ORIGINAL ARTICLE Bacterial Meningitis Impairs Hippocampal Neurogenesis Sandra Hofer, PhD, Denis Grandgirard, PhD, Denise Burri, MSc, Tanja K. Fro ¨hlich, MSc, and Stephen L. Leib, MD Abstract Bacterial meningitis causes persisting neurofunctional sequelae. The occurrence of apoptotic cell death in the hippocampal subgranular zone of the dentate gyrus characterizes the disease in patients and relates to deficits in learning and memory in corresponding exper- imental models. Here, we investigated why neurogenesis fails to regenerate the damage in the hippocampus associated with the per- sistence of neurofunctional deficits. In an infant rat model of bacterial meningitis, the capacity of hippocampal-derived cells to multiply and form neurospheres was significantly impaired compared to that in uninfected littermates. In an in vitro model of differentiating hippo- campal cells, challenges characteristic of bacterial meningitis (i.e. bacterial components, tumor necrosis factor [20 ng/mL], or growth factor deprivation) caused significantly more apoptosis in stem/ progenitor cells and immature neurons than in mature neurons. These results demonstrate that bacterial meningitis injures hippo- campal stem and progenitor cells, a finding that may explain the persistence of neurofunctional deficits after bacterial meningitis. Key Words: Apoptosis, Bacterial meningitis, Dentate gyrus, Differen- tiation, Hippocampal injury, Neurosphere assay, Stem/progenitor cells. INTRODUCTION Survivors of bacterial meningitis experience permanent neurologic sequelae including impairment of learning and memory in up to 50% of cases (1Y3). Importantly, the dis- abilities persist into adulthood (2, 4Y6). Patients who die of bacterial meningitis show neuronal apoptosis in the sub- granular zone of the hippocampal dentate gyrus, a brain structure involved in learning and memory function (7, 8). Identical histopathologic features are observed in corre- sponding animal models (9Y11). Moreover, the severity of learning dysfunction is associated with the extent of hippo- campal cell (HC) apoptosis in experimental pneumococcal meningitis (9, 12, 13). The dentate gyrus is a site of a stem cell niche with continuous formation of new neurons (14). Neurogenesis oc- curs lifelong in predominantly 2 brain regions: the subven- tricular zone lining the lateral ventricles and the subgranular zone, part of the dentate gyrus of hippocampus (15, 16). In the subgranular zone, the stem cells proliferate and differentiate into progenitor cells. These newly formed cells subsequently migrate into the granule layer of the dentate gyrus, differentiate into mature, functional neurons, and become integrated into the hippocampal network. Therefore, the hippocampus is po- tentially well equipped for repair. However, because neuro- logic sequelae of bacterial meningitis related to hippocampus function persist throughout childhood into adulthood, the ca- pacity for self-repair seems insufficient to compensate for the brain damage (2). We hypothesize that the regenerative capacity of the hippocampus is compromised by bacterial meningitis. In this study, we characterized the cellular targets of apoptosis in the dentate gyrus in organotypic HC cultures and in experi- mental pneumococcal meningitis with the aim of assessing whether bacterial meningitis impairs neurogenesis in the hippocampus. MATERIALS AND METHODS Animal Model of Meningitis All animal studies were approved by the Animal Care and Experimentation Committee of the Canton Bern, Switzerland. They followed the Swiss national guidelines for the performance of animal experiments and were con- ducted in accordance with the US Public Health Service’s Policy on Human Care and Use of Laboratory Animals. Eleven-day-old Wistar rats (Charles River, Germany) were injected intracisternally with 10 KL of saline containing 1 10 6 colony-forming units (cfu)/mL of a clinical isolate of Streptococcus pneumoniae (n = 9) (9, 12, 13, 17). Sham- infected, control littermates (n = 7) were injected with an equal volume of sterile, pyrogen-free saline. Cerebrospinal fluid (CSF) samples were obtained 18 hours later by punc- ture of the cisterna magna to document infection. The num- ber of bacteria in the CSF was determined by plating serial dilutions of 10 KL of CSF on blood agar plates (5.17 T 1.17 10 7 cfu/mL). All animals received ceftriaxone (Rocephine; Roche Pharma, Switzerland 100 mg/kg body weight twice a day subcutaneously). At 48 hours after infection, the rats were killed with 100 mg/kg pentobarbital intraperitoneally, brains were removed, and the hippocampi were dissected and processed for isolation and quantitative assessment of neurosphere-forming cells. J Neuropathol Exp Neurol Volume 70, Number 10, October 2011 890 J Neuropathol Exp Neurol Copyright Ó 2011 by the American Association of Neuropathologists, Inc. Vol. 70, No. 10 October 2011 pp. 890Y899 From the Institute for Infectious Diseases, University of Bern, Bern, Switzerland. Send correspondence and reprint requests to: Stephen L. Leib, MD, Neuro- infection Laboratory, Institute for Infectious Diseases, University of Bern, Friedbu ¨hlstrasse 51, PO Box 61, CH-3010 Bern, Switzerland; E-mail: [email protected] Drs Hofer and Grandgirard contributed equally to this work. The in vitro study was financially supported by a grant from the 3R Research Foundation Switzerland (www.forschung3r.ch) (Project 103-06). Addi- tional support was received from the Swiss National Science Foundation (310030-116257). Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Bacterial Meningitis Impairs Hippocampal Neurogenesis
Sandra Hofer, PhD, Denis Grandgirard, PhD, Denise Burri, MSc, Tanja K. Frohlich, MSc,and Stephen L. Leib, MD
AbstractBacterial meningitis causes persisting neurofunctional sequelae.
The occurrence of apoptotic cell death in the hippocampal subgranularzone of the dentate gyrus characterizes the disease in patients andrelates to deficits in learning and memory in corresponding exper-imental models. Here, we investigated why neurogenesis fails toregenerate the damage in the hippocampus associated with the per-sistence of neurofunctional deficits. In an infant rat model of bacterialmeningitis, the capacity of hippocampal-derived cells to multiply andform neurospheres was significantly impaired compared to that inuninfected littermates. In an in vitro model of differentiating hippo-campal cells, challenges characteristic of bacterial meningitis (i.e.bacterial components, tumor necrosis factor [20 ng/mL], or growthfactor deprivation) caused significantly more apoptosis in stem/progenitor cells and immature neurons than in mature neurons.These results demonstrate that bacterial meningitis injures hippo-campal stem and progenitor cells, a finding that may explain thepersistence of neurofunctional deficits after bacterial meningitis.
Key Words: Apoptosis, Bacterial meningitis, Dentate gyrus, Differen-tiation, Hippocampal injury, Neurosphere assay, Stem/progenitor cells.
INTRODUCTIONSurvivors of bacterial meningitis experience permanent
neurologic sequelae including impairment of learning andmemory in up to 50% of cases (1Y3). Importantly, the dis-abilities persist into adulthood (2, 4Y6). Patients who die ofbacterial meningitis show neuronal apoptosis in the sub-granular zone of the hippocampal dentate gyrus, a brainstructure involved in learning and memory function (7, 8).Identical histopathologic features are observed in corre-sponding animal models (9Y11). Moreover, the severity oflearning dysfunction is associated with the extent of hippo-campal cell (HC) apoptosis in experimental pneumococcalmeningitis (9, 12, 13).
The dentate gyrus is a site of a stem cell niche withcontinuous formation of new neurons (14). Neurogenesis oc-
curs lifelong in predominantly 2 brain regions: the subven-tricular zone lining the lateral ventricles and the subgranularzone, part of the dentate gyrus of hippocampus (15, 16). In thesubgranular zone, the stem cells proliferate and differentiateinto progenitor cells. These newly formed cells subsequentlymigrate into the granule layer of the dentate gyrus, differentiateinto mature, functional neurons, and become integrated intothe hippocampal network. Therefore, the hippocampus is po-tentially well equipped for repair. However, because neuro-logic sequelae of bacterial meningitis related to hippocampusfunction persist throughout childhood into adulthood, the ca-pacity for self-repair seems insufficient to compensate for thebrain damage (2).
We hypothesize that the regenerative capacity of thehippocampus is compromised by bacterial meningitis. In thisstudy, we characterized the cellular targets of apoptosis inthe dentate gyrus in organotypic HC cultures and in experi-mental pneumococcal meningitis with the aim of assessingwhether bacterial meningitis impairs neurogenesis in thehippocampus.
MATERIALS AND METHODS
Animal Model of MeningitisAll animal studies were approved by the Animal
Care and Experimentation Committee of the Canton Bern,Switzerland. They followed the Swiss national guidelinesfor the performance of animal experiments and were con-ducted in accordance with the US Public Health Service’sPolicy on Human Care and Use of Laboratory Animals.Eleven-day-old Wistar rats (Charles River, Germany) wereinjected intracisternally with 10 KL of saline containing 1 �106 colony-forming units (cfu)/mL of a clinical isolate ofStreptococcus pneumoniae (n = 9) (9, 12, 13, 17). Sham-infected, control littermates (n = 7) were injected with anequal volume of sterile, pyrogen-free saline. Cerebrospinalfluid (CSF) samples were obtained 18 hours later by punc-ture of the cisterna magna to document infection. The num-ber of bacteria in the CSF was determined by plating serialdilutions of 10 KL of CSF on blood agar plates (5.17 T 1.17 �107 cfu/mL). All animals received ceftriaxone (Rocephine;Roche Pharma, Switzerland 100 mg/kg body weight twicea day subcutaneously). At 48 hours after infection, the ratswere killed with 100 mg/kg pentobarbital intraperitoneally,brains were removed, and the hippocampi were dissectedand processed for isolation and quantitative assessment ofneurosphere-forming cells.
J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011890
J Neuropathol Exp NeurolCopyright � 2011 by the American Association of Neuropathologists, Inc.
Vol. 70, No. 10October 2011pp. 890Y899
From the Institute for Infectious Diseases, University of Bern, Bern, Switzerland.Send correspondence and reprint requests to: Stephen L. Leib, MD, Neuro-
infection Laboratory, Institute for Infectious Diseases, University of Bern,Friedbuhlstrasse 51, PO Box 61, CH-3010 Bern, Switzerland; E-mail:[email protected]
Drs Hofer and Grandgirard contributed equally to this work.The in vitro study was financially supported by a grant from the 3R Research
Foundation Switzerland (www.forschung3r.ch) (Project 103-06). Addi-tional support was received from the Swiss National Science Foundation(310030-116257).
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

Hippocampal Cell Culture and DifferentiationHippocampal cells of rats from P5 and P11 were dis-
sected and dissociated as previously described (18, 19). Forthe induction of differentiation, single cells (P5) were platedonto poly-L-lysineYcoated coverslips (Becton Dickinson AG,Allschwil, Switzerland) at a density of 5 � 104 cells per co-verslip. Defined growth medium conditions (Dulbecco modifiedEagle medium [DMEM]/F12, 1:1; Gibco Life Technologies,Basel, Switzerland), HEPES (15 mmol/L), 10 Kg/mL penicillin-streptomycin-amphotericin B (Bioswisstec AG, Schaffhausen,Switzerland), B27 supplement (Gibco Life Technologies),20 ng/mL fibroblast growth factor (FGF; PeproTech EC Ltd,London, UK), and 20 ng/mL epidermal growth factor (EGF;PeproTech EC Ltd) were used including 20 ng/mL brain-derived neurotrophic factor (BDNF; PeproTech EC Ltd) (20).Cells were cultured at 37-C with 5% CO2 for 21 days.
Collagen-Based Neurosphere AssayThe assay was performed using a protocol adapted
from the procedure of Ma et al (20). After 24 hours in cul-ture, the HC-derived cell suspension was centrifuged for5 minutes at 600 � g, and the pellet was resuspended in1 mL of ice-cold DMEM/F12. Viable cells were countedusing trypan blue exclusion. Four wells of a 24-well tis-sue culture plate coated with poly-HEMA (Sigma, Buchs,Switzerland) were used per condition.
To assess the vulnerability of stem/progenitor cells todeath triggers, cells were challenged with 3 death factorsat defined time points. Five hundred microliters of a semi-solid collagen growth matrix (3 mg/mL collagen (Sigma),PBS pH 7.4, and growth medium including 4 Kg/mL heparin(Stem Cell Technologies, Grenoble, France) containing 5 �104 cells were plated on each well and incubated for 1 hourat 37-C and 5% CO2 in a humidified atmosphere. Once thegel has solidified, 500 KL of neurosphere growth mediumwas added on the top of each gel. The cultures were keptfor 21 days. Half of the growth medium was replaced by
FIGURE 1. In vitro model of hippocampal cell (HC) differ-entiation. (A) The differentiation stage of HC was identifiedby immunoassaying for Nestin (stem cells), doublecortin (DcX)(immature neurons), and microtubule-associated protein 2(MAP2) (neurons) at 1, 7, 14, and 21 days. Neuronal differ-entiation in vitro was documented by a gradual shift in thepredominant staining pattern from Nestin at 1 day to DcX at7 to 14 days to MAP2 at 21 days. (B) Growth factor depriva-tion (GFD) and tumor necrosis factor (TNF) induce caspase3Ydependent apoptosis. Tumor necrosis factor induced apop-tosis early, at 1 to 14 days of differentiation. Apoptosis inducedby GFD was most prominent at 7 and 14 days (p G 0.05). (C)Bacterial components (BC) induce caspase 3Yindependentapoptosis. Hippocampal cells were exposed to BC for 2 hours.Cell death induced by BC was compared with control cellsusing annexin V or apoptosis-inducing factor (AIF) staining.Apoptosis induced by BC was most abundant at 7 days of dif-ferentiation (p G 0.05). Overall, the vulnerability of cells toundergo death after both challenges peaked at 7 days. p valueswere calculated by 1-way analysis of variance followed byTukey multiple comparison post hoc test. Data are presentedas mean T SEM of 3 or more independent experiments.
J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011 Bacterial Meningitis Injures Neural Stem Cells
� 2011 American Association of Neuropathologists, Inc. 891
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

fresh growth medium twice a week. Quantification of theneurospheres was performed by mounting the gels on glassslides after fixation in 10% paraformaldehyde in PBS. Subse-quently, dried gels were stained with cresyl violet, and numbersof neurospheres per gel were counted using ImageJ 1.37v(Wayne Rasband; National Institutes of Health, Bethesda, MD).
Induction of ApoptosisHippocampal cells (P5) were exposed to death stim-
uli characteristic of bacterial meningitis. These consisted ofapplication of bacterial components (BCs) for 2 hours or20 ng/mL tumor necrosis factor (TNF; Sigma) for 24 hoursto the medium, or growth factor deprivation (GFD), that is,removal of FGF, EGF and BDNF for 24 hours at 1, 7, 14, and21 days after initiation of differentiation. For exposure toBC, a clinical isolate of S. pneumoniae (serotype 3) was cul-tured overnight in a brain heart infusion medium. The bacte-rial suspension was diluted and grown to logarithmic phasefor 4.5 hours. After washing with NaCl, bacterial suspens-ion was resuspended to a defined concentration of 1.7 � 107
cfu/mL in DMEM/F12 containing 10 Kg/mL streptomycinand penicillin to cause bacteriolysis. After 4 hours of incu-bation at 37-C with 5% CO2, the suspension was centrifugedat 3000 � g for 10 minutes. The supernatant containing theBCs was added at a volume of 1:5 to the medium of theHC for 2 hours; a longer exposure of BCs resulted in ne-crosis of the HC. Bacterial componentYinduced cell deathwas observed using phase-contrast microscopy. For appli-cation of TNF, 20 ng/mL TNF was added to the mediumfor 24 hours at 37-C with 5% CO2 (21, 22). For GFD,FGF, EGF, and BDNF (20 ng/mL each) were removed fromthe culture medium. staurosporine (STS; 200 nmol/L for2 hours; Sigma) was used as a positive control for induc-tion of apoptosis. Control cells were handled identically ingrowth medium.
For caspase inhibition, z-VAD-FMK (50 KM; Sigma)was used at 7 days after differentiation. After the challenge,coverslips were fixed and stained as follows: for neuronalmarkers (Nestin, mouse-monoclonal, 1:1000; Becton DickinsonAG), doublecortin (DcX, goat-polyclonal, 1:1000; Santa CruzBiotechnology, Heidelberg, Germany), and microtubule-associated protein 2 (MAP2, mouse-monoclonal, 1:10; hybrid-oma supernatant AP14), and for apoptosis markers (caspase 3,rabbit-polyclonal, 1:400; Cell Signaling Technology,Allschwil, Switzerland) apoptosis-inducing factor (AIF,rabbit-polyclonal, 1:100; ProSci, Inc, Lausen, Switzerland),or used for annexin V (1.8 Kg/mL) and propidium iodide(PI) (5 Kg/mL; Sigma) staining using standard protocols.Stained cells were subsequently analyzed at 40� magnifi-cation and photographed with a Zeiss fluorescence micro-scope (Axiophot or Axiovert 135; Zeiss, West Germany)and analyzed with Openlab 4.0.3 (Improvision, UK) andImageJ 1.37v.
Western BlotsHippocampal cells (P5) were induced to form neuro-
spheres, which were then plated on 60-mm dishes and driveninto differentiation as described above. Western blots wereperformed from control and challenged neurosphere-derived
cells. Cells were harvested and lysed with 30 KL of radio-immunoprecipitation assay buffer. Proteins were separatedin an electrophoresis chamber (Mini Protean II; Bio-Rad,Cressier, France) and transferred to a polyvinyl difluoridemembrane (Millipore AG, Zug, Switzerland). After blockingwith 5% milk, the primary antibodies to the following wereapplied at 4-C overnight for immunoblotting: Bax (rabbitantibody kindly provided by Prof. Ch. Borner, PhD, Albert-Ludwigs-University, Freiburg, Germany, at a dilution of1:5000), Bim (rabbit antibody, 1:1000; Millipore), and Mcl-1(rat antibody, gift from Th. Kaufmann, PhD, Institute of Phar-macology, University of Berne, Switzerland, 1:2000). Blotswere incubated with Immobilon Western ChemiluminescentHPR Substrate (Millipore AG), and bands were visualizedby exposure to Fuji RX photo film (Fujifilm Corporation,Tokyo, Japan).
Statistical AnalysisMultigroup comparisons were performed by 1-way
analysis of variance (ANOVA) followed by Tukey post hoctest or 2-way ANOVA and Student t-test to compare themeans of 2 groups of data using Prism 5.01 (level of sig-nificance set to p G 0.05). For Western blots, the relativeamount of each protein was determined quantitatively byImageJ 1.37v and normalized to the expression of A-actin.Data are presented as mean T SEM, and the numbers ofdifferent experiments are indicated for each.
RESULTS
Neuronal Maturation of HC Stem/ProgenitorCell In Vitro
Hippocampal cells were differentiated by culture onpoly-L-lysineYcoated coverslips for 1, 7, 14, and 21 dayswith 20 ng/mL BDNF. At defined time points, the ratios ofthe different developmental stages (stem/progenitors, im-mature, and mature neurons) relative to the total cell pop-ulation were calculated based on counts of cells positive forNestin, DcX, and MAP2, divided by the number of nucleistained with DAPI. Stem and progenitor cells staining posi-tive for Nestin were the predominant cell population on thefirst day (47% T 12%) after starting the differentiation pro-cess; this percentage decreased as a function of cellulardifferentiation over time to a minimum of 21% T 4% at21 days (Fig. 1A). After 7 days of differentiation, 48% T 14%of the cells were immature neurons, indicated by positivestaining for DcX. At 21 days, the percentage of DcX-positivecells (35% T 4%) decreased and 47% T 3% of the cells werepositive for MAP2, a marker of mature neurons.
Caspase 3YDependent Apoptosis of HC by TNFand GFD
The concentration of TNF (20 ng/mL) used corre-sponded to that found in the CSF of infected animals withacute bacterial meningitis (9). Higher concentrations (i.e.200 or 2,000 ng/mL) led to apoptosis in all cell types (datanot shown). No significant difference in apoptosis wasobserved after challenge for 24 or 48 hours (p 9 0.05).Exposure of TNF induced apoptosis at Day 1, peaked at
Hofer et al J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011
� 2011 American Association of Neuropathologists, Inc.892
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

FIGURE 2. The stage of cellular differentiation (Nestin, doublecortin [DcX], microtubule-associated protein 2 [MAP2]) determinesthe vulnerability to undergo apoptosis (annexin V and apoptosis-inducing factor [AIF]) after challenge with death triggers. (AYC)Bacterial components (BC) induce necrosis (propidium iodide) and apoptosis. Apoptotic cells (Arrowhead 1) showed characteristicmorphology with shrunken and condensed nuclei (A), and the appearance of annexin V/propidium iodide (Arrowhead 2; B) andAIF (Arrowhead 1; C). Healthy cells (Arrowhead 3) do not show any of these characteristics. (DYF) Tumor necrosis factor andgrowth factor deprivation induced apoptosis in cells staining positive for doublecortin (DcX) and Nestin but not for microtubuleassociated protein 2 (MAP2). (GYL) After exposure to BC, AIF colocalized in mature neurons (MAP2; I, L), immature neurons (DcX;H, K), and stem/progenitor cells (Nestin; G, J). Scale bar = 50 Km.
J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011 Bacterial Meningitis Injures Neural Stem Cells
� 2011 American Association of Neuropathologists, Inc. 893
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
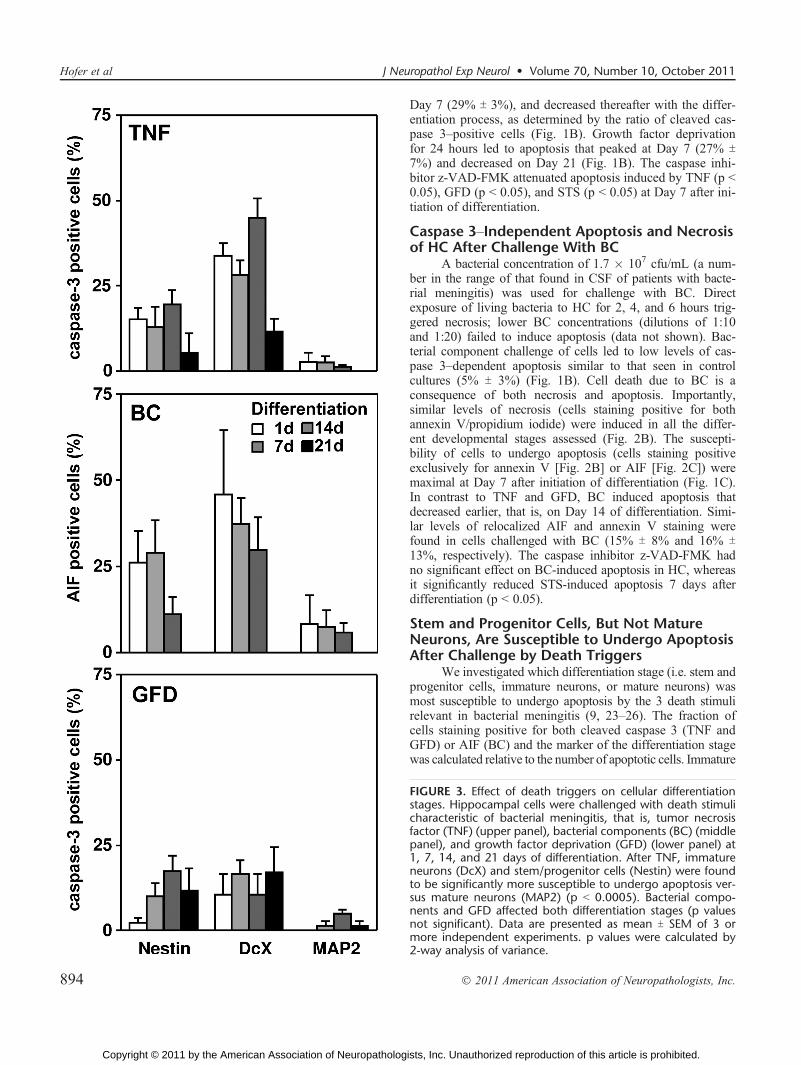
Day 7 (29% T 3%), and decreased thereafter with the differ-entiation process, as determined by the ratio of cleaved cas-pase 3Ypositive cells (Fig. 1B). Growth factor deprivationfor 24 hours led to apoptosis that peaked at Day 7 (27% T7%) and decreased on Day 21 (Fig. 1B). The caspase inhi-bitor z-VAD-FMK attenuated apoptosis induced by TNF (p G0.05), GFD (p G 0.05), and STS (p G 0.05) at Day 7 after ini-tiation of differentiation.
Caspase 3YIndependent Apoptosis and Necrosisof HC After Challenge With BC
A bacterial concentration of 1.7 � 107 cfu/mL (a num-ber in the range of that found in CSF of patients with bacte-rial meningitis) was used for challenge with BC. Directexposure of living bacteria to HC for 2, 4, and 6 hours trig-gered necrosis; lower BC concentrations (dilutions of 1:10and 1:20) failed to induce apoptosis (data not shown). Bac-terial component challenge of cells led to low levels of cas-pase 3Ydependent apoptosis similar to that seen in controlcultures (5% T 3%) (Fig. 1B). Cell death due to BC is aconsequence of both necrosis and apoptosis. Importantly,similar levels of necrosis (cells staining positive for bothannexin V/propidium iodide) were induced in all the differ-ent developmental stages assessed (Fig. 2B). The suscepti-bility of cells to undergo apoptosis (cells staining positiveexclusively for annexin V [Fig. 2B] or AIF [Fig. 2C]) weremaximal at Day 7 after initiation of differentiation (Fig. 1C).In contrast to TNF and GFD, BC induced apoptosis thatdecreased earlier, that is, on Day 14 of differentiation. Simi-lar levels of relocalized AIF and annexin V staining werefound in cells challenged with BC (15% T 8% and 16% T13%, respectively). The caspase inhibitor z-VAD-FMK hadno significant effect on BC-induced apoptosis in HC, whereasit significantly reduced STS-induced apoptosis 7 days afterdifferentiation (p G 0.05).
Stem and Progenitor Cells, But Not MatureNeurons, Are Susceptible to Undergo ApoptosisAfter Challenge by Death Triggers
We investigated which differentiation stage (i.e. stem andprogenitor cells, immature neurons, or mature neurons) wasmost susceptible to undergo apoptosis by the 3 death stimulirelevant in bacterial meningitis (9, 23Y26). The fraction ofcells staining positive for both cleaved caspase 3 (TNF andGFD) or AIF (BC) and the marker of the differentiation stagewas calculated relative to the number of apoptotic cells. Immature
FIGURE 3. Effect of death triggers on cellular differentiationstages. Hippocampal cells were challenged with death stimulicharacteristic of bacterial meningitis, that is, tumor necrosisfactor (TNF) (upper panel), bacterial components (BC) (middlepanel), and growth factor deprivation (GFD) (lower panel) at1, 7, 14, and 21 days of differentiation. After TNF, immatureneurons (DcX) and stem/progenitor cells (Nestin) were foundto be significantly more susceptible to undergo apoptosis ver-sus mature neurons (MAP2) (p G 0.0005). Bacterial compo-nents and GFD affected both differentiation stages (p valuesnot significant). Data are presented as mean T SEM of 3 ormore independent experiments. p values were calculated by2<way analysis of variance.
Hofer et al J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011
� 2011 American Association of Neuropathologists, Inc.894
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

neurons and stem and progenitor cells were found to be mostsusceptible to undergo apoptosis after TNF and GFD, and matureneurons were particularly insensitive (Figs. 2DYF). Bacterialcomponents affected all differentiation stages including mature
neurons (Figs. 2GYL). Two-way ANOVA revealed significantdifferences in vulnerability between the different groups for the3 death triggers. Although both parameters (‘‘stages’’ and ‘‘daysafter differentiation’’) significantly contributed to the observedvariation by TNF challenge, the main determinant explainingthe variation by BC and GFD was ‘‘days after differentiation’’(2-way ANOVA, p G 0.005; Fig. 3).
Death Triggers Impair HC NeurogenesisIn Vitro
The potential of HC-derived cells to multiply andform new colonies in the neurosphere assay was comparedbetween cells challenged in vitro with the 3 death triggers.After GFD (84 T 39, n = 10), no difference in the num-ber of newly formed colonies was observed compared withcells without challenge (84 T 37, n = 10) (p 9 0.05) (Fig. 4A).In contrast, after application of BC (40 T 29, n = 9) andTNF (37 T 19, n = 10), significantly fewer colonies wereobtained versus control cells and cells challenged with GFD(p G 0.005).
Bacterial Meningitis Impairs HC NeurogenesisIn Vivo
To assess whether experimental pneumococcal men-ingitis reduces the number of neurogenic cells in the hippo-campus of infant rats, stem and progenitor cells of individualhippocampi were enumerated using the neurosphere assay.At 3 weeks after plating, significantly fewer colonies devel-oped from HC of animals after pneumococcal meningtis(58 T 20 colonies/35-mm plates, n = 9) versus from HCuninfected control animals (142 T 42 colonies/35-mm plates,n = 7) (p G 0.0001) (Fig. 4B).
Death Triggers Induce Expression ofProapoptotic and Antiapoptotic Proteins
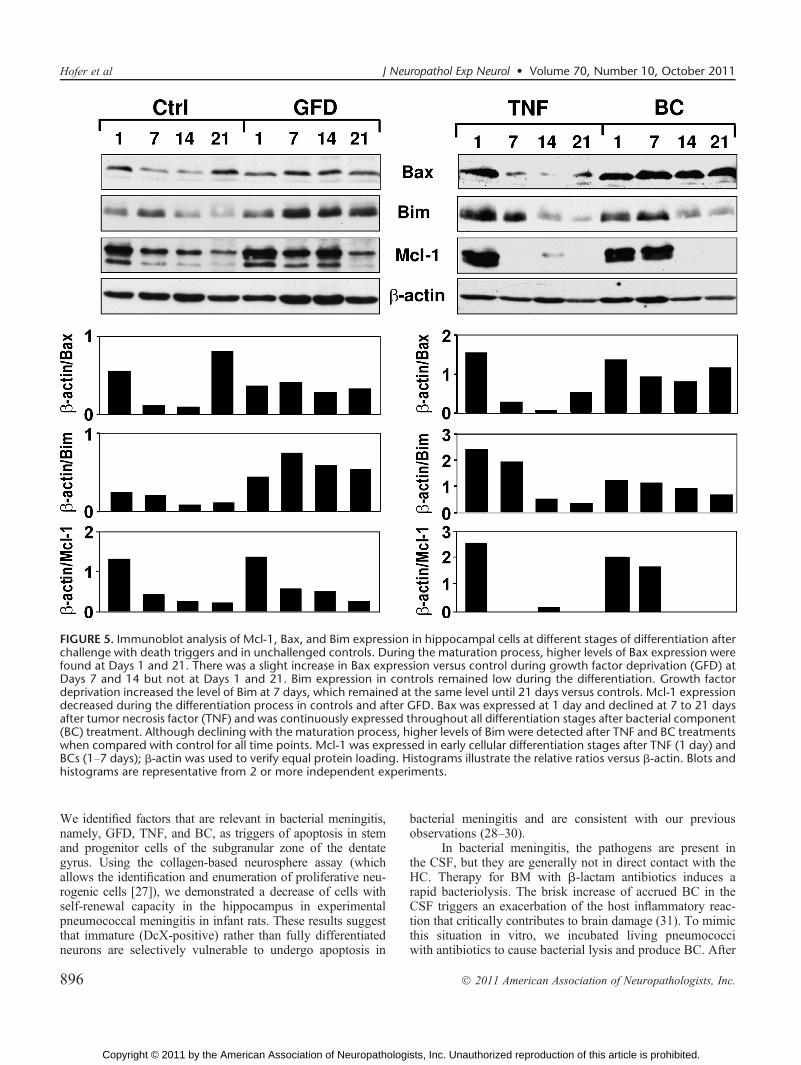
To investigate the molecular mechanisms influencingthe vulnerability of the cells during the apoptotic process, HCextracts were assessed for expression of antiapoptotic andproapoptotic proteins during differentiation by Western blots.During maturation, stronger signals for the proapoptotic pro-tein Bax were found at Days 1 and 21 (Fig. 5). Bax expres-sion was enhanced after application of TNF at 1 day and afterexposure to BC at all days evaluated. A stronger signal forBax expression versus controls was observed during GFD atDays 7 and 14 but not at Days 1 and 21. Levels of the proa-poptotic protein Bim decreased with the maturation processin control cells. Although they also declined with maturation,there were higher levels of Bim after TNF and BC treatmentsversus those in controls at all time points. There were con-tinuously elevated levels of Bim after GFD. In control cells,maximal expression of the antiapoptotic protein Mcl-1 wasobserved at Day 1 with a steady decline until Day 21. For alldeath stimuli used, there was a similar pattern of Mcl-1expression during differentiation.
DISCUSSIONThe results of this study demonstrate that bacterial men-
ingitis injures the neurogenic niche of the HC dentate gyrus.
FIGURE 4. Assessment of the neurogenic potential of hip-pocampal cells (HC) by the collagen-based neurosphere assay.(A) After challenge of HC with bacterial components (BC) andtumor necrosis factor (TNF), fewer colonies were obtainedversus HC from unchallenged controls (p G 0.005). Growthfactor deprivation (GFD) did not affect the yield of proliferat-ing cells (p values not significant). p values were calculatedby one-way analysis of variance followed by Tukey multiplecomparison post hoc test. (B) In an infant rat model of pneu-mococcal meningitis, the colony-forming ability of HC frominfected animals was reduced versus HC cells from mock-infected controls (p G 0.05).
J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011 Bacterial Meningitis Injures Neural Stem Cells
� 2011 American Association of Neuropathologists, Inc. 895
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
We identified factors that are relevant in bacterial meningitis,namely, GFD, TNF, and BC, as triggers of apoptosis in stemand progenitor cells of the subgranular zone of the dentategyrus. Using the collagen-based neurosphere assay (whichallows the identification and enumeration of proliferative neu-rogenic cells [27]), we demonstrated a decrease of cells withself-renewal capacity in the hippocampus in experimentalpneumococcal meningitis in infant rats. These results suggestthat immature (DcX-positive) rather than fully differentiatedneurons are selectively vulnerable to undergo apoptosis in
bacterial meningitis and are consistent with our previousobservations (28Y30).
In bacterial meningitis, the pathogens are present inthe CSF, but they are generally not in direct contact with theHC. Therapy for BM with A-lactam antibiotics induces arapid bacteriolysis. The brisk increase of accrued BC in theCSF triggers an exacerbation of the host inflammatory reac-tion that critically contributes to brain damage (31). To mimicthis situation in vitro, we incubated living pneumococciwith antibiotics to cause bacterial lysis and produce BC. After
FIGURE 5. Immunoblot analysis of Mcl-1, Bax, and Bim expression in hippocampal cells at different stages of differentiation afterchallenge with death triggers and in unchallenged controls. During the maturation process, higher levels of Bax expression werefound at Days 1 and 21. There was a slight increase in Bax expression versus control during growth factor deprivation (GFD) atDays 7 and 14 but not at Days 1 and 21. Bim expression in controls remained low during the differentiation. Growth factordeprivation increased the level of Bim at 7 days, which remained at the same level until 21 days versus controls. Mcl-1 expressiondecreased during the differentiation process in controls and after GFD. Bax was expressed at 1 day and declined at 7 to 21 daysafter tumor necrosis factor (TNF) and was continuously expressed throughout all differentiation stages after bacterial component(BC) treatment. Although declining with the maturation process, higher levels of Bim were detected after TNF and BC treatmentswhen compared with control for all time points. Mcl-1 was expressed in early cellular differentiation stages after TNF (1 day) andBCs (1Y7 days); A-actin was used to verify equal protein loading. Histograms illustrate the relative ratios versus A-actin. Blots andhistograms are representative from 2 or more independent experiments.
Hofer et al J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011
� 2011 American Association of Neuropathologists, Inc.896
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

exposure to BC, HC showed rapid cell death, including cellshrinking and condensation of nuclei, but caspase-3 activationwas not detected and the caspase inhibitor z-VAD-FMK didnot block apoptosis. Bacterial componentYinduced cell death,in this paradigm, is therefore predominantly independent ofcaspase 3 and affects all stages of differentiation includingstem cells, immature neurons, and mature neurons. Theseobservations are in line with our previous in vivo resultsshowing that caspase 3Yindependent injury did not affectimmature neurons selectively but rather clusters of cellsincluding mature neurons relocalizing AIF to their pyknoticcellular nuclei (32). Moreover, in the present in vitro model,the regenerative capacity of HC was reduced by exposure toBC and TNF, suggesting a common mechanism regulating celldeath induced by BC and TNF.
Growth factor deprivation enhanced caspase 3Ydependentapoptosis with a maximum effect at 7 to 14 days of differ-entiation. Growth factors, including neurotrophins such asBDNF, enhance differentiation, induce proliferation, con-trol cell death, and promote the survival of neurons duringdevelopment. Brain-derived neurotrophic factor is of partic-ular interest because its receptor, TrkB, is highly expressedin the hippocampus (33). In a previous study, we demonstratedthat BDNF attenuates caspase 3Ydependent HC apoptosis ininfant rats with bacterial meningitis (24). Downregulation ofnerve growth factor has been demonstrated 30 hours afterinfection in an adult mouse model of pneumococcal menin-gitis (34). Here the increased apoptosis after GFD at 7 to14 days of differentiation coincided with an enhanced cellularexpression of the proapoptotic protein Bim. These findingsare in agreement with studies showing that GFD activates theintrinsic apoptosis pathway leading to increased expressionof proapoptotic proteins such as Bim and Bax that activatecaspase enzymes via the mitochondrial pathway (35, 36).
Tumor necrosis factor exposure induces caspase 3Ydependent apoptosis starting at early stages of differentia-tion, similar to what has been shown in a previous studywhere application of 10 ng/mL of TNF increased apoptosisin adult neural stem cells (37). In an experimental model ofpneumococcal meningitis, inhibition of a TNF-convertingenzyme (TACE, ADAM17) downregulated the CSF concen-tration of TNF, attenuated apoptosis in the dentate gyrus ofthe hippocampus, and preserved spatial learning and mem-ory function (9). Tumor necrosis factor is one of the ex-trinsic death signals that induce apoptosis directly throughactivation of caspase 8 and 3 or via caspase-mediated trans-location of Bid and the release of cytochrome c (38, 39).During differentiation in vitro, the percentage of stem andprogenitor cells decreased as they developed to immatureand subsequently mature neurons. At the protein level, wedemonstrated that on Day 1, when Nestin-positive cells pre-dominate, Mcl-1 was highly expressed and declined steadilythereafter. It has been shown that loss of Mcl-1 in neuronalprogenitor cells results in apoptosis of Nestin-positive cells,whereas in mature neurons, a similar loss of Mcl-1 did notinduce cell death. Mcl-1 has, therefore, been identified as akey regulator of neural precursor cell survival during thetransition from progenitor cells to postmitotic neurons (40).Furthermore, increased neuronal sensitivity to seizures has
been observed in Mcl-1 heterozygous mice (41). Our re-sults suggest that TNF and BC induce apoptosis in neuro-genic cells at an early stage of differentiation, which includestem cells. In contrast, stem and early progenitor cells areprotected from apoptosis under GFD. This finding is consis-tent with results from the neurosphere assay, in which thenumber of colonies obtained after GFD did not differ fromcontrol cultures. The observed lower sensitivity of stem cellstoward GFD-induced apoptosis might be attributed to highexpression levels of Mcl-1. Mcl-1 has been shown to se-quester Bim (42), and its downregulation has been shown tobe required for Bax activation (43).
Tumor necrosis factor and BC induce apoptosis in stemand progenitor cells through a pathway not regulated by Mcl-1.Because Bax knockout animals show increased numbers ofneurons in the CNS, Bax is thought to play a role in neuronalprogrammed cell death (44). Here, Bax expression was upre-gulated by TNF and BC at Day 1 of differentiation, indicatingthat TNF- and BC-induced apoptosis depends on Bax. Simi-lar to Bax, Bim expression was also enhanced by both deathtriggers in early differentiation. This may suggest that Bimcooperates in TNF- and BC-mediated apoptosis in HC stemand progenitor cells. In line with these results, it has recentlybeen shown that Bim is essential for the activation of Baxand Bak-dependent cell death programs (45).
Our present investigations on the role of apoptosisregulators are limited. A number of factors (not yet inves-tigated) could also influence the fate of the cells after deathtriggers in vitro. Apart from the factors explored in the pre-sent study, upregulation of antiapoptotic proteins (e.g. Bcl-XL [46, 47] or XIAP [48]) in mature neurons may explainthe observation that mature neurons were not affected bycaspase 3Ydependent apoptosis. A decline in the expressionof proapoptotic factors (caspases, Apaf-1, Puma) may alsodesensitize the neurons (49). Conversely, noxious stimuliresulting in the elevation of other BH3-only proteins (Bim,Puma) could mediate the vulnerability of HC neurons, assimilarly observed in vivo during proteasome inhibition-mediated apoptosis (50). Finally, posttranslational modifi-cations not investigated in the present study may also accountfor the observed selective vulnerability. In our model ofpediatric pneumococcal meningitis, the number of cells withcolony-forming ability (stem and progenitor cells) decreasedin the infant rats that survived pneumococcal meningi-tis. This finding was validated in an in vitro model of HCdifferentiation. Tumor necrosis factor and GFD induced cas-pase 3Ydependent apoptosis, whereas BC induced caspase3Yindependent death of cells in the neurogenic niche. Ourdata indicate that stem cells and immature neurons are vul-nerable to undergo apoptosis, whereas mature neurons aremore resistant to the death stimuli evaluated. Our resultssuggest that HC injury in bacterial meningitis targets stemand progenitor cells. The impaired neurogenesis in the hip-pocampus may help to understand why neurologic deficitspersist after childhood bacterial meningitis. The pathoge-netic significance of BC, TNF, and GFD in the develop-ment of HC injury demonstrated herein identifies these deathtriggers as therapeutic targets. In consequence, interven-tions that minimize the releases of BCs (31), down-modulate
J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011 Bacterial Meningitis Injures Neural Stem Cells
� 2011 American Association of Neuropathologists, Inc. 897
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

TNF concentration in CSF (9), and increase BDNF (24) rep-resent promising avenues in the search for therapeutic strat-egies to attenuate damage to the neurogenic niche ofthe hippocampus in bacterial meningitis.
ACKNOWLEDGMENTThe authors thank Fransiska Simon, Kevin Oberson,
and Angela Buhlmann for excellent technical support.
REFERENCES1. Merkelbach S, Sittinger H, Schweizer I, et al. Cognitive outcome after
bacterial meningitis. Acta Neurol Scand 2000;102:118Y232. Grimwood K, Anderson P, Anderson V, et al. Twelve year outcomes
following bacterial meningitis: Further evidence for persisting effects.Arch Dis Child 2000;83:111Y16
3. Bedford H, de Louvois J, Halket S, et al. Meningitis in infancy inEngland and Wales: Follow up at age 5 years. BMJ 2001;323:533Y36
4. de Louvois J, Halket S, Harvey D. Neonatal meningitis in England andWales: Sequelae at 5 years of age. Eur J Pediatr 2005;164:730Y34
5. Anderson V, Anderson P, Grimwood K, et al. Cognitive and executivefunction 12 years after childhood bacterial meningitis: Effect of acuteneurologic complications and age of onset. J Pediatr Psychol 2004;29:67Y81
6. Koomen I, Raat H, Jennekens-Schinkel A, et al. Academic and behav-ioral limitations and health-related quality of life in school-age survivorsof bacterial meningitis. Qual Life Res 2005;14:1563Y72
7. Nau R, Soto A, Bruck W. Apoptosis of neurons in the dentate gyrus inhumans suffering from bacterial meningitis. J Neuropathol Exp Neurol1999;58:265Y74
8. Gerber J, Bottcher T, Bering J, et al. Increased neurogenesis after exper-imental Streptococcus pneumoniae meningitis. J Neurosci Res 2003;73:441Y46
9. Leib SL, Clements JM, Lindberg RL, et al. Inhibition of matrix metallo-proteinases and tumour necrosis factor alpha converting enzyme as adju-vant therapy in pneumococcal meningitis. Brain 2001;124:1734Y42
10. Pfister LA, Tureen JH, Shaw S, et al. Endothelin inhibition improvescerebral blood flow and is neuroprotective in pneumococcal meningitis.Ann Neurol 2000;47:329Y35
11. Auer M, Pfister LA, Leppert D, et al. Effects of clinically used anti-oxidants in experimental pneumococcal meningitis. J Infect Dis 2000;182:347Y50
12. Leib SL, Heimgartner C, Bifrare YD, et al. Dexamethasone aggravateshippocampal apoptosis and learning deficiency in pneumococcal menin-gitis in infant rats. Pediatr Res 2003;4:4
13. Loeffler JM, Ringer R, Hablutzel M, et al. The free radical scavengeralpha-phenyl-tert-butyl nitrone aggravates hippocampal apoptosis andlearning deficits in experimental pneumococcal meningitis. J Infect Dis2001;183:247Y52
14. Yagita Y, Kitagawa K, Ohtsuki T, et al. Neurogenesis by progenitor cellsin the ischemic adult rat hippocampus. Stroke 2001;32:1890Y96
15. Cameron HA, Woolley CS, McEwen BS, et al. Differentiation of newlyborn neurons and glia in the dentate gyrus of the adult rat. Neuroscience1993;56:337Y44
16. Eriksson PS, Perfilieva E, Bjork-Eriksson T, et al. Neurogenesis in theadult human hippocampus. Nat Med 1998;4:1313Y17
17. Grandgirard D, Schurch C, Cottagnoud P, et al. Prevention of brain injuryby the nonbacteriolytic antibiotic daptomycin in experimental pneumo-coccal meningitis. Antimicrob Agents Chemother 2007;51:2173Y78
18. Brewer GJ. Isolation and culture of adult rat hippocampal neurons.J Neurosci Methods 1997;71:143Y55
19. Wachs FP, Couillard-Despres S, Engelhardt M, et al. High efficacy ofclonal growth and expansion of adult neural stem cells. Lab Invest 2003;83:949Y62
20. Ma W, Fitzgerald W, Liu QY, et al. CNS stem and progenitor cell dif-ferentiation into functional neuronal circuits in three-dimensional colla-gen gels. Exp Neurol 2004;190:276Y88
21. Bernardino L, Agasse F, Silva B, et al. Tumor necrosis factor-alphamodulates survival, proliferation, and neuronal differentiation in neonatalsubventricular zone cell cultures. Stem Cells 2008;26:2361Y71
22. Wong G, Goldshmit Y, Turnley AM. Interferon-gamma but not TNFalpha promotes neuronal differentiation and neurite outgrowth of murineadult neural stem cells. Exp Neurol 2004;187:171Y77
23. Braun JS, Sublett JE, Freyer D, et al. Pneumococcal pneumolysin andH(2)O(2) mediate brain cell apoptosis during meningitis. J Clin Invest2002;109:19Y27
24. Bifrare YD, Kummer J, Joss P, et al. Brain-derived neurotrophic factorprotects against multiple forms of brain injury in bacterial meningitis.J Infect Dis 2005;191:40Y45
25. Hoffmann O, Mahrhofer C, Rueter N, et al. Pneumococcal cell wall--induced meningitis impairs adult hippocampal neurogenesis. InfectImmun 2007;75:4289Y97
26. Meli DN, Loeffler JM, Baumann P, et al. In pneumococcal meningitis anovel water-soluble inhibitor of matrix metalloproteinases and TNF-alphaconverting enzyme attenuates seizures and injury of the cerebral cortex.J Neuroimmunol 2004;151:6Y11
27. Deleyrolle LP, Reynolds BA. Identifying and enumerating neural stemcells: Application to aging and cancer. Prog Brain Res 2009;175:43Y51
28. Sury MD, Agarinis C, Widmer HR, et al. JNK is activated but doesnot mediate hippocampal neuronal apoptosis in experimental neonatalpneumococcal meningitis. Neurobiol Dis 2008;32:142Y50
29. Grandgirard D, Bifrare YD, Pleasure SJ, et al. Pneumococcal meningitisinduces apoptosis in recently postmitotic immature neurons in the dentategyrus of neonatal rats. Dev Neurosci 2007;29:134Y42
30. Grandgirard D, Steiner O, Tauber MG, et al. An infant mouse model ofbrain damage in pneumococcal meningitis. Acta Neuropathol 2007;114:609Y17
31. Grandgirard D, Oberson K, Buhlmann A, et al. Attenuation of cere-brospinal fluid inflammation by the nonbacteriolytic antibiotic daptomycinversus that by ceftriaxone in experimental pneumococcal meningitis.Antimicrob Agents Chemother 2010;54:1323Y26
32. Bifrare YD, Gianinazzi C, Imboden H, et al. Bacterial meningitis causestwo distinct forms of cellular damage in the hippocampal dentate gyrus ininfant rats. Hippocampus 2003;13:481Y88
33. Masana Y, Wanaka A, Kato H, et al. Localization of trkB mRNA inpostnatal brain development. J Neurosci Res 1993;35:468Y79
34. Tauber SC, Stadelmann C, Spreer A, et al. Increased expression ofBDNF and proliferation of dentate granule cells after bacterial meningitis.J Neuropathol Exp Neurol 2005;64:806Y15
35. Thornberry NA, Lazebnik Y. Caspases: Enemies within. Science 1998;281:1312Y16
36. Letai A. Pharmacological manipulation of Bcl-2 family members tocontrol cell death. J Clin Invest 2005;115:2648Y55
37. Widera D, Mikenberg I, Elvers M, et al. Tumor necrosis factor alphatriggers proliferation of adult neural stem cells via IKK/NF-kappaB sig-naling. BMC Neurosci 2006;7:64
38. Jurgensmeier JM, Xie Z, Deveraux Q, et al. Bax directly induces releaseof cytochrome c from isolated mitochondria. Proc Natl Acad Sci U S A1998;95:4997Y5002
39. Narita M, Shimizu S, Ito T, et al. Bax interacts with the permeabil-ity transition pore to induce permeability transition and cytochromec release in isolated mitochondria. Proc Natl Acad Sci U S A 1998;95:14681Y86
40. Arbour N, Vanderluit JL, Le Grand JN, et al. Mcl-1 is a key regulator ofapoptosis during CNS development and after DNA damage. J Neurosci2008;28:6068Y78
41. Mori M, Burgess DL, Gefrides LA, et al. Expression of apoptosisinhibitor protein Mcl1 linked to neuroprotection in CNS neurons. CellDeath Differ 2004;11:1223Y33
42. Han J, Goldstein LA, Gastman BR, et al. Interrelated roles for Mcl-1 andBIM in regulation of TRAIL-mediated mitochondrial apoptosis. J BiolChem 2006;281:10153Y63
43. Nijhawan D, Fang M, Traer E, et al. Elimination of Mcl-1 is required forthe initiation of apoptosis following ultraviolet irradiation. Genes Dev2003;17:1475Y86
44. White FA, Keller-Peck CR, Knudson CM, et al. Widespread eliminationof naturally occurring neuronal death in Bax-deficient mice. J Neurosci1998;18:1428Y39
Hofer et al J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011
� 2011 American Association of Neuropathologists, Inc.898
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

45. Ren D, Tu HC, Kim H, et al. BID, BIM, and PUMA are essential foractivation of the BAX- and BAK-dependent cell death program. Science2010;330:1390Y93
46. Akhtar RS, Ness JM, Roth KA. Bcl-2 family regulation of neuronaldevelopment and neurodegeneration. Biochim Biophys Acta 2004;1644:189Y203
47. Motoyama N, Wang F, Roth KA, et al. Massive cell death of immaturehematopoietic cells and neurons in Bcl-xYdeficient mice. Science 1995;267:1506Y10
48. Wright KM, Linhoff MW, Potts PR, et al. Decreased apoptosome activitywith neuronal differentiation sets the threshold for strict IAP regulation ofapoptosis. J Cell Biol 2004;167:303Y13
49. Leveille F, Papadia S, Fricker M, et al. Suppression of the intrinsicapoptosis pathway by synaptic activity. J Neurosci 2010;30:2623Y35
50. Tsuchiya T, Bonner HP, Engel T, et al. Bcl-2 homology domain 3-onlyproteins Puma and Bim mediate the vulnerability of CA1 hippo-campal neurons to proteasome inhibition in vivo. Eur J Neurosci2011;33:401Y8
J Neuropathol Exp Neurol � Volume 70, Number 10, October 2011 Bacterial Meningitis Injures Neural Stem Cells
� 2011 American Association of Neuropathologists, Inc. 899
Copyright © 2011 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
Related Documents











