This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Azithromycin inhibits macrophage interleukin-1β production through inhibition ofactivator protein-1 in lipopolysaccharide-induced murine pulmonary neutrophilia☆
Martina Bosnar ⁎,1, Snježana Čužić 1, Berislav Bošnjak 2, Krunoslav Nujić 1, Gabrijela Ergović 1,Nikola Marjanović 1, Ivanka Pašalić 1, Boška Hrvačić 1, Denis Polančec 1,Ines Glojnarić 1, Vesna Eraković Haber 1
GlaxoSmithKline Research Centre Zagreb Limited, Prilaz baruna Filipovića 29, HR-10000 Zagreb, Croatia
a b s t r a c ta r t i c l e i n f o
Article history:Received 15 October 2010Received in revised form 30 November 2010Accepted 14 December 2010Available online 30 December 2010
Keywords:Activator protein-1AzithromycinInterleukin-1βLipopolysaccharide-induced pulmonaryneutrophilia
Macrolide antibiotics, including azithromycin, also possess anti-inflammatory properties. However, themolecular mechanism(s) of activity as well as the target cells for their action have not been unambiguouslyidentified as yet. In this study, the effects of azithromycin on lipopolysaccharide (LPS)-induced pulmonaryneutrophilia were investigated in mice. Using immunohistochemistry, mRNA and specific protein assays, weconfirmed that azithromycin ameliorates LPS-induced pulmonary neutrophilia by inhibiting interleukin-1β(IL-1β) expression and production selectively in alveolar macrophages as well as in LPS-stimulated J774.2macrophage-derived cells in vitro. Inhibition by azithromycin of neutrophilia and IL-1β was accompanied byprevention of nuclear expression of activator protein-1 (AP-1) in both alveolar macrophages and J774.2 cells.The macrolide did not alter nuclear factor kappa B (NF-κB) or extracellular signal-regulated kinase 1/2 (ERK1/2) expression, activation or localization in LPS-stimulated lungs or in J774.2 cells. In conclusion, we haveshown that inhibition of LPS-induced pulmonary neutrophilia and IL-1β concentrations in lung tissuefollowing azithromycin treatment is mediated through effects on alveolar macrophages. In addition, we haveshown for the first time, in an in vivo model, that azithromycin inhibits AP-1 activation in alveolarmacrophages, an action confirmed on J774.2 cells in vitro.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Macrolide antibiotics are natural or semisynthetic polyketides thatconsist of a macrocyclic lactone ring containing 14, 15 or 16 atoms towhich one or more sugars are attached. Erythromycin, a secondarymetabolite isolated from Saccharopolyspora erythraea, was the firstmacrolide to be introduced to clinical use over 50 years ago.Afterwards, several semisynthetic derivatives of erythromycin, likeazithromycin (9-deoxy-9a-aza-9a-methyl-9a-homoerythromycin A),were designed to broaden the antimicrobial spectrum, reducegastrointestinal side effects and increase acid stability and bioavail-ability of this class of antibiotics [1]. Macrolide antibacterials arebroad-spectrum antibiotics that exhibit antibacterial activity againstaerobic Gram-positive bacteria, certain Gram-negative bacteria,
anaerobic bacteria and intracellular pathogens such as Mycoplasma,Chlamydia and Legionella [2]. Nowadays, macrolides are widelyused in the treatment of respiratory and urogenital tract, skin andsoft tissue infections. In addition to their efficacy in the treatment ofbacterial infections, many studies over the last 20 years havedemonstrated that certain macrolides are effective in the treatment ofvarious chronic inflammatory disorders, particularly of the respiratorytract.
The introduction of erythromycin to the treatment of diffusepanbronchiolitis (DPB) in the 1980s drastically increased 10-yearsurvival rate, decreased frequency of exacerbations and restored lungfunction [3]. Subsequently, azithromycin was successfully used in thetreatment of cystic fibrosis (CF), which shares a number of similaritiesin clinical and pathologic characteristics with DPB. In CF patients,azithromycin treatment was shown to significantly improve lungfunction and reduce frequency of exacerbations. Consequently,macrolides, such as erythromycin, clarithromycin and azithromycin,are now first line therapy for DPB and recommended for patients withCF, and they are also being evaluated in the therapy of chronicobstructive lung disease (COPD), chronic sinusitis, asthma, bronchi-ectasis and bronchiolitis obliterans [4–6]. Clinical efficacy of thesemacrolides in diseases such as DPB and CF cannot be directly linked totheir antimicrobial activity [7].
International Immunopharmacology 11 (2011) 424–434
☆ This work was supported by GlaxoSmithKline Research Centre Zagreb Limited.⁎ Corresponding author. Galapagos istraživački centar d.o.o., Prilaz baruna Filipovića
29, HR-10000 Zagreb, Croatia. Tel.: +385 1 8886319; fax: +385 1 8886438.E-mail address: [email protected] (M. Bosnar).
1 Present address: Galapagos istraživački centar d.o.o., Prilaz baruna Filipovića 29,HR-10000 Zagreb, Croatia.
2 Present address: Medical University of Vienna, Department of Dermatology,Experimental Allergy, Lazarettgasse 19/3, A-1090 Wien, Austria.
1567-5769/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.intimp.2010.12.010
Contents lists available at ScienceDirect
International Immunopharmacology
j ourna l homepage: www.e lsev ie r.com/ locate / in t imp

Author's personal copy
A variety of anti-inflammatory effects of erythromycin, clarithro-mycin and azithromycin have been reported in vivo and in vitro,including inhibition of neutrophil activation and recruitment,decrease in cytokine and mucus secretion, inhibition of bacterialquorum sensing and biofilm formation [reviewed in 5,8]. Severalgroups, including ours, have reported that some macrolides effi-ciently inhibit pulmonary neutrophil accumulation and inflamma-tory mediator production induced by lipopolysaccharide (LPS)challenge [9–15]. Moreover, we have suggested that the effects aremediated through inhibition of interleukin-1β (IL-1β) and granulo-cyte-macrophage colony stimulating factor production by macro-phages. In this study, we provide an insight into the cellular andmolecular targets responsible for the in vivo anti-inflammatoryeffects of azithromycin. In order to confirm our hypothesis, IL-1βexpression in lungs was assessed by immunohistochemistry. Inaddition, the influence of azithromycin on activator protein-1 (AP-1),nuclear factor kappa B (NF-κB) and extracellular signal-regulatedkinase 1/2 (ERK1/2), suggested in the literature as possiblemediatorsof its activity [16–19], was also investigated.
2. Materials and methods
2.1. Mice
Studies were performed on 10 week oldmale BALB/cJ mice (CharlesRiver, Lyon, France).Micewere kept onwiremeshfloorswith irradiatedmaize granulate bedding (Scobis Due, Mucedola, Settimo Milanese,Italy) and maintained under standard laboratory conditions (temper-ature 23–24 °C, relative humidity 60±5%, 15 air changes per hour,artificial lightingwith circadian cycle of 12 hours). Pelleted food (VRF-1,Altromin, Charles River, Isaszag, Hungary) and tapwater were providedad libitum. All procedures on animals were approved by the ethicscommittee of GlaxoSmithKline Research Centre Zagreb Limited, andperformed in accordance with the European Economic CommunityCouncil Directive 86/609.
2.2. Materials
LPS from Escherichia coli serotype 0111:B4 was obtained fromSigma Chemical Co. (St Louis, MO). Azithromycin dihydrate was fromPLIVA Inc. (Zagreb, Croatia). Luminex kits and antibodies for enzyme-linked immunosorbentassay (ELISA) were purchased from R&DSystems (Minneapolis, MN). Anti-IL-1β, anti-p-NF-κB (p105/p50) andanti-ERK1/2 antibodies were purchased from Santa Cruz BiotechnologyInc. (Santa Cruz, CA). Anti-AP-1 (c-Jun) was from Sigma ChemicalCo. Anti-p-Jun, anti-ERK and anti-p-ERK antibodies for western blotanalysis were obtained from Cell Signaling Technology (Danvers, MA).All other reagents, if not indicated otherwise,were fromSigmaChemicalCo. (St Louis, MO).
2.3. LPS-induced pulmonary neutrophilia
Experimental pulmonary neutrophilia was induced as describedearlier [10]. Briefly, mice, under light anesthesia, were instilledintranasally with 2 μg LPS from E. coli/60 μl phosphate-bufferedsaline (PBS). Vehicle or azithromycin was administered intraperito-neally (i.p.) 2 h before intranasal challenge with LPS at a dose of200 mg/kg (b.w.). We have previously demonstrated that 600 mg/kgp.o. is an effective dose for inhibition of inflammation in this animalmodel [9]. Based on an oral bioavailability of 35–55% (internal data),the estimated effective dose of azithromycin after i.p. administrationwas calculated to be 200–300 mg/kg. The dosing regimen wasconfirmed as the lowest effective dose according to the results ofpreliminary studies. Azithromycin was first dissolved in dimethly-sulphoxide (DMSO) and then diluted with 0.5% (w/v) methyl-cellulose [final concentration of DMSO was 5% (v/v)]. The solution
obtained was injected in a volume of 20 ml/kg (b.w.). Six animalswere used in each treatment group.
2.4. Bronchoalveolar lavage and determination of total and relative cellnumber in bronchoalveolar lavage fluid (BALF)
Immediately prior (0 h) or at various timepoints after LPS application,the animals were euthanized by an i.p. overdose of Thiopental® (InresaArzneimittel GmbH, Freiburg, Germany). After preparation and cannula-tion of tracheas, bronchoalveolar lavage was performed with coldcalcium- and magnesium-free PBS in a total volume of 1 ml (0.4, 0.3and 0.3 ml). Following bronchoalveolar lavage, lungs were excised andfixed in 10% buffered neutral formalin fixative.
The bronchoalveolar lavage samples were centrifuged (4 °C, 100 g,5 min), the pellet of cells was resuspended in an equal volume of freshPBS containing 2% fetal bovine serum (FBS) and 0.02% ethylenediamine tetra-acetic acid and used for total and differential cellcounts. Total number of cells in BALF was counted with ahematological analyzer (Sysmex SF 3000, Sysmex Co., Kobe, Japan).Differential cell counts were determined by flow cytometry. BALFsamples containing 106 cells/100 μl were stained with CD45-FITC, GR-1-PE (both from Becton Dickinson Pharmingen, Franklin Lakes, NJ)and F4/80 (Caltag Laboratories, Buckingham, UK) antibodies. Afterstaining, samples were fixed with FACS Lysing solution (BectonDickinson Pharmingen). Flow cytometric analysis was performed on aFACSscan (Becton Dickinson Biosciencecs). Forward scatter, sidescatter, and FL1, FL2 and FL3 parameters were measured and 10,000instances of CD45+ events collected. FCS (flow cytometry standards)list mode files were further analyzed using Summit software (DAKO,Glostrup, Denmark).
2.5. Preparation of lung homogenates for determination of cytokines
In separate groups of mice, non-lavaged lungs were collected fordetermination of IL-1β. Lungs were homogenized on ice in PBS withprotease inhibitors (1 μg/ml leupeptin, 2 μg/ml aprotinin, 1 μg/mlpepstatin, and 17 μg/ml phenylmethylsulphonyl fluoride); 4 ml of PBSwith protease inhibitors was added per gram of lung tissue. Homo-genates were centrifuged (4 °C, 2500 g, 15 min) and stored at −80 °Cuntil analysis.
2.6. Measurement of IL-1β in lungs
IL-1β concentration was determined using Fluorokine MAPmultiplex kit (R&D Systems, Minneapolis, MN) according to themanufacturer's protocol. Briefly, 50 μl of samples was incubated withantibody-coated microparticles for 3 h at room temperature. After-wards, washed beads were incubated with biotinylated detectionantibody cocktail for 1 h at room temperature, washed and incubatedfor 30 min with streptavidin–phycoerythrin conjugate. After the finalwash, the microparticles were resuspended in buffer and analyzedwith the Luminex 200TM (Luminex, Austin, TX) and STarStationsoftware v2.3 (Applied Cytometry Systems, Sacramento, CA) using afive-parameter-logistic-curve fitting. The concentration of IL-1β inlung homogenates was further normalized to protein concentration inthe samples and expressed as pg of analyte per mg of protein.
2.7. Immunohistochemistry for IL-1β, AP-1, NF-κB and ERK1/2
Immunohistochemical detection of IL-1β, AP-1, NF-κB and ERK1/2was done on paraffin-embedded tissue sections from all the animalsincluded in the experiment (six per treatment group) using anti-IL-1β(1:25), anti-AP-1 (c-Jun) (1:50), anti-p-NF-κB (p105/p50) (1:500) andanti-ERK1/2 (1:25) antibodies. Antibody binding was detected bysecondary biotinylated antibodies, streptavidin–horseradishperoxidaseconjugate and diaminobenzidine chromogen (HRP-DAB System,
425M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434

Author's personal copy
R&D Systems, Minneapolis, MN) according to the manufacturer'srecommendations. Staining localization and intensity was examinedbyanobserver blinded to theexperimental design andgradedaccordingto the following criteria:
0. No staining1. Weak staining2. Moderate staining3. Strong staining
In borderline cases, an intermediate grade was used (0–1, 1–2,and 2–3), extending the scoring to a total of seven grades.
In addition, the optical densities of all macrophages present onthree random fields were determined using ImageQuant 5.2 software(Molecular Dynamics). The results were presented as optical densitieswith subtracted background values and average value of unchal-lenged animals.
2.8. Cells
The murine monocyte/macrophage cell line, J774.2, was obtainedfrom the European Collection of Cell Cultures (Porton Down,Wiltshire, UK). Cells were grown in Dulbecco's modified Eagle'smedium (DMEM) (Invitrogen, Carlsbad, CA) supplemented with 10%heat-inactivated FBS (BioWest, Nuaillé, France).
2.9. Cell stimulation and inhibition
For all experiments in vitro, azithromycin was dissolved in DMSOat a concentration of 50 mM and further diluted 1000-fold in cellculture medium.
For IL-1β mRNA determination, J774.2 cells were seeded in 24-well plates in DMEM with 10% FBS at a concentration of 3×105 cellsper well. The next day, cells were pre-incubatedwith azithromycin for2 h and stimulated with 1 μg/ml LPS from E. coli serotype 0111:B4(optimal concentration of LPS for stimulation was established inpreliminary experiments, data not shown). At the end of theincubation period, cells were lysed by addition of RLT buffer (Qiagen,Valencia, CA) and stored at −80 °C until assayed.
For intracellular IL-1β, AP-1, NF-κB or ERK1/2 determination, cellswere seeded in 10 cm plates in DMEMwith 10% FBS at a concentrationof 5×106 cells per plate. The next day, cells were pre-incubated withazithromycin for 2 h and stimulated with 1 μg/ml LPS from E. coliserotype 0111:B4. At the end of the incubation period, for westernblot analysis, cells were scraped in lysis buffer (50 mM HEPES,150 mM sodium chloride, 1% Triton X-100, 1 μg/ml leupeptin, 2 μg/mlaprotinin, 1 μg/ml pepstatin, 17 μg/ml phenylmethylsulphonyl fluo-ride, 0.2 mM sodium orthovanadate, 2 mM sodium fluoride, 4 mM β-glycerophosphate, 0.4 mM sodium pyrophosphate, pH 7.4). Forintracellular IL-1β determination and DNA-binding ELISA, cytosolicand nuclear extracts were prepared with Nuclear Extract Kit (ActiveMotif, Carlsbad, CA) according to manufacturer's protocol. Whole celllysates, cytosolic and nuclear extracts were stored at −80 °C untilassayed. All experiments were repeated at least twice with all thesamples taken in triplicate.
2.10. Total RNA extraction, reverse transcription and quantitative real-time PCR
Total RNA was isolated from cells using RNeasy Mini kit (Qiagen)according to the manufacturer's protocol. 1.5 μg RNA was reversetranscribed using SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogen). Quantitative real-time PCR was performed on ABIPRISM 7300 Sequence Detector (Applied Biosystems, Carlsbad, CA) in a30 μl reaction volume containing TaqMan Universal PCR Master Mix(Applied Biosystems), 100 nM primers and fluorescent TaqMan probesand 5 μl of appropriately diluted cDNA template. A non-template control
was included to evaluate DNA contamination. The relative geneexpression (RFR) was calculated by the comparative Ctmethod (RFR=2−ΔΔCT), using GAPDH as the housekeeping gene. Theprimers and the probeswere as follows (F=forward primer, R=reverseprimer, P=probe, all 5′–3′): GAPDH F: TGTGTCCGTCGTGGATCTGA,GAPDH R: CCTGCTTCACCACCTTCTTGA, GAPDH P: CCGCCTGGA-GAAACCTGCCAAGTATG, IL-1β F: AATCTATACCTGTCCTGTGTAATGAAA-GAC , I L - 1β R : TGGGTATTGCTTGGGATCCA , I L - 1β P :CACACCCACCCTGCAGCTGGAGA.
2.11. ELISA
IL-1β concentration in the cytosolic fraction of LPS-stimulatedJ774.2 cells was determined by sandwich ELISA using capture anddetection antibodies according to the manufacturer's instructions.Optical density was measured at 450 nm by use of a microplatereader. Concentration was determined by interpolation from standardcurves with SoftMax Pro v4.3.1 software (Molecular Devices,Sunnyvale, CA).
2.12. DNA-binding ELISA
Activation of AP-1 (p-cJun) and NF-κB (p65) was determined byTransAM kits (Active Motif) according to the manufacturer's protocol.In brief, 5 μg of nuclear extract was added per well of plates coatedwith double-stranded oligonucleotides containing TPA-responsiveelement and NF-κB consensus site, which bind AP-1 and NF-κBdimers, respectively. Following 1 h incubation, plates were washedand samples were incubated for 1 h with antibodies specific for thebound, active form of the transcription factor, c-Jun phosphorylated at
Fig. 1. Azithromycin suppressed LPS-induced pulmonary inflammation. Monocyte/macrophage (CD45High+Gr-1low+/− F4-80+, closed bars) and neutrophil (CD45High+Gr-1High+ F4-80−, open bars) numbers in BALF determined by flow cytometry (A) and IL-1βconcentration in lung homogenates (B) 24 h after challenge with PBS (PBS) or 2 μg of LPS(VEH+LPS). Azithromycin (AZM) was administered intraperitoneally 2 h prior to LPSchallenge at a dose of 200 mg/kg. Data are presented as means±S.E.M. (n=6). Theasterisk (⁎) indicates significant difference (Pb0.05) from LPS-challenged vehicle-treatedanimals, MannWhitney test.
426 M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434
Author's personal copy
Ser73 and p65. At the end of incubation, plates were washed andincubated with horseradish peroxidase-conjugated antibodies. Afterthe final wash, the developing solution was added and absorbancewas read at 450 nm.
2.13. Western blot analysis
Equal amounts of proteins (60 μg), as determined by theBCA method, were separated by SDS-PAGE (NuPAGE 10% Bis–Trisgel, Invitrogen) and transferred to nitrocellulose membrane. Mem-branes were blocked for 1 h in 2% non-fat milk in PBS with 0.02%Tween-20, followed by a 2 h incubation with primary antibodies(ERK1/2, 1:1000 and p-ERK1/2, 1:500, Cell Signaling) in blockingbuffer. Secondary antibodies labeled with horseradish peroxidase(Cell Signaling) were incubated for 1 h in blocking buffer. Detectionwas done with chemiluminescent detection (ECL, GE Healthcare,
Uppsala, Sweden) using film (Lumi film chemifluorescent detectionfilm, Roche, Switzerland). The films were scanned with Storm™/FluorImager (Molecular Dynamics). The optical densities of scannedbands were determined using ImageQuant 5.2 software. The resultswith subtracted background values were expressed as ratios of thedensitometry value of each treatment group to that of negativecontrol (untreated samples) which was always set to equal to 1.
2.14. Determination of protein concentration
Protein concentration in lung homogenates and cell lysates forwestern blot analysis was determined by BCA Protein Assay (ThermoFisher Scientific Inc.) according to the manufacturer's recommenda-tion. Protein concentration in nuclear extracts for DNA-binding ELISAwas determined by 2-D Quant kit (GE Healthcare) according to themanufacturer's protocol.
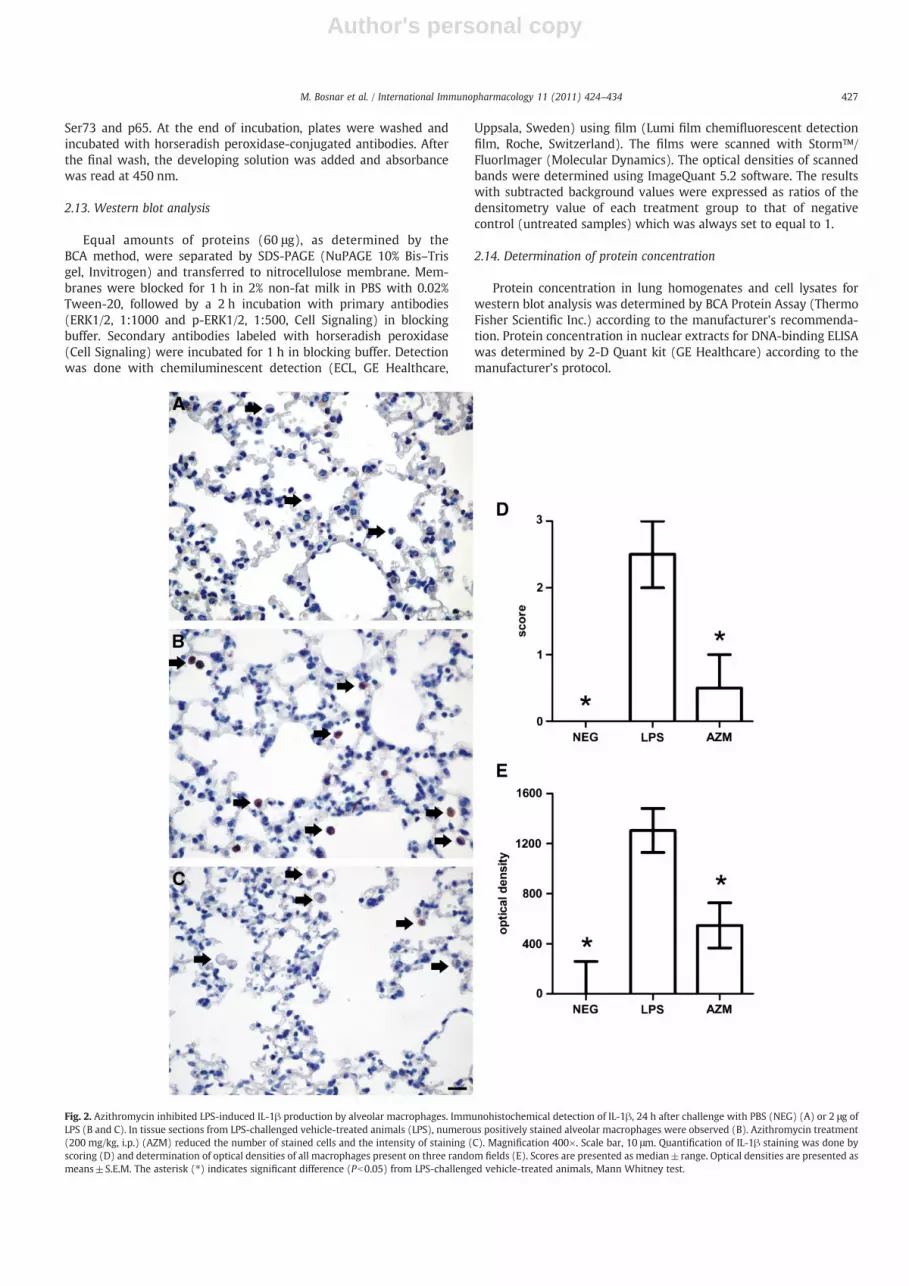
Fig. 2. Azithromycin inhibited LPS-induced IL-1β production by alveolar macrophages. Immunohistochemical detection of IL-1β, 24 h after challenge with PBS (NEG) (A) or 2 μg ofLPS (B and C). In tissue sections from LPS-challenged vehicle-treated animals (LPS), numerous positively stained alveolar macrophages were observed (B). Azithromycin treatment(200 mg/kg, i.p.) (AZM) reduced the number of stained cells and the intensity of staining (C). Magnification 400×. Scale bar, 10 μm. Quantification of IL-1β staining was done byscoring (D) and determination of optical densities of all macrophages present on three random fields (E). Scores are presented as median±range. Optical densities are presented asmeans±S.E.M. The asterisk (⁎) indicates significant difference (Pb0.05) from LPS-challenged vehicle-treated animals, Mann Whitney test.
427M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434
Author's personal copy
2.15. Statistical analysis
All values are presented as means±S.E.M. To define statisticallysignificant differences between vehicle-treated and macrolide-treatedmice after LPS challenge, as well as between vehicle and macrolide-exposed cells in vitro, the data were subjected to Mann Whitney testusing GraphPad Prism version 5.00 for Windows (GraphPad Software,San Diego, CA). The level of significance was set at Pb0.05 in all cases.
3. Results
3.1. Effect of azithromycin on neutrophil accumulation in BALF and IL-1βconcentration in lung homogenates
In order to validate that azithromycin is equally potent insuppressing LPS-induced lung inflammation after intraperitoneal as
it is after oral administration, the effect of i.p. treatment on neutrophilaccumulation in BALF and IL-1β concentration in lung homogenateswas investigated first. In accordance with previously reported results[9], azithromycin treatment (200 mg/kg, i.p.) significantly decreasedneutrophil numbers in BALF (83%) (Fig. 1A) and IL-1β concentrationsin lung homogenates (58%) (Fig. 1B) at 24 h after LPS challenge. Veryfew neutrophils were detected in BALF from healthy animals and IL-1β concentration was below limit of detection (≈3 pg/ml).
3.2. Effect of azithromycin on IL-1β expression in lungs
Based on results from studies in vitro on murine monocyte/macrophage cell line J774.2 and murine epithelial cells MLE-12we have previously suggested that inhibition of IL-1β concentrationsin lung homogenates is caused by influence of azithromycinon alveolar macrophages and not epithelial cells [9]. Therefore,
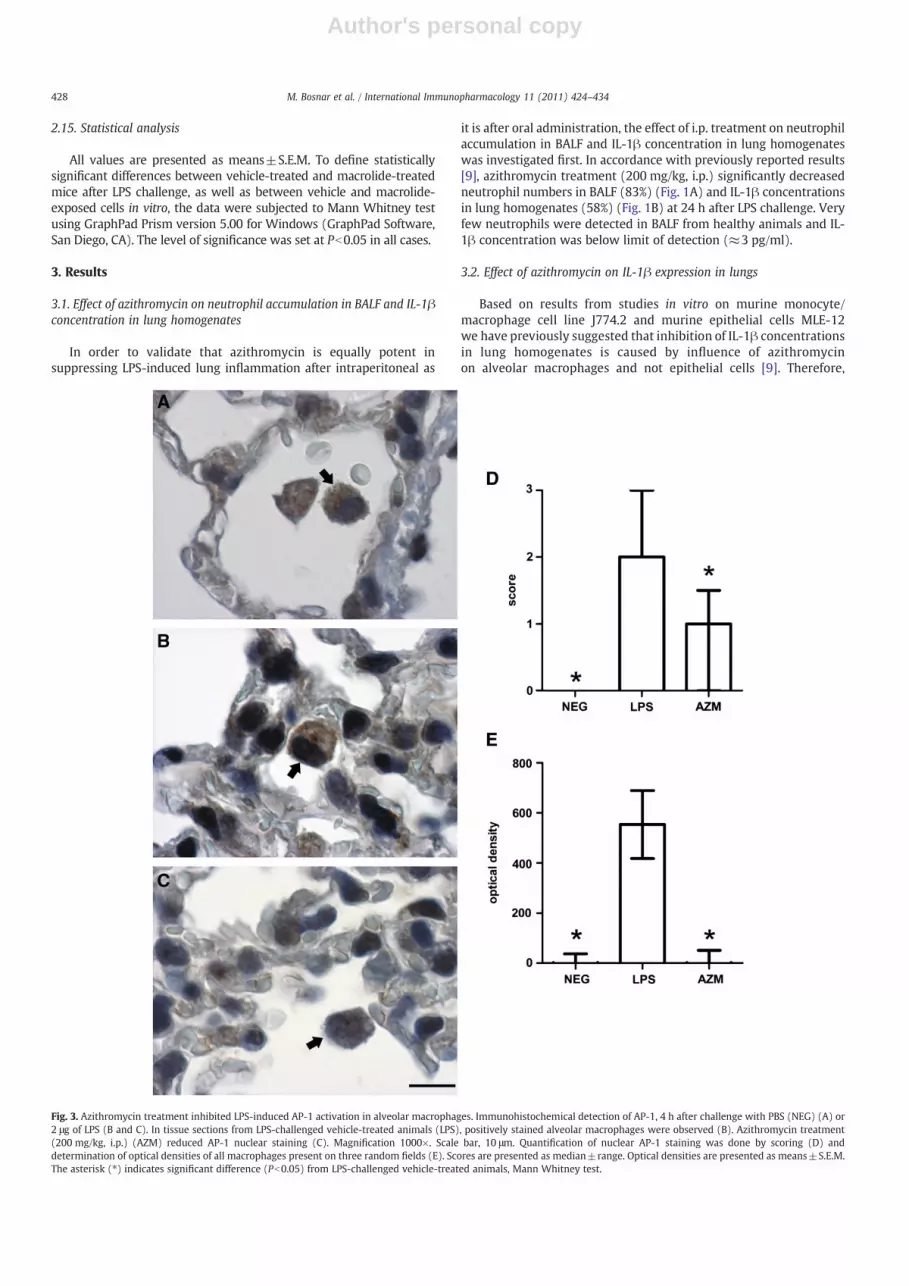
Fig. 3. Azithromycin treatment inhibited LPS-induced AP-1 activation in alveolar macrophages. Immunohistochemical detection of AP-1, 4 h after challenge with PBS (NEG) (A) or2 μg of LPS (B and C). In tissue sections from LPS-challenged vehicle-treated animals (LPS), positively stained alveolar macrophages were observed (B). Azithromycin treatment(200 mg/kg, i.p.) (AZM) reduced AP-1 nuclear staining (C). Magnification 1000×. Scale bar, 10 μm. Quantification of nuclear AP-1 staining was done by scoring (D) anddetermination of optical densities of all macrophages present on three random fields (E). Scores are presented as median±range. Optical densities are presented as means±S.E.M.The asterisk (⁎) indicates significant difference (Pb0.05) from LPS-challenged vehicle-treated animals, Mann Whitney test.
428 M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434

Author's personal copy
immunohistochemistry was used to identify which cells are themajor producers of IL-1β in lungs and confirm the inhibitory activityof azithromycin. Following LPS challenge, only alveolar macrophagesbecame IL-1β positive (Fig. 2B). The intensity of IL-1β staining wasreduced in alveolar macrophages from azithromycin-treated ani-mals (Fig. 2C). The weak expression of IL-1β detected in bronchialepithelial cells of PBS-challenged animals did not change followingLPS administration or azithromycin treatment (not shown).
3.3. Effect of azithromycin on AP-1, NF-κB and ERK1/2 expression andlocalization in alveolar macrophages
To investigate the signaling pathways throughwhich azithromycininhibits IL-1β synthesis in alveolar macrophages, we evaluated AP-1,NF-κB and ERK1/2 expression and localization in the lungs at earlytime points after LPS challenge. In healthy animals, AP-1 was localized
to the cytoplasm of alveolar macrophages (Fig. 3A). Already 30 minafter LPS challenge nuclei of alveolar macrophages became strongly denovo AP-1 positive and remained stained during the next 4 h (Fig. 3B).Azithromycin treatment reduced AP-1 nuclear staining (Fig. 3C).Similar staining pattern was observed in type II alveolar epithelialcells as well.
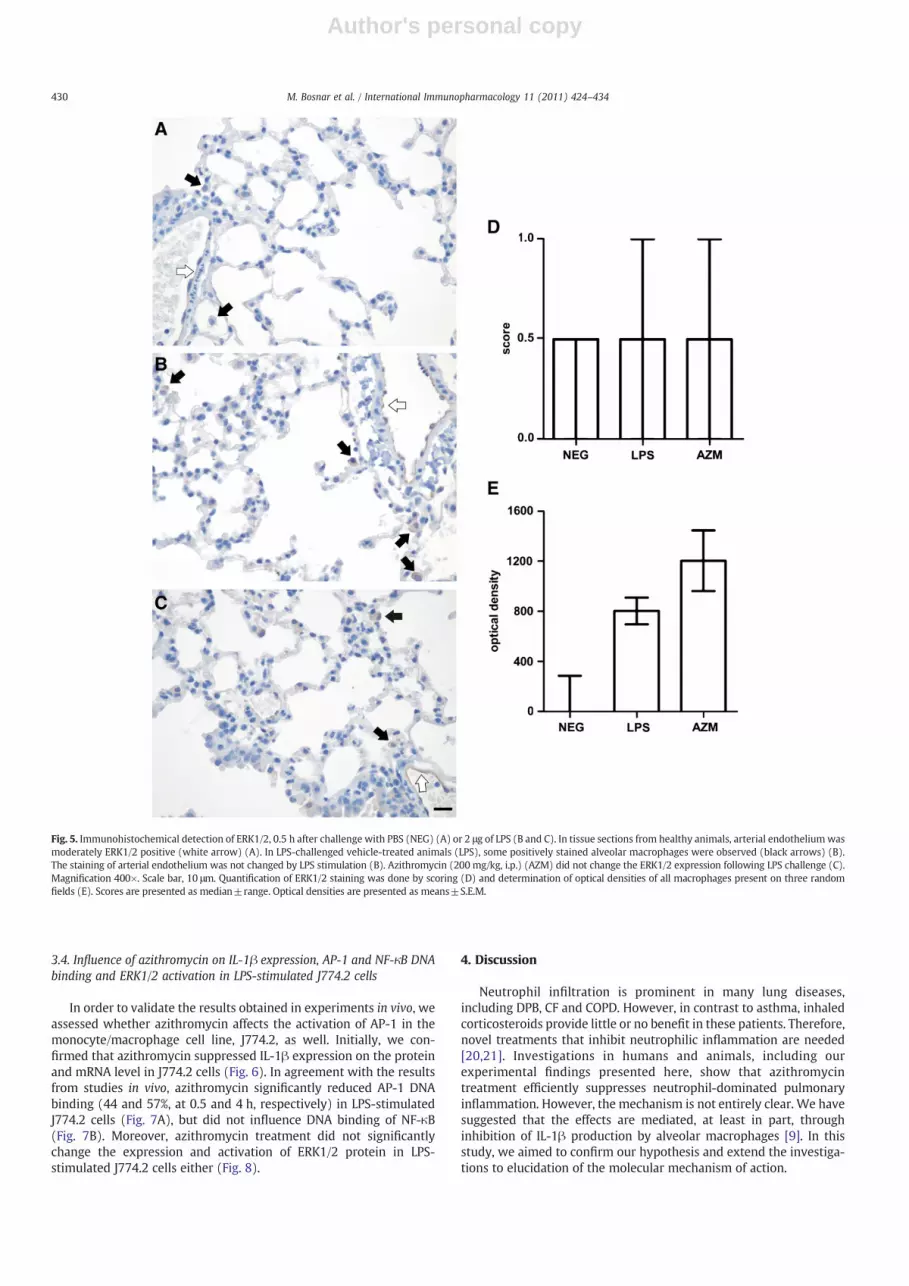
Following LPS challenge, the other two signaling moleculesstudied also changed their expression and/or localization in alveolarmacrophages. NF-κB translocated from cytoplasm to the nuclei ofalveolar macrophages (Fig. 4B), while some alveolar macrophagesalso became positive for ERK1/2 staining (Fig. 5B). However, incontrast to AP-1, azithromycin treatment did not change NF-κB orERK1/2 expression, activation or localization in LPS-challengedanimals (Figs. 4C and 5C). The described changes were observed intissue sections from all animals included in the experiment (six pertreatment group).
Fig. 4. Immunohistochemical detection of NF-κB, 0.5 h after challenge with PBS (NEG) (A) or 2 μg of LPS (B and C). NF-κB translocated from cytoplasm to nuclei of alveolar macrophagesafter i.n. LPS challenge (LPS) (B). Azithromycin (200 mg/kg, i.p.) (AZM) administration did not change the NF-κB localization following LPS challenge (C). Magnification 400×. Scale bar,10 μm.Quantification of nuclear NF-κB stainingwasdoneby scoring (D) and determination of optical densities of allmacrophages present on three random fields (E). Scores are presentedasmedian±range.Optical densities are presentedasmeans±S.E.M. Theasterisk (⁎) indicates significantdifference (Pb0.05) fromLPS-challengedvehicle-treatedanimals,MannWhitneytest.
429M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434
Author's personal copy
3.4. Influence of azithromycin on IL-1β expression, AP-1 and NF-κB DNAbinding and ERK1/2 activation in LPS-stimulated J774.2 cells
In order to validate the results obtained in experiments in vivo, weassessed whether azithromycin affects the activation of AP-1 in themonocyte/macrophage cell line, J774.2, as well. Initially, we con-firmed that azithromycin suppressed IL-1β expression on the proteinand mRNA level in J774.2 cells (Fig. 6). In agreement with the resultsfrom studies in vivo, azithromycin significantly reduced AP-1 DNAbinding (44 and 57%, at 0.5 and 4 h, respectively) in LPS-stimulatedJ774.2 cells (Fig. 7A), but did not influence DNA binding of NF-κB(Fig. 7B). Moreover, azithromycin treatment did not significantlychange the expression and activation of ERK1/2 protein in LPS-stimulated J774.2 cells either (Fig. 8).
4. Discussion
Neutrophil infiltration is prominent in many lung diseases,including DPB, CF and COPD. However, in contrast to asthma, inhaledcorticosteroids provide little or no benefit in these patients. Therefore,novel treatments that inhibit neutrophilic inflammation are needed[20,21]. Investigations in humans and animals, including ourexperimental findings presented here, show that azithromycintreatment efficiently suppresses neutrophil-dominated pulmonaryinflammation. However, the mechanism is not entirely clear. We havesuggested that the effects are mediated, at least in part, throughinhibition of IL-1β production by alveolar macrophages [9]. In thisstudy, we aimed to confirm our hypothesis and extend the investiga-tions to elucidation of the molecular mechanism of action.
Fig. 5. Immunohistochemical detection of ERK1/2, 0.5 h after challenge with PBS (NEG) (A) or 2 μg of LPS (B and C). In tissue sections from healthy animals, arterial endotheliumwasmoderately ERK1/2 positive (white arrow) (A). In LPS-challenged vehicle-treated animals (LPS), some positively stained alveolar macrophages were observed (black arrows) (B).The staining of arterial endothelium was not changed by LPS stimulation (B). Azithromycin (200 mg/kg, i.p.) (AZM) did not change the ERK1/2 expression following LPS challenge (C).Magnification 400×. Scale bar, 10 μm. Quantification of ERK1/2 staining was done by scoring (D) and determination of optical densities of all macrophages present on three randomfields (E). Scores are presented as median±range. Optical densities are presented as means±S.E.M.
430 M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434

Author's personal copy
We have confirmed that azithromycin treatment reduced LPS-induced neutrophil numbers in BALF and IL-1β concentrations in lungtissue. IL-1β is one of the major cytokines involved in the initiationand persistence of inflammation [22] and increased levels of IL-1βhave been reported in induced sputum and BALF from patients withneutrophil-dominated inflammatory disorders of the lungs, such asCOPD, CF and DPB [23–25]. The important role of IL-1β in pulmonaryinflammation has been confirmed in several animal studies. Intra-tracheal administration of IL-1β is sufficient to induce substantial lungneutrophil accumulation [26]. In addition, inducible transgenic miceoverexpressing IL-1β in the lungs developed pulmonary inflammationcharacterized by neutrophil and macrophage infiltration, distalairspace enlargement, and disruption of elastin fibers in alveolarsepta and fibrosis of airway walls and pleura [27]. Consequently,treatment with anti-IL-1β antibodies significantly decreased alveolarmacrophage recruitment in cigarette-smoke exposed mice [28] andanalogous decrease of neutrophils, unfortunately not measured inthat particular research, would have been expected. Similarly, IL-1receptor knockout mice were protected against cigarette-smokeinduced accumulation of inflammatory cells, including neutrophils,in lungs and matrix breakdown [29]. Importance of macrophage-derived IL-1β for neutrophil recruitment following LPS challenge hasbeen reported in other animal models as well. For instance, depletionof mammary alveolar macrophages reduces neutrophil infiltrationand IL-1β concentrations in LPS-induced murine mastitis. In addition,IL-1 receptor gene knockout mice have abrogated neutrophilrecruitment into alveolar and duct spaces of mammary glands [30].Therefore, inhibition of IL-1β production by azithromycin seems to besufficient to reduce neutrophil accumulation in the lungs followingLPS challenge.
We have shown clearly, using immunohistochemistry, thatalveolar macrophages are the major producers of IL-1β followingintranasal LPS administration and thus the target cells of azithromycinanti-inflammatory actions. Accordingly, IL-1β was also inhibited byazithromycin treatment of the murine monocyte/macrophage cellline, J774.2. Inhibition of IL-1β by certain macrolides has beenreported in the clinic as well. IL-1β was the only significantlydecreased pro-inflammatory cytokine in BALF after 12 weeks ofazithromycin treatment of patients with COPD [31]. Erythromycintreatment reduced IL-1β in BALF of DPB patients to levels comparableto those of healthy controls [32,33].
The promoter region in the IL-1β gene contains binding motifs forNF-κB andAP-1 [34,35] aswell as at least three canonical AP-1 enhancerelements [36], which appear to be important in LPS-mediated IL-1βinduction. Furthermore, the activity of AP-1 appears, at least in part, tobe regulated by ERK1/2 signaling molecules [37,38]. We have shownthat following LPS stimulation azithromycin inhibited AP-1 activationand translocation into the nucleus of alveolar macrophages andmacrophage cell line, while having no effects on NF-κB and ERK1/2activation. Since the promoter region in the IL-1β gene contains bindingmotifs for AP-1 [35,36] inhibition of its activation will result in reducedIL-1β production. Having inmind that several studies have identified IL-1β as one of the major cytokines driving neutrophil accumulation inlung tissue [26–29,39,40], inhibition of AP-1 activation and consequentIL-1β production by alveolar macrophages seems to be a sufficient andlikely mechanism by which azithromycin reduced neutrophil accumu-lation in the lungs following LPS challenge (concept summarized inFig. 9).
The effects of clarithromycin, and azithromycin on AP-1, NF-κB andERK1/2 in monocytes/macrophages have so far been investigated only
Fig. 6. Inhibition of IL-1β by azithromycin in LPS-stimulated J774.2 cells. IL-1β protein (A and B) and mRNA (C and D) levels are shown in unstimulated (NEG, dashed bars) and LPS-stimulated J774.2 cells (filled bars). Cells were pre-incubated with 50 μM azithromycin (AZM) (open bars) for 2 h followed by 0.5 h (A and C) or 4 h (B and D) LPS stimulation. Dataare presented as means±S.E.M. from at least two different experiments, all samples being taken in triplicate. The asterisk (⁎) indicates significant difference (Pb0.05) from LPS-stimulated group, Mann Whitney test.
431M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434

Author's personal copy
in vitro. Our results obtained both in vitro and in vivo are in accordancewith previous in vitrofindings showingmacrolide-mediated inhibition ofAP-1 activation and DNA binding in a murine macrophage cell line andhuman monocytes, following LPS+interferon-γ and LPS stimulation,respectively [18,41]. In addition, in line with our data, azithromycin didnot influence ERK1/2 activation in the LPS-stimulated humanmonocyticcell line, THP-1, either [42]. In monocytes and the monocytic cell lines,THP-1 andU-937, clarithromycin inhibited LPS and TNFα-inducedNF-κBactivation [17,18]. Inhibition of NF-κB activation following TNFαstimulation has also been reported in azithromycin-treated trachealaspirate cells from premature infants developing bronchopulmonarydysplasia, as well as, in lung epithelial cell line A549 [43,44]. However, inour study, activation of this transcription factor was not affected eitherin vitro or in vivo. The discrepancymight result fromdifferences in type ofstimulation, species or cell lines used. Moreover, Araki et al. [45]demonstrated in bronchial epithelial cells that azithromycin inhibitedHaemophilus influenzae-induced AP-1, but not NF-κB activation. Azithro-mycin failed to inhibit NF-κB activation following stimulation withChlamydophila penumoniae antigen as well [46]. In addition, conflictingresults on the influence of azithromycin on NF-κB activity were reportedin cystic fibrosis cell lines [16,47,48].
The dose exerting anti-inflammatory effects in this animal model(200 mg/kg) is higher than those exerting therapeutic effects in humans(ranging from 250 mg every other day to 500 mg every day). However,to achieve therapeutic effects in chronic inflammatory diseases,erythromycin, clarithromycin and azithromycin, have to be adminis-
tered for long periods of several months or even years [4]. This mayexplain why, in our acute animal model of inflammation, a single highdose is needed. In animal models of acute inflammation, efficacy can beachieved with lower doses only if administered repeatedly. Roxithro-mycin (2.5 mg/kg) significantly decreased IL-1β and TNFα concentra-tion in lung homogenates of LPS-challenged animals after a 7 weektreatment. Treatment for 5 weeks was not effective [49]. LPS-inducedneutrophil accumulation in lungs of Sprague–Dawley rats was signif-icantly reduced only after 1 week of erythromycin (30 mg/kg) or 4 daysroxithromycin (5 and 10 mg/kg) pretreatment [13,14].
In summary, we have shown that inhibition of LPS-inducedpulmonary neutrophilia and IL-1β concentrations in lung tissuefollowing azithromycin treatment is mediated through effects onalveolar macrophages. In addition, it was shown for the first time, inan in vivo model, that azithromycin inhibits AP-1 activation in alveolarmacrophages, whichmight be the mechanism of observed inhibition ofIL-1β production and consequent effect on neutrophil accumulation. All
Fig. 7. AP-1 (p-cJun) (A) and NF-κB (p65) (B) DNA binding in unstimulated (NEG,dashed bars) and LPS-stimulated J774.2 cells (filled bars). Cells were pre-incubatedwith 50 μM azithromycin (AZM) (open bars) for 2 h followed by 0.5 h or 4 h LPSstimulation. Data are presented as means±S.E.M. from two different experiments, allthe samples being taken in triplicate. The asterisk (⁎) indicates significant difference(Pb0.05) from LPS-stimulated group, Mann Whitney test.
Fig. 8. ERK1/2 activation and expression in unstimulated (NEG, dashed bars) and LPS-stimulated J774.2 cells. Cells were pre-incubated with 50 μM azithromycin (AZM) for2 h followed by 0.5 h or 4 h LPS stimulation. Western blot analysis (A). The imagesshow typical bands appearing at the expected molecular weight. Results of onerepresentative experiment of three are shown. Ratios of the densitometry value of eachtreatment group to that of unstimulated cells (NEG) for p-ERK1/2 (B) and ERK1/2 (C).Data are presented as means±S.E.M. The asterisk (⁎) indicates significant difference(Pb0.05) from LPS-stimulated group, Mann Whitney test.
432 M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434

Author's personal copy
the results obtained in vivo were confirmed in vitro on the murinemonocyte/macrophage cell line, J774.2.
Acknowledgments
We wish to thank Michael J. Parnham for critical reading of themanuscript and Irineja Ćubela, Milka Horvatinčić, Renata Povrženić,Slavica Skender, Marija Škalic, Ksenija Štajcer, and Vedran Vrban forexcellent technical assistance.
References
[1] Whitman MS, Tunkel AR. Azithromycin and clarithromycin: overview andcomparison with erythromycin. Infect Control Hosp Epidemiol 1992;13:357–68.
[2] Zuckerman JM, Qamar F, Bono BR. Macrolides, ketolides, and glycylcyclines:azithromycin, clarithromycin, telithromycin, tigecycline. Infect Dis Clin North Am2009;23:997–1026.
[3] Kudoh S, Azuma A, Yamamoto M, Izumi T, Ando M. Improvement of survival inpatients with diffuse panbronchiolitis treated with low-dose erythromycin. Am JRespir Crit Care Med 1998;157:1829–32.
[4] Crosbie PA, Woodhead MA. Long-termmacrolide therapy in chronic inflammatoryairway diseases. Eur Respir J 2009;33:171–81.
[5] Culic O, Erakovic V, Parnham MJ. Anti-inflammatory effects of macrolideantibiotics. Eur J Pharmacol 2001;429:209–29.
[6] Giamarellos-Bourboulis EJ. Macrolides beyond the conventional antimicrobials: aclass of potent immunomodulators. Int J Antimicrob Agents 2008;31:12–20.
[7] Murphy DM, Forrest IA, Curran D, Ward C. Macrolide antibiotics and the airway:antibiotic or non-antibiotic effects? Expert Opin Investig Drugs 2010;19:401–14.
[8] Shinkai M, Henke MO, Rubin BK. Macrolide antibiotics as immunomodulatorymedications: proposedmechanisms of action. Pharmacol Ther 2008;117:393–405.
[9] Bosnar M, Bosnjak B, Cuzic S, Hrvacic B, Marjanovic N, Glojnaric I, et al.Azithromycin and clarithromycin inhibit lipopolysaccharide-induced murinepulmonary neutrophilia mainly through effects on macrophage-derived granulo-cyte-macrophage colony-stimulating factor and interleukin-1beta. J PharmacolExp Ther 2009;331:104–13.
[10] Ivetic TV, Bosnjak B, Hrvacic B, Bosnar M, Marjanovic N, Ferencic Z, et al. Anti-inflammatory activity of azithromycin attenuates the effects of lipopolysaccharideadministration in mice. Eur J Pharmacol 2006;539:131–8.
[11] Kadota J, Sakito O, Kohno S, Sawa H, Mukae H, Oda H, et al. A mechanism oferythromycin treatment in patients with diffuse panbronchiolitis. Am Rev RespirDis 1993;147:153–9.
[12] Leiva M, Ruiz-Bravo A, Jimenez-Valera M. Effects of telithromycin in in vitro and invivo models of lipopolysaccharide-induced airway inflammation. Chest 2008;134:20–9.
[13] Ou XM, Feng YL, Wen FQ, Wang K, Yang J, Deng ZP, et al. Macrolides attenuatemucus hypersecretion in rat airways through inactivation of NF-kappaB.Respirology 2008;13:63–72.
[14] Sanz MJ, Nabah YN, Cerda-Nicolas M, O'Connor JE, Issekutz AC, Cortijo J, et al.Erythromycin exerts in vivo anti-inflammatory activity downregulating celladhesion molecule expression. Br J Pharmacol 2005;144:190–201.
[15] Tamaoki J, Tagaya E, Yamawaki I, Sakai N, Nagai A, Konno K. Effect of erythromycinon endotoxin-induced microvascular leakage in the rat trachea and lungs. Am JRespir Crit Care Med 1995;151:1582–8.
[16] Cigana C, Nicolis E, Pasetto M, Assael BM, Melotti P. Anti-inflammatory effects ofazithromycin in cystic fibrosis airway epithelial cells. Biochem Biophys ResCommun 2006;350:977–82.
[17] Ichiyama T, Nishikawa M, Yoshitomi T, Hasegawa S, Matsubara T, Hayashi T, et al.Clarithromycin inhibits NF-kappaB activation in human peripheral bloodmononuclear cells and pulmonary epithelial cells. Antimicrob Agents Chemother2001;45:44–7.
[18] Kikuchi T, Hagiwara K, Honda Y, Gomi K, Kobayashi T, Takahashi H, et al.Clarithromycin suppresses lipopolysaccharide-induced interleukin-8 production byhumanmonocytes through AP-1 and NF-kappa B transcription factors. J AntimicrobChemother 2002;49:745–55.
[19] Shinkai M, Foster GH, Rubin BK. Macrolide antibiotics modulate ERK phosphor-ylation and IL-8 and GM-CSF production by human bronchial epithelial cells. Am JPhysiol Lung Cell Mol Physiol 2006;290:L75–85.
[20] Barnes PJ, Stockley RA. COPD: current therapeutic interventions and futureapproaches. Eur Respir J 2005;25:1084–106.
[21] Fitzgerald MF, Fox JC. Emerging trends in the therapy of COPD: novel anti-inflammatory agents in clinical development. Drug Discov Today 2007;12:479–86.
[22] Dinarello CA. Biologic basis for interleukin-1 in disease. Blood 1996;87:2095–147.[23] Carroll TP, Greene CM, Taggart CC, McElvaney NG, O'Neill OJ. Interleukin-1,
neutrophil elastase, and lipopolysaccharide: key pro-inflammatory stimuliregulating inflammation in cystic fibrosis. Curr Respir Med Rev 2005;1:43–67.
[24] Chung KF. Cytokines in chronic obstructive pulmonary disease. Eur Respir J Suppl2001;34:50s–9s.
[25] Ekberg-Jansson A, Andersson B, Bake B, Boijsen M, Enanden I, Rosengren A, et al.Neutrophil-associated activation markers in healthy smokers relates to a fall in DL(CO) and to emphysematous changes on high resolution CT. Respir Med 2001;95:363–73.
[26] Ulich TR, Watson LR, Yin SM, Guo KZ, Wang P, Thang H, et al. The intratrachealadministration of endotoxin and cytokines. I. Characterization of LPS-induced IL-1and TNF mRNA expression and the LPS-, IL-1-, and TNF-induced inflammatoryinfiltrate. Am J Pathol 1991;138:1485–96.
[27] Lappalainen U, Whitsett JA, Wert SE, Tichelaar JW, Bry K. Interleukin-1beta causespulmonary inflammation, emphysema, and airway remodeling in the adultmurine lung. Am J Respir Cell Mol Biol 2005;32:311–8.
[28] Castro P, Legora-Machado A, Cardilo-Reis L, Valenca S, Porto LC, Walker C, et al.Inhibition of interleukin-1beta reduces mouse lung inflammation induced byexposure to cigarette smoke. Eur J Pharmacol 2004;498:279–86.
[29] Churg A, Zhou S, Wang X, Wang R, Wright JL. The role of interleukin-1beta inmurine cigarette smoke-induced emphysema and small airway remodeling. Am JRespir Cell Mol Biol 2009;40:482–90.
[30] Elazar S, Gonen E, Livneh-Kol A, Rosenshine I, Shpigel NY. Neutrophil recruitmentin endotoxin-induced murine mastitis is strictly dependent on mammary alveolarmacrophages. Vet Res 2010;41:10.
[31] Hodge S, Hodge G, Jersmann H, Matthews G, Ahern J, Holmes M, et al.Azithromycin improves macrophage phagocytic function and expression ofmannose receptor in chronic obstructive pulmonary disease. Am J Respir CritCare Med 2008;178:139–48.
[32] Kadota J, Matsubara Y, Ishimatsu Y, Ashida M, Abe K, Shirai R, et al. Significance ofIL-1beta and IL-1 receptor antagonist (IL-1Ra) in bronchoalveolar lavage fluid(BALF) in patients with diffuse panbronchiolitis (DPB). Clin Exp Immunol 1996;103:461–6.
[33] Sakito O, Kadota J, Kohno S, Abe K, Shirai R, Hara K. Interleukin 1 beta, tumornecrosis factor alpha, and interleukin 8 in bronchoalveolar lavage fluid of patientswith diffuse panbronchiolitis: a potential mechanism of macrolide therapy.Respiration 1996;63:42–8.
[34] Cogswell JP, Godlevski MM, Wisely GB, Clay WC, Leesnitzer LM, Ways JP, et al. NF-kappa B regulates IL-1 beta transcription through a consensus NF-kappa B bindingsite and a nonconsensus CRE-like site. J Immunol 1994;153:712–23.
[35] Serkkola E, Hurme M. Synergism between protein-kinase C and cAMP-dependentpathways in the expression of the interleukin-1 beta gene is mediated via theactivator-protein-1 (AP-1) enhancer activity. Eur J Biochem 1993;213:243–9.
[36] Shirakawa F, Saito K, Bonagura CA, Galson DL, Fenton MJ, Webb AC, et al. Thehuman prointerleukin 1 beta gene requires DNA sequences both proximal anddistal to the transcription start site for tissue-specific induction. Mol Cell Biol1993;13:1332–44.
[37] Iles KE, Forman HJ. Macrophage signaling and respiratory burst. Immunol Res2002;26:95–105.
[38] Karin M. The regulation of AP-1 activity by mitogen-activated protein kinases.J Biol Chem 1995;270:16483–6.
Fig. 9. Azithromycin inhibits LPS-induced pulmonary neutrophilia through effects onmacrophage IL-1β and AP-1. Following LPS activation azithromycin inhibited AP-1activation and translocation into the nucleus of alveolar macrophages, while having noeffects on NF-κB and ERK1/2 activation. Since the promoter region in the IL-1β genecontains binding motifs for AP-1, inhibition of its activation will result in reduced IL-1βproduction. IL-1β is one of the major cytokines driving neutrophil accumulation in lungtissue. Therefore, by inhibiting its production azithromycin reduced neutrophilaccumulation in the lungs.
433M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434

Author's personal copy
[39] Saperstein S, Huyck H, Kimball E, Johnston C, Finkelstein J, Pryhuber G. The effectsof interleukin-1beta in tumor necrosis factor-alpha-induced acute pulmonaryinflammation in mice. Mediat Inflamm 2009;2009:958658.
[40] Murugan V, Peck MJ. Signal transduction pathways linking the activation ofalveolar macrophages with the recruitment of neutrophils to lungs in chronicobstructive pulmonary disease. Exp Lung Res 2009;35:439–85.
[41] Yamauchi K, Shibata Y, Kimura T, Abe S, Inoue S, Osaka D, et al. Azithromycinsuppresses interleukin-12p40 expression in lipopolysaccharide and interferon-gamma stimulated macrophages. Int J Biol Sci 2009;5:667–78.
[42] Ikegaya S, Inai K, Iwasaki H, Naiki H, Ueda T. Azithromycin reduces tumor necrosisfactor-alpha production in lipopolysaccharide-stimulated THP-1 monocytic cellsby modification of stress response and p38 MAPK pathway. J Chemother 2009;21:396–402.
[43] Cheung PS, Si EC, Hosseini K. Anti-inflammatory activity of azithromycin asmeasured by its NF-kappaB, inhibitory activity. Ocul Immunol Inflamm 2010;18:32–7.
[44] Aghai ZH, Kode A, Saslow JG, Nakhla T, Farhath S, Stahl GE, et al. Azithromycinsuppresses activation of nuclear factor-kappa B and synthesis of pro-inflammatorycytokines in tracheal aspirate cells from premature infants. Pediatr Res 2007;62:483–8.
[45] Araki N, Yanagihara K, Morinaga Y, Yamada K, Nakamura S, Yamada Y, et al.Azithromycin inhibits nontypeable Haemophilus influenzae-induced MUC5ACexpression and secretion via inhibition of activator protein-1 in human airwayepithelial cells. Eur J Pharmacol 2010;644:209–14.
[46] Morinaga Y, Yanagihara K, Miyashita N, Seki M, Izumikawa K, Kakeya H, et al.Azithromycin, clarithromycin and telithromycin inhibit MUC5AC induction byChlamydophila pneumoniae in airway epithelial cells. Pulm Pharmacol Ther 2009;22:580–6.
[47] Blau H, Klein K, Shalit I, Halperin D, Fabian I. Moxifloxacin but not ciprofloxacin orazithromycin selectively inhibits IL-8, IL-6, ERK1/2, JNK, and NF-kappaB activationin a cystic fibrosis epithelial cell line. Am J Physiol Lung Cell Mol Physiol 2007;292:L343–52.
[48] Cigana C, Assael BM, Melotti P. Azithromycin selectively reduces tumor necrosisfactor alpha levels in cystic fibrosis airway epithelial cells. Antimicrob AgentsChemother 2007;51:975–81.
[49] Suzaki H, Asano K, Ohki S, Kanai K, Mizutani T, Hisamitsu T. Suppressive activity ofa macrolide antibiotic, roxithromycin, on pro-inflammatory cytokine productionin vitro and in vivo. Mediat Inflamm 1999;8:199–204.
434 M. Bosnar et al. / International Immunopharmacology 11 (2011) 424–434
Related Documents











