Plant Cell Physiol. 40(10): 1036-1045 (1999) JSPP © 1999 Biochemical and Morphological Features of Rice Cell Death Induced by Pseudomonas avenae Fang-Sik Che 1 - 4 , Megumi Iwano 1 , Noriko Tanaka 1 , Seiji Takayama 1 , Eiich Minami 2 , Naoto Shibuya 2 , Ikuo Kadota 3 and Akira Isogai' 1 Graduate School of Biological Sciences, Nara Institute of Science and Technology, 8916-5, Takayama Ikoma, Nara, 630-0101 Japan 2 National Institute of Agrobiological Resources, Tsukuba, Ibaraki, 305-8602 Japan 3 National Institute of Agro-Environmental Sciences, Tsukuba, Ibaraki, 305-8604 Japan Pseudomonas avenae is a Gram-negative phytopatho- genic bacterium that causes the symptom of a brown stripe in infected susceptible plants. The host range of P. avenae is wide among the monocotyledonous plants, however, in- dividual strains can infect only one or a few host species. A rice-incompatible strain, N1141, caused rapid cell death in sheath sections and in cultured rice cells. A rice-compatible strain, H8301, also induced cell death, however, this cell death in a compatible interaction was delayed compared to the cell death induced by the N1141 incompatible strain. Inoculation of N1141 strain induced expression of EL2 gene which is thought to be one of the defense-related gene. Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) of cultured rice cells showed that DNA cleavage occurred only in N1141-inoculated rice cells. N1141 strain caused cytoplasmic condensa- tion, shrinkage, and plasma membrane blebbing, all of which are important morphological characteristics of pro- grammed cell death (PCD). In contrast, H8301 strain in- oculated rice cells appeared to show weakening of the cell wall instead of cytoplasm condensation, shrinkage and membrane blebbing. These results suggest that the rapid cell death of rice induced by the incompatible strain is characterized as PCD. Key words: Hypersensitive response — Incompatible in- teraction — Programmed cell death — Pseudomonas ave- nae — TUNEL. In their natural environment, plants are in close con- tact with a vast number of bacterial species, but plant in- fection by bacteria occurs only in certain limited cases. The incompatible interaction often correlated with the elicita- tion of the defense response in plant cells. Several patho- gen-induced plant responses related to resistance have been described: generation of active oxygen species (oxidative burst) (Bolwell et al. 1995, Dixon et al. 1994), the hyper- Abbreviations: HR, hypersensitive response; PCD, pro- grammed cell death; PR, pathogenesis-related; TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling. 4 To whom correspondence should be addressed. Fax, +81-743- 72-5459; E-mail, [email protected] sensitive response (HR) (Klement 1982), induction of the phenylpropanoid pathway (Dixon et al. 1994), biosynthesis of phytoalexins (Gustine et al. 1990), and accumulation of pathogenesis-related (PR) proteins (Dixon et al. 1994). The HR is first described as rapid host cell death in resistant plants upon infection by the fungus Puccinia graminis (Stakman 1915). Recently, the term of the HR has been more frequently used,, and the HR is generally de- fined the rapid, localized cell death that is associated with the active defense of plants against many pathogens such as viruses, bacteria, fungi and nematodes (Keen 1990, Gabriel and Rolfe 1990, Dixon et al. 1994, Dangl et al. 1996, He 1996). The HR elicited by bacteria is readily observed as a collapse of the tissue, including cell death, if high concen- trations (>10 7 cells per ml) of a limited host-range patho- gen such as Pseudomonas syringae are infiltrated into the leaves of non host plants (Turner and Novacky 1974, Kle- ment et al. 1982). The capacities for both eliciting the HR in a non host plant and being pathogenic in a host appear linked. Therefore, the rapid cell death of HR is a hallmark of the resistance response of plants against pathogens. The HR requires active plant metabolism and depends on the activity of transcriptional and translational ma- chinery in the host plant (He et al. 1994). Certain muta- tions in maize and Arabidopsis cause the spontaneous ap- pearance of HR-like cell death (Walbot et al. 1983, Lamb and Dixon 1997). Furthermore, the spontaneous activation of hypersensitive cell death in the absence of pathogens has been reported in transgenic plants that express foreign genes (Culver and Dawson 1991, Hammond-Kosack et al. 1994, Mittler et al. 1995), and hypersensitive cell death is also induced by protein elicitors produced from pathogens such as AVR9, harpins, or PopAl (Wei et al. 1992, He et al. 1993, Arlat et al. 1994, Bauer et al. 1995, Hammond- Kosack et al. 1994). All these reports are favorable towards the existence of cell death genes encoded by the plant genome and regulated by pathogen-related signals. There- fore, there has been considerable speculation that hyper- sensitive cell death might be regulated by the activation of a specific cellular program that is under genetic control. However, the molecular mechanism of hypersensitive cell death and the role of hypersensitive cell death in disease resistance is not fully understood. Programmed cell death (PCD) is a physiological cell 1036 Downloaded from https://academic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Plant Cell Physiol. 40(10): 1036-1045 (1999)JSPP © 1999
Biochemical and Morphological Features of Rice Cell Death Induced byPseudomonas avenae
Fang-Sik Che1-4, Megumi Iwano1, Noriko Tanaka1, Seiji Takayama1, Eiich Minami2, NaotoShibuya2, Ikuo Kadota3 and Akira Isogai'1 Graduate School of Biological Sciences, Nara Institute of Science and Technology, 8916-5, Takayama Ikoma, Nara, 630-0101 Japan2 National Institute of Agrobiological Resources, Tsukuba, Ibaraki, 305-8602 Japan3 National Institute of Agro-Environmental Sciences, Tsukuba, Ibaraki, 305-8604 Japan
Pseudomonas avenae is a Gram-negative phytopatho-genic bacterium that causes the symptom of a brown stripein infected susceptible plants. The host range of P. avenaeis wide among the monocotyledonous plants, however, in-dividual strains can infect only one or a few host species. Arice-incompatible strain, N1141, caused rapid cell death insheath sections and in cultured rice cells. A rice-compatiblestrain, H8301, also induced cell death, however, this celldeath in a compatible interaction was delayed compared tothe cell death induced by the N1141 incompatible strain.Inoculation of N1141 strain induced expression of EL2gene which is thought to be one of the defense-related gene.Terminal deoxynucleotidyl transferase-mediated dUTPnick-end labeling (TUNEL) of cultured rice cells showedthat DNA cleavage occurred only in N1141-inoculatedrice cells. N1141 strain caused cytoplasmic condensa-tion, shrinkage, and plasma membrane blebbing, all ofwhich are important morphological characteristics of pro-grammed cell death (PCD). In contrast, H8301 strain in-oculated rice cells appeared to show weakening of the cellwall instead of cytoplasm condensation, shrinkage andmembrane blebbing. These results suggest that the rapidcell death of rice induced by the incompatible strain ischaracterized as PCD.
Key words: Hypersensitive response — Incompatible in-teraction — Programmed cell death — Pseudomonas ave-nae — TUNEL.
In their natural environment, plants are in close con-tact with a vast number of bacterial species, but plant in-fection by bacteria occurs only in certain limited cases. Theincompatible interaction often correlated with the elicita-tion of the defense response in plant cells. Several patho-gen-induced plant responses related to resistance have beendescribed: generation of active oxygen species (oxidativeburst) (Bolwell et al. 1995, Dixon et al. 1994), the hyper-
Abbreviations: HR, hypersensitive response; PCD, pro-grammed cell death; PR, pathogenesis-related; TUNEL, terminaldeoxynucleotidyl transferase-mediated dUTP nick-end labeling.4 To whom correspondence should be addressed. Fax, +81-743-72-5459; E-mail, [email protected]
sensitive response (HR) (Klement 1982), induction of thephenylpropanoid pathway (Dixon et al. 1994), biosynthesisof phytoalexins (Gustine et al. 1990), and accumulation ofpathogenesis-related (PR) proteins (Dixon et al. 1994).
The HR is first described as rapid host cell death inresistant plants upon infection by the fungus Pucciniagraminis (Stakman 1915). Recently, the term of the HR hasbeen more frequently used,, and the HR is generally de-fined the rapid, localized cell death that is associated withthe active defense of plants against many pathogens such asviruses, bacteria, fungi and nematodes (Keen 1990, Gabrieland Rolfe 1990, Dixon et al. 1994, Dangl et al. 1996, He1996). The HR elicited by bacteria is readily observed as acollapse of the tissue, including cell death, if high concen-trations (>107 cells per ml) of a limited host-range patho-gen such as Pseudomonas syringae are infiltrated into theleaves of non host plants (Turner and Novacky 1974, Kle-ment et al. 1982). The capacities for both eliciting the HRin a non host plant and being pathogenic in a host appearlinked. Therefore, the rapid cell death of HR is a hallmarkof the resistance response of plants against pathogens.
The HR requires active plant metabolism and dependson the activity of transcriptional and translational ma-chinery in the host plant (He et al. 1994). Certain muta-tions in maize and Arabidopsis cause the spontaneous ap-pearance of HR-like cell death (Walbot et al. 1983, Lamband Dixon 1997). Furthermore, the spontaneous activationof hypersensitive cell death in the absence of pathogenshas been reported in transgenic plants that express foreigngenes (Culver and Dawson 1991, Hammond-Kosack et al.1994, Mittler et al. 1995), and hypersensitive cell death isalso induced by protein elicitors produced from pathogenssuch as AVR9, harpins, or PopAl (Wei et al. 1992, He etal. 1993, Arlat et al. 1994, Bauer et al. 1995, Hammond-Kosack et al. 1994). All these reports are favorable towardsthe existence of cell death genes encoded by the plantgenome and regulated by pathogen-related signals. There-fore, there has been considerable speculation that hyper-sensitive cell death might be regulated by the activation ofa specific cellular program that is under genetic control.However, the molecular mechanism of hypersensitive celldeath and the role of hypersensitive cell death in diseaseresistance is not fully understood.
Programmed cell death (PCD) is a physiological cell
1036
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022

Rice cell death induced by Pseudomonas avenae 1037
death process involved in the selective elimination of un-wanted cells. In animal cells, the striking morphologicaland biochemical changes that characterize the most com-monly studied form of PCD, known as apoptosis, includecondensation and fragmentation of the nucleus, cleavageof nuclear DNA at specific sites, plasma membrane bleb-bing, and the fragmentation of the cell into apoptoticbodies (Earnshaw 1995, Cohen 1993, Saville et al. 1993).Recently, plant hypersensitive cell death has been shownto exhibit some of the morphological and biochemicalfeatures of animal cells undergoing PCD. These includefragmentation of nuclear DNA, induction of specific en-donucleases, and condensation and vacuolization of thecytoplasm (Ryerson and Heath 1996, Levine et al. 1996,Pennell and Lamb 1997). On the other hand, it has alsobeen reported that hypersensitive cell death in some casesdoes not clearly show the morphological cellular changesthat characterize mammalian apoptosis (Mittler et al. 1997,Heath 1998). Therefore, it is not clear whether HR-relatedcell death is mediated by a mechanism similar to PCD inanimal cells (Wang et al. 1996, Mittler et al. 1997, Heath1998).
Pseudomonas avenae is a Gram-negative phytopatho-genic bacterium that causes a seedling disease characterizedby the symptom of brown stripes in infected plants (Kadotaet al. 1991). The species P. avenae can infect a wide rangeof monocotyledonous plants including rice, oats, Italianmillet and maize. However, individual strains of the path-ogen infect only one or a few host species (Nishiyama et al.1979, Kadota et al. 1991, 1996). For example, strains iso-lated from rice, such as H8301 (MAFF 301505), can infectrice plants but not other monocotyledonous plants includ-ing finger millet (Kadota et al. 1991), whereas strain N1141(MAFF 301141) isolated from finger millet cannot infectrice plants (Nishiyama et al. 1979). The strict host range ofP. avenae applies to plant species rather than cultivars (e.g.the host range of rice strains such as H83O1 includes all ricecultivars). It has been demonstrated that some examples ofhost species specificity are also controlled by gene-for-genetype interactions, and the induction of the resistance re-sponse in plants is an important factor in the host deter-mination (Heath 1991, Sweigard et al. 1995). However, it isstill not known whether the non host strain of P. avenaeinduces the resistance responses including hypersensitivecell death in plants and whether induction of resistanceresponses is an important factor in host species determi-nation.
Suspension cultured cells have been widely used forinvestigations of the molecular mechanisms involvingplant-pathogen interaction because cultured cells have theadvantage of being relatively homogenous and aseptic.Experimental systems which combine cultured cells withpathogens or elicitors have provided excellent models forstudies of the molecular mechanism in hypersensitive cell
death. Therefore, we examined the relationship betweenthe induction of the defense response, including hypersen-sitive cell death and host species specificity of P. avenae,using cultured rice cells and incompatible or compatiblestrain of P. avenae. We describe here how the P. avenaestrain N1141, which is incompatible with rice, elicitedthe rapid cell death in rice cultured cells accompanied bynuclear DNA cleavage. In addition, we also demonstratethat incompatible strain-infected rice shows a classic setof morphological changes, including cell condensation,plasma membrane shrinkage and blebbing, all of whichthat are important morphological characteristics of pro-grammed cell death.
Materials and Methods
Bacterial strains and cultures—Pseudomonas avenae H8301(MAFF 301505) isolated from rice and strain N1141 (MAFF301141) isolated from finger millet were used in this study (Kadotaet al. 1996). Each strain of P. avenae was maintained on Pseudo-monas F agar plates (Difco, U.S.A.) at 30°C.
Inoculation test—For the inoculation of P. avenae, rice(Oriza sativa L. cv. kinuhikari) was grown for 3 to 4 weeks in anatural-light phytotron at 30/25°C (day/night). One day beforeinoculation, the plants were placed in a growth chamber with a 16h day (200 fiE m"2 s"1 at 30°C) and 8 h night (25°C) cycle and65% relative humidity. The bacteria were suspended in sterilizeddistilled water (lO'cfumP1) and inoculated using the single-needle method (Kadota et al. 1996). A 1 /xl (106 cfu) drop of thebacterial suspension medium was formed on the end of a needleand then the sheath was pricked at a point 2 cm above the soillevel. Control plants were mock-inoculated with sterilized water.Inoculated seedlings were maintained in a growth chamber underthe same conditions. Seven days after inoculation, the pathoge-nicity of each strain was determined by assessing the brown stripedevelopment around the inoculation site.
For determining the growth of P. avenae in rice, seedlingswere inoculated in the same manner. The growth of the H8301and Nl 141 strains was assessed in inoculated plants up to 6 d afterinoculation. Five sets of randomly selected rice plants were har-vested, rinsed thoroughly in sterile water, and homogenized in 10mM MgSO4. Dilution of the homogenate were plated onto Pseu-domonas F agar. After incubation for 48 h at 30°C the number ofcolony-forming units (cfu) was determined.
Cell death detection in leaf sheath sections—Rice seedlingswere grown for 2 weeks in a growth chamber as described above.Leaf sheaths of the seedlings were embedded in agar (0.7% w/v)and then cut into 70 micron section with a Microslicer (D.S.K,Osaka, Japan). Rice stem sections were incubated at 37°C with100/il of 50 mM HEPES-KOH (pH 7.2) with each bacterial strain(lO'cfuml"1) or without bacteria (control). After 2h, the stemsections were stained with 0.1% Evans blue (Turner and Novacky1974) containing 50 mM HEPES-KOH (pH 7.2) for 15min thenwashed extensively with 50 mM HEPES-KOH (pH 7.2) to removeexcess dye and observed by microscopy (Axiophot 2, Carl Zeiss,Germany).
Cell death detection in cultured rice cells—Cell suspensioncultures of rice (Oriza sativa L. cv. Kinmaze), provided by Pro-fessor K. Shimamoto (NAIST, Japan), were grown at 30°C un-der light (Baba 1986). The cells were diluted in fresh medium every
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022
103f Rice cell death induced by Pseudomonas avenge
1 d, and experiments were performed 4 d after transfer. The cul-tured cells were incubated with bacteria (108 cfu ml"') at 30°C atdifferent time lengths after inoculation. Cultured cells werereplaced in 24-well tissue culture plates and incubated for 15 minwith 0.05% Evans blue containing 50 mM HEPES-KOH (pH 7.2),then washed extensively to remove the excess dye. The dye incor-porated in dead cells was extracted by.50% methanol with 1%SDS for 12 h at room temperature, with absorbance at 595 ranused for quantification (Turner and Novacky 1974). A controlexperiment was performed by adding distilled water, which hadthe same volume as that of the bacterial suspension added to therice culture medium.
RNA isolation and gel blot analysis—Cultured rice cells wereinoculated with each bacterial strains (or without bacteria, forcontrol) for cell death detection. Cultured rice cells were groundin liquid nitrogen and total RNA was extracted with ATA (aurin-tricarbosylic acid) (Gonzalez et al. 1980). RNAs (15yUg) wereelectrophoresed on a formaldehyde denaturing 1 % agarose gel in1 x MOPS buffer (20 mM MOPS-KOH, pH 7.0, 5 mM sodiumacetate, and 1 mM EDTA) and blotted onto Hybond-N mem-brane (Amersham), according to standard protocols. Full-lengthcDNA fragments of the EL2 gene were labeled with [32P]dCTPand used as hybridization probes.
TUNEL procedure—-For the TUNEL procedure, culturedrice cells were inoculated with bacterial strains as for the cell deathdetection. Small pieces of inoculated cultured cells were washedthree times with PBS (pH 7.4) and fixed in 4% (v/v) paraform-aldehyde in PBS at 25°C overnight. The cells were washed threetimes with PBS and digested with Proteinase K (10^g ml"1) con-taining 10 mM Tris-HCl (pH 7.5) at 37°C for 30 min and washedthree times with PBS. The cells were subjected to TUNEL usinga terminal deoxynucleotidyl transferase (TdT)-mediated dUTP-based in situ death detection kit (Boeringer Mannheim, Germany)at 37°C for 1 h. After washing with PBS, 4',6-diamidino-2-phyn-ylindole (DAPI) was added to the cells (1 / igmr 1 ) in 50 mMNaCl, 5 mM EDTA and 10 mM Tris-HCl (pH 7.4) and observedby fluorescence microscopy (Carl Zeiss, Germany) using a No. 2filter for DAPI and a NO. 10 filter for TUNEL (Ryerson andHeath 1996). To examine the timing when the DNA cleavage oc-curred after inoculation, quantification of TUNEL-positive nucleiwas performed. All nuclei were detected using DAPI staining at 1/ i g m r 1 on Carl Zeiss Axiophot 2 fluorescence microscopy (CarlZeiss, Germany). The number of TUNEL-positive nuclei andDAPI staining nuclei was determined by counting nuclei withintwenty individual fields. Each determination was done with atleast 3,000 nuclei in each of three independent experiments foreach bacterial infection.
Transmission electron microscopy (TEM)—Cultured cellsinoculated with H83O1 and N1141 strains were fixed with 2.5%glutaraldehyde containing 0.05 M sodium cacodylate (pH 7.0) for2 h at 4°C and then washed with 0.05 M sodium cacodylate (pH7.0). The cells were post-fixed with 1% OsO4 for 2 h on ice. Afterwashing with water, the cells were dehydrated in a graded ace-tone series. The dehydrated cells were embedded in Spurr's resin(TAAB Lab., Aldermaston, U.K.) and sectioned. Ultra-thin sec-tions were stained with lead (Reynolds 1963) and observed undera transmission electron microscope (H-7100, Hitachi, Tokyo,Japan) at an accelerating voltage of 75 kV.
Results
Compatibility of P. avenae H8301 and N1141 strainson rice—When rice plants were inoculated with strain
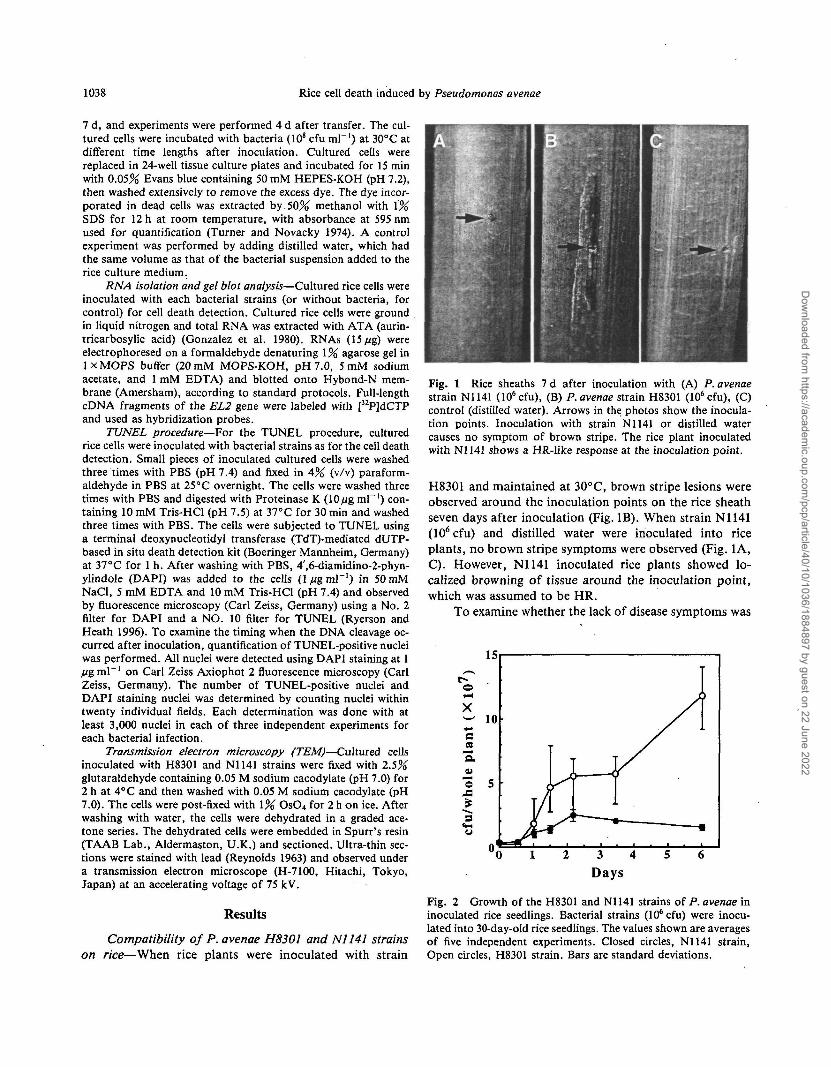
Fig. 1 Rice sheaths 7 d after inoculation with (A) P. avenaestrain N1141 (106cfu), (B) P. avenae strain H8301 (106cfu), (C)control (distilled water). Arrows in the photos show the inocula-tion points. Inoculation with strain N1141 or distilled watercauses no symptom of brown stripe. The rice plant inoculatedwith N1141 shows a HR-like response at the inoculation point.
H8301 and maintained at 30°C, brown stripe lesions wereobserved around the inoculation points on the rice sheathseven days after inoculation (Fig. IB). When strain N1141(106 cfu) and distilled water were inoculated into riceplants, no brown stripe symptoms were observed (Fig. 1A,C). However, N1141 inoculated rice plants showed lo-calized browning of tissue around the inoculation point,which was assumed to be HR.
To examine whether the lack of disease symptoms was
Fig. 2 Growth of the H83O1 and NU41 strains of P. avenae ininoculated rice seedlings. Bacterial strains (106cfu) were inocu-lated into 30-day-old rice seedlings. The values shown are averagesof five independent experiments. Closed circles, N1141 strain,Open circles, H83O1 strain. Bars are standard deviations.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022
Rice cell death induced by Pseudomonas avenae
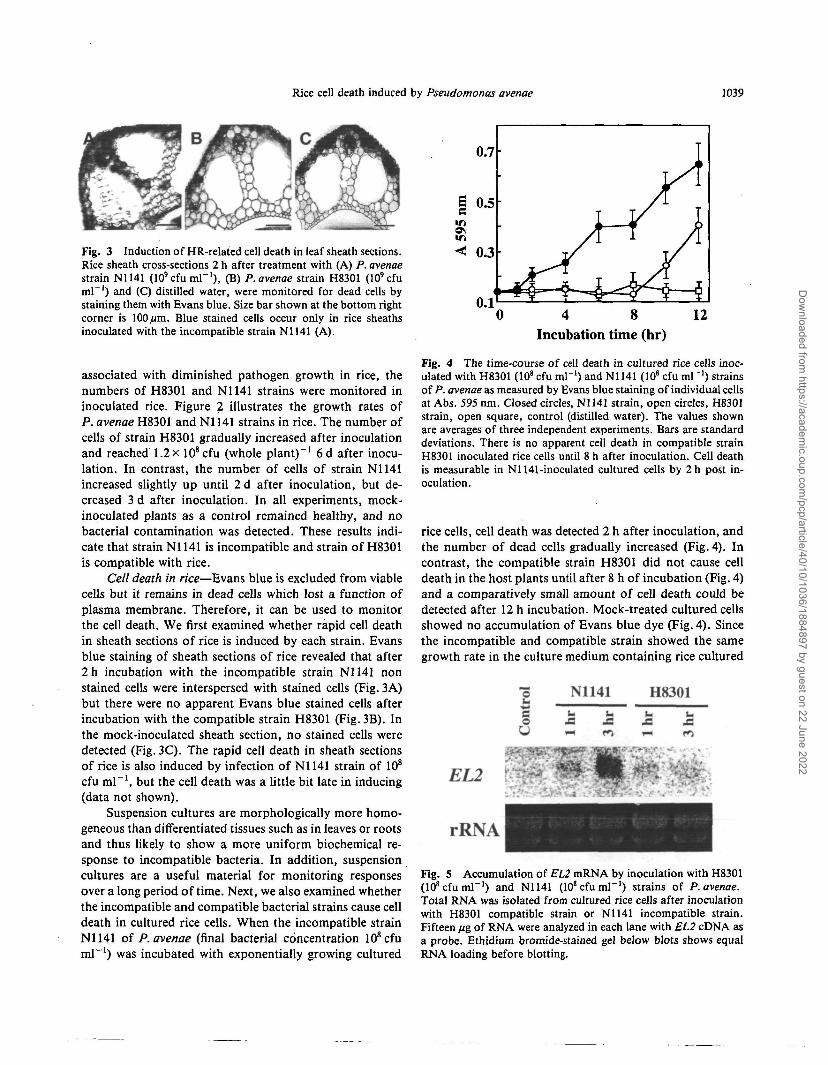
Fig. 3 Induction of HR-related cell death in leaf sheath sections.Rice sheath cross-sections 2 h after treatment with (A) P. avenaestrain N1141 (lO'cfumP1), (B) P. avenae strain H8301 (10'cfuml"1) and (C) distilled water, were monitored for dead cells bystaining them with Evans blue. Size bar shown at the bottom rightcorner is 100/urn. Blue stained cells occur only in rice sheathsinoculated with the incompatible strain N1141 (A).
associated with diminished pathogen growth in rice, thenumbers of H83O1 and N1141 strains were monitored ininoculated rice. Figure 2 illustrates the growth rates ofP. avenae H8301 and N1141 strains in rice. The number ofcells of strain H83O1 gradually increased after inoculationand reached 1.2 x 108 cfu (whole plant)"1 6 d after inocu-lation. In contrast, the number of cells of strain N1141increased slightly up until 2 d after inoculation, but de-creased 3 d after inoculation. In all experiments, mock-inoculated plants as a control remained healthy, and nobacterial contamination was detected. These results indi-cate that strain N1141 is incompatible and strain of H8301is compatible with rice.
Cell death in rice—Evans blue is excluded from viablecells but it remains in dead cells which lost a function ofplasma membrane. Therefore, it can be used to monitorthe cell death. We first examined whether rapid cell deathin sheath sections of rice is induced by each strain. Evansblue staining of sheath sections of rice revealed that after2h incubation with the incompatible strain N1141 nonstained cells were interspersed with stained cells (Fig. 3A)but there were no apparent Evans blue stained cells afterincubation with the compatible strain H83O1 (Fig. 3B). Inthe mock-inoculated sheath section, no stained cells weredetected (Fig. 3C). The rapid cell death in sheath sectionsof rice is also induced by infection of N1141 strain of 108
cfu ml"1, but the cell death was a little bit late in inducing(data not shown).
Suspension cultures are morphologically more homo-geneous than differentiated tissues such as in leaves or rootsand thus likely to show a more uniform biochemical re-sponse to incompatible bacteria. In addition, suspensioncultures are a useful material for monitoring responsesover a long period of time. Next, we also examined whetherthe incompatible and compatible bacterial strains cause celldeath in cultured rice cells. When the incompatible strainN1141 of P. avenae (final bacterial concentration 108cfuml"1) was incubated with exponentially growing cultured
£c
4 8Incubation time (hr)
12
Fig. 4 The time-course of cell death in cultured rice cells inoc-ulated with H8301 (108 cfu ml"1) and N1141 (108 cfu ml"1) strainsof P. avenae as measured by Evans blue staining of individual cellsat Abs. 595 nm. Closed circles, N1141 strain, open circles, H8301strain, open square, control (distilled water). The values shownare averages of three independent experiments. Bars are standarddeviations. There is no apparent cell death in compatible strainH8301 inoculated rice cells until 8 h after inoculation. Cell deathis measurable in Nl 141-inoculated cultured cells by 2 h post in-oculation.
rice cells, cell death was detected 2 h after inoculation, andthe number of dead cells gradually increased (Fig. 4). Incontrast, the compatible strain H83O1 did not cause celldeath in the host plants until after 8 h of incubation (Fig. 4)and a comparatively small amount of cell death could bedetected after 12 h incubation. Mock-treated cultured cellsshowed no accumulation of Evans blue dye (Fig. 4). Sincethe incompatible and compatible strain showed the samegrowth rate in the culture medium containing rice cultured
N1141 H8301
EL2
rRNA
Fig. 5 Accumulation of EL2 mRNA by inoculation with H83O1(108cfuml"') and N1141 (108cfuml"') strains of P. avenae.Total RNA was isolated from cultured rice cells after inoculationwith H8301 compatible strain or N1141 incompatible strain.Fifteen ftg of RNA were analyzed in each lane with EL2 cDNA asa probe. Ethidium bromide-stained gel below blots shows equalRNA loading before blotting.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022
1040 Rice cell death induced by Pseudomonas avenae
cells during this bioassay (data not shown), the rapid celldeath in N1141 inoculated cultured rice cells is caused bythe results that rice cultured cells recognized the incom-patible strain of N1141.
Accumulation of EL2 mRNA in cultured rice cells in-oculated with P. avenae—The EL2 gene is expressed withN-acetylchitoheptaose which induce a set of defense re-actions in cultured rice cells within 15 min (Minami et al.
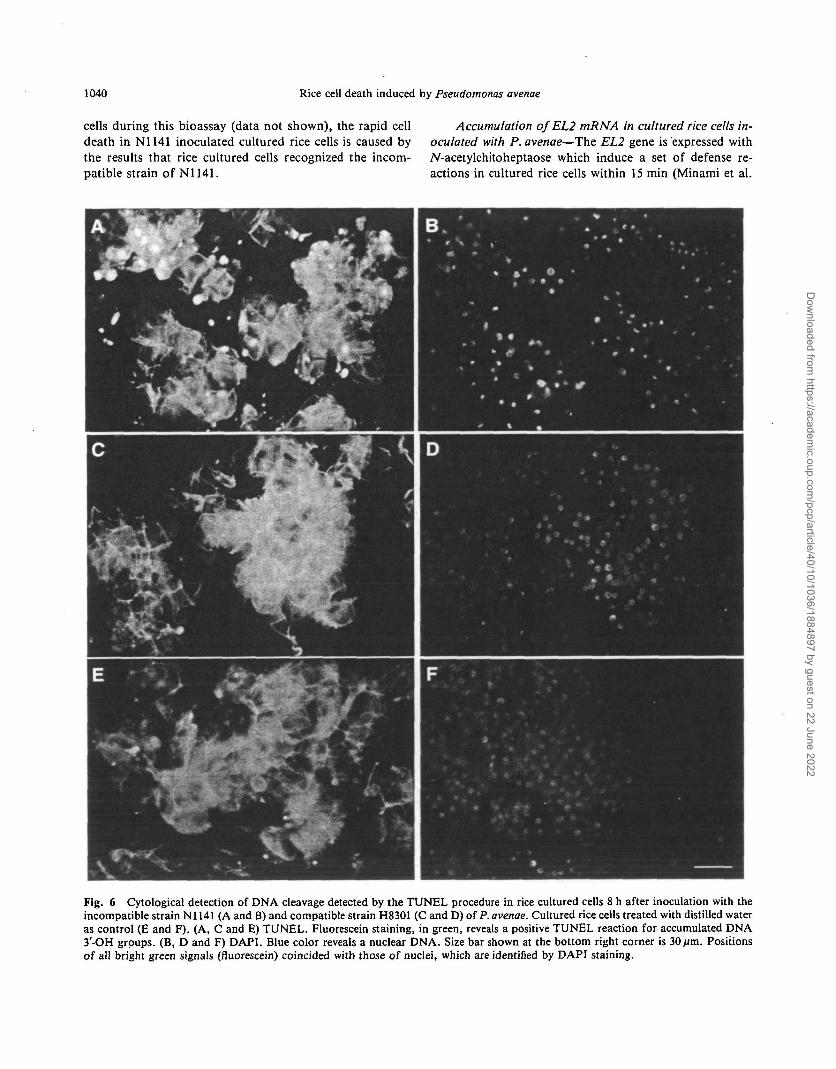
Fig. 6 Cytological detection of DNA cleavage detected by the TUNEL procedure in rice cultured cells 8 h after inoculation with theincompatible strain Nl 141 (A and B) and compatible strain H8301 (C and D) of P. avenae. Cultured rice cells treated with distilled wateras control (E and F). (A, C and E) TUNEL. Fluorescein staining, in green, reveals a positive TUNEL reaction for accumulated DNA3-OH groups. (B, D and F) DAPI. Blue color reveals a nuclear DNA. Size bar shown at the bottom right corner is 30fim. Positionsof all bright green signals (fluorescein) coincided with those of nuclei, which are identified by DAPI staining.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022
Rice cell death induced by Pseudomonas avenae 1041
1996, He et al. 1998). EL2 gene expression correlates withthe rapid expression of other plant defense-related genesuch as phenylalanine ammonia-lyase (PAL), chitinase and/M,3-glucanase. In addition, EL2 gene expression parallelsthe accumulation of a diterpenoid phytoalexin, momilac-tone A (Yamada et al. 1993). Therefore, EL2 gene is thoughtto be one of the defense-related genes such as the gene forpathogenesis-related (PR) proteins (He et al. 1998).
The accumulation of some PR proteins is known to beinduced more rapidly after inoculation in an incompatiblethan in a compatible interaction (Gonstabel and Brisson1992, Midoh and Iwata 1996). Such a differential pattern ofaccumulation is considered to be important in disease re-sistance. Therefore, we examined the time course of the
' accumulation of EL2 mRNA in cultured rice cells afterinoculations with the N1141 and H8301 strains of P. ave-nae. As shown in Fig. 5, the accumulation of the EL2mRNA is induced 3 h after inoculation in an incompatiblestrain, Nl 141, whereas it in H83O1 inoculated cultured cellsis not detected at 3 h after inoculation. The EL2 mRNA inH8301 inoculated cells does not accumulate even at 6 hafter inoculation (data not shown). These results indicatethat the incompatible strain N1141 can induce the expres-sion of the defense-related gene in rice cultured cells, butthe compatible strain H8301 cannot.
Cytologically detectable DNA cleavage in cultured rice, cells inoculated with P. avenae—Recently it has been re-ported that a recurrent feature of the HR is the cleavage ofDNA at specific chromosomal sites by DNA endonucleases(Ryerson and Heath 1996, Wang et al. 1996). Terminaldeoxynucleotidyl transferase-mediated dUTP nick-end la-beling (TUNEL) can be used to quantify the accumulationof DNA 3'-OH groups caused by DNA fragmentation andbreakage (Gavrieli et al. 1992, McCabe et al. 1997). Weperformed TUNEL on cultured rice cells to determinewhether the DNA is cleaved during cell death induced withP. avenae.
The cultured rice cells infected with the incompatiblestrain N1141 had many fluorescein-derived bright greenfluorescence signals at 8 h after inoculation, as detected bythe TUNEL procedure (Fig. 6A). The positions of all brightgreen signals coincided with those of the nuclei, which areidentified by 4',6-diamidino-2-phynylindole (DAPI) stain-ing (Fig. 6B). On the other hand, the cultured rice cells in-fected with the compatible strain H8301 and without bac-teria (control) also had some TUNEL positive nuclei, butthe number of the positive nuclei in H8301 infected ricecells and water-treated control cells was remarkably smallerthan that in N1141 infected cells (Fig.6C, D, E, F). Nofluorescein-derived fluorescence was observed when termi-nal deoxynucleotidyl transferase or the dUTP substratewas omitted from the reaction (data not shown).
Next we examined the timing when the DNA cleavageoccurred after inoculation in the cultured rice cells. Figure
7 shows the percentage of TUNEL-positive nuclei in eachbacterial infected cells at several point after infection.In the cultured rice cells inoculated by the incompatiblestrain N1141, the number of TUNEL-positive nuclei didnot increase until 4 h after inoculation. The percentage ofTUNEL-positive nuclei, however, increased slightly at 6 hafter inoculation, and approximately 18% of nuclei hadFITC-derived fluorescence at 10 and 12 h after inoculation.In contrast, the percentage of TUNEL-positive nuclei inrice cells inoculated with the compatible strain H8301 didnot increase until 10 h after inoculation, and only a smallincrease in the percentage of TUNEL-positive nuclei wasdetected at 12 h after inoculation. Approximately 1.6%of TUNEL-positive nuclei were detected in water-treatedcontrol cells up until 12 h after treatment. These data sug-gest that P. avenae strain Nl 141 which is incompatible withrice, specifically induced DNA cleavage in the nucleus, butthe compatible strain H8301 did not.
Morphological changes in cultured rice cells inocu-lated with P. avenae—Because it has been reported thatprogrammed cell death (PCD) in plants often involvesmorphological changes, including condensation and shrink-age of-the cytoplasm as seen in animal apoptosis (Levineet al. 1996, McCabe et al. 1997), we used transmissionelectron microscopy (TEM) to identify whether infectingrice cultured cells with P. avenae triggered morphologicalchanges such as cell shrinkage and condensation. The cul-tured rice cells were incubated with H8301 and N1141strains, harvested 12 h after incubation, and fixed and em-bedded with resin. When these inoculated rice cells were
usc
Hiooa
pH
[22 N1141
• H8301
2 4 6 8 10 12Time after inoculation
Fig. 7 Time-course of percentage of TUNEL positive nuclei inrice cultured cells inoculated with N1141 (hatched bars) andH8301 (white bars) strains of P. avenae. The percentage ofTUNEL-positive nuclei was determined by counting nuclei withintwenty individual fields. Each determination was done with atleast 3,000 nuclei in each of three independent experiments foreach bacterial infection.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022
1042 Rice cell death induced by Pseudomonas avenae
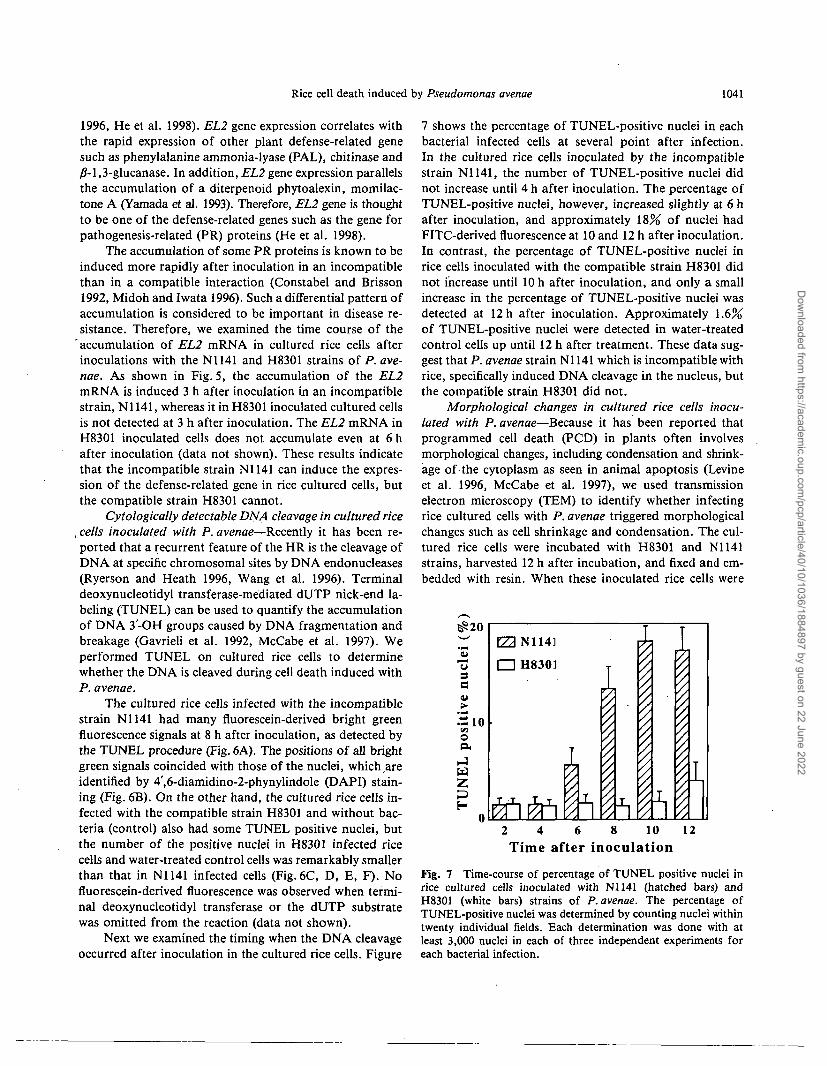
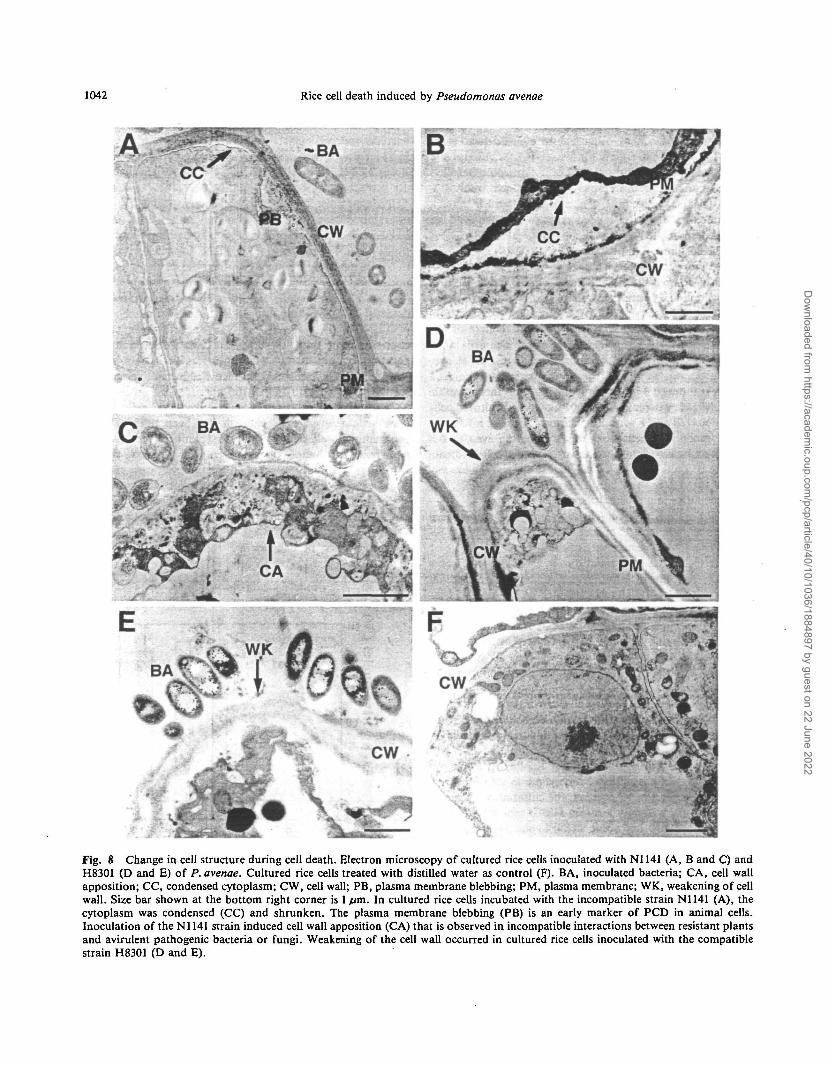
Fig. 8 Change in cell structure during cell death. Electron microscopy of cultured rice cells inoculated with N1141 (A, B and C) andH8301 (D and E) of P. avenae. Cultured rice cells treated with distilled water as control (F). BA, inoculated bacteria; CA, cell wallapposition; CC, condensed cytoplasm; CW, cell wall; PB, plasma membrane blebbing; PM, plasma membrane; WK, weakening of cellwall. Size bar shown at the bottom right corner is 1 /im. In cultured rice cells incubated with the incompatible strain N1141 (A), thecytoplasm was condensed (CC) and shrunken. The plasma membrane blebbing (PB) is an early marker of PCD in animal cells.Inoculation of the N1141 strain induced cell wall apposition (CA) that is observed in incompatible interactions between resistant plantsand avirulent pathogenic bacteria or fungi. Weakening of the cell wall occurred in cultured rice cells inoculated with the compatiblestrain H8301 (D and E).
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022

Rice cell death induced by Pseudomonas avenae 1043
viewed by TEM, bacteria were observed in contact with thecultured rice cells (Fig. 8A, C, D, E). However, the cyto-plasm in the cultured rice cells incubated with N1141 wascondensed and shrunken, but did not cause detectable lysisof either the cell wall or the plasma membrane (Fig. 8A, B).The cultured cells incubated with N1141 also showed wide-spread blebbing (Fig. 8A), disengagement and convolutionof the plasma membrane followed by condensation, shrink-age and shriveling of the cytoplasm (Fig. 8A, B). More-over, inoculation of the N1141 strain induced cell wallapposition (Fig.8C) that is observed in incompatible in-teractions between resistant plants and avirulent patho-genic bacteria or fungi (Koga 1994, Bestwick et al. 1995).These morphological changes could be observed in largenumbers of N1141 infected rice cells, and both condensa-tion and shrinkage in these cells were remarkably observedin the cells which had contact with the bacteria. In con-trast, inoculating rice cells with strain H83O1 appeared toinduce weakening of the cell wall around the bacteria-contacted area (Fig. 8D, E) rather than cytoplasm conden-sation, shrinkage and membrane blebbing. The region ofdegradation in H83O1 inoculated cells included not only themiddle lamella but also the primary or secondary cell wall(Fig. 8D, E). These distinctive morphological changes inN1141 or H83O1 infected rice cultured cells were not ob-served in water-treated control cells (Fig. 8F). Our datasuggest that the distinctive morphological characteristics ofPCD are observed only in N1141 inoculated cultured cells.
Discussion
The potential to elicit necrosis and the ability to mul-tiply in plant tissues are both important elements to beconsidered in determining the compatibility of pathogenicbacteria with certain plant hosts (Klement 1982). Our in-oculation tests showed that strain H8301 of P. avenae hadthe ability to multiply in rice and cause disease, but strainN1141 did not grow in the plant and caused no disease.However, some visible tissue damage which would be char-acterized as HR was present at the point of inoculation ofstrain N1141. These results indicate that strain H83O1 iscompatible and strain N1141 is incompatible with rice. Assuggested by the different response patterns outlined inFig. 2, the fate of rice plant-P. avenae interactions may bedetermined very early after inoculation.
Inoculation of the rice sheath with the incompatiblestrain N1141 caused cell death 2 h after inoculation as as-sayed by Evans blue staining, while the compatible strainH83O1 did not cause cell death in the host plant during thisperiod (Fig. 3). It has been reported that avirulent strainsof phytopathogenic bacteria induced HR-related cell deathin Red Mexican bean detectable with Evans blue staining.The incompatible interaction between Red Mexican beanleaves and the avirulent bacteria Pseudomonassyringae pv.
phaseolicola showed that the dead cells were interspersedwith many living cells (Meier et al. 1993). This finding isin good agreement with our result that rapid cell death inthe rice sheath was induced by the incompatible strain ofP. avenae and that the dead cells were interspersed withmany living cells (Fig. 3). Rapid cell death caused by in-compatible strains was also observed in cultured rice cells.The Nl 141 strain caused rapid cell death in cultured cells 2h after incubation, and the number of dead cells graduallyincreased (Fig. 4). In this study, we also showed that in-oculation of incompatible strain elicits the accumulation ofEL2 mRNA 3 h after inoculation. The EL2 gene is isolatedby cDNA subtraction as a gene responsive within 6 min toTV-acetylchitoheptaose, a potent specific elicitor for phyto-alexin biosynthesis. Because EL2 gene expression corre-lates with the expression of other plant defense-relatedgenes such as phenylalanine ammonia-lyase (PAL), chiti-nase and /M,3-glucanase, the EL2 gene is thought to bea gene related to defense response (Minami et al. 1996).These data indicated that inoculation with the incompati-ble strain N1141 causes the defense response in culturedrice cells and the rapid cell death induced by the N1141strain could be characterized as a hypersensitive cell death.
Hypersensitive cell death is believed to be an activelyprogrammed process in plant cells. Nuclear DNA cleavage,in particular, appears to be a hallmark of animal pro-grammed cell death (PCD), known as apoptosis (Nicholsonet al. 1995). It has been reported that plant DNA cleavageis observed during the hypersensitive cell death triggered bya virus (Mittler et al. 1997), bacteria (Levine et al. 1996,Mittler et al. 1997) and a fungus (Ryerson and Heath 1996).Fragmentation of DNA during HR can be detected in situby reagents that react with the exposed 3'-OH groups onthe nucleosomal units. The assay procedure involves endlabeling the DNA fragments using terminal deoxynucle-otidyl transferase (TdT) with UTP conjugated to a detect-able marker (TUNEL). Inoculation of the incompatiblestrain Nl 141 in cultured rice cells also caused nuclear DNAcleavage as detected by the TUNEL procedure.
Several evidence suggest that death of plant cells dur-ing the HR results from the activation of PCD pathway(Mittler et al. 1997). The recognition of PCD in mam-malian cells has been aided by its usual association witha stereotypical set of morphological changes such as cellcondensation, plasma membrane shrinkage and blebbing(Cohen 1993, Chinnaiyan and Dixit 1996). It has beenreported that the morphological cellular changes that char-acterize mammalian PCD are observed in some cases ofplant HR (Levine et al. 1996, McCabe et al. 1997). In-oculation of Pseudomonas syringae pv. glycinea carryingavrA caused morphological changes in cultured soybeancells including condensation and shrinkage of the cyto-plasm, as well as plasma membrane blebbing (Levine et al.1996). In our present study, transmission electron micros-
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022

1044 Rice cell death induced by Pseudomonas avenae
copy of cultured rice cells showed that inoculation withthe incompatible strain of P. avenae induced shrinkagemanifest by extensive disengagement of the dead proto-plast from the cell wall, condensation of the cytoplasm,and blebbing of the plasma membrane. Moreover, nuclearDNA cleavage clearly occurred in cultured rice cells in-oculated with the incompatible strain N1141. These datasuggest that the rapid cell death induced by the incom-patible strain N1141 is hypersensitive cell death accompa-nied with some of the characteristics of animal PCD. Onthe other hand, the condensation of nucleus which is im-portant characteristic of the apoptosis in animal cell, wasnot observed in N1141 inoculated cells. The hypersensitivecell death which is caused by N1141 inoculation would beinduced by different pathway from animal apoptosis.
Programmed cell death often involves the activationof DNA endonucleases and the appearance of oligonu-cleosome-sized DNA fragments by agarose gel electropho-resis (Estman et al. 1994). However, we could not obtainDNA laddering in rice cultured cells during the incom-patible interaction by agarose gel electrophoresis (datanot shown). It has been shown that oligonucleosome-sizedfragmentation dose not always occur, and PCD in manyanimal and plant cells does not involve the fragmentationof DNA into oligonucleosome-sized pieces (Oberhammeret al. 1993, Schwartz et al. 1993). Furthermore, it has beenshown that in the case of plant PCD in cultured cells con-taining only a small number of dead cells, DNA ladderingcould not be detected by agarose gel electrophoresis (Mc-Cabe et al. 1997). In addition, it has been reported thatoligonucleosomal fragments occurred only with the fungus(Heath 1998). However, there is not enough evidence todecide the reason why DNA laddering was not detected incultured rice cells inoculated with the incompatible strainN1141 of P. avenae.
Cell death in cultured rice cells is induced by not onlythe incompatible strain N1141 but also the compatiblestrain H8301. However, some differences exist in bothtypes of cell death induced by the N1141 and H83O1strains. The cell death induced by the N1141 strain occurs2 h after inoculation, but the cell death induced by theH8301 strain only occurs 10 h after inoculation. In addi-tion, significant TUNEL positive nuclei were not observedin cultured rice cells inoculated with the H8301 strainwithin 10 h after inoculation. TEM observations of cul-tured cells inoculated with H83O1 showed weakening ofthe cell wall rather than the characteristic morphologicalchanges for PCD. Cell wall weakening occurred not only inthe middle lamella, which is the intercellular cement, butalso in the secondary cell wall, which is proximai to thecytosol. These cell wall changes could be observed in largenumbers of cells and the cell wall weakening was clearlyobserved in the cell wall surface which is contacted with thebacteria of H8301. Degradation of the cell wall is caused by
many types of plant pathogenic bacteria and results in li-quefaction of the pectic substances that hold plant cellstogether. The plant cell eventually succumbs to necroticcell death by pathogen attack (Agrios 1997). The cell deathinduced by H8301 strain is not PCD, and this cell deathwould be resulted in catastrophe by attack of compatiblestrain H8301.
In some cases of incompatible interaction, plant cellsdying due to the HR do not clearly show the morphologicalcellular changes that characterize mammalian apoptosis. Ithas been reported that inoculation of Pseudomonas syrin-gae pv. phaseolicola which causes hypersensitive cell deathafter infiltration of suspension cultured lettuce cells inducescell wall alterations including deposition of callose but notshrinkage of plasma membrane and condensation of cyto-plasm (Bestwick et al. 1995, Heath 1998). The reason whythe cell death induced by incompatible pathogens is pro-vided in two type, one is accompanied with a morphologi-cal changes such as condensation, shrinkage and plasmamembrane blebbing and other one is not accompanied withthe morphological change is still not clear. Cell death is anessential part of plant defense and development (Dangl etal. 1996, Pennell and Lamb 1997), and there are likely to beseveral cell death pathways operational in plants. We be-lieve that the examination of the molecular mechanism ofthe cell death induced by incompatible strain of P. avenaewill provide new information about the signaling pathwayof plant PCD.
We thank Professor Ko Shimamoto for providing us withcultured rice cells and for helpful comments and suggestions. Weare also grateful to koichiro Tsuge, Risa Okimatsu and TomokoYoshida for cell death assay, and Yuka Wakasaki for TUNEL andRNA gel blot analysis. This work was supported by the "Researchfor the Future" Program of the Japan Society for the Promotionof Science (JSPS-RFTF96R16001).
References
Agrios, G.N. (1997) How pathogens attack plants. In Plant Pathology, 4thed. Edited by Agrios, G.N. pp. 66-72. Academic Press, San Diego.
Arlat, M., van Gijsegem, F., Heut, J.C., Pernollet, J.C. and Boucher,C.A. (1994) PopAl, a protein which induces a hypersensitivity-like re-sponse on specific petunia genotypes, is secreted via the Hrp pathway ofPseudomonas solariacearum. EMBO J. 13: 543-553.
Baba, A., Hasezawa, S. and Syono, K. (1986) Cultivation of rice proto-plasts and their transformation mediated by Agrobacterium sphero-plasts. Plant Cell Physiol. 27: 463-472.
Bauer, D.W., Wei, Z.M., Beer, S.V. and Collmer, A. (1995) Erwiniachrysanthemi EC 16 harpined,: an elicitor of the hypersensitive responsethat contributes to soft-rot pathogenesis. Mol. Plant-Microbe Interact.8: 484-491.
Bestwick, C.S., Bennett, M.H. and Mansfield, J.W. (1995) Hrp mutant ofPseudomonas syringae pv. phaseolicola induced cell wall alterations butnot membrane damage leading to the hypersensitive reaction in lettuce.Plant Physiol. 108:503-516.
Bolwell, G.P., Butt, V.S., Davies, D.R. and Zimmerlin, A. (1995) Theorigin of the oxidative burst in plants. Free Rad. Res. 23: 517-532.
Chinnaiyan, A.M. and Dixit, V.M. (1996) The cell-death machine. Curr.Biol. 6: 555-562.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022

Rice cell death induced by Pseudomonas avenae 1045
Cohen, J.J. (1993) Apoptosis. Immunol. Today 14: 126-130.Constabel, C.P. and Brisson, N. (1992) The defense-related STH-2 gene
product of potato shows race-specific accumulation after inoculationwith low concentrations of Phytophtora infestans zoospores. Planta188: 289-295.
Culver, N.J. and Dawson, W.O. (1991) Tobacco mosaic virus elicitor coatprotein genes produce a hypersensitive phenotype in transgenic Nico-tiana sylvestris plants. Mol. Plant-Microbe Interact. 4: 458-463.
Dangl, J.L., Dietrich, R.A. and Richberg, M.H. (1996) Death don't haveno mercy: cell death programs in plant-microbe interactions. Plant Cell8: 1793-1807.
Dixon, R.A., Harrison, M.J. and Lamb, C.J. (1994) Early events in theactivation of plant defense responses. Annu. Rev. Phytopathol. 32:479-501.
Earnshaw, W.C. (1995) Apoptosis: lessons from in vitro systems. TrendsCell Biol. 5: 217-220.
Estman, A., Barry, M.A., Demarcq, C , Li, J. and Reynolds, J.E. (1994)Endonucleases associated with apoptosis. In Apoptosis. Edited byMihich, E. and Schimke, R.T. pp. 249-259. Plenum Press, New York.
Gabriel, D.W. and Rolfe, B.G. (1990) Working models of specific recog-nition in plant microbe interactions. Annu. Rev. Phytopathol. 28: 365-391.
Gavrieli, Y., Sherman, Y. and Ben-Sasson, S.A. (1992) Identification ofprogrammed cell death in situ via specific labeling of nuclear DNAfragmentation. J. Cell Biol. I l l : 493-501.
Gonzalez, R.G., Haxo, R.S. and Schleich, T. (1980) Mechanism of actionof polymeric aurintricarboxylic acid, a potent inhibitor of protein-nucleic acid interaction. Biochemistry 19: 4299-4303.
Gustine, D.L., Sherwood, R.T., Moyer, B.G. and Lukezic, F.L. (1990)Metabolites from Pseudomonas corrugata elicit phytoalexin biosynthesisin white clover. Phytopathology 80: 1427-1432.
Hammond-Kosack, K.E., Harrison, K. and Jones, J.D.G. (1994) De-velopmentally regulated cell death on expression of the fungal avirulencegene Avr9 in tomato seedlings carrying the disease-resistance gene Cf-9.Proc. Natl. Acad. Sci. USA 91: 10445-10449.
He, D.Y., Yazaki, Y., Nishizawa, Y., Takai, R., Yamada, K., Sakano, K.,Shibuya, N. and Minami, E. (1998) Gene activation by cytoplasmicacidification in suspension-cultured rice cells in response to the potentelicitor, N-acetylchtoheptaose. Mol-Plant Microbe Intreact. 11: 1167-1174.
He, S.Y. (1996) Elicitation of plant hypersensitive response by bacteria.Plant Physiol. 112: 865-869.
He, S.Y., Bauer, D.W., Collmer, A. and Beer, S.V. (1994) Hypersensitiveresponse elicited by Erwinia amylovora harpin requires active plantmetabolism. Mol. Plant-Microbe Intract. 7: 289-292.
He, S.Y., Huang, H.C. and Collmer, A. (1993) Pseudomonas syringae pv.syringae harpinPlJ: a protein that is secreted via the Hrp pathway andelicits the hypersensitive response in plants. Cell 73: 1255-1266.
Heath, M.C. (1991) The role of gene-for-gene interactions in determina-tion of host species specificity. Phytopathology 81: 127-130.
Heath, M.C. (1998) Apoptosis, programmed cell death and the hypersen-sitive response. Eur. J. Plant Pathol. 104: 117-124.
Kadota, I., Mizuno, A. and Nishiyama, K. (1996) Detection of a proteinspecific to the strain of Pseudomonas avenae Manns 1909 pathogenic torice. Ann. Phytopathol. Soc. Jpn. 62: 425-428.
Kadota, I., Ohuchi, A. and Nishiyama, K. (1991) Serological propertiesand specificity of Pseudomonas avenae Manns 1909, the causal agent ofbacterial brown stripe of rice. Ann. Phytopathal. Soc. Jpn. 57: 268-273.
Keen, N.T. (1990) Gene-for-gene complementarity in plant-pathogen in-teractions. Annu. Rev. Genet. 24: 447-463.
Klement, Z. (1982) Hypersensitivity. In Phytopathogenic Prokaryotes.Editied by Mount, M.S. and Lacy, G.H. Vol. 2. pp. 149-177. AcademicPress, London.
Koga, H. (1994) Hypersensitive death, autofluorescence, and ultrastruc-tural changes in cells of leaf sheaths of susceptible and resistant near-isogenic lines of rice (Pi-z1) in relation to penetration and growth ofPyricularia oryzae. Can. J. Bot. 72: 1463-1477.
Lamb, C. and Dixon, R.A. (1997) The oxidative burst in disease resistance.Annu. Rev. Plant Physiol. Plant Mol. Biol. 48: 251-275.
Levine, A., Pennell, R.I., Alvarez, M.E., Palmer, R. and Lamb, C. (1996)
Calcium-mediated apoptosis in a plant hypersensitive disease resistanceresponse. Curr. Biol. 4: 427-437.
McCabe, P.F., Levine, A., Meijer, P.J., Tapon, N.A. and Pennell, R.I.(1997) A programmed cell death pathway activated in carrot cells cul-tured at low cell density. Plant J. 12: 267-280.
Meier, B.M., Shaw, N. and Slusarenko, A.J. (1993) Spatial and temporalaccumulation of defense gene transcripts in bean (Phaseolus vulgaris)leaves in relation to bacteria-induced hypersensitive cell death. Mol.Plant-Microbe Interact. 6: 453-466.
Midoh, N. and Iwata, M. (1996) Cloning and characterization of a pro-benazole-inducible gene for an intracellular pahogenesis-related proteinin rice. Plant Cell Physiol. 37: 9-18.
Minami, E., Kuchitsu, K., He, D.Y., Kouchi, H., Midoh, N., Ohtsuki, Y.and Shibuya, N. (1996) Two novel genes rapidly and transiently acti-vated in suspension-cultured rice cells by treatment with N-acetylchito-heptaose, a biotic elicitor for phtoalexin production. Plant Cell Physiol.37: 563-567.
Mittler, R., Shulaev, V. and Lam, E. (1995) Coordinated activation ofprogrammed cell death and defense mechanisms in transgenic tobaccoplants expressing a bacterial proton pump. Plant Cell 7: 29-42.
Mittler, R., Simon, L. and Lam, E. (1997) Pathogen-induced programmedcell death in tobacco. J. Cell Sci. 110: 1333-1344.
Nicholson, D.W., Ali, A., Thornberry, N.A., Vaillancourt, J.P., Ding,C.K., Gallant, M., Gareau, Y., Griffin, P.R., Labelle, M., Lazebnik,Y.A., Munday, N.A., Raju, S.M., Smulson, M.E., Yamin, T.T., Yu,V.L. and Miller, D.K. (1995) Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 376: 37-43.
Nishiyama, K., Nishihara, N. and Ezuka, A. (1979) Bacterial brown stripeof ragi caused by Pseudomonas alboprecipitans. Ann. Phytopathol.Soc. Jpn. 45: 25-31.
Oberhammer, F., Wilson, J.W., Dive, C , Morris, I.M., Hickman, J.A.,Wakeling, A.E., Walker, P.R. and Sikorska, M. (1993) Apoptotic deathin epithelial cells: cleavage of DNA to 300 and/or 50 kb fragments priorto or in the absence of internucleosomal fragmentation. EMBO J. 12:3679-3684.
Pennell, R.I. and Lamb, C. (1997) Programmed cell death in plants. PlantCell 9: 1157-1168.
Reynolds, E.S. (1963) The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 38: 1-14.
Ryerson, D.E. and Heath, M.C. (1996) Cleavage of nuclear DNA intooligonucleosomal fragments curing cell death induced by fungal infec-tion or by abiotic treatments. Plant Cell 8: 393-402.
Stakman, E.C. (1915) Relation between Puccinia graminis and plantshighly resistant to its attack. J. Agric. Res. 4: 193-299.
Saville, J., Fadok, V., Henson, P. and Haslett, C. (1993) Phagocyte re-cognition of cells undergoing apoptosis. Immunol. Today 14: 131-136.
Schwartz, L.M., Smith, S.W., Jones, M.E.E. and Osborne, B.A. (1993)Do all programmed cell deaths occur via apoptosis? Proc. Natl. Acad.Sci. USA 90: 980-984.
Sweigard, J.A., Carroll, A.M., Kang, S., Farrall, L., Chumley, F.G. andValent, B. (1995) Identification, cloning, and characterization ofPWL2,a gene for host species specificity in the rice blast fungus. Plant Cell 7:1221-1233.
Turner, J.G. and Novacky, A. (1974) The quantitative relation betweenplant and bacterial cells involved in the hypersensitive reaction. Phyto-pathology 64: 885-890.
Walbot, V., Hoisington, D.A. and Neuffer, M.G. (1983) Disease lesionmimics in maize. In Genetic Engineering of Plant. Edited by Kosuge, T.and Meredith, C. pp. 431-442. Plenum Publishing, New York.
Wang, H., Li, J., Bostock, R.M. and Gilchrist, D.G. (1996) Apoptosis: afunctional paradigm for programmed plant cell death induced by ahost-selective phytotoxin and invoked during development. Plant Cell 8:375-391.
Wei, Z.M., Laby, R.J., Zumoff, C.H., Bauer, D.W., He, S.Y., Collmer,A. and Beer, S.V. (1992) Harpin, elicitor of the hypersensitive responseproduced by the plant pathogen Erwinia amylovora. Science 257: 85-88.
Yamada, A., Shibuya, N., Kodama, O. and Akatsuka, T. (1993) Inductionof phytoalexin formation in suspension-cultured rice cells by N-acetyl-chitooligosaccharides. Biosci. Biotech. Biochem. 57: 405-409.
(Received May 22, 1999; Accepted July 22, 1999)
Dow
nloaded from https://academ
ic.oup.com/pcp/article/40/10/1036/1884897 by guest on 22 June 2022
Related Documents











