Original article Binding of laminin to Paracoccidioides brasiliensis induces a less severe pulmonary paracoccidioidomycosis caused by virulent and low-virulence isolates Denise C. André a , José D. Lopes b , Marcello F. Franco c , Celidéia A.C. Vaz a , Vera L.G. Calich a, * a Departamento de Imunologia, Instituto de Ciências Biomédicas da Universidade de São Paulo, Av. Prof. Lineu Prestes 1730, CEP 05508-900, São Paulo, SP, Brazil b Departamento de Microbiologia, Imunologia e Parasitologia da Faculdade de Medicina da Universidade Federal de São Paulo, São Paulo, Brazil c Departamento de Patologia, Faculdade de Medicina da Universidade Federal de São Paulo, São Paulo, Brazil Received 9 September 2003; accepted 19 February 2004 Available online 15 April 2004 Abstract The pathogenic fungus Paracoccidioides brasiliensis is the causative agent of paracoccidioidomycosis (PCM). This pulmonary mycosis, acquired by inhalation of airborne propagules, may disseminate to several internal organs and tissues, leading to severe disease. Adhesion to host cell components is the first step involved in dissemination of pathogens. Previous studies showed that laminin, the most abundant glycoprotein of the basement membrane, binds to P. brasiliensis yeast cells, enhancing their pathogenicity in the hamster testicle model. As PCM is primarily a pulmonary infection, we studied the influence of previous treatment of yeast cells with laminin on the course of the intratracheal infection of resistant and susceptible mice using high-virulence (Pb18) and low-virulence (Pb265) P. brasiliensis isolates. Laminin treatment did not alter fungal loads, delayed-type hypersensitivity reactions, levels of pulmonary cytokines and production of specific antibodies in any group of Pb18-infected mice. However, early in the infection, a less intense inflammatory reaction was detected in the lungs of the laminin-treated groups. In addition, laminin treatment of Pb265 resulted in a less severe infection as revealed by the lower fungal loads recovered from lungs. Antibody and cytokine levels, however, did not change after laminin treatment. Altogether, our results demonstrate that laminin binding to yeast cells diminishes P. brasiliensis pathogenicity. The lower inflammatory response observed with the virulent isolate and the decreased pulmonary fungal burden with the low-virulence isolate indicate an inhibitory effect of laminin treatment on P. brasiliensis infectivity and interaction with pulmonary host cells or extracellular matrix proteins. © 2004 Elsevier SAS. All rights reserved. Keywords: Paracoccidioides brasiliensis; Laminin; Pathogenicity; Immunity 1. Introduction Paracoccidioides brasiliensis, the causative agent of hu- man paracoccidioidomycosis (PCM), is primarily a respira- tory pathogen, infecting the host through inhalation of air- borne propagules. The great majority of infected subjects develop an asymptomatic pulmonary infection, although some individuals present clinical manifestations which give rise to the polar forms of overt PCM, namely, hyperergic or localized and anergic or disseminated disease [1,2]. Clinical and experimental evidence indicates that cell-mediated im- munity plays a significant role in host defense against P. bra- siliensis infection, whereas high levels of specific antibodies and polyclonal activation of B cells are associated with the most severe forms of the disease [3–9]. Using an isogenic murine model of PCM, our laboratory showed that there were significant differences in susceptibil- ity among inbred strains: A/Sn mice were found to be the most resistant, while B10.A animals were the most suscep- tible to P. brasiliensis infection [5]. Using the intratracheal (i.t.) route of infection, it was observed that A/Sn mice developed a chronic benign, pulmonary-restricted PCM, Abbreviations: CFU, colony-forming units; DTH, delayed-type hyper- sensitivity; ECM, extracellular matrix; i.t, Intratracheal; LN, laminin; Pb, Paracoccidioides brasiliensis; PCM, paracoccidioidomycosis. * Corresponding author. Tel.: +55-11-30917397; fax: +55-11-30917224. E-mail address: [email protected] (V.L.G. Calich). Microbes and Infection 6 (2004) 549–558 www.elsevier.com/locate/micinf © 2004 Elsevier SAS. All rights reserved. doi:10.1016/j.micinf.2004.02.010

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Original article
Binding of laminin to Paracoccidioides brasiliensis inducesa less severe pulmonary paracoccidioidomycosis caused by virulent
and low-virulence isolates
Denise C. André a, José D. Lopes b, Marcello F. Franco c,Celidéia A.C. Vaz a, Vera L.G. Calich a,*
a Departamento de Imunologia, Instituto de Ciências Biomédicas da Universidade de São Paulo, Av. Prof. Lineu Prestes 1730,CEP 05508-900, São Paulo, SP, Brazil
b Departamento de Microbiologia, Imunologia e Parasitologia da Faculdade de Medicina da Universidade Federal de São Paulo, São Paulo, Brazilc Departamento de Patologia, Faculdade de Medicina da Universidade Federal de São Paulo, São Paulo, Brazil
Received 9 September 2003; accepted 19 February 2004
Available online 15 April 2004
Abstract
The pathogenic fungus Paracoccidioides brasiliensis is the causative agent of paracoccidioidomycosis (PCM). This pulmonary mycosis,acquired by inhalation of airborne propagules, may disseminate to several internal organs and tissues, leading to severe disease. Adhesion tohost cell components is the first step involved in dissemination of pathogens. Previous studies showed that laminin, the most abundantglycoprotein of the basement membrane, binds to P. brasiliensis yeast cells, enhancing their pathogenicity in the hamster testicle model. AsPCM is primarily a pulmonary infection, we studied the influence of previous treatment of yeast cells with laminin on the course of theintratracheal infection of resistant and susceptible mice using high-virulence (Pb18) and low-virulence (Pb265) P. brasiliensis isolates.Laminin treatment did not alter fungal loads, delayed-type hypersensitivity reactions, levels of pulmonary cytokines and production of specificantibodies in any group of Pb18-infected mice. However, early in the infection, a less intense inflammatory reaction was detected in the lungsof the laminin-treated groups. In addition, laminin treatment of Pb265 resulted in a less severe infection as revealed by the lower fungal loadsrecovered from lungs. Antibody and cytokine levels, however, did not change after laminin treatment. Altogether, our results demonstrate thatlaminin binding to yeast cells diminishes P. brasiliensis pathogenicity. The lower inflammatory response observed with the virulent isolate andthe decreased pulmonary fungal burden with the low-virulence isolate indicate an inhibitory effect of laminin treatment on P. brasiliensisinfectivity and interaction with pulmonary host cells or extracellular matrix proteins.© 2004 Elsevier SAS. All rights reserved.
Keywords: Paracoccidioides brasiliensis; Laminin; Pathogenicity; Immunity
1. Introduction
Paracoccidioides brasiliensis, the causative agent of hu-man paracoccidioidomycosis (PCM), is primarily a respira-tory pathogen, infecting the host through inhalation of air-borne propagules. The great majority of infected subjectsdevelop an asymptomatic pulmonary infection, although
some individuals present clinical manifestations which giverise to the polar forms of overt PCM, namely, hyperergic orlocalized and anergic or disseminated disease [1,2]. Clinicaland experimental evidence indicates that cell-mediated im-munity plays a significant role in host defense against P. bra-siliensis infection, whereas high levels of specific antibodiesand polyclonal activation of B cells are associated with themost severe forms of the disease [3–9].
Using an isogenic murine model of PCM, our laboratoryshowed that there were significant differences in susceptibil-ity among inbred strains: A/Sn mice were found to be themost resistant, while B10.A animals were the most suscep-tible to P. brasiliensis infection [5]. Using the intratracheal(i.t.) route of infection, it was observed that A/Sn micedeveloped a chronic benign, pulmonary-restricted PCM,
Abbreviations: CFU, colony-forming units; DTH, delayed-type hyper-sensitivity; ECM, extracellular matrix; i.t, Intratracheal; LN, laminin; Pb,Paracoccidioides brasiliensis; PCM, paracoccidioidomycosis.
* Corresponding author. Tel.: +55-11-30917397; fax: +55-11-30917224.E-mail address: [email protected] (V.L.G. Calich).
Microbes and Infection 6 (2004) 549–558
www.elsevier.com/locate/micinf
© 2004 Elsevier SAS. All rights reserved.doi:10.1016/j.micinf.2004.02.010

whereas B10.A mice developed a progressive disseminateddisease [10]. Although the role of CD4+ T lymphocytes is notclearly defined, CD8+ T cells contribute to the resistancemechanisms mounted by both susceptible and resistantmouse strains [11]. Furthermore, macrophage-activating cy-tokines such as IFN-c, TNF-a and IL-12 are required for thedevelopment of protective immunity [12–16].
The adherence of microorganisms to host tissues is acrucial step in the development of infection. Adherence im-plies that the pathogen recognizes carbohydrate or proteinligands on the surface of host cells, or a constituent ofbasement membranes underlying epithelia and endothelia,such as fibrinogen, laminin, collagen or fibronectin [17]. Thelung contains an alveolar basement membrane that lies be-tween alveolar epithelial cells and capillary endothelial cells.In addition, an endothelial basement membrane separatesendothelial cells from interstitial connective tissue cellsalong the tick portion of the alveolar wall [18]. Laminin, themost abundant glycoprotein of the basement membrane, isimplicated in the attachment of a variety of extracellular andintracellular pathogens to host cells [19–22]. It has beenshown that P. brasiliensis yeast cells bind to laminin throughgp-43, its major surface glycoprotein [23], enhancing patho-genicity of fungal cells in a hamster model of infection.Histological analysis of infected hamster testicles showedthat tissue destruction and replacement by P. brasiliensisgranulomas was much more intense in hamsters infectedwith yeast cells coated with laminin than in the animalsinjected with the fungus only.
Since PCM is mainly acquired by the respiratory route, wewanted to know whether laminin coating of P. brasiliensisyeasts would influence the severity of pulmonary infectionand its dissemination to liver and spleen. After i.t. infectionof resistant and susceptible mice with one million yeast cellsof a virulent isolate (Pb18), animals were monitored forseverity of infection in the lungs, liver and spleen and fordelayed-type hypersensitivity (DTH), production of antibod-ies of different isotypes, presence of pro- and anti-inflammatory cytokines at the site of infection and histopa-thology. Experiments were also performed with differentinocula of virulent yeasts (Pb18) as well as with a low-virulence (Pb265) isolate of P. brasiliensis.
2. Materials and methods
2.1. Animals
Groups of 6 to 8 male mice from the susceptible (B10.A)and resistant (A/Sn) strains to P. brasiliensis infection wereused for each period of infection. All the animals were bred atthe University of São Paulo animal facilities and providedwith acidified water and sterilized food and bedding. Themice were bred under specific pathogen-free conditions inenclosed top cages. Mice were 8–11 weeks of age at the timeof infection and procedures involving animals and their carewere conducted in conformity to national and internationalpolicies.
2.2. Fungus
P. brasiliensis isolates Pb18 and Pb265 from our culturecollection, respectively, virulent and slightly virulent for sus-ceptible B10.A mice, were used in this study [24]. To ensurethe maintenance of Pb18 virulence, this isolate was used afterthree serial animal-passages [25]. The isolates were main-tained by weekly sub-cultivation in semisolid Fava Netto’sculture medium [26] at 35 °C and used on the 7th day ofculture. The fungal yeast cells were washed in phosphatebuffered saline (PBS pH 7.2), counted in a hemocytometer,and the concentration was adjusted to the desired fungalnumber. Viability of fungal suspensions, determined by Ja-nus green B vital dye (Merck, Germany) [27], was alwayshigher than 90%.
2.3. Treatment of yeast cells with laminin
Laminin was purified from Engelbrecht-Holm-Swarmmouse tumor cells [28]. Cell suspensions containing 1 × 106
P. brasiliensis yeasts were incubated with 20 µg/ml of lami-nin in PBS for 30 min at room temperature [23]. Controlcultures were incubated with PBS.
2.4. P. brasiliensis infection
Mice were anesthetized and subjected to i.t. P. brasiliensisinfection as previously described [10]. Briefly, after i.p. anes-thesia the animals were infected with 1 × 106 laminin-treatedor untreated P. brasiliensis Pb18 or Pb265 yeast cells, con-tained in 50 µl of PBS, by surgical i.t. inoculation thatallowed dispensing the fungal cells directly into the lungs. Insome experiments, animals were infected with 1 × 105 or 1 ×104 Pb18 yeast cells previously treated or not with laminin.The skin of the animals was then sutured and the mice wereallowed to recover under a heat lamp.
2.5. Assay for organ colony-forming units (CFU)
The numbers of viable microorganisms in the lungs, liverand spleen from experimental and control mice were deter-mined by counting the number of CFU. Animals from eachgroup were sacrificed at adequate intervals after infectionand the enumeration of viable organisms in the three organswas done as previously described [29]. Briefly, aliquots(100 µl) of the cellular suspensions and its serial dilutions ofeach organ were plated on brain heart infusion (BHI) agar(Difco) supplemented with 4% (v/v) horse serum (InstitutoButantan, São Paulo, Brazil) and 5% Pb192 culture filtrate,the latter constituting a source of growth-promoting factor.The plates were incubated at 35 °C and colonies werecounted daily until no increase in counts was observed. Thenumbers (log10) of viable P. brasiliensis colonies per gram oftissue are expressed as mean ± standard error.
2.6. Histopathologic analysis
The left lung of each mouse was removed, fixed in 10%formalin, and embedded in paraffin. Five-micrometer sec-
550 D.C. André et al. / Microbes and Infection 6 (2004) 549–558

tions were stained with hematoxylin-eosin (H&E) for analy-sis of the lesions and silver stained for fungal evaluation.Pathologic changes were analyzed based on the number, size,morphology and cell composition of granulomatous lesions,number of fungi and intensity of the inflammatory infiltrates.The pathological alterations observed in three sections ofeach mouse were semi-quantitatively expressed according tothe severity of lesions: 0, absence of lesions; 1+, up to 25% oflung involvement; 2+, 25–50% and 3+, more than 50%.
2.7. Measurement of cytokines
The mice were infected i.t. with laminin-coated or un-coated P. brasiliensis and their right lungs were removedaseptically and individually disrupted in 4.0 ml of RPMI-1640 medium (Gibco BRL). Supernatants were separatedfrom cell debris by centrifugation at 2000 × g for 15 min,passed through 0.22-µm-pore-size filters (Millipore), andstored at –70 °C. The levels of IL-4, IL-12, IFN-c and TNF-awere measured by capture ELISA using antibody pairs pur-chased from Pharmingen (San Diego, CA). The ELISA pro-cedure was performed according to the manufacturer’s pro-tocol. The concentrations of cytokines were determined withreference to a standard curve for serial twofold dilutions ofmurine recombinant cytokines. The lower limit of each re-combinant standard curve detection was 7.8, 15.0, 20.0 and31.2 pg/ml for IL-4, IL-12, IFN-c and TNF-a, respectively.
2.8. DTH assay
The DTH reactions were always evaluated just beforesacrifice of the same animals used in the CFU assays, by thefootpad test according to previously determined conditions[30]. Briefly, mice were inoculated with 25 µl of the FavaNettos’s antigen [25] and the footpad thickness was mea-sured with a caliper (Mitutoyo Corporation, Tokyo, Japan)immediately before and 24 h after antigen inoculation. Theincrease in thickness was calculated and expressed in milli-meters. Non-infected mice subjected to the footpad test wereused as controls.
2.9. Measurement of serum P. brasiliensis-specific isotypes
Specific isotype levels (total IgG, IgM, IgA, IgG1, IgG2a,IgG2b and IgG3) were measured by a previously describedELISA [10] employing a cell-free antigen [31] preparedusing a pool of different P. brasiliensis isolates (Pb339,Pb265 and Pb18). The average of the optical densities ob-tained using sera from control mice (PBS inoculated), diluted1:20, was considered the cutoff for each respective isotype.Optical densities for each dilution of experimental sera werecompared to the control values. The titer for each sample wasexpressed as the reciprocal of the highest dilution that pre-sented absorbance higher than the cutoff.
2.10. Statistical analysis
Data were analyzed using Student’s t test or two-wayanalysis of variance (ANOVA), depending on the number of
experimental groups [32]. P values under 0.05 were consid-ered significant.
3. Results
3.1. Influence of laminin on the course of pulmonaryinfection and extrapulmonary dissemination
The evolution of disease in B10.A and A/Sn mice infectedwith laminin-coated and uncoated P. brasiliensis (virulentPb18) was monitored by CFU counts in the lungs, liver andspleen at several postinfection periods (2, 4, 8 and 12 weeks).As shown in Fig. 1, no differences in CFU counts were foundin the lungs, liver and spleen of both mouse strains infectedwith laminin-treated or untreated yeast cells. As previouslycharacterized [10], control B10.A mice presented a progres-sive disseminated infection with CFU counts increasing inliver and spleen from week 8 on. On the contrary, controlA/Sn mice presented a regressive pattern of infection and thelow number of yeast cells that disseminated to liver andspleen at weeks 2–8 were no longer detected at week12 postinfection. Whatever the host genetic pattern and dis-ease outcome, our data showed that laminin treatment did notaffect the course and severity of P. brasiliensis infectioncaused by a virulent isolate.
3.2. Laminin diminishes lung inflammation causedby P. brasiliensis infection
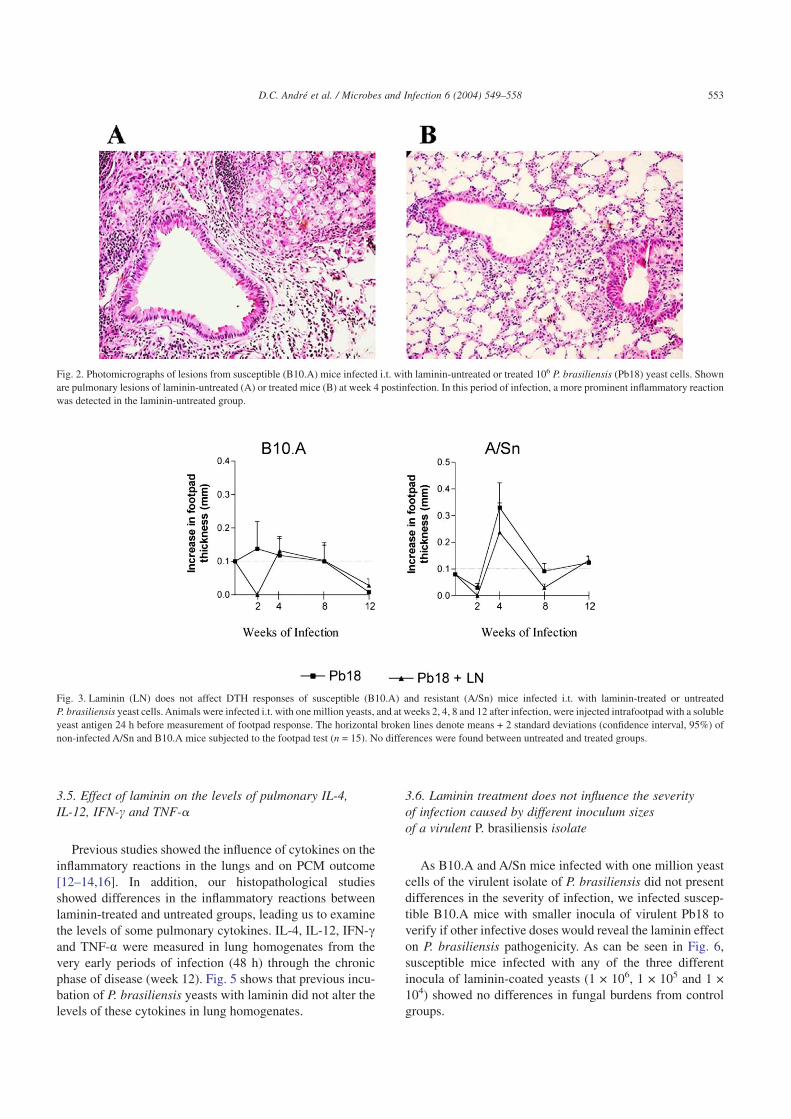
Using a hamster model of infection [23], it was previouslydemonstrated that laminin-treated Pb18 yeast cells caused amuch more intense inflammatory reaction in the testicles ofinfected animals at week 4 of infection. To verify the effect oflaminin on P. brasiliensis pathogenicity, groups of B10.Amice were sacrificed at various postinfection periods and theinflammatory reactions in the lungs were semi-quantitativelyanalyzed. Table 1 shows that laminin-coated yeast cells in-duced less intense inflammatory reactions at weeks 2, 4 and8 postinfection. By week 12, treated and untreated groupspresented equivalent intensity of pulmonary lesions. At week4 after infection the pulmonary lesions in laminin-uncoatedP. brasiliensis-infected groups (Fig. 2A) were randomlydistributed and composed of granulomas of various sizescontaining many fungal cells circumscribed by macroph-ages, lymphocytes and plasma cells, whereas in the laminin-treated groups, lesions were more diffuse and the lung paren-chyma was more preserved (Fig. 2B).
3.3. DTH reactions induced by laminin-coatedand uncoated yeast cells
Since laminin treatment influenced at least partially theseverity of lung inflammation, we studied the effect of lami-nin on the DTH reactions developed by susceptible andresistant mice. As previously characterized [10], susceptibleB10.A mouse behavior was anergy in DTH reactions in thewhole course of infection, whereas resistant A/Sn mice pre-
551D.C. André et al. / Microbes and Infection 6 (2004) 549–558

sented positive DTH reactions. The pre-incubation of P. bra-siliensis yeast cells with laminin did not modify the charac-teristic behavior of either mouse strain (Fig. 3).
3.4. Effect of laminin-binding on the antibody production
We also studied the effect of laminin on specific antibodyproduction (Fig. 4). No differences in the levels of all iso-types tested (IgM, IgA, IgG1, IgG3, IgG2a and IgG2b) werefound between laminin-treated and control groups. In thetreated groups, a trend to lower IgG1, IgG2a and IgG2bproduction at weeks 2 and 4 postinfection was detected; thesedifferences, however, did not reach statistical significance.
Fig. 1. Laminin (LN) treatment does not affect severity of infection of susceptible (B10.A) and resistant (A/Sn) mice to P. brasiliensis infection. Shown isrecovery of CFU from lungs, spleen and liver of mice infected either with laminin-treated or untreated yeasts. The bars depict means ± standard errors of log10
CFU obtained from groups of 6–8 mice at weeks 2, 4, 8 and 12 after infection. No significant differences were detected between laminin-treated and untreatedgroups.
Table 1Median scores of fungal lesions in the lungs of B10.A mice after infectioni.t. with 1 × 106 laminin-coated or uncoated P. brasiliensis (Pb18) yeasts a
Week after infection Pb18 (Median score) Pb18 + laminin(Median score)
2 2+ 1+4 3+ 1+ (P < 0.05)8 3+ 2+12 3+ 3+
a The severity of lesions was semi-quantitatively expressed as 0, absenceof lesions; 1+, up to 25% of lung involvement; 2+, 25–50%; and 3+, morethan 50%. These ratings were based on the number and size of granuloma-tous lesions and the intensity of the inflammatory reactions. Results corres-pond to groups of 6–8 mice and are representative of two separateexperiments.
552 D.C. André et al. / Microbes and Infection 6 (2004) 549–558
3.5. Effect of laminin on the levels of pulmonary IL-4,IL-12, IFN-c and TNF-�
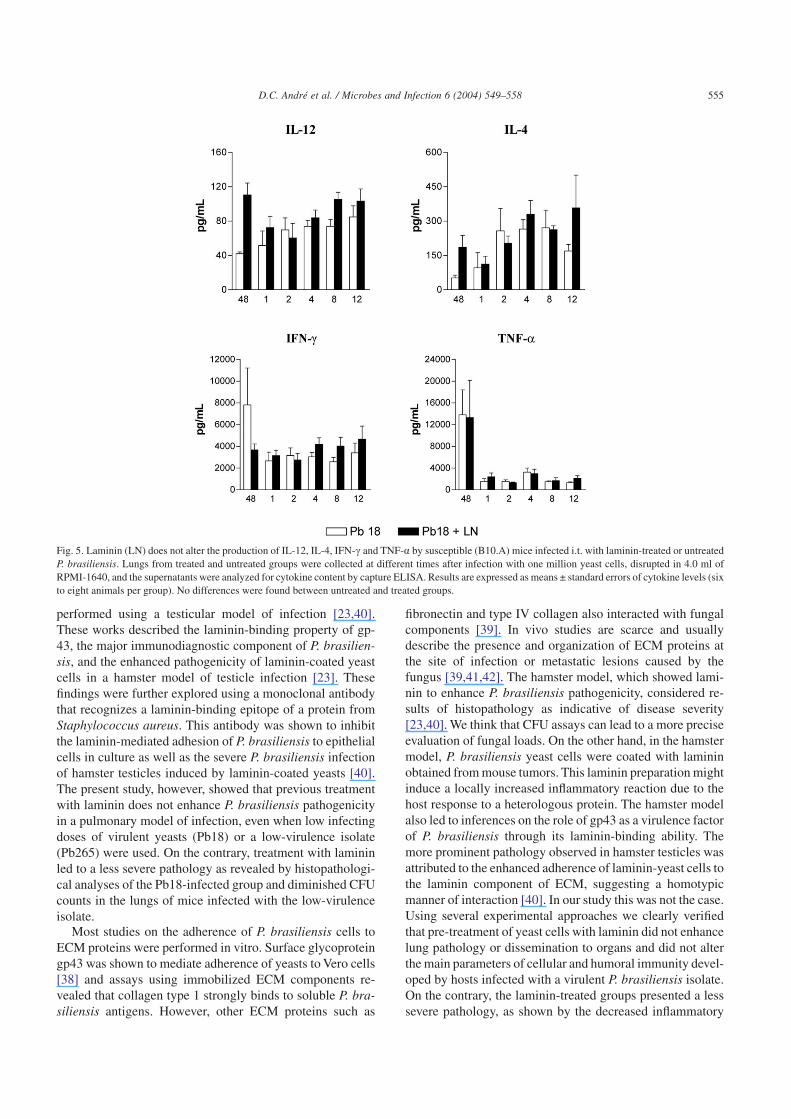
Previous studies showed the influence of cytokines on theinflammatory reactions in the lungs and on PCM outcome[12–14,16]. In addition, our histopathological studiesshowed differences in the inflammatory reactions betweenlaminin-treated and untreated groups, leading us to examinethe levels of some pulmonary cytokines. IL-4, IL-12, IFN-cand TNF-a were measured in lung homogenates from thevery early periods of infection (48 h) through the chronicphase of disease (week 12). Fig. 5 shows that previous incu-bation of P. brasiliensis yeasts with laminin did not alter thelevels of these cytokines in lung homogenates.
3.6. Laminin treatment does not influence the severityof infection caused by different inoculum sizesof a virulent P. brasiliensis isolate
As B10.A and A/Sn mice infected with one million yeastcells of the virulent isolate of P. brasiliensis did not presentdifferences in the severity of infection, we infected suscep-tible B10.A mice with smaller inocula of virulent Pb18 toverify if other infective doses would reveal the laminin effecton P. brasiliensis pathogenicity. As can be seen in Fig. 6,susceptible mice infected with any of the three differentinocula of laminin-coated yeasts (1 × 106, 1 × 105 and 1 ×104) showed no differences in fungal burdens from controlgroups.
Fig. 2. Photomicrographs of lesions from susceptible (B10.A) mice infected i.t. with laminin-untreated or treated 106 P. brasiliensis (Pb18) yeast cells. Shownare pulmonary lesions of laminin-untreated (A) or treated mice (B) at week 4 postinfection. In this period of infection, a more prominent inflammatory reactionwas detected in the laminin-untreated group.
Fig. 3. Laminin (LN) does not affect DTH responses of susceptible (B10.A) and resistant (A/Sn) mice infected i.t. with laminin-treated or untreatedP. brasiliensis yeast cells. Animals were infected i.t. with one million yeasts, and at weeks 2, 4, 8 and 12 after infection, were injected intrafootpad with a solubleyeast antigen 24 h before measurement of footpad response. The horizontal broken lines denote means + 2 standard deviations (confidence interval, 95%) ofnon-infected A/Sn and B10.A mice subjected to the footpad test (n = 15). No differences were found between untreated and treated groups.
553D.C. André et al. / Microbes and Infection 6 (2004) 549–558

3.7. Laminin decreases pathogenicity of a low-virulenceP. brasiliensis isolate (Pb265)
As the high virulence of Pb18 could be masking theinfluence of laminin on the infectivity of P. brasiliensis, weinfected B10.A mice with laminin-treated and untreatedPb265, an isolate that was previously shown to induce a mildand regressive disease in the susceptible strain [16]. Unex-pectedly, previous treatment of Pb265 yeast cells with lami-nin induced a less severe pulmonary infection (Fig. 7). LowerCFU counts were detected in the lungs of the laminin-coatedgroup, but no differences were seen in the levels of pulmo-nary cytokines as well as in the production of P. brasiliensis-specific antibodies (Fig. 8).
4. Discussion
As PCM is acquired by inhalation of airborne propagules,the first step in host–parasite interactions must occur in the
alveoli. The inflammatory mediators induced by early recog-nition of fungal elements by alveolar macrophages and theinitial inflammatory cell influx at the site of infection maylead to epithelial tissue damage. This fact can result in theaccessibility of basement membrane components, namelylaminin, type IV collagen, entactin/nidogen and heparan sul-fate proteoglycans. All these components as well as fibronec-tin and type I collagen present in the interstitial tissue havebeen proposed as ligands for fungi cell adhesins [33–37].
Previous studies characterized some extracellular matrix(ECM) components involved in host–P. brasiliensis interac-tions [23,38–40], and laminin was putatively characterized asan important element, which facilitates P. brasiliensis patho-genicity [23,40]. These facts led us to ask whether laminintreatment of yeast cells would influence pulmonary P. brasil-iensis infection. This question was indeed relevant sinceP. brasiliensis is a lung pathogen and the in vivo studiesshowing the influence of laminin on fungal infection were
Fig. 4. Laminin (LN) does not affect antibody production of susceptible (B10.A) mice infected i.t. with laminin-treated or untreated P. brasiliensis. Seraobtained at weeks 2, 4, 8 and 12 of infection were assayed for IgM, IgA, IgG1, IgG2a, IgG2b and IgG3 by using an isotype-specific ELISA. The bars depictlog2 ± standard errors of serum titers. No differences were found between untreated and treated groups.
554 D.C. André et al. / Microbes and Infection 6 (2004) 549–558
performed using a testicular model of infection [23,40].These works described the laminin-binding property of gp-43, the major immunodiagnostic component of P. brasilien-sis, and the enhanced pathogenicity of laminin-coated yeastcells in a hamster model of testicle infection [23]. Thesefindings were further explored using a monoclonal antibodythat recognizes a laminin-binding epitope of a protein fromStaphylococcus aureus. This antibody was shown to inhibitthe laminin-mediated adhesion of P. brasiliensis to epithelialcells in culture as well as the severe P. brasiliensis infectionof hamster testicles induced by laminin-coated yeasts [40].The present study, however, showed that previous treatmentwith laminin does not enhance P. brasiliensis pathogenicityin a pulmonary model of infection, even when low infectingdoses of virulent yeasts (Pb18) or a low-virulence isolate(Pb265) were used. On the contrary, treatment with lamininled to a less severe pathology as revealed by histopathologi-cal analyses of the Pb18-infected group and diminished CFUcounts in the lungs of mice infected with the low-virulenceisolate.
Most studies on the adherence of P. brasiliensis cells toECM proteins were performed in vitro. Surface glycoproteingp43 was shown to mediate adherence of yeasts to Vero cells[38] and assays using immobilized ECM components re-vealed that collagen type 1 strongly binds to soluble P. bra-siliensis antigens. However, other ECM proteins such as
fibronectin and type IV collagen also interacted with fungalcomponents [39]. In vivo studies are scarce and usuallydescribe the presence and organization of ECM proteins atthe site of infection or metastatic lesions caused by thefungus [39,41,42]. The hamster model, which showed lami-nin to enhance P. brasiliensis pathogenicity, considered re-sults of histopathology as indicative of disease severity[23,40]. We think that CFU assays can lead to a more preciseevaluation of fungal loads. On the other hand, in the hamstermodel, P. brasiliensis yeast cells were coated with lamininobtained from mouse tumors. This laminin preparation mightinduce a locally increased inflammatory reaction due to thehost response to a heterologous protein. The hamster modelalso led to inferences on the role of gp43 as a virulence factorof P. brasiliensis through its laminin-binding ability. Themore prominent pathology observed in hamster testicles wasattributed to the enhanced adherence of laminin-yeast cells tothe laminin component of ECM, suggesting a homotypicmanner of interaction [40]. In our study this was not the case.Using several experimental approaches we clearly verifiedthat pre-treatment of yeast cells with laminin did not enhancelung pathology or dissemination to organs and did not alterthe main parameters of cellular and humoral immunity devel-oped by hosts infected with a virulent P. brasiliensis isolate.On the contrary, the laminin-treated groups presented a lesssevere pathology, as shown by the decreased inflammatory
Fig. 5. Laminin (LN) does not alter the production of IL-12, IL-4, IFN-c and TNF-a by susceptible (B10.A) mice infected i.t. with laminin-treated or untreatedP. brasiliensis. Lungs from treated and untreated groups were collected at different times after infection with one million yeast cells, disrupted in 4.0 ml ofRPMI-1640, and the supernatants were analyzed for cytokine content by capture ELISA. Results are expressed as means ± standard errors of cytokine levels (sixto eight animals per group). No differences were found between untreated and treated groups.
555D.C. André et al. / Microbes and Infection 6 (2004) 549–558

reaction in the lungs. This finding was not associated withdecreased production of pro-inflammatory cytokines(TNF-a, IL-12 and IFN-c), but the low production of otherpro-inflammatory cytokines/chemokines or enhanced levels
of anti-inflammatory mediators could account for the lessintense inflammation observed in the laminin-treated groups.
In several diseases, chemokines appear to play a majorrole in regulating the migration of specific leukocyte subsetsin both the acute and chronic inflammatory processes [43].Indeed, recent studies showed an association between highlevels of RANTES, MCP-1, IP-10 and Mig and mononuclearcell recruitment to the lungs of P. brasiliensis-infectedC57BL/6 mice [44]. To better understand the less intenseinflammatory influx of mononuclear cells induced bylaminin-treated P. brasiliensis, further studies on chemokineproduction as well as on the synthesis of anti-inflammatorycytokines such as IL-10 and TGF-b will be performed in ourlaboratory.
A very elucidative fact was the recovery of lower fungalloads from lungs of mice infected with laminin-treatedPb265. Only with the low-virulence isolate were we able todetect the inhibitory activity of the exogenous laminin onP. brasiliensis infectivity. It is tempting to speculate thatprevious in vitro treatment with laminin covered laminin-binding epitopes on gp43, avoiding their in vivo interactionwith laminin molecules present in damaged alveolar spaces,resulting in lower adherence of fungal cells and decreasedsynthesis of chemotactic mediators and cell adhesion mol-ecules. This effect would result in diminished influx of in-flammatory cells to the lungs of Pb18-infected mice anddecreased fungal burdens in the lungs of Pb265-infectedmice. The protective effect in the Pb18 experiments, how-ever, appeared to be transitory since later in the infection noeffect on disease severity was observed. The diminishedadhesion of yeast cells to host components appeared to besufficient to decrease lung colonization by the low-virulenceisolate, but not the highly virulent Pb18. Furthermore, sev-eral other P. brasiliensis components were shown to bind to
Fig. 6. Effect of different inoculum sizes on the severity of paracoccidioido-mycosis of susceptible (B10.A) mice infected with laminin-treated or un-treated P. brasiliensis. The bars depict means ± standard errors of log10 CFUobtained from groups of 6–8 mice at week 4 after infection with 1 × 106, 1 ×105 or 1 × 104 yeast cells. No significant differences were detected betweenlaminin-treated and untreated groups.
Fig. 7. Laminin (LN) diminishes the infectivity of a low-virulence (Pb265)P. brasiliensis isolate. B10.A mice were infected i.t. with one million yeastcells of Pb265 previously treated or untreated with laminin. The bars depictmeans ± standard errors of log10 CFU obtained from groups of 6–8 mice atweek 4 after infection. Significant differences were detected betweenlaminin-treated and untreated groups (P < 0.05).
556 D.C. André et al. / Microbes and Infection 6 (2004) 549–558

ECM proteins [39] and could account for the yeast adhesionto alveolar basement membrane proteins. Our findings alsosuggest that isolates with different degrees of virulence couldexpress a diverse pattern of adhesins or even different con-centrations of the same fungal adhesin. In conclusion, ourdata appear to be important to the understanding of the initialsteps of pulmonary PCM and indicate that laminin and prob-ably other ECM components are involved in the fungal infec-tion of lungs. Moreover, they demonstrate that binding oflaminin does not enhance pathogenicity of P. brasiliensisyeast cells when the primary site of infection is the lung, as isbelieved to occur in the human disease.
Acknowledgements
We are grateful to T. Alves and B.P. Albe for technicalassistance. This work was supported by a grant (98/13766-0)from the Fundação de Amparo à Pesquisa do Estado de SãoPaulo (FAPESP).
References
[1] M.F. Franco, R.P. Mendes, M. Moscardi-Bachi, M.T. Reskallah-Iwasso, M.R. Montenegro, Paracoccidioidomycosis, Baillières Clin.Trop. Méd. Comum 4 (1989) 185–220.
[2] E. Brummer, E. Castañeda, A. Restrepo, Paracoccidioidomycosis: anupdate, Clin. Microbiol. Rev. 6 (1993) 89–117.
[3] G. Benard, A. Durandy, C.M. Assis, M.A. Hong, N.M. Orii,N.M. Sato, M.J.S. Mendes-Giannini, C.S. Lacaz, A.J.S. Duarte,Responses of T and B lymphocytes to a Paracoccidioides brasiliensiscell wall extract in healthy sensitized and non-sensitized subjects,Amer. J. Trop. Med. Hyg. 53 (1995) 189–194.
[4] V.L.G. Calich, S.S. Kashino, Cytokines produced by susceptible andresistant mice in the course of Paracoccidioides brasiliensis infec-tion, Braz. J. Med. Biol. Res. 31 (1998) 615–623.
[5] V.L.G. Calich, C.A.C. Vaz, E. Burger, Immunity to Paracoccidioidesbrasiliensis infection, Res. Immunol. 149 (1998) 407–416.
[6] A.S.K. Karhawi, A.L. Colombo, R. Salomão, Production of IFN-c isimpaired in patients with paracoccidioidomycosis during active dis-ease and is restored after clinical remission, Med. Mycol. 38 (2000)225–229.
[7] S.S. Kashino, R.A. Fazioli, C. Cafalli-Favati, L.H. Meloni-Bruneri,C.A.C. Vaz, E. Burger, V.L.G. Calich, Resistance to Paracoccidiodesbrasiliensis infection is linked to a preferential Th1 immune responsewhereas susceptibility is associated with absence of IFN-c produc-tion, J. Interf. Cyt. Res. 20 (2000) 89–97.
[8] R.L.R. Mamoni, A.M.S. Nouer, S.A. Oliveira, C.C. Musatti,C.L. Rossi, Z.P. Camargo, M.H.S.L. Blotta, Enhanced production ofspecific IgG4, IgE, IgA and TGF-b in sera from patients with thejuvenile form of paracoccidioidomycosis, Med. Mycol. 40 (2002)1–7.
[9] S.J. Oliveira, R.L. Mamoni, C.C. Musatti, P.M. Papaiodarnau,M.H.S.L. Blotta, Cytokines and lymphocyte proliferation in juvenileand adult forms of paracoccidioidomycosis: comparison with infectedand non-infected controls, Microb. Infect. 4 (2002) 139–144.
[10] L.E. Cano, L.M. Singer-Vermes, C.A.C. Vaz, M. Russo,V.L.G. Calich, Pulmonary paracoccidioidomycosis in resistant andsusceptible mice: relationship among progression of infection, bron-choalveolar cell activation, cellular immune response and specificisotype patterns, Infect. Immun. 63 (1995) 1777–1783.
[11] L.E. Cano, L.M. Singer-Vermes, J.A. Mengel, C.F. Xidieh, C. Arruda,D.C. André, C.A.C. Vaz, E. Burger, V.L.G. Calich, Depletion of CD8T cells in vivo impairs host defense of resistant and susceptible miceto pulmonary paracoccidioidomycosis, Infect. Immun. 68 (2000)352–359.
[12] C. Arruda, M.F. Franco, S.S. Kashino, F.R.F. Nascimento,R.A. Fazioli, C.A.C. Vaz, M. Russo, V.L.G. Calich, IL-12 protectsmice against disseminated infection caused by Paracoccidioides bra-siliensis but enhances pulmonary inflammation, Clin. Immunol. 103(2002) 185–195.
[13] L.E. Cano, S.S. Kashino, C. Arruda, D.C. André, C.F. Xidieh,L.M. Singer-Vermes, C.A.C. Vaz, E. Burger, V.L.G. Calich, Protectiverole of interferon-gamma in experimental pulmonary paracoccidio-idomycosis, Infect. Immun. 66 (1998) 800–806.
[14] G.S. Deepe, L. Romani, V.L.G. Calich, G. Huffnagle, C. Arruda,E.E.I.W. Molinari-Madlum, J.R. Perfect, Knockout mice as experi-mental models of virulence, Med. Mycol. 38 (2000) 87–98.
Fig. 8. Laminin (LN) does not affect antibody production and the levels ofpulmonary cytokines of susceptible (B10.A) mice infected i.t. with laminin-treated or untreated Pb265. (A) Sera obtained at week 4 of infection wereassayed for IgM, IgA, IgG1, IgG2a, IgG2b and IgG3 by using an isotype-specific ELISA. The bars depict log2 ± standard errors of serum titers. Nodifferences were found between untreated and treated groups. (B) Lungswere collected at week 4 after infection, disrupted in 4.0 ml of RPMI-1640,and the supernatants were analyzed for cytokine ( IL-12, IL-4, IFN-c andTNF-a) content by capture ELISA. Results are expressed as means ± stan-dard errors of cytokine levels (six to ten animals per group). No differenceswere found between untreated and treated groups.
557D.C. André et al. / Microbes and Infection 6 (2004) 549–558

[15] F.R.F. Nascimento, V.L.G. Calich, D. Rodrigues, M. Russo, Dual roleof nitric oxide in paracoccidioidomycosis: essential for resistance butoverproduction associated with susceptibility, J. Immunol. 168 (2002)4593–4600.
[16] J.F. Souto, F. Figueiredo, A. Furlanetto, K. Pfeffer, M.A. Rossi,J.S. Silva, Interferon-c and tumor necrosis factor–a determine resis-tance to Paracoccidioides brasiliensis infection, Am. J. Pathol. 156(2000) 1811–1820.
[17] J.M. Patti, B.L. Allen, M.J. McGavin, M. Hook, MSCRAMM—Mediated adherence of microorganisms to host tissues, Review, Annu.Rev. Microbiol. 48 (1994) 585–617.
[18] E.C. Crouch, G.R. Martin, J.S. Brody, in: R.G. Crystal, J.B. West(Eds.), The Lung: Scientific Foundations, Raven Press, New York,1991, pp. 421–438.
[19] G.C. Furtado, M. Slowick, H.K. Kleinman, K.A. Joiner, Lamininenhances binding of Toxoplasma gondii tachyzoïtes to J774 murinemacrophage cells, Infect. Immun. 60 (1992) 2337–2342.
[20] J. McMahon, J. Wheat, M.E. Sobel, R. Pasula, J.F. Downing,W.J. Martin II, Murine laminin binds to Histoplasma capsulatum, apossible mechanism of dissemination, J. Clin. Invest. 96 (1995) 1010–1017.
[21] J.F. Souto, F. Figueiredo, A. Furlanetto, K. Pfeffer, M.A. Rossi,J.S. Silva, Interferon-c and tumor necrosis factor–a determine resis-tance to Paracoccidioides brasiliensis infection, Am. J. Pathol. 156(2000) 1811–1820.
[22] T. Umemoto, I. Nakikawa, Binding of host-associated treponemaproteins to collagens and laminin: a possible mechanism of spiro-chetal adherence to host tissues, Microbiol. Immunol. 38 (1994)655–663.
[23] A.P. Vicentini, J.L. Gesztesi, M.F. Franco, W. Souza, J.Z. Moraes,L.R. Travassos, J.D. Lopes, Binding of Paracoccidioides brasiliensisto laminin through surface glycoprotein gp43 leads to enhancement offungal pathogenesis, Infect. Immun. 62 (1994) 1465–1469.
[24] L.M. Singer-Vermes, E. Burger, M.F. Franco, M. Moschardi-Bacchi,M.J.S. Mendes-Giannini, V.L.G. Calich, Evaluation of the pathoge-nicity and immunogenicity of seven Paracoccidioides brasiliensisisolates in susceptible inbred mice, J. Med. Vet. Mycol. 27 (1989)71–82.
[25] S.S. Kashino, L.M. Singer-Vermes, V.L.G. Calich, E. Burger, Alter-ations in the pathogenicity of one Paracoccidioides brasiliensis iso-late do not correlate with its in vitro growth, Mycopathologia 111(1990) 173–180.
[26] C. Fava-Netto, V.S. Vegas, I.M. Sciannaméa, E.D.B. Guarnieri, Antí-geno polissacarídico do Paracoccidioides brasiliensis. Estudo dotempo de cultivo do P. brasiliensis necessário ao preparo do antí-geno, Rev. Inst. Med. Trop. São Paulo 11 (1969) 177–181.
[27] M.D. Berliner, M.E. Reca, Vital staining of Histoplasma capsulatumwith Janus Green B, Sabouraudia 5 (1966) 26–29.
[28] H.K. Kleinman, F.B. Cannon, G.W. Laurie, J.R. Hassel, M. Aumail-ley, V.P. Terranova, G.R. Martin, M. Dubois-Daleq, Biological activi-ties of laminin, J. Cell. Biochem. 27 (1985) 317–325.
[29] L.M. Singer-Vermes, M.C. Ciavaglia, S.S. Kashino, E. Burger,V.L.G. Calich, The source of the growth-promoting factor(s) affectsthe plating efficiency of Paracoccidioides brasiliensis, J. Med. Vet.Mycol. 30 (1992) 261–264.
[30] R.A. Fazioli, L.M. Singer-Vermes, S.S. Kashino, E. Burger,M. Franco, M. Moschardi-Bacchi, V.L.G. Calich, Delayed-typehypersensitivity response in an isogenic murine model of paracoccid-ioidomycosis, Mycopathologia 126 (1994) 137–146.
[31] Z.P. Camargo, C.P. Taborda, E.G. Rodrigues, L.R. Travassos, The useof cell-free-antigens of Paracoccidioides brasiliensis in serologicaltests, J. Med. Vet. Mycol. 29 (1991) 31–38.
[32] J.H. Zar, Biostatistical Analysis, second ed, Prentice Hall, New Jersey,1984.
[33] M.L. Gil, M.C. Peñalter, J.L. Lopez-Ribot, J.E. O’Connor, J.P. Mar-tinez, Binding of extracellular matrix proteins to Aspergillus fumiga-tus conidia, Infect. Immun. 64 (1996) 5239–5247.
[34] M.K. Hostetter,Adherence molecules in pathogenic fungi, Curr. Opin.Infect. Dis. 9 (1996) 141–145.
[35] A.S. Klotz, R.L. Smith, Gelatin fragments block adherence of Can-dida albicans to extracellular matrix proteins, Microbiology 141(1995) 2681–2684.
[36] J.L. López-Ribot, W.L.J. Chaffin, Binding of extracellular matrixcomponents to Candida albicans, Infect. Immun. 62 (1994) 4564–4571.
[37] M.C. Peñalter, J.E. O’Connor, J.P. Martinez, M.L. Gil, Binding ofhuman fibronectin to Aspergillus fumigatus conidia, Infect. Immun.64 (1996) 1146–1153.
[38] S.A. Hanna, J.L. Monteiro da Silva, M.J.S.M. Giannini, Adherenceand intracellular parasitism of Paracoccidioides brasiliensis in Verocells, Microb. Infect. 2 (2000) 877–884.
[39] M.J.S. Mendes-Giannini, M.L. Taylor, J.B. Bouchara, E. Burger,V.L.G. Calich, E.D. Escalante, S.A. Hanna, H.L. Lenzi,M.P. Machado, M. Myiaji, J.L. Monteiro da Silva, E.M. Mota,A. Restrepo, S. Restrepo, G. Tronchin, L.R. Vicenzi, C.F. Xidieh,E. Zenteno, Pathogenesis II. Fungal responses to host responses:interaction of host cells with fungi, Med. Mycol. 38 (2000) 113–123.
[40] A.P. Vicentini, J.Z. Moraes, J.L. Gesztesi, M.F. Franco, W. De Souza,J.D. Lopes, Laminin-binding epitope on gp43 from Paracoccidioidesbrasiliensis is recognized by a monoclonal antibody raised againstStaphylococcus aureus laminin receptor, J. Med. Vet. Mycol. 35(1997) 37–43.
[41] E. Burger, C.A.C. Vaz, A. Sano, V.L.G. Calich, L.M. Singer-Vermes,C.F. Xidieh, S.S. Kashino, K. Nishimura, M. Miyaji, Paracoccidio-ides brasiliensis infection in nude mice: studies with isolates differingin virulence and definition of their T-cell-dependent and T-cell-independent components, Amer. J. Trop. Med. Hyg. 55 (1996) 235–242.
[42] C.F. Xidieh, H.L. Lenzi, V.L.G. Calich, E. Burger, Influence of thegenetic background on the pattern of lesions developed by resistantand susceptible mice infected with Paracoccidioides brasiliensis,Microbiol. Immunol. 188 (1999) 42–49.
[43] A.D. Luster, Chemokines: chemotactic cytokines that mediate inflam-mation, New Eng. J. Med. 12 (1998) 436–445.
[44] J.T. Souto, J.C. Aliberti, A.P. Campanelli, M. Livonesi,C.M.L. Maffei, B.R. Ferreira, L.R. Travassos, R. Martinez,M.A. Rossi, J.S. Silva, Chemokine production and leukocyte recruit-ment to the lungs of Paracoccidioides brasiliensis-infected mice ismodulated by interferon-c, Am. J. Pathol. 163 (2003) 583–590.
558 D.C. André et al. / Microbes and Infection 6 (2004) 549–558
Related Documents











