. 2. Determination of the molecular weight of WAP by SDSgel electrophoresis. SDS-polyacrylamide slab gel electropho- resis was performed by the method of Laemmli (7) at a 12.5% acrylamide concentration. Reference proteins and WAP-A and WAP- B were co-electrophoresed on the same slab gel and detected by Coomassie blue staining. Molecular weights are plotted relative to the dye front (Rn.
i
I I 80 90 100
ELUTION VOLUME ( ml )
FIG. 3. Determination of the molecular weight of WAP by gel filtration. Bio-Gel P-60 column chromatography (1.0 X 100 cm) was performed as described under “Materials and Methods.” Ftefer- ence proteins and WAP-A and WAP-B were chromatographed sepa- rately and detected by optical absorbance at 280 nm. Molecular weights are plotted uersus the elution volume of each protein.
I I I
6 -
-
I I
0 2 4 6 8 MOLECULAR WEIGHT X
FIG. 4. Determination of the molecular weight of WAP on gels of varying acrylamide concentration. Polyacrylamide slab gel electrophoresis a t pH 9.5 was performed using gels of 15%,16.25%, 17.5%, 18.758, and 20% acrylamide, as described under “Materials and Methods.” Reference proteins and WAP-A and WAP-B were co- electrophoresed on the same slab gel and detected by Coomassie blue staining. The molecular weights are shown in relation to the slopes of lines obtained by plotting log (R, X 100) uersus percentage gel for each protein (R, = migration relative to dye front), as in Fig. 6.
FIG. 5. Electrophoresis of WAP-A and WAP-B preparations
resis a t pH 9.5 in 7 M urea was performed on WAP-A and WAP-B showing minor components. Polyacrylamide slab gel electropho-
samples after Bio-Gel P-60 chromatography. The gel was stained with Coomassie blue. The sample applied in the left column contains 40 pg of WAP-A, the center column contains a mixture of 30 pg of WAP- A and 30 pg of WAP-B, and the column on the right contains 40 g of WAP-B. The cathode and the origin are at the top of the figure.
techniques with estimates of 13,500 by SDS-gel electropho- resis, 15,000 by gel filtration chromatography, and 12,500 by gel electrophoresis.
The gel electrophoresis technique used to measure the molecular weight of WAP reveals two minor bands in addition to the major band in variant and normal WAP samples (Fig. 5). Both minor bands appear to be forms of WAP based on their pink color after staining with Coomassie blue and on their similar molecular weights as shown in Fig. 6. Fig. 6 is a Ferguson plot as modified by Hedrick and Smith (9). When mobilities are plotted as a function of acrylamide concentra- tion, slopes of the resulting lines are proportional to molecular weights and the intercepts to molecular charge. When this procedure is carried out for the major and minor components of WAP-A and WAP-B preparations as shown in Fig. 6, all components exhibited similar slopes and thus possess similar molecular weights. However, each of the four discrete mobility forms observed in Fig. 6 exhibit uniquey-intercepts, suggesting that the mobility difference is due to charge density. It appears that each of the anodal minor components in WAP-A and WAP-B preparations exhibits a mobility different from, but proportional to, the mobility of their respective major WAP component. This observation suggests that the mecha- nism responsible for the electrophoretic difference between WAP-A and WAP-B also affects one, but not both, of the minor components of these preparations. Based on the density of Coomassie stain, each minor band represents approximately 5% of the total WAP. These minor components are separated from the major WAP component by the DEAE-cellulose chromatography step in the WAP purification procedure.
Phosphate and Sugar Content-Whey acidic protein was stained specifically for glycosyl and phosphate residues in order to determine whether these residues are associated with WAP and if they are involved in the WAP-B alteration. Neither WAP-A nor WAP-B stains positively with either periodic acid-Schiff, “Stains-all,” or methyl green dyes. The lack of any staining of WAP with “Stains-all” is an unusual

A Novel Whey Protein from Murine Milk 11513
feature. Control experiments indicate that as little as 20 pg of casein (1% phosphate w/w), when stained with methyl green, and 20 pg of calf fetuin (25% sugar w/w), when stained with the periodic acid-Schiff stain, are easily observed. Since as much as 150 pg of WAP failed to stain with methyl green, it appears that neither WAP-A nor WAP-B contain more than 3.5% carbohydrate by weight. The presence of carbohydrate and phosphate in WAP was further analyzed by enzymatic digestion with either neuraminidase, acid phosphatase, or alkaline phosphatase. These enzymes have no effect on the electrophoretic mobility of either WAP-A or WAP-B, or on the minor forms thereof.
Lipid Content-The presence of lipid in WAP was deter- mined by staining on gels with Sudan Black B (16). Approxi- mately 100 pg of WAP-A and WAP-B stains as well for lipid as does 20 pg of human serum low density lipoprotein. In order to determine the nature of the lipid associated with WAP, samples of WAP-A and WAP-B were delipidated by chloroform/methanol extraction (17) and the chloroform phases were analyzed for triglyceride and cholesterol using a Technicon automatic analyzer. The results of the lipid analysis in Table I1 indicate that triglyceride and cholesterol together account for 2045% percent of the total mass of WAP. Human low density lipoprotein was used as a positive control and was 65% lipid by mass. WAP-A and WAP-B are nearly identical in their lipid contents, the small differences in triglyceride and cholesterol being within background variation of the instru- ment.
2.2 L PERCENT ACRYLAMIDE
FIG. 6. Ferguson plots of electrophoretic components of WAP-A and WAP-B preparations. Polyacrylamide slab gel elec- trophoresis of WAP-A and WAP-B samples pusled by Bio-Gel P-60 chromatography was performed using gels of 15%, 16.25%, 17.576, 18.75%, and 20% acrylamide as described under “Materials and Meth- ods.” The log of the relative mobilities (Rm) were plotted as a function of the per cent acrylamide in the gels. The WAP-A forms are desig- nated by closed symbols and WAP-B by open symbols: A, A, fast migrating minor forms; 0, 0, major bands, U, m, slow migrating minor forms.
TABLE I1 Lipid analysis of WAP-A and WAP-B
Each sample contained 200 pg of Lowry protein and was prepared by the lipid extraction technique of Folch et al. (17). The chloroform phases were evaporated under nitrogen and analyzed for triglyceride and cholesterol content using an AAII Technicon automatic analyzer.
Sample Protein Cholesterol Tridvceride ~~~
clg
Low density lipoprotein 200 315 85 a-Lactalbumin 200 10 5
200 43 200
26 40 30
WAP-A WAP-B
TABLE 111 Amino acid residues of whey acidicprotein
Amino aeid WAP-A WAP-B nzol amino aeidlrnol WAP“
Aspartic acid 8.4 f 0.2b 8.3 f 0.2 Threonine 6.3 +- 0.1 6.2 rt 0.1 Serine 6.7 f 0.0 7.0 & 0.1 Glutamic acid 12.5 f 0.1 12.4 f 0.2 Proline 9.1 -c 0.2 9.0 % 0.1 Glycine 6.0 k 0.1 6.0 +. 0.2 Alanine 3.5 rt 0.1 Cysteine
3.5 +. 0.1 5.8 rt 0.1 5.2 k 0.1
Valine 6.4 f 0.1 6.0 0.1 Methionine 6.0 2 0.1 6.1 5~ 0.1 Isoleucine 5.0 2 0.1 5.0 * 0.1 Leucine 3.4 2 0.0 3.5 f 0.1 Tyrosine N.D.‘ N.D. Phenylalanine 2.5 a 0.2 Histidine
2.5 5 0.1 N.D. N.D.
Lysine 3.6 & 0.1 3.5 k 0.0 Arginine 2.3 k 0.1 NHs
3.0 k 0.1 12.2 f. 0.4 12.0 & 0.4
Moles of WAP were calculated based on a molecular weight of
bValues are mean f S.D. from three separate determinations of
e N.D., not detectable.
To test whether WAP has a serum lipoprotein-like density, WAP was centrifuged for 20 h in a potassium bromide solution of 1.21 specific gravity. Following centrifugation, the majority of immunoprecipitable WAP-A and WAP-B is found at the bottom of the centrifuge tube. This result suggests that the density of WAP is greater than that of serum lipoproteins. Another feature of WAP which distinguishes it from other lipoproteins is its existence in solution as a monomer (Fig. 3).
In order to determine whether lipid affects the electropho- retic migration of WAP-A and WAP-B, samples of each were delipidated by chloroform/methanol (17) and by tetrameth- ylurea extraction (5) and electrophoresed on pH 9.5 acrylam- ide gels (8). Delipidated WAP-A and WAP-B exhibit mobili- ties similar to nondelipidated samples and delipidation does not effect the relative mobility difference between variant and normal WAP.
Amino Acid Content-The amino acid compositions of purified samples of WAP-A and WAP-B are shown in Table 111. WAP contains high levels of glutamate, aspartate, and proline and no tryosine or histidine. The amino acid compo- sition of WAP does not resemble that of any previously reported milk protein from other species (1). A comparison of the amino acid compositions of WAP-A and WAP-B suggests that WAP-B may contain one more arginine and one less cysteine than WAP-A (Table 111). This difference is consistent with the electrophoretic and isoelectric point differences be- tween the two forms of WAP. However, the absolute cysteine values may be somewhat unreliable since a stable derivative of cysteine was not prepared by carboxymethylation or per- formic acid oxidation.
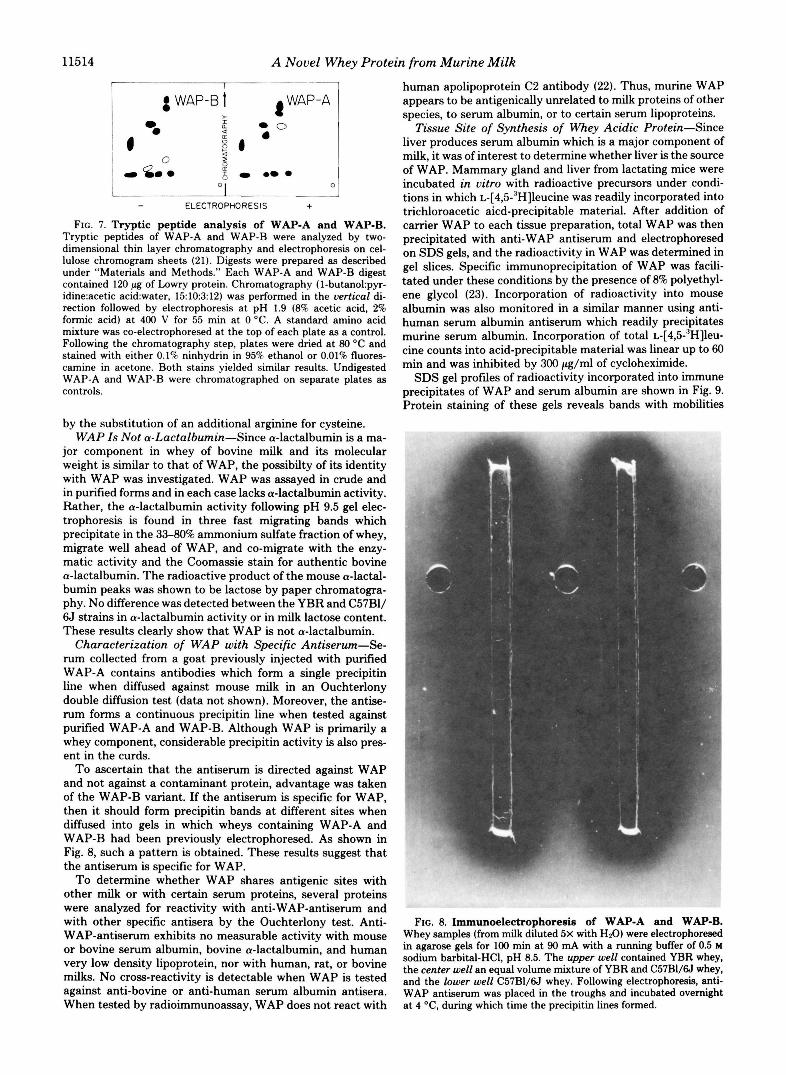
Tryptic Peptide Analysis-To test the possibility that WAP-€3 differs from WAP-A by a cysteine to arginine substi- tution, tryptic digests were prepared from variant and normal WAP. If correct, WAP-B digests should yield two unique peptides when compared with those of WAP-A. Peptide pat- terns after thin layer chromatography and electrophoresis of WAP-A and WAP-B digests are shown in Fig. 7. Two peptides are unique to the WAP-B sample and one to WAP-A as predicted for an arginine substitution. The WAP-B peptides display a reduced hydrophobicity in the chromatography di- mension and an increased positive charge by electrophoresis. This result strongly argues that WAP-€3 differs from WAP-A
14,500.
milk pooled from 3 mice of each strain.
11514 A Novel Whey Protein from Murine Milk
I I I
! a -. 0 6 WAP-6 1 (WAP-A 1
- ELECTROPHORESIS + FIG. 7. Tryptic peptide analysis of WAP-A and WAP-B.
Tryptic peptides of WAP-A and WAP-B were analyzed by two- dimensional thin layer chromatography and electrophoresis on cel- lulose chromogram sheets (21). Digests were prepared as described under "Materials and Methods." Each WAP-A and WAP-B digest contained 120 pg of h w r y protein. Chromatography (I-butano1:pyr- idine:acetic acid:water, 1510312) was performed in the uertical di- rection followed by electrophoresis at pH 1.9 (8% acetic acid, 2% formic acid) at 400 V for 55 min at 0 "C. A standard amino acid mixture was co-electrophoresed at the top of each plate as a control. Following the chromatography step, plates were dried at 80 O C and stained with either 0.1% ninhydrin in 95% ethanol or 0.01% fluores- camine in acetone. Both stains yielded similar results. Undigested WAP-A and WAP-B were chromatographed on separate plates as controls.
by the substitution of an additional arginine for cysteine. WAP Is Not a-Lactalbumin-Since a-lactalbumin is a ma-
jor component in whey of bovine milk and its molecular weight is similar to that of WAP, the possibilty of its identity with WAP was investigated. WAP was assayed in crude and in purified forms and in each case lacks a-lactalbumin activity. Rather, the a-lactalbumin activity following pH 9.5 gel elec- trophoresis is found in three fast migrating bands which precipitate in the 33-80% ammonium sulfate fraction of whey, migrate well ahead of WAP, and co-migrate with the enzy- matic activity and the Coomassie stain for authentic bovine a-lactalbumin. The radioactive product of the mouse a-lactal- bumin peaks was shown to be lactose by paper chromatogra- phy. No difference was detected between the YBR and C57B1/ 6J strains in a-lactalbumin activity or in milk lactose content. These results clearly show that WAP is not a-lactalbumin.
Characterization of WAP with Specific Antiserum-Se- rum collected from a goat previously injected with purified WAP-A contains antibodies which form a single precipitin line when diffused against mouse milk in an Ouchterlony double diffusion test (data not shown). Moreover, the antise- rum forms a continuous precipitin line when tested against purified WAP-A and WAP-B. Although WAP is primarily a whey component, considerable precipitin activity is also pres- ent in the curds.
To ascertain that the antiserum is directed against WAP and not against a contaminant protein, advantage was taken of the WAP-B variant. If the antiserum is specific for WAP, then it should form precipitin bands at different sites when diffused into gels in which wheys containing WAP-A and WAP-B had been previously electrophoresed. As shown in Fig. 8, such a pattern is obtained. These results suggest that the antiserum is specific for WAP.
To determine whether WAP shares antigenic sites with other milk or with certain serum proteins, several proteins were analyzed for reactivity with anti-WAP-antiserum and with other specific antisera by the Ouchterlony test. Anti- WAP-antiserum exhibits no measurable activity with mouse or bovine serum albumin, bovine a-lactalbumin, and human very low density lipoprotein, nor with human, rat, or bovine milks. No cross-reactivity is detectable when WAP is tested against anti-bovine or anti-human serum albumin antisera. When tested by radioimmunoassay, WAP does not react with
human apolipoprotein C2 antibody (22). Thus, murine WAP appears to be antigenically unrelated to milk proteins of other species, to serum albumin, or to certain serum lipoproteins.
Tissue Site of Synthesis of Whey Acidic Protein-Since liver produces serum albumin which is a major component of milk, it was of interest to determine whether liver is the source of WAP. Mammary gland and liver from lactating mice were incubated in vitro with radioactive precursors under condi- tions in which ~-[4,5-"H]leucine was readily incorporated into trichloroacetic aicd-precipitable material. After addition of carrier WAP to each tissue preparation, total WAP was then precipitated with anti-WAP antiserum and electrophoresed on SDS gels, and the radioactivity in WAP was determined in gel slices. Specific immunoprecipitation of WAP was facili- tated under these conditions by the presence of 8% polyethyl- ene glycol (23). Incorporation of radioactivity into mouse albumin was also monitored in a similar manner using anti- human serum albumin antiserum which readily precipitates murine serum albumin. Incorporation of total ~-[4,5-'H]leu- cine counts into acid-precipitable material was linear up to 60 min and was inhibited by 300 pg/ml of cycloheximide.
SDS gel profiles of radioactivity incorporated into immune precipitates of WAP and serum albumin are shown in Fig. 9. Protein staining of these gels reveals bands with mobilities
.*-
R
FIG. 8. Immunoelectrophoresis of WAP-A and WAP-B. Whey samples (from milk diluted 5x with HzO) were electrophoresed in agarose gels for 100 min at 90 mA with a running buffer of 0.5 M sodium barbital-HCI, pH 8.5. The upper well contained YBR whey, the center well an equal volume mixture of YBR and C57B1/6J whey, and the lower well C57B1/6J whey. Following electrophoresis, anti- WAP antiserum was placed in the troughs and incubated overnight at 4 O C , during which time the precipitin lines formed.

A Novel Whey Protein from Murine Milk 11515
identical with the light and heavy chains of goat IgG, to WAP, and to mouse serum albumin. A large peak of radioactivity was associated with the serum albumin band of liver but not with that of mammary gland (Fig. 9), an observation in agreement with the known tissue site of synthesis of serum albumin (2). Conversely, anti-WAP immunoprecipitates from mammary gland but not liver explants contained radioactivity which co-migrated with WAP (Fig. 9). These results argue strongly that WAP is synthesized by the mammary gland, but not the liver. In addition to the major radioactivity peak precipitated with anti-WAP antiserum from mammary gland, two additional peaks were seen with molecular weights of 34,000 and 27,000 (Fig. 9). Because of their co-precipitation with anti-WAP antiserum, these may represent precursors of WAP.
80 Sn’ ts I 60L n
14 12
8
4
‘0 .25 .50 7 5 1.00
Rm FIG. 9. Radioactivity profiles of SDS-gels after electropho-
resis of dissolved immunoprecipitates of mouse WAP and serum albumin after [‘HJleucine incorporation in tissue slices of liver and mammary gland. Liver and mammary gland tissue slices from C57B1/6J mice were incubated with ~-[4,5-’H]leucine and portions of each were treated separately with anti-WAP antiserum and anti-human serum albumin antiserum as described under “Ma- terials and Methods.” Immune precipitates were collected and elec- trophoresed on SDS-polyacrylamide gels. Following staining, the gels were frozen on Dry-Ice and sliced into 2-mm sections. Radioactivity in gel slices was determined by liquid scintillation spectrometry. Radioactivity is plotted as a function of relative mobility (Rm). The closed circles represent radioactivity in an immune precipitate pre- pared with anti-WAP antiserum and open circles represent that prepared with anti-human serum albumin antiserum.
TABLE IV Incorporation of radioactivity into immune-precipitated WAP and
total protein in mouse mammary gland explants Mammary gland explants (0.2 g) were labeled with radioactive
amino acids and [3*P]orthophosphate and analyzed for total acid- precipitable and specific WAP radioactivity as described under “Ma- terials and Methods.” WAP radioactivity was determined by immu- noprecipitation of WAP and measuring that radioactivity in such immunoprecipitates which co-migrates with authentic WAP on SDS gels (as in Fig. 9). WAP radioactivity was immunoprecipitated from an extract of the entire 0.2 g of mammary gland explant prepared after sampling 10 pl of homogenate in triplicate for total acid-precip- itable protein radioactivity.
(1) WAP ra- (7,) Total pro-
explants ity in explants Label dioactzvlty in tein radioactiv- (1)/(2) X 100
cpm/g cpm/g x ~-[4,5-~H]Leucine 19,950 160,141 0.120 ~-[3,4-~H]Proline 4,829 1,031 4.680 ~-[’~C]Tyrosine 192 8,450 0.020 [32P]Orthophosphate 40 43,154 0.001
Consistent with the view that the radioactivity migrating with WAP represents WAP synthesis are the results of an- other experiment in which mammary gland explants were incubated with ~-[3,4-~H]prolie, ~-[‘~C]tyrosine, or [”2p]or- thophosphate. Based on our knowledge of WAP composition, it was predicted that 3H, but not I4C or 32P, should co-migrate with stainable WAP protein on SDS gels. The results of these experiments are presented in Table IV.
The very low level of radioactivity incorporated into the M , = 13,000 peak with labeled tyrosine in contrast to the sub- stantial incorporation with labeled leucine or proline corre- lates well with the levels of these amino acids in WAP (Table 111). Moreover, no 32P radioactivity was associated with WAP, which is consistent with our observation that WAP is not a phosphoprotein.
DISCUSSION
We have identified a protein of previously undescribed characteristics in murine milk which we call WAP since it is the major whey protein of murine milk and because it exhibits an acidic isoelectric point. The molecular weight of WAP is approximately 14,000 and WAP accounts for a minimum of 2.4% of the mass of total protein in mouse milk. WAP is apparently unique to the mouse since no protein with similar properties has been described in the milk of other species and since goat anti-WAP antiserum does not cross-react with any component of bovine, rat, or human milk.
Based on its positive stain with Sudan Black B and its content of triglycerides and cholesterol (Table 11), WAP ap- pears to be a lipoprotein, with 20-25% of the mass as lipid. However, some samples of WAP were found which did not stain as strongly as others with Sudan Black B, suggesting that heterogeneity may exist in the amount of lipid associated with WAP. WAP is different from common serum lipoproteins since it exists in solution as a monomer and does not float after equilibrium centrifugation in a KBr solution of 1.21 specific gravity. The amount of lipid associated with WAP is also much lower than that of serum lipoproteins.
Immune precipitates of WAP from mammary gland and liver explants incubated with radioactive amino acids show that WAP is synthesized by the mammary gland but not by the liver (Fig. 9). Specific WAP synthesis represents 0.12% of the incorporated [’Hlleucine and 4.68% of the [3H]proline incorporated into total protein in the mammary gland. In agreement with the mammary site of synthesis, whey acidic protein is not detectable in mouse serum by either immuno- logic reactivity or by gel electrophoresis. Taken together, these results suggest that WAP is synthesized in the mammary gland although it cannot be ruled out that WAP is also synthesized in the liver in amounts too low to be immunolog- ically detectable.
What we have designated as WAP was previously proposed by Green and Pastewka (24) to be mouse a-lactalbumin. WAP is now shown to have an amino acid composition very different from that of lactalbumin of other species, and to contain no lactose synthetase B-protein activity. In addition, WAP is completely separated from a-lactalbumin by ammonium sul- fate fractionation and by electrophoresis. We show that mouse lactose synthetase B-protein activity is localized in three fast migrating bands of the 33-80% ammonium sulfate fraction of whey when electrophoresed in pH 9.5 gels containing urea. The fastest migrating mouse a-lactalbumin band stains as a glycoprotein with “Stains-All’’ and the other two bands stain as naked proteins in a manner consistent with the a-lactal- bumin of other species (19).
The absence of detectable phosphate (Table IV) and the solubility of WAP at acid pH makes it unlikely that WAP is

11516 A Novel Whey Protein from Murine Milk
a casein. Nevertheless, WAP may be related to casein since anti-WAP immune precipitates of in uitro-labeled mammary gland protein on SDS gels display possible percursor forms of WAP with molecular weights equivalent to as,,- and p-casein (Fig. 9) and WAP exhibits a high proline content similar to casein (1).
Whey acidic protein has several properties in common with human serum very low density lipoproteins. Very low density lipoprotein is associated with triglyceride and cholesterol, as is WAP (16). Also, the apo-C lipoproteins from very low density lipoprotein exhibit a molecular weight of 10,000 and have low levels of histidine and tyrosine similar to WAP (25). The isoelectric points of the apo-C lipoproteins are also similar to WAP: apo-C-I1 = 4.79, apo-C-IIIo = 4.93, apo-C-III1 = 4.7, and apo-C-1112 = 4.53, compared to wild type WAP with PI = 4.7. Serum apo-C-I1 activates and apo-C-I11 inhibits bovine milk lipoprotein lipase activity (25,26). In order to determine the possible relationship between these proteins, we tested for cross-reactivity using specific anti-WAP antisera by Ouchter- lony analysis and by using radioimmunoassay of human apo- C-11. No cross-reactivity is found. These data, and the absence of WAP in mouse serum, indicates that WAP is probably not a serum very low density lipoprotein. Nevertheless, these experiments do not rule out a possible relationship between WAP and very low density lipoprotein.
Proteins similar to WAP have been reported in bovine colostrum (27) and in rat milk (14). The bovine colostral protein M-1 (28) is found in whey, exhibits an amino acid composition similar to WAP including a lack of tyrosine and histidine, and has the same molecular weight as WAP (27). However, M-1 is a phosphoglycoprotein and is found only in colostrum. If WAP is the murine equivalent of bovine "1, then its expression would have diverged at some develop- mental switch which allows it to be expressed throughout lactation (29). Another protein described in rat milk exhibits similarities to WAP and M-1 in molecular weight and amino acid composition (14). Designated P-protein, this component of rat milk is phosphorylated like M-1 and, like WAP, exhibits the unusual pink color when stained with Coomassie blue and viewed by reflected light. WAP, however, differs from M-1 and P-protein in that it lacks carbohydrate and phosphate and contains lipid. Since P-protein, M-1, and WAP have nearly identical amino acid compositions, they may represent species-specific forms of the same naked protein. No functions have been ascribed to any of these proteins.
A unique property of WAP is its pink character when stained with Coomassie blue and viewed by reflected light. Other proteins have been reported from human saliva (30) and from rat milk (14) with similar Coomassie staining prop- erties. The basis of this unusual staining property is not understood, although each of these proteins lacks tyrosine and histidine.
The electrophoretic variation in WAP was identified in the YBR strain of mice during a screen for milk protein variants (31). Amino acid and tryptic peptide analyses of WAP-A and WAP-B suggest that this difference may result from a cysteine to arginine amino acid substitution. The tryptic peptide maps of WAP-A and WAP-B, however, cannot distinguish whether the substituted amino acid is lysine or arginine, nor do they distinguish the amino acid unique to WAP-A. Other obser- vations consistent with a cysteine to arginine substitution are the relative isoelectric points of WAP-A and WAP-B, the faster electrophoretic mobility of WAP-B at acid pH, and the slower mobility of WAP-B at basic pH. No differences are observed in molecular weights between WAP-A and WAP-B. The proposed cysteine to arginine substitution is consistent with a simple base substitution in the genetic code (32). A
transition in position 1 of either of the cysteine codons UGU or UGC would produce the arginine codons CGU or CGC, respectively.
In a separate study, we demonstrate that two alleles of an autosomal locus are responsible for the difference between WAP-A and WAP-B (31). We designate this locus as Wap with the alleles WapA and Wap". We now propose that Wap is the structural gene for whey acidic protein.
Acknowledgments-We thank Dr. Verne Chapman of Roswell Park Memorial Institute, Buffalo, New York, for supplying mice of the YBR strain, Dr. M. Kashyap of the University of Cincinnati College of Medicine for electrofocusing equipment, and Dr. Clark West of Children's Hospital Research Foundation, Cincinnati, Ohio for anti-human serum albumin antiserum.
REFERENCES 1. Jenness, R. (1974) in Lactation, A Comprehensive Treatise (Lar-
son, B. L., and Smith, V. R., eds) Vol. 3, pp. 3-96, Academic Press, New York
2. McKenzie, H. A., ed (1971) Milk Proteins, Vols. 1 and 2, Academic Press, New York
3. Thompson, M. P., and Farrell, H. M., Jr. (1974) in Lactation, A Comprehensive Treatise (Larson, B. L., and Smith, V. R. eds) Vol. 3, pp. 109-132, Academic Press, New York
4. McBurney, J. J., Meir, H., and Hoag, W . G. (1964) J. Lab. Clin. Med. 64,485-487
5. Kane, J . P., Sata, T., Hamilton, R. L., and Havel, R. J. (1975) J. Clin. Invest. 56, 1622-1634
6. Albers, J. J., and Scanu, A. M. (1971) Biochim. Biophys. Acta
7. Laemmli, U. K. (1970) Nature 227,680-685 8. Feldman, M. K., and Hohmann, P. (1971) Int. J. Biochem. 2,477-
9. Hedrick, J. L., and Smith, A. J. (1968) Arch. Biochem. Biophys.
236,29-37
480
126,155-164 10. Davis, B. J. (1964) Ann. N.Y. Acad. Sci. 121,404-427 11. Segrest, J. P., and Jackson, R. L. (1972) Methods Enzymol. 28B,
12. Green, M. R., Pastewka, J. V., and Peacock, A. C . (1973) Anal.
13. Cutting, J. A., and Roth, T. R. (1973) Anal. Biochem. 54,386-394 14. McKenzie, R. M., and Larson, B. L. (1978) J. Dairy Sci. 61, 723-
728 15. Pastewka, J. V., Reed, R. A., Ness, A. T., and Peacock, A. C.
(1973) Anal. Biochem. 51, 152-158 16. Noble, R. P. (1968) J. Lipid Res. 9,693-700 17. Folch, J., Lees, M., and Sloane Stanley, G. H. (1957) J. Biol.
18. Brew, K., Vanaman, T. C., and Hill, R. L. (1968) Proc. Nutl.
19. Brew, K., and Campbell, P. N. (1967) Biochem. J. 102,265-274 20. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
21. Stephens, R. E. (1978) Anal. Biochem. 84, 116-126 22. Kashyap, M L., Srivastava, L. S., Chen, C. Y., Perisutti, J.,
Campbell, M., Lutmer, F., and Glueck, C. J. (1977) J. Clin. Invest. 60, 171-180
23. Creighton, W . D., Lambert, P. H., and Miescher, P. A. (1973) J. Immunol. 111, 1219-1226
24. Green, M. R., and Pastewka, J. V. (1975) J. Dairy Sci. 59, 207- 215
25. Hael, R. J., Fielding, C. J., Olirecrona, T., Shore, V. G., Fielding, P. E., and Egelrud, T. (1973) Biochemistry 12, 1828-1833
26. Miller, A. L., and Smith, L. C. (1973) J. Biol. Chem. 248, 3359- 3362
27. Bezkorovainy, A. (1965) Arch. Biochem. Biophys. 110,558-567 28. Bezkorovainy, A., and Grohlich, D. (1969) Biochem. J. 115,817-
29. Morris, I. G. (1969) J. Immunol. 17, 139-149 30. Henkin, R. I., Lippoldt, R. E., Bilstad, J., Wolf, R. O., Lum, C. K.
31. Piletz, J. E., and Ganschow, R. E. (1981) Biochem. Genet., in
32. Crick, F. H. C. (1968) J. Mol. Biol. 38, 367-379
54-63
Biochem. 56,43-51
Chem. 226,497-509
Acad. Sci. U. S. A. 59,491-497
(1951) J. Biol. Chem. 193,265-275
822
L., and Edelhoch, H. (1978) J. Biol. Chem. 253, 7556-7565
press
Related Documents











