www.rsc.org/crystengcomm Volume 9 | Number 12 | December 2007 | Pages 1131–1270 CrystEngComm COVER ARTICLE Addadi et al. Asprich mollusk shell protein: in vitro experiments aimed at elucidating function in CaCO 3 crystallization HOT ARTICLE de Leeuw and Rabone Molecular dynamics simulations of the interaction of citric acid with the hydroxyapatite (0001) and (010) surfaces in an aqueous environment HOT ARTICLE Pokroy and Aizenberg Calcite shape modulation through the lattice mismatch between the self-assembled monolayer template and the nucleated crystal face

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.rsc.org/crystengcomm Volume 9 | Number 12 | December 2007 | Pages 1131–1270
CrystEngComm
COVER ARTICLEAddadi et al.Asprich mollusk shell protein: in vitro experiments aimed at elucidating function in CaCO3 crystallization
HOT ARTICLEde Leeuw and RaboneMolecular dynamics simulations of the interaction of citric acid with the hydroxyapatite (0001) and (010) surfaces in an aqueous environment
HOT ARTICLEPokroy and AizenbergCalcite shape modulation through the lattice mismatch between the self-assembled monolayer template and the nucleated crystal face

This paper is published as part of a CrystEngComm themed issue on:
Biomineralisation
Guest edited by Lia Addadi The Weizmann Institute of Science, Israel
Published in issue 12, 2007 of CrystEngComm
Images reproduced by permission of Lia Addadi (outside) and James De Yoreo (inside) Other papers published in this issue include: Molecular dynamics simulations of the interaction of citric acid with the hydroxyapatite (0001) and (01-10) surfaces in an aqueous environment N. H. de Leeuw and J. A. L. Rabone, CrystEngComm, 2007, DOI: 10.1039/b710974a
Synthesis-dependant structural variations in amorphous calcium carbonate Raymond S. K. Lam, John M. Charnock, Alistair Lennie and Fiona C. Meldrum, CrystEngComm, 2007, DOI: 10.1039/b710895h
Calcite shape modulation through the lattice mismatch between the self-assembled monolayer template and the nucleated crystal face Boaz Pokroy and Joanna Aizenberg, CrystEngComm, 2007, DOI: 10.1039/b710294a
Fine structure of nacre revealed by solid state 13C and 1H NMR Christian Jäger and Helmut Cölfen, CrystEngComm, 2007, DOI: 10.1039/b708600h
Visit the CrystEngComm website for cutting-edge crystal engineering research
www.rsc.org/crystengcomm

Asprich mollusk shell protein: in vitro experiments aimed at elucidatingfunction in CaCO3 crystallization
Yael Politi,a Julia Mahamid,a Harvey Goldberg,b Steve Weinera and Lia Addadi*a
Received 27th June 2007, Accepted 11th September 2007
First published as an Advance Article on the web 17th September 2007
DOI: 10.1039/b709749b
Acidic proteins are key components of the organic matrix of many biologically formed minerals
and are therefore thought to play an important role in their formation. Here we study the effect of
one unusually acidic protein of the Asprich family, associated with mollusk shell prismatic layer,
on the precipitation of CaCO3 in vitro. We show that Asprich induces and transiently stabilizes the
deposition of amorphous calcium carbonate (ACC). Asprich also induces the formation of ACC
when adsorbed onto chitin, a major component of the intracrystalline organic matrix of the
prismatic layer. Based on this evidence, combined with previous studies on the forming prisms in
the shell layer, we suggest that the in vivo function of Asprich is inducing and stabilizing ACC
particles and inhibiting their uncontrolled crystallization until they undergo secondary nucleation
on the growing prisms.
Introduction
The formation of crystals from an amorphous precursor phase
is a common phenomenon in biomineralization.1,2 Organisms
from various phyla use this strategy to form different
crystalline materials, including the transformation of a
disordered ferrihydrite phase into magnetite,3 amorphous
calcium phosphate into carbonated apatite,4 and amorphous
calcium carbonate (ACC) into either calcite5–7 or aragonite.8,9
Organisms must invest energy to reversibly stabilize these
intrinsically unstable transient amorphous phases. Possible
advantages of this strategy are the efficient transport of ions to
the mineralization site, providing alternative pathways for the
synthesis of otherwise difficult-to-form phases and for the
production of skeletal materials in complex shapes.2,10
The mechanisms of formation of the mature crystalline
phase involve several distinct processes: transient stabilization
of the amorphous phase, subsequent transformation into a
stable crystalline phase, polymorph selection and crystal
orientation. In the case of ACC, polymorph selection may
occur at the initial stage based on the observation that the
transient amorphous phase already has the nascent order of
the polymorph into which it will transform.11,12 The nucleation
stage may occur by epitaxial nucleation on a preexisting
crystalline seed5,13,14 or be induced by specific nucleating
proteins in the organic matrix.15 Control over these processes
needs to be exerted at every stage. Much of this control is
assumed to be exercised by highly charged glycoproteins, many
of which are rich in aspartic acid.16–20 Here we investigate the
role of a family of such proteins, called Asprich,21 present in
the calcitic prisms of the shells from the mollusk Atrina rigida.
The Asprich proteins are abundant in the calcite prisms, and
are therefore thought to play a significant role in the formation
of the mineral. They are composed of several domains,
including a calcium binding domain, a domain that possibly
binds Mg ions, and long stretches that are enriched in aspartic
acid, including sequences of poly-Asp. The acidic amino acids
constitute more than 50% of the sequence and around 30% are
hydrophobic amino acids.21 Nudelman et al.14 characterized
the structure and organization of the organic matrix in the
mature and forming prismatic shell layer of the mollusk Atrina
rigida. They show that the intracrystalline matrix is composed
of a network of chitin fibers, and that the prisms grow by
depositing spherical mineral particles on the fibrous scaffold.
Using immuno-labeling, they show that Asprich proteins
are associated with the initially deposited mineral particles.
Interestingly, each prism in the A. rigida prismatic layer
diffracts X-rays as a very well ordered single calcite crystal.22
Here we show that one protein from the Asprich family
(Asprich-c) transiently stabilizes ACC during the precipitation
of CaCO3 from solution. The influence of the protein on the
different stages of crystal growth is characterized, as well as its
effect on crystal nucleation when adsorbed on a glass substrate
and chitin fibers. We suggest that these observations are
relevant to the function of the protein in vivo.
Results
Asprich (His-22) cDNA clone c was expressed in competent
E. coli XL-10 cell systems, and a series of in vitro assays were
performed in which the effect of the expressed protein on the
precipitation of CaCO3 was studied. Note that it is not known
whether native Asprich extracted from the shell is post-
translationally modified. The Asprich used in this study was
expressed in prokaryotic cells, and was thus devoid of post-
translational modifications. The effect of a synthetic poly-
aspartic acid peptide (poly-Asp) was analysed for comparison.
The length of the peptide (8.4 KDa) is of the same order of
aDepartment of Structural Biology, Weizmann Institute of Science,Rehovot 76100, IsraelbCIHR Group in Skeletal Development and Remodeling, Schulich Schoolof Medicine & Dentistry, University of Western Ontario, London,Ontario, Canada N6A 5C1
PAPER www.rsc.org/crystengcomm | CrystEngComm
This journal is � The Royal Society of Chemistry 2007 CrystEngComm, 2007, 9, 1171–1177 | 1171
magnitude as the length of the protein (21.8 KDa). All
experiments were performed using equal molar concentrations
of total amino acids for Asprich and poly-Asp, rather than
using protein/peptide molar concentrations.
Slow precipitation experiments with increasing
concentrations of Asprich
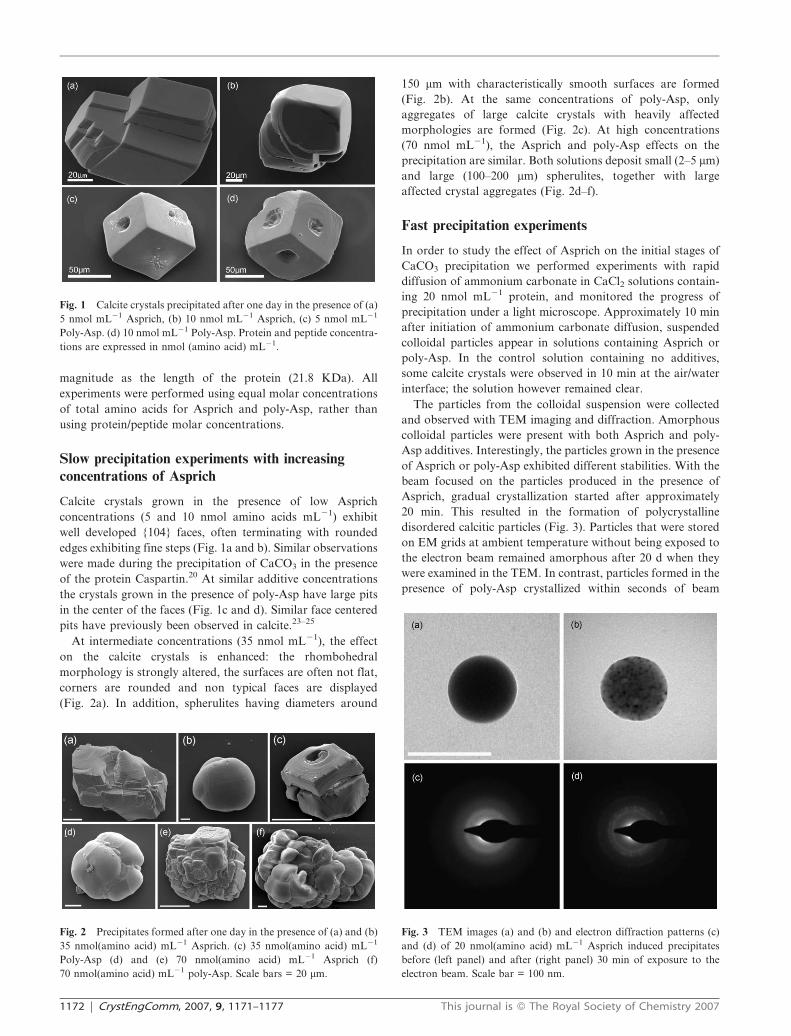
Calcite crystals grown in the presence of low Asprich
concentrations (5 and 10 nmol amino acids mL21) exhibit
well developed {104} faces, often terminating with rounded
edges exhibiting fine steps (Fig. 1a and b). Similar observations
were made during the precipitation of CaCO3 in the presence
of the protein Caspartin.20 At similar additive concentrations
the crystals grown in the presence of poly-Asp have large pits
in the center of the faces (Fig. 1c and d). Similar face centered
pits have previously been observed in calcite.23–25
At intermediate concentrations (35 nmol mL21), the effect
on the calcite crystals is enhanced: the rhombohedral
morphology is strongly altered, the surfaces are often not flat,
corners are rounded and non typical faces are displayed
(Fig. 2a). In addition, spherulites having diameters around
150 mm with characteristically smooth surfaces are formed
(Fig. 2b). At the same concentrations of poly-Asp, only
aggregates of large calcite crystals with heavily affected
morphologies are formed (Fig. 2c). At high concentrations
(70 nmol mL21), the Asprich and poly-Asp effects on the
precipitation are similar. Both solutions deposit small (2–5 mm)
and large (100–200 mm) spherulites, together with large
affected crystal aggregates (Fig. 2d–f).
Fast precipitation experiments
In order to study the effect of Asprich on the initial stages of
CaCO3 precipitation we performed experiments with rapid
diffusion of ammonium carbonate in CaCl2 solutions contain-
ing 20 nmol mL21 protein, and monitored the progress of
precipitation under a light microscope. Approximately 10 min
after initiation of ammonium carbonate diffusion, suspended
colloidal particles appear in solutions containing Asprich or
poly-Asp. In the control solution containing no additives,
some calcite crystals were observed in 10 min at the air/water
interface; the solution however remained clear.
The particles from the colloidal suspension were collected
and observed with TEM imaging and diffraction. Amorphous
colloidal particles were present with both Asprich and poly-
Asp additives. Interestingly, the particles grown in the presence
of Asprich or poly-Asp exhibited different stabilities. With the
beam focused on the particles produced in the presence of
Asprich, gradual crystallization started after approximately
20 min. This resulted in the formation of polycrystalline
disordered calcitic particles (Fig. 3). Particles that were stored
on EM grids at ambient temperature without being exposed to
the electron beam remained amorphous after 20 d when they
were examined in the TEM. In contrast, particles formed in the
presence of poly-Asp crystallized within seconds of beam
Fig. 1 Calcite crystals precipitated after one day in the presence of (a)
5 nmol mL21 Asprich, (b) 10 nmol mL21 Asprich, (c) 5 nmol mL21
Poly-Asp. (d) 10 nmol mL21 Poly-Asp. Protein and peptide concentra-
tions are expressed in nmol (amino acid) mL21.
Fig. 2 Precipitates formed after one day in the presence of (a) and (b)
35 nmol(amino acid) mL21 Asprich. (c) 35 nmol(amino acid) mL21
Poly-Asp (d) and (e) 70 nmol(amino acid) mL21 Asprich (f)
70 nmol(amino acid) mL21 poly-Asp. Scale bars = 20 mm.
Fig. 3 TEM images (a) and (b) and electron diffraction patterns (c)
and (d) of 20 nmol(amino acid) mL21 Asprich induced precipitates
before (left panel) and after (right panel) 30 min of exposure to the
electron beam. Scale bar = 100 nm.
1172 | CrystEngComm, 2007, 9, 1171–1177 This journal is � The Royal Society of Chemistry 2007

exposure, yielding crystals with individually identifiable single
crystal electron diffraction patterns (Fig. 4). When unexposed
samples were observed after 10 d, they were all crystalline.
Similar colloidal particle suspensions were collected 30 min
after the initiation of the experiment and examined by FTIR.
The crystallinity of the samples can be established by following
the n4 in-plane bending vibration at 713 cm21, which is
extremely broad in samples of stable ACC. The ratio of the
intensities of the out-of-plane bending and in-plane bending
vibrations In2/In4 is around 3 in fully crystallized geological or
synthetic calcite, and increases with increasing disorder in the
structure.5,26 The spectra of the particles precipitated in the
presence of Asprich indicated a mixture of ACC and calcite
and in one case only ACC was detected (Fig. 5b). The particles
formed in the presence of poly-Asp under the same conditions
were all calcitic and had In2/In4 ratios around 2.6, implying that
they were fully crystalline (Fig. 5a).
The precipitates that accumulated at the bottom of the wells
containing suspensions formed in the presence of Asprich,
consist of 0.05–5 mm spherulites and calcite rhombohedra. The
smallest particles (Fig. 6a, inset) are presumably colloidal
particles that adhered to the glass, while the larger particles
result from fusion of the former. Interestingly, the {104} faces
of the calcite crystals show relatively high steps covering part
of the face. The steps have very flat surfaces and rough edges.
These steps seem to advance by addition and fusion of
spherulites (colloidal particles) to their edges (Fig. 6b, c).
Spherulites and crystal aggregates of different sizes also preci-
pitate in the presence of poly-Asp (Fig. 7), however, the above
mentioned unique form of crystallization was not observed.
Nucleation assays
Asprich protein and poly-Asp peptide were adsorbed on
silicon coated glass slides by incubation of the slides with a
drop of 50 nmol mL21 protein/peptide solution. The solution
was removed, the slides were floated on top of a CaCl2solution and precipitation was induced. After approximately
1 h the slides were collected and the precipitates were observed
Fig. 4 TEM image (a) and electron diffraction pattern of
20 nmol(amino acid) mL21 poly-Asp induced precipitates before (b)
and after (c) exposure of a few seconds to the electron beam.
Fig. 5 Normalized FTIR spectra of suspended particles grown in the
presence of 20 nmol(amino acid) mL21 poly-Asp (a) and Asprich (b).
Fig. 6 Precipitates accumulated on the glass at the bottom of the well after 30 min of fast precipitation, in the presence of 20 nmol(amino acid)
mL21 Asprich. (a) Large spherulites. Inset: small spherulite similar to spherulites in Fig. 3. scale bar = 50 nm; (b) a crystal displaying the typical
rhombohedral morphology, with colloidal particles accumulating on the steps. (c) Higher magnification of (b), arrows point to colloidal particles.
This journal is � The Royal Society of Chemistry 2007 CrystEngComm, 2007, 9, 1171–1177 | 1173

using a light microscope. The influence of the protein or
peptide on crystal nucleation was evaluated by examining
only the crystals nucleated on the slides, and avoiding crystals
that were homogeneously nucleated in the bulk solution.
Moreover, because the only protein present is adsorbed on the
glass substrate, there should be no effect of the protein on
crystal growth.
In the control containing no additive, calcite crystals
nucleated over the whole area of the glass (Fig. 8c). In
contrast, nucleation of calcite crystals was almost totally
inhibited by the presence of Asprich adsorbed on the glass
slide. This was inferred from the fact that the area where the
protein-containing drop was adsorbed contained relatively
few crystals (Fig. 8a). Nucleation was also inhibited by the
presence of poly-Asp adsorbed on the glass, but to a much
lesser extent (Fig. 8b). However without knowing the amount
of the respective protein adsorption to the surface, we cannot
determine inhibitory potencies. It is possible that the difference
in the degree of inhibition between poly-Asp and Asprich is
related to the difference in the extent to which the poly-Asp
adsorbs compared to the Asprich.
Protein adsorbed on chitin
Chitin and acidic macromolecules are the two major compo-
nents of the intracrystalline organic matrix assembly in the
prismatic layer.14 There are also indications that Asprich is
associated with chitin at the early stage of mineralization.14
In order to test for the combined effect of the two components
on calcium carbonate deposition, we performed precipitation
experiments on a chitin substrate to which Asprich was
adsorbed, in a manner similar to the method described by
Falini et al.27 The results are compared with poly-Asp
adsorbed on chitin, and a chitin only substrate.
In the absence of adsorbed macromolecules calcite crystals
were only present on the chitin surface and no precipitation
occurred within the chitin sheaths. The presence of either
poly-Asp or Asprich adsorbed on the chitin fibers resulted in
precipitation occurring inside the chitin, but the resultant
crystals were significantly different. In the presence of poly-
Asp most of the crystals were affected rhombohedra (Fig. 9c)
Fig. 7 Precipitates accumulated on the glass at the bottom of the well
after 30 min of fast precipitation, in the presence of 20 nmol(amino
acid) mL21 poly-Asp. (a) Large spherulites; (b) stepped calcite crystals
delimited by regular {104} faces. Scale bar = 10 mm.
Fig. 8 Calcite crystals nucleated on siliconized glass slides. The slides were pre-incubated with a drop of (a) Asprich protein, (b) poly-Asp peptide
and (c) no additive, and placed on top of a CaCl2 solution. The dashed line marks the borders where the drop was incubated.
Fig. 9 CaCO3 Precipitation on chitin substrate with Asprich and poly-Asp adsorbed. (a) Spherulite precipitated on chitin substrate incubated with
Asprich. (b) A magnification of the interior of the spherulite showing the mineral spheres and an area with extensive fiber mineralization. (c) A
calcite crystal precipitated on chitin substrate incubated with poly-Asp.
1174 | CrystEngComm, 2007, 9, 1171–1177 This journal is � The Royal Society of Chemistry 2007

and some vaterite spherulites. In contrast, a large number of
spherulites formed in the presence of Asprich protein. A close
examination of the interior of the spherulites revealed that the
crystals grew around the chitin fibers and that the chitin is
decorated with small spherical mineral particles (Fig. 9b). In
the presence of both Asprich and poly-Asp we found many
places in which the chitin fibers are mineralized. Similar
observations were made by Kato and Amamiya28 using chitin
and polyanions, e.g., polyAsp, polyGlu and poly(acrylic acid).
Experimental
Cloning and expression of Asprich
Asprich (His-22) cDNA obtained from the Atrina cDNA
library was cloned into the pET28 expression vector
(Novagen). The vector added an N-terminal 6xHis tag and a
TEV cleavage site to the Asprich cDNA sequence. The amino
acid sequence of the expressed protein is as described (protein
sequence ‘‘c’’). The recombinant DNA procedures were carried
out using methods described by Sambrok 1989.29 The coding
sequence was confirmed by DNA sequencing.
Plasmids were first transfected into competent E. coli XL-10
cells. Colonies were picked and the sequence confirmed. E. coli
BL21 (DE3) were transformed with the expression plasmids,
and grown in phosphate-buffered Super Broth (SB) supple-
mented with 15 mg mL21 kanamycin and 0.4% glucose to an
absorbance of 0.6–0.9. After induction with 2 mM isopropyl-
b-D-thiogalactopyranoside, cultures were grown for a further
4 h, cells recovered by centrifugation, resuspended and then
sonicated in denaturing binding buffer (5 mM imidazole, 0.5 M
NaCl, 0.02 M Tris/HCl, 6 M urea, pH 7.9) as described.14 The
extract was applied to a His-bind column (Novagen), and
Asprich eluted with 0.5 M imidazole-containing elution buffer.
Fractions containing Asprich were pooled and subjected to
purification by fast protein liquid chromatography (FPLC)
using established protocols. Proteins were purified by chro-
matography on a Q-Sepharose Fast Flow column followed by
gel filtration on a Superdex 200PG column (GE Biosciences).
Purification of Asprich was monitored by electrophoresis on
12.5% SDS polyacrylamide gels, stained with Stains-all and
silver nitrate as described. Fractions containing Asprich were
dialyzed against 0.01 M ammonium bicarbonate buffer,
lyophilized in aliquots, and protein content determined by
amino acid analysis. Yield of intact Asprich (y95% purity)
was approximately 400 mg L21 of initial culture media.
Poly-aspartic-acid peptide, mean MW(vis) = 8400 Da, was
purchased from Sigma.
Precipitation experiments
Slow precipitation. Synthetic crystals were grown in Nunc
multi-well dishes (24 wells; 1.5 cm diameter) by diffusion of
ammonium carbonate vapor into calcium chloride solutions.
Glass cover slips were placed on the bottom of each well. A
total volume of 0.5 mL of 10 mM CaCl2 (Merck, A grade)
solution, was introduced into each well. Poly-Asp or Asprich
were added to reach different final concentrations: 5, 10, 35
and 70 nmol (amino acids) mL21, corresponding to 0.55, 1.1,
3.86 and 7.7 mg mL21. Each well was sealed separately with
aluminium foil, pierced with a 25 G needle and sealed with
Parafilm. Control experiments were performed in parallel with
10 mM CaCl2 and no additives. The multi-well dish was placed
in a closed desiccator containing ammonium carbonate vapor
for one day at room temperature.
Fast precipitation. These experiments were performed in
a Nunc multi-well dish, where the central wells contained
the crystallization solutions with 10 mM CaCl2 and
20 nmol (amino acids) mL21 poly-Asp or Asprich additives,
and a CaCl2 control. Four vials containing ammonium
carbonate powder were placed at the corners of the plate.
The vials were covered with aluminium foil and sealed with
Parafilm. One hole was punctured in each of the vials with a
25 G needle. The plates were covered, sealed with Parafilm and
placed under a light microscope to follow the precipitation.
FTIR spectrometry. For the fast precipitation procedure, the
reaction was stopped when the solution became cloudy,
normally after approximately 30 min. The suspension was
transferred with a glass pipette into an Eppendorf tube, and
centrifuged for 3 min at 14000 rpm. The pellet was re-
suspended in ethanol and placed in an agate mortar. After the
ethanol evaporated, the precipitate was lightly ground and
KBr pellets were prepared and analysed by FTIR spectrometry
(Nicolet 380).
Transmission electron microscopy (TEM). (FEI, Philips,
T12). The suspension was collected 10 min after diffusion
was initiated, centrifuged as above and the pellet was re-
suspended in ethanol. A drop of the ethanol suspension was
evaporated on a TEM grid.
Precipitations on a chitin substrate. b-Chitin was obtained
from the pen of the squid Loligo sp. (Mediterranean Sea) and
purified by reflux in 1 M NaOH solution, following Darmon
et al.30 It was then washed extensively with water and stored
dry. Pieces of chitin (0.5 x 0.5 cm) were incubated overnight
with 50 nmol (amino acids) mL21 Asprich or poly-Asp in
10 mM CaCl2 solution. The control chitin was incubated
with CaCl2 solution only. The chitin was then washed with
deionized water and placed in wells containing 0.5 mL 10 mM
CaCl2 solution. The crystallization was performed in the same
way as in the fast crystallization described above, but with no
additives in the solution.
Scanning electron microscopy (SEM). (Philips, XL30
FESEM FEG). The glass slides from each well were rinsed
in purified water and mounted on an aluminum stub with
double-sided carbon tape and Au/Pd sputtered.
Nucleation assay. A drop of 10 mM CaCl2 solution with or
without 50 nmol (amino acids) mL21 Asprich or poly-Asp
was placed in the center of a 12 mm siliconized glass slide
(Hampton Research). The incubation was performed in a
closed box containing open water vials, for 4–6 h at room
temperature. The slide was then rinsed with water, placed
face down on a 10 mM CaCl2 solution in a multi-well dish
and crystallization was initiated as described above for fast
This journal is � The Royal Society of Chemistry 2007 CrystEngComm, 2007, 9, 1171–1177 | 1175

precipitation. After 1 h the slides were collected, rinsed with
water and viewed with a light microscope (Nikon, Eclipse
E600 Pol).
Discussion
We show that Asprich protein inhibits calcite nucleation and
growth. It also induces and stabilizes an otherwise unstable
amorphous calcium carbonate phase. The stabilization is
transient and the mineral eventually transforms into calcite.
The effects of both Asprich and poly-Asp on calcite crystal
growth are not specific for certain crystallographic planes.
Nevertheless, there are significant differences in the manner
Asprich and poly-Asp interact with the growing crystals. The
rounded crystal edges observed during slow growth of calcite
in the presence of low concentrations of Asprich in solution
may result from reversible adsorption of Asprich on high-
energy steps and kink sites on the growing crystal and their
subsequent accumulation at the crystal edges. Alternatively,
they may also result from the addition of colloidal ACC
particles to the growing steps, as observed during the fast
growth experiments. In contrast, poly-Asp induces the forma-
tion of deep pits in the center of the {104} crystal faces, the
so-called ‘hopper crystals’. This morphology may arise from
adsorption of inhibitors on sites of emerging screw dislocations
or from Mullins–Sekerka instabilities, as observed for calcite
grown in the presence of simple alcohols.25 At higher additive
concentrations, the respective effects of poly-Asp and Asprich
become more pronounced, until deposition of spherulites,
characteristic of crystal formation through transient ACC
precursors, is observed for both additives. This occurs however
with Asprich at concentrations lower than with poly-Asp.
During fast precipitation experiments, both Asprich and
poly-Asp promote the formation of a colloidal suspension in
solution before the onset of crystallization in the control
experiment. The ACC precipitates formed in the presence of
Asprich, when stored in air, were stable for at least 20 d,
whereas ACC formed in the presence of poly-Asp crystallized
after 4 to 10 d. After 30 min in water, the former particles
were amorphous, while the latter were crystalline, based on
infrared analysis. Interestingly, colloidal particles formed in
the presence of Asprich, were observed to incorporate onto
layers of growing calcite single crystals. Although we do not
have direct in situ evidence that the particles adding to the
crystal are still amorphous, it is reasonable to assume that once
the colloids come in contact with the crystal they diffuse along
the surface to steps, where they undergo secondary nucleation
to become part of the growing single crystal.
Polyanions, such as poly-aspartate, poly-acrylate and poly-
carboxylate in general are well known for their ability to inhibit
calcite nucleation and induce the formation of amorphous
phases.31–33 Nevertheless, the potency of the Asprich protein is
found to be much higher at lower concentrations compared to
poly-Asp.
ACC may form in the presence of Asprich by inhibition of
nucleation and subsequent increase of the supersaturation
levels in the solution. This has been proposed for other
poly-anions,34 acidic macromolecules extracted from other
biominerals,35–37 and occurs during fast precipitation even
without additives.33 Moreover, Asprich, being negatively
charged, recruits calcium ions and thus locally increases the
supersaturation level such that ACC may precipitate. Asprich
is composed of y60% acidic residues, and y30% hydrophobic
residues. The poly-Asp peptide used here thus has approxi-
mately twice the negative charge/amino-acid concentration
compared to Asprich. When used at the same concentration
(in moles amino acids) mL21 as Asprich, poly-Asp’s ability
to inhibit calcite nucleation is lower. We deduce from this
that the potency of Asprich protein is not solely related to its
polyanionic nature, and that its composition and/or sequence
and structure play a significant role in ACC induction and
stabilization.
A mechanistic interpretation of the results presented must
provide an explanation for four independent effects observed
here in the presence of Asprich: inhibition of crystal growth,
inhibition of de novo crystal nucleation, induction of ACC and
stabilization of ACC particles. Inhibition of crystal growth as
well as concentration of calcium carbonate leading to its pre-
cipitation can both be explained by the poly-anionic character
of the protein that concentrates calcium (and subsequently
carbonate) ions and is adsorbed on crystal steps because of
its multiple charge.38 Inhibition of de novo nucleation by
mixed hydrophilic–hydrophobic peptides and proteins has
been suggested to arise from the hydrophobic domain acting as
a diffusion barrier for lattice ion adsorption to the incipient
nuclei.39,40 This would, however, require sufficient protein
coverage of the crystallization nuclei that could not be easily
achieved at the protein concentration used, unless there is
protein aggregation. Besides, Asprich adsorbed on hydro-
phobic glass also inhibits calcite nucleation much more than
the hydrophobic glass itself. This excludes this explanation in
the present case.
Some interesting information also related to the nucleation-
inhibiting mechanism can be deduced from the Asprich effect
on ACC stabilization. Interestingly, the IR spectra of the
colloidal particles show a broad absorption around the n4
absorption frequency. This is characteristic of both synthetic
and biogenic stable ACC minerals, whereas in transient ACC
this peak has reduced intensity, but remains relatively sharp.
Stable ACC contains structural water in proportions of
one molecule of water per CaCO3, while transient ACC is
practically anhydrous.10 We suggest that, in vitro, the Asprich-
induced ACC phase contains bound water, and that this
structural water plays a role in the stabilization of the phase.
Practically, the particles cannot crystallize unless water is
expelled. The hydrophobic part of the protein may then
conceivably contribute to stabilization of ACC by causing
water ordering. This would be similar to the role suggested in
the inhibition of ice formation by antifreeze proteins.41
Considering the above information on the effect of Asprich
on calcium carbonate deposition in vitro, what can be
concluded concerning the function(s) of Asprich in mollusk
shell prismatic layer formation? Nudelman et al.14 showed that
the deposition cycle leading to formation of the large calcite
crystals in growing A. rigida prismatic layer starts with
secretion of a chitin meshwork, which is then decorated with
20–30 nm particles of calcium carbonate containing Asprich.
While the particles become denser, they fuse with the existing
1176 | CrystEngComm, 2007, 9, 1171–1177 This journal is � The Royal Society of Chemistry 2007

calcite crystal. The final product is a single crystal of calcite.
We suggest that the particles are Asprich-stabilized ACC
particles introduced by the mantle cells into the mineralization
compartment, where they undergo epitaxial nucleation on
the preexisting calcite prism. The in vivo function of Asprich
would be to induce and stabilize ACC particles and to inhibit
uncontrolled calcite nucleation, until the mineral particles
come into contact with the preexisting calcite prism. The
crystallization process would then be similar to the sequence
observed during in vitro crystallization (Fig. 6), where initial
formation of transient ACC particles, followed by their fusion
to growing calcite crystals, occurs. In addition, the in vitro
experiments performed in the presence of chitin support a
possible function of the interaction between Asprich and chitin
in the formation of the biogenic mineral.
As yet there is no direct evidence for the use of a transient
ACC precursor phase in adult bivalve shell biomineralization,
although this has been suggested.14,15,42 It has been shown to
occur in the formation of the larval mollusk shell.8,9 In vitro
studies such as the one described here and by Gotliv et al.43
show that the proteins involved in the mineralization process
of the prismatic and nacreous layers induce the formation of
ACC phase that subsequently crystallizes into either calcite or
aragonite. It is therefore likely that adult bivalves such as
Atrina rigida, also use this apparently widespread strategy of
biomineralization. We note that this strategy can also be
exploited for the design of new synthetic crystalline materials
with complex shapes and superior materials properties.
Acknowledgements
We are especially grateful to Hong H. Chen for preparing the
recombinant Asprich protein. We thank Irit Nudelman for
help in experiments, Dr Talmon Arad for his help in the
electron microscopy work and Fabio Nudelman for useful
discussions. We thank the Israeli Ministry of Science and
the Minerva Foundation for financial support. L.A. is the
incumbent of the Dorothy and Patrick Gorman Professorial
Chair of Biological Ultrastructure, and S.W. is the incumbent
of the Dr Trude Burchardt Professorial Chair of Structural
Biology.
References
1 H. A. Lowenstam and S. Weiner, On Biomineralization, OxfordUniversity Press, New York, 1989.
2 S. Weiner, I. Sagi and L. Addadi, Science, 2005, 309, 1027.3 K. M. Towe and H. A. Lowenstam, J. Ultrastruct. Res., 1967, 17, 1.4 H. A. Lowenstam and S. Weiner, Science, 1985, 227, 51.5 E. Beniash, J. Aizenberg, L. Addadi and S. Weiner, Proc. R. Soc.
London, Ser. B, 1997, 264, 461.6 Y. Politi, T. Arad, E. Klein, S. Weiner and L. Addadi, Science,
2004, 306, 1161.7 R. Dillaman, S. Hequembourg and M. Gay, J. Morph., 2005, 263,
356.
8 J. C. Marxen, W. Becker, D. Finke, B. Hasse and M. Epple,J. Molluscan Studies, 2003, 69, 113.
9 I. M. Weiss, N. Tuross, L. Addadi and S. Weiner, J. Exp. Zool.,2002, 293, 478.
10 L. Addadi, S. Raz and S. Weiner, Adv. Mater., 2003, 15, 959.11 B. Hasse, H. Ehrenberg, J. Marxen, W. Becker and M. Epple,
Chem.–Eur. J., 2000, 6, 3679.12 Y. Politi, Y. Levi-Kalisman, S. Raz, F. Wilt, L. Addadi, S. Weiner
and I. Sagi, Adv. Funct. Mater., 2006, 16, 1289.13 H. Theel, Nova Acta Res. Soc. Sci. Upsala, 1892, 15, 1.14 F. Nudelman, H. H. Chen, H. A. Goldberg, S. Weiner and
L. Addadi, Faraday Discuss., 2007, 136, 9.15 L. Addadi, D. Joester, F. Nudelman and S. Weiner, Chem.–Eur. J.,
2006, 12, 980; F. Nudelman, B. A. Gotliv, L. Addadi and S. Weiner,J. Struct. Biol., 2006, 153, 176.
16 S. Weiner, Calcif. Tissue Int., 1979, 29, 163.17 H. A. Lowenstam, Bull. Marine Sci., 1989, 452, 243.18 D. Tsukamoto, I. Sarashina and K. Endo, Biochem. Biophys. Res.
Commun., 2004, 320, 1175.19 A. Veis and A. Perry, Biochemistry, 1967, 6, 2409.20 F. Marin, R. Amons, N. Guichard, M. Stiger, A. Hecker,
G. Luquet, C. Layrolle, G. Alcaraz, C. Riondet andP. Westbroek, J. Biol. Chem., 2005, 280, 33895.
21 B. Gotliv, N. Kessler, J. L. Sumerel, D. E. Morse, N. Tuross,L. Addadi and S. Weiner, ChemBioChem, 2005, 6, 304.
22 A. Berman, J. Hanson, L. Leiserowitz, T. F. Koetzle, S. Weinerand L. Addadi, Science, 1993, 259, 776.
23 J. Lahiri, G. Xu, D. M. Dabbs, N. Yao, A. Aksay and J. T. Groves,J. Am. Chem. Soc., 1997, 119, 5449.
24 L. A. Gower and D. A. Tirrell, J. Cryst. Growth, 1998, 191, 153.25 S. R. Dickinson and K. M. McGrath, J. Mater. Chem., 2003, 13,
928.26 R. Gueta, A. Natan, L. Addadi, S. Weiner, K. Refson and
L. Kronik, Ang. Chem., 2006, 119, 295.27 G. Falini, S. Albeck, S. Weiner and L. Addadi, Science, 1996,
271, 67.28 T. Kato and T. Amamiya, Chem. Lett., 1999, 3, 199.29 J. Sambrok, F. E. Fritsch and T. Maniatis, Molecular cloning:
A Laboratory Manual, Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY, 1989.
30 S. E. Darmon and K. M. Rudall, Discuss. Faraday Soc., 1950, 9,251.
31 L. B. Gower and D. J. Odum, J. Cryst. Growth, 2000, 210, 719.32 G. F. Xu, N. Yao, I. A. Aksay and J. T. Groves, J. Am. Chem.
Soc., 1998, 120, 11977.33 J. Rieger, T. Frechen, G. Cox, W. Heckmann, C. Schmidt and
J. Thieme, Faraday Discuss., 2007, 136, 265.34 H. Colfen and S. Mann, Angew. Chem., 2003, 42, 2350.35 J. Aizenberg, G. Lambert, L. Addadi and S. Weiner, Adv. Mater.,
1996, 8, 222.36 S. Raz, P. Hamilton, F. Wilt, S. Weiner and L. Addadi, Adv. Funct.
Mater., 2003, 13, 480.37 B. Pokroy, E. Zolotoyabko and N. Adir, Biomacromolecules, 2006,
7, 550.38 S. Elhadj, J. J. De Yoreo, J. R. Hoyer and P. M. Dove, Proc. Natl.
Acad. Sci. U. S. A., 2006, 103, 19237.39 D. I. Hay, E. C. Moreno and D. H. Schlesinger, Inorg. Persp. Biol.
Med., 1979, 2, 271.40 C. S. Sikes and A. P. Wheeler, Chemical Aspects of Regulation
of Mineralization, University of South Alabama PublicationServices, 1988.
41 Z. Jia and P. L. Davis, TRENDS Biochem., 2002, 27, 101.42 N. Nassif, N. Pinna, N. Gehrke, M. Antonietti, C. Jager and
H. Colfen, Proc. Natl. Acad. Sci. U. S. A., 2005, 102, 12563.43 B. A. Gotliv, L. Addadi and S. Weiner, ChemBioChem, 2003, 4,
522.
This journal is � The Royal Society of Chemistry 2007 CrystEngComm, 2007, 9, 1171–1177 | 1177
Related Documents











