Automatic and Controlled Processing of Melodic Contour and Interval Information Measured by Electrical Brain Activity Laurel J. Trainor 1,2 , Kelly L. McDonald 2 , and Claude Alain 2,3 Abstract & Most work on how pitch is encoded in the auditory cortex has focused on tonotopic (absolute) pitch maps. However, melodic information is thought to be encoded in the brain in two different ‘‘relative pitch’’ forms, a domain-general contour code (up/down pattern of pitch changes) and a music-specific interval code (exact pitch distances between notes). Event- related potentials were analyzed in nonmusicians from both passive and active oddball tasks where either the contour or the interval of melody–final notes was occasionally altered. The occasional deviant notes generated a right frontal positivity peaking around 350 msec and a central parietal P3b peaking around 580 msec that were present only when participants focused their attention on the auditory stimuli. Both types of melodic information were encoded automatically in the absence of absolute pitch cues, as indexed by a mismatch negativity wave recorded during the passive conditions. The results indicate that even in the absence of musical training, the brain is set up to automatically encode music-specific melodic information, even when absolute pitch information is not available. & INTRODUCTION How sound information is represented in the auditory cortex is not entirely understood. It is clear that there are tonotopic maps in the primary auditory cortex whereby different pitches are represented in different cortical areas in an orderly fashion (e.g., Woods, Alho, & Allgazi, 1993; Pantev, Hoke, Lutkenhoner, & Lehnertz, 1989; Pantev et al., 1995). However, the information in melodies is not so much in the absolute frequencies or pitches of the tones, but rather in the pitch relations or pitch distances between the tones—a melody retains its identity whether rendered in a high or a low pitch range, as long as the relative pitch relations are maintained. At present, the stage of processing at which relative pitch information is extracted is not known. We investigate this question in a group of nonmusicians. There are reasons to expect that the auditory cortex might be setup up to extract music-specific information. Music is an integral part of every human society (Wallin, Merker, & Brown, 2000), and there is evidence that melodic music-making has a long history in human cultural evolution (Kuneg & Turk, 2000). There is also evidence that across languages and cultures parents use music to communicate to young infants long before they understand language (Trehub & Trainor, 1998). At the same time, music is often considered to be a ‘‘frill,’’ an evolutionarily unimportant accident in the development of speech perception (e.g., Pinker, 1994). Brain imaging studies (PET and fMRI) have clearly shown that a large number of brain regions are involved in melodic pro- cessing, including the temporal and frontal regions, the supplementary motor areas, and the thalamus (e.g., Halpern & Zatorre, 1999; Mirz et al., 1999; Platel et al., 1997; Zatorre, Halpern, Perry, Meyer, & Evans, 1996). Although more studies implicate the right than the left hemisphere in musical processing, both hemispheres are clearly involved (e.g., Peretz & Herbert, 2000; Patel & Peretz, 1997; Zatorre, 1984; Zatorre et al., 1996), and musical input stimulates the perceptual, cognitive, me- mory, and emotional systems. Melodic information is thought to be encoded in two basic forms (e.g., Liegeois-Chauvel, Peretz, Babaie, La- guitton, & Chauvel, 1998; Peretz & Morais, 1987; Peretz, Morais, & Bertelson, 1987; Peretz, 1990; Peretz & Babaie, 1992; Edworthy, 1985; Dowling, 1978, 1982; Bever & Chiarello, 1974). The contour code consists of the up/ down pattern of pitch changes without regard to the size of those changes. The interval code consists of the exact pitch distances between successive tones. Contour in- formation is not specific to music, but is crucial to both speech prosody and melodic structure (Patel, Peretz, Tramo, & Labrecque, 1998), whereas exact interval information is specific to music. Contour detection appears to be a more universal process than interval 1 McMaster University, Canada, 2 Baycrest Centre for Geriatric Care, Canada, 3 University of Toronto © 2002 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 14:3, pp. 430–442

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Automatic and Controlled Processing of Melodic Contourand Interval Information Measured by Electrical
Brain Activity
Laurel J Trainor12 Kelly L McDonald2 and Claude Alain23
Abstract
amp Most work on how pitch is encoded in the auditory cortexhas focused on tonotopic (absolute) pitch maps Howevermelodic information is thought to be encoded in the brain intwo different lsquolsquorelative pitchrsquorsquo forms a domain-general contourcode (updown pattern of pitch changes) and a music-specificinterval code (exact pitch distances between notes) Event-related potentials were analyzed in nonmusicians from bothpassive and active oddball tasks where either the contour orthe interval of melodyndashfinal notes was occasionally altered Theoccasional deviant notes generated a right frontal positivity
peaking around 350 msec and a central parietal P3b peakingaround 580 msec that were present only when participantsfocused their attention on the auditory stimuli Both types ofmelodic information were encoded automatically in theabsence of absolute pitch cues as indexed by a mismatchnegativity wave recorded during the passive conditions Theresults indicate that even in the absence of musical trainingthe brain is set up to automatically encode music-specificmelodic information even when absolute pitch information isnot available amp
INTRODUCTION
How sound information is represented in the auditorycortex is not entirely understood It is clear that thereare tonotopic maps in the primary auditory cortexwhereby different pitches are represented in differentcortical areas in an orderly fashion (eg Woods Alho ampAllgazi 1993 Pantev Hoke Lutkenhoner amp Lehnertz1989 Pantev et al 1995) However the information inmelodies is not so much in the absolute frequencies orpitches of the tones but rather in the pitch relations orpitch distances between the tonesmdasha melody retains itsidentity whether rendered in a high or a low pitch rangeas long as the relative pitch relations are maintained Atpresent the stage of processing at which relative pitchinformation is extracted is not known We investigatethis question in a group of nonmusicians
There are reasons to expect that the auditory cortexmight be setup up to extract music-specific informationMusic is an integral part of every human society (WallinMerker amp Brown 2000) and there is evidence thatmelodic music-making has a long history in humancultural evolution (Kuneg amp Turk 2000) There is alsoevidence that across languages and cultures parents usemusic to communicate to young infants long before theyunderstand language (Trehub amp Trainor 1998) At the
same time music is often considered to be a lsquolsquofrillrsquorsquo anevolutionarily unimportant accident in the developmentof speech perception (eg Pinker 1994) Brain imagingstudies (PET and fMRI) have clearly shown that a largenumber of brain regions are involved in melodic pro-cessing including the temporal and frontal regions thesupplementary motor areas and the thalamus (egHalpern amp Zatorre 1999 Mirz et al 1999 Platel et al1997 Zatorre Halpern Perry Meyer amp Evans 1996)Although more studies implicate the right than the lefthemisphere in musical processing both hemispheresare clearly involved (eg Peretz amp Herbert 2000 Patel ampPeretz 1997 Zatorre 1984 Zatorre et al 1996) andmusical input stimulates the perceptual cognitive me-mory and emotional systems
Melodic information is thought to be encoded in twobasic forms (eg Liegeois-Chauvel Peretz Babaie La-guitton amp Chauvel 1998 Peretz amp Morais 1987 PeretzMorais amp Bertelson 1987 Peretz 1990 Peretz amp Babaie1992 Edworthy 1985 Dowling 1978 1982 Bever ampChiarello 1974) The contour code consists of the updown pattern of pitch changes without regard to the sizeof those changes The interval code consists of the exactpitch distances between successive tones Contour in-formation is not specific to music but is crucial to bothspeech prosody and melodic structure (Patel PeretzTramo amp Labrecque 1998) whereas exact intervalinformation is specific to music Contour detectionappears to be a more universal process than interval
1 McMaster University Canada 2 Baycrest Centre for GeriatricCare Canada 3University of Toronto
copy 2002 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 143 pp 430ndash442
detection in that both infants and musically untrainedadults readily detect when the contour changes buthave more difficulty perceiving changes between theintervals of consecutive notes in unfamiliar melodies(eg Trehub Trainor amp Unyk 1993 Bartlett amp Dowling1980 Dowling 1978 Cuddy amp Cohen 1976) Moreoveryoung children correctly reproduce the contour ofsimple melodies before they correctly reproduce theintervals (Dowling 1982) At the same time the intervalrepresentation is vitally important in musical structurebecause unlike the contour representation it allows theemergence of the specific scale structures musical keysand harmonic structures that distinguish different musi-cal systems and must be learned (eg Trainor amp Trehub1992 1994)
Basic sound features such as pitch duration andloudness are encoded automatically in the auditorycortex In humans this has been established primarilywith studies of the mismatch negativity (MMN) com-ponent of the event-related brain potential (ERP)(Naatanen Tervaniemi Sussman Paavilainen amp Win-kler 2001 Picton Alain Otten Ritter amp Achim 2000)This component is elicited in an oddball paradigm inwhich a repeating standard sound occasionally deviatesin pitch duration loudness timbre or location Thedeviant sounds generate activity in the auditory cortexbetween about 150 and 250 msec after stimulus onsetwhich is measured as an increased negativity at the frontof the scalp and an increased positivity at sites below theSylvian fissure in comparison to the frequent standardsounds (eg Picton et al 2000 Schroger 1998Naatanen 1992) MMN is typically recorded when sub-jects are not paying attention to the sounds as it canoverlap with and thus be obscured by componentsmodulated by attention Recent evidence indicates thatnot only are there circuits in the auditory cortex thatrespond automatically to differences in basic soundfeatures but that there are also circuits that respondto changes in sound patterns (Naatanen et al 2001Picton et al 2000) such as when one tone of a repeat-ing sequence is altered (eg Alain Cortese amp Picton1999) MMN even occurs to pattern deviants in which nonew tones are introduced such as when one note of thepattern is repeated (eg Alain Woods amp Ogawa 1994Alain Achim amp Woods 1999)
Of most interest in the present context are reportsthat the pitch contour of tone pairs (ie whether risingor falling) is encoded (ie MMN is seen) even when theabsolute pitch changes from instance to instance(Paavilainen Jaramillo amp Naatanen 1998 TervaniemiMaury amp Naatanen 1994 Sa arinen PaavilainenSchroger Tervaniemi amp Naatanen 1992) This suggeststhat lsquolsquorelativersquorsquo pitch contour is encoded automaticallyGiven the central role of contour in both speech andmusical prosody this is perhaps not surprising Asstated above however the essence of musical pitch isthe pitch intervalmdashthe exact pitch distance between
tones regardless of the absolute pitch of the tonesWhether the auditory cortex encodes pitch intervals inthe absence of attention has not been tested previouslyalthough it is a fundamental question in the study ofhow music is processed in the brain If nonmusiciansprocess changes in pitch interval automatically andwithout attention it would indicate that their auditorycortex is set up whether through innate processes orincidental musical exposure to process music-specificinformation
The neural correlates supporting contour and intervalprocessing have been examined recently in two separatestudies using ERPs but neither included a passive con-dition to allow the assessment of automatic processingUsing an oddball paradigm Schiavetto Cortese andAlain (1999) found that a larger N2 wave (peaking atabout 200 msec) was elicited for the identification ofcontour deviants than for interval deviants in nonmusi-cians The N2 complex consists of two components theN2a or MMN which occurs whenever the brain detectsan occasional deviant stimulus in a sequence of standardauditory stimuli whether or not the auditory stimuli aretask-relevant and the N2b which is also elicited bydeviant stimuli but only when they are task-relevant(Rugg amp Coles 1995) Because the ERPs in the Schia-vetto et al study were measured only during an activelistening task (ie participants attended to the stimuliand pressed buttons to identify the deviant stimuli) it isdifficult to evaluate whether the enhanced N2 wave forcontour over interval deviants reflects differences intask-relevant stimulus categorization (N2b) or in anautomatic change-detection process (MMN) Previousstudies have shown that the MMN amplitude increaseswith increasing deviant discriminability (Naatanen1992) Given that response times were slower for detect-ing interval than contour deviants in the Schiavetto et alstudy it is possible that the enhanced N2 for contourdeviants reflects a larger MMN wave associated withcontour than with interval deviants Alternatively it ispossible that neither contour nor interval deviants areprocessed automatically and the N2 difference foundreflects solely differences in controlled processes andstimulus categorization
Trainor Desjardins and Rockel (1999) comparedERPs from musicians and nonmusicians across contourand interval tasks using an oddball paradigm but exam-ined later ERP components In separate blocks of trialsparticipants responded as to whether or not there wasan occasional change (deviant) in the contour of a set ofmelodies with rising contours or whether or not therewas an occasional change (deviant) in the interval of amelody presented in transposition (ie starting at differ-ent pitches) The detection of both contour and intervalviolations was associated with a positive wave over thefrontal lobe peaking between 311 and 340 msec Thiswave referred to as a P3a preceded the P3b recordedover the parietal region which peaked between 383 and
Trainor McDonald and Alain 431
441 msec While the P3a and P3b recorded during thecontour task were similar in amplitude latency andtopography in musicians and nonmusicians they weredelayed and smaller in nonmusicians during the intervaltask These findings suggest that melodic contour isprocessed similarly in musicians and nonmusicians butthat discriminating changes in interval depends onmusical training
The frontal positivity reported by Trainor et al (1999)bears more investigation A frontal positivity is typicallyelicited either by occasional salient stimuli that occur inan unattended channel (eg Escera Alho Winkler ampNaatanen 1998 Alho Escera Diaz Yago amp Serra 1997Alho et al 1998 Squires Squires amp Hillyard 1975) or byoccasional salient stimuli in an attended channel that areto be ignored (ie not responded to) (eg Comercheroamp Polich 1998 Katayama amp Polich 1998 Holdstock ampRugg 1993 1995 Knight 1984) Thus this componentis thought to be related to the inadvertent capture ofattention by salient events and so it is of great interestto an investigation of automatic versus controlled pro-cesses for melody Although frontal P3 components havenot been studied often in relation to musical processingrobust frontal positivities have also been observed inone other study (Janata 1995) and frontal positivitiesare evident in the waveforms from other musical studiesalthough they were not analyzed (eg Besson amp Faita1995) This suggests that both contour and intervalchanges may trigger automatic attentional P3a pro-cesses However this interpretation needs to be treatedwith caution because the frontal positivities observed inthese musical studies were generated to occasionalattended stimuli that were targets not to stimuli thatwere to be ignored Furthermore in nonmusical pro-cessing P3as are not always found in all subjects (Polich1988 Squires et al 1975) and are strongest for deviantsthat are very different from the standard stimuli (egComerchero amp Polich 1998 Katayama amp Polich 1998Schroger amp Wolff 1998) On the other hand the targetsin the musical cases were sometimes rather subtly differ-ent from the standards (eg the interval task of Trainoret al 1999) again suggesting that the frontal positivitiesin the musical case may not map directly onto P3asdescribed in other literatures In the present study weexamine the frontal positivity in both active and passivemusical tasks in order to determine whether it functionslike the P3a described for other stimuli or whether itreflects some other process
The final ERP component of interest in the presentinvestigation is the P3b the large parietally centeredpeak that is found to infrequent target stimuli (iestimuli requiring a response) in an attended channelbut not to infrequent stimuli in an unattended channelThis component is thought to be related to consciousstimulus evaluation and the updating of working mem-ory (eg see Katayama amp Polich 1998 Verleger 1997Stuss Picton Cerri Leech amp Stethem 1992 Donchin amp
Coles 1988 Johnson-Laird 1988) and has been found inmany studies requiring the detection of various deviantsin musical stimuli (eg Schiavetto et al 1999 Trainoret al 1999 Besson amp Faotildeta 1995 Besson 1998 Janata1995 Cohen Granot Pratt amp Barneah 1993 Levett ampMartin 1992 Paller McCarthy amp Wood 1992 Besson ampMacar 1987)
In order to examine both early and later corticalcomponents related to the processing of pitch contourand interval information we used the set of five-notecontour melodies from the Trainor et al (1999) studyEach of these melodies started on a different pitch andcontained different pitch intervals but all melodies hada rising contour (see Figure 1) On 20 of trials the lasttone was changed so that the final interval descendedrather than ascended Thus this task cannot be done byrelying on an absolute pitch representation of themelodies For the interval task we created a melodyconsisting of the first five notes of the rising major scale(see Figure 1) On different trials this melody wasrandomly transposed to start on any of the 12 notes ofthe Western scale On 20 of trials the final note of themelody was changed to form a different final intervalsuch that the contour always ascended As in the con-tour task this interval task cannot be performed usingan absolute pitch representation
Each participant completed both an active (buttonpressed as to whether a standard or deviant melody waspresented) and a passive (read a book while the melo-dies were presented) version of each of the contour andinterval tasks The passive task allowed us to examinewhether contour and interval information are encodedautomatically (presence of MMN) as well as whether thefrontal P3 reported in the Trainor et al (1999) studyindexes automatic or controlled processes of musicalinformation
In summary we investigated automatic and con-trolled processes in sensory and working memoryrepresentations of melodic contour and interval infor-mation On the basis of previous simple two-tonecontour (Paavilainen et al 1998 Tervaniemi et al1994 Saarinen et al 1992) and the domain-generalnature of contour processing we expected contourviolations to elicit MMN However because intervalprocessing depends to a much greater extent onmusical experience (eg Peretz amp Babaie 1992 Dow-ling 1982 Bever amp Chiarello 1974) and tonality(Edworthy 1985) and because it is thought to bespecialized to the musical system (Patel amp Peretz1997) an automatic relative-pitch encoding of intervalinformation as reflected by MMN would be moresurprising in nonmusicians The absence of an MMNresponse in nonmusicians to interval violations wouldsuggest that automatic interval processing does notdevelop universally and would be consistent withthe idea that musical processing is not importantenough for the allocation of extensive resources for
432 Journal of Cognitive Neuroscience Volume 14 Number 3
automatic extraction of musical features at sensorylevels of auditory processing On the other hand ifthe nonmusically trained person extracts intervalinformation automatically in the absence of absolutepitch information this would suggest that the brainplaces high value on music-specific pitch patterninformation
RESULTS
Performance
The tasks were relatively easy with participants correctlyidentifying the melody as standard or deviant in morethan 90 of the trials in both contour and interval tasks
(Table 1) Although performance was high in both casesit was significantly better for the contour than for theinterval task F(110) = 609 p lt 05 The difference inreaction times (RTs) between the contour and intervaltasks was not significant although the trend was forshorter response time in the former case (Table 1) Inthe interval task the final note could either be in key orout of key There were no significant differences ineither accuracy or RT between these conditions
ERP Results
Passive Listening
Figure 2 shows group mean ERPs during passive listen-ing for the contour and interval tasks In both tasksstandards and deviants generated an N1 An analysis ofvariance (ANOVA) at Fz with task (contour interval)and stimulus type (standard deviant) as factors re-vealed that the N1 was later in deviant than in standardstimuli [main effect of stimulus type F(110) = 617 p lt05] and that this difference was larger in the contour(means of 127 msec SD = 55 and 117 msec SD =158 respectively) than in the interval (means of 122msec SD = 126 and 121 msec SD = 129 respec-tively) task [Task pound Stimulus Type interaction F(110)= 675 p lt 05]
Using the mean amplitude between 95 and 135 msecas the dependent measure a similar ANOVA revealedthat the N1 was larger in the contour than in the intervaltask F(110) = 1553 p lt 01 and that it was largerfor deviant than for standard stimuli F(110) = 1117p lt 01 The latter effect is likely due to overlap betweenthe N1 and MMN (see below) components elicited byoddball stimuli
Contour and interval deviant stimuli generated anMMN that was maximal at the frontal sites and reversedpolarity at the mastoids (Figure 3) The MMN recordedat the midline electrode (Fz) was delayed in the interval(mean = 180 msec SD = 270) compared with thecontour (mean = 150 msec SD = 227) task F(110) =1077 p lt 01 Within the interval task the MMN wasearlier for the whole tone than for the semitonechanges F(110) = 989 p lt 01 (means of 1466 and1906 msec respectively)
The mean amplitude was calculated for 40-msecwindows around the contour and interval means forstandard and deviant stimuli There were no differences
Table 1 Behavioral Performance for the Contour and IntervalTasks
Contour Task Interval Task
Correct 9496 (444) 9175 (703)
RT (msec) 570 (113) 604 (80)
Standard deviations are shown in parentheses
Figure 1 The contour (upper panel) and interval (lower panel)stimuli In each case the first four notes of the melodies (commonsequence) are shown followed by the standard and deviant (oddball)terminal notes In the contour case interval size changes from melodyto melody but the standard terminal notes always rise whereas theoddball terminal notes always fall In the interval case deviant terminalnotes either go outside the key of the common sequence (secondndashlastbar) or remain in the key of the common sequence (last bar) but thecontour is always rising
Trainor McDonald and Alain 433
in amplitude between the whole-tone and semitonechanges in the interval task so the data were collapsedacross this factor An ANOVA with task (contour inter-
val) and stimulus type (standard deviant) revealed thatdeviants were indeed more negative than standardsF(110) = 5083 p lt 001 and that contour was more
Figure 2 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the passive contour andinterval tasks In this and thesubsequent figures the thickbar shows the onset of the finalmelody note
434 Journal of Cognitive Neuroscience Volume 14 Number 3
negative than interval F(110) = 2794 p lt 001 Therewas no interaction between task and stimulus typehowever indicating that the MMN was of comparablesize in the contour and interval tasks (see Figure 3)
An examination of the head maps (Figure 4) showsthat the MMN was fronto-centrally distributed over thescalp in the contour task while it was greater over theright hemisphere in the interval task However therewere no significant topological differences betweencontour and interval MMN as revealed in analyses usingall electrodes as well as analyses using only midline oronly lateral line electrodes
As expected no P3b was evident in the passive listen-ing tasks of either contour or interval Interestinglythere was also no evidence of a frontal positivity in thepassive listening task
Active Listening
Figure 5 shows group mean ERPs during active listeningfor the contour and interval tasks Because no significantdifferences were found for any ERP components be-tween the whole tone and semitone conditions in theinterval task the following analyses are collapsed acrossthis factor
In both contour and interval tasks standards anddeviants generated an N1 peaking at 119 msec (SD =127) poststimulus An ANOVA with task (contour
interval) and stimulus type (standard deviant) as fac-tors showed that the N1 measured at Fz was larger(mean amplitude between 95 and 135 msec post stim-ulus) in the contour than in the interval task F(110) =886 p lt 02 and larger for deviants than for standardsF(1 10) = 920 p lt 02 The latter effect is probablydue to the fact that the N2 wave elicited by the oddballstimuli was partly superimposed on the N1 deflection(see below)
Figure 3 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during pas-sive listening
Contour Interval
Figure 4 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for the peak MMN during contour(150 msec) and interval (180 msec) tasks Isocontour spacing is 01 mV
Trainor McDonald and Alain 435
The ERPs to deviant stimuli were characterized by anN2 wave that was superimposed on the P2 deflectionThe N2 wave was maximal over the fronto-central regionso the following analyses were done at Fz The N2
peaked earlier for the contour than for the intervaltask F(110) = 1791 p lt 002 The mean amplitudecalculated in 40-msec windows around the grand ave-rage peaks for contour (170ndash210 msec) and interval
Figure 5 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the active contour and in-terval tasks
436 Journal of Cognitive Neuroscience Volume 14 Number 3
(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

detection in that both infants and musically untrainedadults readily detect when the contour changes buthave more difficulty perceiving changes between theintervals of consecutive notes in unfamiliar melodies(eg Trehub Trainor amp Unyk 1993 Bartlett amp Dowling1980 Dowling 1978 Cuddy amp Cohen 1976) Moreoveryoung children correctly reproduce the contour ofsimple melodies before they correctly reproduce theintervals (Dowling 1982) At the same time the intervalrepresentation is vitally important in musical structurebecause unlike the contour representation it allows theemergence of the specific scale structures musical keysand harmonic structures that distinguish different musi-cal systems and must be learned (eg Trainor amp Trehub1992 1994)
Basic sound features such as pitch duration andloudness are encoded automatically in the auditorycortex In humans this has been established primarilywith studies of the mismatch negativity (MMN) com-ponent of the event-related brain potential (ERP)(Naatanen Tervaniemi Sussman Paavilainen amp Win-kler 2001 Picton Alain Otten Ritter amp Achim 2000)This component is elicited in an oddball paradigm inwhich a repeating standard sound occasionally deviatesin pitch duration loudness timbre or location Thedeviant sounds generate activity in the auditory cortexbetween about 150 and 250 msec after stimulus onsetwhich is measured as an increased negativity at the frontof the scalp and an increased positivity at sites below theSylvian fissure in comparison to the frequent standardsounds (eg Picton et al 2000 Schroger 1998Naatanen 1992) MMN is typically recorded when sub-jects are not paying attention to the sounds as it canoverlap with and thus be obscured by componentsmodulated by attention Recent evidence indicates thatnot only are there circuits in the auditory cortex thatrespond automatically to differences in basic soundfeatures but that there are also circuits that respondto changes in sound patterns (Naatanen et al 2001Picton et al 2000) such as when one tone of a repeat-ing sequence is altered (eg Alain Cortese amp Picton1999) MMN even occurs to pattern deviants in which nonew tones are introduced such as when one note of thepattern is repeated (eg Alain Woods amp Ogawa 1994Alain Achim amp Woods 1999)
Of most interest in the present context are reportsthat the pitch contour of tone pairs (ie whether risingor falling) is encoded (ie MMN is seen) even when theabsolute pitch changes from instance to instance(Paavilainen Jaramillo amp Naatanen 1998 TervaniemiMaury amp Naatanen 1994 Sa arinen PaavilainenSchroger Tervaniemi amp Naatanen 1992) This suggeststhat lsquolsquorelativersquorsquo pitch contour is encoded automaticallyGiven the central role of contour in both speech andmusical prosody this is perhaps not surprising Asstated above however the essence of musical pitch isthe pitch intervalmdashthe exact pitch distance between
tones regardless of the absolute pitch of the tonesWhether the auditory cortex encodes pitch intervals inthe absence of attention has not been tested previouslyalthough it is a fundamental question in the study ofhow music is processed in the brain If nonmusiciansprocess changes in pitch interval automatically andwithout attention it would indicate that their auditorycortex is set up whether through innate processes orincidental musical exposure to process music-specificinformation
The neural correlates supporting contour and intervalprocessing have been examined recently in two separatestudies using ERPs but neither included a passive con-dition to allow the assessment of automatic processingUsing an oddball paradigm Schiavetto Cortese andAlain (1999) found that a larger N2 wave (peaking atabout 200 msec) was elicited for the identification ofcontour deviants than for interval deviants in nonmusi-cians The N2 complex consists of two components theN2a or MMN which occurs whenever the brain detectsan occasional deviant stimulus in a sequence of standardauditory stimuli whether or not the auditory stimuli aretask-relevant and the N2b which is also elicited bydeviant stimuli but only when they are task-relevant(Rugg amp Coles 1995) Because the ERPs in the Schia-vetto et al study were measured only during an activelistening task (ie participants attended to the stimuliand pressed buttons to identify the deviant stimuli) it isdifficult to evaluate whether the enhanced N2 wave forcontour over interval deviants reflects differences intask-relevant stimulus categorization (N2b) or in anautomatic change-detection process (MMN) Previousstudies have shown that the MMN amplitude increaseswith increasing deviant discriminability (Naatanen1992) Given that response times were slower for detect-ing interval than contour deviants in the Schiavetto et alstudy it is possible that the enhanced N2 for contourdeviants reflects a larger MMN wave associated withcontour than with interval deviants Alternatively it ispossible that neither contour nor interval deviants areprocessed automatically and the N2 difference foundreflects solely differences in controlled processes andstimulus categorization
Trainor Desjardins and Rockel (1999) comparedERPs from musicians and nonmusicians across contourand interval tasks using an oddball paradigm but exam-ined later ERP components In separate blocks of trialsparticipants responded as to whether or not there wasan occasional change (deviant) in the contour of a set ofmelodies with rising contours or whether or not therewas an occasional change (deviant) in the interval of amelody presented in transposition (ie starting at differ-ent pitches) The detection of both contour and intervalviolations was associated with a positive wave over thefrontal lobe peaking between 311 and 340 msec Thiswave referred to as a P3a preceded the P3b recordedover the parietal region which peaked between 383 and
Trainor McDonald and Alain 431
441 msec While the P3a and P3b recorded during thecontour task were similar in amplitude latency andtopography in musicians and nonmusicians they weredelayed and smaller in nonmusicians during the intervaltask These findings suggest that melodic contour isprocessed similarly in musicians and nonmusicians butthat discriminating changes in interval depends onmusical training
The frontal positivity reported by Trainor et al (1999)bears more investigation A frontal positivity is typicallyelicited either by occasional salient stimuli that occur inan unattended channel (eg Escera Alho Winkler ampNaatanen 1998 Alho Escera Diaz Yago amp Serra 1997Alho et al 1998 Squires Squires amp Hillyard 1975) or byoccasional salient stimuli in an attended channel that areto be ignored (ie not responded to) (eg Comercheroamp Polich 1998 Katayama amp Polich 1998 Holdstock ampRugg 1993 1995 Knight 1984) Thus this componentis thought to be related to the inadvertent capture ofattention by salient events and so it is of great interestto an investigation of automatic versus controlled pro-cesses for melody Although frontal P3 components havenot been studied often in relation to musical processingrobust frontal positivities have also been observed inone other study (Janata 1995) and frontal positivitiesare evident in the waveforms from other musical studiesalthough they were not analyzed (eg Besson amp Faita1995) This suggests that both contour and intervalchanges may trigger automatic attentional P3a pro-cesses However this interpretation needs to be treatedwith caution because the frontal positivities observed inthese musical studies were generated to occasionalattended stimuli that were targets not to stimuli thatwere to be ignored Furthermore in nonmusical pro-cessing P3as are not always found in all subjects (Polich1988 Squires et al 1975) and are strongest for deviantsthat are very different from the standard stimuli (egComerchero amp Polich 1998 Katayama amp Polich 1998Schroger amp Wolff 1998) On the other hand the targetsin the musical cases were sometimes rather subtly differ-ent from the standards (eg the interval task of Trainoret al 1999) again suggesting that the frontal positivitiesin the musical case may not map directly onto P3asdescribed in other literatures In the present study weexamine the frontal positivity in both active and passivemusical tasks in order to determine whether it functionslike the P3a described for other stimuli or whether itreflects some other process
The final ERP component of interest in the presentinvestigation is the P3b the large parietally centeredpeak that is found to infrequent target stimuli (iestimuli requiring a response) in an attended channelbut not to infrequent stimuli in an unattended channelThis component is thought to be related to consciousstimulus evaluation and the updating of working mem-ory (eg see Katayama amp Polich 1998 Verleger 1997Stuss Picton Cerri Leech amp Stethem 1992 Donchin amp
Coles 1988 Johnson-Laird 1988) and has been found inmany studies requiring the detection of various deviantsin musical stimuli (eg Schiavetto et al 1999 Trainoret al 1999 Besson amp Faotildeta 1995 Besson 1998 Janata1995 Cohen Granot Pratt amp Barneah 1993 Levett ampMartin 1992 Paller McCarthy amp Wood 1992 Besson ampMacar 1987)
In order to examine both early and later corticalcomponents related to the processing of pitch contourand interval information we used the set of five-notecontour melodies from the Trainor et al (1999) studyEach of these melodies started on a different pitch andcontained different pitch intervals but all melodies hada rising contour (see Figure 1) On 20 of trials the lasttone was changed so that the final interval descendedrather than ascended Thus this task cannot be done byrelying on an absolute pitch representation of themelodies For the interval task we created a melodyconsisting of the first five notes of the rising major scale(see Figure 1) On different trials this melody wasrandomly transposed to start on any of the 12 notes ofthe Western scale On 20 of trials the final note of themelody was changed to form a different final intervalsuch that the contour always ascended As in the con-tour task this interval task cannot be performed usingan absolute pitch representation
Each participant completed both an active (buttonpressed as to whether a standard or deviant melody waspresented) and a passive (read a book while the melo-dies were presented) version of each of the contour andinterval tasks The passive task allowed us to examinewhether contour and interval information are encodedautomatically (presence of MMN) as well as whether thefrontal P3 reported in the Trainor et al (1999) studyindexes automatic or controlled processes of musicalinformation
In summary we investigated automatic and con-trolled processes in sensory and working memoryrepresentations of melodic contour and interval infor-mation On the basis of previous simple two-tonecontour (Paavilainen et al 1998 Tervaniemi et al1994 Saarinen et al 1992) and the domain-generalnature of contour processing we expected contourviolations to elicit MMN However because intervalprocessing depends to a much greater extent onmusical experience (eg Peretz amp Babaie 1992 Dow-ling 1982 Bever amp Chiarello 1974) and tonality(Edworthy 1985) and because it is thought to bespecialized to the musical system (Patel amp Peretz1997) an automatic relative-pitch encoding of intervalinformation as reflected by MMN would be moresurprising in nonmusicians The absence of an MMNresponse in nonmusicians to interval violations wouldsuggest that automatic interval processing does notdevelop universally and would be consistent withthe idea that musical processing is not importantenough for the allocation of extensive resources for
432 Journal of Cognitive Neuroscience Volume 14 Number 3
automatic extraction of musical features at sensorylevels of auditory processing On the other hand ifthe nonmusically trained person extracts intervalinformation automatically in the absence of absolutepitch information this would suggest that the brainplaces high value on music-specific pitch patterninformation
RESULTS
Performance
The tasks were relatively easy with participants correctlyidentifying the melody as standard or deviant in morethan 90 of the trials in both contour and interval tasks
(Table 1) Although performance was high in both casesit was significantly better for the contour than for theinterval task F(110) = 609 p lt 05 The difference inreaction times (RTs) between the contour and intervaltasks was not significant although the trend was forshorter response time in the former case (Table 1) Inthe interval task the final note could either be in key orout of key There were no significant differences ineither accuracy or RT between these conditions
ERP Results
Passive Listening
Figure 2 shows group mean ERPs during passive listen-ing for the contour and interval tasks In both tasksstandards and deviants generated an N1 An analysis ofvariance (ANOVA) at Fz with task (contour interval)and stimulus type (standard deviant) as factors re-vealed that the N1 was later in deviant than in standardstimuli [main effect of stimulus type F(110) = 617 p lt05] and that this difference was larger in the contour(means of 127 msec SD = 55 and 117 msec SD =158 respectively) than in the interval (means of 122msec SD = 126 and 121 msec SD = 129 respec-tively) task [Task pound Stimulus Type interaction F(110)= 675 p lt 05]
Using the mean amplitude between 95 and 135 msecas the dependent measure a similar ANOVA revealedthat the N1 was larger in the contour than in the intervaltask F(110) = 1553 p lt 01 and that it was largerfor deviant than for standard stimuli F(110) = 1117p lt 01 The latter effect is likely due to overlap betweenthe N1 and MMN (see below) components elicited byoddball stimuli
Contour and interval deviant stimuli generated anMMN that was maximal at the frontal sites and reversedpolarity at the mastoids (Figure 3) The MMN recordedat the midline electrode (Fz) was delayed in the interval(mean = 180 msec SD = 270) compared with thecontour (mean = 150 msec SD = 227) task F(110) =1077 p lt 01 Within the interval task the MMN wasearlier for the whole tone than for the semitonechanges F(110) = 989 p lt 01 (means of 1466 and1906 msec respectively)
The mean amplitude was calculated for 40-msecwindows around the contour and interval means forstandard and deviant stimuli There were no differences
Table 1 Behavioral Performance for the Contour and IntervalTasks
Contour Task Interval Task
Correct 9496 (444) 9175 (703)
RT (msec) 570 (113) 604 (80)
Standard deviations are shown in parentheses
Figure 1 The contour (upper panel) and interval (lower panel)stimuli In each case the first four notes of the melodies (commonsequence) are shown followed by the standard and deviant (oddball)terminal notes In the contour case interval size changes from melodyto melody but the standard terminal notes always rise whereas theoddball terminal notes always fall In the interval case deviant terminalnotes either go outside the key of the common sequence (secondndashlastbar) or remain in the key of the common sequence (last bar) but thecontour is always rising
Trainor McDonald and Alain 433
in amplitude between the whole-tone and semitonechanges in the interval task so the data were collapsedacross this factor An ANOVA with task (contour inter-
val) and stimulus type (standard deviant) revealed thatdeviants were indeed more negative than standardsF(110) = 5083 p lt 001 and that contour was more
Figure 2 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the passive contour andinterval tasks In this and thesubsequent figures the thickbar shows the onset of the finalmelody note
434 Journal of Cognitive Neuroscience Volume 14 Number 3
negative than interval F(110) = 2794 p lt 001 Therewas no interaction between task and stimulus typehowever indicating that the MMN was of comparablesize in the contour and interval tasks (see Figure 3)
An examination of the head maps (Figure 4) showsthat the MMN was fronto-centrally distributed over thescalp in the contour task while it was greater over theright hemisphere in the interval task However therewere no significant topological differences betweencontour and interval MMN as revealed in analyses usingall electrodes as well as analyses using only midline oronly lateral line electrodes
As expected no P3b was evident in the passive listen-ing tasks of either contour or interval Interestinglythere was also no evidence of a frontal positivity in thepassive listening task
Active Listening
Figure 5 shows group mean ERPs during active listeningfor the contour and interval tasks Because no significantdifferences were found for any ERP components be-tween the whole tone and semitone conditions in theinterval task the following analyses are collapsed acrossthis factor
In both contour and interval tasks standards anddeviants generated an N1 peaking at 119 msec (SD =127) poststimulus An ANOVA with task (contour
interval) and stimulus type (standard deviant) as fac-tors showed that the N1 measured at Fz was larger(mean amplitude between 95 and 135 msec post stim-ulus) in the contour than in the interval task F(110) =886 p lt 02 and larger for deviants than for standardsF(1 10) = 920 p lt 02 The latter effect is probablydue to the fact that the N2 wave elicited by the oddballstimuli was partly superimposed on the N1 deflection(see below)
Figure 3 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during pas-sive listening
Contour Interval
Figure 4 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for the peak MMN during contour(150 msec) and interval (180 msec) tasks Isocontour spacing is 01 mV
Trainor McDonald and Alain 435
The ERPs to deviant stimuli were characterized by anN2 wave that was superimposed on the P2 deflectionThe N2 wave was maximal over the fronto-central regionso the following analyses were done at Fz The N2
peaked earlier for the contour than for the intervaltask F(110) = 1791 p lt 002 The mean amplitudecalculated in 40-msec windows around the grand ave-rage peaks for contour (170ndash210 msec) and interval
Figure 5 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the active contour and in-terval tasks
436 Journal of Cognitive Neuroscience Volume 14 Number 3
(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

441 msec While the P3a and P3b recorded during thecontour task were similar in amplitude latency andtopography in musicians and nonmusicians they weredelayed and smaller in nonmusicians during the intervaltask These findings suggest that melodic contour isprocessed similarly in musicians and nonmusicians butthat discriminating changes in interval depends onmusical training
The frontal positivity reported by Trainor et al (1999)bears more investigation A frontal positivity is typicallyelicited either by occasional salient stimuli that occur inan unattended channel (eg Escera Alho Winkler ampNaatanen 1998 Alho Escera Diaz Yago amp Serra 1997Alho et al 1998 Squires Squires amp Hillyard 1975) or byoccasional salient stimuli in an attended channel that areto be ignored (ie not responded to) (eg Comercheroamp Polich 1998 Katayama amp Polich 1998 Holdstock ampRugg 1993 1995 Knight 1984) Thus this componentis thought to be related to the inadvertent capture ofattention by salient events and so it is of great interestto an investigation of automatic versus controlled pro-cesses for melody Although frontal P3 components havenot been studied often in relation to musical processingrobust frontal positivities have also been observed inone other study (Janata 1995) and frontal positivitiesare evident in the waveforms from other musical studiesalthough they were not analyzed (eg Besson amp Faita1995) This suggests that both contour and intervalchanges may trigger automatic attentional P3a pro-cesses However this interpretation needs to be treatedwith caution because the frontal positivities observed inthese musical studies were generated to occasionalattended stimuli that were targets not to stimuli thatwere to be ignored Furthermore in nonmusical pro-cessing P3as are not always found in all subjects (Polich1988 Squires et al 1975) and are strongest for deviantsthat are very different from the standard stimuli (egComerchero amp Polich 1998 Katayama amp Polich 1998Schroger amp Wolff 1998) On the other hand the targetsin the musical cases were sometimes rather subtly differ-ent from the standards (eg the interval task of Trainoret al 1999) again suggesting that the frontal positivitiesin the musical case may not map directly onto P3asdescribed in other literatures In the present study weexamine the frontal positivity in both active and passivemusical tasks in order to determine whether it functionslike the P3a described for other stimuli or whether itreflects some other process
The final ERP component of interest in the presentinvestigation is the P3b the large parietally centeredpeak that is found to infrequent target stimuli (iestimuli requiring a response) in an attended channelbut not to infrequent stimuli in an unattended channelThis component is thought to be related to consciousstimulus evaluation and the updating of working mem-ory (eg see Katayama amp Polich 1998 Verleger 1997Stuss Picton Cerri Leech amp Stethem 1992 Donchin amp
Coles 1988 Johnson-Laird 1988) and has been found inmany studies requiring the detection of various deviantsin musical stimuli (eg Schiavetto et al 1999 Trainoret al 1999 Besson amp Faotildeta 1995 Besson 1998 Janata1995 Cohen Granot Pratt amp Barneah 1993 Levett ampMartin 1992 Paller McCarthy amp Wood 1992 Besson ampMacar 1987)
In order to examine both early and later corticalcomponents related to the processing of pitch contourand interval information we used the set of five-notecontour melodies from the Trainor et al (1999) studyEach of these melodies started on a different pitch andcontained different pitch intervals but all melodies hada rising contour (see Figure 1) On 20 of trials the lasttone was changed so that the final interval descendedrather than ascended Thus this task cannot be done byrelying on an absolute pitch representation of themelodies For the interval task we created a melodyconsisting of the first five notes of the rising major scale(see Figure 1) On different trials this melody wasrandomly transposed to start on any of the 12 notes ofthe Western scale On 20 of trials the final note of themelody was changed to form a different final intervalsuch that the contour always ascended As in the con-tour task this interval task cannot be performed usingan absolute pitch representation
Each participant completed both an active (buttonpressed as to whether a standard or deviant melody waspresented) and a passive (read a book while the melo-dies were presented) version of each of the contour andinterval tasks The passive task allowed us to examinewhether contour and interval information are encodedautomatically (presence of MMN) as well as whether thefrontal P3 reported in the Trainor et al (1999) studyindexes automatic or controlled processes of musicalinformation
In summary we investigated automatic and con-trolled processes in sensory and working memoryrepresentations of melodic contour and interval infor-mation On the basis of previous simple two-tonecontour (Paavilainen et al 1998 Tervaniemi et al1994 Saarinen et al 1992) and the domain-generalnature of contour processing we expected contourviolations to elicit MMN However because intervalprocessing depends to a much greater extent onmusical experience (eg Peretz amp Babaie 1992 Dow-ling 1982 Bever amp Chiarello 1974) and tonality(Edworthy 1985) and because it is thought to bespecialized to the musical system (Patel amp Peretz1997) an automatic relative-pitch encoding of intervalinformation as reflected by MMN would be moresurprising in nonmusicians The absence of an MMNresponse in nonmusicians to interval violations wouldsuggest that automatic interval processing does notdevelop universally and would be consistent withthe idea that musical processing is not importantenough for the allocation of extensive resources for
432 Journal of Cognitive Neuroscience Volume 14 Number 3
automatic extraction of musical features at sensorylevels of auditory processing On the other hand ifthe nonmusically trained person extracts intervalinformation automatically in the absence of absolutepitch information this would suggest that the brainplaces high value on music-specific pitch patterninformation
RESULTS
Performance
The tasks were relatively easy with participants correctlyidentifying the melody as standard or deviant in morethan 90 of the trials in both contour and interval tasks
(Table 1) Although performance was high in both casesit was significantly better for the contour than for theinterval task F(110) = 609 p lt 05 The difference inreaction times (RTs) between the contour and intervaltasks was not significant although the trend was forshorter response time in the former case (Table 1) Inthe interval task the final note could either be in key orout of key There were no significant differences ineither accuracy or RT between these conditions
ERP Results
Passive Listening
Figure 2 shows group mean ERPs during passive listen-ing for the contour and interval tasks In both tasksstandards and deviants generated an N1 An analysis ofvariance (ANOVA) at Fz with task (contour interval)and stimulus type (standard deviant) as factors re-vealed that the N1 was later in deviant than in standardstimuli [main effect of stimulus type F(110) = 617 p lt05] and that this difference was larger in the contour(means of 127 msec SD = 55 and 117 msec SD =158 respectively) than in the interval (means of 122msec SD = 126 and 121 msec SD = 129 respec-tively) task [Task pound Stimulus Type interaction F(110)= 675 p lt 05]
Using the mean amplitude between 95 and 135 msecas the dependent measure a similar ANOVA revealedthat the N1 was larger in the contour than in the intervaltask F(110) = 1553 p lt 01 and that it was largerfor deviant than for standard stimuli F(110) = 1117p lt 01 The latter effect is likely due to overlap betweenthe N1 and MMN (see below) components elicited byoddball stimuli
Contour and interval deviant stimuli generated anMMN that was maximal at the frontal sites and reversedpolarity at the mastoids (Figure 3) The MMN recordedat the midline electrode (Fz) was delayed in the interval(mean = 180 msec SD = 270) compared with thecontour (mean = 150 msec SD = 227) task F(110) =1077 p lt 01 Within the interval task the MMN wasearlier for the whole tone than for the semitonechanges F(110) = 989 p lt 01 (means of 1466 and1906 msec respectively)
The mean amplitude was calculated for 40-msecwindows around the contour and interval means forstandard and deviant stimuli There were no differences
Table 1 Behavioral Performance for the Contour and IntervalTasks
Contour Task Interval Task
Correct 9496 (444) 9175 (703)
RT (msec) 570 (113) 604 (80)
Standard deviations are shown in parentheses
Figure 1 The contour (upper panel) and interval (lower panel)stimuli In each case the first four notes of the melodies (commonsequence) are shown followed by the standard and deviant (oddball)terminal notes In the contour case interval size changes from melodyto melody but the standard terminal notes always rise whereas theoddball terminal notes always fall In the interval case deviant terminalnotes either go outside the key of the common sequence (secondndashlastbar) or remain in the key of the common sequence (last bar) but thecontour is always rising
Trainor McDonald and Alain 433
in amplitude between the whole-tone and semitonechanges in the interval task so the data were collapsedacross this factor An ANOVA with task (contour inter-
val) and stimulus type (standard deviant) revealed thatdeviants were indeed more negative than standardsF(110) = 5083 p lt 001 and that contour was more
Figure 2 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the passive contour andinterval tasks In this and thesubsequent figures the thickbar shows the onset of the finalmelody note
434 Journal of Cognitive Neuroscience Volume 14 Number 3
negative than interval F(110) = 2794 p lt 001 Therewas no interaction between task and stimulus typehowever indicating that the MMN was of comparablesize in the contour and interval tasks (see Figure 3)
An examination of the head maps (Figure 4) showsthat the MMN was fronto-centrally distributed over thescalp in the contour task while it was greater over theright hemisphere in the interval task However therewere no significant topological differences betweencontour and interval MMN as revealed in analyses usingall electrodes as well as analyses using only midline oronly lateral line electrodes
As expected no P3b was evident in the passive listen-ing tasks of either contour or interval Interestinglythere was also no evidence of a frontal positivity in thepassive listening task
Active Listening
Figure 5 shows group mean ERPs during active listeningfor the contour and interval tasks Because no significantdifferences were found for any ERP components be-tween the whole tone and semitone conditions in theinterval task the following analyses are collapsed acrossthis factor
In both contour and interval tasks standards anddeviants generated an N1 peaking at 119 msec (SD =127) poststimulus An ANOVA with task (contour
interval) and stimulus type (standard deviant) as fac-tors showed that the N1 measured at Fz was larger(mean amplitude between 95 and 135 msec post stim-ulus) in the contour than in the interval task F(110) =886 p lt 02 and larger for deviants than for standardsF(1 10) = 920 p lt 02 The latter effect is probablydue to the fact that the N2 wave elicited by the oddballstimuli was partly superimposed on the N1 deflection(see below)
Figure 3 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during pas-sive listening
Contour Interval
Figure 4 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for the peak MMN during contour(150 msec) and interval (180 msec) tasks Isocontour spacing is 01 mV
Trainor McDonald and Alain 435
The ERPs to deviant stimuli were characterized by anN2 wave that was superimposed on the P2 deflectionThe N2 wave was maximal over the fronto-central regionso the following analyses were done at Fz The N2
peaked earlier for the contour than for the intervaltask F(110) = 1791 p lt 002 The mean amplitudecalculated in 40-msec windows around the grand ave-rage peaks for contour (170ndash210 msec) and interval
Figure 5 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the active contour and in-terval tasks
436 Journal of Cognitive Neuroscience Volume 14 Number 3
(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

automatic extraction of musical features at sensorylevels of auditory processing On the other hand ifthe nonmusically trained person extracts intervalinformation automatically in the absence of absolutepitch information this would suggest that the brainplaces high value on music-specific pitch patterninformation
RESULTS
Performance
The tasks were relatively easy with participants correctlyidentifying the melody as standard or deviant in morethan 90 of the trials in both contour and interval tasks
(Table 1) Although performance was high in both casesit was significantly better for the contour than for theinterval task F(110) = 609 p lt 05 The difference inreaction times (RTs) between the contour and intervaltasks was not significant although the trend was forshorter response time in the former case (Table 1) Inthe interval task the final note could either be in key orout of key There were no significant differences ineither accuracy or RT between these conditions
ERP Results
Passive Listening
Figure 2 shows group mean ERPs during passive listen-ing for the contour and interval tasks In both tasksstandards and deviants generated an N1 An analysis ofvariance (ANOVA) at Fz with task (contour interval)and stimulus type (standard deviant) as factors re-vealed that the N1 was later in deviant than in standardstimuli [main effect of stimulus type F(110) = 617 p lt05] and that this difference was larger in the contour(means of 127 msec SD = 55 and 117 msec SD =158 respectively) than in the interval (means of 122msec SD = 126 and 121 msec SD = 129 respec-tively) task [Task pound Stimulus Type interaction F(110)= 675 p lt 05]
Using the mean amplitude between 95 and 135 msecas the dependent measure a similar ANOVA revealedthat the N1 was larger in the contour than in the intervaltask F(110) = 1553 p lt 01 and that it was largerfor deviant than for standard stimuli F(110) = 1117p lt 01 The latter effect is likely due to overlap betweenthe N1 and MMN (see below) components elicited byoddball stimuli
Contour and interval deviant stimuli generated anMMN that was maximal at the frontal sites and reversedpolarity at the mastoids (Figure 3) The MMN recordedat the midline electrode (Fz) was delayed in the interval(mean = 180 msec SD = 270) compared with thecontour (mean = 150 msec SD = 227) task F(110) =1077 p lt 01 Within the interval task the MMN wasearlier for the whole tone than for the semitonechanges F(110) = 989 p lt 01 (means of 1466 and1906 msec respectively)
The mean amplitude was calculated for 40-msecwindows around the contour and interval means forstandard and deviant stimuli There were no differences
Table 1 Behavioral Performance for the Contour and IntervalTasks
Contour Task Interval Task
Correct 9496 (444) 9175 (703)
RT (msec) 570 (113) 604 (80)
Standard deviations are shown in parentheses
Figure 1 The contour (upper panel) and interval (lower panel)stimuli In each case the first four notes of the melodies (commonsequence) are shown followed by the standard and deviant (oddball)terminal notes In the contour case interval size changes from melodyto melody but the standard terminal notes always rise whereas theoddball terminal notes always fall In the interval case deviant terminalnotes either go outside the key of the common sequence (secondndashlastbar) or remain in the key of the common sequence (last bar) but thecontour is always rising
Trainor McDonald and Alain 433
in amplitude between the whole-tone and semitonechanges in the interval task so the data were collapsedacross this factor An ANOVA with task (contour inter-
val) and stimulus type (standard deviant) revealed thatdeviants were indeed more negative than standardsF(110) = 5083 p lt 001 and that contour was more
Figure 2 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the passive contour andinterval tasks In this and thesubsequent figures the thickbar shows the onset of the finalmelody note
434 Journal of Cognitive Neuroscience Volume 14 Number 3
negative than interval F(110) = 2794 p lt 001 Therewas no interaction between task and stimulus typehowever indicating that the MMN was of comparablesize in the contour and interval tasks (see Figure 3)
An examination of the head maps (Figure 4) showsthat the MMN was fronto-centrally distributed over thescalp in the contour task while it was greater over theright hemisphere in the interval task However therewere no significant topological differences betweencontour and interval MMN as revealed in analyses usingall electrodes as well as analyses using only midline oronly lateral line electrodes
As expected no P3b was evident in the passive listen-ing tasks of either contour or interval Interestinglythere was also no evidence of a frontal positivity in thepassive listening task
Active Listening
Figure 5 shows group mean ERPs during active listeningfor the contour and interval tasks Because no significantdifferences were found for any ERP components be-tween the whole tone and semitone conditions in theinterval task the following analyses are collapsed acrossthis factor
In both contour and interval tasks standards anddeviants generated an N1 peaking at 119 msec (SD =127) poststimulus An ANOVA with task (contour
interval) and stimulus type (standard deviant) as fac-tors showed that the N1 measured at Fz was larger(mean amplitude between 95 and 135 msec post stim-ulus) in the contour than in the interval task F(110) =886 p lt 02 and larger for deviants than for standardsF(1 10) = 920 p lt 02 The latter effect is probablydue to the fact that the N2 wave elicited by the oddballstimuli was partly superimposed on the N1 deflection(see below)
Figure 3 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during pas-sive listening
Contour Interval
Figure 4 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for the peak MMN during contour(150 msec) and interval (180 msec) tasks Isocontour spacing is 01 mV
Trainor McDonald and Alain 435
The ERPs to deviant stimuli were characterized by anN2 wave that was superimposed on the P2 deflectionThe N2 wave was maximal over the fronto-central regionso the following analyses were done at Fz The N2
peaked earlier for the contour than for the intervaltask F(110) = 1791 p lt 002 The mean amplitudecalculated in 40-msec windows around the grand ave-rage peaks for contour (170ndash210 msec) and interval
Figure 5 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the active contour and in-terval tasks
436 Journal of Cognitive Neuroscience Volume 14 Number 3
(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3
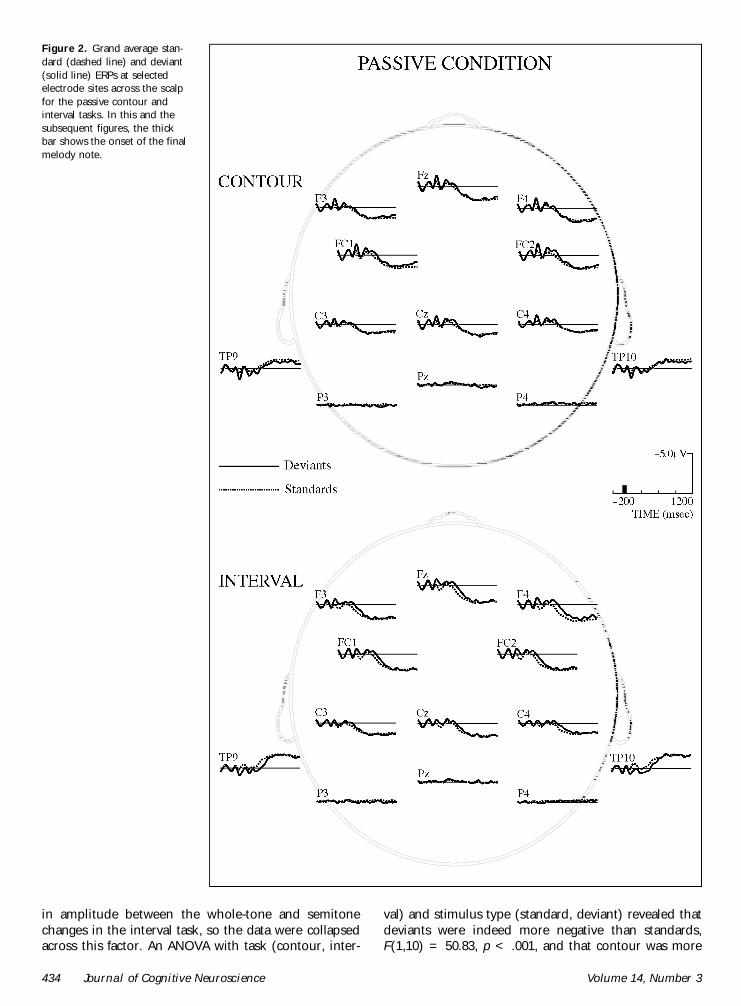
in amplitude between the whole-tone and semitonechanges in the interval task so the data were collapsedacross this factor An ANOVA with task (contour inter-
val) and stimulus type (standard deviant) revealed thatdeviants were indeed more negative than standardsF(110) = 5083 p lt 001 and that contour was more
Figure 2 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the passive contour andinterval tasks In this and thesubsequent figures the thickbar shows the onset of the finalmelody note
434 Journal of Cognitive Neuroscience Volume 14 Number 3
negative than interval F(110) = 2794 p lt 001 Therewas no interaction between task and stimulus typehowever indicating that the MMN was of comparablesize in the contour and interval tasks (see Figure 3)
An examination of the head maps (Figure 4) showsthat the MMN was fronto-centrally distributed over thescalp in the contour task while it was greater over theright hemisphere in the interval task However therewere no significant topological differences betweencontour and interval MMN as revealed in analyses usingall electrodes as well as analyses using only midline oronly lateral line electrodes
As expected no P3b was evident in the passive listen-ing tasks of either contour or interval Interestinglythere was also no evidence of a frontal positivity in thepassive listening task
Active Listening
Figure 5 shows group mean ERPs during active listeningfor the contour and interval tasks Because no significantdifferences were found for any ERP components be-tween the whole tone and semitone conditions in theinterval task the following analyses are collapsed acrossthis factor
In both contour and interval tasks standards anddeviants generated an N1 peaking at 119 msec (SD =127) poststimulus An ANOVA with task (contour
interval) and stimulus type (standard deviant) as fac-tors showed that the N1 measured at Fz was larger(mean amplitude between 95 and 135 msec post stim-ulus) in the contour than in the interval task F(110) =886 p lt 02 and larger for deviants than for standardsF(1 10) = 920 p lt 02 The latter effect is probablydue to the fact that the N2 wave elicited by the oddballstimuli was partly superimposed on the N1 deflection(see below)
Figure 3 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during pas-sive listening
Contour Interval
Figure 4 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for the peak MMN during contour(150 msec) and interval (180 msec) tasks Isocontour spacing is 01 mV
Trainor McDonald and Alain 435
The ERPs to deviant stimuli were characterized by anN2 wave that was superimposed on the P2 deflectionThe N2 wave was maximal over the fronto-central regionso the following analyses were done at Fz The N2
peaked earlier for the contour than for the intervaltask F(110) = 1791 p lt 002 The mean amplitudecalculated in 40-msec windows around the grand ave-rage peaks for contour (170ndash210 msec) and interval
Figure 5 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the active contour and in-terval tasks
436 Journal of Cognitive Neuroscience Volume 14 Number 3
(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3
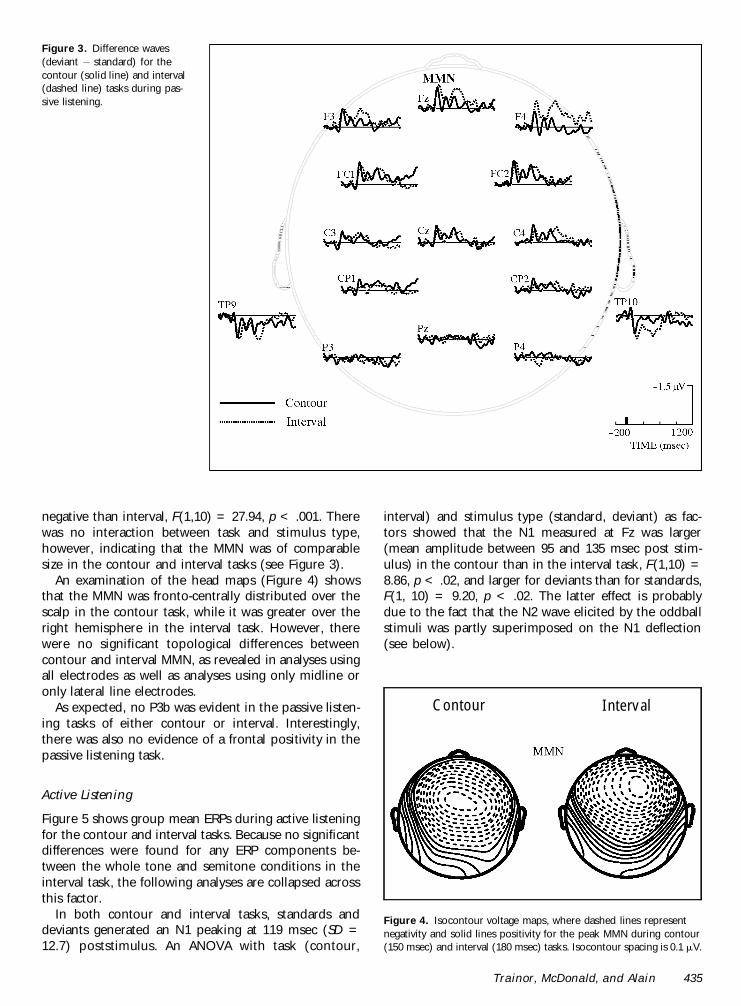
negative than interval F(110) = 2794 p lt 001 Therewas no interaction between task and stimulus typehowever indicating that the MMN was of comparablesize in the contour and interval tasks (see Figure 3)
An examination of the head maps (Figure 4) showsthat the MMN was fronto-centrally distributed over thescalp in the contour task while it was greater over theright hemisphere in the interval task However therewere no significant topological differences betweencontour and interval MMN as revealed in analyses usingall electrodes as well as analyses using only midline oronly lateral line electrodes
As expected no P3b was evident in the passive listen-ing tasks of either contour or interval Interestinglythere was also no evidence of a frontal positivity in thepassive listening task
Active Listening
Figure 5 shows group mean ERPs during active listeningfor the contour and interval tasks Because no significantdifferences were found for any ERP components be-tween the whole tone and semitone conditions in theinterval task the following analyses are collapsed acrossthis factor
In both contour and interval tasks standards anddeviants generated an N1 peaking at 119 msec (SD =127) poststimulus An ANOVA with task (contour
interval) and stimulus type (standard deviant) as fac-tors showed that the N1 measured at Fz was larger(mean amplitude between 95 and 135 msec post stim-ulus) in the contour than in the interval task F(110) =886 p lt 02 and larger for deviants than for standardsF(1 10) = 920 p lt 02 The latter effect is probablydue to the fact that the N2 wave elicited by the oddballstimuli was partly superimposed on the N1 deflection(see below)
Figure 3 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during pas-sive listening
Contour Interval
Figure 4 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for the peak MMN during contour(150 msec) and interval (180 msec) tasks Isocontour spacing is 01 mV
Trainor McDonald and Alain 435
The ERPs to deviant stimuli were characterized by anN2 wave that was superimposed on the P2 deflectionThe N2 wave was maximal over the fronto-central regionso the following analyses were done at Fz The N2
peaked earlier for the contour than for the intervaltask F(110) = 1791 p lt 002 The mean amplitudecalculated in 40-msec windows around the grand ave-rage peaks for contour (170ndash210 msec) and interval
Figure 5 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the active contour and in-terval tasks
436 Journal of Cognitive Neuroscience Volume 14 Number 3
(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

The ERPs to deviant stimuli were characterized by anN2 wave that was superimposed on the P2 deflectionThe N2 wave was maximal over the fronto-central regionso the following analyses were done at Fz The N2
peaked earlier for the contour than for the intervaltask F(110) = 1791 p lt 002 The mean amplitudecalculated in 40-msec windows around the grand ave-rage peaks for contour (170ndash210 msec) and interval
Figure 5 Grand average stan-dard (dashed line) and deviant(solid line) ERPs at selectedelectrode sites across the scalpfor the active contour and in-terval tasks
436 Journal of Cognitive Neuroscience Volume 14 Number 3
(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

(195ndash235 msec) tasks did not differ between tasks butwas larger for deviants than for standards F(110) =1759 p lt 002 (see Figures 5 and 6) There were nosignificant differences in scalp distribution for N2 com-ponents between the contour and interval tasks
At the frontal sites measured N2 waves in active tasksare thought to reflect superimposed MMN and N2bcomponents whereas the concurrent positivity at mas-toid sites (TP9 TP10) is thought to be a more puremeasure of MMN because the N2b does not reversepolarity here (Naatanen 1992) An ANOVA with condi-tion (passive active) and task (contour interval) asfactors yielded an almost significant effect of conditionF(110) = 479 p = 053 on the amplitude of the N2MMN peaks at Fz On the other hand an ANOVA withcondition (passive active) task (contour interval) andelectrode (TP9 TP10) as factors yielded no significanteffects Thus no significant effect of listening conditionon MMN was found but there was an N2b effect that wasmodulated by attention
Deviant stimuli also generated a frontal positivitypeaking at about 350 msec poststimulus (Figures 5and 6) At Fz the frontal positivity peaked earlier forthe contour task than for the interval task F(110) =1077 p lt 008 with means of 317 (SD = 442) and 379msec (SD = 577) respectively An ANOVA with task(contour interval) and stimulus type (standard devi-ant) as factors and the mean amplitude in 40-msec
windows around the peak as the dependent measurerevealed that deviants were indeed more positive thanstandards [main effect of stimulus type F(110) =1387 p lt 004] and that the interval stimuli weremore positive than the contour stimuli [main effect oftask F(110) = 536 p lt 04] The Task pound StimulusType interaction approached significance F(110) =460 p lt 06 indicating that the frontal positivitytended to be bigger in the interval than in the contourtask
There were no significant topographical differences inthe frontal positivity across the scalp between the con-tour and interval tasks according to an ANOVA withmean amplitude in a 40-msec window around the peak(ie 297ndash337 msec for contour and 359ndash399 msec forinterval) as the dependent measure and task (contourinterval) hemisphere (left right) and electrode (F34F78 AF34 FC12) as factors (Figure 7 upper panel)The frontal positivity was centered on the right side forboth contour and interval tasks however as indicatedby a significant effect of hemisphere F(110) = 531p lt 05
Deviant stimuli generated a large P3b wave maximalover the posterior regions (Figures 5 and 6) Althoughthe P3b at Pz tended to be earlier in the contour (mean= 548 msec SD = 706) than in the interval (mean =612 msec SD = 1077) task the large variance precludedthis effect from reaching significance Using the mean in
Figure 6 Difference waves(deviant iexcl standard) for thecontour (solid line) and interval(dashed line) tasks during ac-tive listening
Trainor McDonald and Alain 437
a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

a 100-msec windows centered at the peak an ANOVAwith task (contour interval) and stimulus type (stand-ard deviant) as factors revealed that deviants weresignificantly larger than standards F(110) = 1408 p lt004 but that there was no difference in P3b amplitudebetween the contour and interval tasks
Figure 7 lower panel shows the P3b scalp distribu-tion ANOVAs using all electrodes as well as analysesusing only midline and only lateral line electrodesrevealed no significant differences in topography be-tween the contour and interval tasks
DISCUSSION
Changes in both contour and interval were processedautomatically in the auditory cortices of nonmusiciansin the absence of absolute frequency information asrevealed by the presence of MMN Contour processingis important in both the music and the speech domains(Patel et al 1998) so it is not very surprising that thereare automatic mechanisms for its extraction Thepresent finding extends previous reports of MMN forcontour changes in two-tone sequences (Paavilainenet al 1998 Tervaniemi et al 1994 Saarinen et al1992) to multitone sequences Precise interval process-ing on the other hand is specific to the musical systemand is greatly affected by musical training (Patel et al1998 Peretz amp Morais 1988 Bever amp Chiarello 1974)
Thus the existence of automatic interval extractionmechanisms in musically untrained listeners suggeststhat musical information is important to the brainotherwise there would be no need for the automaticextraction of such detailed information about musicalpitch patterns This finding is new and indicates thatnot only does the auditory cortex encode sound fea-tures (pitch duration loudness location timbre) se-quential pattern information and abstract contourinformation but it also encodes the exact pitch distan-ces between tones in the absence of absolute pitchinformation
In general it might be expected that the magnitude ofthe MMN should reflect the magnitude of the deviationInterestingly there were no significant differences be-tween contour and interval deviants in this regardsuggesting that interval changes are just as salient ascontour changes at this level of processing There werehowever speed-of-processing differences with earlierMMN for contour than for interval changes suggestingthat more time is needed to extract interval than con-tour information Within interval processing there wereno significant differences in the magnitude of the MMNfor the larger whole tone over the smaller semitonedifferences This could potentially reflect the interactionof two forces Because the whole tone difference islarger than the semitone difference larger MMN mightbe expected in the whole tone case however thesemitone difference might be more salient because itgoes outside the key of the melody whereas the wholetone difference does not leading to the expectation forlarger MMN in the semitone case In terms of speed ofprocessing the size of the interval change appears to bemost important because the larger whole tone changesgenerated earlier MMN than the smaller semitonechanges It would be interesting to examine these issuesin musicians for whom musical key relations are likelyto be more salient
No evidence of attentional modulation of the MMNwas found for either contour or interval processingHowever it could be argued that the passive task (read-ing a book) was not sufficiently distracting to elicit acomplete lack of attention to the stimuli It is possiblethat with a more demanding distracter task the effectsof attention on MMN might have emerged (WoldorffHillyard Gallen Hampson amp Bloom 1998 Alain ampWoods 1997) In any case the effects of controlledprocessing on MMN for melodic processing are small ifthey exist at all On the other hand the effect ofcontrolled processing on both the frontal positivityand the P3b was profoundmdashthese components werecompletely absent in the passive tasks
Changes in both contour and interval produced afrontal right-sided positivity around 350 msec afterstimulus onset consistent with the findings of Trainoret al (1999) and Janata (1995) This component waspresent however only in the active and not in the
Contour Interval
Figure 7 Isocontour voltage maps where dashed lines representnegativity and solid lines positivity for frontal positivity (isocontourspacing of 03 mV) and P3b (isocontour spacing of 08 mV) at thepeak of each component (contour and interval latencies are 318 and379 msec for frontal positivity and 547 and 613 msec for P3b)
438 Journal of Cognitive Neuroscience Volume 14 Number 3
passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

passive condition It is unlikely that this frontal positi-vity represents the same processes as those of the P3athat have been described in the literature As discussedin the Introduction P3as are typically elicited either byoccasional salient stimuli that occur in an unattendedchannel (eg Escera et al 1998 Alho et al 1997 1998Squires et al 1975) or by occasional salient stimuli inan attended channel that are to be ignored (egComerchero amp Polich 1998 Katayama amp Polich 1998Holdstock amp Rugg 1993 Holdstock amp Rugg 1995Knight 1984) The frontal positivity in the present studyoccurred only in an attended channel and not in anunattended channel and it occurred to deviants thatwere targets
There are likely several generators of P3a waveformsincluding the frontal temporoparietal and hippocampalregions (eg Halgren Marinkovic amp Chauvel 1998Chao amp Knight 1996 Alain Richer Achim amp Saint-Hilaire 1989 Knight Scabini Woods amp Clayworth1989 Knight 1984) and one recent ERP study hasseparated two P3a components a centrally dominantcomponent peaking at 230 msec and a right frontallydominant component peaking at 315 msec (Escera et al1998) Escera et al (1998) suggest that the later compo-nent is generated primarily in the right frontal areas andreflects the actual orienting of attention whereas theearlier component reflects the capture of attention It ispossible that the right-sided frontal positivity observedin the present study reflects the actual orienting ofattention In this case it would seem that musical stimuliare particularly powerful in their ability to engage theattentional system Further research will have to addresswhether the frontal positivities observed in the presentstudy are specific to musical processing specific tocomplex pattern processing or are simply produced inforced choice tasks with complex stimuli
Some researchers have argued that contour andinterval information are processed separately in thebrain perhaps even in different hemispheres (egLiegeois-Chauvel et al 1998 Peretz amp Morais 1987Peretz et al 1987 Peretz 1990 Peretz amp Babaie 1992Edworthy 1985 Dowling 1978 1982 Bever amp Chiarello1974) However using ERPs as the measure we foundno significant topographical differences in MMN frontalpositivity or P3b when participants processed contourversus interval information (see also Schiavetto et al1999 Trainor et al 1999) It is possible that ERPmeasures are simply not sensitive enough to pick upsmall differences in the locations of the brain activityIndeed the head maps of Figure 5 suggest a more right-sided focus for interval than for contour MMN Howeverthe lack of significance suggests that differences in thelocation of the underlying generators if they exist arelikely to be small and may vary considerably from personto person The results do appear to be inconsistent withthe exclusive use of different hemispheres to performcontour and interval processing
The results of the present study show that both pitchcontour and music-specific pitch interval information areprocessed automatically in nonmusicians Thus we havediscovered a new type of information relative pitchdistance or pitch interval that elicits MMN In otherwords although there are tonotopic (absolute) pitchrepresentations in the auditory cortex the presentresults imply that there are also cortical circuits encod-ing interval information independent of the absolutepitch information Because interval information is spe-cific to musical processing the present results imply thatthe auditory system is set up to extract music-specificmelodic information
METHODS
Participants
Fifteen nonmusically trained adults participated in thestudy but four were excluded from the data analysis(one participant was unavailable for the interval portionof the study and three responded correctly to less than50 of deviants) The 11 remaining subjects (5 men6 women) ranged in age from 21 to 42 years (mean =289 years) All participants had no more than 4 years ofmusical training (mean = 136 years range = 0ndash4) andnone had perfect pitch For those subjects who did havesome musical training an average of 114 years (range 8ndash14 years) had passed since training stopped All parti-cipants were right-handed and reported normal hearing
Stimuli Apparatus and Procedure
In both the contour and the interval tasks there werestandard and oddball sets of melodies Each melodyconsisted of five notes Each note was a digitally re-corded file created with the Korg MIDI Tonefile 01RWusing piano timbre and sampled at 441 kHz Theappropriate five notes for each melody were played insuccession with note-to-note onsets of 300 msec for atotal melody length of 1500 msec Stimuli were pre-sented with a 16-bit Tucker Davis System and TDH 50Pheadphones at about 75 dB SPL
In the contour task all seven standard melodies wereascending in pitch but had different starting notes andcontained different intervals between notes (seeFigure 1) For each standard melody an oddball melodywas created in which the last note was changed to forma descending interval In other words the contour andinterval information was identical between standard anddeviant melodies up to the last note In the interval taskthe standard set consisted of one melody (the first fivenotes of the major scale) that was transposed to the 12major keys on different trials In order to maintain asense of Western tonality on successive trials the trans-position was either by a perfect fifth (seven semitones)or a perfect fourth (five semitones) There were two
Trainor McDonald and Alain 439
oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

oddball melodies for each transposition of the standardmelody one in which the final note was raised by a tone(and thus stayed within the key of the melody) and onein which it was lowered by a semitone (and thus wentoutside the key of the melody) Again the contour andinterval information was identical between standard anddeviant melodies up to the last note
For both contour and interval tasks standard anddeviant melodies were played on 80 and 20 of trialsrespectively (Figure 1) The order of trials was pseudo-random with the constraints that at least two standardtrials occurred between each oddball trial and that thesame melody did not occur twice in succession Melo-dies were separated by 2000 msec
All participants completed a passive and an activelistening task for both the contour and interval taskshalf participated in the contour task first and half in theinterval task first and within those groups half werepresented with the passive versions first and half theactive versions first In the passive listening task theparticipants read a book of their choice and ignoredthe stimuli In the active listening task the participantswere required on each trial to make a response as towhether the melody was from the standard or oddballset by pressing the number lsquolsquo1rsquorsquo or lsquolsquo2rsquorsquo on the numberpad of the keyboard with their right hand No feedbackwas provided on their performance
Electrophysiological Recording and Analysis
The electroencephalogram was recorded from an arrayof 47 electrodes including those from the standard10ndash20 placement Vertical and horizontal eye move-ments were recorded with electrodes at the outer canthiand at the superior and inferior orbit Electrophysio-logical signals were digitized continuously (bandpass005ndash50 Hz 250-Hz sampling rate) via NeuroScanSynAmps and stored for offline analysis During therecording all electrodes were referenced to Cz for dataanalysis they were re-referenced to an average referenceand the electrode Cz was reinstated
The analysis epoch included 200 msec of baselineactivity before the onset of the final melody note ofthe sequence and 1200 msec of poststimulus activityTrials contaminated by excessive peak-to-peak deflec-tion (plusmn 150 mV) at the channels not adjacent to the eyeswere automatically rejected before averaging The ERPswere then averaged separately for each site stimulustype and listening task ERPs were digitally lowpass-filtered to attenuate frequencies above 32 Hz
Prior to the experiment a set of ocular calibrationsignals was obtained from the participant From this setaveraged ERPs were calculated for vertical and horizontaleye movements as well as for eyeblinks Using principalcomponent analysis a set of components that bestexplained the eye movements was extracted using brainelectrical source analysis software The ocular source
components were then subtracted from the experi-mental ERPs to minimize ocular contamination (Berg ampScherg 1994 Lins Picton Berg amp Scherg 1993)
ERP components were quantified by computing meanvalues in selected latency regions relative to the meanamplitude of the 200-msec prestimulus activity Scalptopographies using 40 electrodes (omitting the perio-cular electrodes) were statistically analyzed after scalingthe amplitudes to eliminate amplitude differencesbetween tasks (McCarthy amp Wood 1985) All measure-ments were subjected to ANOVAs with repeated meas-ures The original degrees of freedom for all analysesare reported throughout the article Type I errorsassociated with inhomogeneity of variance were con-trolled by decreasing the degrees of freedom using theGreenhousendashGeisser epsilon and the probability esti-mates are based on these reduced degrees of freedom
Acknowledgments
This research was supported by a CIHR grant to LJT and anNSERC grant to CA We thank Yu He and Ryan Van Lieshoutfor technical assistance
Reprint requests should be sent to Dr Laurel J TrainorDepartment of Psychology McMaster University HamiltonON Canada L8S 4K1 or via e-mail ljtmcmasterca
REFERENCESAlain C Achim A amp Woods D L (1999) Separate memory-
related processing for auditory frequency and patternsPsychophysiology 36 737ndash744
Alain C Cortese F amp Picton T W (1999) Event-relatedactivity associated with auditory pattern processingNeuroReport 10 2429ndash2434
Alain C Richer F Achim A amp Saint-Hilaire J M (1989)Human intracerebral potentials associated with targetnovel and omitted auditory stimuli Brain Topography 1237ndash245
Alain C amp Woods D L (1997) Attention modulatesauditory pattern memory as indexed by event-related brainpotentials Psychophysiology 34 534ndash546
Alain C Woods D L amp Ogawa K H (1994) Brain indices ofautomatic pattern processing NeuroReport 6 140ndash144
Alho K Escera C Diaz R Yago E amp Serra J M (1997)Effects of involuntary auditory attention on visual taskperformance and brain activity NeuroReport 8 3233ndash3237
Alho K Winkler I Escera C Huotilainen M Virtanen JJaaskelainen I P Pekkonen E amp Ilmoniemi R J(1998) Processing of novel sounds and frequency changesin the human auditory cortex Magnetoencephalographicrecordings Psychophysiology 35 211ndash224
Bartlett J C amp Dowling W J (1980) Recognition oftransposed melodies A key-distance effect in developmentalperspective Journal of Experimental Psychology HumanPerception and Performance 6 501ndash515
Berg P amp Scherg M (1994) A multiple source approach tothe correction of eye artifacts Electroencephalography andClinical Neurophysiology 90 229ndash241
Besson M (1998) Meaning structure and time in languageand music Current Psychology of Cognition 17 921ndash950
Besson M amp Faotildeta F (1995) An event related potential (ERP)study of musical expectancy Comparison of musicians with
440 Journal of Cognitive Neuroscience Volume 14 Number 3
nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

nonmusicians Journal of Experimental Psychology HumanPerception and Performance 21 1278ndash1296
Besson M amp Macar F (1987) An event-related potentialanalysis of incongruity in music and other non-linguisticcontexts Psychophysiology 24 14ndash25
Bever T G amp Chiarello R J (1974) Cerebral dominance inmusicians and nonmusicians Science 185 537ndash539
Chao L L amp Knight R T (1996) Prefrontal and posteriorcortical activation during auditory working memoryCognitive Brain Research 4 27ndash37
Cohen D Granot R Pratt H amp Barneah A (1993)Cognitive meanings of musical elements as disclosed byevent-related potential (ERP) and verbal experimentsMusic Perception 11 153ndash184
Comerchero M D amp Polich J (1998) P3a perceptualdistinctiveness and stimulus modality Cognitive BrainResearch 7 41ndash48
Cuddy L L amp Cohen A J (1976) Recognition of transposedmelodic sequences Quarterly Journal of ExperimentalPsychology 28 255ndash270
Donchin E amp Coles M G H (1988) Is the P300 component amanifestation of context updating Behavioral and BrainSciences 11 357ndash374
Dowling W J (1978) Scale and contour Two components ofa theory of memory for melodies Psychological Review 85341ndash354
Dowling W J (1982) Contour in context Comments onEdworthy Psychomusicology 2 47ndash48
Edworthy J (1985) Interval and contour in melody processingMusic Perception 2 375ndash388
Escera C Alho K Winkler I amp Naatanen R (1998) Neuralmechanisms of involuntary attention to acoustic novelty andchange Journal of Cognitive Neuroscience 10 590ndash604
Halgren E Marinkovic K amp Chauvel P (1998) Generators ofthe late cognitive potentials in auditory and visual oddballtasks Electroencephalography and ClinicalNeurophysiology 106 156ndash164
Halpern A R amp Zatorre R J (1999) When that tune runsthrough your head A PET investigation of auditory imageryfor familiar melodies Cerebral Cortex 9 697ndash704
Holdstock J S amp Rugg M D (1993) Dissociation ofauditory P300 differential brain response to target andnovel nontarget stimuli In H J Heinze T F Munte ampG R Mangun (Eds) New developments in event-relatedpotentials (pp 71ndash78) Boston Birkhauser
Holdstock J S amp Rugg M D (1995) The effect of attentionon the P300 deflection elicited by novel sounds Journal ofPsychophysiology 9 18ndash31
Janata P (1995) ERP measures assay the degree of expectancyviolation of harmonic contexts in music Journal of CognitiveNeuroscience 7 153ndash164
Johnson-Laird P N (1988) How to reason syllogistically InA M Collins amp E E Smith (Eds) Readings in cognitivescience A perspective from psychology and artificial intel-ligence (pp 424ndash439) San Mateo CA Morgan Kaufmann
Katayama J amp Polich J (1998) Stimulus context determinesP3a and P3b Psychophysiology 35 23ndash33
Knight R T (1984) Decreased response to novel stimuli afterprefrontal lesions in man Electroencephalography andClinical Neurophysiology Evoked Potentials 59 9ndash20
Knight R T Scabini D Woods D L amp Clayworth C C(1989) Contributions of temporalndashparietal junction to thehuman auditory P3 Brain Research 502 109ndash116
Kuneg D amp Turk I (2000) New perspectives on thebeginnings of music Archeological and musicologicalanalysis of a middle paleolithic bone lsquolsquoflutersquorsquo In N LWallin B Merker amp S Brown (Eds) The origins ofmusic (pp 235ndash268) Cambridge MIT Press
Levett C amp Martin F (1992) The relationship betweencomplex music stimuli and the late components of theevent-related potential Psychomusicology 11 125ndash140
Liegeois-Chauvel C Peretz I Babaie M Laguitton V ampChauvel P (1998) Contribution of different cortical areasin the temporal lobes to music processing Brain 1211853ndash1867
Lins O G Picton T W Berg P amp Scherg M (1993)Ocular artifacts in EEG and event-related potentials I Scalptopography Brain Topography 6 51ndash63
McCarthy G amp Wood C C (1985) Scalp distributions ofevent-related potentials An ambiguity associated withanalysis of variance models Electroencephalography andClinical Neurophysiology 62 203ndash208
Mirz F Ovesen T Ishizu K Johannsen P Madsen SGjedde A amp Pedersen C B (1999) Stimulus-dependentcentral processing of auditory stimuli ScandinavianAudiology 28 161ndash169
Naatanen R (1992) Attention and brain function HillsdaleNJ Erlbaum
Naatanen R Tervaniemi M Sussman E Paavilainen P ampWinkler I (2001) lsquoPrimitive intelligencersquo in the auditorycortex Trends in Neuroscience 24 283ndash288
Paavilainen P Jaramillo M amp Naatanen R (1998) Binauralinformation can converge in abstract memory tracesPsychophysiology 35 483ndash487
Paller K A McCarthy G amp Wood C C (1992) Event-relatedpotentials elicited by deviant endings to melodiesPsychophysiology 29 202ndash206
Pantev C Bertrand O Eulitz C Verkindt C Hampson SSchuierer G amp Elbert T (1995) Specific tonotopic orga-nizations of different areas of the human auditory cortexrevealed by simultaneous magnetic and electric recordingsElectroencephalography and Clinical Neurophysiology 9426ndash40
Pantev C Hoke B Lutkenhoner B amp Lehnertz K (1989)Tonotopic organization of the auditory cortex Pitch versusfrequency representation Science 246 486ndash488
Patel A D amp Peretz I (1997) Is music autonomous fromlanguage A neuropsychological appraisal In I Deliege ampJ A Sloboda (Eds) Perception and cognition of music(pp 191ndash215) Hove England Erlbaum
Patel A D Peretz I Tramo M J amp Labrecque R (1998)Processing prosodic and musical patterns A neuropsycho-logical investigation Brain and Language 61 123ndash144
Peretz I (1990) Processing of local and global musicalinformation by unilateral brain-damaged patients Brain113 1185ndash1205
Peretz I amp Babaie M (1992) The role of contour andintervals in the recognition of melody parts Evidence fromcerebral asymmetries in musicians Neuropsychologia 30277ndash292
Peretz I amp Herbert S (2000) Toward a biological account ofmusic experience Brain and Cognition 42 131ndash134
Peretz I amp Morais J (1987) Analytic processing in theclassification of melodies as same or differentNeuropsychologia 25 645ndash652
Peretz I amp Morais J (1988) Determinants of laterality formusic Towards an information processing account InK Hugdahl (Ed) Handbook of dichotic listening Theorymethods and research (pp 323ndash358) Chichester EnglandWiley
Peretz I Morais J amp Bertelson P (1987) Shifting eardifferences in melody recognition through strategyinducement Brain and Cognition 6 202ndash215
Picton T W Alain C Otten L J Ritter W amp Achim A(2000) Mismatch negativity Different water in the sameriver Audiology amp Neuro-Otology 5 111ndash139
Trainor McDonald and Alain 441
Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3

Pinker S (1994) The language instinct New York WilliamMorrow and Co
Platel H Price C Baron J-C Wise R Lambert J Frack-owiak R S J Lechevalier B amp Eustache F (1997) Thestructural components of music perception Afunctional anatomical study Brain 120 229ndash243
Polich J (1988) Bifurcated P300 peaks P3a and P3b revisitedJournal of Clinical Neurophysiology 5 287ndash294
Rugg M D amp Coles M G H (1995) Electrophysiology ofmind Event-related brain potentials and cognition(vol 25) Oxford Oxford University Press
Saarinen J Paavilainen P Schroger E Tervaniemi M ampNaatanen R (1992) Representation of abstract attributesof auditory stimuli in the human brain NeuroReport 31149ndash1151
Schiavetto A Cortese F amp Alain C (1999) Global and localprocessing of musical sequences and event-related brainpotential study NeuroReport 10 2467ndash2472
Schroger E (1998) Measurement and interpretation ofthe mismatch negativity Behavior Research MethodsInstruments and Computers 30 131ndash145
Schroger E amp Wolff C (1998) Behavioral and electro-physiological effects of task-irrelevant sound change Anew distraction paradigm Cognitive Brain Research 771ndash87
Squires K C Squires N K amp Hillyard S A (1975) Twovarieties of long latency positive waves evoked by unpre-dictable auditory stimuli in man Electroencephalographyand Clinical Neurophysiology 38 387ndash401
Stuss D T Picton T W Cerri A M Leech E E amp StethemL L (1992) Perceptual closure and object identificationElectrophysiological responses to incomplete picturesBrain and Cognition 19 253ndash266
Tervaniemi M Maury S amp Naatanen R (1994) Neuralrepresentations of abstract stimulus features in the humanbrain as reflected by the mismatch negativity NeuroReport5 844ndash846
Trainor L J Desjardins R N amp Rockel C (1999) Acomparison of contour and interval processing in musiciansand nonmusicians using event-related potentials AustralianJournal of Psychology 51 147ndash153
Trainor L J amp Trehub S E (1992) A comparison of infantsrsquoand adultsrsquo sensitivity to western musical structure Journalof Experimental Psychology Human Perception andPerformance 18 394ndash402
Trainor L J amp Trehub S E (1994) Key membership andimplied harmony in Western tonal music Developmentalperspectives Perception and Psychophysics 56 125ndash132
Trehub S E amp Trainor L J (1998) Singing to infantsLullabies and play songs In C Rovee-Collier L P Lipsitt ampH Hayne (Eds) Advances in infancy research (vol 12pp 43ndash77) Stamford CT Ablex
Trehub S E Trainor L J amp Unyk A M (1993) Music andspeech perception in the first year of life In H W Reese ampL P Lipsitt (Eds) Advances in child development andbehavior (vol 24 pp 1ndash35) New York Academic Press
Verleger R (1997) On the utility of P3 latency as an index ofmental chronometry Psychophysiology 34 131ndash156
Wallin N L Merker B amp Brown S (2000) The origins ofmusic Cambridge MIT Press
Woldorff M G Hillyard S A Gallen C C Hampson S R ampBloom F E (1998) Magnetoencephalographic recordingsdemonstrate attentional modulation of mismatch-relatedneural activity in human auditory cortex Psychophysiology35 283ndash292
Woods D L Alho K amp Algazi A (1993) Intermodal selectiveattention Evidence for processing in tonotopic auditoryfields Psychophysiology 30 287ndash295
Zatorre R J (1984) Musical perception and cerebral functionA critical view Music Perception 2 196ndash221
Zatorre R J Halpern A R Perry D W Meyer E amp EvansA C (1996) Hearing in the mindrsquos ear A PET investigationof musical imagery and perception Journal of CognitiveNeuroscience 8 29ndash46
442 Journal of Cognitive Neuroscience Volume 14 Number 3
Related Documents











