Eur. J. Biochem. 229,494-502 (1995) 0 FEBS 1995 Assignment and secondary-structure determination of monomeric bovine seminal ribonuclease employing computer-assisted evaluation of homonuclear three-dimensional ‘H-NMR spectra Annamaria D’URSI’, Hartmut OSCHKINAT’, Christian CIESLAR’, Delia PICONE’, Giuseppe D’ALESSI04, Pietro AMODEO’ and Piero A. TEMUSSI’ ’ Dipartimento di Chimica, Universiti di Napoli Federico 11, Napoli, Italy ’ EMBL, Heidelberg, Germany ’ MPI fur Biochemie, Martinsried bei Munchen, Germany Dipartimento 1 Chimica Organica e Biologica, Universiti di Napoli Federico 11, Napoli, Italy (Received 6 February 1995) - EJB 95 0179/3 Monomeric bovine seminal ribonuclease (mBS-RNase), the subunit of dimeric bovine seminal ribo- nuclease (BS-RNase), is an unusual monomer: for its structural stability, its catalytic activity, which is even higher than that of the parent dimeric enzyme, and for its role as an intermediate in the refolding of dimeric BS-RNase. Here we present the proton NMR assignment and secondary-structure determina- tion of mBS-RNase, with a comparison of its structure to the structure of its parent protein, and to the structure of RNase A, a homologue with more than 80% identity in amino acid sequence. Proton NMR assignment was performed using a computer-assisted procedure, through a partially automated analysis of homonuclear three-dimensional spectra [Oschkinat, H., Holak, T. A. & Cieslar, C. (1991) Biopolymers 31, 699-7121. The secondary structures of mBS-RNase, of the A chain of dimeric BS-RNase, and of RNase A, are found to be similar. Significant differences are found instead, between mBS-RNase and RNase A in the more flexible stretches of the molecule, where a higher number of substitutions is present. Furthermore, a preliminary tertiary-structure model is reported, showing that the overall folding of mBS- RNase is closer to that of RNase A rather than that of (dimeric) BS-RNase. Keywords. NMR ; automatic assignment ; protein conformation ; ribonucleases. Bovine seminal ribonuclease (BS-RNase, see [l] for a re- view), produced in bull seminal vesicles, is endowed, like other RNases of the RISBASE group [l], with special, i.e. non-cata- lytic, biological activities, including an anti-tumor action, re- cently confirmed in vivo [2]. BS-RNase is the only dimeric pro- tein of the pancreatic-type super-family, with the subunits linked by two disulfide bridges and non-covalent forces 13, 41. BS- RNase subunit (mBS-RNase) is homologous to bovine pancre- atic RNase A (RNase A), with a primary structure more than 80 % identical, including the amino acid residues found at the active site of RNase A [5]. The enzyme is isolated as a mixture of three isoenzymic forms, az, ap and p2, generated by the spon- taneous partial transformation of p-type into a-type subunits. This consists in the selective deamidation of the Am67 of p- type subunit, which may transform into Asp or PAsp, as the Correspondence to A. D’Ursi, Dipartimento di Chimica, UniversitB Fax: +39 81 5527771. Abbreviations. BS-RNase, bovine seminal ribonuclease; DQF- COSY, double quantum filtered correlation spectroscopy; mBS-RNase, monomeric bovine seminal ribonuclease ; NOE, nuclear Overhauser ef- fect; NOESY, nuclear Overhauser effect spectroscopy ; RNase A, bovine pancreatic ribonuclease ; TOCSY, total correlation spectroscopy ; TPPI, time proportional phase incrementation. ; 2D, two-dimensional ; 3D, three-dimensional ; SA, simulate annealing. di Napoli Federico 11, via Mezzocannone 4, 1-80134 Napoli, Italy Enzyme. Ribonuclease (EC 3.1.27.5). cyclic imide intermediate involving Gly68 may spontaneously hydrolyze on either side of the nitrogen [6]. In the structure of BS-RNase obtained by X-ray crystallogra- phy [7, 81 and recently refined to 0.19 nm [9], the two subunits have been found to exchange their N-terminal segments, a struc- tural motif reminiscent of that proposed by Crestfield [lo] for dimerized RNase A. Recently, it has been found that two quater- nary structures, in equilibrium with each other, are accessible to native BS-RNase : one, characterized by the above-mentioned exchange of N-terminal segments; the other in which no ex- change between subunits takes place [ll]. Only the structure with exchange of the N-terminal ends displays the allosteric ki- netics characteristic of native BS-RNase [12], apparently for the composite nature of the catalytic sites resulting from the pres- ence, at each catalytic site, of residues essential to catalysis and located, in the exchanged N-terminal ends, such as Hisl2, and in the main subunit body, such as Hisll9. No data are available for the structure of BS-RNase charac- terized by the absence of exchange of N-terminal ends, a struc- ture in which the two subunits associate as independent mono- mers, nor for mBS-RNase, the isolated monomer of BS-RNase. It should be stressed that mBS-RNase is an unusual monomer: for its structural stability [4], its catalytic activity, which is even higher than that of the parent dimeric enzyme [12], and for its role as an intermediate in the refolding of dimeric BS-RNase [13]. The availability of structural data on mBS-RNase would thus not only provide useful information on the monomeric form

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Biochem. 229,494-502 (1995) 0 FEBS 1995
Assignment and secondary-structure determination of monomeric bovine seminal ribonuclease employing computer-assisted evaluation of homonuclear three-dimensional ‘H-NMR spectra Annamaria D’URSI’, Hartmut OSCHKINAT’, Christian CIESLAR’, Delia PICONE’, Giuseppe D’ALESSI04, Pietro AMODEO’ and Piero A. TEMUSSI’
’ Dipartimento di Chimica, Universiti di Napoli Federico 11, Napoli, Italy ’ EMBL, Heidelberg, Germany ’ MPI fur Biochemie, Martinsried bei Munchen, Germany
Dipartimento 1 Chimica Organica e Biologica, Universiti di Napoli Federico 11, Napoli, Italy
(Received 6 February 1995) - EJB 95 0179/3
Monomeric bovine seminal ribonuclease (mBS-RNase), the subunit of dimeric bovine seminal ribo- nuclease (BS-RNase), is an unusual monomer: for its structural stability, its catalytic activity, which is even higher than that of the parent dimeric enzyme, and for its role as an intermediate in the refolding of dimeric BS-RNase. Here we present the proton NMR assignment and secondary-structure determina- tion of mBS-RNase, with a comparison of its structure to the structure of its parent protein, and to the structure of RNase A, a homologue with more than 80% identity in amino acid sequence. Proton NMR assignment was performed using a computer-assisted procedure, through a partially automated analysis of homonuclear three-dimensional spectra [Oschkinat, H., Holak, T. A. & Cieslar, C. (1991) Biopolymers 31, 699-7121. The secondary structures of mBS-RNase, of the A chain of dimeric BS-RNase, and of RNase A, are found to be similar. Significant differences are found instead, between mBS-RNase and RNase A in the more flexible stretches of the molecule, where a higher number of substitutions is present. Furthermore, a preliminary tertiary-structure model is reported, showing that the overall folding of mBS- RNase is closer to that of RNase A rather than that of (dimeric) BS-RNase.
Keywords. NMR ; automatic assignment ; protein conformation ; ribonucleases.
Bovine seminal ribonuclease (BS-RNase, see [ l ] for a re- view), produced in bull seminal vesicles, is endowed, like other RNases of the RISBASE group [l], with special, i.e. non-cata- lytic, biological activities, including an anti-tumor action, re- cently confirmed in vivo [2]. BS-RNase is the only dimeric pro- tein of the pancreatic-type super-family, with the subunits linked by two disulfide bridges and non-covalent forces 13, 41. BS- RNase subunit (mBS-RNase) is homologous to bovine pancre- atic RNase A (RNase A), with a primary structure more than 80 % identical, including the amino acid residues found at the active site of RNase A [5]. The enzyme is isolated as a mixture of three isoenzymic forms, az, ap and p2, generated by the spon- taneous partial transformation of p-type into a-type subunits. This consists in the selective deamidation of the Am67 of p- type subunit, which may transform into Asp or PAsp, as the
Correspondence to A. D’Ursi, Dipartimento di Chimica, UniversitB
Fax: +39 81 5527771. Abbreviations. BS-RNase, bovine seminal ribonuclease; DQF-
COSY, double quantum filtered correlation spectroscopy; mBS-RNase, monomeric bovine seminal ribonuclease ; NOE, nuclear Overhauser ef- fect; NOESY, nuclear Overhauser effect spectroscopy ; RNase A, bovine pancreatic ribonuclease ; TOCSY, total correlation spectroscopy ; TPPI, time proportional phase incrementation. ; 2D, two-dimensional ; 3D, three-dimensional ; SA, simulate annealing.
di Napoli Federico 11, via Mezzocannone 4, 1-80134 Napoli, Italy
Enzyme. Ribonuclease (EC 3.1.27.5).
cyclic imide intermediate involving Gly68 may spontaneously hydrolyze on either side of the nitrogen [6].
In the structure of BS-RNase obtained by X-ray crystallogra- phy [7, 81 and recently refined to 0.19 nm [9], the two subunits have been found to exchange their N-terminal segments, a struc- tural motif reminiscent of that proposed by Crestfield [lo] for dimerized RNase A. Recently, it has been found that two quater- nary structures, in equilibrium with each other, are accessible to native BS-RNase : one, characterized by the above-mentioned exchange of N-terminal segments; the other in which no ex- change between subunits takes place [l l] . Only the structure with exchange of the N-terminal ends displays the allosteric ki- netics characteristic of native BS-RNase [12], apparently for the composite nature of the catalytic sites resulting from the pres- ence, at each catalytic site, of residues essential to catalysis and located, in the exchanged N-terminal ends, such as Hisl2, and in the main subunit body, such as Hisll9.
No data are available for the structure of BS-RNase charac- terized by the absence of exchange of N-terminal ends, a struc- ture in which the two subunits associate as independent mono- mers, nor for mBS-RNase, the isolated monomer of BS-RNase. It should be stressed that mBS-RNase is an unusual monomer: for its structural stability [4], its catalytic activity, which is even higher than that of the parent dimeric enzyme [12], and for its role as an intermediate in the refolding of dimeric BS-RNase [13]. The availability of structural data on mBS-RNase would thus not only provide useful information on the monomeric form

D'Ursi et al. ( E m J. Biochem. 229) 495
of BS-RNase, it would also provide insights into the structure of native dimeric BS-RNase made up of independent monomers which do not exchange their N-terminal segments.
Here we report the secondary-structure determination in so- lution of mBS-RNase by NMR spectroscopy using a combina- tion of homonuclear two-dimensional (2D) and three-dimen- sional (3D) NMR techniques. Since our material was isolated from natural sources, we used exclusively homonuclear 3D ex- periments, performing the complete proton assignment with the aid of an automatic method. The automatic assignment program based on homonuclear 3D spectra had already been tested on bovine pancreatic trypsin inhibitor [ 141, but it seemed interesting to try the same approach on mBS-RNase, whose size is at the limit of those normally solved by 2D methods.
MATERIALS AND METHODS
Protein purification. BS-RNase was purified from bull seminal vesicles as previously described [15]. To avoid side- reactions (involving the spontaneous deamidation of Asn67) during NMR data acquisition, Asn67 of native dimeric BS- RNase (2 mM) was selectively deamidated by incubation at 37"C, pH 8.4 [6], giving rise to two [Asp671 isomers, the first one (70%) containing a normal peptide bond, the other (30%) containing an isopeptide bond between Asp67 and Gly68. mBS- RNase was prepared by selective reduction of the interchain di- sulfides and carboxymethylation of the exposed sulfydryls as previously described [4].
NMR measurements. Samples for NMR measurements were prepared by dissolving 20 mg protein either in 500 pl H,Ol D,O (90: 10, by vol.) or 500 ml D,O. The pH was set to 5 with HCI. Measurements were performed on Bruker AM-400, AM- 500, and AMX-600 spectrometers at 300 K. Time proportional phase incrementation (TPPI) was applied to achieve quadrature detection in the virtual dimension [16].
One-dimensional protonic experiments were performed at different times, ranging from 5 min up to 2 weeks. Correlation spectroscopy (COSY) double quantum filtered (DQF-COSY) [17] and NOE spectroscopy (NOESY) [18] spectra were re- corded 2 h and 2 weeks after sample dissolution in D,O.
A set of two-dimensional spectra was recorded; COSY, total correlation spectroscopy (TOCSY) [19], with a 27-ms mixing time and NOESY (90-ms mixing time) all with presaturation; TOCSY and NOESY with a jump-return read pulse for water suppression [20]. The 2D spectra were typically recorded with a spectral width of 8333 Hz, 102412048 data points were ac- quired in t&,.
3D-TOCSY-TOCSY [21] and TOCSY-NOESY [22] spectra were also recorded; the TOCSY-TOCSY pulse sequence was implemented with two different spin lock sequences; MLEV 17 and MLEV 4 [14]. The length of each mixing time was 20-ms. A four-step phase cycle was employed with 256124011024 data points in t,lt,lt3 with four scanslincrement. The spin lock used for the TOCSY-NOESY experiment was also MLEV 17. Water suppression was achieved with a 1-1 pulse at the end of the sequence; an eight-step phase cycle was used. The experiment was recorded with 200/208/1024 data points in tlltZlt3 with eight scans for increments. 3D spectra were processed in subcubes of 512151211024 data points used for the whole spectrum. Linear prediction was applied to predict data points up to double the valve of the recorded ones, and extensive baseline correction was applied to all three dimensions.
Automatic assignment procedure. The number of 3D cross-peaks contained in a 3D spectrum makes manual evalua- tion a tedious procedure. Therefore, the 3D cross-peak pattern
has been recognized by a specific program, based on subsequent automatic evaluation of TOCSY-TOCSY and TOCSY-NOESY data, as described in greater detail in [14].
Spin system analysis. In the automated procedure 3D TOCSY-TOCSY is used to find the sets of signals belonging to individual amino acids. The aim is to assign the NH, C,H, and C,H resonances, and to find the chemical shifts of the other protons belonging to the same coupling network. The program consists of five steps. The first step is a simple clean up of the peak lists to remove artifacts from fllt2 noise and those around the signal of the residual water. In the second step, all symmetri- cal cross-peaks are treated as one. The simplified cross-peaks are subjected to the third step, which leads to generation of structures with four different chemical shifts, with the condition that each chemical shift is contained in at least two different 3D cross-peaks. The fourth step is the construction of clusters connecting all sets of four different chemical shifts that have one cross-peak in common. The fifth step consists of the inspec- tion of the clusters, involving the control of the chemical shifts, and the interpretation of the peak amplitudes to find the NH, C,H, and C,H resonances. The final result is a list containing all residues with at least four chemical shifts.
Secondary structure recognition. In the analysis of protein NMR spectra, secondary-structure recognition is usually coinci- dent with sequential assignment, performed through the evalua- tion of amplitudes of characteristic cross-peaks in NOESY spectra. It has been shown that 3D TOCSY-NOESY spectra con- tain even more characteristic patterns. This allows discrimina- tion between different possible connectivities with greater cer- tainty.
For P-strand and P-sheet-like conformations, NOE involving the distance d,, are most significant. They give rise to two very strong cross-peaks of the type C,H(i+ l)/NH(i+ 1 jIC<H(i) and NH(i)/C,H(ij/NH(i+l) in F,IFZIF,, which yield the sequential assignment in a straightforward manner. Tight turns and a heli- ces give rise to cross-peaks of the type C,H(i+I)/NH(i)l NH(i+l) These peaks however, are expected to be rather weak for residues located in helices, for which connectivities of the type C,WC,H/NH are more likely to be found. In the same area of the spectrum, cross-peaks indicating the transfer C,H/C,W NH are expected to occur. Obviously, areas of proteins without a regular secondary structure may show all three types of these cross-peaks or none.
Sequential assignment. The residues are characterized by the frequencies of NH, C,H, and C,,H, the information of the type of spin system, and a number for a more convenient character- ization later in the output of the result. First, pairs of residues are formed utilizing the interresidue cross-peaks given in a list. Next, small sequences are made up whenever pairs with the same amino acid are found at the beginning and end of the small sequence. If there are two different possible connections, both solutions are kept. The distinction between good and bad partial sequences is made in the last step, when the partial sequences are fitted to the amino acid sequence of the protein. With increasing length of the partial sequences the number of solu- tions found are reduced and bad partial sequences no longer fit into the sequence.
Selection of the results is performed in a simple manner: in every solution the number of connections is counted and those solutions that contain the longer partial sequences are, therefore, rated higher.
Structure calculation. Preliminary 3D structure refinement has been performed by using only 630 NOE involving backbone NH. The X-ray structure of the A chain of BS-RNase [9] has been partially energy minimized, applying only short range NOE-derived distance restraints. The folding of the N-terminal

496 D’Ursi et al. (EUK J. Biochern. 229)
region has been obtained by a 120-ps simulated annealing (SA) simulation [23] performed by rigidly restraining the position of the residues following Tyr25, and including long-range restraints involving the first 25 residues. A final 100-ps molecular dynam- ics simulation, without position restraints from Cys26 on, but with full application of the whole set of NOE-derived restraints has led to the tentative structure discussed below.
RESULTS AND DISCUSSION
Resonance assignment. The strategy that allowed full proton resonance assignment of mBS-RNase consists of a combination of the automated procedure described in Materials and Methods and of a standard, manual analysis to verify and complete the automated procedure.
In the automated procedure, 3D TOCSY-TOCSY evaluation employed peak lists from two different parts of the spectrum, one comprising the aliphatic region in all three dimensions, the other comprising the aliphatic region in F1 and F2, and the NH region in F3. This led to the identification of 60% of the spin systems. The remaining spin systems could not be automatically identified owing to some specific problems: the primary struc- ture of mBS-RNase includes a high number of long-chain resi- dues, such as Lys and Arg, for which side-chain signals were often severely overlapped, so that it was not easy to discriminate automatically between different spin systems to produce dif- ferent clusters; the short mixing times used in the 3D TOCSY- TOCSY prevented the observation of 3D peaks in most cases of normally weak signals. A manual comparison with 2D TOCSY and COSY data, using lists from automated peak picking, was helpful to identify many of the Arg and Lys residues and some Pro, Ile, Leu.
The poor definition of some spin systems in the TOCSY- NOESY spectrum due to the reasons previously described, led us to follow, in the second part of the automatic procedure, a slightly different scheme. The output of the analysis of the TOCSY-NOESY did not include many long sequences of well connected amino acid residues, but rather a high number of short pairs, connected by 3D cross-peaks of significant intensity. The pairs were submitted to an accurate checking step, based on evaluation of cross-peaks connection and on a survey of the con- sistency. Subsequently, the best pairs, judged from their reliabil- ity, were fitted in the sequence in a preliminary step, to have the correct sequential assignment.
To complete the sequential assignment, it proved helpful to employ an interactive procedure using both 3D spectra with the automatic program and 2D spectra with a manual analysis. This was necessary because the exclusive use of homonuclear 3D spectra implies some specific difficulties : problems linked to computer memory and the high number of peaks present in 3D spectra may lead to complete deletion of noisy regions of 3D spectra; in 3D spectra the peaks normally have lower intensity than in 2D spectra. The consequence is that the automated analy- sis is not very helpful for residues belonging to regions of the protein with no regular secondary structure, because many sig- nals of these residues, that are also generally weak in 2D spectra, are often undetectable in 3D spectra. The identification of helical strands is also problematic due to the weakness of NH-C,H con- nectivities and to the higher resolution required to detect NH- NH effects.
Thus, the automated procedure led to the assignment of most of the B sheet and some loop regions of the protein; the remain- ing part containing the helical segments and the less regular parts requested the use of basically manual work. Table 1 sum- marizes all chemical-shift assignments.
Table 1. ‘H chemical shifts of the assigned proton resonances of mo- nomeric bovine seminal ribonuclease at pH 5.0 and 300 K. Chemical shifts are referenced to the water resonance at 4.70 ppm, and are accurate to -+-0.01 ppm. (a) and (b) indicate the observable split resonances of the protein containing Asp or PAsp at position 67, respectively.
Re- Chemical shift of sidue
NH aCH PCH, G L others
PPm
K1 E2 s 3 A4 A5 A6 K7 F8 E9 R10 Q11 H12 MI3 D14 S15 GI 6 D17 S18 PI9 s20 s21 s22 S23 N24 Y25 C26 N27 L28 M29 M30 C3 1 C32 R33 K34 M35 T36 Q37 G38 K39 C40 K41
P42 v43 N44 T45 F46 v47 H48 Ex9 S50 L5 1 A52 D53 v54 K55 A56 v57 C58 s59 Q60
8.52 8.87 8.98 8.65 8.04 8.74 8.04 7.93 8.37 8.41 7.82 8.02 8.76 8.94 8.53 7.95 8.28
8.09 8.37 8.26 8.48 8.48 7.68 7.78 7.78 7.47 8.40 8.72 6.66 8.86 8.17 7.71 8.21 7.56 7.27 8.34 7.22 8.87 7.70
6.96 8.78 7.14 9.33 9.34 8.85 6.61 9.59 8.79 8.47 7.84 7.88 9.00 7.55 7.58 6.81 7.50 7.72
4.39 2.0711.91 4.48 2.7612.64 4.20 1.56 4.35 1.47 4.20 1.58 4.02 2.0011.81 4.42 3.4512.95 3.73 2.1312.24 4.17 2.0311.98 3.79 1.9911.33 4.92 2.5311.75 5.46 2.7812.20 4.98 2.3112.33 4.22 3.8513.71 4.0513.90 4.67 2.611263 4.26 3.6513.61 4.17 1.5911.20 4.30 3.7113.60 4.36 3.9213.89 4.55 4.0013.88 4.48 4.0913.97 4.94 2.9312.78 3.91 3.2812.76 3.77 3.0712.1 1 4.40 23612.90 4.04 1 S911.57 4.36 1.3410.86 4.26 1.3710.48 4.60 3.0913.01 4.70 3.0312.16 4.43 1.9011.88 4.04 1.8611.27 4.56 2.3512.13 5.21 4.84 4.51 2.0412.02 4.2513.20 3.54 1.6211.49 4.39 3.2012.97 4.40 2.2111.19
4.61 2.4112.09 5.29 2.11 4.76 2.3412.07 5.14 2.20 4.88 2.1212.59 4.22 2.50 5.32 3.42 4.76 1.4611.44 4.22 4.2514.05 4.08 1.7711.75 3.94 1.37 4.35 2.9312.45 3.69 2.16 3.62 1.8111.17 3.99 1.44 3.18 2.17 3.51 2.6612.52 4.57 3.9213.90 4.56 2.4112.37
2.4712.07
1.4111.29
2.9612.37 1.6211.54 2.7011.10
2.3112.21
1.5111.53
1.43 2.3212.30 2.3611.70
2.3712.39
1.18 2.4012.34
1.1110.92
11921190
2.1 812.16 0.9710.95
0.72
1.0210.91
2.5512.53
1.59
1.0911.06 1.6911.63
1.1110.72
2.6912.67
eCH, 3.1213.23 67.011~ 6.84117.00
XH, 3.22l3.08
C,H 7.751C4H 6.72
NH, 7.8316.80
SCH, 3.2213.13
NH, 7.5811.12 66.9916.43
NH, 7.6316.95 XH, 0.8010.79
6 1.4U1.12
6CH, 1.4411.12 ; EE’CH, 3.02 SCH, 4.0513.91
6 6.8Uc 6.90lc 6.51
C,H 8.00lC4H 6.62
X H , : 1.0910.99

D'Ursi et al. (EUK J. Biochem. 229) 497
Table 1. Continuation
Re- Chemical shift of sidue
NH K H PCH, yCH, others
PPm
K61 7.55 4.24
K62 V63 T64 C65 C65 K66 K66
8.42 4.05 8.33 4.49 7.97 4.26
(a) 8.81 4.44 @) 4.46 (a) 10.91 3.99 (b) 11.13 3.97
1.9311.80 1.3811.19 JCH, 1.1211.10; eCH, 2.9612.84
1.6411.62 1.5311.51 X H , 1.1911.11 2.11 0.6910.39 3.86 1.26 3.4012.89 3.3212.83 1.9211.78 1.5411.49 1.8911.75 1.51
D67 G68 Q69 T70 N71 C72 Y73 Q74 s75 K76
s77 T78 M79 R80 I81 T82 D83 C84 R85 E86 T87 G88 S89 S90 K91 Y92 P93 N94 c95 A96 Y97 K98 T99 TlOO QlOl v102 El03 K104 H105 I106 I107 V108 A109 CllO G l l l G112 K113 PI14 S115 V116 PI17 V118 H119 F120 D121
8.01 8.02 8.1 1 8.56 8.22 7.44 8.95 9.48 9.30 8.94
8.60 8.15 8.52 9.50 8.69 9.29 9.27 8.78 8.21 8.73 8.12 8.79 7.62 7.01 7.55 9.30
8.91 7.88 8.56 9.38 9.45 9.07 8.83 8.83 8.66 8.08 8.35 9.20 8.47 8.32 8.90 8.61 8.78 9.12 8.22 7.78
8.21 8.51
8.95 7.65 9.18 8.62
4.44 2.9812.57 4.2413.47 4.33 2.1411.93 4.48 4.39 4.84 3.0212.82 5.44 2.2612.06 5.09 2.9212.15 5.40 1.8711.67 4.70 4.7214.80 4.18 13511.83
4.87 3.9213.73 3.11 3.65 4.63 1.8311.81 4.36 1.9211.72 5.29 1.81 5.03 3.92 5.00 2.5712.46 5.92 2.8912.54 5.42 1.9411.83 4.29 2.0312.01 4.50 4.47 3.9613.90 4.48 3.8113.77 3.93 3.8213.61 4.41 1.7111.69 3.73 3.3912.76 3.08 1.1311.11 4.96 2.7012.68 4.82 2.9212.90 4.70 1.15 4.88 2.4912.39 4.74 1.9111.74 5.21 4.23 4.63 4.08 5.18 2.0711.93 4.61 2.04 5.42 1.8311.66 4.78 2.2711.39 4.41 3.2013.14 4.99 1.76 5.28 1.45 4.83 2.13 5.28 1.45 5.67 3.2912.14 4.4813.89 4.4313.65 4.37 1.6911.61 4.59 2.3111.85 3.94 3.5913.57 4.86 2.08 4.51 0.9410.13 4.51 2.26 5.34 3.5712.99 4.26 2.6312.56 4.63 2.2512.05
2.3612.13 1.16
6 6.701~ 6.55 2.4411.68
1.4911.47 XH, 1.7111.69; ECH, 2.95
0.72
1.7011.68 XH, 3.2913.12 1.2510.92 yCH, 0.80I6CH3 0.73 1.25
1.7911.70 6CH, 3.2913.22 2.2812.25 1.09
1.3411.32
1.3811.36 3.3513.27 Ar : 6 6.891~ 6.96
NH, 7.6916.92
6 6.4618 7.02 1.4311.34 1.32 1.23 2.34: 2.31 0.90 : 0.80 2.1 112.00 1.5911.57
1.5611.42 yCH, 1.02 6CH3 1.00 1.1311.21 yCH, 0.73 yCH, 0.62 0.7610.65
C,H 8.521C4H 7.35
1.5911.31 E 2.93 1.8411.73 Z H , 3.5813.43
0.8310.81 1.5211.50 6CH, 3.6013.34 0.7710.57
C,H 7.02 6 6.62 E 6.97 < 6.64
Table 1. Continuation
Re- Chemical shift of sidue -
NH aCH PCH, $H, others
PPm
A122 7.48 4.47 1.33 S123 8.08 5.32 3.8313.77 V124 8.70 4.31 1.99 0.7510.78
t JP6 I (Asp 67 isomer)
P6 (iso-Asp 61 isomer)
I I l l
11.0 10.0
w2 (PPm
Fig. 1. Portion of 600-MHz NOESY spectrum of mBS-RNase in H,O/ D,O (90:lO; by vol.) at pH5.0, 300K; mixing time 100 ms. The spectral region shows the cross-peaks connected to Lys66 NH. They are split in two different sets, originating from the deamidation of the protein at Asn67, that leads to both a normal and an isopeptide Asp67-Gly68 bond.
As the region around 4.70 ppm is partially bleached by the irradiation of the water resonance, it was not possible to observe the spin systems of Glu49 and Ala96 that have the frequency of the C,H, respectively, at 4.76 pprn and 4.70 ppm, and for Ser75 that has C,H and CpH2, at 4.70 pprn and 4.72 pprn, respectively. For residues adjacent to Cys31 and Cys32, that do not have strong signals, and do not show a pattern of effects indicating a regular secondary structure, no connections were found in the 3D spectra from which the sequential assignments were derived.
Fig. 1 contains a portion of a 600-MHz NOESY spectrum of mBS-RNase in H20, showing the splitting of the NH proton of Lys66 into two signals (10.91 ppm and 11.13 ppm) of different intensities ; the two signals originate from the deamidation of

498 D'Ursi et al. (Eul: J. Biochern. 229)
I A d
c*4 'I
Q
J
8 I I I I I 5.5 5.0 4.5 4.0 3.5 3.0
. 3.0
3.5
4.0
4.5 r\
8 a v
3- 5.0
5.5
0 2 (PPm)
Fig.2. Partial 400-MHz NOESY spectrum of mBS-RNase in D,O at pH 5.0, 300 K ; mixing time 100 ms. This spectral region contains some of the crucial effects (labeled) for the assignment of proline isomerism as well as some diagnostic interchain C,H-C,H cross-peaks.
Asn67 that leads to two different isomers, containing a normal Asn67-Gly68 peptide bond and an isopeptide bond in different amounts [6]. The possibility of observing this isomerism is fa- vored by the fact that the low-field region showing the splitting of the NH proton of Lys66 is free from other resonances, whereas the direct observation of splittings on signals of Asp67 is precluded by severe resonance overlaps, in particular by the accidental superposition of the NH resonances of Asp67 and Gly68. The splitting of the resonances is also detectable for the side chain of Lys66 and for C,H and C,WC,H of Cys65, so that not only the residues immediately adjacent to Asp67 are involved.
Manual work was also necessary to observe typical NOE for the stereospecific assignment [24] of four of the five proline residues. Fig. 2 shows a region of a D,O 2D NOESY spectrum containing some of the crucial effects for the assignment of pro- line isomerism as well as some diagnostic interchain effects. On the basis of C,H-C,H (i , i+l) and NH-C,aH (i,i+l) cross-peaks, we assigned cis isomerism to prolines 93, 114, while trans iso- merism was observed for Proll7, on the basis of C,H-C,H, (i,i+l) and NH-C,H, (i,i+l) NOE, consistently with the find- ings for RNAse A [25-301 and of BS-RNase [7, 81. Prol9, which is not present in RNAse A, was also assigned trans iso- merism. In the case of Pro42, we were not able to detect une- quivocally either the cross-peaks diagnostic for the cis or trans isomerism, because C,H-C,H (Lys41 -Pro42) and NH-C,H (Lys41-Pro42) effects fall in a region of severe resonance over- laps. A preference in favor of trans isomerism is furnished by the agreement with the structure of RNase A and of BS-FWase.
Secondary Structure. A qualitative analysis of NOE can suffice to identify the secondary structure of the whole protein and to make a detailed comparison with the corresponding secondary
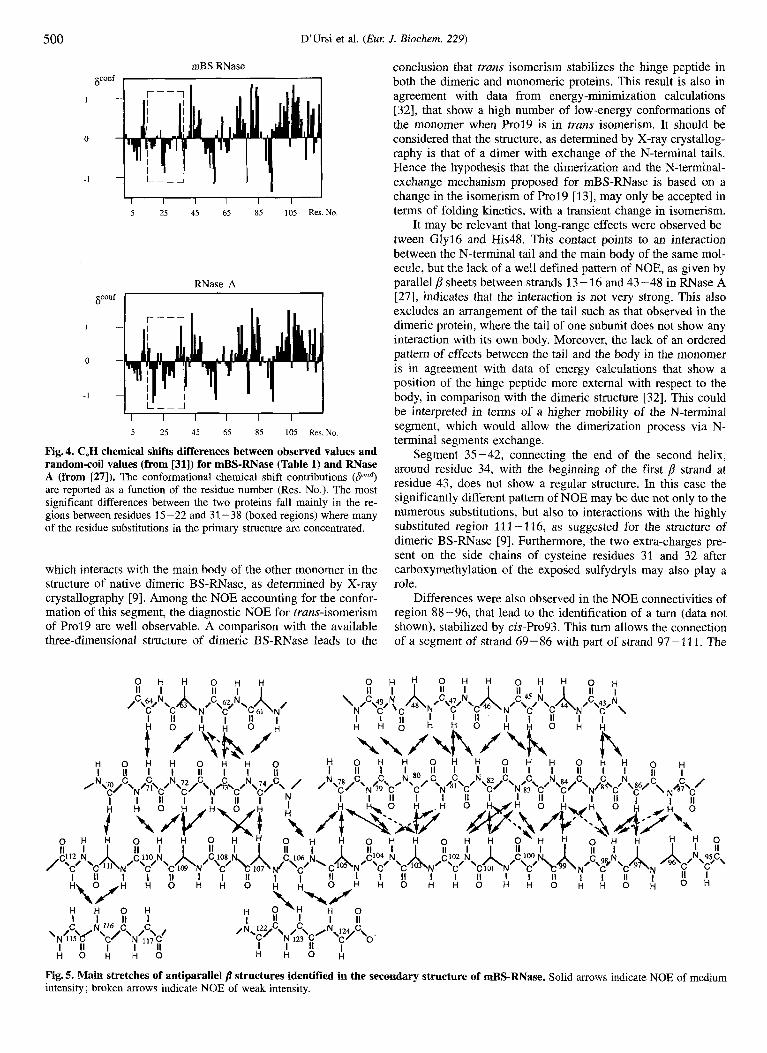
structure of RNase A. Sequential and medium-range connectivi- ties observed in the NOESY spectrum of mBS-RNAse, that can be used to identify helices, /3 sheets and turns [24], are summa- rized in the bar diagram of Fig. 3. Fig. 3 also shows the corre- sponding diagram for RNase A 1271 and the primary structures of both proteins. The general pattern of NOE is the same for both proteins, indicating also for mBS-RNase a secondary struc- ture characterized by long stretches of p strands and by three a- helical segments. The main differences in NH-NH and NH-C,H effects with respect to those observed for RNase A [27-301 are concentrated in regions of primary structure with a larger number of substitutions. As a consequence, these regions corre- spond to segments of different secondary structure, as pointed out by the bar diagram of Fig. 3. The same observation is con- firmed by comparing the C,H chemical shifts of mBS-RNase and RNase A, since it is known that C,H chemical shifts are dependent on the protein secondary structure [31]. Fig. 4 reports the C,H chemical shifts differences between observed values and random coil values [31] for mBS-RNase and RNase A as a function of the sequence position. The most significant differ- ences between the two proteins fall in the regions between resi- dues 15-22 and 31 -38 (Fig. 4) where many of the residue sub- stitutions in the primary structure are concentrated.
The main p-strand regions of mBS-RNase, identified by stretches of daN and lack of sequential dNN connectivities, are shown schematically in Fig. 5 ; they are located in segments 43 - 49, 61-64, 70-74, 78-87 and 94-112. The secondary struc- ture of these regions is very similar to that of dimeric BS-RNase [8, 91 and to that of the corresponding segments of RNase A, as shown by crystallographic [25, 261 and NMR studies [27-301.
A number of long-range NOE confirms the relative arrange- ment of the sheets. In particular, as shown in Fig. 5, numerous NH-NH and NH-C,H inter-strand NOE were identified, whereas

D'Ursi et al. (Eur. J. Biochem. 229)
BSRNase
RNase A
K E S A A A K F E R Q H M D S G NS P S S S S N Y C N L M M C C R K M T Q G K C
K E T A A A K F E R Q H M D S S T S A A S S S N Y C N Q M M K S R N L T K D R C
499
1 10 20 30 40
daN(i,i+l)BS RNase
daN(i,i+l)RNase A
- - - daN(i,i+Z)BS RNase -- - -- - daN(i,i+S)BS RNase - -- -
dNN(i,i+l)BS RNase
dNN(i,i+l)RNate A
dNN(i,i+2)BS RNase - - da&i+3)BS RNase - - - -
BS RNase K P V N T F V H E S L A D V K A V C S Q K K V T C K D G Q T N C Y Q S K S T M R RNase A K P V N T F V H E S L A D V Q A V C S Q K N V A C K N G Q T N C Y Q S Y S T M S
50 60 70 SO
d a N(i, i+l)BS RNase
daN(i,i+l)RNase A 0
d aN(1,1+2)BS RNase - - - - - - - -- -- d aN(i,i+3)BS RNase --
dNN(i, i+l)BS RNase I
dNN(i,i+I)RNase A
- - dNN(i,i+2)BS RNase - - - - - - - dap(i,i+3)BS RNase -
BSRNase RNaseA
I T D C R E T G S S K Y P N C A Y K T T Q V E K H I I V A C G G K P S V P V H F D A S V I T D C R E T G S S K Y P N C A Y K T T Q A N K H I I V A C E G N P Y V P V H F D A S V
90 100 110 120
d aN(i,i+l)BS RNase
daN(i,i+l)RNase A
daN(i,i+2)BS RNase
daN(i.i+3)BS RNase
dNN(i,i+l)BS RNase I - dNN(i.i+l)RNase A c;]
dNN(i,i+2)BS RNase - - da,P(i,i+3)BS RNase
Fig. 3. Sequential and long-range connectivities observed in the NOESY spectrum of mBS-Rh'ase, used for the characterization of the secondary structure (dark grey bars). The corresponding pattern of the connectivities reported for RNase A (from [27]) is shown for comparison (light grey bars).
CaH-C,H inter-strand correlations, that were partially undetect- able in H,O since some C,H were bleached by irradiation of the water resonance, were observed in D,O solution (Fig. 2). NH- C,H signals belonging to VaI63, Asn71, Tyr73, Gln74, Met79, Ile81, Asp83, Cys84, Arg85, Val102, LyslO4 and Val108 resi- dues are still present in a COSY-DQF spectrum recorded two weeks after dissolution in D,O. As can be seen in Fig. 5, the arrangement of the p sheets is consistent with the solvent inac- cessibility of the NH of these residues.
Along segments 4-12, 25-31 and 51-60, three helices were identified by the observation of strong NH-NH and me- dium C,aH-NH (i,i +1) NOESY connectivities, as well as by
C,aH-CFH (i,i + 3) and NWNH(i,i + 3) NOE for most of the residues. The helicoidal conformations were found to be similar to those of RNase A, even if more regular patterns were observ- able in the 51 -60 helix of mBS-RNase.
Segments 13-24 and 32-42, linking regular helices, are characterized by pseudo-helical conformations. They do not show a distinct pattern of NOE as for a regular helix, since the sequential NWNH effects between adjacent residues are not ac- companied by all other diagnostic helicoidal effects.
The first non-regular segment contains the hinge peptide 16- 22 whose conformation is particularly interesting since it connects helix 25-31 with the first a-helical region, i.e. 4-12,
500 D'Ursi et al. (Eur: J. Biochem. 229)
mBS RNase cconf I 0
1
0
- 1
I I I I I I 5 25 45 65 85 105 Res.No.
RNase A gconf fi
5 25 45 65 85 105 Res.No.
Fig. 4. CmH chemical shifts differences between observed values and random-coil values (from [31]) for mBS-RNase (Table 1) and RNase A (from [27]). The conformational chemical shift contributions (P"? are reported as a function of the residue number (Res. No.). The most significant differences between the two proteins fall mainly in the re- gions between residues 15-22 and 31-38 (boxed regions) where many of the residue substitutions in the primary structure are concentrated.
which interacts with the main body of the other monomer in the structure of native dimeric BS-RNase, as determined by X-ray crystallography [9]. Among the NOE accounting for the confor- mation of this segment, the diagnostic NOE for trans-isomerism of Pro19 are well observable. A comparison with the available three-dimensional structure of dimeric BS-RNase leads to the
conclusion that trans isomerism stabilizes the hinge peptide in both the dimeric and monomeric proteins. This result is also in agreement with data from energy-minimization calculations [32], that show a high number of low-energy conformations of the monomer when Pro19 is in trans isomerism. It should be considered that the structure, as determined by X-ray crystallog- raphy is that of a dimer with exchange of the N-terminal tails. Hence the hypothesis that the dimerization and the N-terminal- exchange mechanism proposed for mBS-FWase is based on a change in the isomerism of Pro19 [13], may only be accepted in terms of folding kinetics, with a transient change in isomerism.
It may be relevant that long-range effects were observed be- tween Gly16 and His48. This contact points to an interaction between the N-terminal tail and the main body of the same mol- ecule, but the lack of a well defined pattern of NOE, as given by parallel p sheets between strands 13-16 and 43-48 in RNase A [27], indicates that the interaction is not very strong. This also excludes an arrangement of the tail such as that observed in the dimeric protein, where the tail of one subunit does not show any interaction with its own body. Moreover, the lack of an ordered pattern of effects between the tail and the body in the monomer is in agreement with data of energy calculations that show a position of the hinge peptide more external with respect to the body, in comparison with the dimeric structure [32]. This could be interpreted in terms of a higher mobility of the N-terminal segment, which would allow the dimerization process via N- terminal segments exchange.
Segment 35-42, connecting the end of the second helix, around residue 34, with the beginning of the first /? strand at residue 43, does not show a regular structure. In this case the significantly different pattern of NOE may be due not only to the numerous substitutions, but also to interactions with the highly substituted region 111-116, as suggested for the structure of dimeric BS-RNase [9]. Furthermore, the two extra-charges pre- sent on the side chains of cysteine residues 31 and 32 after carboxymethylation of the exposed sulfydryls may also play a role.
Differences were also observed in the NOE connectivities of region 88-96, that lead to the identification of a turn (data not shown), stabilized by cis-Pro93. This turn allows the connection of a segment of strand 69-86 with part of strand 97 - 11 1. The
Fig. 5. Main stretches of antiparallel fi structures identified in the secondary structure of mBS-RNase. Solid arrows indicate NOE of medium intensity ; broken arrows indicate NOE of weak intensity.

D’Ursi et al. fEux .I. Biochem. 229) 501
Fig. 6. Superposition of the ribbon structures of mBS-RNase (black) and of the A chain of dimeric BS-RNase (light gray). The mBS-RNase model has been obtained, as described in Materials and Methods, with 630 NOE effects involving backbone NH; the model of the A chain of dimeric BS-RNases derived from the X-ray structure [9].
C-terminal p structure, characterized as in RNase A by an irreg- ular connection between segments 115-124 and 106-110, ow- ing to the substitutions Glul11-.Glylll, Asnl13+Lys113 and Serl15+Tyrl15 in the loop 11 1 - 116 that connects the two seg- ments, has a different pattern of NOE. The sequential effect be- tween Tyrl15 NH and Pro114 C,H is absent in RNase A, and can be observed in mBS-RNase; moreover, we observe two NH- NH effects between residues 111-112 and 112-113, whereas no NH-NH effect is present in this region for RNase A.
For the segments 88-96 and 111-116, a different arrange- ment of the loops, compared with those of RNase A, has also been found for dimeric BS-RNase [9]. This shows that the con- formations of these flexible segments, only partially involved in the stability of tertiary structure, are strongly influenced by short-term interactions among the residues.
Loop 63-69, stabilized by the S-S linkage between Cis65 and Cis72 and connecting the two anti-parallel strands 61-64 and 70-74, is characterized by a nearly regular type I p turn. As already pointed out, 2D spectra analysis shows that the NH proton of Lys66, belonging to this segment, is split in two signals of different intensities ; corresponding to two protein isomers, containing a normal or an isopeptide bond in different amounts. This suggests that two different local conformations are gener- ated from the presence of a Asp67 residue in one conformer and p Asp67 in the other conformer. The rigidity of the loop, due to the 65 -72 disulfide bridge, makes the effect even more evident.
A preliminary tertiary-structure model of mBS-RNase was obtained from the X-ray structure of the A chain of BS-RNase [9] by applying only short-range-NOE-derived distance re- straints. The folding of the N-terminal region was obtained by a 120-ps SA simulation 1231 performed by rigidly restraining the position of the residues following Tyr25, and including long- range restraints involving the first 25 residues. A final 100-ps molecular dynamics simulation, with the whole set of NOE-de-
rived restraints, led to the tentative structure shown below. Fig. 6 shows the ribbon representation of the best fit superposition of mBS-RNase and of BS-RNase obtained excluding, in the best fitting calculation, the first 26 residues. It can be seen that both proteins have a similar secondary structure but for the N-termi- nal segment that in the monomeric protein is folded over the main body of the protein, whereas in the dimer it is folded over the main body of the other subunit. It is clear that the folding of mBS-RNase is closer to that of RNase A [30] than to that of (dimeric) BS-RNase.
Conclusions. Proton NMR assignment and secondary-structure determination of monomeric bovine seminal ribonuclease were performed with 3D homonuclear NMR techniques. Computer- assisted analysis of 3D homonuclear spectra was thus tested in this case on a protein of considerable dimensions. The secondary structure of mBS-RNase seems very similar to that of BS-RNase and of RNase A, even though some differences were detected in segments with a high number of substitutions. It may be interest- ing to note that the pattern of NOE observed between residues belonging to the N-terminal tail and the body of the molecule indicates that these two domains of the protein do not interact very strongly, as in the case of RNase A. This could account for a higher mobility of the N-terminal segment, which would allow the dimerization process via N-terminal segments exchange. However, a preliminary 3D model, obtained on the basis of a small number of NOE, indicates that the global 3D structure of mBS-RNase is closer to that of RNase A [30] than to that of (dimeric) BS-RNase.
Characteristic split signals of the residues belonging to the 65-72 loop, confirm the presence of mixture of isomers con- taining a Asp67 or p Asp67, probably in two different conforma- tions.
No differences were observed, arising from the isomerism of Prol9, between mBS-RNase and native dimeric BS-RNase characterized by the exchange of N-terminal segments. This leads to the conclusion that the different conformation of the peptide segment that links the exchangeable N-terminal a helix with the main body of the molecule is not necessarily linked to a different isomerism of Prol9, as previously proposed [13].
A detailed tertiary structure determination, using a complete list of NOE, is presently under way in our laboratory.
We wish to thank Prof. V. Barone for the use of AMBER on his computer.
REFERENCES 1. D’Alessio, G., Di Donato, A,, Parente, A. & Piccoli, R. (1992) Sem-
inal RNase: a unique member of the ribonuclease superfamily, Trends Biochem. Sci. 16, 104-106.
2. Laccetti, P., Spalletti-Cemia, D., Portella, G., De Corato, P., D’Ales- sio, G. & Vecchio, G. (1994) Seminal ribonuclease inhibits tumor growth and reduces the metastatic potential of Lewis Lung carci- noma, Cancer Res. 54, 4253-4256.
3. Di Donato, A. & D’Alessio, G. (1973) Interchain disulfide bridges in ribonuclease BS-1, Biochem. Biophys. Res. Commun. 55,919- 928.
4. D’Alessio, G., Malomi, M. C. & Parente, A. (1975) Dissociation of bovine seminal ribonuclease into catalytically active monomers by selective reduction and alkylation of the intersubunit disulfide bridges, Biochemistry 14, 1116-1122.
5 . Richard, F. M. & Wyckoff, H. W. (1971) Bovine pancreatic ribo- nuclease, in The enzymes, 3rd edn (Boyer, P. D., ed.) vol. 4, pp. 647 - 806, Academic Press, New York.
6. Di Donato, A,, Galletti, P. & D’Alessio, G. (1986) Selective deami- dation and enzymatic methylation of seminal ribonuclease, Bio- chemistry 25, 8361-8368.

502 D’Ursi et al. IEur J. Biochem. 229)
7. Capasso, S., Giordano, F., Mattia, C. A., Mazzarella, L. & Zagari, A. (1983) Refiniment of the structure of bovine seminal ribo- nuclease, Biopolymers 22, 327-332.
8. Mazzarella, L., Mattia, C. A., Capasso, S. & Di Lorenzo, G. (1987) Composite active sites in bovine seminal ribonuclease, Gaz. Chim. Ital. I 1 7, 91 -97.
9. Mazzarella, L., Capasso, S., Demasi, D., Di Lorenzo, G., Mattia, C. A. & Zagari, A. (1993) Bovine seminal ribonuclease: structure at 1.9 A Resolution, Acta Crystallogr. Sect. D 49, 389-402.
10. Crestfield, A. M. (1967) Homologous and hybrid dimers of ribo- nuclease A and its carboxymethylhistidine derivatives, J. Biol. Chem. 242, 3279-3284.
11. Piccoli, R., Tambumni, M., Piccialli, G., Di Donato, A., Parente, A. & D’Alessio, G. (1992) The dual-mode quaternary structure of seminal RNase, Proc. Natl Acad. Sci. USA 89, 1870-1874.
12. Piccoli, R., Di Donato, A. & D’Alessio, G. (1988) Co-operativity in seminal ribonuclease function, Biochem. J. 253, 329 -336.
13. Parente, A. & D’Alessio, G. (1985) Reacquisition of quaternary strucure by fully reduced and denaturated seminal ribonuclease, Eur. J. Biochem. 149, 381 -387.
14. Oschkinat, H., Holak, T. A. & Cieslar, C. (1991) Assignment of protein NMR spectra in the light of homonuclear 3D spectros- copy: an automatable procedure based on 3D TOCSY-TOCSY and 3D TOCSY-NOESY, Biopolymers 31, 699-712.
15. Tamburrini, M., Piccoli, R., De Prisco, R., Di Donato, A. & DAles- sio, G. (1986) Fast and high-yelding procedures for the isolation of bovine seminal RNase, Ital. J. Biochem. (Engl. Ed.) 35, 22- 32.
16. Marion, D. & Wuthrich, K. (1983) Application of phase sensitive two-dimensional correlated spectroscopy (COSY) for measure- ments of proton-proton spin-spin coupling constants, Biochem. Biophys. Res. Commun. 113, 967-974.
17. Piantini, U., Soerensen, 0. W. & Ernst, R. R. (1982) Multiple quantum filters for elucidating NMR coupling networks, J. Am. Chem. SOC. 104, 6800-6801.
18. Jeener, J., Meyer, B. H., Bachman, P. & Ernst, R. R. (1979) Investi- gation of exchange processes by two-dimensional NMR spectros- copy, J. Chem. Phys. 71, 4546-4553
19. Bax, A. & Davis, D. G. (1985) MLEV-17- based two dimensional homonuclear magnetization transfer spectroscopy, J. Magn. Re- son. 65, 355-360.
20. Plateau, P. & Gueron, M. (1982) Exchangeable proton NMR without base-line distortion, using new strong pulse sequences, J. Am. Chem. SOC. 104,7310-7311.
21. Cieslar, C., Holak, T. A. & Oschkinat, H. (1990) 3D TOCSY- TOCSY processing using Linear prediction, as a potential tech- nique for automated assignment, J. Magn. Reson. 89, 84-90.
22. Oschkinat, H., Clore, G. M. & Gronenbom, A. M. (1988) Three- dimensional homonuclear Hartmann-Hahn-nuclear Overhauser enhancement spectroscopy in H,O and its applications to proteins, J. Magn. Reson. 81, 212-219.
23. Pearlman, D. A., Case, D. A., Caldwell, J. C., Seibel, G. L., Singh, U. C., Weiner, P. & Kollman, A. (1991) AMBER 4.0, University of California, San Francisco.
24. Wuthrich, K. (1986) in NMR of proteins and nucleic acids (John Wiley & Sons, Inc., eds) pp. 162-175, New York.
25. Kartha, G., Bello, J. & Harker, D. (1967) Tertiary structure of Ribo- nuclease, Nature 213, 862-865.
26. Avey, H. P., Boles, M. O., Carlisle, C. H., Evans, S. A., Moms, S. J., Palmer, R. A., Woolhouse, B. A. & Shall, S . (1967) Structure of ribonuclease, Nature 213, 557-562.
27. Rico, M., Bruix, M., Santoro, J., Gonzalez, C., Neira, J. L., Nieto, J. L. & Herranz, J. (1989) Sequential H-NMR assignment and solution structure of bovine pancreatic ribonuclease A, Eur. J. Biochem. 183, 623-638.
28. Robertson, A. D., Purisima, E. O., Eastman, M. A. & Scheraga, H. A. (1989) Proton NMR assignment and regular backbone struc- ture of bovine pancreatic ribonuclease A in aqueous solution, Bio- chemistry 28, 5930-5938.
29. Rico, M., Santoro, J., Gonzales, C., Bruix, M., Neira, J. L., Nieto, J. L. & Herranz, J. (1991) 3D Structure of bovine pancreatic ribo- nuclease A in aqueous solution: An approach to tertiary structure determination from a small basis of H-NMR NOE correlations, J. Biomol. NMR 1, 283-298.
30. Santoro, J., Gonzales, C., Bruix, M., Neira, J. L., Nieto, J. L., Her- ranz, J. & Rico, M. (1993) High resolution three-dimentional structure of ribonuclease A in solution by Nuclear Magnetic Res- onance Spectroscopy, J. Mol. Biol. 229, 722-734.
31. Wishart, D. S., Sykes, B. D. & Richards, F. M. (1992) The chemical shift index: a fast simple method for the assignment of protein secondary structure through NMR spectroscopy, Biochemistry 31,
32. Mazzarella, L., Vitagliano, L., Zagari, A. & Capasso S. (1993) Flexi- bility of the peptide 16-22 in Ribonucleases, Proc. 3rd Znt. Meet. Ribonucleases: Chemistry, Biology and Biotechnology, Capri 1993, L7.
1647 - 1651.
Related Documents











