BEHAVIORAL EFFECTS OF NEUROTENSIN Tj.B. van Wimersma Greidanus, M.C.G. van Praag, R. Kalmann, G.J.E. Rinkel, G. Croiset, E.C. Hoeke, M.A.H. van Egmond, and M. Fekete Riidolf Magnus Institute for Pharmacology University of Utrecht, Medical Fuculty 3521 GD Utrecht. the Netherlands I NTKODUCTION Numerous reports in literature deal with central nervous system effects of the tridecapeptide neurotensin. ' Most of these reports concern the effects of this neuropeptide on body t e m p e r a t ~ r e , ~ - ~ on locomotor activity '.:' and on responsive- ness to a painful s t i m u l ~ s . ~ - ~ Relatively small quantities of neurotensin induce hypothe~mia,~-~ diminish locomotor activity,2, '' and potentiate some actions of barbiturates.'.'' In addition, neurotensin increases reaction time of mice in a hot- plate t e ~ t . ~ - ~ Neurotensin exerts these effects after central, mostly intracerebro- ventricular, administration but not following systemic injection. This is probably due to the poor permeability of the blood-brain barrier to penetration by this neuropeptide. ' In order to further evaluate the influence of neurotensin on the central nervous system and to extend its behavioral profile, we examined the effects of this neuro- peptide in a variety of behavioral tests, such as open field behavior, and active as well as passive avoidance behavior. Moreover, the effects of neurotensin on responsiveness to electric footshock were studied. MATERIAL AND METHODS Male rats of an inbred Wistar strain, weighing 120-160 g, were used. A poly- ethylene cannula was placed in the right lateral ventricle of the brain for intra- cerebroventricular (i .c.v.) administration of neurotensin or artificial cerebrospinal fluid (CSF), which was used as a placebo. The operation was performed 3 or 4 days before behavioral experimentation. Locomotor activity of the animals was studied in a circular open field (diameter 75 cm) during 3-min sessions held on each of 3 consecutive days. The behavioral parameters used during the sessions were ambulation (frequency of crossings of lines drawn on the floor of the open field), rearing, grooming, and defe~ation.~ Neurotensin was i.c.v. administered in doses of 3 ng, 30 ng, 300 ng, 3 pg, and 10 pg 1 hour before each session. In addition haloperidol was administered in a dose of 10 p g in a separate group of animals as well. For studying the effect of neurotensin on amphetamine-induced increase in locomotor activity, a separate experiment was performed in which amphetamine was intraperitoneally (i.p.) injected 1 hour before a single behavioral observation in a dose of 3 mg/kg. Neurotensin was i.c.v. injected at the same time in a dose of 300 ng. Responsiveness of rats to electric footshock (EFS) was determined by scoring 3 19 O77-8923 / 82 / MOO-03 19 $0 I .75/ 0 0 1982. NYAS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

BEHAVIORAL EFFECTS OF NEUROTENSIN
Tj.B. van Wimersma Greidanus, M.C.G. van Praag, R. Kalmann, G.J.E. Rinkel, G. Croiset, E.C. Hoeke,
M.A.H. van Egmond, and M. Fekete
Riidolf Magnus Institute for Pharmacology University of Utrecht, Medical Fuculty
3521 GD Utrecht. the Netherlands
I NTKODUCTION
Numerous reports in literature deal with central nervous system effects of the tridecapeptide neurotensin. ' Most of these reports concern the effects of this neuropeptide on body t e m p e r a t ~ r e , ~ - ~ on locomotor activity '.:' and on responsive- ness to a painful s t i m u l ~ s . ~ - ~ Relatively small quantities of neurotensin induce hypothe~mia ,~-~ diminish locomotor activity,2, '' and potentiate some actions of barbiturates.'.'' In addition, neurotensin increases reaction time of mice in a hot- plate t e ~ t . ~ - ~ Neurotensin exerts these effects after central, mostly intracerebro- ventricular, administration but not following systemic injection. This is probably due to the poor permeability of the blood-brain barrier to penetration by this neuropeptide. '
In order to further evaluate the influence of neurotensin on the central nervous system and to extend its behavioral profile, we examined the effects of this neuro- peptide in a variety of behavioral tests, such as open field behavior, and active as well as passive avoidance behavior. Moreover, the effects of neurotensin on responsiveness to electric footshock were studied.
MATERIAL AND METHODS
Male rats of an inbred Wistar strain, weighing 120-160 g, were used. A poly- ethylene cannula was placed in the right lateral ventricle of the brain for intra- cerebroventricular (i .c.v.) administration of neurotensin or artificial cerebrospinal fluid (CSF), which was used as a placebo. The operation was performed 3 or 4 days before behavioral experimentation.
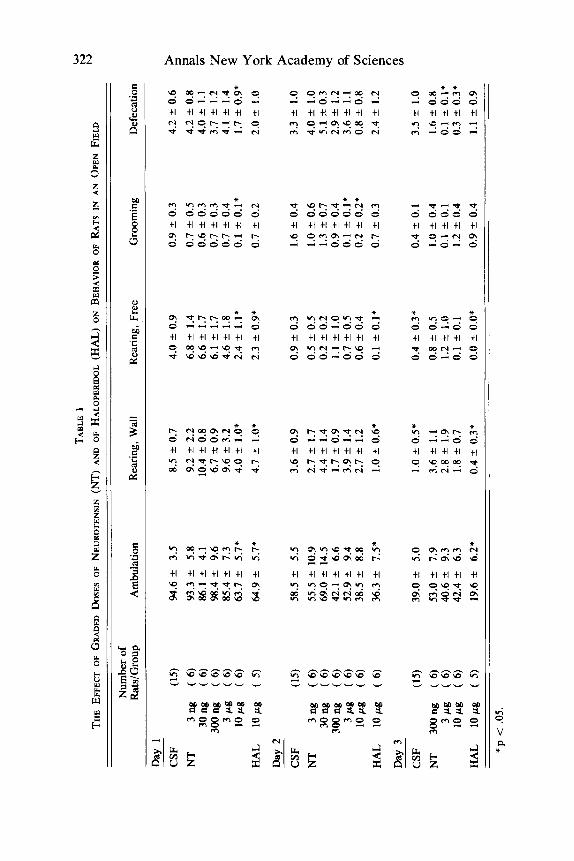
Locomotor activity of the animals was studied in a circular open field (diameter 75 cm) during 3-min sessions held on each of 3 consecutive days. The behavioral parameters used during the sessions were ambulation (frequency of crossings of lines drawn on the floor of the open field), rearing, grooming, and defe~at ion .~ Neurotensin was i.c.v. administered in doses of 3 ng, 30 ng, 300 ng, 3 pg, and 10 pg 1 hour before each session. In addition haloperidol was administered in a dose of 10 pg in a separate group of animals as well.
For studying the effect of neurotensin on amphetamine-induced increase in locomotor activity, a separate experiment was performed in which amphetamine was intraperitoneally (i.p.) injected 1 hour before a single behavioral observation in a dose of 3 mg/kg. Neurotensin was i.c.v. injected at the same time in a dose of 300 ng.
Responsiveness of rats to electric footshock (EFS) was determined by scoring
3 19 O77-8923 / 82 / MOO-03 19 $0 I .75/ 0 0 1982. NYAS

320 Annals New York Academy of Sciences
their percentages of jerk/run/jump reactions, flinches, and no responses to various shock levels, ranging from 0.03 1 mA to 0.187 mA.8 Ten different shock levels were used and each shock level was presented twice. The 20 EFS’s were presented in a randomly futed order. The duration of each EFS presentation was 1 sec and the interval between presentations was 20 sec. The effect of neurotensin on respon- siveness to EFS was studied in an acute experiment using graded doses (3 ng, 30 ng, 300 ng, and 3 pg) of the peptide i.c.v. injected 1 hour before the session. Also the influence of naltrexone and naloxone (1 pg, i.c.v.) on the neurotensin (300 ng, i.c.v.) induced change in responsiveness to EFS was investigated. In addition it was studied whether tolerance developed to the neurotensin-induced change in responsiveness to EFS by measuring the responsiveness to EFS of rats treated with 300 ng neurotensin (i.c.v.) for three consecutive days.
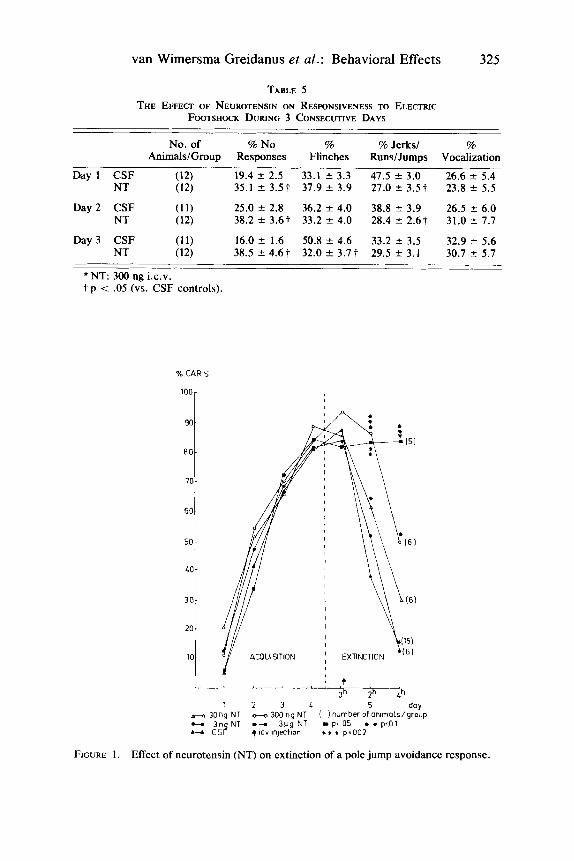
Active avoidance behavior was studied in a pole-jump, one-way, active avoid- ance situation. Rats were trained to jump onto a pole in order to avoid a scrambled EFS of .20 mA. The light of a bulb placed on top of the box was used as a conditioning stimulus (CS), which was presented 5 sec before the unconditioning stimulus (US) of EFS. As soon as the rat made an avoidance or an escape re- sponse, the presentation of the CS or of the CS/US combination was terminated immediately. Acquisition sessions of 10 trialslday with a mean intertrial interval of 60 sec were run on 4 consecutive days. On day 5 , extinction trials were run during which only the CS was presented. During extinction, the CS presentation was terminated as soon as the animal made a response. In case an animal did not respond to CS presentation within 5 sec, the light was switched off after 5 sec. Three extinction sessions of 10 trials/session were performed with a 2-hr i n t e ~ a l . ~ Neurotensin was administered in doses of 3 ng, 30 ng, 300 ng, and 3 pg immedi- ately after the first extinction session.
Passive avoidance behavior was studied in a simple step-through situation. lo
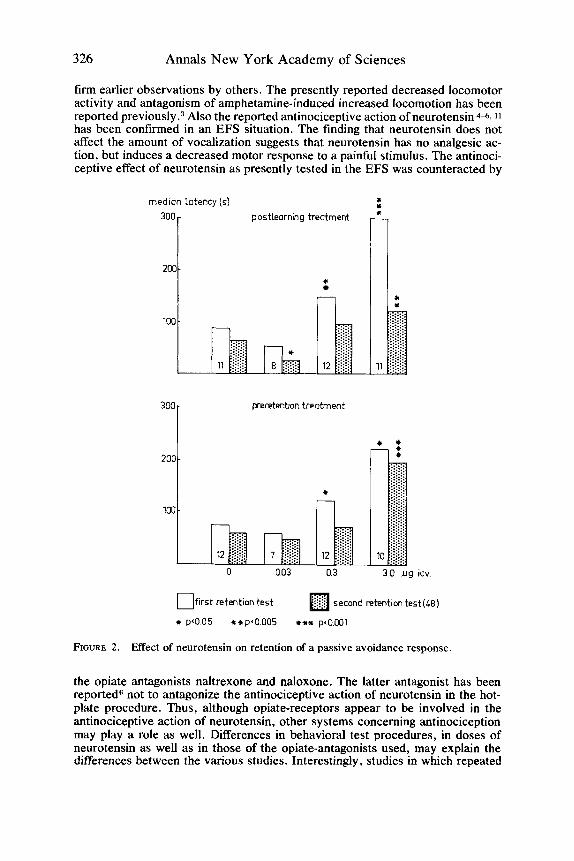
The apparatus consisted of a dark compartment to which an illuminated elevated platform was attached. After adaptation to the situation, the initial latency of rats to enter the dark compartment was registered during 1 trial, on day 1 and during 3 trials on day 2. Immediately after entering the dark compartment during the third trial on day 2, the rats received a scrambled EFS (.25 mA) during 2 sec (learning trial). Avoidance latencies were registered during retention sessions performed at 24 hours and at 48 hours after the learning trial. A maximal observation time of 300 sec was used during the retention trials. Neurotensin was i.c.v. administered in doses of 30 ng, 300 ng, and 3 pg either immediately after the learning trial or 1 hr before the first retention session.
For statistical evaluation of the results, Student’s t-test, Mann-Whitney’s U-test or Dunnett’s test were used as appropriate.
RESULTS
Locomotor Activity in the Open Field
Only the highest dose of neurotensin induced a significant effect on the be- havior of rats in an open field. Ambulation as well as rearing, grooming, and defecation were significantly diminished after treatment with 10 Fg neurotensin during the first day of observation. The other doses applied were all ineffective in this respect. On the second day, this effect of neurotensin was less pronounced and only a significant reduction of ambulation and of grooming was observed. On

van Wimersma Greidanus et al.: Behavioral Effects 32 1
the third day, no effects of neurotensin were seen in this behavioral situation, except a reduction of defecation was noticed (TABLE 1). In contrast, the neurolep- tic agent haloperidol induced a continuous reduction in ambulation during the three days of observation.
Neurotensin appeared to antagonize the stimulatory influence of amphetamine on ambulation and rearing in an open field. Amphetamine induced a marked increase in ambulation and rearing. This effect was not observed when amphe- tamine was administered in combination with neurotensin (TABLE 2).
Responsiveness to Electric Footshock
In the acute experiment, neurotensin exerts a dose-dependent decrease in responsiveness to electric footshock. This was demonstrated by the increased percentage of “no responses” and a decreased percentage of jerks/run/jumps following neurotensin administration (TABLE 3). The change in responsiveness to EFS was not accompanied by changes in the amount of vocalization. If naltrexone or naloxone (1 pg i.c.v.) was given at the same time as neurotensin (300 ng), the decreased responsiveness to EFS as induced by neurotensin was antagonized by the opiate antagonists (TABLE 4).
Observation of the neurotensin effect on responsiveness to EFS during three consecutive days revealed that not only on the first day of observation but also on the second and third day, neurotensin decreased responsiveness to EFS. This was illustrated by the increased percentage of no responses on days 2 and 3 and by the reduction of intense or weak motor reaction on days 2 and 3 respectively (TABLE 5). In addition, no differences in vocalization were found.
Active Avoidance Behavior
Neurotensin induced a dose-dependent inhibition of extinction of a conditioned avoidance response (CAR). Placebo-treated rats displayed percentages of 38% and 15% during the two extinction sessions performed at 2 and 4 hours after treatment. 30 ng of the neuropeptide resulted in a mean of 63% CAR’s during the first extinction session after treatment and 29% CAR’s during the second extinction session. These percentages amounted to 87% and 50% for the dose of 300 ng and to 83% and 84% for the dose of 3 pg (FIG. 1).
Passive Avoidunce Behavior
Neurotensin increased passive avoidance latencies. Treatment of rats with 300 ng neurotensin immediately after the learning trial as well as before the first retention session induced an increase in the median latency score during the first retention session only. A dose of 3 pg induced an increased latency during the first and the second retention sessions (FIG. 2). A low dose (3 ng) of neuroten- sin, when administered after the learning trial, induced a weak but significant attenuation of passive avoidance behavior at the second retention session.
Annals New York Academy of Sciences
* '9 091P?b.s 9
t I +I +I +I +I +I +I p! N.?"?? 9
0 0 - - - 0 - -t - t - t m - t - N
* c? Y c ? c ? 9 1 p! 0 0 0 0 0 0 0 +I t I t l tl +I +I +I s ?'9"?1" 0 0 0 0 0 0 0
* c? Y p ! 9 Y - ? ' 0 0 0 - 0 0 0
t I +I +I +I +I +I +I 2 y-?sz 2
* 2 Y?-?sr 3 +I ti +I +I +I t l +I '9 ?9r:Q!? 9 m c c b - m N -
* * 9 091? s - 000 0 t i +I +I +I t I Y '9-7- ? m - 0 0 -
- 9"9 9 0 000 0 +I t I t I +I +I p: 9"p! Q! 0 -0- 0
* * Y O ?
+I +I +I +I t I wee-
2 0-0 0 o- ;o 8
* * 0 --0 0 Y Y o ! ? c?
* ??$&! +I +I +I +I +I 9 9'99 '9 s 9
$ V a *
TA
BL
E 1
TH
E E
FFE
CT
OF GRADED
DO
SES O
F N
EW
RO
TE
NSI
N
(NT
) A
ND
O
F H
AL
OPE
R~W
L
(HA
L)
ON
B
EH
AV
IOR
O
F R
ATS
IN A
N
OPE
N FI
EL
D

TA
BL
E 2
TH
E E
FFE
CT
O
F N
EU
RO
TE
NSI
N
* O
N
AM
PHE
TA
MIN
E
t-im
ucm
AC
TIV
ITY
O
F R
ATS
IN A
N O
PEN
FIE
LD
Num
ber
of
i.c.
v./i
.p.
Rat
s/G
roup
A
mbu
latio
n R
eari
ng, W
all
Rea
ring
, Fre
e G
room
ing
Def
ecat
ion
CSF
/sal
ine
( 8)
C
SF/A
mph
. (
7)
137.
9 2
11
.1$
NT
/Am
ph.
( 8)
N
T/s
alin
e (1
5)
86.6
r
6.1
* NT
: 300
ng
i.c.
v.
t AM
PH:
0.3
mg
i.p.
$p
< .0
5 (v
s. C
SFis
alin
e co
ntro
ls).
5.1 2 0
.8
3.6
2 1
.0
84.8
2
9.2
10.8
f 2
.8
8.5
2 2
.0
0.1
-r 0.1
3.4
%
1.3
5.6 2 0
.6
102.
6 -r-
5.7
10.3
f 1
.7
6.5 f 2
.4
0.2 f 0
.1
0.1
2 0
.1
0.3
+. 0.
2
19.3
% 3
.7$
7.5 f 0
.8
14.6
% 2
.6$
5.7 ’’
1.0
w
h)
w

324 Annals New York Academy of Sciences
TABLE 3 THE EFFECT OF GRADED DOSES OF NEUROTENSIN (NT) ON RESFQNSIVENESS TO
ELECTRIC FOOTSHOCK
Number of % % % Jerks/ % Animals/Group No Responses Flinches Runs/ Jumps Vocalization
CSF 12.9 f 2.0 33.4 t 3.0 53.6 f 2.8 22.9 t 4.3 21.3 f 4.0 29.7 f 3.0 49.0 f 1.9 19.2 f 9.0 25.3 f 3.0* 26.0 f 3.5 48.7 f 3.0 22.1 * 6.6 33.7 f 3.0t 28.0 f 2.5 38.3 f 2.0" 20.0 f 6.9 27.6 f 2.4t 31.2 f 4.7 41.1 f 3.0* 22.5 f 5.7
NT 3ng ( 5 ) 30 ng (5 )
300 ng (5 ) 3 Pg (5 )
* p < .05 (vs. CSF controls). t p < .02 (vs. CSF controls).
DISCUSSION
Some effects of neurotensin have been claimed to be shared by neuroleptic agents.3, I ' It has been reported that neurotensin exerts effects in various pharma- cological and behavioral tests in which neuroleptic agents are active. It appeared that neurotensin releases prolactin and that bilateral nucleus accumbens injec- tions of neurotensin, like the neuroleptic drug haloperidol, significantly antago- nized increased locomotor activity and rearing induced by amphetamine .3f However, unlike neuroleptic drugs, neurotensin seems not to inhibit the binding of 3H-spiroperidol to dopamine receptors in rat brain membrane preparations from the nucleus caudatus or the nucleus a c c ~ m b e n s . ~ Thus, neurotensin seems to share several, but not all, properties with neuroleptic agents.3
Generally the results obtained from the presently described experiments con-
TABLE 4
INDUCED DECREASED RESFQNSIVENESS TO ELECTRIC FOOTSHOCK THE EFFECT OF NALTREXONE* AND OF NALOXONEt ON NEUROTENSIN$-
No. of % No % % Jerks/ % Animals/Group Responses Flinches Runs/Jumps Vocalization
15.8 f 2.0 30.8 t 3.4 53.4 t 4.0 22.9 f 4.3 21.7 f 3.1 34.2 f 1.6 44.1 f 2.1 18.3 rt 3.9 44.2 f 2.81) 24.2 t 3.3 31.6 f 4.91 20.0 2 6.9 24.1 * 2.4 27.5 t 2.5 48.4 2 2.5 20.8 f 5.3
CSF (12) 17.0 f 1.4 39.6 f 1.4 43.4 f 1.1 45.8 t 4.0 NALO (12) 17.4 t 1.4 38.6 f 1.3 44.4 f 1.7 36.5 f 3.0 NT (9) 25.0 f 1.27 39.8 f 2.6 35.2 rt 2.21 40.7 t 2.2 NT + NALO (10) 20.8 f 1.6 37.9 f 1.8 41.3 f 2.3 41.7 f 3.4
CSF ( 5 ) NALT (5 ) NT (5 ) NT + NALT (5 )
*NALT: 1 pg i.c.v. t NALO: 1 pg i.c.v. $NT: 300 ng i.c.v. 1 p < .05 (vs. CSF controls as well as vs. NT + NALT or NT + NALO). ( p < .02 (vs. CSF controls as well as vs. NT + NALO). IIp < .01 (vs. CSF controls as well as vs. NT + NALT).
van Wimersma Greidanus el af.: Behavioral Effects 325
1oor
90.
8 0 -
70
60.
50
40
30-
20.
10.
TABLE 5 THE EFFECT OF NEUROTENSIN ON RESPONSIVENESS TO ELECTRIC
FOOTSHOCK DURING 3 CONSECUTIVE DAYS
No. of % No % % Jerks/ % Anirnals/Group Responses Flinches RundJurnps Vocalization
Day 1 CSF (12) 19.4 2 2.5 33.1 f 3.3 47.5 2 3.0 26.6 f 5.4 NT (12) 35.1 f 3.5t 37.9 2 3.9 27.0 f 3.57 23.8 2 5.5
Day2 CSF (11) 25.0 f 2.8 36.2 f 4.0 38.8 f 3.9 26.5 f 6.0 NT (12) 38.2 2 3.67 33.2 ? 4.0 28.4 f 2.67 31.0 f 7.7
Day3 CSF (1 1) 16.0 f 1.6 50.8 f 4.6 33.2 f 3.5 32.9 f 5.6 NT (12) 38.5 f 4.6t 32.0k 3.7t 29.5 f 3.1 30.7 f 5.7
* NT: 300 ng i.c.v. t p < .05 (vs. CSF controls).
-
-
-
Qh 2h Lh
1 2 3 4 5 day w 30ng NT - 300ng NT ( )numberof onimals/group c 3n NT CI 3yg NT p i 05 + p . O 1 4~ CSF t ICY injection b t p a 002
FIGURE 1. Effect of neurotensin (NT) on extinction of a pole jump avoidance response.
3 26 Annals N e w York Academy of Sciences
200-
firm earlier observations by others. The presently reported decreased locomotor activity and antagonism of amphetamine-induced increased locomotion has been reported p rev i~us ly .~ Also the reported antinociceptive action of neurotensin 4 4 . I I has been confirmed in an EFS situation. The finding that neurotensin does not affect the amount of vocalization suggests that neurotensin has no analgesic ac- tion, but induces a decreased motor response to a painful stimulus. The antinoci- ceptive effect of neurotensin as presently tested in the EFS was counteracted by
median latency (s) t t
postlearning treatment
ff
--I I I:
preretention treatment 300r
n 0 0.03 0.3
n f i r s t retention test
.:.:.: :.:.:.
...... .:.:.: :::::: .:.:.: ......
3.0 ,ug icv
second retention test( l8)
* ~ ‘ 0 0 5 **p.0.005 *** p<O.001
FIGURE 2. Effect of neurotensin on retention of a passive avoidance response.
the opiate antagonists naltrexone and naloxone. The latter antagonist has been reported6 not to antagonize the antinociceptive action of neurotensin in the hot- plate procedure. Thus, although opiate-receptors appear to be involved in the antinociceptive action of neurotensin, other systems concerning antinociception may play a role as well. Differences in behavioral test procedures, in doses of neurotensin as well as in those of the opiate-antagonists used, may explain the differences between the various studies. Interestingly, studies in which repeated

van Wimersma Greidanus et a / . : Behavioral Effects 327
testing and treatment with neurotensin were performed did not reveal a develop- ment of tolerance to the antinociceptive effect of neurotensin.
Studies performed to further evaluate the behavioral activity of neurotensin did not indicate that neurotensin has in general a neuroleptic-like action. Neuroleptics like haloperidol applied to rats submitted to active pole-jump avoidance behavior induces a facilitation of extinction of a conditioned avoidance ~esponse . '~ How- ever neurotensin inhibits this extinction in a dose-dependent way, that is, opposite to what may be expected from treatment with a neuroleptic drug. In addition, neuroleptics attenuate passive avoidance behavior. l3 The results of the present experiments reveal that neurotensin administration during the acquisition as well as retention of the passive avoidance response generally results in increased latencies during retention sessions of this avoidance response. Thus, in fact neuro- tensin enhances passive avoidance behavior.
Whether or not the inhibitory effect of neurotensin on extinction of a CAR and the stimulatory effect of this neuropeptide on passive avoidance behavior are direct effects of neurotensin or mediated by a stimulation of the release of other neuropeptides (e.g., vasopressin and/or ACTH I*) by neurotensin remains to be elucidated.
In order to determine the possible physiological role of endogenous neuroten- sin in behavior, a preliminary experiment was performed in which the amount of biologically available neurotensin was reduced by i.c.v. application of antiserum to this neuropeptide. Undiluted neurotensin antiserum (anti-NT)* was i.c.v. in- jected in an amount of 3 p,l immediately after the first extinction session of the pole-jump avoidance response and the effect of this treatment was observed 4 hours later in a second extinction session. A tendency toward a facilitated extinc- tion of conditioned avoidance behavior following administration of the antiserum was observed. This tendency is opposite to the marked effect found after treatment with neurotensin itself. Since binding capacity and binding specificity of the anti- serum used are not known with respect to this kind of application, no conclusion can be drawn yet concerning the role of centrally available endogenous neuroten- sin in brain processes related to avoidance behavior.
In summary, it may be concluded that neurotensin shares some, but not all, effects with neuroleptic substances. In fact, it appears that neurotensin exerts some effects opposite to those induced by neuroleptics.
REFERENCES
1. NEMEROFF, C. B., D. LUTTINGER & A. J. PRANCE. 1980. Neurotensin: Central nerv- ous system effects of a neuropeptide. TINS September: 212-215.
2. NEMEROFF, C. B., G . BISSETTE, A. J. PRANCE JR., P. T. LOOSEN, T. S. BARLQW & M. A. LIPTON. 1W7. Neurotensin: Central nervous system effects of a hypothalamic peptide. Brain Res. 128: 4854%.
3. NEMEROFF, C. B. 1980. Neurotensin: Perchance an endogenous neuroleptic? Biol. Psychiatry IS: 283-302.
4. MARTIN, G. E., T. NARUSE & N. L. PAPP. 1981. Antinociceptive and hypothermic actions of neurotensin administered centrally in the rat. Neuropeptides 1: 447-454.
5. CLINESCHMIDT, B. V. & J. C. MCGUFFIN. 1977. Neurotensin administered intracis- ternally inhibits responsiveness of mice to noxious stimuli. Eur. J. Pharmacol. 46: 395-3%.
*Anti-NT was kindly supplied by Dr. R. E. Carraway, University of Massachusetts.

328 Annals New York Academy of Sciences
6. CLINESCHMIDT, B. V., I. C. MCGUFFIN & P. B. BUNTING. 1979. Neurotensin: Antino- cisponsive action in rodents. Eur. J. Pharmacol. 54: 129-139.
extinction of conditioned behavior. Prop. Brain Res. 32: 221-235. 8. GISPEN, W. H., A VAN DER POEL & TI. B. VAN WIMERSMA GREDANUS. 1973.
Pituitary-adrenal influences on behavior: Responses to test situations with or without electric footshock. Physiol. Behav. 10: 345-350.
9. VAN WIMERSMA GREDANUS, TI. B. & D. DE WIED. 1971. Effects of systemic and intracerebral administratin of two opposite acting ACTH-related peptides on extinc- tion of conditioned avoidance behavior. Neuroendocrinology 7: 291-301.
Retention of a passive avoid- ance response as a function of the intensity and duration of electric shock. Psychon. Sci. 26: 125-128.
11. ERVIN, G. N., L. S. BIRKEMO, C. B. NEMEROFF& A. J. PRANGE JR. 1981. Neurotensin blocks certain amphetamine-induced behaviors. Nature 291: 73-76.
12. RIVIER, C., M. BROWN & W. VALE. 1977. Effect of neurotensin, substance P and morphine sulfate on the secretion of prolactin and growth hormone in the rat. Endo- crinology 100: 751-754.
13. Kovics, G. L. & D. DE WIED. 1978. Effects of amphetamine and haloperidol on avoidance behavior and exploratory acitivity. Eur. J. Pharmacol. 53: 103-107.
14. LEEMAN, S. E., E. A. MROZ & R. E. CARRAWAY. 1977. Substance P and neurotensin. I n Peptides in Neurobiology. H. Gainer, Ed. Plenum Press. New York. pp. 99-144.
7. WEIINEN, J . A. W. M. & J. L. SLANGEN. 1970. Effects of ACTH analogues on
10. ADER, R., J. A. W. M. WEIJNEN & P. MOLEMAN. 1972.
DISCUSSION OF THE PAPER
E. A. ZIMMERMAN (Columbia University, New York): As you well know one of the sources of the vasopressin for memory may be the paraventricular nucleus which projects to many areas. I am thinking about Dr. D. Kahn’s demonstration of neurotensin cells next to the PVN. Could those neurotensin cells be regulating the vasopressin pathway?
TJ. B. VAN WIMERSMA GREIDANUS (University of Utrecht, the Netherlands): The only experiment related to your question we have done is to inject neurotensin ICV and measure at one single time point vasopressin levels in blood. We found a tendency towards an increase, but plasma levels of vasopressin may not really be related to changes in memory.
A. J. DUNN (University of Florida, Gainesville): Regarding extinction of pole jump avoidance, it seems to me that there are a number of peptides that seem to affect it one way or another: ACTH, vasopressin, endorphins and their deriva- tives, and neurotensin. Can you compare the potency of neurotensin with ACTH and vasopressin?
GREIDANUS: After i.c.v. injection neurotensin is 1&25 times less active than, for instance, vasopressin and the endorphins.
DUNN: Is it comparable to ACTH? GREIDANUS: ACTH data showed were based on peripheral injections and gen-
erally we need 100 times less of a peptide after i.c.v. injection than after systemic injection.
D. KAHN (Columbia University, New York): To follow up on Dr. Zimmerman’s comments, one suggestion would be to examine whether neurotensin’s effects are mediated through vasopressin by looking at a vasopressin-deficient rat. Similarly if the NT effect is mediated through ACTH, one might look at an MSG-treated rat.
GREIDANUS: I agree.

van Wimersma Greidanus et al.: Behavioral Effects 329
A. J . PRANCE, JR. (University of North Carolina, Chapel Hill): How would
GREIDANUS: I really do not know. M. D. HIRSCH (Roche Institute of Molecular Biology, Nutley, NJ) : I recall some
data that compared several peptides and TRH had about 50% of the activity of ACTH I-Lo in the pole jumping test.
GREIDANUS: Indeed, there was an effect of TRH, but it was only one single experiment in which the effects of TRH, LHRH and ACTH were compared.
S. FIELDING (Hoechst-Roussel, Somerville, NJ) : Was there an escape compo- nent in the pole jump avoidance response?
GREWANUS: No, because we injected the peptide only during the extinction. An animal has no reason to escape because a shock is never applied during extinction. So, we never injected the peptide prior to the stimulus either in active or in passive avoidance behavior. The peptide was never present during the presentation of electric foot shock, either in active or in passive avoidance behavior.
FIELDING: In evaluating neuroleptics in these kinds of procedures we usu- ally look at a separation between the avoidance and the escape response. If there is wide separation between then it seems to point to a specific response for neuroleptics.
TRH compare with neurotensin in these two models?
Related Documents











