Agricultural and Forest Entomology (2012), 14, 207–215 DOI: 10.1111/j.1461-9563.2011.00560.x Attraction to monoterpenes and beetle-produced compounds by syntopic Ips and Dendroctonus bark beetles and their predators Richard W. Hofstetter, Monica L. Gaylord, Sharon Martinson∗ and Michael R. Wagner School of Forestry, Northern Arizona University, Flagstaff, AZ 86011, U.S.A. and ∗ Department of Biological Sciences, Dartmouth College, Hanover, NH 03755, U.S.A. Abstract 1 Bark beetles are significant mortality agents of conifers. Four beetle species, the pine engraver Ips pini, the six-spined pine engraver Ips calligraphus sub. ponderosae, the southern pine beetle Dendroctonus frontalis, and the western pine beetle Dendroctonus brevicomis, cohabitate pines in Arizona. 2 A pheromone trapping study in ponderosa forests of Arizona determined the attraction of beetles to conspecific and heterospecific pheromone components in the presence and absence of host volatiles, and tested whether predators differ in their attraction to combinations of pheromone components and tree monoterpenes. 3 All four bark beetle species differed in their responses to heterospecific lures and monoterpenes. Ips calligraphus was the only species that increased in trap catches when heterospecific lures were added. Heterospecific lures did not inhibit the attraction of either Dendroctonus or Ips species. The replacement of myrcene with α-pinene increased the attraction of Dendroctonus, whereas the addition of α-pinene had mixed results for Ips. The prominent predators Temnochila chlorodia and Enoclerus lecontei were more attracted to the I. pini lure than the D. brevicomis lure, and the combination of the two lures with α-pinene was most attractive to both predator species. 4 Cross attraction and limited inhibition of bark beetles to heterospecific pheromones suggest that some of these species might use heterospecific compounds to increase successful location and colonization of trees. Predator responses to treatments suggest that tree volatiles are used to locate potential prey and predators are more responsive to Ips than to Dendroctonus pheromone components in Arizona. Keywords Kairomones, pine engraver, pheromones, southern pine beetle, western pine beetle. Introduction Bark beetles in the genera Dendroctonus and Ips commonly colonize phloem and cambium tissues of ponderosa pine (Pinus ponderosa Laws.) in North America (Wood, 1982; Cibrian- Tovar et al., 1995). In Arizona, Ips are considerable agents of ponderosa mortality, often causing higher mortality rates than Dendroctonus species (USDA Forest Service, 2002–2010). Ips and Dendroctonus species, however, are often observed to col- onize the same host tree (i.e. syntopic) (Breece et al., 2008; Wallin et al., 2008). Four bark beetles species, the pine engraver Ips pini Say, the six-spined pine engraver Ips calligraphus sub. ponderosae Swaine, the southern pine beetle Dendroctonus Correspondence: Richard W. Hofstetter. Tel.: +1 928 523 6452; fax: +1 928 523 1080; e-mail: [email protected] frontalis Zimmerman, and the western pine beetle Dendroc- tonus brevicomis LeConte (Coleoptera: Curculionidae, Scolyti- nae), are abundant in ponderosa pine forests of central Arizona (Wood, 1982; Williams et al., 2008). Pre-attack olfactory mechanisms for avoiding competition would be mutually advantageous to species competing for the same resource when beetle densities are high (Amezaga & Rodriguez, 1998). Competitive displacement via beetle pheromones has been observed between multiple bark beetle genera: I. pini and D. ponderosae Hopkins in lodgepole pine in northwest U.S.A. (Rankin & Borden, 1991; Safranyik et al., 1996; Boone et al., 2008); Dendroctonus rufipennis and Ips tri- dens in spruce in British Columbia, Canada (Poland & Borden, 1998), Ips paraconfusus and D. brevicomis in ponderosa pine in California (Byers & Wood, 1980, 1981); and Pityogenes chalcographus and Ips typographus in Europe (Byers, 1993; © 2012 The Authors Agricultural and Forest Entomology © 2012 The Royal Entomological Society

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Agricultural and Forest Entomology (2012), 14, 207–215 DOI: 10.1111/j.1461-9563.2011.00560.x
Attraction to monoterpenes and beetle-produced compounds bysyntopic Ips and Dendroctonus bark beetles and their predators
Richard W. Hofstetter, Monica L. Gaylord, Sharon Martinson∗ and Michael R. Wagner
School of Forestry, Northern Arizona University, Flagstaff, AZ 86011, U.S.A. and ∗Department of Biological Sciences, Dartmouth College, Hanover,
NH 03755, U.S.A.
Abstract 1 Bark beetles are significant mortality agents of conifers. Four beetle species,the pine engraver Ips pini, the six-spined pine engraver Ips calligraphus sub.ponderosae, the southern pine beetle Dendroctonus frontalis, and the western pinebeetle Dendroctonus brevicomis, cohabitate pines in Arizona.
2 A pheromone trapping study in ponderosa forests of Arizona determined theattraction of beetles to conspecific and heterospecific pheromone components inthe presence and absence of host volatiles, and tested whether predators differ intheir attraction to combinations of pheromone components and tree monoterpenes.
3 All four bark beetle species differed in their responses to heterospecific luresand monoterpenes. Ips calligraphus was the only species that increased in trapcatches when heterospecific lures were added. Heterospecific lures did not inhibitthe attraction of either Dendroctonus or Ips species. The replacement of myrcenewith α-pinene increased the attraction of Dendroctonus, whereas the addition ofα-pinene had mixed results for Ips. The prominent predators Temnochila chlorodiaand Enoclerus lecontei were more attracted to the I. pini lure than the D. brevicomislure, and the combination of the two lures with α-pinene was most attractive to bothpredator species.
4 Cross attraction and limited inhibition of bark beetles to heterospecific pheromonessuggest that some of these species might use heterospecific compounds to increasesuccessful location and colonization of trees. Predator responses to treatmentssuggest that tree volatiles are used to locate potential prey and predators are moreresponsive to Ips than to Dendroctonus pheromone components in Arizona.
Keywords Kairomones, pine engraver, pheromones, southern pine beetle, westernpine beetle.
Introduction
Bark beetles in the genera Dendroctonus and Ips commonlycolonize phloem and cambium tissues of ponderosa pine (Pinusponderosa Laws.) in North America (Wood, 1982; Cibrian-Tovar et al., 1995). In Arizona, Ips are considerable agents ofponderosa mortality, often causing higher mortality rates thanDendroctonus species (USDA Forest Service, 2002–2010). Ipsand Dendroctonus species, however, are often observed to col-onize the same host tree (i.e. syntopic) (Breece et al., 2008;Wallin et al., 2008). Four bark beetles species, the pine engraverIps pini Say, the six-spined pine engraver Ips calligraphus sub.ponderosae Swaine, the southern pine beetle Dendroctonus
Correspondence: Richard W. Hofstetter. Tel.: +1 928 523 6452; fax:+1 928 523 1080; e-mail: [email protected]
frontalis Zimmerman, and the western pine beetle Dendroc-tonus brevicomis LeConte (Coleoptera: Curculionidae, Scolyti-nae), are abundant in ponderosa pine forests of central Arizona(Wood, 1982; Williams et al., 2008).
Pre-attack olfactory mechanisms for avoiding competitionwould be mutually advantageous to species competing forthe same resource when beetle densities are high (Amezaga& Rodriguez, 1998). Competitive displacement via beetlepheromones has been observed between multiple bark beetlegenera: I. pini and D. ponderosae Hopkins in lodgepole pinein northwest U.S.A. (Rankin & Borden, 1991; Safranyik et al.,1996; Boone et al., 2008); Dendroctonus rufipennis and Ips tri-dens in spruce in British Columbia, Canada (Poland & Borden,1998), Ips paraconfusus and D. brevicomis in ponderosa pinein California (Byers & Wood, 1980, 1981); and Pityogeneschalcographus and Ips typographus in Europe (Byers, 1993;
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society

208 R. W. Hofstetter et al.
Lieutier et al., 2004). Alternatively, colonization of healthytrees requires hundreds to thousands of beetles (Safranyik et al.,1975; Berryman, 1976; Raffa & Berryman, 1983), and coop-eration among species could increase the chance of successfulcolonization of host trees, particularly when bark beetle densi-ties are low (Davis & Hofstetter, 2009). In this case, multiplespecies might have positive taxis toward pheromones emittedby heterospecific species (Symonds & Elgar, 2004; Davis &Hofstetter, 2009). Cross attraction is known to occur in syntopicbark beetle species (Smith et al., 1990; Payne et al., 1991), suchas I. paraconfusus Lanier and I. calligraphus in California (Vite& Gara, 1962).
Insect predators of bark beetles exploit bark beetlepheromones (as kairomones) to locate prey (Dahlsten &Stephen, 1974; Billings & Cameron, 1984; Boone et al., 2008).Predator species can be species-specific in their preference ofpheromones or more general, preferring beetle pheromone com-ponents or tree volatiles (Seybold et al., 2006). Predators canbe more or less attracted to volatiles from host trees depend-ing on host selection behaviour of their prey (e.g. bark beetlesattacking living versus dead host trees). For example, predatorsof tree-killing bark beetles use different suites of chemical cues,such as pheromones from beetles or host tree semiochemicals,to locate their prey, and predators of secondary bark beetlesoften target bark beetle pheromones and disregard host treevolatiles (Reeve et al., 2009). Research in northern Arizona hasfound that predators are collected more in traps baited with thelure for I. pini than the standard lure for D. brevicomis (Gay-lord et al., 2006; Williams et al., 2009). How predators respondto a combination of beetle pheromones with and without thepresence of particular monoterpenes in Arizona is not known.
In the present study, we focus solely on the attractiveness ofknown aggregation pheromone components from I. pini (Birchet al., 1980; Teale et al., 1991; Seybold et al., 1995; Steed& Wagner, 2008), D. brevicomis (Wood et al., 1976; Byers,1983; Hofstetter et al., 2008) and D. frontalis (Renwick &Vite, 1968; Hofstetter et al., 2008), which occur during the ini-tial colonization phase. The objectives were to determine therelative attraction of Ips and Dendroctonus beetles to conspe-cific and heterospecific aggregation pheromones in the presenceor absence of host volatiles, and determine whether preda-tors and competitors differ in the attraction to multiple barkbeetle aggregation pheromones and tree monoterpene combi-nations. We hypothesize that: (i) the presence of heterospecificpheromone components will reduce bark beetle capture and (ii)predator captures will be highest in traps with both bark beetlepheromone components. We tested our hypotheses by combin-ing tree monoterpenes with either or both I. pini lure and D.brevicomis lure components. The I. pini lure is attractive toboth Ips pini and I. calligraphus (Renwick & Vite, 1972; Milleret al., 1996) and the D. brevicomis lure is attractive to bothD. brevicomis and D. frontalis amongst other Dendroctonusspecies (Gaylord et al., 2006; Hofstetter et al., 2008; Williamset al., 2008).
Materials and methods
A multicomponent pheromone trapping experiment was con-ducted at three different sites in natural ponderosa pine (Pinus
Table 1 Study sites in Arizona. Each trap array contained eight traptreatments
Study site ElevationNumber oftrap arrays Latitude and longitude
Woody Mountain 2235 m 3 35◦08′44 N 111◦46′06 WLake Mary 2268 m 2 34◦52′01 N 111◦25′49 WRim 2140 m 3 34◦28′09 N 111◦27′27 W
ponderosae Douglas ex. Lawson) stands within the CoconinoNational Forest of central and northern Arizona from 25 Julyto 6 September 2005 (Table 1). At each site, two or three lin-ear arrays of eight 12-unit funnel traps (Lindgren, 1983) [PheroTech Inc. (now Contech Enterprises), Canada] were set up with50 m between traps and >2 km between blocks. Each trap washung on metal conduit at least 3 m from the nearest tree. Bot-toms of traps were 1 m above ground. Each trap was randomlyallocated to a treatment and the location was re-randomizedeach sample period. Lures were attached to the middle fun-nel of the trap and a 3 × 3 cm section of Spectracide Bug Stoppest strip (18.6% Dichlorvos, United Industries Corp., St Louis,Missouri) was placed into each collection cup to kill capturedinsects and reduce predation by predatory insects. Trap catcheswere collected every 3–4 days. All beetles were identified tospecies or genus, except for large wood borers (Cerambycidaeand Buprestidae) and click beetles (Elateridae), which wereonly identified to family.
Information for each lure component is provided inTable 2. The eight treatments tested were: (i) blank con-trol (no lure); (ii) I. pini lure (I) (ipsdienol and lanierone)(Steed & Wagner, 2008); (iii) I. pini lure + low release ofα-pinene (IL); (iv) I. pini lure + high release of α-pinene(IH); (v) D. brevicomis lure (W) (exo-brevicomin + frontalin+ myrcene); (vi) D. brevicomis lure with myrcene replacedby low α-pinene (WL); (vii) D. brevicomis lure with myrcenereplaced by high release α-pinene (WH); and (viii) I. pinilure + D. brevicomis lure with myrcene replaced by highrelease α-pinene (WIH). High release rates for the α-pinenewere in the range of the release rates reported from fresh pon-derosa chips (1–3 days post chipping) and the low release ratewas slightly lower than the release rates reported from slash(Fettig et al., 2006).
Analysis of variance was performed on the four target barkbeetles, as well as known predators for which substantialnumbers (>500 individuals for the duration of the trappingperiod) were captured. Data were analyzed using a split-plot analysis of repeated measures design (sas proc mixed;SAS Institute, Cary, North Carolina). Site and Lure andtheir interaction were considered as independent effects. Datawere transformed (
√0.375 + y) resulting in homoscedasticity
(Rao, 1998), although raw means and SEs are shown. Wheresignificant treatment effects occurred (α = 0.05), differenceswere separated by Tukey’s honestly significant difference onleast squared means (Carmer & Swanson, 1973). The blankcontrol trap data were not included in our analyses becausetoo few beetles (mean range of 0–0.04 beetles/trap/day) werecollected (Reeve & Strom, 2004).
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215

Attraction of pheromones to beetles and predators 209
Table 2 Compounds and their source, chemical purity, release devices, enantiomeric composition, and release characteristics of semiochemicalstested in field experiments
Compounds usedin treatments Source Chemical purity (%) Release device
Enantiomericcomposition (+, −)
Release rate(mg/day) (SD)
Frontalin Phero Techa 99 Bubble cap 50 : 50 2.0 (0.2)Ipsdienol Phero Tech 95 Bubble cap 3 : 97 0.2 (0.05)exo-Brevicomin Phero Tech 99 Bubble cap 50 : 50 1.5 (0.2)Lanierone Phero Tech Bubble cap Not chiral 0.2 (0.05)α-Pinene (low rate) Sigma-Aldrichb 99 Polyethylene bottlec 81 : 19 2.2 (0.3)α-Pinene (high rate) Sigma-Aldrich 99 Polyethylene bottles 81 : 19 21.7 (0.5)Myrcene Phero Tech 93 Polyethylene bottle Not chiral 1.2 (1.0)
Release rates are based on loses during field conditions.aPhero Tech Inc. (now Contech Enterprises).bSt. Louis, Missouri.c15-mL polyethylene tubes [Phero Tech Inc. (now Contech Enterprises)].
Table 3 Total numbers of beetles collected from trapping study duringSummer 2005 in the Coconino National Forest, Arizona
Insect Family Guild Total
Dendroctonus adjunctus Curculionidae Herbivores 36Dendroctonus
approximatus(Scolytinae) 92
Dendroctonusbrevicomis
502
Dendroctonusfrontalis
2489
Dendroctonus valens 105Hylastes spp. 123Ips calligraphus 2248Ips lecontei 54Ips pini 2323Orthotomicus latidens 15
Metallic woodborers Buprestidae Herbivores/predators
20
Longhorn beetles Cerambycidae 211Cossonus sp. Curculionidae 4Corticeus sp. Tenebrionidae 3Click beetle spp. Elateridae 174
Enoclerus spp.a Cleridae Predators 837Platysoma spp. Histeridae 52Temnochila chlorodia Trogositidae 1677Tenebroides sp. Trogositidae 38
Elacatis sp. Othniidae Unknown 872Total: 11 875
aEnoclerus lecontei: 95% of the catch; Enoclerus sphegeus: 5% ofthe catch.Insects shown in bold represent species that were analyzed in thepresent study.
Results
A total of 11 875 beetles over a 42-day period were cap-tured, representing more than 22 insect species (Table 3).Bark beetles and bark beetle predators accounted for 67%and 29% of the insects collected, respectively. The remaining4% of insects consisted of Buprestidae, Cerambycidae, Elateri-dae and other bark-inhabiting insects. The most abundant barkbeetle species was I. calligraphus followed by D. frontalis,
I. pini and D. brevicomis. Low numbers of Hylastes spp.,Dendroctonus valens LeConte, Dendroctonus approximatusHopkins, Ips lecontei Swaine, Dendroctonus adjunctus Bland-ford, and Orthotomicus latidens LeConte (listed in order ofabundance) were also captured but not analyzed. The mostabundant bark beetle predator was Temnochila chlorodia (Man-nerheim, 1843) (Coleoptera: Trogositidae) followed by Eno-clerus spp. (Coleoptera: Cleridae). Several Enoclerus sphegeuswere captured and analyzed together with Enoclerus leconteinumbers. Other predators captured, although in low numbers,included Tenebroides and Platysoma species. A large numberof Elacatis sp. (Coleoptera: Othniidae), a potential competitor(Fukuda, 1962), were also captured but not analyzed.
Dendroctonus bark beetles
Trap catches of D. frontalis exhibited a significant lure bysite interaction (Fig. 1 and Table 4); thus, sites were analyzedindependently. There was a significant effect of lure within eachsite (d.f. = 6, P < 0.0001 for each site; F = 13.99 for LakeMary, F = 25.46 for Woody Mountain, F = 32.35 for Rim).At all sites, the highest numbers of D. frontalis were collectedin the WH treatments, whereas few beetles were attracted toI, IL or IH traps. Replacement of myrcene with α-pinene inthe WL and WH traps increased trap catches, although themagnitude varied by site. The WIH treatment reduced trapcatches slightly from that of the WH treatment, although thedifference was not significant.
Dendroctonus brevicomis attraction to lures showed a signif-icant interaction by site (Table 4). Within each site, there was asignificant difference in lure attraction (P < 0.0001, d.f. = 6,for all three sites; F = 10.94 for Lake Mary, F = 12.06 forWoody Mountain, F = 7.7 for Rim) (Fig. 2). The highestattraction was to the WL and WH treatments. The additionof the I lure to the WH lure decreased the attraction of D. bre-vicomis, although the difference was not significant. The I lurewith or without terpenes was not attractive to D. brevicomis.
Ips bark beetles
The two most abundant Ips beetles in our traps, I. calligraphusand I. pini, had a significant site by lure interaction (Table 4).
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215

210 R. W. Hofstetter et al.
(a) Lake Mary
D. frontalis/trap/day D. frontalis/trap/day D. frontalis/trap/day
0.0 0.5 1.0 1.5 2.0 2.5 3.0
Lu
re
Lu
re
Lu
re
I
IL
IH
W
WL
WH
WIH
(b) Rim
0 2 4 6 8
I
IL
IH
W
WL
WH
WIH
(c) Woody Mtn.
0 1 2 3 4 5
I
IL
IH
W
WL
WH
WIHab
a
a
bc
c
c
c
c
c
c
b
c
ab
a
d
cd
d
a
ab
bc
b
Figure 1 Mean ± SEM of Dendroctonus frontalis beetles collected in Lindgren funnel traps with different lure and terpene combinations at (a) LakeMary, (b) Rim and (c) Woody Mountain sites in ponderosa pine forests in northern Arizona over 6 weeks in 2005. Lure treatments were (I) Ips pinilure (−97/+3 ipsdienol and lanierone); (IL) I. pini lure + low release of α-pinene (IH) I. pini lure + high release of α-pinene, (W) D. brevicomis lure(exo-brevicomin + frontalin + myrcene); (WL) D. brevicomis lure with myrcene replaced by low α-pinene, (WH) D. brevicomis lure with myrcene replacedby high release α-pinene and (WIH) I. pini lure + D. brevicomis lure with myrcene replaced by high release α-pinene. Blank control trap data were notincluded. Means followed by the same lowercase letter are not significantly different at P = 0.05 (analysis of variance).
Table 4 Results for split plot analysis of beetle trap catches
Species
Lure × Sited.f. = 12P-value(F-ratio)
Site d.f. = 2P-value(F-ratio)
Lure d.f. = 6P-value(F-ratio)
Dendroctonusfrontalis
<0.0001* (3.78)
Dendroctonusbrevicomis
0.0494* (1.77)
Ips calligraphus <0.0001* (11.53)Ips pini <0.0001* (8.40)Temnochila
chlorodia0.82 (0.63) 0.22 (1.51) <0.0001* (14.90)
Enoclerus spp. 0.42 (1.04) 0.0002* (8.82) <0.0001* (13.45)
Orthotomicus latidens (LeConte) and I. lecontei were alsocaptured, although there were too few to draw conclusions onlure preferences.
Ips calligraphus showed a significant difference in attractionto lures within all three sites (P = 0.0014, d.f. = 6, F = 3.83for Lake Mary; P = 0.0006, d.f. = 6, F = 4.13 for WoodyMountain; P < 0.0001, d.f. = 6, F = 12.76 for the Rim site)(Fig. 3) and, overall, showed a significant attraction to the Itreatment with high release rate of α-pinene (=IH). Additionof the W lure to the IH lure (=WIH) increased trap collectionsat two of the three sites, although the difference was notsignificant. At all three sites, few beetles were attracted to theW lure.
Ips pini showed a significant difference in attraction tolures within each site (P < 0.0001, d.f. = 6 for all three sites;F = 9.62 for Lake Mary, F = 23.73 for Woody Mountain,F = 34.69 for Rim). At all three sites, there was no attractionto the W lure (Fig. 4). Impact of the addition of the terpenelure on catches of I . pini varied by site. Addition of the Wlure to the IH lure did not significantly affect trap collectionsrelative to the IH lure alone.
Bark beetle predators
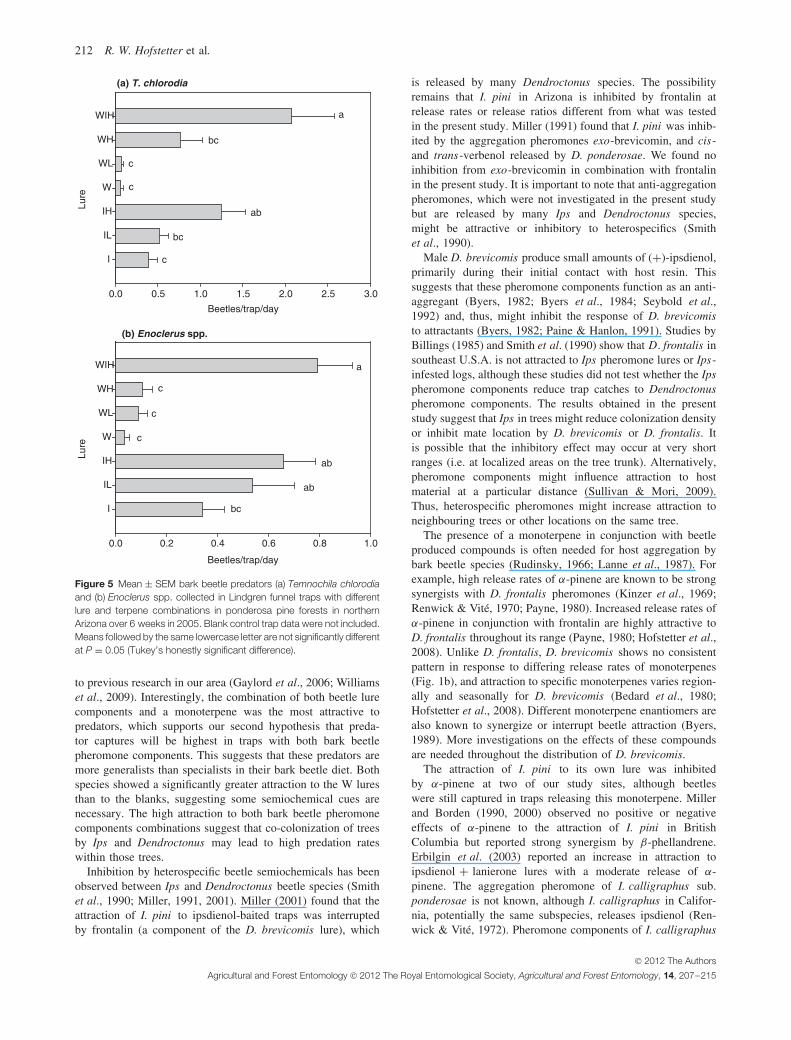
There was no significant interaction of lure by site for barkbeetle predators (Table 4). Overall, the prominent predators(T. chlorodia and Enoclerus spp.) were more attracted to the Ilure than the W lure (Fig. 5), and the WIH treatment collectedthe highest number of both predator species. Addition of α-pinene to the W lure and I lure generally increased trap catchesof both predators compared with lures without the terpene,and the presence of high release rate of α-pinene significantlyincreased captures of T. chlorodia. For Enoclerus, site alsohad a significant effect on trap catches, with Lake Mary andWoody Mountain sites collecting significantly higher numbersof beetles than the Rim site (mean ± SEM: 0.49 ± 0.10, 0.34 ±0.05 and 0.40 ± 0.03 for Lake Mary, Woody Mountain andRim, respectively).
Discussion
The presence of heterospecific pheromone components did notreduce or increase the capture of heterospecific bark beetles(i.e. a lack of strong support for hypothesis 1). There are manyreported instances of cross attraction to pheromone componentsproduced by other syntopic species of bark beetles (Lanier &Wood, 1975; Hedden et al., 1976; Svihra et al., 1980; Caneet al., 1990; Smith et al., 1990; Poland & Borden, 1994). InArizona, however, Dendroctonus and Ips species appeared toshow little cross attraction to pheromone components of othergenera. Additionally, attraction to their own pheromone wasslightly deterred by the presence of heterospecific pheromonecomponents, although this decrease was not significant for allspecies.
Many predators are habitat specialists in that they areattracted to bark beetle-killed trees, and might not specializeon a specific prey species but prey on many insects inside trees(Raffa, 2001). The predator species in the present study weremore attracted to the I lure than the W lure, which corresponds
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215

Attraction of pheromones to beetles and predators 211
(a) Lake Mary
D. brevicomis/trap/day D. brevicomis/trap/day D. brevicomis/trap/day
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4
Lu
re
Lu
re
Lu
re
I
IL
IH
W
WL
WH
WIH
(b) Rim
0.0 0.2 0.4 0.6 0.8
I
IL
IH
W
WL
WH
WIH
(c) Woody Mtn.
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
I
IL
IH
W
WL
WH
WIH
c
c
c
abc
a
ab
bc
c
c
c
bc
c
c
ab
a
abc bc
c
ab
a
ab
Figure 2 Mean ± SEM of Dendroctonus brevicomis beetles collected in Lindgren funnel traps with different lure and terpene combinations at (a) LakeMary, (b) Rim and (c) Woody Mountain sites in ponderosa pine forests in northern Arizona over 6 weeks in 2005. Blank control trap data were notincluded. Means followed by the same lowercase letter are not significantly different at P = 0.05 (analysis of variance).
(a) Lake Mary
I. calligraphus/trap/day
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
Lu
re
Lu
re
Lu
re
I
IL
IH
W
WL
WH
WIH
(b) Rim
I. calligraphus/trap/day
0 2 4 6 8 10 12 14
I
IL
IH
W
WL
WH
WIH
(c) Woody Mtn.
I. calligraphus/trap/day
0.00 0.02 0.04 0.06 0.08 0.10 0.12 0.14
I
IL
IH
W
WL
WH
WIH
b c
a
ab
ab a a
bc b
ab ab ab
a ab
c
c
b
b
b
b
b
Figure 3 Mean ± SEM of Ips calligraphus beetles collected in Lindgren funnel traps with different lure and terpene combinations at (a) Lake Mary,(b) Rim and (c) Woody Mountain sites in ponderosa pine forests in northern Arizona over 6 weeks in 2005. Blank control trap data were not included.Means followed by the same lowercase letter are not significantly different at P = 0.05 (analysis of variance).
(a) Lake Mary
I. pini/trap/day I. pini/trap/dayI. pini/trap/day0.0 0.5 1.0 1.5 2.0
Lure
Lure
Lure
I
IL
IH
W
WL
WH
WIH
b) Rim
0 2 4 6 8
I
IL
IH
W
WL
WH
WIH
(c) Woody Mtn.
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8
I
IL
IH
W
WL
WH
WIH
b
a
a
a
a a
a
a
b
b
b
b
b
c
c
c
b
ab
c
c
c
Figure 4 Mean ± SEM of Ips pini beetles collected in Lindgren funnel traps with different lure and terpene combinations at (a) Lake Mary, (b) Rimand (c) Woody Mountain sites in ponderosa pine forests in northern Arizona over 6 weeks in 2005. Blank control trap data were not included. Meansfollowed by the same lowercase letter are not significantly different at P = 0.05 (analysis of variance).
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215
212 R. W. Hofstetter et al.
(a) T. chlorodia
Beetles/trap/day
0.0 0.5 1.0 1.5 2.0 2.5 3.0
Lure
I
IL
IH
W
WL
WH
WIH
(b) Enoclerus spp.
Beetles/trap/day
0.0 0.2 0.4 0.6 0.8 1.0
Lure
I
IL
IH
W
WL
WH
WIH a
ab
a
ab
bc
c
c
c
c
ab
bc
bc
c
c
Figure 5 Mean ± SEM bark beetle predators (a) Temnochila chlorodiaand (b) Enoclerus spp. collected in Lindgren funnel traps with differentlure and terpene combinations in ponderosa pine forests in northernArizona over 6 weeks in 2005. Blank control trap data were not included.Means followed by the same lowercase letter are not significantly differentat P = 0.05 (Tukey’s honestly significant difference).
to previous research in our area (Gaylord et al., 2006; Williamset al., 2009). Interestingly, the combination of both beetle lurecomponents and a monoterpene was the most attractive topredators, which supports our second hypothesis that preda-tor captures will be highest in traps with both bark beetlepheromone components. This suggests that these predators aremore generalists than specialists in their bark beetle diet. Bothspecies showed a significantly greater attraction to the W luresthan to the blanks, suggesting some semiochemical cues arenecessary. The high attraction to both bark beetle pheromonecomponents combinations suggest that co-colonization of treesby Ips and Dendroctonus may lead to high predation rateswithin those trees.
Inhibition by heterospecific beetle semiochemicals has beenobserved between Ips and Dendroctonus beetle species (Smithet al., 1990; Miller, 1991, 2001). Miller (2001) found that theattraction of I. pini to ipsdienol-baited traps was interruptedby frontalin (a component of the D. brevicomis lure), which
is released by many Dendroctonus species. The possibilityremains that I. pini in Arizona is inhibited by frontalin atrelease rates or release ratios different from what was testedin the present study. Miller (1991) found that I. pini was inhib-ited by the aggregation pheromones exo-brevicomin, and cis-and trans-verbenol released by D. ponderosae. We found noinhibition from exo-brevicomin in combination with frontalinin the present study. It is important to note that anti-aggregationpheromones, which were not investigated in the present studybut are released by many Ips and Dendroctonus species,might be attractive or inhibitory to heterospecifics (Smithet al., 1990).
Male D. brevicomis produce small amounts of (+)-ipsdienol,primarily during their initial contact with host resin. Thissuggests that these pheromone components function as an anti-aggregant (Byers, 1982; Byers et al., 1984; Seybold et al.,1992) and, thus, might inhibit the response of D. brevicomisto attractants (Byers, 1982; Paine & Hanlon, 1991). Studies byBillings (1985) and Smith et al. (1990) show that D. frontalis insoutheast U.S.A. is not attracted to Ips pheromone lures or Ips-infested logs, although these studies did not test whether the Ipspheromone components reduce trap catches to Dendroctonuspheromone components. The results obtained in the presentstudy suggest that Ips in trees might reduce colonization densityor inhibit mate location by D. brevicomis or D. frontalis. Itis possible that the inhibitory effect may occur at very shortranges (i.e. at localized areas on the tree trunk). Alternatively,pheromone components might influence attraction to hostmaterial at a particular distance (Sullivan & Mori, 2009).Thus, heterospecific pheromones might increase attraction toneighbouring trees or other locations on the same tree.
The presence of a monoterpene in conjunction with beetleproduced compounds is often needed for host aggregation bybark beetle species (Rudinsky, 1966; Lanne et al., 1987). Forexample, high release rates of α-pinene are known to be strongsynergists with D. frontalis pheromones (Kinzer et al., 1969;Renwick & Vite, 1970; Payne, 1980). Increased release rates ofα-pinene in conjunction with frontalin are highly attractive toD. frontalis throughout its range (Payne, 1980; Hofstetter et al.,2008). Unlike D. frontalis, D. brevicomis shows no consistentpattern in response to differing release rates of monoterpenes(Fig. 1b), and attraction to specific monoterpenes varies region-ally and seasonally for D. brevicomis (Bedard et al., 1980;Hofstetter et al., 2008). Different monoterpene enantiomers arealso known to synergize or interrupt beetle attraction (Byers,1989). More investigations on the effects of these compoundsare needed throughout the distribution of D. brevicomis.
The attraction of I. pini to its own lure was inhibitedby α-pinene at two of our study sites, although beetleswere still captured in traps releasing this monoterpene. Millerand Borden (1990, 2000) observed no positive or negativeeffects of α-pinene to the attraction of I. pini in BritishColumbia but reported strong synergism by β-phellandrene.Erbilgin et al. (2003) reported an increase in attraction toipsdienol + lanierone lures with a moderate release of α-pinene. The aggregation pheromone of I. calligraphus sub.ponderosae is not known, although I. calligraphus in Califor-nia, potentially the same subspecies, releases ipsdienol (Ren-wick & Vite, 1972). Pheromone components of I. calligraphus
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215

Attraction of pheromones to beetles and predators 213
found in eastern U.S.A. are (–)-ipsdienol, trans-verbenol andcis-verbenol at elution ratios of 2 : 2.5 : 1, respectively (Viteet al., 1972, 1978). The present study shows that the combina-tion of the I and W lure with α-pinene (=WIH) was the mostattractive lure combination for I . calligraphus ponderosae. Ineastern U.S.A., I. calligraphus is not attracted to D. frontalispheromone components (Dixon & Payne, 1980; Smith et al.,1990) or D. frontalis-infested logs (Vite et al., 1964; Birchet al., 1980; Svihra, 1982; but see also Dixon & Payne 1980).In the present study, captures of this beetle increased withan increase in the release rate of α-pinene. Renwick and Vite(1972) indicated that the presence of host odour is an impor-tant component in the attraction of I. calligraphus to trees. Thepositive response of I. calligraphus to α-pinene suggests thatthis beetle in Arizona might attack wounded or even healthyhosts (Paine et al., 1981).
Given the extensive ponderosa pine stands in Arizona and thediverse community of bark beetles that persist there, long-termand wide-ranging interactions among these species likely occur.Such communities would more likely persist if there were ben-efits of multispecies associations on attacked trees (Davis et al.,2009). Ayres et al. (2001) discusses three mechanisms by whichbark beetle individuals could benefit from multispecies associa-tions: predator swamping, group attack and host location. Eachspecies can benefit from one or several of these mechanisms.For example, greater attack rates by bark beetles likely resultsin reduced predation per capita. Abrams et al. (1998) describeda similar situation as apparent mutualism. Both Dendroctonusand Ips probably share a guild of predators but predator attrac-tion to specific pheromones differs (Williams et al., 2009).Therefore, bark beetle species emitting pheromones that areless attractive to predators would suffer greater predation byco-colonizing a host tree with another species (Boone et al.,2008). Alternatively, Dendroctonus might benefit from multi-species aggregations if their densities are low and tree resistanceis high (Davis & Hofstetter, 2009).
The two dominant bark beetle predators varied in theirresponses to tree terpenes. High levels of host terpenes stronglysynergized attraction of T. chlorodia to beetle pheromones atthe same time as having no significant effect on the Eno-clerus species. Thus, T. chlorodia might target more aggressivebeetle species such as Dendroctonus spp., whereas Enoclerusmight target secondary bark beetles such as Ips spp., althoughthe results from the pheromone studies do not support thishypothesis. Niche partitioning by these predators likely reducesintraguild predation (Boone et al., 2008).
Conclusions
Low numbers of bark beetle species appear to be the normthrough ponderosa forests of Arizona (Sanchez-Martinez &Wagner, 2002; Gaylord et al., 2006, 2010). Only duringwidespread drought have bark beetles become sufficiently abun-dant to cause significant tree mortality across Arizona forests(USDA Forest Service, 2002–2010). During drought condi-tions, interactions among the beetle species studied might differfrom that reported in the present study. Pheromone compo-nents not tested or spacing of lures at distances varying from
those used in the present study might show differing attractionor deterrence to these species. Also, volatiles from microbesintroduced by heterospecific beetles might attract or deter latercolonizing species. For example, yeasts and fungi introduced byD. brevicomis can produce ethanol and 3-carene (Davis et al.,2010), which might influence attraction to host trees. The use ofW lures could have minimal effects on predator populations.In addition, estimates of predator populations based on stud-ies using only the W lure will underestimate their abundancecompared with studies using the I lure.
Acknowledgements
We thank Elizabeth Alden, Laine Smith, James Steward,Gabe DeGomez and Greg Zausen for their help in the fieldand sorting insects. We appreciate the efforts of the Schoolof Forestry – Northern Arizona University and the RockyMountain Research Station for providing us with laboratoryfacilities, as well as Centennial Forest, State Lands and NationalForest Service lands for use of field sites. We thank JoelMcMillin and Tom DeGomez for discussion of ideas. Weacknowledge our funding sources: Dartmouth College CrammerFund (to S. Martinson), Western Bark Beetle Initiative andUSDA Forest Service Rocky Mountain Research StationJoint-Venture Agreements 04-JV-11221615-270 and 05-PA-11221615-104.
References
Abrams, P.A., Holt, R.D. & Roth, J.D. (1998) Apparent competitionor apparent mutualism? Shared predation when populations cycle.Ecology, 79, 201–212.
Amezaga, I. & Rodriguez, M.A. (1998) Resource partitioning of foursympatric bark beetles depending on swarming dates and tree species.Forest Ecology and Management, 109, 127–135.
Ayres, B.D., Ayres, M.P., Abrahamson, M.D. & Teale, S.A. (2001)Resource partitioning and overlap in three sympatric species of Ipsbark beetles (Coleoptera: Scolytidae). Oecologia, 128, 443–453.
Bedard, W.D., Tilden, P.E., Lindahl, K.Q. Jr, Wood, D.L. &Rauch, P.A. (1980) Effects of verbenone and trans-verbenol onthe response of Dendroctonus brevicomis to natural and syntheticattractant in the field. Journal of Chemical Ecology, 6, 997–1013.
Berryman, A.A. (1976) Theoretical explanation of mountain pine beetledynamics in lodgepole pine forests. Environmental Entomology, 5,1225–1233.
Billings, R.F. (1985) Southern pine beetles and associated insects.Effects of rapidly-released host volatiles on response to aggregationpheromones. Zeitschrift fur Angewandte Entomologie, 99, 483–491.
Billings, R.F. & Cameron, R.S. (1984) Kairomonal responses ofColeoptera, Monochamus titillator (Cerambycidae), Thanasimusdubius (Cleridae) and Temnochila virescens (Trogositidae), tobehavioral chemicals of southern pine bark beetles (Coleoptera:Scolytidae). Environmental Entomology, 13, 1542–1548.
Birch, M.C., Light, D.M., Wood, D.L. et al. (1980) Pheromonal attrac-tion and allomonal interruption of Ips pini in California by the 2enantiomers of ipsdienol. Journal of Chemical Ecology, 6, 703–717.
Boone, C.K., Six, D.L. & Raffa, K.F. (2008) The enemy of my enemyis still my enemy: competitors add to predator load of a tree-killingbark beetle. Agricultural and Forest Entomology, 10, 411–421.
Breece, C.R., Kolb, T.E., Dickson, B.G., McMillin, J.D. & Clancy,K.M. (2008) Prescribed fire effects on bark beetle activity and tree
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215

214 R. W. Hofstetter et al.
mortality in southwestern ponderosa pine forests. Forest Ecology andManagement, 255, 119–128.
Byers, J.A. (1982) Male-specific conversion of the host plant com-pound, myrcene, to the pheromone, (+)-ipsdienol, in the bark beetle,Dendroctonus brevicomis. Journal of Chemical Ecology, 8, 363–371.
Byers, J.A. (1983) Influence of sex, maturity and host substances onpheromones in the guts of the bark beetles, Ips paraconfusus andDendroctonus brevicomis. Journal of Insect Physiology, 29, 5–13.
Byers, J.A. (1989) Chemical ecology of bark beetles. Experientia, 45,271–283.
Byers, J.A. (1993) Avoidance of competition by spruce bark beetles,Ips typographus and Pityogenes chalcographus. Experientia, 49,272–275.
Byers, J.A. & Wood, D.L. (1980) Interspecific inhibition of theresponse of the bark beetles, Dendroctonus brevicomis and Ipsparaconfusus to their pheromones in the field. Journal of ChemicalEcology, 6, 149–164.
Byers, J.A. & Wood, D.L. (1981) Interspecific effects of pheromoneson the attraction of the bark beetles, Dendroctonus brevicomis andIps paraconfusus in the laboratory. Journal of Chemical Ecology, 7,9–18.
Byers, J.A., Wood, D.L., Craig, J. & Hendry, L.B. (1984) Attractiveand inhibitory pheromones produced in the bark beetle, Dendroc-tonus brevicomis, during host colonization: regulation of inter-and intraspecific competition. Journal of Chemical Ecology, 10,861–877.
Cane, J.H., Wood, D.L. & Fox, J.W. (1990) Ancestral semiochemicalattraction persists for adjoining populations of sibling Ips barkbeetles (Coleoptera: Scolytidae). Journal of Chemical Ecology, 16,993–1013.
Carmer, S.G. & Swanson, M.R. (1973) Evaluation of ten pairwisemultiple comparison procedures by Monte Carlo methods. Journalof the American Statistics Association, 68, 66–74.
Cibrian-Tovar, D., Mendez-Montiel, J.T., Campos-Bolanos, R., Yates,H.O. & Flores-Lara, J. (1995) Insectos Forestales de Mexico.Universidad Autonoma Chapingo, Publication #6, Mexico.
Dahlsten, D.L. & Stephen, M. (1974) Natural enemies and insectassociates of the mountain pine beetle, Dendroctonus ponderosae(Coleoptera: Scolytidae), in sugar pine. Canadian Entomologist, 106,1211–1217.
Davis, T.S. & Hofstetter, R.W. (2009) Effects of gallery density andspecies ratio on the fitness and fecundity of two sympatric barkbeetles (Coleoptera: Curculionidae). Environmental Entomology, 38,639–650.
Davis, T.S., Hofstetter, R.W., Klepzig, K.D., Foster, J.T. & Keim, P.(2010) Sympatry and allopatry of beetle hosts influences thecompetitive interactions among fungal mutualists. Journal of Yeastand Fungal Research, 1, 118–126.
Dixon, W.N. & Payne, T.L. (1980) Attraction of entomophagousassociate insects of the southern pine beetle to beetle- and host tree-produced volatiles. Journal of Georgia Entomological Society, 15,378–389.
Erbilgin, N., Powell, J.S. & Raffa, K.F. (2003) Effect of varyingmonoterpene concentrations on the response of Ips pini (Coleoptera:Scolytidae) to its aggregation pheromone: implications for pestmanagement and ecology of bark beetles. Agricultural and ForestEntomology, 5, 269–274.
Fettig, C.J., McMillin, J.D., Anhold, J.A., Hamud, S.M., Borys, R.R.,Dabney, C.P. & Seybold, S.J. (2006) The effects of mechanical fuelreduction treatments on the activity of bark beetles (Coleoptera:Scolytidae) infesting ponderosa pine. Forest Ecology and Manage-ment, 230, 55–68.
Fukuda, A. (1962) Description of the larvae of Elacatis kraatze Reitter(Elacatidae, Coleoptera). Kontyu, 30, 17–21.
Gaylord, M.L., Kolb, T.E., Wallin, K.F. & Wagner, M.R. (2006)Seasonality and lure preference of bark beetles (Curculionidae:Scolytinae) and associates in a northern Arizona ponderosa pineforest. Environmental Entomology, 35, 37–47.
Gaylord, M.L., Hofstetter, R.W. & Wagner, M.R. (2010) Impacts ofsilvicultural thinning treatments on bark beetle trap captures andtree attacks during low bark beetle populations in ponderosa pineforests of northern Arizona. Journal of Economic Entomology, 103,1693–1703.
Hedden, R., Vite, J.P. & Mori, K. (1976) Synergistic effect of apheromone and a kairomones on host selection and colonization byIps avulsus. Nature, 261, 696–697.
Hofstetter, R.W., Chen, Z., Gaylord, M.L., McMillin, J.D. & Wag-ner, M.R. (2008) Synergistic effects of α-pinene and exo-brevicominon pine bark beetles and associated insects in Arizona. Journal ofApplied Entomology, 132, 387–397.
Kinzer, G.W., Fentiman, A.F., Page, T.F., Foltz, R.L., Vite, J.P. &Pitman, G.B. (1969) Bark beetle attractants: identification, synthesisand field bioassay of a new compound isolated from Dendroctonus.Nature, 221, 477–478.
Lanier, G.N. & Wood, D.L. (1975) Specificity of response topheromones in the genus Ips. Journal of Chemical Ecology, 1, 9–13.
Lanne, B.S., Schlyter, F., Byers, J.A. et al. (1987) Differences inattraction to semiochemicals present in sympatric pine shoot beetles,Tomicus minor and T. piniperda. Journal of Chemical Ecology, 13,1045–1067.
Lieutier, F., Day, K.R., Battisti, A., Gregoire, J.C. & Evans, H.F.(2004) Bark and Wood Boring Insects in Living Trees in Europe,a Synthesis. Springer, The Netherlands.
Miller, D.R. (1991) Reproductive and ecological isolation: commu-nity structure in the use of semiochemicals by pine bark beetles(Coleoptera: Scolytidae). PhD Thesis. Simon Fraser University.
Miller, D.R. (2001) Frontalin interrupts attraction of Ips pini(Coleoptera: Scolytidae) to ipsdienol. Canadian Entomologist, 133,407–408.
Miller, D.R. & Borden, J.H. (1990) B -Phellandrene: Kairomone forpine engraver, Ips pini (Say) (Coleoptera: Scolytidae). Journal ofChemical Ecology, 16, 2519–2531.
Miller, D.R. & Borden, J.H. (2000) Dose-dependent and species-specific responses of pine bark beetles (Coleoptera: Scolytidae) tomonoterpenes in association with pheromones. Canadian Entomolo-gist, 132, 183–195.
Miller, D.R., Borden, J.H. & Slessor, K.N. (1996) Enantiospecificpheromone production and response profiles for populations ofpine engravers, Ips pini (Say) (Coleoptera: Scolytidae), in BritishColumbia. Journal of Chemical Ecology, 22, 2157–2172.
Paine, T.D. & Hanlon, C.C. (1991) Response of Dendroctonus brevi-comis and Ips paraconfusus (Coleoptera: Scolytidae) to combina-tions of synthetic pheromone attractants and inhibitors verbenoneand ipsdienol. Journal of Chemical Ecology, 17, 2163–2176.
Paine, T.D., Birch, M.C. & Svihra, P. (1981) Niche breadth andresource partitioning by four sympatric species of bark beetles(Coleoptera: Scolytidae). Oecologia, 48, 1–6.
Payne, T.L. (1980) Life history and habits. The Southern Pine Beetle(ed. by R. C. Thatcher, J. L. Searcy, J. E. Coster and G. D. Hertel),pp. 7–28. USDA Forest Service, Technical Bulletin 1631, Washing-ton, District of Columbia.
Payne, T.L., Smith, M.T., Birch, M.C. & Ascoli, A. (1991) Interspe-cific olfactory communication in the southern pine bark beetle guild.Forest Insect Guilds: Patterns of Interaction with Host Trees (ed. byY. N. Baranchikov, W. J. Mattson, F. P. Hain and Payne, T. L.), pp.352–370. United States Department of Agriculture Forest Service,Radnor, Pennsylvania.
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215

Attraction of pheromones to beetles and predators 215
Poland, T.M. & Borden, J.H. (1994) Semiochemical-based communi-cation in interspecific interactions between Ips pini (Say) and Pityo-genes knechteli (Swaine) (Coleoptera: Scolytidae) in lodgepole pine.Canadian Entomologist, 126, 269–276.
Poland, T.M. & Borden, J.H. (1998) Competitive exclusion ofDendroctonus rufipennis induced by pheromone of Ips tridens andDryocoetes affabar (Coleoptera: Scolytidae). Journal of EconomicEntomology, 91, 1150–1161.
Raffa, K.F. (2001) Mixed messages across multiple trophic levels:the ecology of bark beetle chemical communication systems.Chemoecology, 11, 49–65.
Raffa, K.F. & Berryman, A.A. (1983) The role of host plant resistancein the colonization behavior and ecology of bark beetles (Coleoptera:Scolytidae). Ecological Monographs, 53, 27–49.
Rankin, L.J. & Borden, J.H. (1991) Competitive interactions betweenthe mountain pine beetle and the pine engraver in lodgepole pine.Canadian Journal of Forest Research, 21, 1029–1036.
Rao, P.V. (1998) Statistical Research Methods in the Life Sciences.Brooks/Cole Publishing Company, Pacific Grove, California.
Reeve, J.D. & Strom, B.L. (2004) Statistical problems encounteredin trapping studies of Scolytids and associated insects. Journal ofChemical Ecology, 30, 1575–1590.
Reeve, J.D., Strom, B.L., Rieske, L.K., Ayres, B.D. & Costa, A.(2009) Geographic variation in prey preference in bark beetle preda-tors. Ecological Entomology, 34, 183–192.
Renwick, A.A. & Vite, J.P. (1968) Isolation of the populationaggregating pheromone of the southern pine beetle. Contributionsfrom the Boyce Thompson Institute, 24, 65–68.
Renwick, J.A.A. & Vite, J.P. (1970) Systems of chemical communi-cation in Dendroctonus. Contributions from the Boyce ThompsonInstitute, 24, 283–292.
Renwick, J.A.A. & Vite, J.P. (1972) Pheromones and host volatilesthat govern aggregation of the six-spined engraver beetle, Ipscalligraphus. Journal of Insect Physiology, 18, 1215–1219.
Rudinsky, J.A. (1966) Observations of olfactory behavior of scolytidbeetles (Coleoptera: Scolytidae) associated with Douglas-fir forests.Zeitschrift fur Angeandte Entomologie, 58, 356–361.
Safranyik, L., Shripton, D.M. & Whitney, H.S. (1975) An interpreta-tion of the interaction between lodgepole pine, the mountain pinebeetle and its associated blue stain fungi in western Canada. Man-agement of Lodgepole Pine Ecosystems (ed. by D. M. Baumgartner),pp. 406–428. Washington State University Cooperative ExtensionService, Pullman, Washington.
Safranyik, L., Shore, T.L. & Linton, D.A. (1996) Ipsdienol andlanierone increase Ips pini Say (Coleoptera: Scolytidae) attack andbrood density in lodgepole pine infested by mountain pine beetle.Canadian Entomologist, 128, 199–207.
Sanchez-Martinez, G. & Wagner, M.R. (2002) Bark beetle communitystructure under four ponderosa pine forest stand conditions innorthern Arizona. Forest Ecology and Management, 170, 145–160.
Seybold, S.J., Teale, S.A., Wood, D.L., Zhang, A.J., Webster, F.X.,Lindahl, K.Q. Jr & Kubo, I. (1992) The role of lanierone in thechemical ecology of Ips pini (Coleoptera: Scolytidae) in California.Journal of Chemical Ecology, 18, 2305–2329.
Seybold, S.J., Ohtsuka, T., Wood, D.L. & Kudo, I. (1995) The Enan-tiomeric composition of ipsdienol: a chemotaxonomic charac-ter for North American populations of Ips spp. in the pinigroup (Coleoptera: Scolytidae). Journal of Chemical Ecology, 21,995–1016.
Seybold, S.J., Huber, D.P.W., Lee, J.C., Graves, A.D. & Bohlmann, J.(2006) Pine monoterpenes and pine bark beetles: a marriage of con-venience for defense and chemical communication. PhytochemistryReview, 5, 143–178.
Smith, M.T., Paine, T.L. & Birch, M.C. (1990) Olfactory-based behav-ioral interactions among five species in the southern pine bark beetlegroup. Journal of Chemical Ecology, 16, 3317–3331.
Steed, R.E. & Wagner, M.R. (2008) Seasonal pheromone responseby Ips pini in northern Arizona and western Montana, U.S.A.Agriculture and Forest Entomology, 10, 189–203.
Sullivan, B.T. & Mori, K. (2009) Spatial displacement of release pointcan enhance activity of an attractant pheromone synergist of a barkbeetle. Journal of Chemical Ecology, 35, 1222–1233.
Svihra, P. (1982) Influence of opposite sex on attraction produced bypioneer sex of four bark beetle species cohabiting pine in the southernUnited States. Journal of Chemical Ecology, 8, 373–378.
Svihra, P., Paine, T.D. & Birch, M.C. (1980) Interspecific olfactorycommunications in southern pine beetles. Naturwissenschaften, 67,518–519.
Symonds, M.R.E. & Elgar, M.A. (2004) Species overlap, speciationand the evolution of aggregation in bark beetles. Ecology Letters, 7,202–212.
Teale, S.A., Webster, F.X., Zhang, A. & Lanier, G.N. (1991) Lanierone:a new pheromone component from Ips pini (Coleoptera: Scolytidae)in New York. Journal of Chemical Ecology, 17, 1159–1176.
USDA Forest Service (2002–2010) Forest Insect and disease conditionsin the United States. Research Health Protection. United StatesDepartment of Agriculture Forest Service [WWW document]. URLhttp://www.fs.fed.us/r3/resources/health/fid_surveys.shtml [accessedin 2010].
Vite, J.P. & Gara, R.I. (1962) Volatile attractants from ponderosa pineattacked by bark beetles (Coleoptera: Scolytidae). Contributions fromthe Boyce Thompson Institute, 21, 251–273.
Vite, J.P., Gara, R.I. & van Sheller, H.D. (1964) Field observations onthe response of bark beetles infesting southern pines. Contributionfrom the Boyce Thompson Institute, 22, 461–470.
Vite, J.P., Bakke, A. & Renwick, J.A.A. (1972) Pheromones in Ips(Coleoptera: Scolytidae): occurrence and production. CanadianEntomologist, 104, 1967–1975.
Vite, J.P., Ohloff, G. & Billings, R.F. (1978) Pheromonal chirality andintegrity of aggregation response in southern species of the barkbeetle Ips sp. Nature, 272, 817–818.
Wallin, K.F., Kolb, T.E., Skov, K.R. & Wagner, M.R. (2008) Forestmanagement treatments, tree resistance, and bark beetle resourceutilization in ponderosa pine forests of northern Arizona. ForestEcology and Management, 255, 3263–3269.
Williams, K.K., McMillin, J.D., DeGomez, T.E., Clancy, K.M. &Miller, A. (2008) Influence of elevation on bark beetle (Coleoptera:Curculionidae, Scolytinae) community structure and flight periodicityin ponderosa pine forests of Arizona. Environmental Entomology, 37,94–109.
Williams, K.K., McMillin, J.D. & DeGomez, T.E. (2009) Relative andseasonal abundance of three bark beetle predators (Coleoptera:Trogositidae, Cleridae) across an elevation gradient in ponderosapine forests of north central Arizona. Western North AmericanNaturalist, 69, 351–363.
Wood, S.L. (1982) The Bark and Ambrosia Beetles of North and CentralAmerica (Coleoptera: Scolytidae), a Taxonomic Monograph. GreatBasin Naturalist Memoirs, Vol. 6. Brigham Young University, Provo,Utah.
Wood, D.L., Browne, L.E., Ewing, B. et al. (1976) Western pine bee-tle: specificity among enantiometers of male and female componentsof an attractant pheromone. Science, 192, 896–898.
Accepted 30 October 2011First published online 31 January 2012
© 2012 The Authors
Agricultural and Forest Entomology © 2012 The Royal Entomological Society, Agricultural and Forest Entomology, 14, 207–215

Related Documents











