Awareness of the saccade goal in oculomotor selection: Your eyes go before you know Wieske van Zoest * , Mieke Donk Department of Cognitive Psychology, Vrije Universiteit Amsterdam, The Netherlands article info Article history: Received 10 December 2009 Available online 24 April 2010 Keywords: Saccadic eye movements Awareness Saccade goal Saliency Oculomotor capture abstract The aim of the present study was to investigate how saccadic selection relates to people’s awareness of the saliency and identity of a saccade goal. Observers were instructed to make an eye movement to either the most salient line segment (Experiment 1) or the only right- tilted element (Experiment 2) in a visual search display. The display was masked contin- gent on the first eye movement and after each trial observers indicated whether or not they had correctly selected the target. Whereas people’s awareness concerning the saliency of the saccade goal was generally low, their awareness concerning the identity was high. Observers’ awareness of the saccade goal was not related to saccadic performance. Whereas saccadic selection consistently varied as a function of saccade latency, people’s awareness concerning the saliency or identity of the saccade goal did not. The results sug- gest that saccadic selection is primarily driven by subconscious processes. Ó 2010 Elsevier Inc. All rights reserved. 1. Introduction Saccadic movements or saccades are rapid changes in eye position that occur three or four times each second. The pur- pose of saccades is to bring information to the central area of the retina called the fovea where spatial acuity is best. During the more or less stationary periods between saccades, called fixations, information is acquired for processing and the landing point of the next saccade may be determined. Whereas it is generally acknowledged that saccadic programming, for the most part, operates below the level of conscious awareness (Findlay & Gilchrist, 2003; Haggard, 2005; Mokler & Fischer, 1999; Westheimer & Mitchell, 1969), the role of awareness in the saccadic selection of information is less determined. The aim of the present study was to investigate the extent to which observers’ awareness concerning a saccade goal is related to sacc- adic selection performance. It is possible to make a distinction between at least three different views with respect to this issue. First, it may be the case that observers are at all times aware of the information that is overtly selected for closer inspection. In other words, observers are aware of the property of the saccade goal that is selected. For example, if an observer selects a saccade goal on the basis of its color, the observer will be aware of this color; and likewise for other properties such as orientation, stim- ulus-saliency and location. The idea here is that objects or locations selected for foveal viewing reach observer’s awareness via the attentional system. This proposal is based on the idea that attention is tightly related to the saccadic system (Belo- polsky, Kramer, & Theeuwes, 2008; Deubel & Schneider, 1996; Klein, 1980; Klein & Pontefract, 1994; Kowler, Anderson, Dosher, & Blaser, 1995; Rizzolatti, Riggio, Dascola, & Umilta, 1987; Sheliga, Riggio, Craighero, & Rizzolatti, 1995). Accordingly, many theories assume that a shift in covert attention precedes the eye movement to a location (Deubel & Schneider, 1996; 1053-8100/$ - see front matter Ó 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.concog.2010.04.001 * Corresponding author. Address: Department of Psychology, Vrije Universiteit Amsterdam, Van der Boechorststraat 1, 1081 BT Amsterdam, The Netherlands. E-mail address: [email protected] (W. van Zoest). Consciousness and Cognition 19 (2010) 861–871 Contents lists available at ScienceDirect Consciousness and Cognition journal homepage: www.elsevier.com/locate/concog

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Consciousness and Cognition 19 (2010) 861–871
Contents lists available at ScienceDirect
Consciousness and Cognition
journal homepage: www.elsevier .com/locate /concog
Awareness of the saccade goal in oculomotor selection: Your eyes gobefore you know
Wieske van Zoest *, Mieke DonkDepartment of Cognitive Psychology, Vrije Universiteit Amsterdam, The Netherlands
a r t i c l e i n f o
Article history:Received 10 December 2009Available online 24 April 2010
Keywords:Saccadic eye movementsAwarenessSaccade goalSaliencyOculomotor capture
1053-8100/$ - see front matter � 2010 Elsevier Incdoi:10.1016/j.concog.2010.04.001
* Corresponding author. Address: Department oNetherlands.
E-mail address: [email protected] (W. van Z
a b s t r a c t
The aim of the present study was to investigate how saccadic selection relates to people’sawareness of the saliency and identity of a saccade goal. Observers were instructed to makean eye movement to either the most salient line segment (Experiment 1) or the only right-tilted element (Experiment 2) in a visual search display. The display was masked contin-gent on the first eye movement and after each trial observers indicated whether or not theyhad correctly selected the target. Whereas people’s awareness concerning the saliency ofthe saccade goal was generally low, their awareness concerning the identity was high.Observers’ awareness of the saccade goal was not related to saccadic performance.Whereas saccadic selection consistently varied as a function of saccade latency, people’sawareness concerning the saliency or identity of the saccade goal did not. The results sug-gest that saccadic selection is primarily driven by subconscious processes.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Saccadic movements or saccades are rapid changes in eye position that occur three or four times each second. The pur-pose of saccades is to bring information to the central area of the retina called the fovea where spatial acuity is best. Duringthe more or less stationary periods between saccades, called fixations, information is acquired for processing and the landingpoint of the next saccade may be determined. Whereas it is generally acknowledged that saccadic programming, for the mostpart, operates below the level of conscious awareness (Findlay & Gilchrist, 2003; Haggard, 2005; Mokler & Fischer, 1999;Westheimer & Mitchell, 1969), the role of awareness in the saccadic selection of information is less determined. The aimof the present study was to investigate the extent to which observers’ awareness concerning a saccade goal is related to sacc-adic selection performance.
It is possible to make a distinction between at least three different views with respect to this issue. First, it may be thecase that observers are at all times aware of the information that is overtly selected for closer inspection. In other words,observers are aware of the property of the saccade goal that is selected. For example, if an observer selects a saccade goalon the basis of its color, the observer will be aware of this color; and likewise for other properties such as orientation, stim-ulus-saliency and location. The idea here is that objects or locations selected for foveal viewing reach observer’s awarenessvia the attentional system. This proposal is based on the idea that attention is tightly related to the saccadic system (Belo-polsky, Kramer, & Theeuwes, 2008; Deubel & Schneider, 1996; Klein, 1980; Klein & Pontefract, 1994; Kowler, Anderson,Dosher, & Blaser, 1995; Rizzolatti, Riggio, Dascola, & Umilta, 1987; Sheliga, Riggio, Craighero, & Rizzolatti, 1995). Accordingly,many theories assume that a shift in covert attention precedes the eye movement to a location (Deubel & Schneider, 1996;
. All rights reserved.
f Psychology, Vrije Universiteit Amsterdam, Van der Boechorststraat 1, 1081 BT Amsterdam, The
oest).

862 W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871
Henderson, 1992; Hoffman & Subramaniam, 1995; Kowler et al., 1995; Schneider & Deubel, 1995). Although the relationshipbetween attention and awareness is still a matter of debate, attention has been proposed to act as a requisite for awareness(e.g., Posner, 1994). For example, Lamme (2003, 2004) has argued that attention may act as a gate towards a representationthat can be consciously reported. Given the relation between attention and eye movements, and between attention andawareness, attention may provide the critical link between awareness and eye movements. If attention precedes the eyesto a selected location and if attention acts as a requisite for awareness, observers should always be aware of the informationthey select for foveal viewing.
Second, it may be that awareness of the saccade goal is related to the response latency of the saccade. Observers may notbe aware of the information that drives a response when the response is executed shortly following display presentation.However, as times passes and information about the goal location accrues, observers may be become increasingly awareof this information and responses may become increasingly based on this conscious information. Accordingly, observers’awareness may develop over time and be related to whether responses were triggered earlier or later in the stream of pro-cessing (Johnson, Van Beers, & Haggard, 2002). This distinction between short-latency and long-latency responses is corrob-orated by differences in the time-course between stimulus- and goal-driven mechanisms of selection. Whereas earlyselection is primarily automatic and determined by visually salient properties of a display, late selection tends to be morevolitional and controlled by the goals and intentions of the observer (Donk & van Zoest, 2008; van Zoest & Donk, 2005,2006, 2008; van Zoest, Donk, & Theeuwes, 2004). With regards to observers’ awareness, it may be the case that observersare unaware of the saccade goal when they make fast stimulus-driven eye movements, yet are aware of the saccade goalwhen they make slow goal-driven eye movement (however, see Sumner & Husain, 2008). Various studies demonstrate thatpeople do not have much conscious recollection concerning the saccade goal when eye movements are stimulus-driven. Spe-cifically, observers may not be aware of the fact that they make eye movements directed to salient objects in the visual field(Belopolsky et al., 2008; Kramer, Hahn, Irwin, & Theeuwes, 2000; Theeuwes, Kramer, Hahn, & Irwin, 1998; Zhaoping, 2008).Experimental support for the idea that observers are aware of the saccade goal in goal-driven selection is less readily avail-able. However, this seems to be related to the naturalness of the idea rather than to the lack of evidence. Processes thatunderlie visual awareness are very similar to those that regulate voluntary movements (e.g., Hallett, 2007). As a conse-quence, it is often assumed that per definition, goal-driven selection is accompanied by awareness of the saccade goal. Thus,according to this second view, observers’ awareness depends on saccade latency and may in part be driven by differences inthe time-course of stimulus-driven and goal-driven control of selection.
A third view on the relationship posits that people are unaware of the information that guides saccadic selection. Inline with this view, is evidence showing that observers are unaware of the location of a saccade goal even though theywere making a voluntary eye movement. Such findings are primarily derived from studies using a ‘double-step paradigm’(Bridgeman, Hendry, & Stark, 1975; Bridgeman, Kirch, & Sperling, 1981; Bridgeman, Lewis, Heit, & Nagle, 1979; Castiello,Paulignan, & Jeannerod, 1991; Johnson & Haggard, 2005; Prablanc & Martin, 1992; Pélisson, Prablanc, Goodale, & Jeann-erod, 1986). In this task, participants are required to make a saccade to a visual target. However, during the saccadiceye movement the visual target may unpredictably shift a small distance. The results typically show that even thoughobservers fail to detect the shift of the visual target, participants were still able to saccade or point correctly to the centerof the target (Bridgeman et al., 1975, 1979). This work demonstrates that observers are not always aware of the visualinformation that guides the oculomotor system (see also, Deubel, Irwin, & Schneider, 1999; Koch & Tsuchiya, 2007).According to the third view, this may be the rule rather than the exception. Moreover, this may not only apply to infor-mation concerning the location of a saccade goal but also apply to other properties of the saccade goal, such as the color,orientation and saliency.
To briefly summarize, there are different perspectives regarding the relationship between awareness of the saccade goaland saccadic selection performance. However, little work has been conducted to investigate the differences between theseviews systematically. The aim of the present study was to investigate the extent to which observers were aware of the sac-cade goal that is selected for further processing. More specifically, in the present study we investigated whether observerswere aware of the saliency (Experiment 1) and the orientation (Experiment 2) of a saccade goal and investigated the rela-tionship between observers’ awareness and oculomotor performance. In Experiment 1 participants were instructed to selectthe most visually salient element in the display. In Experiment 2, participants were instructed to search for a pre-specifiedidentity, a line oriented to the right. Eye movements were monitored and the proportion of correct eye movements to thetarget was analyzed as a function of saccade latency. Contingent on the first eye movement, the search display was masked.Participants’ awareness of the saccade goal was tested after each trial. In Experiment 1, participants indicated whether or notthey correctly moved their eyes to the most salient element. In Experiment 2, participants indicated whether or not theycorrectly moved their eyes to the right-tilted element. Participants’ report indicating whether an eye movement was cor-rectly directed to the target was used to calculate the sensitivity index d-prime. d-Prime was also analyzed as a functionof saccade latency to investigate whether observers’ awareness varied as a function of the time after the presentation ofthe stimulus display. Note that with saccade goal we refer to the information at the landing position of the first eye move-ment, irrespective of whether the correct target or incorrect distractor was selected.
Based on the three perspectives discussed above we can formulate three predictions. If participants are fully aware of theinformation they select for foveal viewing (view I), we predict that observer’s overall awareness of the saccade goal will behigh. Furthermore, this high level-of-awareness should characterize saccadic selection independent of whether observerswere fast or slow to select the saccade goal. If observers’ awareness depends on the saccade latency (view II), we predict that

W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871 863
the degree of awareness will depend on whether observers were fast or slow to make an eye movement. In this case, aware-ness is predicted to increase with saccade latency. Short-latency eye movements that are primarily driven by stimulus-dri-ven control should occur with little awareness of the saccade goal. As saccade latency increases and selection becomes moregoal-driven, the level-of-awareness of the saccade goal is predicted to become higher. If saccadic selection occurs completelyoutside awareness (view III), we predict that observers will be scarcely aware of the saccade goal. Furthermore, this low le-vel-of-awareness should not depend on saccade latency.
2. Experiment 1
2.1. Method
Experiment 1 investigated the extent to which observers’ awareness of the relative saliency of the saccade goal is relatedto saccadic target selection. Observers were presented with displays containing multiple homogenously oriented back-ground lines and two singletons, each defined by a different orientation-contrast relative to the background lines. The dif-ference between the singleton and the background elements was either 20� or 70�. Participants were given the instruction tomake an eye movement to the visually most salient element in the display which was specified as the singleton with thelargest orientation-contrast relative to the background elements. The most salient singleton, the target, was defined by anorientation contrast of 70� relative to the background lines. The non-salient singleton was defined by an orientation contrastof 20� relative to the background lines. To disentangle the singletons’ identities (i.e., specific orientations) from their relativesaliency, the orientation of the background lines was varied over trials. That is, the background lines were either vertically orhorizontally oriented. This manipulation prevented observers from using identity information to infer relative saliency andencouraged the use of low-level information to guide their search. Critically, as soon as the start of a saccadic movement wasdetected, the search display was masked such that participants did no longer have access to the display information oncethey had landed at the saccade goal. Following each trial, participants were asked to indicate whether they thought theymoved their eyes correctly to the most salient element or not.
2.1.1. ParticipantsEight paid volunteers (aged 18–25 years, average age 20.5 years) participated in Experiment 1. All participants had nor-
mal or corrected-to-normal vision.
2.1.2. ApparatusA Pentium IV computer with a processor speed of 2.3 GHz controlled the timing of the events. Displays were pre-
sented on an Iiyama 2100 SVGA monitor with a resolution of 1280 � 1024 pixels and a 100-Hz refresh rate. Eye move-ments were recorded by means of an Eyelink II tracker (SR Research Ltd.) with a 500 Hz temporal resolution and a 0.2�of visual angle spatial resolution. The system uses an infrared video-based tracking technology to compute the pupilcentre and pupil size of both eyes. An infrared head motion tracking system was also used. All subjects were testedin a sound-attenuated, dimly-lit room with their heads resting on a chinrest. The monitor was located at eye level75 cm from the chinrest.
2.1.3. StimuliThe search array consisted of multiple line segments, two of which were differently oriented than the homogeneous
group of background lines. One singleton was oriented at either 20� or �20� and the other singleton was oriented at either70� or �70�. The background lines were either horizontally or vertically oriented line segments. The line segments were pre-sented in a 17 � 17 square matrix with a raster width of 17.4� � 17.4� of visual angle. The two singletons could be presentedat six potential locations in such a way that in one display the circular angle between the two singletons was 180�. The po-tential singleton locations were all at the same retinal eccentricity (5.3� of visual angle). Elements had an approximate lengthof 0.76� and width of 0.15�.
2.1.4. Design and procedureParticipants first viewed a calibration display consisting of nine points in a square array, which were fixated sequentially.
The eye-tracking system was calibrated at the start of the experiment and following each block. In order to start each trialparticipants maintained fixation on a central dot (0.25�). Participants then pressed the spacebar in order to apply a drift cor-rection and to begin the trial with the presentation of a small fixation point (0.1�) for 1000 ms followed by the search display(see Fig. 1). Observers had the task to make a speeded eye movement towards the most salient element upon appearance ofthe search display. However, the search display was masked contingent to the eye movement, such that when the saccadeended, the search display was no longer visible and participants viewed the mask. The mask display was composed of a ras-ter of 17 � 17 individual masks, each one consisting of multiple superimposed line segments that were orientated at 0�, 20�,�20�, 70�, �70�, and 90�. The mask was presented for 1000 ms. After each trial participants were asked to indicate whetherthey thought they had correctly selected the most salient element (press/-key) or had incorrectly selected the least salientelement (press z-key). This response was probed by a presentation of a display with the question ‘‘did you correctly moveyour eyes?”, and a reminder of the key responses available for report.

Fig. 1. An example of a trial sequence.
864 W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871
There were a total of 24 practice trials and 384 experimental trials. The position of the most salient singleton (6), the ori-entation of the most salient singleton (2), the orientation of the least salient singleton (2), and the orientation of the back-ground lines (2) were randomly varied within blocks of trials. Participants were encouraged to take a break half way duringthe experiment (after 192 trials) and feedback regarding saccade latency was provided every 24 trials. To ensure that par-ticipants fully understood the task both verbal and written instructions were provided.
2.1.5. Data analysisAn eye movement was considered a saccade when either eye velocity exceeded 35�/s or eye acceleration exceeded 9500�/
s2. Saccade latency was defined as the interval between the display of the search display and the initiation of a saccadic eyemovement. If the saccade latency was shorter than 80 ms (anticipation error), or longer than 600 ms, the trial was removedfrom analysis. The initial saccade was assigned to a target or distractor if the endpoint of the initial saccade was within 2.4�of visual angle of the particular target or distractor position. If the saccade did not reach either the location of the target orthe distractor, the trial was excluded from further analysis.
2.2. Results
In 8.5% of the trials the initial saccade latency was shorter than 80 ms and in 2.6% of the trials saccade latency was higherthan 600 ms. Of the initial saccades 4.8% missed either the location of the target or the distractor.
2.2.1. Eye movement dataIn line with the instructions, the results showed that the salient singleton was significantly more often selected than the
non-salient singleton, 58.0% vs. 42.0%, t(7) = 7.59, p < .0005. Moreover, saccade latencies to salient singletons were shorter(mean 268.7 ms) than latencies directed to non-salient singletons (mean 279.8 ms), t(7) = 2.67, p = .032.
The proportion of eye movements directed correctly towards the target was analyzed as a function of saccade latency. Foreach participant, an individual distribution of the initial saccade latencies was calculated irrespective of the saccade desti-nation. Distributions were divided into five quintiles based on saccade latency and the proportion of correct eye movementswas calculated for each quintile. For each quintile the average saccade latency was calculated (see Fig. 2). An analysis of var-
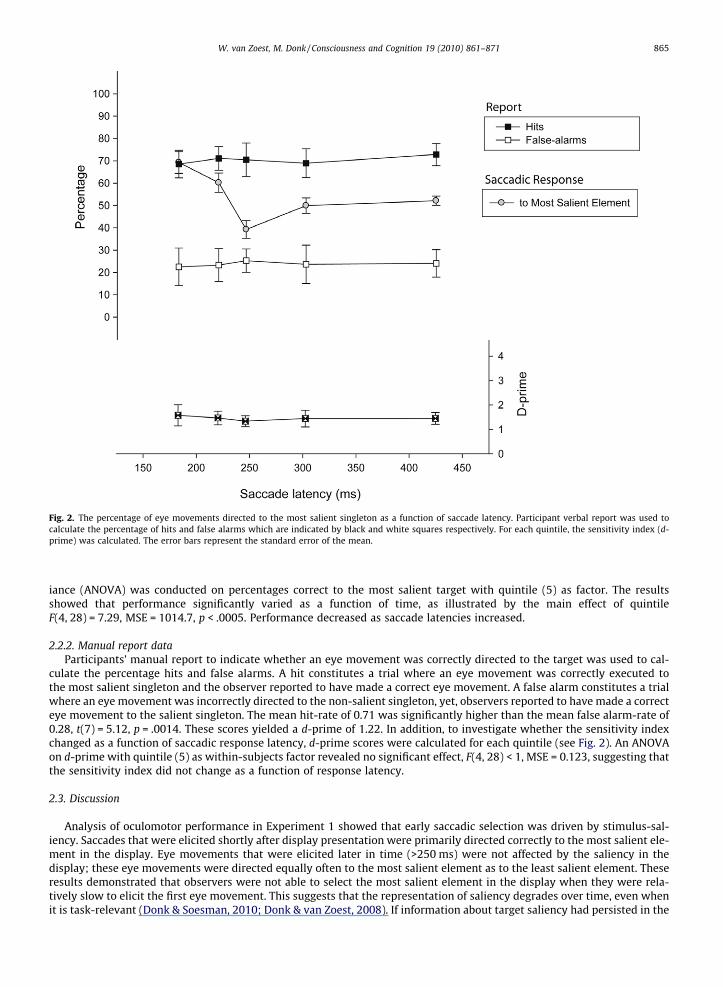
Fig. 2. The percentage of eye movements directed to the most salient singleton as a function of saccade latency. Participant verbal report was used tocalculate the percentage of hits and false alarms which are indicated by black and white squares respectively. For each quintile, the sensitivity index (d-prime) was calculated. The error bars represent the standard error of the mean.
W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871 865
iance (ANOVA) was conducted on percentages correct to the most salient target with quintile (5) as factor. The resultsshowed that performance significantly varied as a function of time, as illustrated by the main effect of quintileF(4, 28) = 7.29, MSE = 1014.7, p < .0005. Performance decreased as saccade latencies increased.
2.2.2. Manual report dataParticipants’ manual report to indicate whether an eye movement was correctly directed to the target was used to cal-
culate the percentage hits and false alarms. A hit constitutes a trial where an eye movement was correctly executed tothe most salient singleton and the observer reported to have made a correct eye movement. A false alarm constitutes a trialwhere an eye movement was incorrectly directed to the non-salient singleton, yet, observers reported to have made a correcteye movement to the salient singleton. The mean hit-rate of 0.71 was significantly higher than the mean false alarm-rate of0.28, t(7) = 5.12, p = .0014. These scores yielded a d-prime of 1.22. In addition, to investigate whether the sensitivity indexchanged as a function of saccadic response latency, d-prime scores were calculated for each quintile (see Fig. 2). An ANOVAon d-prime with quintile (5) as within-subjects factor revealed no significant effect, F(4, 28) < 1, MSE = 0.123, suggesting thatthe sensitivity index did not change as a function of response latency.
2.3. Discussion
Analysis of oculomotor performance in Experiment 1 showed that early saccadic selection was driven by stimulus-sal-iency. Saccades that were elicited shortly after display presentation were primarily directed correctly to the most salient ele-ment in the display. Eye movements that were elicited later in time (>250 ms) were not affected by the saliency in thedisplay; these eye movements were directed equally often to the most salient element as to the least salient element. Theseresults demonstrated that observers were not able to select the most salient element in the display when they were rela-tively slow to elicit the first eye movement. This suggests that the representation of saliency degrades over time, even whenit is task-relevant (Donk & Soesman, 2010; Donk & van Zoest, 2008). If information about target saliency had persisted in the

866 W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871
visual system, correct selection of the salient location should have been possible regardless of whether saccadic target selec-tion would have been quick or slow.
Analysis of observers’ manual report regarding saccade goal awareness revealed three important finding. First, overallsensitivity was low suggesting that observers were poorly able to indicate whether they made an eye movement to the mostsalient item or not. Second, awareness of the selected response was not related to saccadic performance. In other words,observers’ awareness of the selected item did not depend on whether observers correctly selected the most salient targetor not. Third, observers’ awareness of the saccade goal did not vary as a function of saccade latency. Regardless of whetherparticipants had viewed the display for a short or long duration before making the first eye movement, awareness of thesaccade goal was low. Based on the second view outlined in the introduction, it was predicted that awareness of the saccadegoal may develop as a function of time. Accordingly, it was predicted that observers would have had a higher level-of-aware-ness regarding the saccade goal when they had more time to process the display. However, timing of the response did notaffect the observers’ awareness of the saccade goal. To sum up, the results suggest that eye movements are elicited with min-imal awareness of the relative saliency in the display. The results of Experiment 1 are not in line with the idea that awarenessof the saccade goal is invariably high (view I), nor with the idea that awareness is related to saccade latency (view II). Exper-iment 1 is mostly in line with the idea that observers are unaware of the information that guides saccadic selection (view III).Experiment 2 was set up to investigate to what extent observers are aware of the identity of a saccade goal.
3. Experiment 2
3.1. Method
Participants were instructed to make an eye movement to the orientation singleton that was tilted to the right. The right-tilted orientation singleton was either a salient or a non-salient singleton in the display. Note that in the latter case there is aconflict between stimulus-driven and goal-driven influences. Similar to Experiment 1, the search display was masked con-tingent to the eye movement, such that participants viewed a mask when their eyes landed on the saccade goal. Followingeach trial participants were asked to indicate whether they had or had not correctly made an eye movement to the right-tilted target.
3.1.1. ParticipantsTen paid volunteers (aged 18–22 years, average age 20.7 years) participated in Experiment 2. All participants had normal
or corrected-to-normal vision. One participant was excluded due to an excess of anticipation errors, yielding a total of nineparticipants in Experiment 2.
3.1.2. Apparatus and stimuliThe apparatus was identical to that in Experiment 1. The stimuli presented were identical, except that the two singletons
presented were never oriented in the same direction. Singletons always pointed in opposite directions, e.g., if one singletonwas tilted to the right (e.g., 20�), the other was left-tilted (e.g., �70�). This way, there were never two right-titled or two left-tilted singletons; there was always one right-tilted element present in the display, either rotated at 20� or at 70�.
3.1.3. Design and procedureParticipants were given the instruction to make an eye movement to the singleton element that was tilted to the right.
This singleton element was either a non-salient element, when the absolute difference between the right-tilted singletonand the nontargets was 20� or was a salient singleton, that is, when the absolute difference between the right-tilted singletonand the nontargets was 70�. Analyses were performed separately for the salient target and non-salient target.
There was a total of 24 practice trials and 384 experimental trials. The position of the most salient singleton (6), the sal-iency of the right-tilted element (2), and the orientation of the background lines (2) were randomly varied within blocks oftrials. Except for the above, the procedure was identical to that of Experiment 1.
3.2. Results
In 9.8% of the trials the initial saccade latency was shorter than 80 ms and in 3.5% of the trials saccade latency was higherthan 600 ms; of the initial saccades 11.4% missed either the location of the target or the distractor.
3.2.1. Eye movement dataThe results showed that the right-tilted salient target was significantly more often correctly selected than the right-tilted
non-salient target, 61.5% vs. 42.6%, t(8) = 9.47, p < .0001. Saccade latencies of eye movements directed to the salient right-tilted targets were shorter than saccade latencies directed to non-salient right-tilted targets, 213.1 ms vs. 240.7 ms,t(8) = 3.94, p = .0043.
The analysis of the data as a function of time showed that the percentage of correct eye movements depended on thesaliency of the right-tilted target, as well as on saccade latency (see Fig. 3). These observations were supported by an ANOVA
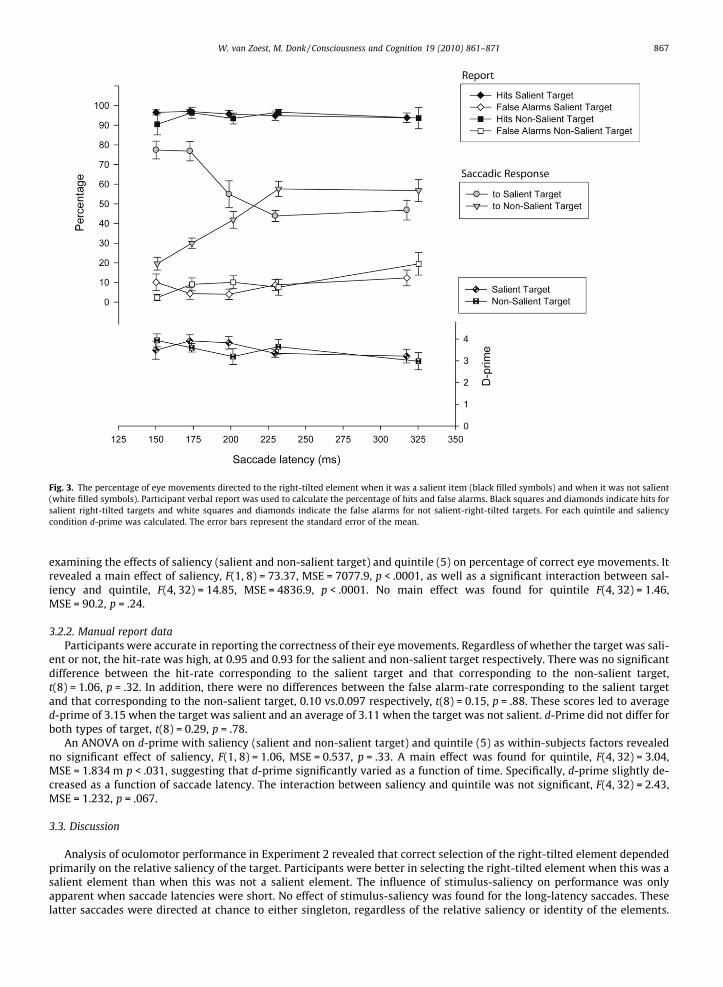
Fig. 3. The percentage of eye movements directed to the right-tilted element when it was a salient item (black filled symbols) and when it was not salient(white filled symbols). Participant verbal report was used to calculate the percentage of hits and false alarms. Black squares and diamonds indicate hits forsalient right-tilted targets and white squares and diamonds indicate the false alarms for not salient-right-tilted targets. For each quintile and saliencycondition d-prime was calculated. The error bars represent the standard error of the mean.
W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871 867
examining the effects of saliency (salient and non-salient target) and quintile (5) on percentage of correct eye movements. Itrevealed a main effect of saliency, F(1, 8) = 73.37, MSE = 7077.9, p < .0001, as well as a significant interaction between sal-iency and quintile, F(4, 32) = 14.85, MSE = 4836.9, p < .0001. No main effect was found for quintile F(4, 32) = 1.46,MSE = 90.2, p = .24.
3.2.2. Manual report dataParticipants were accurate in reporting the correctness of their eye movements. Regardless of whether the target was sali-
ent or not, the hit-rate was high, at 0.95 and 0.93 for the salient and non-salient target respectively. There was no significantdifference between the hit-rate corresponding to the salient target and that corresponding to the non-salient target,t(8) = 1.06, p = .32. In addition, there were no differences between the false alarm-rate corresponding to the salient targetand that corresponding to the non-salient target, 0.10 vs.0.097 respectively, t(8) = 0.15, p = .88. These scores led to averaged-prime of 3.15 when the target was salient and an average of 3.11 when the target was not salient. d-Prime did not differ forboth types of target, t(8) = 0.29, p = .78.
An ANOVA on d-prime with saliency (salient and non-salient target) and quintile (5) as within-subjects factors revealedno significant effect of saliency, F(1, 8) = 1.06, MSE = 0.537, p = .33. A main effect was found for quintile, F(4, 32) = 3.04,MSE = 1.834 m p < .031, suggesting that d-prime significantly varied as a function of time. Specifically, d-prime slightly de-creased as a function of saccade latency. The interaction between saliency and quintile was not significant, F(4, 32) = 2.43,MSE = 1.232, p = .067.
3.3. Discussion
Analysis of oculomotor performance in Experiment 2 revealed that correct selection of the right-tilted element dependedprimarily on the relative saliency of the target. Participants were better in selecting the right-tilted element when this was asalient element than when this was not a salient element. The influence of stimulus-saliency on performance was onlyapparent when saccade latencies were short. No effect of stimulus-saliency was found for the long-latency saccades. Theselatter saccades were directed at chance to either singleton, regardless of the relative saliency or identity of the elements.

868 W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871
Overall, the eye movement data revealed that participants were unable to overtly select the right-tilted element on the basisof its identity.
The analysis of observers’ manual report regarding saccade goal awareness revealed three important finding. First, overallsensitivity was high suggesting that observers were very well able to indicate whether they made a correct eye movement tothe right-tilted item or not. Even for saccades that were triggered very shortly after display presentation (e.g., 150 ms)observers were almost perfectly able to indicate whether they had correctly or incorrectly selected the predefined target.Second, awareness was not related to saccadic performance. Observers’ awareness of the saccade goal did not depend onwhether observers correctly selected the right-tilted target or not. Third, observers’ awareness of the saccade goal variedas a function of saccade latency. However, the relationship between awareness and latency was opposite to what we hadpredicted on the basis of the second view that we outlined in the introduction. The results revealed that as saccade latencyincreased, participants were less able to indicate the orientation of the saccade goal. It was predicted that awareness wouldbe positively associated to saccade latency, such that when observers have more time to process the display, awareness ofthe to-be-selected singleton would increase. However, the results demonstrated, surprisingly, that observers were signifi-cantly less sensitive to the identity of the saccade goal when saccade latencies were higher.
To sum up, the results suggest that eye movements are elicited with a high level-of-awareness of the orientation of thesaccade goal. The results of Experiment 2 are not in line with the idea that awareness of the saccade goal is invariably low(view III), nor with the idea that awareness is positively related to saccade latency (view II). Experiment 2 is mostly in linewith the idea that observers are aware of the information that guides saccadic selection (view I).
4. General discussion
In two experiments, we investigated the extent to which observers’ awareness of the saccade goal was related to saccadicselection performance. Participants were required to saccade to the either the most salient line element (Experiment 1) orthe line element that was tilted to the right (Experiment 2) in a search display consisting of two singletons and multiplehomogenously oriented background lines. The search display was masked contingent on the start of the first saccade suchthat the display was no longer visible the moment the eyes landed on the saccade goal. Following each trial participants indi-cated whether they had correctly made a saccade to the target or not. Participants’ report was used to investigate the level-of-awareness of the saccade goal.
Experiment 1 revealed that observers were poorly aware of the relative saliency of the saccade goal. In contrast to theresults obtained in Experiment 1, Experiment 2 revealed that observers had a high level-of-awareness concerning the iden-tity (i.e., orientation) of the saccade goal. This combination of results is not in line with the idea that awareness of the saccadegoal is invariably high (view I), nor with the idea that awareness of the saccade goal is invariably low (view III). The currentwork demonstrates that observers’ awareness may depend on the property observers are asked to report. Further researchmay provide insight in the relationship between the awareness of other attributes of a saccade goal, like color and position.
Importantly, in neither Experiment 1 nor Experiment 2 was the level-of-awareness of the overtly selected element relatedto oculomotor performance. For example, in Experiment 1 participants were correctly selecting the most salient elementwhen saccade latencies where short. However, participants were relatively unaware of the fact that they were making thesecorrect eye movements. Though observers obtained a high level of oculomotor performance early during selection, they wereunable to maintain this level at longer saccade latencies. And even though oculomotor behavior changed over time, people’sawareness did not vary as a function of saccade latency. In Experiment 2, the results showed that observers were very wellable to identify the orientation of the saccade goal; in fact, they were able to do so already after 150 ms of viewing the dis-play before it was masked. However, this high level-of-awareness of the saccade goal early during selection did not enableparticipants to guide the eye movement system to the correct target location at longer latencies. The findings from the pres-ent study suggest that there is no relationship between awareness of the saccade goal and saccadic eye movementperformance.
The results indicated that irrespective of the instruction given to the participants – thus, regardless of whether partici-pants were instructed to search for the target on the basis of saliency or identity – early saccadic selection was primarilydriven by stimulus-saliency. In both experiments the effect of stimulus-saliency dissipated over time and no influence ofstimulus-saliency was found for the high-latency saccades. More specifically, in Experiment 2, saccades elicited shortly afterdisplay presentation were directed to the most salient element, regardless of whether it was tilted to the right or left. Thisresulted in a high level of saccadic performance when the right-tilted target was the most salient item, but a low level ofperformance when the right-tilted target was not salient. Later in time, selection was no longer driven by stimulus-saliency(see also, Donk & Soesman, 2010; Donk & van Zoest, 2008; Hunt, von Mühlenen, & Kingstone, 2007; van Zoest & Donk, 2006,2008; van Zoest, Hunt, & Kingstone, 2010). However, while these results show that the mode of selection consistently chan-ged as a function of saccade latency, no associated differences in awareness were observed. The present results are not in linewith the idea that awareness of the saccade goal increases with saccade latency and may depend on whether selection wasstimulus-driven or goal-driven (view II).
Previous studies have demonstrated that participants may be unaware of the fact that they make erroneous stimulus-dri-ven eye movement to irrelevant abrupt onsets (Belopolsky et al., 2008; Kramer, Hahn, Irwin, & Theeuwes, 1999; Theeuweset al., 1998). However, there is an important distinction to make here regarding the awareness of events in oculomotor cap-

W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871 869
ture. On the one hand, there is the awareness regarding the saccade goal, the abrupt onset; on the other hand, there is theawareness of the incorrect eye movement in the direction of the abrupt onset. The majority of studies that have investigatedawareness of saccadic selection have looked at awareness of the initial direction of the eye movement. For example, Belo-polsky et al. (2008) showed that participants may be partially aware of the fact that in some trials their eyes moved incor-rectly to the onset before moving onto the target. In this study, participants were more likely to report an initial misdirectionof the eyes to the onset than that they failed to report it. Accordingly, it was concluded that stimulus-driven selection doesnot always occur outside awareness. Studies on anti-saccades have provided support for the idea that stimulus-driven orreflexive saccadic selection may occur outside awareness. In the anti-saccade task participants are required to make aneye movement to a location in space opposite to the location of a salient visual target (Hallet, 1978; Hallet & Adams,1980). Results show that incorrect reflexive pro-saccades directed to the visual target often occur outside observers’ aware-ness (Endrass, Reuter, & Kathmann, 2007; Mokler & Fischer, 1999; Nieuwenhuis, Ridderinkhof, Blom, Band, & Kok, 2001).However, similar to Belopolsky et al. (2008), observers’ awareness of erroneous pro-saccades concern the initial directionof the eye movement and this is considerably different from observers’ awareness of the saccade goal.
One study that has looked at awareness of the saccade goal is the study by Kramer et al. (2000) in which the luminance ofan abrupt onset was varied as a means to investigate how observers’ awareness of an irrelevant onset relates to awareness inoculomotor capture. In this study, the abrupt onset presented was either equiluminant with the other stimuli in the displayor brighter than the other stimuli. It was hypothesized that the awareness of the onset distractor would depend on the lumi-nance of the onset: observers were predicted to be able to report the bright onset but not the equiluminant onset. Partici-pants were questioned after the experiment whether they had noticed the presentation of the abrupt onset during thepresentation of the search displays and whether they had made an eye movement to this item. Although the results revealedthat both the equiluminant and the bright onset captured the eyes, observers reported never to have made incorrect eyemovements in the direction of the onset distractor. Importantly, as predicted, while observers reported never having seenthe equiluminant abrupt onset, all observers reported to have seen the brighter abrupt onset. These results suggest that eventhough observers were aware of the irrelevant bright onset distractor, involuntary saccadic selection towards this distractoroccurred completely outside awareness (Kramer et al., 2000).
The present study is different from the aforementioned study in a number of ways. In the current study observers’ aware-ness concerned awareness of the saccade goal only and participants were not questioned about the movement of the eye.Also, instead of questioning the participants in a general way after the experiment was finished, participants in the presentstudy were asked to report on their responses following each trial. The present design required observers to respond both incase of a correct and incorrect saccade. In Kramer et al. (2000), observers’ awareness concerned the irrelevant distractor only;the awareness regarding information at the pre-specified target location was never questioned. In the present study, unlikeprevious work on awareness and saccadic selection (Belopolsky et al., 2008; Kramer et al., 2000; Theeuwes et al., 1998), thesearch display was masked contingent on the first eye movement. This ensured that the awareness concerning the target ordistractor was for the most part attained in the time limited to the time before the first initial eye movement, and was notattained later during the trial.
The present study demonstrated that observers’ high level-of-awareness concerning the orientation of the saccade goaldid not benefit performance; orientation awareness was not related to saccadic selection. These findings are in line with theidea that eye movements are mediated by a system that operates independently from the visual system which enables con-scious perception of objects in the visual field. Studies of the neurophysiology of the primate visual system provide evidencefor the existence of two functionally distinct neural streams that mediate processing of visual information (e.g., Ungerleider& Mishkin, 1982). Whereas a dorsal system runs from V1 to the posterior parietal cortex and is assumed to be primarily in-volved in spatial processing and visuomotor performance, the ventral stream runs from V1 to the inferior temporal cortexand is assumed to be involved in object identification and recognition. Supporting evidence for this dual-route hypothesiswas found in studies of neuropsychological patients (Goodale & Milner, 1992; Goodale, Milner, Jakobson, & Carey, 1991; Mil-ner & Goodale, 1995). The results of the present study are in line with the idea that (unaware) oculomotor behavior mayprimarily rely on dorsal stream processing while conscious report relies on ventral stream processing with the result thatboth a largely independent (see, Andersen, 1989). However, recent evidence questions the functional independence of thetwo pathways (e.g., Dassonville & Bala, 2004; de Grave, Smeets, & Brenner, 2006; Eckstein, Beutter, Pham, Shimozaki, &Stone, 2007; Franz & Gegenfurtner, 2008). Moreover, pathways controlling saccadic movements are manifold. These includevisual cortical pathways involving the frontal eye fields (Schall & Hanes, 1993), the lateral intraparietal cortex (Goldberg, Bis-ley, Powell, Gottlieb, & Kusunoki, 2002) as well as subcortical routes involving the superior colliculus (McPeek & Keller,2002). Evidence furthermore suggests a role for the ventral stream in the programming of saccadic eye movements (Mazer& Gallant, 2003; Moore, 1999). As such, there may be alternative ways to explain the obtained dissociation between oculo-motor behavior and awareness. Additional research may provide insight in the neural substrate that could explain the inde-pendence between saccadic selection and awareness found in the present study (e.g., Desmurget & Sirigu, 2009; Desmurgetet al., 2009).
The present study demonstrated that people awareness concerning the saccade goal is not directly related to saccadicselection. Previous studies have similarly demonstrated that oculomotor behavior may be unrelated to awareness. For in-stance Bridgeman et al. (1975) demonstrated that people may initiate an eye movement to a visual event that they didnot perceive. However, findings like these have been primarily reported with displaced stimuli (Deubel, Schneider, & Bridg-eman, 1996), displays that are presented binocularly (Zhaoping, 2008), visual illusions (Dassonville & Bala, 2004; de’Sperati

870 W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871
& Baud-Bovy, 2008), and in patient populations (Zihl, 1980). The present study shows that also when people perform a rel-ative simple visual search task, eye movement behavior is strongly dissociated from conscious awareness.
Acknowledgement
WvZ was supported by a grant from the Netherlands Organization for Scientific Research.
References
Andersen, R. A. (1989). Visual and eye movement functions of the posterior parietal cortex. Annual Review of Neuroscience, 12, 377–403.Belopolsky, A. V., Kramer, A. F., & Theeuwes, J. (2008). The role of awareness in processing of oculomotor capture: Evidence from event-related potentials.
Journal of Cognitive Neuroscience, 20(12), 2285–2297.Bridgeman, B., Hendry, D., & Stark, L. (1975). Failure to detect displacement of the visual world during saccadic eye movements. Vision Research, 15(6),
719–722.Bridgeman, B., Kirch, M., & Sperling, A. (1981). Segregation of cognitive and motor aspects of visual function using induced motion. Perception &
Psychophysics, 29(4), 336–342.Bridgeman, B., Lewis, S., Heit, G., & Nagle, M. (1979). Relation between cognitive and motor-oriented systems of visual position perception. Journal of
Experimental Psychology: Human Perception and Performance, 5(4), 692–700.Castiello, U., Paulignan, Y., & Jeannerod, M. (1991). Temporal dissociation of motor responses and subjective awareness. A study in normal subjects. Brain,
114(Pt 6), 2639–2655.Dassonville, P., & Bala, J. K. (2004). Perception, action, and roelofs effect: A mere illusion of dissociation. PLoS Biology, 2(11), e364.de Grave, D. D., Smeets, J. B., & Brenner, E. (2006). Why are saccades influenced by the Brentano illusion? Experimental Brain Research, 175(1), 177–182.Desmurget, M., Reilly, K. T., Richard, N., Szathmari, A., Mottolese, C., & Sirigu, A. (2009). Movement intention after parietal cortex stimulation in humans.
Science, 324(5928), 811–813.Desmurget, M., & Sirigu, A. (2009). A parietal–premotor network for movement intention and motor awareness. Trends in Cognitive Sciences, 13(10),
411–419.de’Sperati, C., & Baud-Bovy, G. (2008). Blind saccades: An asynchrony between seeing and looking. Journal of Neuroscience, 28(17), 4317–4321.Deubel, H., Irwin, D. E., & Schneider, W. X. (1999). The subjective direction of gaze shifts long before the saccade. In W. Becker, H. Deubel, & T. Mergner (Eds.),
Current oculomotor research: Physiological and psychological aspects. New York, London: Plenum Publishers.Deubel, H., & Schneider, W. A. (1996). Saccade target selection and object recognition: Evidence for a common attentional mechanism. Vision Research,
36(12), 1827–1837.Deubel, H., Schneider, W. X., & Bridgeman, B. (1996). Postsaccadic target blanking prevents saccadic suppression of image displacement. Vision Research,
36(7), 985–996.Donk, M., & Soesman, L. (2010). Salience is only briefly represented: Evidence from probe-detection performance. Journal of Experimental Psychology: Human
Perception and Performance, 36(2), 286–302.Donk, M., & van Zoest, W. (2008). Effects of salience are short-lived. Psychological Science, 19(7), 733–739.Eckstein, M. P., Beutter, B. R., Pham, B. T., Shimozaki, S. S., & Stone, L. S. (2007). Similar neural representations of the target for saccades and perception
during search. Journal of Neuroscience, 27(6), 1266–1270.Endrass, T., Reuter, B., & Kathmann, N. (2007). ERP correlates of conscious error recognition: Aware and unaware errors in an antisaccade task. European
Journal of Neuroscience, 26(6), 1714–1720.Findlay, J. M., & Gilchrist, I. D. (2003). Active vision. New York: Oxford University Press.Franz, V. H., & Gegenfurtner, K. R. (2008). Grasping visual illusions: Consistent data and no dissociation. Cognitive Neuropsychology, 25(7), 920–950.Goldberg, M. E., Bisley, J., Powell, K. D., Gottlieb, J., & Kusunoki, M. (2002). The role of the lateral intraparietal area of the monkey in the generation of
saccades and visuospatial attention. Annals of the New York Academy of Sciences, 956, 205–215.Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways for perception and action. Trends in Neurosciences, 15(1), 20–25.Goodale, M. A., Milner, A. D., Jakobson, L. S., & Carey, D. P. (1991). A neurological dissociation between perceiving objects and grasping them. Nature,
349(6305), 154–156.Haggard, P. (2005). Conscious intention and motor cognition. Trends in Cognitive Sciences, 9(6), 290–295.Hallet, P. E. (1978). Primary and secondary saccades to goals defined by instructions. Vision Research, 18, 1279–1296.Hallett, M. (2007). Volitional control of movement: the physiology of free will. Clinical Neurophysiology, 118(6), 1179–1192.Hallet, P. E., & Adams, B. D. (1980). The predictability of saccadic latency in a novel voluntary oculomotor task. Vision Research, 20, 329–339.Henderson, J. M. (1992). Visual attention and eye movement control during reading and picture viewing. In K. Rayner (Ed.), Eye movements and visual
cognition (pp. 261–283). Berlin: Springer.Hoffman, J. E., & Subramaniam, B. (1995). The role of visual attention in saccadic eye movements. Perception & Psychophysics, 37(6), 787–795.Hunt, A. R., von Mühlenen, A., & Kingstone, A. (2007). The time course of attentional and oculomotor capture reveals a common cause. Journal of
Experimental Psychology: Human Perception and Performance, 33(2), 271–284.Johnson, H., & Haggard, P. (2005). Motor awareness without perceptual awareness. Neuropsychologia, 43(2), 227–237.Johnson, H., Van Beers, R. J., & Haggard, P. (2002). Action and awareness in pointing tasks. Experimental Brain Research, 146(4), 451–459.Klein, R. M. (1980). Does oculomotor readiness mediate cognitive control of visual attention? In R. Nickerson (Ed.), Attention and performance. Hillsdale, NJ:
Lawrence Erlbaum.Klein, R. M., & Pontefract, A. (1994). Does oculomotor readiness mediate cognitive control of visual attention? Revisited! In C. Umilta & M. Moscovitch (Eds.),
Attention and performance 15: Conscious and nonconscious information processing. Attention and performance series (pp. 333–350). Cambridge, MA: TheMIT Press.
Koch, C., & Tsuchiya, N. (2007). Attention and consciousness: Two distinct brain processes. Trends in Cognitive Sciences, 11(1), 16–22.Kowler, E., Anderson, E., Dosher, B., & Blaser, E. (1995). The role of attention in the programming of saccades. Vision Research, 35(13), 1897–1916.Kramer, A. F., Hahn, S., Irwin, D. E., & Theeuwes, J. (1999). Attentional capture and aging: Implications for visual search performance and oculomotor control.
Psychology and Aging, 14(1), 135–154.Kramer, A. F., Hahn, S., Irwin, D. E., & Theeuwes, J. (2000). Age differences in the control of looking behavior: Do you know where your eyes have been?
Psychological Science, 11(3), 210–217.Lamme, V. A. (2003). Why visual attention and awareness are different. Trends in Cognitive Sciences, 7(1), 12–18.Lamme, V. A. (2004). Separate neural definitions of visual consciousness and visual attention; a case for phenomenal awareness. Neural Networks, 17(5–6),
861–872.Mazer, J. A., & Gallant, J. L. (2003). Goal-related activity in V4 during free viewing visual search. Evidence for a ventral stream visual salience map. Neuron,
40(6), 1241–1250.McPeek, R. M., & Keller, E. L. (2002). Saccade target selection in the superior colliculus during a visual search task. Journal of Neurophysiology, 88, 2019–2034.Milner, A. D., & Goodale, M. A. (1995). The visual brain in action. Oxford, England: Oxford University Press.Mokler, A., & Fischer, B. (1999). The recognition and correction of involuntary prosaccades in an antisaccade task. Experimental Brain Research, 125, 511–516.

W. van Zoest, M. Donk / Consciousness and Cognition 19 (2010) 861–871 871
Moore, T. (1999). Shape representations and visual guidance of saccadic eye movements. Science, 285(5435), 1914–1917.Nieuwenhuis, S., Ridderinkhof, K. R., Blom, J., Band, G. P., & Kok, A. (2001). Error-related brain potentials are differentially related to awareness of response
errors: Evidence from an antisaccade task. Psychophysiology, 38(5), 752–760.Pélisson, D., Prablanc, C., Goodale, M. A., & Jeannerod, M. (1986). Visual control of reaching movements without vision of the limb. II. Evidence of fast
unconscious processes correcting the trajectory of the hand to the final position of a double-step stimulus. Experimental Brain Research, 62(2), 303–311.Posner, M. I. (1994). Attention: The mechanisms of consciousness. Proceedings of the National Academy of Sciences of the United States of America, 91(16),
7398–7403.Prablanc, C., & Martin, O. (1992). Automatic control during hand reaching at undetected two-dimensional target displacements. Journal of Neurophysiology,
67(2), 455–469.Rizzolatti, G., Riggio, L., Dascola, I., & Umilta, C. (1987). Reorienting attention across the horizontal and vertical meridians: Evidence in favor of a premotor
theory of attention. Neuropsychologia, 25(1-A), 31–40.Schall, J. D., & Hanes, D. P. (1993). Neural basis of saccade target selection in frontal eye field during visual search. Nature, 366(6454), 467–469.Schneider, W. X., & Deubel, H. (1995). Visual attention and saccadic eye movements: Evidence for obligatory and selective spatial coupling. In J. M. Findlay,
R. Walker, & R. W. Kentridge (Eds.), Eye movement research: Mechanism, processes and applications. Amsterdam: Elsevier Science BV.Sheliga, B. M., Riggio, L., Craighero, L., & Rizzolatti, G. (1995). Spatial attention-determined modifications in saccade trajectories. Neuroreport: An
International Journal for the Rapid Communication of Research in Neuroscience, 6(3), 585–588.Sumner, P., & Husain, M. (2008). At the edge of consciousness: Automatic motor activation and voluntary control. Neuroscientist, 14(5), 474–486.Ungerleider, L. G., & Mishkin, M. (Eds.). (1982). Analysis of visual behavior. Cambridge, MA: MIT Press.Theeuwes, J., Kramer, A. F., Hahn, S., & Irwin, D. E. (1998). Our eyes do not always go where we want them to go: Capture of the eyes by new objects.
Psychological Science, 9(5), 379–385.van Zoest, W., & Donk, M. (2005). The effects of salience on saccadic target selection. Visual Cognition, 2(2), 353–375.van Zoest, W., & Donk, M. (2006). Saccadic target selection as a function of time. Spatial Vision, 19(1), 61–76.van Zoest, W., & Donk, M. (2008). Goal-driven modulation as a function of time in saccadic target selection. Quarterly Journal of Experimental Psychology,
61(10), 1553–1572.van Zoest, W., Donk, M., & Theeuwes, J. (2004). The role of stimulus-driven and goal-driven control in saccadic visual selection. Journal of Experimental
Psychology: Human Perception and Performance, 30(4), 746–759.van Zoest, W., Hunt, A. R., & Kingstone, A. (2010). Representations in visual cognition: It’s about time. Current Directions in Psychological Science.Westheimer, G., & Mitchell, D. E. (1969). The sensory stimulus for disjunctive eye movements. Vision Research, 9(7), 749–755.Zhaoping, L. (2008). Attention capture by eye of origin singletons even without awareness—A hallmark of a bottom-up saliency map in the primary visual
cortex. Journal of Vision.Zihl, J. (1980). ‘‘Blindsight”: Improvement of visually guided eye movements by systematic practice in patients with cerebral blindness. Neuropsychologia,
18(1), 71–77.
Related Documents











