Bioactivity studies on atypical natural opioid hexapeptides processed from proenkephalin (PENK) precursor polypeptides Engin Bojnik a,1 , Patrycja Kleczkowska b , Ezequiel Marron Fernandez de Velasco c , Maïthé Corbani d , Fruzsina Babos e , Andrzej W. Lipkowski b , Anna Magyar e , Sandor Benyhe a, ⁎ a Institute of Biochemistry, Biological Research Centre, Hungarian Academy of Sciences, 6726 Szeged, Temesvari krt 62, Hungary b Mossakowski Medical Research Centre, Polish Academy of Sciences, Department of Neuropeptides, Warsaw, Poland c Department of Pharmacology, University of Minnesota, Minneapolis, MN 55455, USA d Institue of Functional Genomics, CNRS UMR 5203-INSERM U.661, University of Montpellier, 141 rue de la Cardonille, 34094 Montpellier Cedex 05, France e MTA-ELTE Research Group of Peptide Chemistry, Hungarian Academy of Sciences, Eötvös Lorand University, Budapest, Hungary abstract article info Article history: Received 3 February 2014 Received in revised form 2 June 2014 Accepted 6 June 2014 Available online 16 June 2014 Keywords: Enkephalin hexapeptides Proenkephalin evolution Natural chemical library Mu and delta opioid receptors Species differences Endogenous opioids are derived from four related polypeptide precursors: proenkephalin (PENK), prodynorphin (PDYN), pronociceptin (PNOC) and proopiomelanocortin (POMC). In mammals PENK encodes for four copy of Met-enkephalin, one octapeptide Met-enkephalin-Arg-Gly-Leu, one heptapeptide Met-enkephalin-Arg-Phe and a single copy of Leu-enkephalin. Our detailed bioinformatic search on the existing PENK sequences revealed several atypical hexapeptide Met-enkephalins in different vertebrate animals. They are located either in the second enkephalin unit or in the seventh enkephalin core position at the C-terminus. Altogether four different hexapeptide sequences were obtained representing eleven animal species: Met-enkephalin-Arg 6 (YGGFM R) in the bird zebra finch, Met-enkephalin-Asp 6 (YGGFM D), Met-enkephalin-Ile 6 (YGGFM I) in zebrafish; and Met-enkephalin-Ser 6 (YGGFM S) in two pufferfish species. All novel peptides were chemically synthesized and studied in receptor binding and G-protein activation assays performed on rat brain membranes. The four novel enkephalins were equipotent in stimulating G-proteins. Affinities of the peptides determined by equilibrium competition assays in receptor binding experiments were statistically different. At the MOP receptors the highest affinity (K i 4 nM) was obtained with the zebra finch peptide Met-enkephalin-Arg 6 . The pufferfish Met-enkephalin-Ser 6 ex- hibited the highest affinity (K i 6.7 nM) at the DOP receptor. Phylogenetic neuropeptide libraries, defined here as a collection of mutationally different species variants of orthologous and paralogous peptide sequences, represent the natural molecular diversity of the neuropeptides. Such libraries can provide a wide range of structural information establishing comparative functional analyses. Since DNA sequencing data are rapidly increasing, more development in the natural peptide library concept is expected. © 2014 Elsevier Inc. All rights reserved. 1. Introduction The opioid system is involved in pain and reward mechanisms and consists of four receptors and several peptides. In mammals the mu- (MOP), kappa- (KOP), delta- (DOP) and ORL-1 (NOP) receptor genes constitute the opioid/ORL-1 receptor gene family. Endogenous ligands for these receptors are the opioid peptides. Natural opioid pep- tides are derived from four polypeptide precursors: proenkephalin (PENK) (Noda et al., 1982; Gubler et al., 1982), prodynorphin (PDYN) (Kakidani et al., 1982; Horikawa et al., 1983), pronociceptin (PNOC) (Mollereau et al., 1996; Nothacker et al., 1996) and proopiomelanocortin (POMC) (Nakanishi et al., 1979). Enkephalins are endogenous pentapep- tides with opioid receptor activity (Hughes et al., 1975). The source for these naturally occurring sequences is the precursor polypeptide termed proenkephalin (PENK) (Comb et al., 1982; Noda et al., 1982). A second- ary source for Leu-enkephalins is the α-neo-endorphin/dynorphin A/ rimorphin precursor PDYN (Traynor, 1987). The structural organization of PENK includes an N-terminal signal peptide sequence followed by a cysteine rich domain and then the coding region for the mature oligopeptides interrupted by short non-processing segments. Mammali- an PENK contains seven copies of enkephalins bordered by basic dipep- tide repeats, such as Arg-Arg, Lys-Lys or Lys-Arg (RR, KK, KR). These dibasic residues represent recognition sites for the processing endopep- tidase enzymes, called prohormone convertases (Hook et al., 2008). Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35 ⁎ Corresponding author at: Institute of Biochemistry, Biological Research Centre, Hungarian Academy of Sciences, H-6701 Szeged, P.O. Box 521, Hungary. Tel.: +36 62 432 099; fax: +36 62 433 432. E-mail address: [email protected] (S. Benyhe). 1 Current address: Department of Surgery, Oncology and Gastroenterology, Section of Oncology and Immunology, University of Padova, Padova, Italy. http://dx.doi.org/10.1016/j.cbpb.2014.06.002 1096-4959/© 2014 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Comparative Biochemistry and Physiology, Part B journal homepage: www.elsevier.com/locate/cbpb

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part B
j ourna l homepage: www.e lsev ie r .com/ locate /cbpb
Bioactivity studies on atypical natural opioid hexapeptides processedfrom proenkephalin (PENK) precursor polypeptides
Engin Bojnik a,1, Patrycja Kleczkowska b, Ezequiel Marron Fernandez de Velasco c, Maïthé Corbani d,Fruzsina Babos e, Andrzej W. Lipkowski b, Anna Magyar e, Sandor Benyhe a,⁎a Institute of Biochemistry, Biological Research Centre, Hungarian Academy of Sciences, 6726 Szeged, Temesvari krt 62, Hungaryb Mossakowski Medical Research Centre, Polish Academy of Sciences, Department of Neuropeptides, Warsaw, Polandc Department of Pharmacology, University of Minnesota, Minneapolis, MN 55455, USAd Institue of Functional Genomics, CNRS UMR 5203-INSERM U.661, University of Montpellier, 141 rue de la Cardonille, 34094 Montpellier Cedex 05, Francee MTA-ELTE Research Group of Peptide Chemistry, Hungarian Academy of Sciences, Eötvös Lorand University, Budapest, Hungary
⁎ Corresponding author at: Institute of BiochemistrHungarian Academy of Sciences, H-6701 Szeged, P.O. B432 099; fax: +36 62 433 432.
E-mail address: [email protected] (S. Benyhe).1 Current address: Department of Surgery, Oncology a
Oncology and Immunology, University of Padova, Padova
http://dx.doi.org/10.1016/j.cbpb.2014.06.0021096-4959/© 2014 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 3 February 2014Received in revised form 2 June 2014Accepted 6 June 2014Available online 16 June 2014
Keywords:Enkephalin hexapeptidesProenkephalin evolutionNatural chemical libraryMu and delta opioid receptorsSpecies differences
Endogenous opioids are derived from four related polypeptide precursors: proenkephalin (PENK), prodynorphin(PDYN), pronociceptin (PNOC) and proopiomelanocortin (POMC). In mammals PENK encodes for four copy ofMet-enkephalin, one octapeptide Met-enkephalin-Arg-Gly-Leu, one heptapeptide Met-enkephalin-Arg-Pheand a single copy of Leu-enkephalin. Our detailed bioinformatic search on the existing PENK sequences revealedseveral atypical hexapeptideMet-enkephalins in different vertebrate animals. They are located either in the secondenkephalin unit or in the seventh enkephalin core position at the C-terminus. Altogether four different hexapeptidesequences were obtained representing eleven animal species: Met-enkephalin-Arg6 (YGGFMR) in the bird zebrafinch, Met-enkephalin-Asp6 (YGGFMD), Met-enkephalin-Ile6 (YGGFMI) in zebrafish; and Met-enkephalin-Ser6
(YGGFMS) in two pufferfish species. All novel peptides were chemically synthesized and studied in receptorbinding and G-protein activation assays performed on rat brain membranes. The four novel enkephalins wereequipotent in stimulating G-proteins. Affinities of the peptides determined by equilibrium competition assaysin receptor binding experiments were statistically different. At the MOP receptors the highest affinity(Ki 4 nM)was obtainedwith the zebra finch peptideMet-enkephalin-Arg6. The pufferfishMet-enkephalin-Ser6 ex-hibited the highest affinity (Ki 6.7 nM) at the DOP receptor. Phylogenetic neuropeptide libraries, defined here as acollection ofmutationally different species variants of orthologous and paralogous peptide sequences, represent thenatural molecular diversity of the neuropeptides. Such libraries can provide a wide range of structural informationestablishing comparative functional analyses. SinceDNA sequencing data are rapidly increasing,more developmentin the natural peptide library concept is expected.
© 2014 Elsevier Inc. All rights reserved.
1. Introduction
The opioid system is involved in pain and reward mechanismsand consists of four receptors and several peptides. In mammalsthe mu- (MOP), kappa- (KOP), delta- (DOP) and ORL-1 (NOP) receptorgenes constitute the opioid/ORL-1 receptor gene family. Endogenousligands for these receptors are the opioid peptides. Natural opioid pep-tides are derived from four polypeptide precursors: proenkephalin
y, Biological Research Centre,ox 521, Hungary. Tel.: +36 62
nd Gastroenterology, Section of, Italy.
(PENK) (Noda et al., 1982; Gubler et al., 1982), prodynorphin (PDYN)(Kakidani et al., 1982; Horikawa et al., 1983), pronociceptin (PNOC)(Mollereau et al., 1996; Nothacker et al., 1996) and proopiomelanocortin(POMC) (Nakanishi et al., 1979). Enkephalins are endogenous pentapep-tides with opioid receptor activity (Hughes et al., 1975). The source forthese naturally occurring sequences is the precursor polypeptide termedproenkephalin (PENK) (Comb et al., 1982; Noda et al., 1982). A second-ary source for Leu-enkephalins is the α-neo-endorphin/dynorphin A/rimorphin precursor PDYN (Traynor, 1987). The structural organizationof PENK includes an N-terminal signal peptide sequence followed by acysteine rich domain and then the coding region for the matureoligopeptides interrupted by short non-processing segments. Mammali-an PENK contains seven copies of enkephalins bordered by basic dipep-tide repeats, such as Arg-Arg, Lys-Lys or Lys-Arg (RR, KK, KR). Thesedibasic residues represent recognition sites for the processing endopep-tidase enzymes, called prohormone convertases (Hook et al., 2008).

30 E. Bojnik et al. / Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35
Upon cleavage terminal basic amino acids are typically removed. Sevenopioid coremotifs existwithin PENK andMet-enkephalin units are pres-ent as repetitive sequences with a copy number of six while one singlecopy of Leu-enkephalin is also present at the sixth core position. Besidethe pentapeptides, two extendedMet-enkephalin sequence forms exist:the fourth opioid motif is the octapeptide Met-enkephalin-Arg-Gly-Leu(MERGL) and the PENK C-terminal is indeed a heptapeptide with thestructure of Met-enkephalin-Arg-Phe (MERF).
Among multiple opioid peptides the enkephalins have the widesttissue distribution. According to phylogenetic analyses the PENK geneis considered to be the primordial opioid propeptide gene (Doreset al., 1993). This is the ‘proenkephalin hypothesis’. Gene duplicationis a recurring theme in the evolution of vertebrate polypeptidehormones and neuropeptides. It has been proposed that during the evo-lution of the opioid peptides at least two duplication events occurred inthe most ancestral PENK gene (Dores et al., 1993). Consistent with theproenkephalin hypothesis, the first gene duplication event wouldhave produced the POMC gene, which is the common precursor forthe non-opioid adrenocorticotrop hormone (ACTH) and the threemelanocyte stimulating hormones (α, β, γ MSHs) and the opioiduntriacontapeptide β-endorphin. A further duplication event wouldhave given rise to the PDYN gene. The proenkephalin hypothesisand the evolution of the opioid peptide precursor genes have beendescribed in more details (Khalap et al., 2005; Roberts et al., 2007;Komorowski et al., 2012). Another scenario for gene duplications hasbeen proposed more recently. Opioid peptide gene family memberswere investigated using a combination of sequence-based phylogenyand chromosomal locations of the peptide genes in various vertebrates(Sundstrom et al., 2010). The results showed that the ancestral PENKgene gave rise to two additional copies in the genome doublings. Thefourth member was generated by a local gene duplication, as thegenes encoding POMC and PNOC are located on the same chromosomein several teleost and bird genomes that have been studied. In conclu-sion, the system of opioid peptides and receptors was largely formedby the genome doublings that took place early in vertebrate evolution.
Opioid precursor polypeptides are well conserved, yet they exhibitremarkable structural variations even close to the opioid core sequencemotifs. As a result, the set of mammalian enkephalins (Met-enkephalin,Leu-enkephalin, Met-enkephalin-Arg-Gly-Leu and Met-enkephalin-Arg-Phe) can be extended mutationally by novel variant members. Thus,two new pentapeptides Ile-enkephalin (YGGFI) and Phe-enkephalin(YGGFF) have been described from PDYN cDNA sequences of Africanclawed frog and African lungfish, respectively (Bojnik et al., 2009).Analyzing the fourth octapeptide region of PENK revealed the existenceof four novel orthologues including YGGFMRSL (mouse, Australian lung-fish, African lungfish), YGGFMRGY (three frog species), YGGFMRSV(chicken) and YGGFMNGF (bullhead shark). All four octapeptides werechemically synthesized and studied in biochemical pharmacologicalassays (Bojnik et al., 2010). Further bioinformatic analysis of theC-terminal tail in PENK of 56 vertebrates revealed new orthologues tothe heptapeptide Met-enkephalin-Arg-Phe (Bojnik et al., 2011). Fournovel heptapeptides with single or double amino acid replacementswere identified among 15 animals, such as YGGFMGY (zebrafish),YGGFMRY(newt), YGGFMKF (hedgehog tenrek) andYGGFMRI (commonmudpuppy).
Previously, we have identified novel opioid penta-, hepta- and octa-peptides within PENK sequences among different animal species andhave shown that they possess good tomoderate opioid receptor bindingaffinity (Bojnik et al., 2009; 2010; 20121). Recent database [NCBI directsubmissions by Wada,K. and Jarvis,E.D. 2005; Dores et al., 2002;Gonzalez-Nuñez et al., 2003)] searching for PENK sequences revealedthe presence of unusual hexapeptide enkephalins in various vertebrateanimals. The aim of the current study was to collect all presently avail-able PENK encoded hexapeptide enkephalin sequences from differentspecies. The bioinformatically identified sequence variants were chem-ically synthesized and studied further in biochemical pharmacological
assays. Here we describe and compare the bioactivity pattern of fourdistinct novel peptides that occur naturally in vertebrates and locatedat the second or seventh opioid core positions of PENK.
2. Materials and methods
2.1. Animals
Albino Wistar rats (Rattus norvegicus; 250–300 g body mass) weremaintained in the local animal house of the Biological Research Center(BRC, Szeged, Hungary). Animals were kept in groups of six, allowedfree access to tap water and standard rodent food and kept on a12:12-h dark/light cycle until the time of sacrifice. Animal studieswere controlled according to the Hungarian Act for the Protection ofAnimals in Research (XXVIII.tv. 32.§) and the European CommunitiesCouncil Directives (86/609/ECC). The number of rats and their sufferingwere minimized during our experiments.
2.2. Chemicals and radioligands
[3H]DAMGO ([D-Ala2,NMePhe4,Gly5-ol]enkephalin; 41 Ci/mmol) and[3H]DIDII (Tyr1,Ile5,6deltorphin-2; 48 Ci/mmol) were prepared andtritiated in the Isotope Laboratory of BRC (Radiolab), Szeged as describedpreviously (Nevin et al., 1994). Guanosine-5-[γ-35S]-triphosphate ([35S]GTPγS; 1200 Ci/mmol) was obtained from the Isotope Institute Ltd. (Bu-dapest, Hungary). Captopril was purchased from The Squibb Institute forMedical Research (Princeton, NJ, USA). Polyethyleneimine (PEI), EDTA(ethylenediamine tetraacetic acid), Tris(hydroxymethyl)amino-methane(Tris), guanosine 5′-diphosphate (GDP), unlabeled GTPγS, and the pepti-dase inhibitors phosphoramidon and bestatin were purchased fromSigma-Aldrich (St. Louis, MO, USA).
2.3. Peptide synthesis
The identified novel hexapeptides were chemically synthesized.MammalianMet-enkephalin used as a control enkephalinwas obtainedfrom Bachem Feinbiochimica, Bubendorf, Switzerland. The followingendogenous hexapeptides YGGFMS (Pufferfish-type-1), YGGFMI(Zebrafish-type), YGGFMD (Pufferfish-type-2) and YGGFMR (Zebrafinch-type) were synthesized in the Research Group for PeptideChemistry of the Hungarian Academy of Sciences, Budapest, Hungary.Crude peptides were prepared by solid phase peptide synthesis(Fmoc/tBu strategy, 2-Cl-trityl resin, SYRO, MultiSyntech synthesizer,Witten, Germany). Crude peptides were purified by analytical reversedphase high performance liquid chromatography (HPLC, SynergiMAX-RP, Phenomenex, Inc. Berlin, Germany) and preparative fastprotein liquid chromatography (FPLC, Pharmacia, Uppsala, Sveden;Vydac C18 column, Hesperia, CA, USA). The structures were analyzedand confirmed by mass spectrometry (Bruker Corp. Esquire 3000Quadrupole plus ion trap machine).
2.4. Preparation of rat brain membrane fractions
Membrane-rich fractions from brains of inbred Wistar rats wereprepared as described previously (Benyhe et al., 1997). Briefly, fullbrains without cerebella were rapidly removed and homogenized(Braun teflon/glass homogenizator) in 30 vol (mL/g wet mass) of50 mM Tris–HCl buffer pH 7.4. The homogenate was centrifuged at40,000 g for 20 min (Sorvall RC5C superspeed centrifuge, SS34 rotor).The pellet was resuspended in fresh buffer (30 vol) by vortexing andthen incubated for 30 min at 37 °C to remove endogenous opioid pep-tides. The membrane suspension was again centrifuged as describedabove. The final pellet was suspended in 5 mL/g (volume/wet mass)Tris–HCl buffer (pH 7.4) containing 0.32 M sucrose. Five milliliteraliquots weremade and immediately frozen under liquid nitrogen. Sam-ples were stored at −80 °C. On the day of the experiments membranes

31E. Bojnik et al. / Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35
were thawed and washed by centrifugation to remove sucrose. The finalmembrane pellets were suspended in the assay buffer and used immedi-ately in the binding experiments.
2.5. Opioid receptor binding experiments
MOP and DOP receptor binding experiments were performed induplicates. The samples were composed of 50 mM Tris–HCl buffer(pH 7.4), 1 mg/mL bovine serum albumin (BSA) and 0.2–0.4 mg/mLmembrane protein, 0.8–1.3 nM radioligand in the absence or thepresence of unlabelled peptides. The following endopeptidase inhibitorscaptopril, bestatin and phosphoramidon (1 μM each) were included inthe assay mixtures to inhibit endogenous peptidase enzymes(Hiranuma et al., 1998). Incubations at 24 °C for 45 min were started bythe addition of membrane proteins. Reactions were terminated by rapidfiltration followed by extensive washings (3 × 5 mL chilled buffer)through Whatman GF/C glass fiber filters. The filtering/washing cycleswere carried out with an automated cell harvester (Brandel ModelM24R). Bound radioactivitiesweremeasured inUltimaGold™MVscintil-lation cocktail (Perkin Elmer,Waltham,MA, USA) using a Packard Tricarb2300TR liquid scintillation analyzer with 60% counting efficiency.
2.6. [35S]GTPγS binding assays
Agonist induced-receptor mediated G-protein activation wasmeasured as described by Traynor and Nahorsky (1995) with slightmodifications. Diluted rat brain membrane fractions (10–15 μg ofprotein/assay tube) were incubated at 30 °C for 60 min in Tris–EGTAbuffer (pH 7.4) containing 50 mM Tris–HCl, 1 mM EGTA, 3 mMMgCl2,100 mM NaCl, 0.05 nM [35S]GTPγS, 30 μM GDP and increasing concen-trations of the peptides tested. The final volume was 1 mL. Incubationswere started by the addition of the radioligand. Non-stimulated activity(total binding)wasmeasured in the absence of tested compounds, non-specific binding was determined in the presence of 100 μM unlabelledGTPγS. Bound and free [35S]GTPγS were separated by vacuum filtrationthrough Whatman GF/B glass fiber filters with a Brandel M24R cellharvester. After extensive washings with 3 × 5 mL ice-cold buffer filterdisks were removed from the sheet and the bound radioactivity wasdetected in UltimaGOLD™ (Perkin Elmer) scintillation cocktail with aPackard TriCarb 2300TR counter. G-protein activation is given as per-centage over the specific [35S]GTPγS binding obtained in the absenceof receptor ligands (basal activity).
2.7. Data analysis
Radioligand binding assays were carried out in duplicate and the[35S]GTPγS binding experiments were performed in triplicate. Boundradioactivity data were processed by non-linear regression analysiswith GraphPad Prism software version 5.00 for Windows, GraphPadSoftware, San Diego CA, USA, www.graphpad.com research softwarepackage using standard office computers. Data statistics were analyzedby GraphPad InStat, version 3.06).
2.8. Cellular expression and internalization of hMOP–EGFP receptors
Human embryonic kidney cells (HEK293) were maintained andtransfected with 20 μg of pEGFP–MOPr vector as described previously(Bojnik et al., 2011). Cultured HEK cells were treated with opioidpeptides (1 μM) for 1 h at 37 °C. The internalization of the hMOP–EGFPreceptors was followed in living cells in real time at room temperatureby sequential laser scanning under the Olympus Cell-R epi-fluorescencemicroscope using a 20× objective and a combination of filter set for GFP.
3. Results
3.1. Bioinformatic analysis revealed the PENK-related hexapeptides
The analysis of public nucleotide and protein databases (Ensembl,PubMed Protein) reveals the existence of several orthologous sequencesto the known mammalian consensus enkephalin structures (Fig. 1).Thus, 35 vertebrate sequences were analyzed and a C-terminal aminoacid deletion combined with non-synonymous point mutations resultedin four atypical hexapeptide enkephalins in six fish, amphibian and birdspecies. Among these four atypical hexapeptides, twowere located in thefirst opioid and the other two in the seventh opioid core units withinPENK of: the zebrafish Danio rerio, the ocellate tiger pufferfish Takifugurubripes, the green spotted puffer Tetraodon nigroviridis, and the three-spined stickleback Gasterosteus aculeatus, Tyr-Gly-Gly-Phe-Met-Asp(YGGFMD, seventh unit), the common mudpuppy Necturus maculosusand the zebra finch Taeniopygia guttata Tyr-Gly-Gly-Phe-Met-Arg(YGGFMR, seventh unit). A second opioid core unit of PENK encodesfor Tyr-Gly-Gly-Phe-Met-Ser (YGGFMS, second unit) in three polyploidfish species, Takifugu, Tetraodon and Gasterosteus. At the same positionTyr-Gly-Gly-Phe-Met-Ile (YGGFMI) encoded for zebrafish PENK. In addi-tion, endopeptidase recognition motifs were analyzed for their distribu-tion frequencies at different positions (Table 1) and theywere divided asan N-terminal and C-terminal side as distribution varies depending onterminal side. Overall, the most abundant cleavage site is determinedby the KR motif (272 out of total 423 cleavage sites, 64%), although KKand RR dibasic repeats were also frequently found (94 and 57 out of423, 22% and 14% respectively). The KR exclusively represents theN-terminal cleavage site for almost all enkephalin units within PENK(193 out of 230, 83%), except for the first Met-enkephalin, that is KK(35 out of 230, 15%). In the C-terminal site sequence of the cleavagesites are more homogenously distributed (Table 1). KR is mainlypresent in the first, fourth and sixth Met-enkephalin segments (79 outof 193, 41%), while KK is present solely in the second and third Met-enkephalin segments (59 out of 193, 31%). Interestingly, C-terminalsite encodes for the RR cleavage site (55 out of 193, 28%) that is almostabsent in the N-terminal side. RK border sequence was not observed atall. The alignment results of the full PENK sequences that contain theseatypical opioid peptides and their endopeptidase cleavage sites areshown and compared to human PENK in Fig. 1.
3.2. Opioid receptor binding experiments
In order to explore the opioid receptor binding properties of thesenovel atypical opioid hexapeptides, we have chemically synthesized.Firstly we have tested their μ-opioid (MOP) receptor affinity using[3H]DAMGO [Tyr-D-Ala-Gly-NMePhe-Gly5-ol]enkephalin binding assays.Their binding properties were compared to those of unlabelled DAMGO(Fig. 2. left panel). Each of the hexapeptide enkephalin displayedmoderate affinity in heterologous competition experiments with [3H]DAMGO, while the homologous ligand exhibited the highest affinityin these experiments. The rank order of potencies was DAMGO N
YGGFMR N YGGFMS N YGGFMI N YGGFMD (Table 2). A statisticalevaluation by one-way analysis of variance of the individual IC50 valuesfollowed by Bonferroni's multiple comparisons test (GraphPad InStat,version 3.06) showed that the affinities of YGGFMI and YGGFMD dif-fered significantly (p b 0.001) from the affinity of DAMGO.
We next explored the binding properties of the hexapeptides atthe δ-opioid (DOP) receptors using a radiolabeled frog-skin peptideligand analogue, Ile5,6deltorphin-2 with the following structure:[3H]Tyr-D-Ala-Phe-Glu-Ile-Ile-Gly-NH2. [3H]Ile5,6deltorphin-2, a syntheticdisubstituted deltorphin derivative is a highly specific agonist for theDOPreceptors (Nevin et al., 1994). Consistent with a DOP receptor preferenceof the mammalian enkephalins, the novel hexapeptides displayedgood to moderate affinities in the DOP receptor selective assays (Fig. 2.right panel, Table 2). The rank order of affinities was Ile5,6deltorphin-2

TetraodonPENK ----------PMAVPTHCRCVWMFLA--GMCVSLVAGTDCRKECALCVYHLLGQQSTFSS 48TakifuguPENK KSALSVGLLNPMAVPTHCSCIWMFLA--GMCVSLVAGTDCRKECALCVYHLLGQQSTISS 58GasterosteusPENK -----------MAAPAHSSCVWILVLVLGACVSQVVGTECGKECALCVYRLLAQQSGFSS 49Danio1PENK -----------MALMMNS---WWTVA-LSACLVLMVRAECGRDCALCVYRLLR-QTDIDT 44HumanPENK -----------MARFLT--LCTWLLL-LGPGLLATVRAECSQDCATCSYRLVRPAD-INF 45Zebra finchPENK -----------MALLLR--LGCSLLA-LSACLLPRARADCGRDCAACAYRLGPRAG-IHP 45NecturusPENK -----------MALLLLKHCCCLLLVLAA-CLIGAVRSECSEDCAACTFRLGHQAD-INP 47XenopusPENK -----------MGLGARRCCFFLLMFA---SLSVAIRADCTKDCASCALHLGQQTE-INS 45
TetraodonPENK LTCSLECEEEAKNAP-LDAESARELEQGAEG---AMTMEDVDTTL-------------PG 91TakifuguPENK LTCSLECEGELDSQK-LHLCQDFILEKAKNAPLNAESAHELEQEA-------------EG 104GasterosteusPENK PTCSLECKGGLDTQK-LRLCRDFLIEEEHNIPLNADPLQPQDQEASDEATADEDDSPSPQ 108Danio1PENK LACTLECEGSVDSRK-IEICKNILTEEDRLA---IDSLK-QDEES-------------AN 86HumanPENK LACVMECEGKLPSLKIWETCKELLQLSKPELPQDGTSTLRENSKPE------------ES 93Zebra finchPENK LACTLECEGKLPSAKAWETCKELLQLTKLDLSEDGNIAPGDKKELD------------EN 93NecturusPENK LTCTLECEGKLPSS-----------QAWDTFKEFLQVIKLDLVQQD---GQKDPAGDEK- 90XenopusPENK LACTLECEGKLPSA-----------KAWGTCKELLQLTKVDSVQDG----EKYQDNNDS- 89
1st / 2nd 3rdME / Hexapeptide & ME ME /deletion
TetraodonPENK QQLAKKYGGFMKRYGGFMSRRD--VPEGALEHPSDP-----------------DEEENIR 132TakifuguPENK HQLAKKYGGFMKRYGGFMSRRD--MPEGALEHPLNP-----------------DEEETIR 145GasterosteusPENK HLLAKKYGGFMKRYGGFMSRRSP-SQEGLLEDPGNQ-----------------DEEESVR 150Danio1PENK HVLAKKYGGFMKRYGGFMIKKAAEIGTGAPAESDGTGAISKKYGGFMKKAD--DGAEDQQ 144HumanPENK HLLAKRYGGFMKRYGGFMKKMDELYPMEPEEEANGSEILAKRYGGFMKKDAEEDDSLANS 153Zebra finchPENK HLLAKKYGGFMKRYGGFMKKMDELYRAEPEDEANGGEMLAKRYGGFMKKDSD-DDALANS 152NecturusPENK HLLAKKYGGFMKRYGGFMKKMDELYTVEPEEEYNG-EILAKRYGGFMKKDSD---EATDL 146XenopusPENK HYAAKKYGGFMKRYGGFMKKMDELYHAEPEEDDAGGEILAKKYGGFMKKEYD---SDRDA 146
4thME / Octapeptide
TetraodonPENK LEILKILNAAAVHGSE-----GGGKAGEEGKRYGGFMRRAEEGAAQGDLLEAVLGRGLK 186TakifuguPENK LDILKILNAAAGHDSE-----GAGKAGEEVKRYGGFMRRAEEGVAQGDLLEAVLGRGLK 199GasterosteusPENK LEILKILNAAPEQGGE-----GPG--GDAVKRYGGFMRRAGGGVAQGDLLEAVLGRGLK 202Danio1PENK VELLREILRVGLSSES-----DDQPDGDMVKRYGGFMRSVQENT----------GRDLH 188HumanPENK SDLLKELLETGDNRERSHHQDGSDNEEEVSKRYGGFMRGLKRSPQLED-----EAKELQ 207Zebra finchPENK SDLLKELLGTGDSPEAAHYREINENDGDVSKRYGGFMRSIKRSPELED-----EAKELQ 206NecturusPENK LRELLGTADGGNPEAGRYRDSANESQGEVSKRYGGFMREVKR-----SPEMEEGAKDLH 200XenopusPENK SDLLELLATSGDPESAIYHDNNSETPGEMNKRYGGFMRGYRR-----STDLEDETRGIQ 200
5th 6th 7thME LE / ME / Relics Hexapeptide/Heptapeptide
TetraodonPENK KRYGGFMRRVGRPEWLVDSSKR-GGVLKRAWG---------SDN-----DLQKRYGGFMD- 225TakifuguPENK KRYGGFMRRVGRPEWLVDSNKR-GGELKRAWE---------NGN-----DLQKRYGGFMD- 238GasterosteusPENK KRYGGFMRRVGRPEWLVDSSK-SGGMLKRAWE---------SGS-----ELQKRYGGFMD- 241Danio1PENK KRYGGFMRRVGRPDWL-DNQK-SGGLLKRTWE---------EGGETALPDMQKRYGGFMD- 231HumanPENK KRYGGFMRRVGRPEWWMDYQKRYGGFLKRFAE-ALPSDEEGESYSKEVPEMEKRYGGFMRF 260Zebra finchPENK KRYGGFMRRVGRPEWWLDYQKRYGGFLKRFADSILPSEEDGETYSKEVPEMEKRYGGFMR- 260NecturusPENK KRYGGFMRRVGRPEWWLDYQKRYGGFMRRFADYSAPSDEDGESYSKEIPEMEKRYGGFMRF 254XenopusPENK KRYGGFMRRVGRPEWWQDYQKRYGGFMRRFTDSFLPSEEDGESYSKENPDMEKRYGGFMRF 254
Fig. 1. Multiple alignment of various proenkephalin sequences expressing hexapeptide enkephalins. Green spotted pufferfish (Tetraodon nigroviridis; ENSTNIG00000018138), Japanesepufferfish (Takifugu rubripes; ENSTRUP00000025825), three-spined stickleback (Gasterosteus aculeatus; ENSGACG00000014732), Clawed frog (Xenopus laevis; AAB20686), zebrafish(Danio rerio; A8E7S2 and NP_878303.1), human (Homo sapiens; AAH32505.1) zebra finch (Taeniopygia guttata; gb|ACH43852.1), common mudpuppy (Necturus maculosus;AAN46361). The processable opioid segments are in bold and underlined.Mutated amino acid positions in comparisonwith the human PENK are coloredwith red. Endoproteolytic cleav-age sites are shown in blue. Only 100% identical amino acid positions are shaded in gray. (For interpretation of the references to color in thisfigure legend, the reader is referred to thewebversion of this article.)
32 E. Bojnik et al. / Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35
N YGGFMSN YGGFMI N YGGPMRN YGGFMD. IC50 valueswere comparedby one-way analysis of variance followed by Bonferroni's multiple com-parisons test (Table 2.). IC50 values of YGGPMR (p b 0.01) and YGGFMD
Table 1Distribution frequency of dibasic endopeptidase cleavage sites in different proenkephalins.
N-terminal side C-terminal side
Enkephalin position KR KK RR RK KR KK RR RK
First (Met) 5 27 – – (33) – – –
Second (Met) (33) – – – – 30 –
Third (Met) 22 7 2 – 29Fourth (Met) octapeptide 34 1 – – 24 – 9 –
Fifth (Met) 33 – – – 1 – 34 –
Sixth (Leu) 32 – – – 21 – 12 –
Seventh (Met) heptapeptide 34 – – – – – – –
Total 193 35 2 – 79 59 55 –
% of total 84 15 1 – 41 31 28 –
35 vertebrate species with full, or partial cDNA or genomic sequences were analyzed. ThecommonKR sequence interconnecting thefirst and second enkephalin units is in parenthesis.
(p b 0.001) differed significantly from that of the homologous ligand,Ile5,6deltorphin-2. μ/δ Receptor binding ratios were within a narrowscale ranging 0.32–2.9 (Table 2., last column) indicating that thepeptides exhibit rather a small preference, than a large selectivity, fora particular receptor type.
3.3. [35S]GTPγS binding experiments
Since hexapeptides could bind to opioid receptor and since thesereceptors are G-protein coupled receptors and induce their cellularsignaling through their G-proteins, we have studied if they are able tostimulate G-proteins. All novel hexapeptide enkephalins effectivelystimulated the G-proteins by increasing the binding of [35S]GTPγS onrat brain membranes indicating the agonist properties of the ligandstested (Fig. 3). In terms of maximal stimulation, YGGFMI caused thehighest effect (189%), but all other peptides were also decidedly effec-tive. The maximal stimulation levels (efficacy, %Emax) and the potency(pEC50) values of the ligands are summarized in Table 3. An extra sumof squares F test revealed that the best fit values for %Emax and pEC50
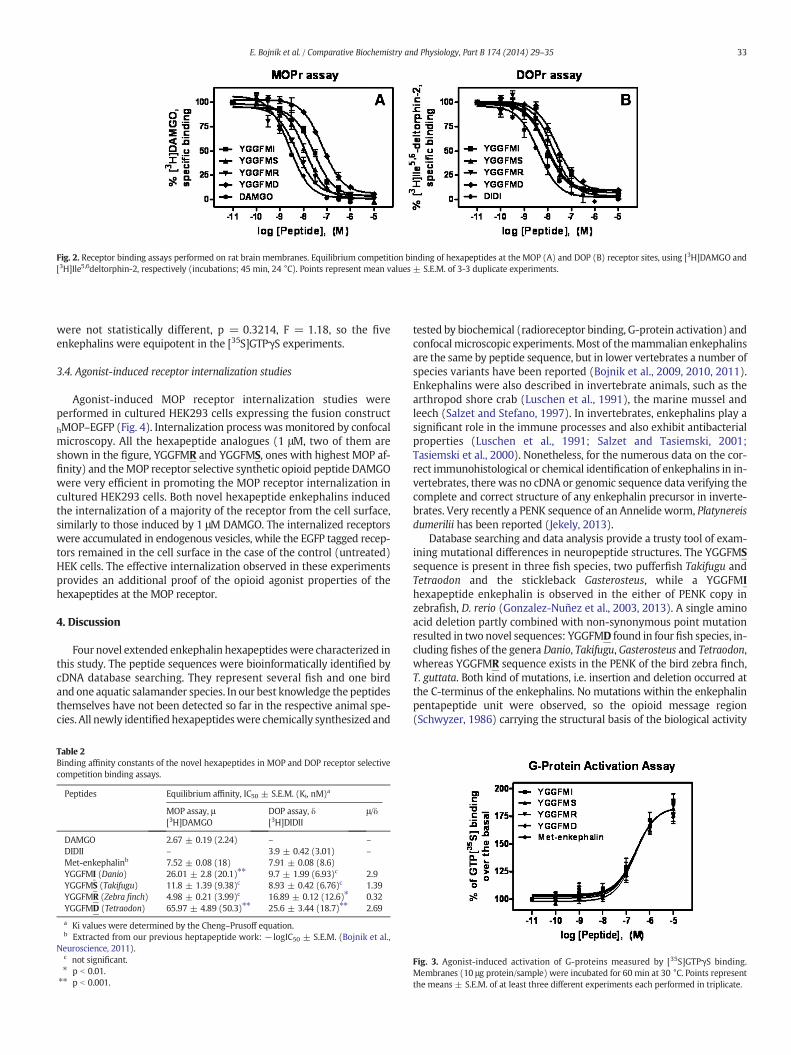
Fig. 2. Receptor binding assays performed on rat brain membranes. Equilibrium competition binding of hexapeptides at the MOP (A) and DOP (B) receptor sites, using [3H]DAMGO and[3H]Ile5,6deltorphin-2, respectively (incubations; 45 min, 24 °C). Points represent mean values ± S.E.M. of 3-3 duplicate experiments.
33E. Bojnik et al. / Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35
were not statistically different, p = 0.3214, F = 1.18, so the fiveenkephalins were equipotent in the [35S]GTPγS experiments.
3.4. Agonist-induced receptor internalization studies
Agonist-induced MOP receptor internalization studies wereperformed in cultured HEK293 cells expressing the fusion constructhMOP–EGFP (Fig. 4). Internalization process wasmonitored by confocalmicroscopy. All the hexapeptide analogues (1 μM, two of them areshown in the figure, YGGFMR and YGGFMS, ones with highest MOP af-finity) and theMOP receptor selective synthetic opioid peptide DAMGOwere very efficient in promoting the MOP receptor internalization incultured HEK293 cells. Both novel hexapeptide enkephalins inducedthe internalization of a majority of the receptor from the cell surface,similarly to those induced by 1 μM DAMGO. The internalized receptorswere accumulated in endogenous vesicles, while the EGFP tagged recep-tors remained in the cell surface in the case of the control (untreated)HEK cells. The effective internalization observed in these experimentsprovides an additional proof of the opioid agonist properties of thehexapeptides at the MOP receptor.
4. Discussion
Four novel extended enkephalin hexapeptideswere characterized inthis study. The peptide sequences were bioinformatically identified bycDNA database searching. They represent several fish and one birdand one aquatic salamander species. In our best knowledge the peptidesthemselves have not been detected so far in the respective animal spe-cies. All newly identifiedhexapeptideswere chemically synthesized and
Table 2Binding affinity constants of the novel hexapeptides in MOP and DOP receptor selectivecompetition binding assays.
Peptides Equilibrium affinity, IC50 ± S.E.M. (Ki, nM)a
MOP assay, μ[3H]DAMGO
DOP assay, δ[3H]DIDII
μ/δ
DAMGO 2.67 ± 0.19 (2.24) – –
DIDII – 3.9 ± 0.42 (3.01) –
Met-enkephalinb 7.52 ± 0.08 (18) 7.91 ± 0.08 (8.6)YGGFMI (Danio) 26.01 ± 2.8 (20.1)⁎⁎ 9.7 ± 1.99 (6.93)c 2.9YGGFMS (Takifugu) 11.8 ± 1.39 (9.38)c 8.93 ± 0.42 (6.76)c 1.39YGGFMR (Zebra finch) 4.98 ± 0.21 (3.99)c 16.89 ± 0.12 (12.6)⁎ 0.32YGGFMD (Tetraodon) 65.97 ± 4.89 (50.3)⁎⁎ 25.6 ± 3.44 (18.7)⁎⁎ 2.69
a Ki values were determined by the Cheng–Prusoff equation.b Extracted from our previous heptapeptide work: −logIC50 ± S.E.M. (Bojnik et al.,
Neuroscience, 2011).c not significant.⁎ p b 0.01.⁎⁎ p b 0.001.
tested by biochemical (radioreceptor binding, G-protein activation) andconfocalmicroscopic experiments. Most of themammalian enkephalinsare the same by peptide sequence, but in lower vertebrates a number ofspecies variants have been reported (Bojnik et al., 2009, 2010, 2011).Enkephalins were also described in invertebrate animals, such as thearthropod shore crab (Luschen et al., 1991), the marine mussel andleech (Salzet and Stefano, 1997). In invertebrates, enkephalins play asignificant role in the immune processes and also exhibit antibacterialproperties (Luschen et al., 1991; Salzet and Tasiemski, 2001;Tasiemski et al., 2000). Nonetheless, for the numerous data on the cor-rect immunohistological or chemical identification of enkephalins in in-vertebrates, there was no cDNA or genomic sequence data verifying thecomplete and correct structure of any enkephalin precursor in inverte-brates. Very recently a PENK sequence of an Annelide worm, Platynereisdumerilii has been reported (Jekely, 2013).
Database searching and data analysis provide a trusty tool of exam-ining mutational differences in neuropeptide structures. The YGGFMSsequence is present in three fish species, two pufferfish Takifugu andTetraodon and the stickleback Gasterosteus, while a YGGFMIhexapeptide enkephalin is observed in the either of PENK copy inzebrafish, D. rerio (Gonzalez-Nuñez et al., 2003, 2013). A single aminoacid deletion partly combined with non-synonymous point mutationresulted in two novel sequences: YGGFMD found in four fish species, in-cluding fishes of the genera Danio, Takifugu, Gasterosteus and Tetraodon,whereas YGGFMR sequence exists in the PENK of the bird zebra finch,T. guttata. Both kind of mutations, i.e. insertion and deletion occurred atthe C-terminus of the enkephalins. No mutations within the enkephalinpentapeptide unit were observed, so the opioid message region(Schwyzer, 1986) carrying the structural basis of the biological activity
Fig. 3. Agonist-induced activation of G-proteins measured by [35S]GTPγS binding.Membranes (10 μg protein/sample) were incubated for 60 min at 30 °C. Points representthe means ± S.E.M. of at least three different experiments each performed in triplicate.
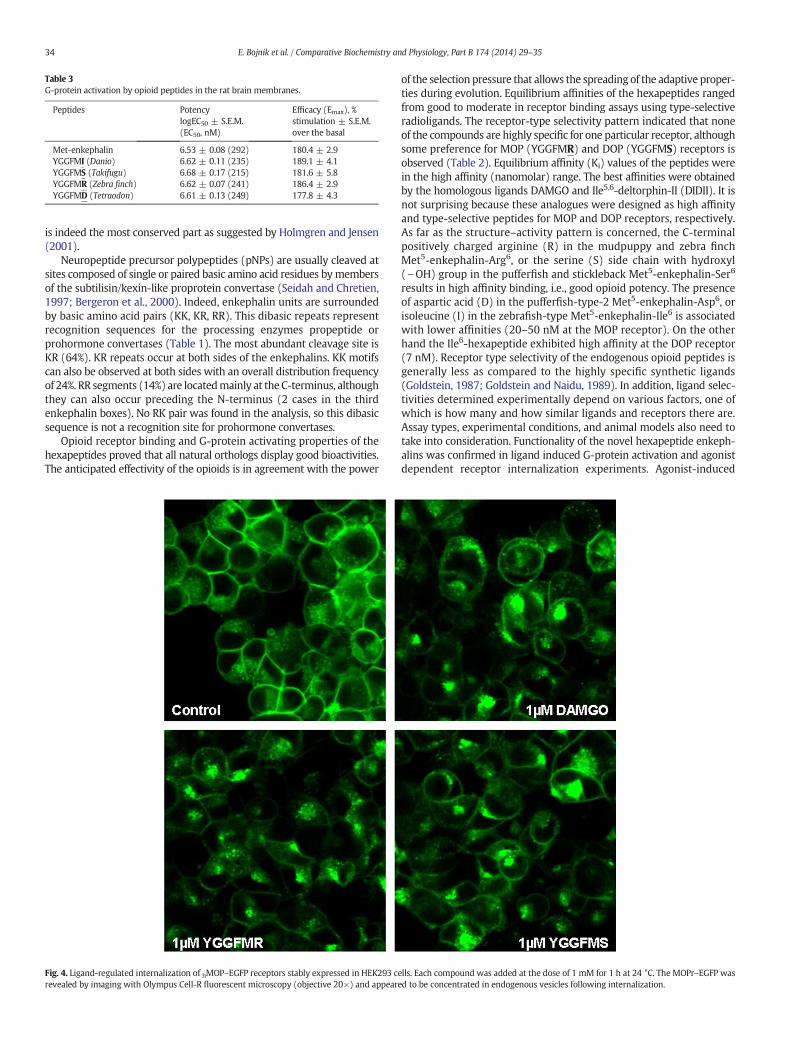
Table 3G-protein activation by opioid peptides in the rat brain membranes.
Peptides PotencylogEC50 ± S.E.M.(EC50, nM)
Efficacy (Emax), %stimulation ± S.E.M.over the basal
Met-enkephalin 6.53 ± 0.08 (292) 180.4 ± 2.9YGGFMI (Danio) 6.62 ± 0.11 (235) 189.1 ± 4.1YGGFMS (Takifugu) 6.68 ± 0.17 (215) 181.6 ± 5.8YGGFMR (Zebra finch) 6.62 ± 0.07 (241) 186.4 ± 2.9YGGFMD (Tetraodon) 6.61 ± 0.13 (249) 177.8 ± 4.3
34 E. Bojnik et al. / Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35
is indeed the most conserved part as suggested by Holmgren and Jensen(2001).
Neuropeptide precursor polypeptides (pNPs) are usually cleaved atsites composed of single or paired basic amino acid residues bymembersof the subtilisin/kexin-like proprotein convertase (Seidah and Chretien,1997; Bergeron et al., 2000). Indeed, enkephalin units are surroundedby basic amino acid pairs (KK, KR, RR). This dibasic repeats representrecognition sequences for the processing enzymes propeptide orprohormone convertases (Table 1). The most abundant cleavage site isKR (64%). KR repeats occur at both sides of the enkephalins. KK motifscan also be observed at both sides with an overall distribution frequencyof 24%. RR segments (14%) are locatedmainly at the C-terminus, althoughthey can also occur preceding the N-terminus (2 cases in the thirdenkephalin boxes). No RK pair was found in the analysis, so this dibasicsequence is not a recognition site for prohormone convertases.
Opioid receptor binding and G-protein activating properties of thehexapeptides proved that all natural orthologs display good bioactivities.The anticipated effectivity of the opioids is in agreement with the power
Fig. 4. Ligand-regulated internalization of hMOP–EGFP receptors stably expressed in HEK293 crevealed by imaging with Olympus Cell-R fluorescent microscopy (objective 20×) and appeare
of the selection pressure that allows the spreading of the adaptive proper-ties during evolution. Equilibrium affinities of the hexapeptides rangedfrom good to moderate in receptor binding assays using type-selectiveradioligands. The receptor-type selectivity pattern indicated that noneof the compounds are highly specific for one particular receptor, althoughsome preference for MOP (YGGFMR) and DOP (YGGFMS) receptors isobserved (Table 2). Equilibrium affinity (Ki) values of the peptides werein the high affinity (nanomolar) range. The best affinities were obtainedby the homologous ligands DAMGO and Ile5,6-deltorphin-II (DIDII). It isnot surprising because these analogues were designed as high affinityand type-selective peptides for MOP and DOP receptors, respectively.As far as the structure–activity pattern is concerned, the C-terminalpositively charged arginine (R) in the mudpuppy and zebra finchMet5-enkephalin-Arg6, or the serine (S) side chain with hydroxyl(\OH) group in the pufferfish and stickleback Met5-enkephalin-Ser6
results in high affinity binding, i.e., good opioid potency. The presenceof aspartic acid (D) in the pufferfish-type-2 Met5-enkephalin-Asp6, orisoleucine (I) in the zebrafish-type Met5-enkephalin-Ile6 is associatedwith lower affinities (20–50 nM at the MOP receptor). On the otherhand the Ile6-hexapeptide exhibited high affinity at the DOP receptor(7 nM). Receptor type selectivity of the endogenous opioid peptides isgenerally less as compared to the highly specific synthetic ligands(Goldstein, 1987; Goldstein and Naidu, 1989). In addition, ligand selec-tivities determined experimentally depend on various factors, one ofwhich is how many and how similar ligands and receptors there are.Assay types, experimental conditions, and animal models also need totake into consideration. Functionality of the novel hexapeptide enkeph-alins was confirmed in ligand induced G-protein activation and agonistdependent receptor internalization experiments. Agonist-induced
ells. Each compound was added at the dose of 1 mM for 1 h at 24 °C. The MOPr–EGFP wasd to be concentrated in endogenous vesicles following internalization.

35E. Bojnik et al. / Comparative Biochemistry and Physiology, Part B 174 (2014) 29–35
stimulation of G-proteins by the four compounds was found to be effi-cient and very similar, while the internalization assays revealed thatthe hexapeptides at 1 μM concentration effectively internalized EGFP-fused MOP receptors in cultured HEK 293 cell lines. These data togetherconfirm that all newly identified hexapeptide sequences indeed carryopioid-related biological activity.
Summarizing the results, the receptor binding and G-proteinactivating properties of four novel, non-mammalian enkephalins weredescribed and compared to those of the well-known Met-enkephalin.Among the four structures tested, the zebra finch peptide YGGFMR ex-hibited the highest affinities in theMOP receptor sensitive binding assays,while the fish peptide YGGFMS displayed the highest affinity at the DOPreceptor. Met-enkephalin and the four new peptides were equipotent inthe G-protein stimulation experiments. The novel endogenously occur-ring sequences represent further examples of the natural diversity ob-served within the opioid peptide family. Studying these peptides withnon-conventional structures is important from both phylogenetic andchemical–biochemical points of view. An increasing number of such spe-cies variants of opioid peptides are expected since bioinformatic data areemerging rapidly from DNA sequencing efforts.
Acknowledgments
This work was supported by TÁMOP— 4.2.2A-11/1KONV-2012-0024,2012-052 and OTKA 108518 grants.
References
Benyhe, S., Farkas, J., Tóth, G., Wollemann, M., 1997. Met5-enkephalin-Arg6-Phe7, an en-dogenous neuropeptide, binds to multiple opioid and non-opioid sites in rat brain.J. Neurosci. Res. 48, 249–258.
Bergeron, F., Leduc, R., Day, R., 2000. Subtilase-like pro-protein convertases: from molec-ular specificity to therapeutic applications. J. Mol. Endocrinol. 24, 1–22.
Bojnik, E., Magyar, A., Tóth, G., Bajusz, S., Borsodi, A., Benyhe, S., 2009. Binding studies ofnovel, non-mammalian enkephalins, structures predicted from frog and lungfishbrain cDNA sequences. Neuroscience 158, 867–874.
Bojnik, E., Babos, F., Magyar, A., Borsodi, A., Benyhe, S., 2010. Bioinformatic and biochemicalstudies on the phylogenetic variability of proenkephalin-derived octapeptides. Neuro-science 165, 542–552.
Bojnik, E., Boynik, E., Corbani, M., Babos, F., Magyar, A., Benyhe, S., 2011. Phylogeneticdiversity and functional efficacy of the C-terminally expressed heptapeptide unit inthe opioid precursor polypeptide proenkephalin A. Neuroscience 178, 56–67.
Comb, M., Seeburg, P.H., Adelman, J., Eiden, L., Herbert, E., 1982. Primary structure of thehuman Met- and Leu-enkephalin precursor and its mRNA. Nature 295, 663–666.
Dores, R.M., McDonald, L.K., Goldsmith, A., Deviche, P., Rubin, D.A., 1993. The phylogeny ofenkephalins: speculations on the origins of opioid precursors. Cell. Physiol. Biochem.3, 231–244.
Dores, R.M., Lecaude, S., Bauer, D., Danielson, P.B., 2002. Analyzing the evolution of theopioid/orphanin gene family. Mass Spectrom. Rev. 21, 1–24.
Goldstein, A., 1987. Binding selectivity profiles for ligands of multiple receptor types:focus on opioid receptors. Trends Pharmacol. Sci. 8, 456–458.
Goldstein, A., Naidu, A., 1989. Multiple opioid receptors: ligand selectivity profiles andbinding site signatures. Mol. Pharmacol. 36, 265–272.
Gonzalez-Nuñez, V., Gonzalez Sarmiento, R., Rodriguez, R.E., 2003. Characterization ofzebrafish proenkephalin reveals novel opioid sequences. Mol. Brain Res. 114, 31–39.
Gonzalez-Nuñez, V., Jimenez Gonzalez, A., Barreto-Valer, K., Rodriguez, R.E., 2013. In vivoregulation of the mu opioid receptor: role of the endogenous opioid agents. Mol.Med. 19, 7–17.
Gubler, U., Seeburg, P., Hoffman, B.J., Gage, L.P., Udenfriend, S., 1982.Molecular cloning es-tablishes proenkephalin as precursor of enkephalin-containing peptides. Nature 295,206–208.
Hiranuma, T., Kitamura, K., Taniguchi, T., Kobayashi, T., Tamaki, R., Kanai, M., Akahori, K.,Iwao, K., Oka, T., 1998. Effects of three peptidase inhibitors, amastatin, captopril andphosphoramidon, on the hydrolysis of [Met5]-enkephalin-Arg6-Phe7 and other opioidpeptides. Naunyn Schmiedebergs Arch. Pharmacol. 357, 276–282.
Holmgren, S., Jensen, J., 2001. Evolution of vertebrate neuropeptides. Brain Res. Bull. 55,723–735.
Hook, V., Funkelstein, L., Douglas, L., Bark, S., Wegrzyn, J., Hwang, S.-R., 2008. Proteases forprocessing proneuropeptides into peptide neurotransmitters and hormones. Annu.Rev. Pharmacol. Toxicol. 48, 393–423.
Horikawa, S., Takai, T., Toyosato, M., Takahashi, H., Noda, M., Kakidani, H., Kubo, T., Hirose,T., Inayama, S., Hayashida, H., Miyata, T., Numa, S., 1983. Isolation and structural orga-nization of the human preproenkephalin B gene. Nature 306, 611–614.
Hughes, J., Smith, T.H., Kosterlitz, H.W., Fothergill, L.A., Morgan, B.A., Morris, H.R., 1975.Identification of two related pentapeptides from the brain with potent opiate agonistactivity. Nature 258, 577–579.
Jekely, G., 2013. Global view of the evolution and diversity of metazoan neuropeptide sig-naling. Proc. Natl. Acad. Sci. U. S. A. 110, 8702–8707.
Kakidani, H., Furutani, Y., Takahashi, H., Noda, M., Morimoto, Y., Hirose, T., Asai, M.,Inayama, S., Nakanishi, S., Numa, S., 1982. Cloning and sequence analysis of cDNAfor porcine beta-neo-endorphin/dynorphin precursor. Nature 298, 245–249.
Khalap, A., Bagrosky, B., Lecaude, S., Youson, J., Danielson, P., Dores, R.M., 2005. Trends inthe evolution of the proenkephalin and prodynorphin genes in gnathostomes. Ann.N. Y. Acad. Sci. 1040, 22–37.
Komorowski, L.K., Lecaude, S.G., Westring, C.G., Danielson, P.B., Dores, R.M., 2012. Evolu-tion of gnathostome prodynorphin and proenkephalin: characterization of a sharkproenkephalin and prodynorphin cDNAs. Gen. Comp. Endocrinol. 177, 353–364.
Luschen, W., Buck, F., Willig, A., Jaros, P.P., 1991. Isolation, sequence-analysis, andphysiological-properties of enkephalins in the nervous-tissue of the shore crabCarcinus maenas L. Proc. Natl. Acad. Sci. U. S. A. 88, 8671–8675.
Mollereau, C., Moisand, C., Butour, J.L., Parmentier, M., Meunier, J.-C., 1996. Structure,tissue distribution, and chromosomal localization of the prepronociceptin gene.Proc. Natl. Acad. Sci. U. S. A. 93, 8666–8670.
Nakanishi, S., Inoue, A., Kita, T., Nakamura, M., Chang, A.C.Y., Cohen, S.N., Numa, S., 1979.Nucleotide sequence of cloned cDNA for bovine corticotropin-beta-lipotropin precursor.Nature 278, 423–427.
Nevin, S.T., Kabasakal, L., Ötvös, F., Tóth, G., Borsodi, A., 1994. Binding characteristics of thenovel highly selective delta agonist, [3H]Ile5,6deltorphin II. Neuropeptides 26,261–265.
Noda, M., Furutani, Y., Takahashi, H., Toyosato, M., Hirose, T., Inayama, S., Nakanishi, S.,Numa, S., 1982. Cloning and sequence analysis of cDNA for bovine adrenalpreproenkephalin. Nature 295, 202–206.
Nothacker, H.P., Reinscheid, R.K., Mansour, A., Henningsen, R.A., Ardati, A., Monsma Jr., F.J.,Watson, S.J., Civelli, O., 1996. Primary structure and tissue distribution of theorphanin FQ precursor. Proc. Natl. Acad. Sci. U. S. A. 93, 8677–8682.
Roberts, E., Shoureshi, P., Kozak, K., Szynskie, L., Baron, A., Lecaude, S., Dores, R.M., 2007.Tracking the evolution of the proenkephalin gene in tetrapods. Gen. Comp.Endocrinol. 153, 189–197.
Salzet, M., Stefano, G.B., 1997. Invertebrate proenkephalin: delta opioid binding sites inleech ganglia and immunocytes. Brain Res. 768, 224–232.
Salzet, M., Tasiemski, A., 2001. Involvement of proenkephalin-derived peptides in immunity.Dev. Comp. Immunol. 25, 177–185.
Seidah, N.G., Chretien, M., 1997. Eukaryotic protein processing: endoproteolysis ofprecursor proteins. Curr. Opin. Biotechnol. 8, 602–607.
Sundstrom, G., Dreborg, S., Larhammar, D., 2010. Concomitant duplications of opioidpeptide and receptor genes before the origin of jawed vertebrates. PLoS One 5,e10512.
Schwyzer, R., 1986. Molecular mechanism of opioid receptor selection. Biochemistry 25,6335–6342.
Tasiemski, A., Verger-Bocquet, M., Cadet, M., Goumon, Y., Metz-Boutigue, M.-H., Aunis, D.,Stefano, G.B., Salzet, M., 2000. Proenkephalin A-derived peptides in invertebrate in-nate immune processes. Mol. Brain Res. 76, 237–252.
Traynor, J.R., 1987. Prodynorphin as a source of [Leu] enkephalin. Trends Pharmacol. Sci.8, 47–48.
Traynor, J.R., Nahorsky, S.R., 1995. Modulation by μ-opioid agonists of guanosine-5′-O-(3-[γ-[35S]thio)triphosphate binding to membranes from human neuroblastoma SH-SY5Y cells. Mol. Pharmacol. 47, 848–854.
Related Documents











