Biochemistry 2 Spr 2015; Lecture TCA ( Krebs) & Glyoxylate Cycle There is no reason for this except I love Paris!

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochemistry 2 Spr 2015; Lecture TCA ( Krebs) & Glyoxylate Cycle
There is no reason for this except I love Paris!!

What flows from the TCA cycle to ETS is the NADH. It is the oxidation of NADH drives the production of ATP. The TCA intermediates function in other pathways.

THE starting substrate for the TCA cycle is acetylCoA, that can be derived from pyruvate or fatty acids through β-oxidation. • Why is this called an amphibolic pathway? • Is this only found in aerobes? • In aerobes it is an oxidative pathway, in anaerobes is it a reductive pathway? 3NAD+ + FAD+ +GDP + Pi + Acetyl-CoA 3NADH + FADH2 + GTP + CoA + 2CO2

Electron micrographs of the E. coli pyruvate dh multienzyme complex, approx 300 angrstroms. Range of size is 300 to 500 A. Stucturally as well as mechanistically there is not difference between the prokaryotic Pdh & the eukaryotic Pdh. The difference is in the number of peptides of each specific peptide in the protein. The Pdh is a model for Keto acid dh, such as αKGdh is an identical structure but E1 has a different specificity.

The product of this pathway is a highly reactive substrate for the TCA cycle, acetyl-CoA. It will condense with OAA to form citrate. The rate limiting substrate for this pathway is NAD+. It is reduced by the enzyme lipoamide dh, and then feeds its H+ (NADH is oxidized) into the ETS to generate ATP. 4H+ produced reduces 1 O2 molecule.


This pathway is a multienzyme complex that functions as a structure. The relationship of E1, E2 & E3 and how lipoamide acts as a structural extension of the transacetylase.

Lipoamide forms a 14A arm that binds the acetyl group that is transferred from TPP in the the E1 active site, and bring it to the E2 active site where the acetyl group is transferred to CoA. It is a transfer from the thiol of lipoamide to the thiol of CoA.

Structure of E1 from P. putida branched-chain a-keto acid
dehydrogenase. An a2b2 heterotetrameric protein.
Structure of dihydrolipoamide dehydrogenase (E3) from P. putida in complex with FAD+ and NAD+. A homodimeric enzyme.

Structure of E! from P. putida branched-chain a-keto acid dehydrogenase. A surface diagram of the active site
region.
Structure of dihydrolipoamide dehydrogenase (E3) from P. putida in complex with FAD+ and NAD+. The enzyme’s active site region. Cys 43 & 48 are found on an destorted α-helixes and a deep FAD+ binding site with NAD+ very close. The phenolic group of Tyr 181 protects the e- transfer. FAD+ has a reduction potential of V≈0, and NAD+ Εo’ of -0.315V.

Structure of a trimer of A. vinelandii dihydrolipoyl transacetylase (E2) catalytic domains. Lipoamide is assoc with Lys residue on E2 protein.



Factors controlling the activity of the Pdh product inhibition.
Factors controlling the activity of the Pdh. Covalent modification in the eukaryotic complex.

Factors regulating the activity of pyruvate dehydrogenase, (Pdh). Pdh activity is regulated by its state of phosphorylation, being most active in the dephosphorylated state. Phosphorylation of Pdh is catalyzed by a specific Pdh kinase. The activity of the kinase is enhanced when cellular energy charge is high which is reflected by an increase in the level of ATP, NADH and acetyl-CoA. Conversely, an increase in pyruvate strongly inhibits Pdh kinase. Additional negative effectors of Pdh kinase are ADP, NAD+ and CoASH, the levels of which increase when energy levels fall. The regulation of Pdh phosphatase is not completely understood but it is known that Mg2+ and Ca+ activate the enzyme. In adipose tissue insulin increases Pdh activity and in cardiac muscle Pdh activity is increased by catecholamines.

CONTROL, REGULATION AND ACTIVITY OF Pdh. Two products of the complex, NADH and acetyl-CoA, are negative allosteric effectors on Pdh-a, the non-phosphorylated, active form of Pdh. These effectors reduce the affinity of the enzyme for pyruvate, thus limiting the flow of carbon through the Pdh complex. In addition, NADH and acetyl-CoA are powerful positive effectors on Pdh kinase, the enzyme that inactivates Pdh by converting it to the phosphorylated Pdh-b form. Since NADH and acetyl-CoA accumulate when the cell energy charge is high, it is not surprising that high ATP levels also up-regulate Pdh kinase activity, reinforcing down-regulation of Pdh activity in energy-rich cells. Note, however, that pyruvate is a potent negative effector on Pdh kinase, with the result that when pyruvate levels rise, Pdh-a will be favored even with high levels of NADH and acetyl-CoA. Concentrations of pyruvate which maintain Pdh in the active form (Pdh-a) are sufficiently high so that, in energy-rich cells, the allosterically down-regulated, high Km form of Pdh is nonetheless capable of converting pyruvate to acetyl-CoA. With large amounts of pyruvate in cells having high energy charge and high NADH, pyruvate carbon will be directed to the 2 main storage forms of carbon---glycogen via gluconeogenesis and fat production via fatty acid synthesis---where acetyl-CoA is the principal carbon donor. Although the regulation of Pdh-b phosphatase is not well understood, it is quite likely regulated to maximize pyruvate oxidation under energy-poor conditions and to minimize Pdh activity under energy-rich conditions. The enzyme complex then is the gate keeper for the entrance into the TCA cycle and can be considered one of the rate-limiting steps ultimately of aerobic respiration.

The TCA cycle showing enzymes, substrates and products. The GTP generated during the succinate thiokinase (succinyl-CoA synthetase) reaction is equivalent to a mole of ATP by virtue of the presence of nucleoside diphosphokinase. The 3 moles of NADH and 1 mole of FADH2 generated during each round of the cycle feed into the oxidative phosphorylation pathway. Each mole of NADH leads to 3 moles of ATP and each mole of FADH2 leads to 2 moles of ATP. Therefore, for each mole of pyruvate which enters the TCA cycle, 12 moles of ATP can be generated.
The overall stoichiometry of the TCA cycle is: acetyl-CoA + 3NAD+ + FAD+ + GDP + Pi + 2H2O ----> 2CO2 + 3NADH + FADH2 + GTP + 2H+ + HSCoA
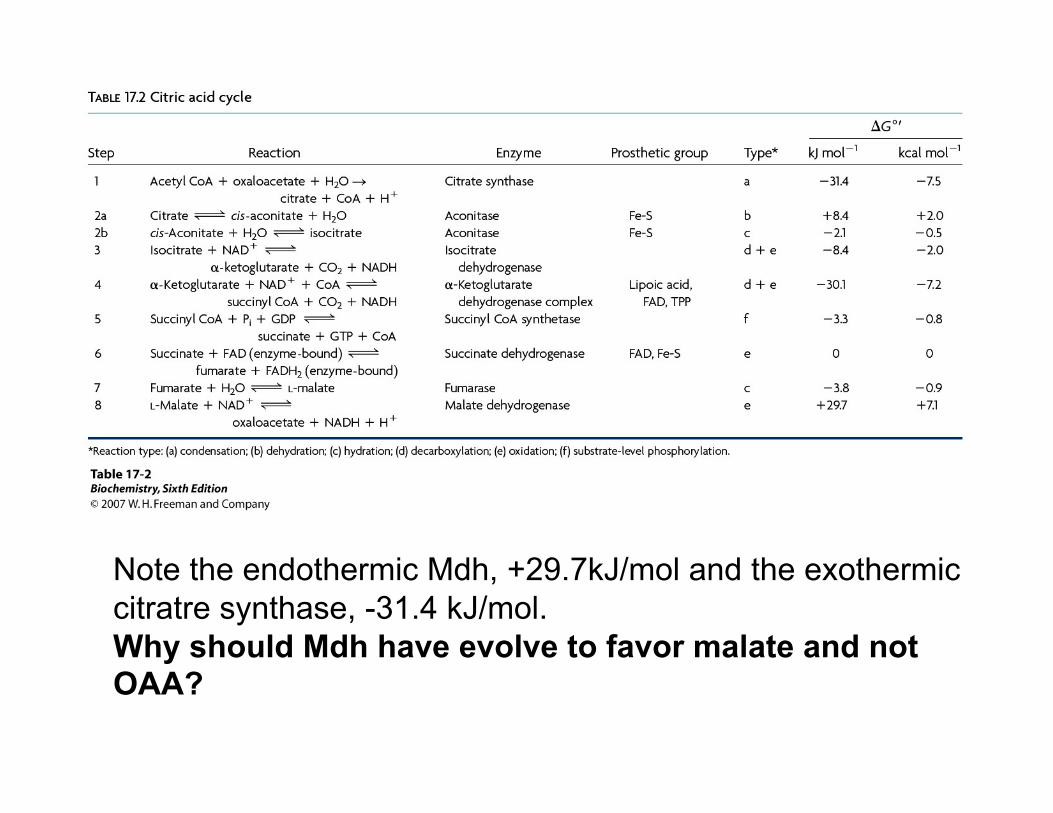
Note the endothermic Mdh, +29.7kJ/mol and the exothermic citratre synthase, -31.4 kJ/mol. Why should Mdh have evolve to favor malate and not OAA?

Regulation of the citric acid cycle.
Standard Free Energy changes (ΔG°) and physiological Free Energy changes (ΔG) of citric acid cycle reactions.

Structure of citrate synthase. The flexible domain of each subunit undergoes a large conformational change on binding OAA, creating a binding site for acetyl-CoA. (a) Open form of the enzyme alone; (b) closed form with bound OAA and a stable analog of acetyl-CoA. In these representations one subunit is colored tan and one green.

MECHANISM; Citrate synthase.
In the mammalian citrate synthase rxn,OAA binds first, in a strictly ordered reaction sequence. This binding triggers a conformation change that opens up the binding site for acetyl-CoA. OAA is specifically oriented in the active site of citrate synthase by interaction of its two carboxylates with two positively charged Arg residues (not shown here).

Iron-sulfur center in aconitase. The iron-sulfur center is in red, the citrate molecule in blue. Three Cys residues of the enzyme bind three iron atoms; the fourth iron is bound to one of the carboxyl groups of citrate and also interacts non-covalently with a hydroxyl group of citrate (dashed bond). A basic residue (:B) in the enzyme helps to position the citrate in the active site. The iron-sulfur center acts in both substrate binding and catalysis. This reaction is to convert the tertiary alc to a an easily oxidized secondary alc, isocitrate.

Isocitrate dehydrogenase. In this reaction, the substrate, isocitrate, loses one carbon by oxidative decarboxylation. This occurs with the oxidation of isocitrate to oxalosuccinate Reducing NAD+ to NADH, then the decarboxylation of oxalosucc to α-KG.

This is another member of the Keto acid dh enzymes like Pdh. The E1 is the αKG dh.

The succinyl-CoA synthetase reaction. In step 1 a phosphoryl group replaces the CoA of succinyl-CoA bound to the enzyme, forming a high-energy acyl phosphate. In step 2 the succinyl phosphate donates its phosphoryl group to a His residue of the enzyme, forming a high-energy phosphohistidyl enzyme. In step 3 the phosphoryl group is transferred from the His residue to the terminal phosphate of GDP (or ADP), forming GTP (or ATP).

The succinyl-CoA synthetase reaction. Active site of succinyl-CoA synthetase of E. coli. The active site includes part of both the α (blue) and the β (brown) subunits. The power helices (blue, brown) place the partial positive charges of the helix dipole near the phosphate group of P–His246 in the α chain, stabilizing the phosphohistidyl enzyme. The bacterial and mammalian enzymes have similar amino acid sequences and three-dimensional structures.

In the reverse rxn this is fumerate reductase. Facultative aerobes Like E.coli have both a succinate dh & fumerate reductase gene, but structually they are identical. In the mitochondria this is a trans-membrane bound protein that is part of electron transport, complex II.

3NAD + FAD +GDP + Pi + Acetyl-CoA 3NADH + FADH2 + GTP + CoA + 2CO2
The NADH and FADH formed are the substrates for the ETS and oxidative- phoshorylation.

Human cells contain almost equal amounts of mitochondrial and cytosolic PEPCK so this second reaction can occur in either cellular compartment. For gluconeogenesis to proceed, the OAA produced by PC needs to be transported to the cytosol. However, no transport mechanism exist for its' direct transfer and OAA will not freely diffuse. Mitochondrial OAA can become cytosolic via three pathways, conversion to PEP (as indicated above through the action of the mitochondrial PEPCK), transamination to aspartate or reduction to malate, all of which are transported to the cytosol. If OAA is converted to PEP by mitochondrial PEPCK, it is transported to the cytosol where it is a direct substrate for gluconeogenesis and nothing further is required. Transamination of OAA to aspartate allows the aspartate to be transported to the cytosol where the reverse transamination occurs yielding cytosolic OAA. This transamination reaction requires continuous transport of glutamate into, and a-ketoglutarate out of, the mitochondrion. Therefore, this process is limited by the availability of these other substrates. Either of these latter two reactions will predominate when the substrate for gluconeogenesis is lactate. Whether mitochondrial decarboxylation or transamination occurs is a function of the availability of PEPCK or transamination intermediates. Mitochondrial OAA can also be reduced to malate in a reversal of the TCA cycle reaction catalyzed by malate dehydrogenase (MDH). The reduction of OAA to malate requires NADH, which will be accumulating in the mitochondrion as the energy charge increases. The increased energy charge will allow cells to carry out the ATP costly process of gluconeogenesis. The resultant malate is transported to the cytosol where it is oxidized to OAA by cytosolic MDH which requires NAD+ and yields NADH.

What is the reason for this shuttle pathway? How does this pathway give the cell an advantage?



The NADH produced during the cytosolic oxidation of malate to OAA. Conversion of pyruvate to PEP requires the action of two mitochondrial enzymes. The first is an ATP-requiring reaction catalyzed by pyruvate carboxylase, (PC). As the name of the enzyme implies, pyruvate is carboxylated to form OAA. The CO2 in this reaction is in the form of bicarbonate (HCO3
2-) . This reaction is an anaplerotic reaction since it can be used to fill-up the TCA cycle. The second enzyme in the conversion of pyruvate to PEP is PEP carboxykinase (PEPCK). PEPCK requires GTP in the decarboxylation of OAA to yield PEP. Since PC incorporated CO2 into pyruvate and it is subsequently released in the PEPCK reaction, no net OAA is utilized during the glyceraldehyde-3-phosphate dehydrogenase reaction of glycolysis. The coupling of these two oxidation-reduction reactions is required to keep gluconeogenesis functional when pyruvate is the principal source of carbon atoms. The conversion of OAA to malate predominates when pyruvate (derived from glycolysis or amino acid catabolism) is the source of carbon atoms for gluconeogenesis. When in the cytoplasm, OAA is converted to PEP by the cytosolic version of PEPCK. Hormonal signals control the level of PEPCK protein as a means to regulate the flux through gluconeogenesis.



A significant amount of metabolic energy can come from amino acid metabolism, particularly under conditions of starvation. The metabolism of amino acids occurs through common metabolic intermediates, many of them part of or linked to the Krebs cycle. The intermediates like pyruvate, oxaloacetate, fumarate, succinyl-CoA and a-ketobutyrate all can contribute to the net synthesis of glucose through gluconeogenesis. The amino acids that are degraded into these intermediates are called glucogenic. Some amino acids are degraded directly to gluconeogenic intermediates, while others contribute to gluconeogenesis more indirectly, such as through conversion to oxaloacetate through the Krebs cycle. Other common intermediates in amino acid metabolism are acetyl-CoA and acetoacetate. Animals do not have a metabolic pathway to convert these intermediates into glucose. Instead the metabolic fate of acetyl-CoA and acetoacetate is the production of fatty acids or ketone bodies. Amino acids that are metabolized to produce acetyl-CoA and acetoacetate are called ketogenic. Several of the amino acids do not fall cleanly into one group or another, but are both ketogenic and glucogenic. For example, isoleucine metabolism produces both acetyl-CoA, which makes it ketogenic, but it also produces succinyl-CoA, which contributes to glucose production.



The amino acids may be grouped for discussion on the basis of the specific keto acid products of their deamination. The 3-carbon a-keto acid pyruvate is produced from alanine, cysteine, glycine, serine, & threonine. Alanine deamination via Transaminase directly yields pyruvate

The 4-carbon Krebs Cycle intermediate oxaloacetate is produced from aspartate & asparagine. Aspartate deamination via transaminase directly yields oxaloacetate . Aspartate also is converted to fumarate in the Urea Cycle. Fumarate is then converted by Krebs Cycle enzymes in the cytosol (fumerase) to malate, and by Mdh to oxaloacetate.

The 4-carbon Krebs Cycle intermediate succinyl-CoA is produced from isoleucine, valine, & methionine. Propionyl-CoA, which is an intermediate on these pathways, is also a product of β-oxidation of fatty acids with an odd number of C atoms.

The 5-carbon Krebs Cycle intermediate a-ketoglutarate is produced from arginine, glutamate, glutamine, histidine, and proline. Glutamate deamination, via glutamate dehydrogenase or transaminase, directly yields α-ketoglutarate
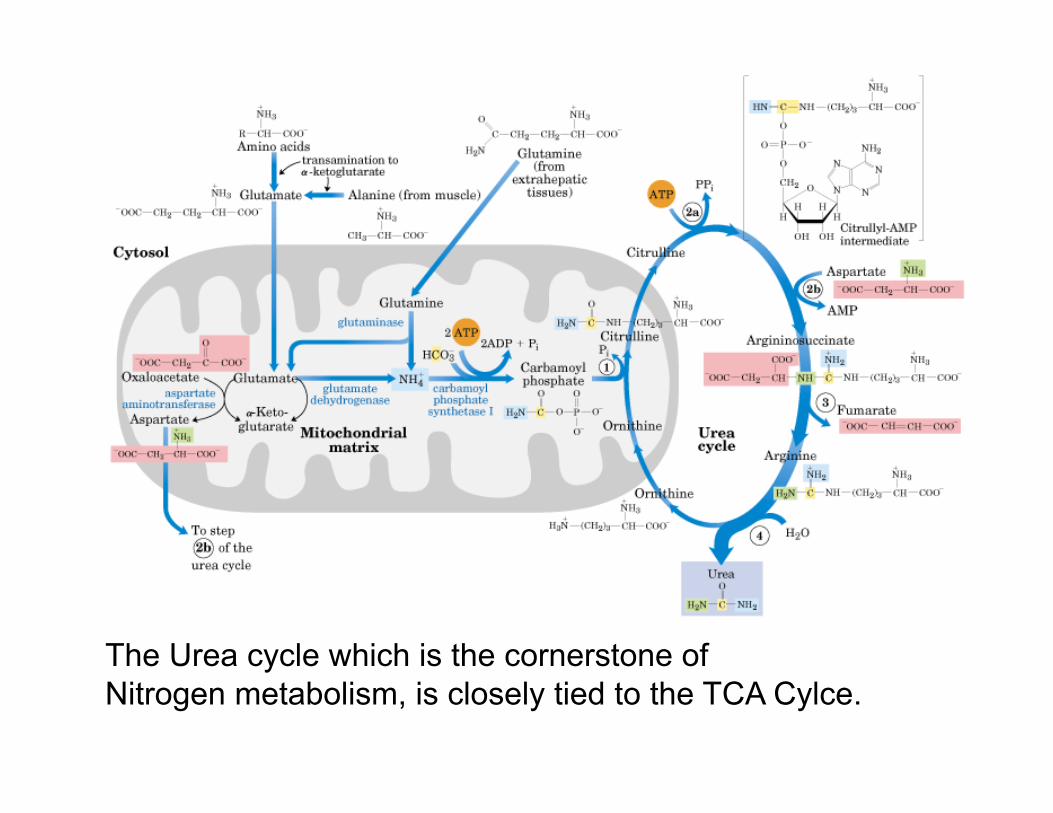
The Urea cycle which is the cornerstone of Nitrogen metabolism, is closely tied to the TCA Cylce.


Glyoxylate cycle!


What makes this pathway Unique is that it produces Succinate & glyoxylate from Isocitrate. It is An incomplete TCA cycle that has two unique enzymes? What are they? What advantage does this pathway give to a cell?

Isocitrate lyase is an enzyme that functions at a branch point of carbon metabolism and diverts isocitrate through a carbon-conserving pathway, the glyoxylate cycle, bypassing the two decarboxylative steps of the tricarboxylic acid cycle that convert isocitrate to succinyl-CoA. The glyoxylate molecule formed by isocitrate lyase reaction condenses with one acetyl-CoA to produce L-malate in the subsequent step catalyzed by malate synthase (the other enzyme tipical of the glyoxylate cycle). acetyl-CoA + glyoxylate + H2O = L-malate + CoA

The isocitrate used in the first reaction of the glyoxylate cycle is restored by the action of three enzymes characteristic of Krebs cycle (malate dehydrogenase, citrate synthase and cis-aconitase) on L-malate, with the utilization of a second molecule of acetyl-CoA. The glyoxylate cycle thus enables the synthesis of a mole of succinate from two moles of acetate (as acetyl-CoA), being the overall net reaction: 2 acetyl-CoA + 2H2O + NAD+ = succinate + 2CoA + NADH + H+
The glyoxylate cycle replenishes intermediates of the Krebs cycle and conserves carbon that would otherwise be oxidized and lost to biosynthetic pathways, with the final result of a net conversion of fats to carbohydrates. This replenishing function has been termed anaplerotic (Kornberg, 1966) and probably plays an essential role in growth of microorganisms on fatty acids, in germination of oil-rich seedlings and in development of certain animal embryos. Isocitrate lyase is an enzyme that functions at a branch point of carbon metabolism and diverts isocitrate through a carbon-conserving pathway, the glyoxylate cycle, bypassing the two decarboxylative steps of the tricarboxylic acid cycle that convert isocitrate to succinyl-CoA.

In what organisms would this sort of TCA cycle function? It appears that the two main products of this pathway are succinyl-CoA and αKG.
We will come back to the TCA Cycle in numerous future chapters.

IN a reductive pathway what are the possible end products?
PNAS-2004-Smith

PNAS-2004-Smith
Related Documents