
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biotechnology and Bioprocess Engineering 15: 696-703 (2010)
DOI 10.1007/s12257-009-3126-9
Biochemical Characterization and Potential for Textile Dye
Degradation of Blue Laccase from Aspergillus ochraceus NCIM-1146
Amar A. Telke, Avinash A. Kadam, Sujit S. Jagtap, Jyoti P. Jadhav, and Sanjay P. Govindwar
Received: 23 November 2009 / Revised: 12 December 2009 / Accepted: 4 January 2010
© The Korean Society for Biotechnology and Bioengineering and Springer 2010
Abstract In our study, we produced intracellular blue
laccase by growing the filamentous fungus Aspergillus
ochraceus NCIM-1146 in potato dextrose broth. The
enzyme was then purified 22-fold to a specific activity
of 4.81 U/mg using anion-exchange and size exclusion
chromatography. The molecular weight of purified laccase
was estimated as 68 kDa using sodium dodecyl sulfate
polyacrylamide gel electrophoresis. The enzyme showed
maximum substrate specificity toward 2,2'-Azinobis, 3-
ethylbenzothiazoline-6-sulfonic acid than any other sub-
strate. The optimum pH and temperature for laccase activity
were 4.0 and 60ºC, respectively. The purified enzyme was
stable up to 50ºC, and high laccase activity was maintained
at pH 5.0 ~ 7.0. Laccase activity was strongly inhibited by
sodium azide, EDTA, dithiothreitol, and L-cysteine. Puri-
fied laccase decolorized various textile dyes within 4 h in
the absence of redox mediators. HPLC and FTIR analysis
confirmed degradation of methyl orange. The metabolite
formed after decolorization of methyl orange was charac-
terized as p-N,N'-dimethylamine phenyldiazine using GC-
MS.
Keywords: Aspergillus ochraceus NCIM-1146, laccase,
textile dyes, decolorization, redox mediators
1. Introduction
The decolorization of textile dyes can be performed with
various microorganisms, including fungi, actinomycetes,
algae, and bacteria. Fungi and bacteria especially are wide-
ly used for the biodegradation of textile dyes. Biodegrada-
tion ability is associated with the production of oxido-
reductive enzymes such as lignin peroxidase and laccase
[1]. Most of the research concerning bioremediation is
centered on a single fungal species, Phanerochaete chryso-
sporium, which is known to metabolize a wide range of
xenobiotic compounds [2]. Ascomycetes such as Asper-
gillus niger, Aspergillus fumigatus, Aspergillus oryzae, and
Aspergillus olliaceus strain 212C are capable of decolori-
zing a wide range of structurally different dyes and are
more effective than Phanerochaete chrysosporium [3-6].
Our earlier reports showed that the decolorization of textile
dyes using Aspergillus ochraceus NCIM-1146 is associated
with laccase [7].
Laccases are the most abundant members of the multi-
copper protein family, which also includes tyrosinases,
monoxygenases, and dioxygenases [8]. Phylogenetically,
these enzymes have developed from small sized prokar-
yotic azurins to eukaryotic plasma proteins, such as cerulo-
plasmin [9]. Laccases contain four histidine-rich copper-
binding domains, which coordinate copper atoms types 1-
3 that differ in their environment and spectroscopic
properties [10]. Laccases are categorized as blue laccase
(laccase containing four copper sites) and laccase lacking
type-1 copper site [11]. Laccases are the model enzymes
for multi-copper oxidases and participate in the cross-
linking of monomers, degradation of polymers and ring
cleavage of aromatic compounds [12]. Further, they possess
great biotechnological potential because of their wide
reaction capabilities and broad substrate specificities.
Promising applications for laccases include biosensors for
drug analysis and phenols in tea, polymer synthesis, textile-
dye bleaching, bioremediation, and pulp bleaching [13-16].
The majority of previous studies have focused on laccase-
Amar A. Telke, Avinash A. Kadam, Sujit S. Jagtap, Jyoti P. Jadhav, SanjayP. Govindwar*Department of Biochemistry, Shivaji University, Kolhapur 416004, IndiaTel: +91-231-2609152; Fax: +91-231-2691533E-mail: [email protected]
RESEARCH PAPER

Biochemical Characterization and Potential for Textile Dye Degradation of Blue Laccase from Aspergillus ochraceus NCIM-1146 697
producing Trametes versicolor, Pleurotus ostreatus, and
Phanerochaete chrysosporium [17]. However, there is only
limited research on intracellular laccase produced by Asper-
gillus species such as Aspergillus nidulans [18,19]. In the
present work, we report the biochemical properties of blue
laccase from Aspergillus ochraceus NCIM-1146 as well as
its mechanism of textile dye degradation.
2. Materials and Methods
2.1. Dyestuff and chemicals
2,2'-Azinobis, 3-ethylbenzothiazoline-6-sulfonic acid (ABTS),
o-tolidine, hydroquinone, pyrogallol, guaiacol, L-DOPA,
and commassie brilliant blue R-250 were obtained from
SRL Chemicals, India. Veratryl alcohol, p-cresol, o-dianisi-
dine, L-tyrosine, sodium azide, L-cysteine, EDTA, dithi-
othreitol, Methyl orange, peptone, yeast extract and agar
powder were obtained from Hi-Media laboratory, India.
Protein markers were obtained from Bangalore Genei,
India. Textile dyes were obtained from local industry at
Ichalkaranji, India. All chemicals were of the highest purity
and of analytical grade.
2.2. Microorganism and culture conditions
A. ochraceus NCIM-1146 was obtained from the National
Center for Industrial Microorganisms, Pune, India. The
stock culture was maintained on potato-dextrose agar slants
at 4ºC.
For the enzyme production, two fungal discs (8 mm
diameter) of 4-day-old culture were inoculated into 250
mL Erlenmeyer flasks containing 100 mL of PDB (potato-
dextrose broth) medium with 200 g/L of peeled potatoes
and 5.0 g/L of yeast extract, followed by incubation for 96
h at 30ºC with shaking.
2.3. Preparation of crude enzyme
The fungal mycelium was collected by filtering the 96 h
growth culture of A. ochraceus NCIM-1146. The mycelium
was suspended in 50 mM potassium phosphate buffer (pH
7.0) and homogenized using a mortar and pestle and later
a homogenizer. The homogenized sample was centrifuged
at 9,000 rpm for 15 min under cold conditions. The super-
natant obtained after centrifugation was used as a source of
crude enzyme.
2.4. Enzyme activity
Laccase activity was measured in the reaction mixture
(2.0 mL) containing 100 µM ABTS and 20 mM sodium
acetate buffer (pH 4.0). The reaction was started by the
addition of 0.2 mL of enzyme solution [20]. The substrate
oxidation was monitored at 420 nm (ε420 = 0.036/µM/cm).
One unit of enzyme activity was defined as the amount of
enzyme required to produce 1 µM of oxidized product per
min. The protein concentration of the crude sample was
determined by the Lowry method using bovine serum
albumin as a standard protein [21]. The protein concent-
ration was monitored based on the absorbance at 280 nm
after anion-exchange and size exclusion chromatography.
2.5. Purification of laccase
DEAE-anion exchange chromatography was carried out
using an automated Econo purification system (Bio-rad).
The crude enzyme was directly applied to a DEAE-cellu-
lose anion exchange column (cylindrical glass column
15 cm in height and 1 cm in diameter) equilibrated with
20 mM potassium phosphate buffer (pH 8.0) at a flow rate
of 1 mL/min. The retained proteins were eluted with a
linear NaCl gradient former (0 ~ 0.4 M). Size exclusion
chromatography was carried out using a cylindrical glass
column packed with Biogel P100 (50 cm height and 1 cm
diameter). The column was equilibrated with 20 mM pota-
ssium phosphate buffer (pH 7.0).
2.6. Gel electrophoresis, spectrum, and copper content
Native and sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) were carried out on 5%
stacking and 11% resolving gels using a Genei vertical
electrophoresis system (Bangalore Genei, Pvt. Ltd., India).
Protein bands were stained with commassie brilliant blue
R-250. The molecular mass of purified laccase was deter-
mined by calculating the relative mobility of standard pro-
tein markers, such as phosphorylase b (97.40 kDa), bovine
serum albumin (66.0 kDa), ovalbumin (43.0 kDa), carbonic
anhydrase (29.0 kDa), soybean trypsin inhibitor (20.10
kDa), and lysozyme (14.30 kDa). Activity staining was
carried out by incubating the gel after native PAGE at room
temperature in 20 mM sodium acetate buffer (pH 6.5) with
1 mM L-DOPA [22].
Purified laccase (10 µM) in 20 mM sodium acetate buffer
(pH 4.0) was subjected for wavelength scan (200 ~ 800
nm) by a UV-visible spectrophotometer (U-2800, Hitachi,
Japan). The copper content was determined using an atomic
absorption spectrophotometer (Perkin Elmer, model no.
4100).
2.7. Effect of pH and temperature on laccase activity
and stability
The enzyme obtained after size exclusion chromatography
was used for the biochemical characterizations. The effect
of pH on laccase activity was determined within a pH
range of 2.5 ~ 8.5 using ABTS as a substrate at room
temperature (30ºC). The optimum temperature for purified
laccase was examined over the temperature range of 20 ~

698 Biotechnology and Bioprocess Engineering 15: 696-703 (2010)
80°C with ABTS as a substrate and at optimal pH.
The effect of temperature and pH on laccase stability
was studied by incubating 3.61 U of laccase at various
temperatures between 20 and 60ºC at pH 7.0 or at various
pHs between 3.0 and 9.0 at 25ºC. Sodium citrate and
sodium acetate buffers were used for maintaining the pH
between 3.0 and 6.0, potassium phosphate buffer for main-
taining the pH between 7.0 and 8.0, and sodium carbonate-
sodium bicarbonate buffer for pH 9.0 ~ 10.0. Aliquots
were transferred after specific intervals into a cuvette con-
taining 100 µM ABTS and 20 mM sodium acetate buffer
(pH 4.0) in order to determine residual laccase activity.
2.8. Substrate specificity and kinetics of laccase
Substrate specificity was studied using nonphenolic and
phenolic compounds, such as veratryl alcohol, o-tolidine,
ABTS, pyrogallol, guaiacol, hydroquinone, tyrosine, p-
cresol, o-dianisidine, and L-DOPA, as substrates. The rate
of substrate oxidation was determined by measuring the
absorbance increase with the molar extinction coefficient
(εm) obtained from the literature [11-23]. One unit of
enzyme activity was defined as the amount of enzyme
required to increase 1.0 ABS unit/min.
2.9. Effect of metal salts, salinity, and inhibitor on laccase
activity
We studied the effect of different metal salts (1 mM; MgCl2,
CaCl2, MnSO4, ZnSO4, HgCl2, FeSO4, and CuSO4), salinity
(1 to 500 mM) and inhibitors (1 and 5 mM; EDTA,
dithiothreitol, L-cysteine, and sodium azide) on laccase
activity. The enzyme (3.05 U) was incubated at various
concentrations of metal salts, NaCl and inhibitor for
15 min. Laccase activity was determined using ABTS as a
substrate.
2.10. Decolorization of textile dyes using pure laccase
The reaction mixture (5 mL) for the decolorization of
textile dyes contained 50 mg/L of textile dyes, 20 mM
sodium acetate buffer (pH 4.0), and 10 µM of pure enzyme
solution. The reaction mixture was incubated at 40ºC with
shaking. Heat inactivated enzyme was used in the control
experiment. All experiments were run in triplicate and the
average value was calculated.
2.11. Extraction and analysis of metabolites formed
after decolorization
The decolorization was monitored at λmax of respective dye
using a Hitachi U-2800 spectrophotometer. In order to
elucidate the mechanism of dye decolorization, we analy-
zed the degradation metabolites of pure azo dye Methyl
orange using HPLC, FTIR, and GC-MS. The metabolites
formed after decolorization of Methyl orange were extract-
ed three times with 10 mL of ethyl acetate with vigorous
shaking. The combined organic phase was filtered over
Na2SO4 on filter paper and concentrated in a rotary vacuum
evaporator. HPLC analysis was carried out (Waters model
no. 2690) on a C18 column (symmetry, 4.6 × 250 mm)
using the isocratic method with a 10 min run time. The
mobile phase used was methanol at a flow rate of 0.75 mL/
min with a UV detector at 280 nm. FTIR analysis was
performed in the mid IR region of 400 ~ 4,000/cm with
16 scan speed. The pellets were prepared using spectro-
scopically pure KBr (5:95) and fixed in a sample holder.
GC-MS analysis was carried out using a QP 5000 mass
spectrophotometer (Shimadzu model no. U-2800). The
ionization voltage was 70 eV. Gas chromatography was
conducted in temperature programming mode with a Re-
steck column (0.25 mm × 30 mm; XTI-5). The initial column
temperature was 40ºC for 4 min, which was then increased
linearly at 10ºC per min up to 270ºC and held for 4 min.
The temperature of the injection port was 275ºC. GC-MS
interface was maintained at 300ºC. The helium was used as
carrier gas at a flow rate of 1 mL/min with a 30 min run
time. The compounds were identified on the basis of mass
spectra using the NIST library of GC-MS (version 1.10
beta Shimadzu).
3. Results and Discussion
3.1. Purification of laccase
The majority of fungi produce both intracellular and extra-
cellular laccases. The localization of laccases seems to be
associated with the physiological functions of the enzymes
Table 1. Summary of purification of laccase from A. ochraceous NCIM-1146
Purification stepsTotal activity
(U)Total protein
(mg)Specific activity
(U/mg)Purification
foldYield(%)
Crude enzyme 55 255 0.215 − 100
DEAE-anion exchanger chromatography 28 8.98 3.10 15 50
Size exclusion chromatography 8.0 1.66 4.81 22 15
U- units.DEAE-diethylaminoethyl.
Biochemical Characterization and Potential for Textile Dye Degradation of Blue Laccase from Aspergillus ochraceus NCIM-1146 699
[24]. Optimal laccase production was observed after 96 h
growth of A. ochraceus NCIM-1146. The intracellular lac-
case was purified using DEAE-cellulose anion exchange
and size exclusion chromatography. The enzyme was eluted
at a NaCl concentration of 0.25 M from the DEAE-cellu-
lose anion exchange column. The procedure yielded 1.66
mg of pure protein, and the recovery of total laccase
activity was 15% with 22-fold purification (Table 1).
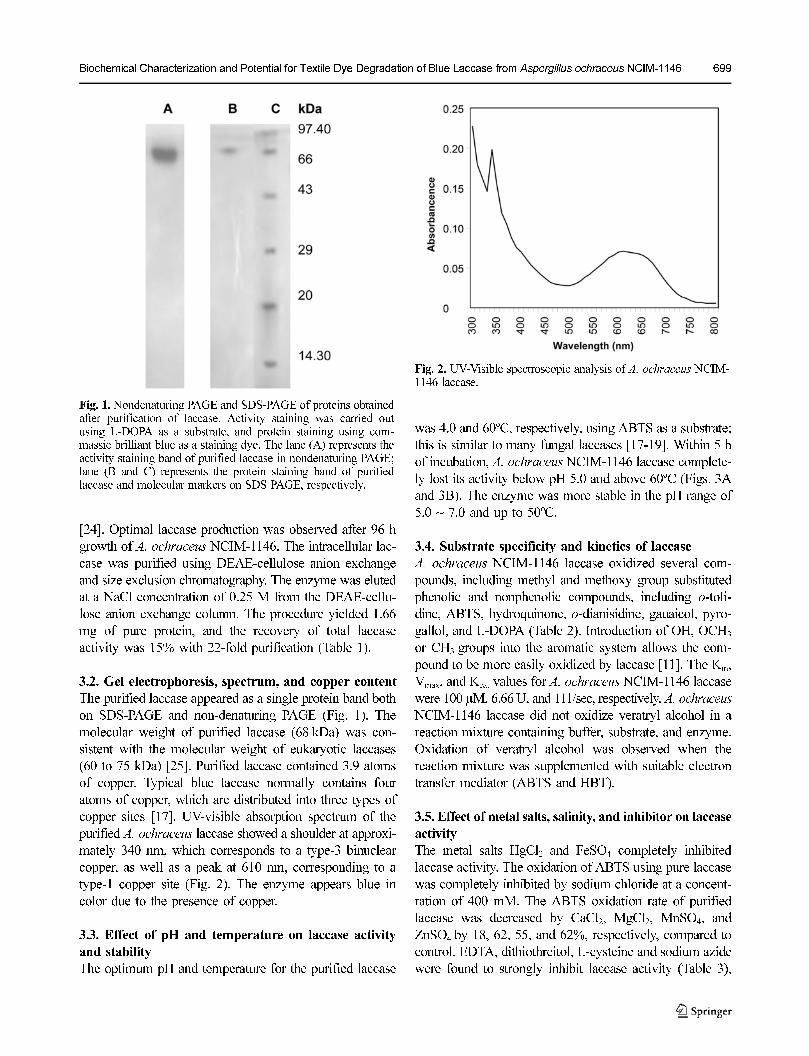
3.2. Gel electrophoresis, spectrum, and copper content
The purified laccase appeared as a single protein band both
on SDS-PAGE and non-denaturing PAGE (Fig. 1). The
molecular weight of purified laccase (68 kDa) was con-
sistent with the molecular weight of eukaryotic laccases
(60 to 75 kDa) [25]. Purified laccase contained 3.9 atoms
of copper. Typical blue laccase normally contains four
atoms of copper, which are distributed into three types of
copper sites [17]. UV-visible absorption spectrum of the
purified A. ochraceus laccase showed a shoulder at approxi-
mately 340 nm, which corresponds to a type-3 binuclear
copper, as well as a peak at 610 nm, corresponding to a
type-1 copper site (Fig. 2). The enzyme appears blue in
color due to the presence of copper.
3.3. Effect of pH and temperature on laccase activity
and stability
The optimum pH and temperature for the purified laccase
was 4.0 and 60ºC, respectively, using ABTS as a substrate;
this is similar to many fungal laccases [17-19]. Within 5 h
of incubation, A. ochraceus NCIM-1146 laccase complete-
ly lost its activity below pH 5.0 and above 60ºC (Figs. 3A
and 3B). The enzyme was more stable in the pH range of
5.0 ~ 7.0 and up to 50ºC.
3.4. Substrate specificity and kinetics of laccase
A. ochraceus NCIM-1146 laccase oxidized several com-
pounds, including methyl and methoxy group substituted
phenolic and nonphenolic compounds, including o-toli-
dine, ABTS, hydroquinone, o-dianisidine, gauaicol, pyro-
gallol, and L-DOPA (Table 2). Introduction of OH, OCH3
or CH3 groups into the aromatic system allows the com-
pound to be more easily oxidized by laccase [11]. The Km,
Vmax, and Kcat values for A. ochraceus NCIM-1146 laccase
were 100 µM, 6.66 U, and 111/sec, respectively. A. ochraceus
NCIM-1146 laccase did not oxidize veratryl alcohol in a
reaction mixture containing buffer, substrate, and enzyme.
Oxidation of veratryl alcohol was observed when the
reaction mixture was supplemented with suitable electron
transfer mediator (ABTS and HBT).
3.5. Effect of metal salts, salinity, and inhibitor on laccase
activity
The metal salts HgCl2 and FeSO4 completely inhibited
laccase activity. The oxidation of ABTS using pure laccase
was completely inhibited by sodium chloride at a concent-
ration of 400 mM. The ABTS oxidation rate of purified
laccase was decreased by CaCl2, MgCl2, MnSO4, and
ZnSO4 by 18, 62, 55, and 62%, respectively, compared to
control. EDTA, dithiothreitol, L-cysteine and sodium azide
were found to strongly inhibit laccase activity (Table 3),
Fig. 1. Nondenaturing PAGE and SDS-PAGE of proteins obtainedafter purification of laccase. Activity staining was carried outusing L-DOPA as a substrate, and protein staining using com-massie brilliant blue as a staining dye. The lane (A) represents theactivity staining band of purified laccase in nondenaturing PAGE;lane (B and C) represents the protein staining band of purifiedlaccase and molecular markers on SDS PAGE, respectively.
Fig. 2. UV-Visible spectroscopic analysis of A. ochraceus NCIM-1146 laccase.

700 Biotechnology and Bioprocess Engineering 15: 696-703 (2010)
similar to previously reported results [26].
3.6. Decolorization of textile dyes and analysis of obtained
metabolites
Earlier reports have demonstrated the expression of intra-
cellular laccase during the development of Aspergillus
nidulans fruitbody [18,19]. Previously, the involvement of
laccase in the decolorization of textile dyes by Aspergillus
ochraceus was shown [20]. A. ochraceus NCIM-1146 lac-
case decolorized textile dyes in the absence of redox
mediators with a decolorization efficiency from 56 to 90%
(Table 4). Trametes versicolor laccase showed 27 and 40%
decolorization in the presence of HBT after 5 and 24 h of
treatment, respectively, whereas only 7 and 15% was
observed in the absence of mediator [27]. The textile dyes
reactive navy blue HER, reactive yellow 84-A and methyl
orange showed absorbance peaks at 610, 460, and 470 nm
respectively (Fig. 4). The fact that the decolorized sample
did not show corresponding absorbance peaks suggests
decolorization of textile dyes (Fig. 4). The difference in the
Fig. 3. The pH (A) and temperature (B) stability of purified
laccase. Enzyme activity after 0 (◆), 1 (■), 2 (▲), 3 (◇), 4 (□),and 5 h (△). Data points represent the mean of three independentreplicates, standard error of the mean (SE) is indicated by errorbars.
Table 2. The oxidation of phenolic and nonphenolic substrates bypurified laccase from A. ochraceous NCIM-1146
Substratesλmax
(nm)Specific activity*
ABTS 420 0.505
o-tolidine 366 0.498
2,6-dimethoxyphenol 468 0.260
Veratryl alcohol 310 ND
Hydroquinone 248 0.333
L-DOPA 475 0.223
o-Dianisidine 460 0.111
Guaiacol 465 0.238
Pyrogallol 450 0.215
p-Cresol 400 ND
Tyrosine 278 ND
*U/mg of protein/min. ND: Not detected.
Table 3. Influence of inhibitors on the laccase activity
InhibitorsRelative activity (%)
1 mM 5 mM
Control 100 100
Sodium azide 18 09
EDTA ND ND
Dithiothreitol 09 ND
L-Cystein 09 ND
ND: Not detected.
Table 4. Decolorization potential of A. ochraceus NCIM-1146 laccase
Dyes Dye class C.I. No. CAS No. λmax (nm) Decolorization (%)*
Reactive navy blue HER Vinyl sulfone NA 77905-32-5 610 90
Reactive golden yellow HER Sulfonated monoazo NA 61951-85-7 460 90
Methyl orange Sulfonated monoazo 13025 547-58-0 470 56
*The decolorization was measured after 4 h of incubation.

Biochemical Characterization and Potential for Textile Dye Degradation of Blue Laccase from Aspergillus ochraceus NCIM-1146 701
FTIR spectra of Methyl orange and metabolites obtained
after decolorization suggests that biodegradation occurred.
The FTIR spectrum of Methyl orange showed peaks at
569/cm for S-O stretching vibrations, 702/cm for C-H
stretching vibrations, 844/cm for C-N bending vibrations,
1031/cm for S-O stretching vibrations, and 1168/cm for N-
CH3 stretching vibrations. The group frequency region
showed specific peaks for functional groups, peaks at
1367/cm for N=N stretching vibrations, 1518/cm for N-
CH3 bending vibrations, 1602/cm for C-H bending vibra-
tions, 2914/cm for C-H stretching vibrations in -CH3 group,
and 3481/cm for O-H stretching vibrations (Fig. 5). On the
other hand, the FTIR spectrum of metabolites obtained
after decolorization of methyl orange showed peaks at
1032/cm for N-H stretching vibrations, 1117/cm for C-O
stretching vibrations, 1464/cm for C=C stretching vibra-
tions, 1610/cm for C-C stretching vibrations, 174/cm for
C=O stretching vibrations, 2854/cm for -CH3 stretching
vibrations, 2920/cm for -CH2 stretching vibrations, and
3429/cm for N-H stretching vibrations (Fig. 5). The absence
of a peak at 1367/cm suggests that the azo bond was
cleaved. HPLC analysis of methyl orange showed a peak at
Fig. 4. Spectroscopic analysis for decolorization of reactive navy blue HER (A) reactive golden yellow HER (B) and methyl orange (C).
Spectrum of initial sample (◆) and spectrum of decolorized sample (▲). Data points represent the mean of three independent replicates,standard error of the mean (SE) is indicated by error bars.
Fig. 5. FTIR spectrum of methyl orange and metabolites obtained after decolorization.

702 Biotechnology and Bioprocess Engineering 15: 696-703 (2010)
2.26 min, whereas metabolites obtained after decolorization
showed peaks at 3.39, 3.49, and 3.84 min. The difference
in retention time between methyl orange and metabolites
formed after decolorization implies the biodegradation of
methyl orange into different metabolites. The metabolite
formed after the degradation of methyl orange was identi-
fied as p-N,N'-dimethylamine phenyldiazine using GC-MS
(Table 5).
The first step in the decolorization of azo dyes using
laccase is the formation of an electron-deficient reaction
center (carbocation). Carbocation creates a highly reactive
intermediate, which is often subject to nucleophillic attack
by nucleophiles such as -OH, -SO3 or halogen ions, result-
ing into asymmetric cleavage of the azo bond [11]. The
biodegradation of methyl orange involves asymmetric cleav-
age of the azo bond, resulting in formation of a p-N,N'-
dimethylamine phenyldiazine intermediate and a p-hydr-
oxybenzene sulfonic acid intermediate (Fig. 6).
4. Conclusion
Compared to the considerable research conducted on azo
dyes, the information exists on its biodegradation mechanism
is limited. In this paper, we describe the mechanism for azo
dye degradation by intracellular blue laccase of A. ochraceus
NCIM-1146, which has potential for environmental appli-
cations.
Acknowledgements
The authors are thankful to the University Grants Commi-
ssion, New Delhi for the financial assistance. Authors also
thank to Common Facility Center, Shivaji University,
Kolhapur, India for GC-MS facility.
References
1. Kalyani, D. C., P. S. Patil, J. P. Jadhav, and S. P. Govindwar(2008) Biodegradation of reactive textile dye Red BLI by an iso-lated bacterium Pseudomonas sp. SUK1. Bioresour. Technol. 99:4635-4641.
2. Cameron, M., S. Timofeevski, and S. Aust (2000) Mini-review:Enzymology of Phanerochaete chrysosporium with respect tothe degradation of recalcitrant compounds and xenobiotics. Appl.Microbiol. Biotechnol. 54: 751-758.
3. Assadi, M. M. and M. R. Jahangiri (2001) Textile wastewatertreatment by Aspergillus niger. Desalination 141: 1-6.
Table 5. GC-MS data of metabolites obtained after decolorization of Methyl orange
MetaboliteRt. time
(min)
Mw
(m/z)Mass spectrum
p-N,N'-dimethylamine phenyldiazine
25.08 149
Fig. 6. Proposed pathway for biodegradation of methyl orangeusing A. ochraceus NCIM-1146 laccase. [I]: Methyl orange; [II]:postulated p-hydroxybenzene sulfonic acid intermediate; and [III]:p-N,N'-dimethyl phenyldiazine intermediate.

Biochemical Characterization and Potential for Textile Dye Degradation of Blue Laccase from Aspergillus ochraceus NCIM-1146 703
4. Sharma, P., L. Singh, and N. Dilbaghi (2009) Response surfacemethodological approach for the decolorization of simulated dyeeffluent using Aspergillus fumigatus fresenius. J. Hazard. Mater.161: 1081-1086.
5. Corso, C. R. and A. C. M. Almeida (2009) Bioremediation ofdyes in textile effluents by Aspergillus oryzae. Microb. Ecol. 57:384-390.
6. Khelifi, E., L. Ayed, H. Bouallagui, Y. Touhami, and M. Hamdi(2009) Effect of nitrogen and carbon sources on Indigo andCongo red decolorization by Aspergillus alliaceus strain 121C. J.Hazard. Mater. 163: 1056-1062.
7. Saratale, G. D., S. D. Kalme, and S. P. Govindwar (2006) Decol-orization of textile dyes by Aspergillus ochraceus NCIM-1146.Ind. J. Biotechnol. 5: 407-410.
8. Sharma, P., R. Goel, and N. Capalash (2007) Bacterial laccases.World J. Microbiol. Biotechnol. 23: 823-832.
9. Claus, H. (2003) Laccases and their occurrence in prokaryotes.Arch. Microbiol. 179: 145-150.
10. Messerschmidt, A. and R. Huber (1990) The blue oxidases,ascorbate oxidase, laccase and ceruloplasmin. Modelling andstructural relationships. Eur. J. Biochem. 187: 341-352.
11. Telke, A. A., D. C. Kalyani, U. U. Jadhav, G. K. Parshetti, and S.P. Govindwar (2009) Purification and characterization of anextracellular laccase from a Pseudomonas sp. LBC1 and itsapplication for the removal of bisphenol A. J. Mol. Catal. B:Enzymatic 61: 252-260.
12. Kawai, S., T. Umezawa, and T. Higuchi (1988) Degradationmechanisms of phenolic β-1 lignin substructure model compoundsby laccase of Coriolus versicolor. Arch. Biochem. Biophys. 262:99-110.
13. Wesenberg, D., I. Kyriakides, and S. Agathos (2003) White-rotfungi and their enzymes for the treatment of industrial dye efflu-ents. Biotechnol. Advan. 22: 161-187.
14. Claus, H. (2004) Laccases: Structure, reactions, distribution.Micron 35: 93-96.
15. Palonen, H. and L. Viikari (2004) Role of oxidative enzymatictreatments on enzymatic hydrolysis of softwood. Biotechnol.Bioeng. 86: 550-557.
16. Murugesan, K., A. Dhamija, I. Nam, Y. Kim, and Y. Chang (2007)
Decolorization of Reactive black 5 by laccase: Optimization byresponse surface methodology. Dyes Pigments. 75: 176-184.
17. Baldrian, P. (2006) Fungal laccases-occurrence and properties.FEMS Microbiol. Rev. 30: 215-242.
18. Kurtz, M. and S. Champe (1982) Purification and characteri-zation of the Conidial Laccase of Aspergillus nidulans. J. Bacte-riol. 151: 1338-1345.
19. Scherer, M. and R. Fischer (1998) Purification and characteriza-tion of laccase II of Aspergillus nidulans. Arch. Microbiol. 170:78-84.
20. Parshetti, G. K., S. D. Kalme, S. S. Gomare, and S. P. Govindwar(2007) Biodegradation of Reactive blue-25 by Aspergillus ochra-ceus NCIM-1146. Bioresour. Technol. 98: 3638-3642.
21. Lowry, O. H., N. J. Rosebrough, A. L. Farr, and R. L. Randall(1951) Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193: 265-275.
22. Eisenman, H., M. Mues, S. Weber, S. Frases, S. Chaskes, G. Ger-fen, and A. Casadevall (2007) Cryptococcus neoformans laccasecatalyses melanin synthesis from both D- and L-DOPA. Micro-biol. 153: 3954-3962.
23. Nagai, M., T. Sato, H. Watanabe, K. Saito, M. Kawata, and H.Enei (2002) Purification and characterization of an extracellularlaccase from the edible mushroom Lentinula edodes and decol-orization of chemically different dyes. Appl. Microbiol. Biotech-nol. 60: 327-335.
24. Blaich, R. and K. Esser (1975) Function of enzymes in wooddestroying fungi. Arch. Microbiol. 103: 271-277.
25. Ko, E. M., Y. E. Leem, and H. T. Choi (2001) Purification, andcharacterization of laccase isozymes from the white-rot basidio-mycete Ganoderma lucidum. Appl. Microbiol. Biotechnol. 57:98-102.
26. Sadhasivam, S., S. Savitha, K. Swaminathan, and F. Lin (2008)Production, purification and characterization of mid-redox poten-tial laccase from a newly isolated Trichoderma harzianum WL1.Proc. Biochem. 43: 736-742.
27. Minussi, R. C., G. M. Pastorea, and N. Durán (2007) Laccaseinduction in fungi and laccase/N-OH mediator systems applied inpaper mill effluent. Bioresour. Technol. 98: 158-164.
Related Documents











