ORIGINAL PAPER Biochemical and molecular characterization of Cronobacter spp. (formerly Enterobacter sakazakii) isolated from foods Imrich Turcovsky ´ • Kristı ´na Kunikova ´ • Hana Drahovska ´ • Eva Kaclı ´kova ´ Received: 1 March 2010 / Accepted: 6 July 2010 / Published online: 17 July 2010 Ó Springer Science+Business Media B.V. 2010 Abstract The aim of this study was to identify and characterize Cronobacter spp. isolated from a range of foods. A total of 71 Cronobacter strains were isolated from 602 foods in our laboratory. The highest contamination was observed in foods of plant origin, e.g. spices, teas, chocolate, nuts, pastries and vegetables. On the basis of genus and species identification performed using genus-specific PCR, 16S rRNA sequencing and AFLP genotyping, most of the strains belonged to Cronobacter sakazakii. Biochemical profiling by the tests included in API 20E, complemented with relevant additional tests, classified the strains into 13 biogroups. AFLP genotyping facilitated discrimination of six main groups at the 70% similarity level and strain grouping correlated clearly with species identification. Our results indicate that molecular typing by AFLP may be applied as a useful tool not only for direct comparison of Cronobacter isolates, providing trace- ability, but also for the reliable species classification. Moreover, tracing of these bacteria in a wider variety of foods should be important to enhance the knowl- edge of their transmission. Keywords Cronobacter Á Characterization Á Food isolates Á Biotyping Á AFLP genotyping Introduction Enterobacter sakazakii previously referred to as ‘yellow pigmented E. cloacae’, was defined as a new species in 1980 (Farmer et al. 1980) and 15 biogroups were described based on biochemical character- ization. Members of this species were considered relatively phenotypically and genotypically hetero- geneous (Lehner et al. 2004), a 16th biogroup has been reported and the existence of several genetic groups has been demonstrated based on 16S rRNA gene sequence analysis (Iversen et al. 2006). E. sak- azakii strains were divided into separate groups based on f-AFLP fingerprints, ribopatterns and full- length 16S rRNA sequences. DNA–DNA hybridiza- tion revealed several genomospecies (Iversen et al. 2007) and, subsequently, led to classification of these bacteria into six species within the new Cronobacter genus (Iversen et al. 2008a). Subsequently, the six Cronobacter species were supported by multilocus sequence analysis (Kuhnert et al. 2009; Baldwin et al. 2009) and a differentiation system based on PCR targeting the rpoB gene was developed (Stoop et al. I. Turcovsky ´ Á E. Kaclı ´kova ´(&) Department of Microbiology and Molecular Biology, Food Research Institute, PO Box 25, 82475 Bratislava, Slovakia e-mail: [email protected] K. Kunikova ´ Á H. Drahovska ´ Department of Molecular Biology, Faculty of Natural Sciences, Comenius University, Bratislava, Slovakia 123 Antonie van Leeuwenhoek (2011) 99:257–269 DOI 10.1007/s10482-010-9484-7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Biochemical and molecular characterization of Cronobacterspp. (formerly Enterobacter sakazakii) isolated from foods
Imrich Turcovsky • Kristına Kunikova •
Hana Drahovska • Eva Kaclıkova
Received: 1 March 2010 / Accepted: 6 July 2010 / Published online: 17 July 2010
� Springer Science+Business Media B.V. 2010
Abstract The aim of this study was to identify and
characterize Cronobacter spp. isolated from a range
of foods. A total of 71 Cronobacter strains were
isolated from 602 foods in our laboratory. The
highest contamination was observed in foods of plant
origin, e.g. spices, teas, chocolate, nuts, pastries and
vegetables. On the basis of genus and species
identification performed using genus-specific PCR,
16S rRNA sequencing and AFLP genotyping, most
of the strains belonged to Cronobacter sakazakii.
Biochemical profiling by the tests included in
API 20E, complemented with relevant additional
tests, classified the strains into 13 biogroups. AFLP
genotyping facilitated discrimination of six main
groups at the 70% similarity level and strain grouping
correlated clearly with species identification. Our
results indicate that molecular typing by AFLP may
be applied as a useful tool not only for direct
comparison of Cronobacter isolates, providing trace-
ability, but also for the reliable species classification.
Moreover, tracing of these bacteria in a wider variety
of foods should be important to enhance the knowl-
edge of their transmission.
Keywords Cronobacter � Characterization �Food isolates � Biotyping � AFLP genotyping
Introduction
Enterobacter sakazakii previously referred to as
‘yellow pigmented E. cloacae’, was defined as a new
species in 1980 (Farmer et al. 1980) and 15 biogroups
were described based on biochemical character-
ization. Members of this species were considered
relatively phenotypically and genotypically hetero-
geneous (Lehner et al. 2004), a 16th biogroup has
been reported and the existence of several genetic
groups has been demonstrated based on 16S rRNA
gene sequence analysis (Iversen et al. 2006). E. sak-
azakii strains were divided into separate groups
based on f-AFLP fingerprints, ribopatterns and full-
length 16S rRNA sequences. DNA–DNA hybridiza-
tion revealed several genomospecies (Iversen et al.
2007) and, subsequently, led to classification of these
bacteria into six species within the new Cronobacter
genus (Iversen et al. 2008a). Subsequently, the six
Cronobacter species were supported by multilocus
sequence analysis (Kuhnert et al. 2009; Baldwin et al.
2009) and a differentiation system based on PCR
targeting the rpoB gene was developed (Stoop et al.
I. Turcovsky � E. Kaclıkova (&)
Department of Microbiology and Molecular Biology,
Food Research Institute, PO Box 25,
82475 Bratislava, Slovakia
e-mail: [email protected]
K. Kunikova � H. Drahovska
Department of Molecular Biology, Faculty of Natural
Sciences, Comenius University, Bratislava, Slovakia
123
Antonie van Leeuwenhoek (2011) 99:257–269
DOI 10.1007/s10482-010-9484-7

2009). The whole genome sequence of the Cronob-
acter sakazakii ATCC BAA-894 has been recently
published and its genome was compared with repre-
sentatives of all Cronobacter species by microarray
analysis. It was shown that about 55% of genes were
common to C. sakazakii strains and 43% were
common to all Cronobacter strains (Kucerova et al.
2010).
Cronobacter spp. are considered to be opportunis-
tic pathogens, implicated in particularly severe food-
borne diseases in neonates and infants (Farber 2004;
Mullane et al. 2007; Townsend et al. 2008). Symp-
toms include bacteraemia, necrotizing enterocolitis
and meningitis, with fatality rates as high as 80%
(Lai 2001). Although these microorganisms are widely
distributed in the environment (Kandhai et al. 2004),
dried infant milk formula has been implicated as the
vehicle of transmission in many clinical manifesta-
tions (Nazarowec-White and Farber 1997; van Acker
et al. 2001; Gurtler et al. 2005). More recently,
infections in adults have been reported, in particular
among the elderly and immunocompromised patients
(Ray et al. 2007; See et al. 2007). More than 500
Cronobacter infections were reported in United
Kingdom during 1999–2007; 90% of them occurred
in persons older than 15 years (FAO/WHO report
2008). For persons with a decreased immunity, in
particular neonates, elderly and persons with severe
underlying diseases, the occurrence of Cronobacter
spp. in food and environment may represent a hazard
to health (Lai 2001; Friedemann 2007).
The primary reservoirs for subsequent food con-
tamination remain undefined due to the ubiquitous
nature of these bacteria. Cronobacter strains were
isolated from a wide range of foods including milk,
cheese, dried foods, meats, water, vegetables, rice,
tea, herbs and spices (Iversen and Forsythe 2004;
Edelson-Mammel et al. 2005; Friedemann 2007),
various food production environments and house-
holds (Kandhai et al. 2004; Gurtler et al. 2005). The
spectrum of Cronobacter-contaminated foods covers
both raw and processed foods and the kind of
processing is not restricted to dry products (Friede-
mann 2007).
Isolation and detection of Cronobacter spp. is
based on standardised microbiological procedures of
enrichment followed by the isolation of colonies on
chromogenic media. Confirmation of these colonies
is necessary to verify the identity of the isolates.
Commercial biochemical systems or PCR-based
assays are available for food and clinical microbio-
logical laboratories (Fanjat et al. 2007; Stoop et al.
2009; Druggan and Iversen 2009). Generally, pheno-
type analysis may provide unreliable identification as
long as it is based on unstable expression of the
markers (Drudy et al. 2006; Druggan and Iversen
2009).
DNA-based methods offer an alternative approach,
enabling a direct comparison of isolates in outbreaks.
Molecular methods may facilitate the trace back of
outbreak isolates from clinical samples to the source
of contamination and provide useful tools to target
control strategies to reduce the risk of transmission
(Drudy et al. 2006).
In this study, 98 strains belonging to the genus
Cronobacter, including 71 strains isolated in our
laboratory, were identified and characterized using
biochemical profiling and molecular techniques,
namely genus-specific real-time PCR, AFLP typing
and partial 16S rRNA sequencing. Since Cronobact-
er spp. are ubiquitous in many environments, a wide
range of foods was analysed.
Materials and methods
Isolation of Cronobacter spp. from food
Cronobacter strains were isolated from various food
samples analysed within the frame of a surveillance
programme for coliforms in the State Veterinary and
Food Institute, Bratislava, Slovakia in 2006–2009
(Table 1). Strains were isolated from 5 replicates of a
25 g test portion of each food sample according to the
standard method specified in ISO/TS 22964:2006
using the two-step enrichment consisting of pre-
enrichment (1/10) in Buffered Peptone Water (Merck,
Darmstadt, Germany) and selective enrichment
(1/100) at 45�C in modified Lauryl Sulfate Tryptose
(mLST) broth (Merck) with additional NaCl (Merck)
to the final concentration of 0.5 mol l-1 and vanco-
mycin 10 mg l-1 (Fluka, Buchs, Switzerland). Pre-
sumptive Cronobacter strains were isolated as typical
green–blue colonies grown on Chromogenic Entero-
bacter sakazakii agar, DFI formulation (Oxoid,
Basingstoke, England) or Chromocult Enterobac-
ter sakazakii (Merck) based on the expression of
a-glucosidase activity. Identification of Cronobacter
258 Antonie van Leeuwenhoek (2011) 99:257–269
123
spp. was confirmed by real-time PCR targeting the
dnaG gene according to Seo and Brackett (2005).
Our collection of strains was supplemented
with 22 strains from the American Type Culture
Collection (ATCC, Manassas, VA, USA), Belgian
Co-ordinated Collections of Microorganisms in Gent
(BCCM/LMG Bacteria Collection) and from Czech
Collection of Microorganisms (CCM, Brno, Czech
Republic). Five further strains isolated from fruit
powder, kindly provided by Professor Roger Stephan
from the Institute of Food Safety and Hygiene (ILS),
Vetsuisse Faculty University of Zurich, Switzerland,
were also added to the collection. All strains were
maintained in 20% glycerol solution at -18�C or
freeze-dried for long-term storage. Bacteria were
cultured in Luria–Bertani or Brain Heart broth, and
on Brain Heart agar (Merck) at 37�C. Strains of
Cronobacter spp. used in the study are listed in
Table 2.
Phenotypic characterization and biotyping
Production of the yellow pigment was tested on
Tryptone Soya Agar (Oxoid) supplemented with bile
salts (Fluka) with exposure to light during incubation.
In addition to the results derived from biochemical
reactions included in the biochemical test kit API 20E
(bioMerieux, Marcy l’Etoile, France), the following
tests were performed using conventional methods.
Tests for the acid production from carbohydrates were
examined in broth base (peptone 10 g l-1, yeast
extract 1 g l-1 and NaCl 5 g l-1) with phenol red
indicator (0.018 g l-1). Filter-sterilized dulcitol and
methyl-a-D-glucopyranoside (both from Fluka), at a
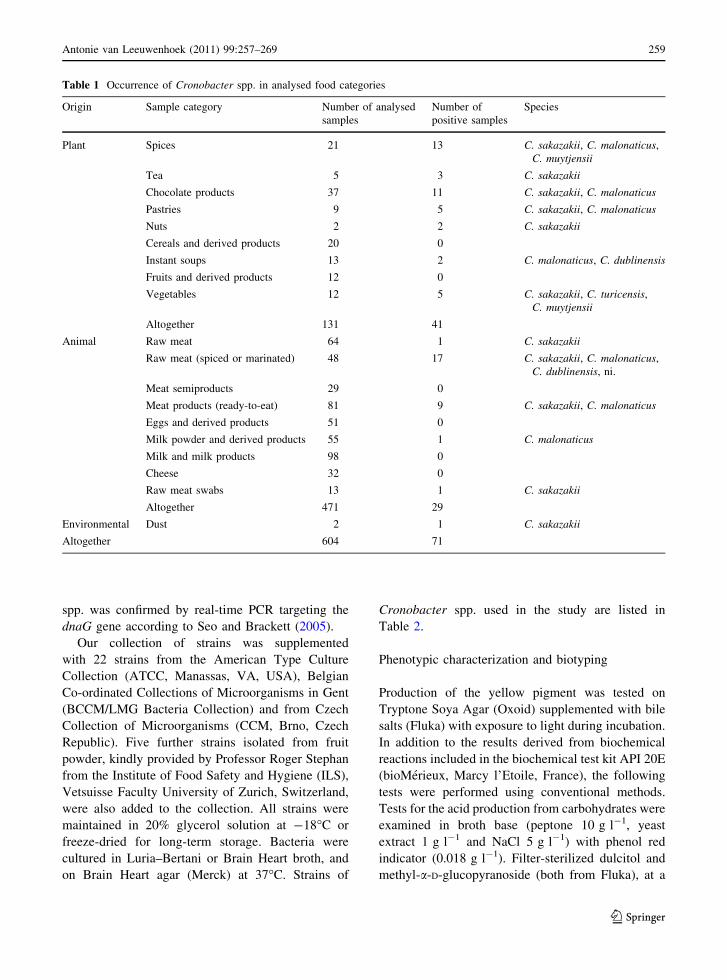
Table 1 Occurrence of Cronobacter spp. in analysed food categories
Origin Sample category Number of analysed
samples
Number of
positive samples
Species
Plant Spices 21 13 C. sakazakii, C. malonaticus,
C. muytjensii
Tea 5 3 C. sakazakii
Chocolate products 37 11 C. sakazakii, C. malonaticus
Pastries 9 5 C. sakazakii, C. malonaticus
Nuts 2 2 C. sakazakii
Cereals and derived products 20 0
Instant soups 13 2 C. malonaticus, C. dublinensis
Fruits and derived products 12 0
Vegetables 12 5 C. sakazakii, C. turicensis,
C. muytjensii
Altogether 131 41
Animal Raw meat 64 1 C. sakazakii
Raw meat (spiced or marinated) 48 17 C. sakazakii, C. malonaticus,
C. dublinensis, ni.
Meat semiproducts 29 0
Meat products (ready-to-eat) 81 9 C. sakazakii, C. malonaticus
Eggs and derived products 51 0
Milk powder and derived products 55 1 C. malonaticus
Milk and milk products 98 0
Cheese 32 0
Raw meat swabs 13 1 C. sakazakii
Altogether 471 29
Environmental Dust 2 1 C. sakazakii
Altogether 604 71
Antonie van Leeuwenhoek (2011) 99:257–269 259
123
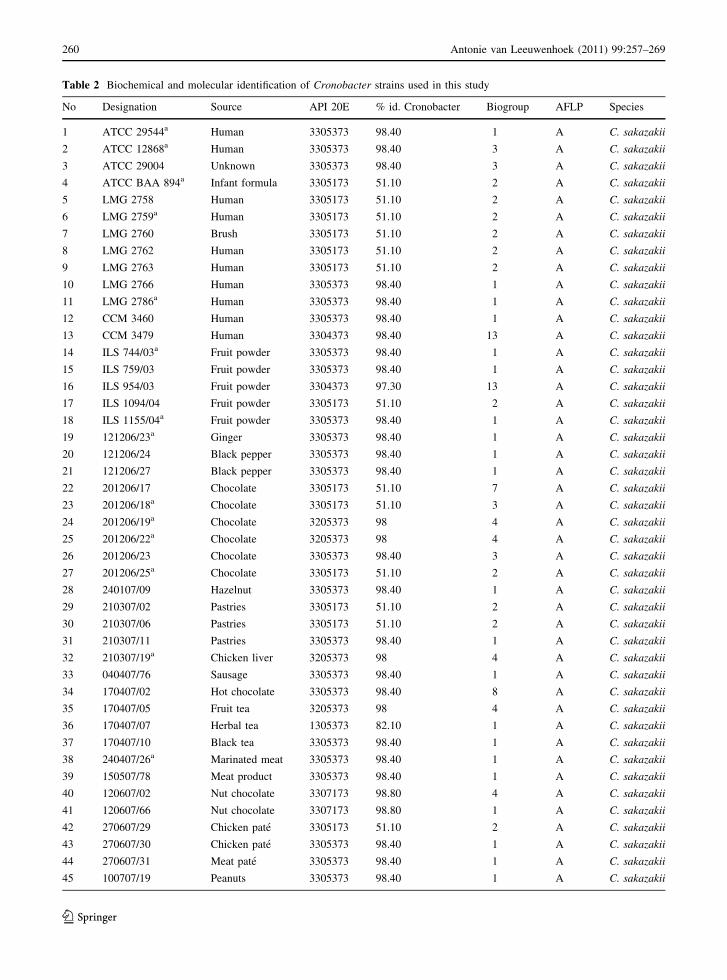
Table 2 Biochemical and molecular identification of Cronobacter strains used in this study
No Designation Source API 20E % id. Cronobacter Biogroup AFLP Species
1 ATCC 29544a Human 3305373 98.40 1 A C. sakazakii
2 ATCC 12868a Human 3305373 98.40 3 A C. sakazakii
3 ATCC 29004 Unknown 3305373 98.40 3 A C. sakazakii
4 ATCC BAA 894a Infant formula 3305173 51.10 2 A C. sakazakii
5 LMG 2758 Human 3305173 51.10 2 A C. sakazakii
6 LMG 2759a Human 3305173 51.10 2 A C. sakazakii
7 LMG 2760 Brush 3305173 51.10 2 A C. sakazakii
8 LMG 2762 Human 3305173 51.10 2 A C. sakazakii
9 LMG 2763 Human 3305173 51.10 2 A C. sakazakii
10 LMG 2766 Human 3305373 98.40 1 A C. sakazakii
11 LMG 2786a Human 3305373 98.40 1 A C. sakazakii
12 CCM 3460 Human 3305373 98.40 1 A C. sakazakii
13 CCM 3479 Human 3304373 98.40 13 A C. sakazakii
14 ILS 744/03a Fruit powder 3305373 98.40 1 A C. sakazakii
15 ILS 759/03 Fruit powder 3305373 98.40 1 A C. sakazakii
16 ILS 954/03 Fruit powder 3304373 97.30 13 A C. sakazakii
17 ILS 1094/04 Fruit powder 3305173 51.10 2 A C. sakazakii
18 ILS 1155/04a Fruit powder 3305373 98.40 1 A C. sakazakii
19 121206/23a Ginger 3305373 98.40 1 A C. sakazakii
20 121206/24 Black pepper 3305373 98.40 1 A C. sakazakii
21 121206/27 Black pepper 3305373 98.40 1 A C. sakazakii
22 201206/17 Chocolate 3305173 51.10 7 A C. sakazakii
23 201206/18a Chocolate 3305173 51.10 3 A C. sakazakii
24 201206/19a Chocolate 3205373 98 4 A C. sakazakii
25 201206/22a Chocolate 3205373 98 4 A C. sakazakii
26 201206/23 Chocolate 3305373 98.40 3 A C. sakazakii
27 201206/25a Chocolate 3305173 51.10 2 A C. sakazakii
28 240107/09 Hazelnut 3305373 98.40 1 A C. sakazakii
29 210307/02 Pastries 3305173 51.10 2 A C. sakazakii
30 210307/06 Pastries 3305173 51.10 2 A C. sakazakii
31 210307/11 Pastries 3305373 98.40 1 A C. sakazakii
32 210307/19a Chicken liver 3205373 98 4 A C. sakazakii
33 040407/76 Sausage 3305373 98.40 1 A C. sakazakii
34 170407/02 Hot chocolate 3305373 98.40 8 A C. sakazakii
35 170407/05 Fruit tea 3205373 98 4 A C. sakazakii
36 170407/07 Herbal tea 1305373 82.10 1 A C. sakazakii
37 170407/10 Black tea 3305373 98.40 1 A C. sakazakii
38 240407/26a Marinated meat 3305373 98.40 1 A C. sakazakii
39 150507/78 Meat product 3305373 98.40 1 A C. sakazakii
40 120607/02 Nut chocolate 3307173 98.80 4 A C. sakazakii
41 120607/66 Nut chocolate 3307173 98.80 1 A C. sakazakii
42 270607/29 Chicken pate 3305173 51.10 2 A C. sakazakii
43 270607/30 Chicken pate 3305373 98.40 1 A C. sakazakii
44 270607/31 Meat pate 3305373 98.40 1 A C. sakazakii
45 100707/19 Peanuts 3305373 98.40 1 A C. sakazakii
260 Antonie van Leeuwenhoek (2011) 99:257–269
123
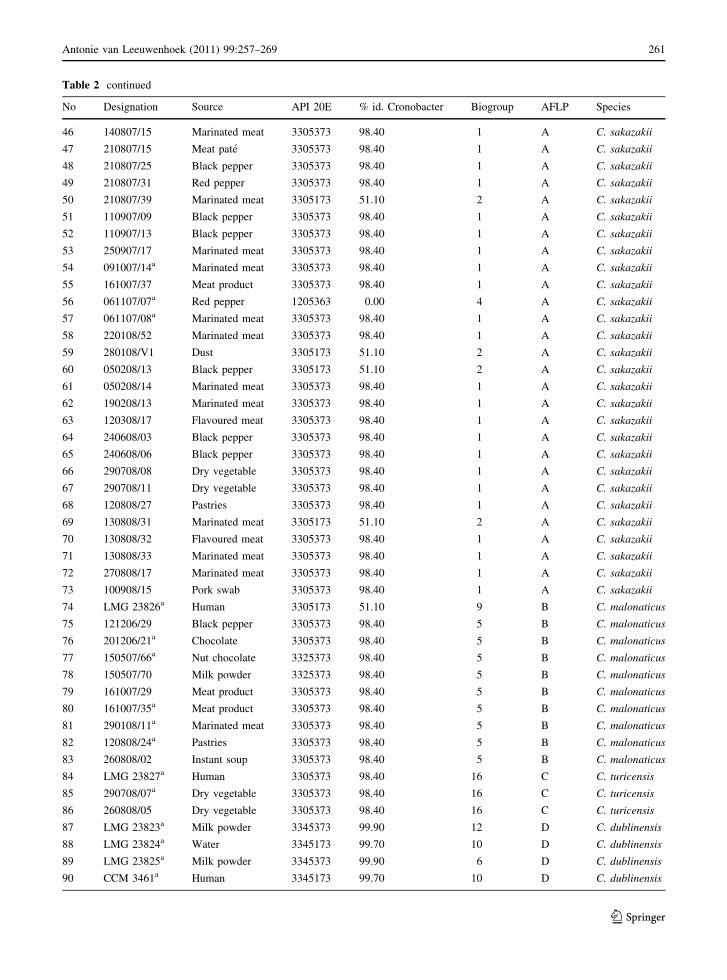
Table 2 continued
No Designation Source API 20E % id. Cronobacter Biogroup AFLP Species
46 140807/15 Marinated meat 3305373 98.40 1 A C. sakazakii
47 210807/15 Meat pate 3305373 98.40 1 A C. sakazakii
48 210807/25 Black pepper 3305373 98.40 1 A C. sakazakii
49 210807/31 Red pepper 3305373 98.40 1 A C. sakazakii
50 210807/39 Marinated meat 3305173 51.10 2 A C. sakazakii
51 110907/09 Black pepper 3305373 98.40 1 A C. sakazakii
52 110907/13 Black pepper 3305373 98.40 1 A C. sakazakii
53 250907/17 Marinated meat 3305373 98.40 1 A C. sakazakii
54 091007/14a Marinated meat 3305373 98.40 1 A C. sakazakii
55 161007/37 Meat product 3305373 98.40 1 A C. sakazakii
56 061107/07a Red pepper 1205363 0.00 4 A C. sakazakii
57 061107/08a Marinated meat 3305373 98.40 1 A C. sakazakii
58 220108/52 Marinated meat 3305373 98.40 1 A C. sakazakii
59 280108/V1 Dust 3305173 51.10 2 A C. sakazakii
60 050208/13 Black pepper 3305173 51.10 2 A C. sakazakii
61 050208/14 Marinated meat 3305373 98.40 1 A C. sakazakii
62 190208/13 Marinated meat 3305373 98.40 1 A C. sakazakii
63 120308/17 Flavoured meat 3305373 98.40 1 A C. sakazakii
64 240608/03 Black pepper 3305373 98.40 1 A C. sakazakii
65 240608/06 Black pepper 3305373 98.40 1 A C. sakazakii
66 290708/08 Dry vegetable 3305373 98.40 1 A C. sakazakii
67 290708/11 Dry vegetable 3305373 98.40 1 A C. sakazakii
68 120808/27 Pastries 3305373 98.40 1 A C. sakazakii
69 130808/31 Marinated meat 3305173 51.10 2 A C. sakazakii
70 130808/32 Flavoured meat 3305373 98.40 1 A C. sakazakii
71 130808/33 Marinated meat 3305373 98.40 1 A C. sakazakii
72 270808/17 Marinated meat 3305373 98.40 1 A C. sakazakii
73 100908/15 Pork swab 3305373 98.40 1 A C. sakazakii
74 LMG 23826a Human 3305173 51.10 9 B C. malonaticus
75 121206/29 Black pepper 3305373 98.40 5 B C. malonaticus
76 201206/21a Chocolate 3305373 98.40 5 B C. malonaticus
77 150507/66a Nut chocolate 3325373 98.40 5 B C. malonaticus
78 150507/70 Milk powder 3325373 98.40 5 B C. malonaticus
79 161007/29 Meat product 3305373 98.40 5 B C. malonaticus
80 161007/35a Meat product 3305373 98.40 5 B C. malonaticus
81 290108/11a Marinated meat 3305373 98.40 5 B C. malonaticus
82 120808/24a Pastries 3305373 98.40 5 B C. malonaticus
83 260808/02 Instant soup 3305373 98.40 5 B C. malonaticus
84 LMG 23827a Human 3305373 98.40 16 C C. turicensis
85 290708/07a Dry vegetable 3305373 98.40 16 C C. turicensis
86 260808/05 Dry vegetable 3305373 98.40 16 C C. turicensis
87 LMG 23823a Milk powder 3345373 99.90 12 D C. dublinensis
88 LMG 23824a Water 3345173 99.70 10 D C. dublinensis
89 LMG 23825a Milk powder 3345373 99.90 6 D C. dublinensis
90 CCM 3461a Human 3345173 99.70 10 D C. dublinensis
Antonie van Leeuwenhoek (2011) 99:257–269 261
123

final concentration of 0.5% were used. Gas production
from glucose was determined using Durham tubes.
Malonate utilization was determined using Malonate
broth (ammonium sulphate 1 g l-1, dipotassium phos-
phate 0.6 g l-1, monopotassium phosphate 0.4 g l-1,
NaCl 2 g l-1, sodium malonate 3 g l-1 and bromoth-
ymol blue indicator 0.025 g l-1, pH 6.8). Motility at
36�C was determined in Motility medium (tryptose
10 g l-1, NaCl 5 g l-1, agar 5 g l-1) with 0.05 g l-1
triphenyltetrazoliumchloride allowing the pink col-
our-visualized diffusion of motile cells from stab
inoculation. Nitrate reduction was determined using
Nitrate Reduction Kit (Fluka) and the addition of zinc
dust to confirm the presence of unreduced nitrate in
tubes with negative results. Methyl-red test was
performed by addition of indicator (0.1 g in 300 ml
of 95% ethanol) to 48-h cultures in 5 ml of Glucose
phosphate broth (peptone 10 g l-1, dipotassium phos-
phate 5 g l-1 and glucose 5 g l-1) in tubes.
Sequencing of 16S rRNA gene
The chromosomal DNA was isolated using the
DNeasy Blood and Tissue kit (Qiagen, Hilden,
Germany) according to manufacturers instructions.
A fragment of the 16S rRNA gene was ampli-
fied with bacterial universal primers 27f/685r (Lane
1991). DNA sequencing was performed using the
automatic DNA analyzer ABI Prism 3130 Avant
(Applied Biosystems, Foster City, CA, USA) using
Big Dye Terminator 3.1 kit (Applied Biosystems).
Final sequences (one representative for each unique
sequence) were deposited into the EMBL database
under accession numbers FN539026–FN539035.
Amplified fragment length polymorphism
AFLP was performed according to the procedure
described previously (Pangallo et al. 2008). 200 ng of
the genomic DNA was digested with 5 U of EcoRI
and 2 U of MseI in a 10 ll reaction mixture
containing 19 T4 DNA ligase buffer (New England
Biolabs, Ipswich, USA), 0.05 mol l-1 NaCl and
0.1 mg ml-1 bovine serum albumin at 37�C for 1 h.
Then, 5 ll of the fresh ligation mixture containing
50 pmol each of the EcoRI adapter (ctcgtagactgcg-
tacc/aattggtacgcagtctac) and MseI adapter (gacgat-
gagtcctgag/tactcaggactcat), and 1 U of T4 DNA
ligase were added and the incubation continued for
3 h. Subsequently, the mixture was diluted 20 times
with TE buffer (10 mmol l-1 Tris HCl, 0.1 mmol l-1
EDTA, pH 8) and stored at -20�C. PCR was per-
formed in a 20 ll volume, which contained 10 pmol
of EcoA primer (gactgcgtaccaattca, FAM-labelled),
10 pmol of MseC primer (gatgagtcctgagtaac),
200 lmol l-1 of each dNTP (Promega, Madison,
USA), 1 U Taq DNA polymerase (New England
Biolabs), 19 PCR buffer and 2 ll template DNA.
The thermocycler program consisted of 2 min at
72�C, 10 cycles (30 s at 94�C, 30 s at 65�C, 90 s at
72�C, the annealing temperature decreased by 1�C
with each cycle), 25 cycles (30 s at 94�C, 30 s at
56�C, 90 s at 72�C), and a final polymerization for
30 min at 60�C. PCR products were separated in the
automatic genetic analyzer ABI Prism 3130 with
LIZ-500 size standard (Applied Biosystems). The
AFLP profiles were analysed by the BioNumerics
Software (Applied Maths, Belgium). The levels of
similarity between AFLP fingerprints were calculated
Table 2 continued
No Designation Source API 20E % id. Cronobacter Biogroup AFLP Species
91 140807/04 Marinated meat 3345373 99.90 6 D C. dublinensis
92 260808/01a Instant soup 3345173 99.70 10 D C. dublinensis
93 040407/32a Spiced meat 3305373 98.40 1 E Ni
94 ATCC 51329a Unknown 3345373 99.90 15 F C. muytjensii
95 LMG 2787 Human 3345373 99.90 15 F C. muytjensii
96 LMG 2788a Human 3345373 99.90 15 F C. muytjensii
97 051206/07 Caraway 3345373 99.90 15 F C. muytjensii
98 260808/07a Dry vegetable 3247373 99.40 15 nt C. muytjensii
a Strains were analyzed by 16S rRNA gene sequencing; ni not identified; nt untypable
262 Antonie van Leeuwenhoek (2011) 99:257–269
123

on the basis of Dice’s coefficient (at a position
tolerance of 0.2% and optimization at 0) and cluster
analysis was done by UPGMA.
Results
Isolation, identification and phenotypic
characterization of Cronobacter strains
Seventy-one Cronobacter strains were isolated and
identified from 602 food and 2 environmental sam-
ples sorted for this study to 19 sample categories
(Table 1) by processing of five test portions from
each sample. Among the analysed samples, Cronob-
acter strains were isolated from 41 of 131 (31.3%)
samples of plant origin and from 29 of 471 (6.2%)
samples of animal origin; one strain was isolated
from dust.
All 71 putative Cronobacter strains which grew as
green–blue colonies on the chromogenic medium
were positively identified as Cronobacter spp. by the
genus-specific real-time PCR. All but one were
determined to be ‘‘E. sakazakii’’ using API 20E. All
strains produced a yellow pigment.
The panel of biochemical tests contained in the
API 20E gave 10 different profiles resulting in per-
centage identification (% id.) values from 51.1% for
19 inositol-negative C. sakazakii and Cronobact-
er malonaticus to 99.9% for most Cronobacter
muytjensii and some Cronobacter dublinensis isolates.
One strain (061107/07) was misidentified as Esche-
richia vulneris at 61.5% id and a 0.0% id. value for
‘‘E. sakazakii’’. However, this isolate was positive in
the genus-specific real time PCR and clustered with
C. sakazakii by AFLP (Fig. 1). All 22 Cronobacter
spp. culture collection strains and five strains from
ILS grew as green–blue colonies on the chromogenic
medium, produced the yellow pigment on Tryptone
soya agar with bile salts and were positive in the real-
time PCR confirmation. By API 20E, these strains
were identified as ‘‘E. sakazakii’’ with 51.1–99.9% id.
Additional biochemical tests such as acid production
from dulcitol and methyl-a-D-glucopyranoside, mal-
onate utilization, gas production from glucose,
methyl red and motility tests facilitated classification
of the strains into 13 different biogroups according to
Farmer et al. (1980) and Iversen et al. (2006)
(Table 2).
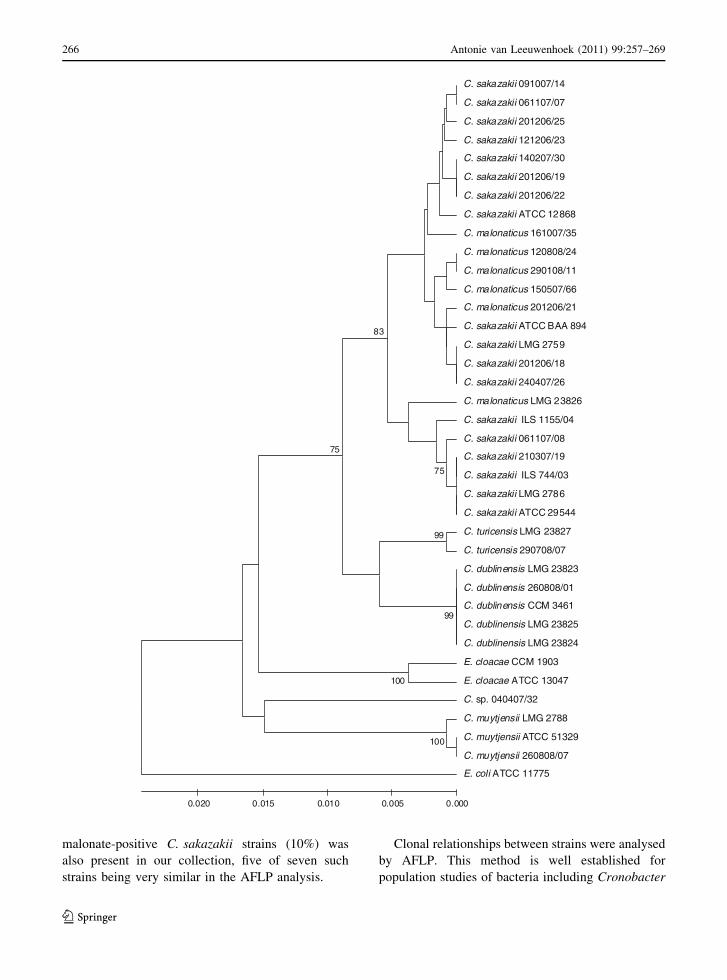
AFLP subtyping
The clonality of 71 Cronobacter spp. food isolates, 22
collection strains and 5 identified isolates was deter-
mined by AFLP. By repeated analysis of the strain
C. sakazakii ATCC 12868 (CCM 1902), 95–100%
similarity of Dice’s coefficient was obtained for
all runs. AFLP-profile similarity of different strains
reached values from 60 to 100%. Strains were
grouped into 46 clusters at a similarity level of 90%.
Six main groups (designated A–F) were clearly
distinguished at the 70% similarity level; these strain
grouping being in concordance with their species
identification and biochemical properties (Fig. 1).
From the total number of 98 Cronobacter strains,
the largest AFLP cluster A contained 73 C. sakazakii
strains including the type strain ATCC 29544 (LMG
5740). Other Cronobacter species were separated into
the remaining AFLP clusters: ten strains of C. malo-
naticus in cluster B, three Cronobacter turicensis
strains in cluster C, six C. dublinensis strains in
cluster D and four C. muytjensii strains in cluster F.
One strain (040407/32) had a low similarity to all
other Cronobacter strains and was separated to its
own cluster E, while API 20E identified this strain as
‘E. sakazakii’ with 98.4% id. value corresponding to
C. sakazakii type strains profile (3305373) and bio-
type 1 using additional biochemical tests. The strain
C. muytjensii 260808/07 was untypable by AFLP.
Partial sequencing of 16S rRNA gene
For confirmation of species identification, a fragment
of 657 bp from the beginning of the 16S rRNA gene
was sequenced from 27 Cronobacter strains. Eight
additional sequences of collection strains were
downloaded from the NCBI database (Fig. 2). The
similarity of all Cronobacter spp. strains reached
95.5% and the sequences were grouped according to
the species: C. dublinensis (5 strains, 100% similar-
ity), C. turicensis (2 strains, 99.9% similarity),
C. muytjensii (3 strains, 99.7% similarity). C. sak-
azakii (18 strains) and C. malonaticus (6 strains)
could not be distinguished by partial 16S rRNA
sequencing and formed one common cluster with
98.3% similarity. Cronobacter sp. 040407/32, which
had a unique profile in AFLP, formed an independent
branch also in the 16S rRNA sequence analysis being
adjacent to C. muytjensii sequences.
Antonie van Leeuwenhoek (2011) 99:257–269 263
123

264 Antonie van Leeuwenhoek (2011) 99:257–269
123

Discussion
Cronobacter spp. (formerly E. sakazakii) have
emerged as a rare cause of life-threatening neonatal
infections. The microorganism has been detected in
powdered infant milk formula products and improper
handling during its rehydration has been implicated
in several clinical cases. Considering the indication
that Cronobacter spp. may cause infection in both
infants and adults, evaluation of a wide variety of
foods may be important to reveal the possible routes
for its transmission.
In our study, 71 strains of Cronobacter genus
isolated from a variety of foods in our laboratory and
5 other isolates were identified and compared with 22
strains from collections. All strains were analysed
using biochemical profiling and molecular tech-
niques, namely genus-specific real-time PCR, AFLP
typing and, for selected strains, sequencing of
16S rRNA.
During food screening, more than 600 food
products other than infant formula showed on average
12% positivity. However, this value may be an
underestimate, as the method specified in ISO/TS
22964 was used for isolation as the best alternative
available in 2006, when the study has started. This
method is now known to miss some strains unable to
grow in mLST media and/or at 45�C (Iversen and
Forsythe 2007; Druggan and Iversen 2009). Advanced
procedures have been recently recommended for
effective isolation of Cronobacter spp., e.g. combi-
nation of Cronobacter screening broth (CSB) and
chromogenic agar (Iversen et al. 2008b), and alter-
natively Cronobacter enrichment broth (CEB) may
be effectively used in conjunction with selective-
differential agar (O’Brien et al. 2009a).
Cronobacter spp. occurrence in samples was
unevenly distributed in different food types, the
highest contamination being observed in foods of
plant origin, e.g. spices, teas, chocolate, nuts, pastries
and vegetables. It is also interesting that the majority
of positive samples of animal origin contained plant
ingredients, namely herbs and spices. This corre-
sponds to the observations of other authors:
Cronobacter contamination was particularly detected
in fresh and dried vegetables and spices (Iversen and
Forsythe 2004; Baumgartner et al. 2009; Jaradat et al.
2009) and in cereal-based infant drinks and foods
(O’Brien et al. 2009b; Chap et al. 2009). The ability
of Cronobacter strains to associate with plants was
recently experimentally confirmed for environmental
as well as clinical strains (Schmid et al. 2009). Their
presence in dried foods (spices, chocolate, pastries)
may be due to the increased desiccation resistance of
Cronobacter strains (Breeuwer et al. 2003; Edelson-
Mammel et al. 2005; Osaili and Forsythe 2009).
Real-time PCR targeting the dnaG gene of the
MMS operon according to Seo and Brackett (2005) is
an extensively utilized Cronobacter spp. identifica-
tion method (Druggan and Iversen 2009). In our study
it was used as a principal identification test for
Cronobacter genus confirmation and it 100% agreed
with AFLP and 16S rRNA sequencing classification.
API 20E biochemical identification system (bio-
Merieux) was employed as a simple commercially
available tool. It proved to be highly accurate for
Cronobacter identification at the genus level as all
but one isolate were correctly identified (99% relative
sensitivity based upon the analysis of 98 Cronobacter
strains). Most of the analysed Cronobacter strains
(80%) fell within the range from 98.0 to 99.9% id.
values. At the same time, the id. value of 98.4% for
C. sakazakii type strain ATCC 29544 was lower than
for C. muytjensii strains with id. values of 99.9%.
Surprisingly, the lowest % id. value of 51.1%, which
is considered a doubtful result by the API 20E
software, was associated with the group of 19
inositol-negative C. sakazakii and C. malonaticus
strains, as negative inositol fermentation is a typical
attribute of about 25% of Cronobacter strains
(Farmer et al. 1980). Inability to correctly identify
some Cronobacter strains by the API20E was previ-
ously reported (Iversen et al. 2004). For this reason,
additional tests such as production of acid from
dulcitol and methyl-a-D-glucopyranoside, as well as
utilization of malonate according to Iversen et al.
(2007) were necessary for reliable genus level
classification. Further additional tests such as motility
test, gas production and methyl red tests allowed the
identification of Cronobacter spp. (E. sakazakii)
biogroups (Farmer et al. 1980; Iversen et al. 2006).
It was interesting that apart from all the C. malo-
naticus isolates, a relatively high proportion of
Fig. 1 Dendrogram of AFLP profiles of Cronobacter strains.
AFLP profiles of 97 Cronobacter strains with species
separation at the 70% similarity level are shown in the
dendrogram. E. cloacae CCM 1903 is used as an outgroup
b
Antonie van Leeuwenhoek (2011) 99:257–269 265
123
malonate-positive C. sakazakii strains (10%) was
also present in our collection, five of seven such
strains being very similar in the AFLP analysis.
Clonal relationships between strains were analysed
by AFLP. This method is well established for
population studies of bacteria including Cronobacter
C. sakazakii 091007/14
C. sakazakii 061107/07
C. sakazakii 201206/25
C. sakazakii 121206/23
C. sakazakii 140207/30
C. sakazakii 201206/19
C. sakazakii 201206/22
C. sakazakii ATCC 12868
C. malonaticus 161007/35
C. malonaticus 120808/24
C. malonaticus 290108/11
C. malonaticus 150507/66
C. malonaticus 201206/21
C. sakazakii ATCC BAA 894
C. sakazakii LMG 2759
C. sakazakii 201206/18
C. sakazakii 240407/26
C. malonaticus LMG 23826
C. sakazakii ILS 1155/04
C. sakazakii 061107/08
C. sakazakii 210307/19
C. sakazakii ILS 744/03
C. sakazakii LMG 2786
C. sakazakii ATCC 29544
C. turicensis LMG 23827
C. turicensis 290708/07
C. dublinensis LMG 23823
C. dublinensis 260808/01
C. dublinensis CCM 3461
C. dublinensis LMG 23825
C. dublinensis LMG 23824
E. cloacae CCM 1903
E. cloacae ATCC 13047
C. sp. 040407/32
C. muytjensii LMG 2788
C. muytjensii ATCC 51329
C. muytjensii 260808/07
E. coli ATCC 11775
99
75
100
99
100
83
75
0.0000.0050.0100.0150.020
266 Antonie van Leeuwenhoek (2011) 99:257–269
123

spp. (Iversen et al. 2007). In our study, a high
discriminatory power of AFLP was confirmed, as the
98 strains were separated into 46 different clusters at
a similarity level of 90%. With a few exceptions,
different strains simultaneously isolated from differ-
ent batches of the same food did not share any genetic
similarity, which may reflect the fact that Cronob-
acter spp. contamination occurs at various occasions
from different sources. At a similarity level of 70%,
six main groups were clearly distinguished in the
AFLP dendrogram, and strain grouping was in
concordance with their species identification as
confirmed by biogroups and 16S rRNA sequencing.
With a few exceptions, strains that were highly
similar according to the AFLP analysis had the same
biogroup assignation. Despite using different restric-
tion endonucleases in their AFLP protocol, Iversen
et al. (2007) observed similar species specific clus-
tering of the AFLP profiles of Cronobacter strains.
The AFLP-based similarity between strains belong-
ing to the same species reached 73–83%, the most
heterogeneous being three strains of C. turicensis. In
C. dublinensis, no clear delineation between subspe-
cies dublinensis, lactaridi and lausannensis was
observed, which might have been due to the low
number of analysed strains. Our results indicate that
AFLP typing may be applied as a useful tool not only
for direct comparison of Cronobacter isolates pro-
viding the traceability, but also for the reliable
species identification and classification. Partial
sequencing of 16S rRNA gene was further used for
confirmation of species identification and in all cases
we observed strain clustering identical with main
species delineating AFLP groups. The only excep-
tions were C. sakazakii and C. malonaticus, which
belonged to one common 16S rRNA cluster.
However, due to low variability in the sequenced
region and due to some polymorphism between 16S
rRNA copies in the same strain (e.g. 1–6 nucleotide
variants are present between six 16S rRNA genes in
the genome of C. sakazakii ATTC BAA 894,
Kucerova et al. 2010), the relationship between these
two species could not be properly distinguished from
the dendrogram (many branches are supported with
low bootstrap values).
Among Cronobacter spp. food isolates, strains of
C. sakazakii dominated with 79% occurrence. This
species was followed by C. malonaticus (12% iso-
lates), while representatives of other species were
isolated only rarely. Similar results were published
previously, e.g., preferential isolation of C. sakazakii
from dairy products was observed by El-Sharoud
et al. (2009). This confirms the predominance of this
species among food isolates (Iversen et al. 2007;
Chap et al. 2009).
One strain (040407/32) isolated from spiced meat
possessed a unique AFLP profile and differed from
all other Cronobacter spp. also in 16S rRNA gene
sequence. Detailed taxonomic classification of this
strain will be the subject of our further study.
In conclusion, we can conclude that Cronobacter
spp. are frequently present in various foods, in
particular fresh and dry foods of plant origin.
Although it is harmless for most of the population,
it can pose risks for immunocompromised consumers.
Tracing of these bacteria in a wide variety of foods is
important to reveal the possible routes for its
transmission.
Acknowledgments This work was supported by Slovak
Research and Development Agency under the contract no.
APVV-27-009705 and by Slovak Ministry of Education under
the contract no. VEGA 1/0344/10.
References
Baldwin A, Loughlin M, Caubilla-Barron J, Kucerova E,
Manning G, Dowson C, Forsythe S (2009) Multilocus
sequence typing of Cronobacter sakazakii and Cronob-acter malonaticus reveals stable clonal structures with
clinical significance which do not correlate with biotypes.
BMC Microbiol 9:223
Baumgartner A, Grand M, Liniger M, Iversen C (2009)
Detection and frequency of Cronobacter spp. (Entero-bacter sakazakii) in different categories of ready-to-eat
foods other than infant formula. Int J Food Microbiol
136:189–192
Fig. 2 Dendrogram of partial 16S rRNA sequences of Cronob-acter strains. The 657 bp fragment from the beginning of the
16S rRNA gene was compared in 35 Cronobacter strains.
E. cloacae ATCC 13047 (AJ51469), E. cloacae CCM 1903
(FN539035) and E. coli ATCC 11775 (X80725) were used as
outgroups. Sequences of strains ATCC BAA 894 (NC_009778),
ATTC 29544 (EF 059843), ATCC 51329 (EF059845), LMG
23823 (EF059892), LMG 23824 (EF059841), LMG 23825
(EF059838), LMG 23826 (EF059881), LMG 23827 (EF059891)
were obtained from GenBank database. The other strains were
sequenced in our study (FN539026–FN539034). UPGMA
clustering by MEGA 4 software (Tamura et al. 2007) was used
in dendrogram construction and bootstrap values greater than 75
were shown
b
Antonie van Leeuwenhoek (2011) 99:257–269 267
123

Breeuwer P, Lardeau A, Peterz M, Joosten HM (2003) Des-
iccation and heat tolerance of Enterobacter sakazakii.J Appl Microbiol 95:967–973
Chap J, Jackson P, Siqueira R, Gaspar N, Quintas C, Park J,
Osaili T, Shaker R, Jaradat Z, Hartantyo SHP, Abdullah
Sani N, Estuningsih S, Forsythe SJ (2009) International
survey of Cronobacter sakazakii and other Cronobacterspp. in follow up formulas and infant foods. Int J Food
Microbiol 136:185–188
Drudy D, O’Rourke M, Murphy M et al (2006) Characteriza-
tion of a collection of Enterobacter sakazakii isolates
from environmental and food sources. Int J Food Micro-
biol 110:127–134
Druggan P, Iversen C (2009) Culture media for the isolation of
Cronobacter spp. Int J Food Microbiol 136:169–178
Edelson-Mammel SG, Porteous MK, Buchanan RL (2005)
Survival of Enterobacter sakazakii in a dehydrated pow-
dered infant formula. J Food Prot 68:1900–1902
El-Sharoud WM, O’Brien S, Negredo C, Iversen C, Fanning S,
Healy B (2009) Characterization of Cronobacter recov-
ered from dried milk and related products. BMC Micro-
biol 9:24–32
Fanjat N, Leclercq A, Joosten H, Robichon D (2007) Com-
parison of the phenotyping methods ID 32E and VITEK 2
compact GN with 16 rDNA gene sequencing for the
identification of Enterobacter sakazakii. J Clin Microbiol
45:2048–2050
FAO/WHO report (2008) Enterobacter sakazakii (Cronobacterspp.) in powdered follow-up formulae: meeting report.
Microbiological risk assessment series no. 15. Rome,
Italy, 90 pp
Farber JM (2004) Enterobacter sakazakii—new foods for
thought? Lancet 363:5–6
Farmer JJ III, Asbury MA, Hickman FW, Brenner DJ, The
Enterobacteriaceae Study Group (1980) Enterobactersakazakii: a new species of ‘‘Enterobacteriaceae’’ isolated
from clinical specimens. Int J Syst Bacteriol 30:569–584
Friedemann M (2007) Enterobacter sakazakii in food and
beverages (other than infant formula and milk powder).
Int J Food Microbiol 116:1–10
Gurtler JB, Kornacki JL, Beuchat LR (2005) Enterobactersakazakii: a coliform of increased concern to infant health.
Review. Int J Food Microbiol 104:1–34
Iversen C, Forsythe S (2004) Isolation of Enterobacter sak-azakii and other Enterobacteriaceae from powdered infant
formula milk and related products. Food Microbiol
21:771–777
Iversen C, Forsythe SJ (2007) Comparison of media for the
isolation of Enterobacter sakazakii. Appl Environ
Microbiol 73:48–52
Iversen C, Druggan P, Forsythe S (2004) A selective differ-
ential medium for Enterobacter sakazakii, a preliminary
study. Int J Food Microbiol 96:133–139
Iversen C, Waddington M, Farmer JJ III, Forsythe S (2006)
The biochemical differentiation of Enterobacter sakazakiigenotypes. BMC Microbiol 6:94–100
Iversen C, Lehner A, Mullane N et al (2007) The taxonomy
of Enterobacter sakazakii: proposal of a new genus
Cronobacter gen. nov. and descriptions of Cronobactersakazakii comb. nov. Cronobacter sakazakii subsp. sak-azakii, comb. nov., Cronobacter sakazakii subsp.
malonaticus subsp. nov., Cronobacter turicensis sp. nov.,
Cronobacter muytjensii sp. nov., Cronobacter dublinen-sis sp. nov. and Cronobacter genomospecies 1. BMC
Evol Biol 7:64–78
Iversen C, Mullane N, McCardell B, Tall BD, Lehner A,
Fanning S, Stephan R, Joosten H (2008a) Cronobactergen. nov., a new genus to accommodate the biogroups of
Enterobacter sakazakii, and proposal of Cronobactersakazakii gen nov., comb. nov., Cronobacter malonaticussp. nov., Cronobacter turicensis sp. nov., Cronobactermuytjensii sp. nov., Cronobacter dublinensis sp. nov.,
Cronobacter genomospecies 1 and three subspecies, Cro-nobacter dublinensis subsp. dublinensis subsp. nov.,
Cronobacter dublinensis subsp. lausannensis subsp. nov.
and Cronobacter dublinensis subsp. lactaridi subsp.
nov. Int J Syst Evol Microbiol 58:1442–1447
Iversen C, Druggan P, Schumacher S, Lehner A, Feer C,
Gschwend K, Joosten H, Stephan R (2008b) Development
of a novel screening method for the isolation of ‘‘Cro-nobacter’’ spp. (Enterobacter sakazakii). Appl Environ
Microbiol 74:2550–2553
Jaradat ZW, Ababneh QO, Saadoun IM, Samara NA,
Rashdan AM (2009) Isolation of Cronobacter spp. (for-
merly Enterobacter sakazakii) from infant food, herbs and
environmental samples and the subsequent identification
and confirmation of the isolates using biochemical, chro-
mogenic assays, PCR and 16S rRNA sequencing. BMC
Microbiol 9, art. no. 225
Kandhai MC, Reij MW, Gorris LG, Guillaume-Gentil O,
Schothorst M (2004) Occurrence of Enterobacter sak-azakii in food production environments and households.
Lancet 363:39–40
Kucerova E, Clifton SW, Xia X-Q, Long F, Porwollik S, Fulton
L, Fronick C, Minx P, Kyung K, Warren W, Fulton R,
Feng D, Wollam A, Shah N, Bhonagiri V, Nash WE,
Hallsworth-Pepin K, Wilson RK, McClelland M,
Forsythe SJ (2010) Genome sequence of Cronobactersakazakii BAA-894 and comparative genomic hybridiza-
tion analysis with other Cronobacter species. PLoS ONE
5, art. no. e9556
Kuhnert P, Korczak BM, Stephan R, Joosten H, Iversen C
(2009) Phylogeny and prediction of genetic similarity of
Cronobacter and related taxa by multilocus sequence
analysis (MLSA). Int J Food Microbiol 136:152–158
Lai KK (2001) Enterobacter sakazakii infections among neo-
nates, infants, children, and adults: case reports and a
review of the literature. Medicine 80:113–122
Lane DJ (1991) 16S/23S rRNA sequencing. In: Stackenbrandt E,
Goodfellow M (eds) Nucleic acid techniques in bacterial
systematics. Wiley, New York, pp 115–148
Lehner A, Tasara T, Stephan R (2004) 16S rRNA gene based
analysis of Enterobacter sakazakii strains from different
sources and development of a PCR assay for identifica-
tion. BMC Microbiol 4:43–49
Mullane NR, Iversen C, Healy B, Walsh C, Whyte P, Wall PG,
Quinn T, Fanning S (2007) Enterobacter sakazakii an
emerging bacterial pathogen with implications for infant
health. Minerva Pediatr 59:137–148
Nazarowec-White M, Farber JM (1997) Thermal resistance of
Enterobacter sakazakii in reconstituted dried-infant for-
mula. Lett Appl Microbiol 24:9–13
268 Antonie van Leeuwenhoek (2011) 99:257–269
123

O’Brien S, Healy B, Negredo C, Fanning S, Iversen C (2009a)
Evaluation of a new one-step enrichment in conjunction
with a chromogenic medium for the detection of Cro-nobacter spp. (Enterobacter sakazakii) in powdered infant
formula. J Food Prot 72:1472–1475
O’Brien S, Healy B, Negredo C, Anderson W, Fanning S,
Iversen C (2009b) Prevalence of Cronobacter species
(Enterobacter sakazakii) in follow-on infant formulae and
infant drinks. Lett Appl Microbiol 48:536–541
Osaili T, Forsythe S (2009) Desiccation resistance and per-
sistence of Cronobacter species in infant formula. Int J
Food Microbiol 136:214–220
Pangallo D, Drahovska H, Harichova J, Karelova E, Chova-
nova K, Aradska J, Ferianc P, Turna J, Timko J (2008)
Evaluation of different PCR-based approaches for the
identification and typing of environmental enterococci.
Antonie van Leeuwenhoek 93:193–203
Ray P, Das A, Gautam V, Jain N, Wig JD, Sharma M (2007)
Postoperative nosocomial Enterobacter sakazakii sepsis.
ANZ J Surg 77:915–916
Schmid M, Iversen C, Gontia I, Stephan R, Hofmann A,
Hartmann A, Jha B, Eberl L, Riedel K, Lehner A (2009)
Evidence for a plant-associated natural habitat for Cro-nobacter spp. Res Microbiol 160:608–614
See KC, Than HA, Tang T (2007) Enterobacter sakazakiibacteraemia with multiple splenic abscesses in a 75-year
old woman: a case report. Age Ageing 36:595–596
Seo KH, Brackett RE (2005) Rapid, specific detection of
Enterobacter sakazakii in infant formula using a real-time
PCR. J Food Prot 68:59–63
Stoop B, Lehner A, Iversen C, Fanning S, Stephan R (2009)
Development and evaluation of rpoB based PCR systems
to differentiate the six proposed species within the genus
Cronobacter. Int J Food Microbiol 136:165–168
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4:
Molecular Evolutionary Genetics Analysis (MEGA)
software version 4.0. Mol Biol Evol 24:1596–1599
Townsend S, Hurrell E, Forsythe S (2008) Virulence studies of
Enterobacter sakazakii isolates associated with a neonatal
intensive care unit outbreak. BMC Microbiol 8:64–72
Van Acker J, de Smet F, Muyldermans G, Bougatef A, Naes-
sens A (2001) Outbreaks of necrotizing enterocolitis
associated with Enterobacter sakazakii in powdered milk
formula. J Clin Microbiol 39:293–297
Antonie van Leeuwenhoek (2011) 99:257–269 269
123
Related Documents











