REGULAR ARTICLE Aspects of soil lichen biodiversity and aggregation interact to influence subsurface microbial function Andrea P. Castillo-Monroy & Matthew A. Bowker & Pablo García-Palacios & Fernando T. Maestre Received: 4 February 2014 /Accepted: 28 August 2014 /Published online: 18 September 2014 # Springer International Publishing Switzerland 2014 Abstract Background and aims Many previous studies have evaluated aboveground–heterotrophic belowground in- teractions such as plant-soil feedbacks, plant- mycorrhizal fungi associations or plant-actinorhizal symbioses. However, few studies have used biocrusts, which are specialized soil communities of autotrophic cyanobacteria, mosses, lichens and non-photosynthetic fungi and bacteria that are prevalent in drylands world- wide. These communities largely influence ecosystem functioning, and can be used as a model system for studying above-belowground interactions. In this study, we evaluated how biocrusts affect the functional diver- sity and biomass of microbial diversities beneath biocrusts. Methods We performed two microcosm experiments using biocrust-forming lichens where we manipulated their biotic attributes to test independently the effects of species richness (from two to eight species), composi- tion, evenness (maximal and low evenness) and spatial pattern (clumped and random distribution) on the micro- bial catabolic profile and microbial functional diversity. Results Microcosms with a random pattern had a higher microbial catabolic profile than those with a clumped pattern. Significant richness × evenness × pattern and richness × evenness interactions were found when ana- lyzing microbial catabolic profile and biomass, respec- tively. Microcosms with a random pattern, intermediate number of species, and maximal evenness level had higher microbial catabolic profile. At the maximal even- ness level, assemblages had higher microbial catabolic profile and microbial biomass when they contained four species. The richness × evenness × pattern interaction was the most informative predictor of variations in microbial catabolic profile. Conclusions Our results indicate that soil microorgan- isms are influenced by biocrusts, just as they are influ- enced by plants, and highlight the importance of higher order interactions among species richness, evenness, and spatial pattern as drivers of microbial communities. The Plant Soil (2015) 386:303–316 DOI 10.1007/s11104-014-2256-9 Responsible Editor: Ute Skiba. Electronic supplementary material The online version of this article (doi:10.1007/s11104-014-2256-9) contains supplementary material, which is available to authorized users. A. P. Castillo-Monroy : F. T. Maestre Área de Biodiversidad y Conservación, Departamento de Biología y Geología, Escuela Superior de Ciencias Experimentales y Tecnología, Universidad Rey Juan Carlos, Móstoles 28933, Spain F. T. Maestre e-mail: [email protected] A. P. Castillo-Monroy (*) Departamento de Ciencias Naturales, Universidad Técnica Particular de Loja, San Cayetano Alto s/n, Loja, Ecuador e-mail: [email protected] M. A. Bowker School of Forestry, Northern Arizona University, 200 E. Pine Knoll, PO Box 15018, Flagstaff, AZ 86011, USA e-mail: [email protected] P. García-Palacios Centre d’Ecologie Fonctionnelle & Evolutive, CEFE-CNRS, 1919 route de Mende, Montpellier 34293, France e-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REGULAR ARTICLE
Aspects of soil lichen biodiversity and aggregation interactto influence subsurface microbial function
Andrea P. Castillo-Monroy & Matthew A. Bowker &
Pablo García-Palacios & Fernando T. Maestre
Received: 4 February 2014 /Accepted: 28 August 2014 /Published online: 18 September 2014# Springer International Publishing Switzerland 2014
AbstractBackground and aims Many previous studies haveevaluated aboveground–heterotrophic belowground in-teractions such as plant-soil feedbacks, plant-mycorrhizal fungi associations or plant-actinorhizalsymbioses. However, few studies have used biocrusts,which are specialized soil communities of autotrophic
cyanobacteria, mosses, lichens and non-photosyntheticfungi and bacteria that are prevalent in drylands world-wide. These communities largely influence ecosystemfunctioning, and can be used as a model system forstudying above-belowground interactions. In this study,we evaluated how biocrusts affect the functional diver-sity and biomass of microbial diversities beneathbiocrusts.Methods We performed two microcosm experimentsusing biocrust-forming lichens where we manipulatedtheir biotic attributes to test independently the effects ofspecies richness (from two to eight species), composi-tion, evenness (maximal and low evenness) and spatialpattern (clumped and random distribution) on the micro-bial catabolic profile and microbial functional diversity.Results Microcosms with a random pattern had a highermicrobial catabolic profile than those with a clumpedpattern. Significant richness × evenness × pattern andrichness × evenness interactions were found when ana-lyzing microbial catabolic profile and biomass, respec-tively. Microcosms with a random pattern, intermediatenumber of species, and maximal evenness level hadhigher microbial catabolic profile. At the maximal even-ness level, assemblages had higher microbial catabolicprofile and microbial biomass when they contained fourspecies. The richness × evenness × pattern interactionwas the most informative predictor of variations inmicrobial catabolic profile.Conclusions Our results indicate that soil microorgan-isms are influenced by biocrusts, just as they are influ-enced by plants, and highlight the importance of higherorder interactions among species richness, evenness, andspatial pattern as drivers of microbial communities. The
Plant Soil (2015) 386:303–316DOI 10.1007/s11104-014-2256-9
Responsible Editor: Ute Skiba.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11104-014-2256-9) contains supplementarymaterial, which is available to authorized users.
A. P. Castillo-Monroy : F. T. MaestreÁrea de Biodiversidad y Conservación, Departamento deBiología y Geología, Escuela Superior de CienciasExperimentales y Tecnología, Universidad Rey Juan Carlos,Móstoles 28933, Spain
F. T. Maestree-mail: [email protected]
A. P. Castillo-Monroy (*)Departamento de Ciencias Naturales, Universidad TécnicaParticular de Loja, San Cayetano Alto s/n, Loja, Ecuadore-mail: [email protected]
M. A. BowkerSchool of Forestry, Northern Arizona University,200 E. Pine Knoll, PO Box 15018, Flagstaff,AZ 86011, USAe-mail: [email protected]
P. García-PalaciosCentre d’Ecologie Fonctionnelle & Evolutive, CEFE-CNRS,1919 route de Mende, Montpellier 34293, Francee-mail: [email protected]

results also emphasize the importance of studying severalbiotic attributes simultaneously when studying biocrust-soil microorganism interactions, as in nature, communityproperties do not exert their influence in isolation.
Keywords Richness . Evenness . Spatial pattern .
Microbial biomass . Basal respiration .Microbialcommunities and biological soil crusts
Introduction
Soil organisms are intimately linked to the plant com-munity. Not only do plants (in a broad sense, inclusiveof terrestrial photo-autotrophs outside of kingdomPlantae) provide resources such as carbon and shelterto the community, but their tissues also act as a host formany soil organisms, such as herbivores, pathogens,and symbionts (Wardle et al. 2004; Donoso et al.2013). A large body of literature has studied specifictypes of autotrophic aboveground–heterotrophic below-ground interactions, such as plant-soil feedbacks (Bever2002; Kardol et al. 2006; Vivanco and Austin 2008),plant-mycorrhizal fungi associations (van der Heijdenet al. 1998; Johnson et al. 2005; Antoninka et al. 2009)or plant-actinorrhizal symbiosis (Tjepkema et al. 2000;Schwintzer and Tjepkema 2001). These interactionshave also been addressed in drylands (McHugh andGehring 2006), which collectively constitute ~41 % ofthe terrestrial surface in the form of arid, semi-arid, andsub-humid deserts, steppes, woodlands and grasslands(Maestre et al. 2012a). Biological soil crusts (biocrustshereafter) are a highly specialized community of auto-trophic cyanobacteria, mosses, and, lichens, and non-photosynthetic fungi and bacteria that usually cover thesoil surface of the open spaces located between plantpatches in drylands worldwide (Belnap and Lange2003). These communities have been seldom studiedas the autotroph component of aboveground–below-ground interactions, despite that they can be of particularutility in determining which of several properties ofautotrophic communities exert the greatest influenceon belowground heterotrophs (Bowker et al. 2010).
Biocrusts exert a strong influence on ecosystemmultifunctionality (i.e. ability of communities to simul-taneously sustain multiple ecosystem functions(Bowker et al. 2010, 2011, 2013; Maestre et al.2012b). They contribute to carbon and nitrogen fixation(Lange et al. 1992; Belnap 2002), confer resistance to
erosion (Chaudhary et al. 2009), and modulate soilnutrient mineralization, total C released by soil respira-tion and the water runoff-infiltration balance (Castillo-Monroy et al. 2010, 2011a; Eldridge et al. 2010). Inaddition, they exert a strong influence on communitiessuch as vascular plants (Green et al. 2008), microfauna(Neher et al. 2009), fungi (Bates et al. 2010) and bacteria(Yeager et al. 2004; Castillo-Monroy et al. 2011b).
Recent research suggests that biocrusts may serve asa useful model system for evaluating biodiversity-ecosystem functioning relationships in soils (Bowkeret al. 2010; Maestre et al. 2012b). Biocrusts may havesimilar value as a model for plant-soil interactions, astheir visible components are taxonomically well-definedand the manipulation of their attributes (e.g., biodiver-sity, spatial pattern and composition) in experiments isfully feasible (Maestre et al. 2012b, Bowker et al. 2014).The biodiversity and ecological importance of biocrust-associated bacteria and fungi is well documented (e.g.Billings et al. 2003; Yeager et al. 2004, Gundlapally andGarcia-Pichel 2006; Bates et al. 2010), as are patterns ofmicrobial community abundance, distribution, or com-position (Steven et al. 2013) and temporal patterns(Johnson et al. 2012) in biocrusts and underlying soils.However, only a few studies have examined how par-ticular attributes of the autotrophic components ofbiocrusts, such as their abundance or biodiversity, influ-ence the abundance and/or functioning of the associatedmicrobial communities (Castillo-Monroy et al. 2011b;García-Palacios et al. 2011; Maestre et al. 2012b;Bowker et al. 2013). In this study we evaluated, forthe first time and using a manipulative experimentalapproach, how different biodiversity components (spe-cies composition, richness, and evenness) and the spatialpattern of biocrust autotrophs affect the catabolic pro-file, functional diversity, biomass and basal respirationof the soil microbial communities beneath them.
We previously showed that species richness, compo-sition and spatial pattern affected multiple ecosystemfunctions such as organic C, total N, N availability, β-glucosidase activity, although the magnitude and direc-tion of their effects varied with the particular function,experiment and soil depth considered (Maestre et al.2012b). Here we present new measurements from thisexperiment including microbial catabolic profile, micro-bial functional diversity, basal respiration and activemicrobial biomass, and test the hypothesis that soilmicrobial functional diversity is influenced by the attri-butes of biocrust-forming autotrophs.
304 Plant Soil (2015) 386:303–316

Material and methods
Experimental design
Two manipulative microcosm experiments were con-ducted at the plant growth facilities of the Rey JuanCarlos University (Móstoles, Central Spain, 40° 20′28″ N, 3° 52′58″ O, 650 m a.s.l.) under natural light,temperature and rainfall conditions between June 2006and December 2008. Soil and biocrust-forming lichenspecies for these experiments were collected from gyp-sum outcrops located in the surroundings of theUniversity. The species used in the experiments wereselected among the pool of the ten most common lichenspecies found in gypsum outcrops from Central Spain(Maestre et al. 2008; Castillo-Monroy et al. 2010):Acarospora nodulosa (Dufour) Hue., Collema crispum(Hudson) Weber., Diploschistes diacapsis (Ach.)Lumbsch, Squamarina lentigera (Weber) Poelt.,Fulgensia subbractaceata (Nyl.) Poelt., Leprariamembranaceum (Dickson) Vainio., Psora decipiens(Hedw.) Hoffm., Cladonia convoluta (Lam.) Anders.,Squamarina cartilaginea (With.) P. James. and Toniniasedifolia (Scop.) Timdal.
We created microcosms with lichen colonies placedover the surface of soil, with a constant lichen cover of60 %, but imposed variation in species richness, even-ness, spatial patterning (aggregated vs. random), anddifferent composition (Fig. 1; Maestre et al. 2012b fordetails). The first experiment (hereafter Composition
experiment) was designed to independently test for theeffects of species richness (Ri), species composition(Co), and spatial pattern (Pa) on microbial catabolicprofile and functional diversity. Four Co levels werenested within two Ri levels (four and eight species).Each combination of Co and Ri was established undertwo Pa: clumped and random. Each combination of Ri(2), Co (4) and Pa (2) was replicated six times for a totalof 96 microcosms (Appendix A). The second experi-ment (hereafter Evenness experiment) was set up toindependently test for the effects of Ri (two, four andeight species), species evenness (Ev) (maximalevenness vs. communities with a geometricdistribution of abundances among species; Wilsey andPolley 2004) and Pa (clumped and random). Each com-bination Ri (3), Ev (2) and Pa (2) was replicated sixtimes for a total of 72 microcosms (Appendix B).
Characterization of microbial communities and activity
At the end of the experiment, a composite soil sample(0–2 cm depth) from the portion of the microcosmcovered by lichens was obtained. The lichens wereremoved prior to sampling, and the samples were storedin plastic bags at −80 °C until analyses.
We analyzed soil heterotrophic microbial functionaldiversity using MicroResp™ (Campbell et al. 2003).This is a whole-soil method based on community levelphysiological profiles obtained by testing different car-bon sources that vary in structural complexity and that
Fig. 1 Artificial biocrust microcosm created using the “mosaic”technique. Examples of microcosms with a random (a) andclumped (b) spatial pattern and intact lichen pieces collected from
the field, cut into homogeneous 0.5 cm-side square fragments (c).See Maestre et al. 2012a for details
Plant Soil (2015) 386:303–316 305

provide information about the ability of the microbialcommunities to catabolize different C sources (Oren andSteinberger 2008a). We modified the protocol of theMicroResp™ system to include C-compounds producedby lichens (e.g., phenolic acids). We used amino acids(L-alanine, L-cysteine-HCl and N-acetyl-glucosamine),carbohydrates (D-fructose, D-galactose, D-glucose andL-arabinose), carboxylic acids (citric acid, L-malic acid,oxalic acid and γ amino butyric acid), phenolic acids(orcinol and anthraquinone) and fatty acids (Tween 80and dextrin). In functional terms, the substrate utiliza-tion rates of the carbon sources correspond to the cata-bolic attributes of different soil microbial functionalgroups (Zak et al. 1994). Even if we cannot assessmicrobial communities in relation to taxonomic or phy-logenetic diversity (Øvreås 2000), we can useMicroResp™ data to interpret changes inmicrobial func-tional diversity (Oren and Steinberger 2008a; García-Palacios et al. 2011). Prior to MicroResp™ analyses,defrosted soils were introduced into deep well platesand pre-incubated for 5 days at 25 °C. The moisturewithin the plates was corrected to 40 % water holdingcapacity to condition the soils and reestablish activemicrobial populations. The plates were then incubatedfor 6 h and read at 595 nm. The results were calculatedon the basis of the 16th substrate (water), which repre-sents the basal respiration, as explained in García-Palacios et al. (2011). Although potential changes inmicrobial communities may have occurred due tofreeze-thaw cycles, samples are still comparable be-cause all the soils used in this study were subjected tothe same storage conditions.
Microbial catabolic profile was characterized as thedata matrix of the respiration of the carbon sources used.Using these data, we also calculated the Shannon-Weaver index as H′= −∑Pi (ln Pi), where Pi is theactivity of a particular carbon source/the sum of activityof all carbon sources (Yu et al. 2012). Thus, high H′values would indicate a greater capacity of the microbialcommunity to catabolize a gradient of C recalcitrance.
Control wells were amended with water only, whichprovided basal respiration and glucose-induced respira-tion; this was assumed to be proportional to activemicrobial biomass (Oren and Steinberger 2008b, BenSassi et al. 2012). The metabolic quotient (qCO2) wasalso calculated as the ratio between basal respiration andmicrobial biomass (Oren and Steinberger 2008b). Alldependent variables used in the entire study were mea-surements from the MicroResp assay.
Statistical analyses
We analyzed the results of the two experiments sepa-rately. To evaluate the effects of the different factorsevaluated on univariate (H′, basal respiration, microbialbiomass, qCO2) and multivariate (microbial catabolicprofile) response variables obtained from MicroResp,we used a three-way nested ANOVA/MANOVA modelin the Composition experiment (Ri and Pa were consid-ered fixed factors, while Co was a random factor nestedwithin Ri), and a factorial ANOVA/MANOVAmodel inthe Evenness experiment (Ri, Pa and Ev were consid-ered fixed factors). In the Composition experiment,species richness was tested against composition, speciescomposition(richness) and the spatial pattern ×composition(richness) interaction were tested againstthe error term, and spatial pattern and the spatial pattern× species richness interaction were tested against thespatial pattern × composition(richness) interaction(Doncaster and Davey 2007). In the Evenness experi-ment, all the factors were tested against the error term.The semiparametric PERMANOVA approach(Anderson 2001) was preferred to traditional ANOVAbecause our response variables did not follow ANOVA/MANOVA assumptions (normality and homogeneity ofvariances). All PERMANOVA analyses were conduct-ed using the Euclidean distance and 9999 permutationsof the raw data. To aid in the interpretation of the effectsof the factors when analyzing multivariate MicroRespdata (microbial catabolic profile), we also conducted aprincipal coordinate analysis (PCO; Anderson et al.2008) using the Euclidean distance.
We also used structural equation models (SEM;Grace 2006), which can be helpful in determiningthe relative influence of multiple factors on the re-sponse variables. SEMs have a variety of uses, includ-ing improved causal inference from observational data,and partitioning of direct and indirect effects (Shipley2000; Grace 2006). They are also useful for answeringquestions related to relative importance of factors be-cause effect sizes can be described in a commoncurrency, the path coefficient. A path coefficientexpressed the strength of the effect of one variableupon another. In their standardized format, rangingfrom 0 to 1, path coefficients are equivalent to partialcorrelation coefficients or regression weights.Associated with each path coefficient estimate is aprobability test that estimates the probability that thepath estimate is equal to zero.
306 Plant Soil (2015) 386:303–316

SEMs can evaluate the interrelationships amongmul-tiple variables, but does not directly analyze multivariateresponses. To express the microbial catabolic profiledata in a format compatible with this technique, weconducted some data reduction using a PrincipalComponents Analysis (PCA) on the correlation matrixof the different carbon sources used in the MicroRespanalyses. In both Experiments, the first component ex-plained a large proportion of the variation, (65 and 69%,respectively), thus it was used as a parsimonious sum-mary of these data.
Our SEMs are constructed similarly to a multipleregression with experimental factors exerting an in-fluence on the PCA components. However, theydiffer from multiple regressions in several ways; i)spatial pattern and evenness were coded as 0 or 1; ii)composition was treated using a composite variable.Composite variables have multiple uses, but herethey function to sum together the effects of thelevels of a categorical variable, which are represent-ed by dummy variables. This use of the compositevariable is a graphical and numerical interpretationalaid that does not alter the underlying model (Grace2006). It simply sums together the effects of multi-ple conceptually related variables upon another, col-lapsing them into a single path coefficient; iii) weconstructed an interaction term for the Ri effect inthe Evenness experiment. First we viewed a plot ofmeans and confidence intervals of the microbialPCA axis as a function of this interaction term. Itwas clear that the samples driving this interactiveeffect were those with a random spatial pattern, highevenness, and intermediate richness. Thus the sim-plest method to express this was to create anotherbinary coded variable, wherein samples with thistreatment combination were coded as “1” and therest were coded as “0”; iv) richness and compositionwere explicitly allowed to covary because they arecorrelated due to nesting in the Composition exper-iment; and v) the Ri × Ev × Pa term was allowed tocovary with the main effects in the Evenness exper-iment, because it is mathematically derived fromthem.
An overall goodness-of-fit test is usually per-formed with a SEM. It tests whether the proposedstructure of the model is a reasonable representationof the causal relationships underlying the correla-tions among variables. In our case, the causal struc-ture of the system is known with certainty because
most variables are experimental factors which canaffect a response, but cannot be affected by a re-sponse. Nevertheless, we verified model fit, usingthe traditional χ2 goodness-of-fit test to assure thatwe had not misspecified the models. Unlike mostprobability tests, this goodness-of-fit tests the prob-ability that the model fits the data (or more precise-ly, that the covariance matrix implied by the modelstructure is similar to the covariance matrix deriveddirectly from the data). Upon verifying fit, we obtainedestimates of path coefficients and associated probabilitylevels. PERMANOVA, SEM, and PCA analyses wereconducted using PERMANOVA + for PRIMER statis-tical package (PRIMER-E Ltd., Plymounth MarineLaboratory, UK), Amos 18.0 statistical software (SPSSInc., Chicago, IL, USA), and JMP 4.0 (SAS InstituteInc., Cambridge, Ma, USA), respectively. The rawMicroResp data from our experiments are availablefrom figshare (Castillo-Monroy and Maestre 2014; seeappendices I and J for treatment summaries).
Results
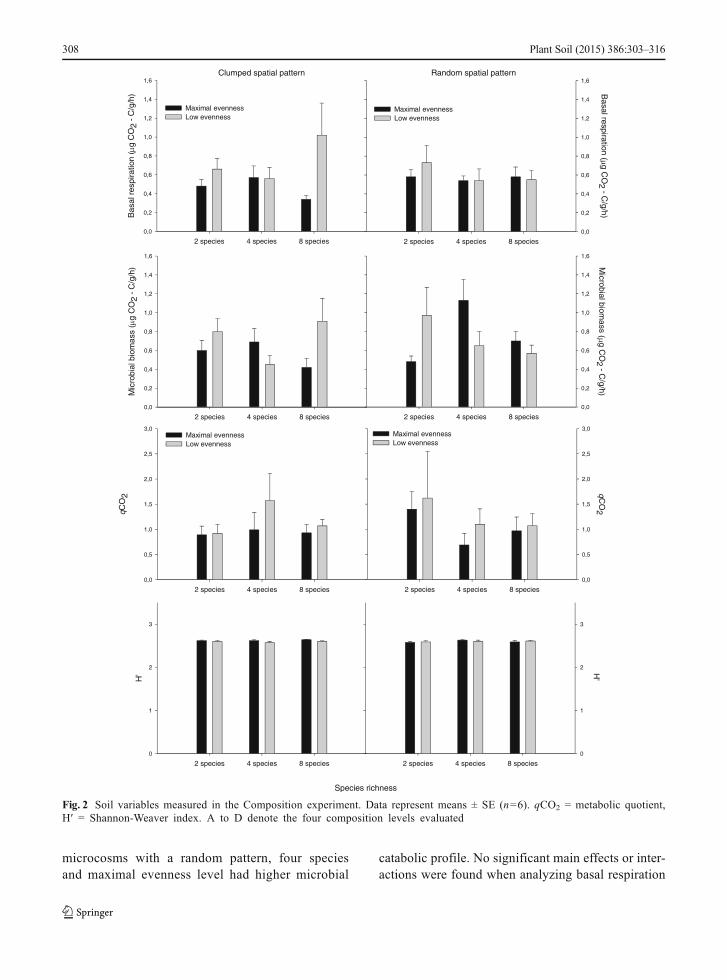
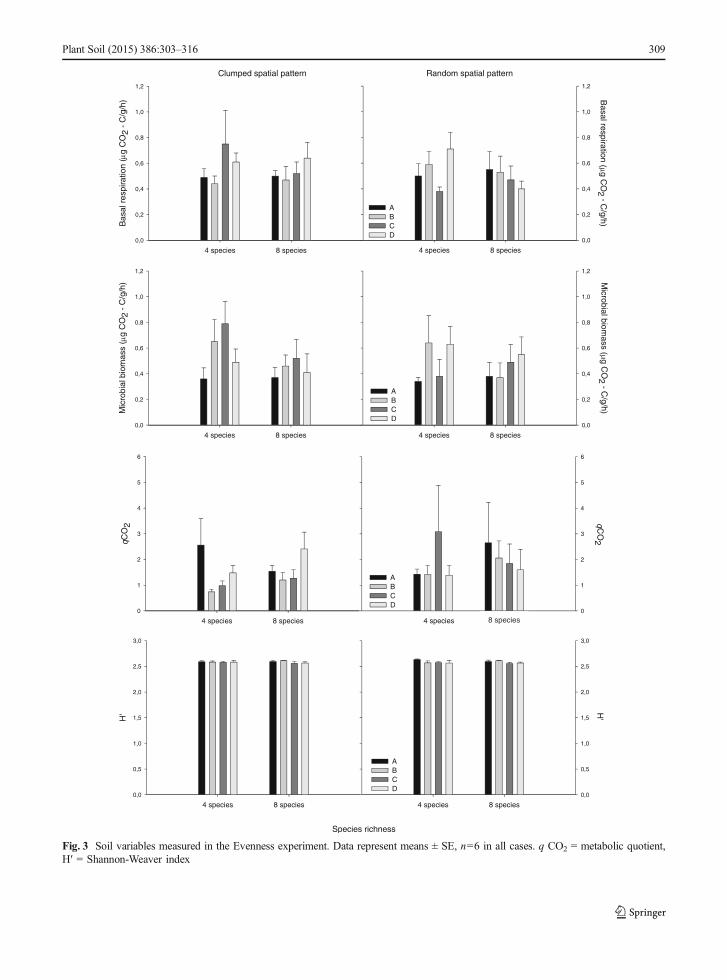
We did not find any significant effect of attributes ofbiocrust on the soil microbial catabolic profile, H′,microbial biomass, basal respiration and metabolicquotient when analyzing data from the Compositionexperiment (Fig. 2, Appendices C, E). However, wefound a significant effect of Pa (F1,60 = 3.111, P=0.042) on microbial catabolic profile in the Evennessexperiment; randomly patterned samples had highermicrobial catabolic profile that those with a clumpedpattern. Significant Ri × Ev × Pa (F1,60 = 2.915, P=0.025) and Ri × Ev (F1,60 = 3.114, P=0.021) interac-tions were also found when analyzing microbial cata-bolic profile in the Evenness experiment (Fig. 3,Appendices D, F). A significant Ri × Ev interactionwas also found when analyzing microbial biomassfrom this experiment (F1,60 = 5.248, P=0.008). Weinvestigated these interactions by conducting sepa-rate PERMANOVAs for each Ev level. At the max-imal evenness level, assemblages had higher micro-bial catabolic profile and biomass when theycontained four species (Appendix H). At this even-ness level, a significant effect of Pa was also found(F1,30 = 3.889, P=0.027), with randomly patternedmicrocosms having higher microbial catabolic pro-file than those with a clumped pattern. Overall,
Plant Soil (2015) 386:303–316 307
microcosms with a random pattern, four speciesand maximal evenness level had higher microbial
catabolic profile. No significant main effects or inter-actions were found when analyzing basal respiration
Clumped spatial pattern
2 species 4 species 8 species
Bas
al r
espi
ratio
n (
g C
O2
- C
/g/h
)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
Maximal evenness Low evenness
Random spatial pattern
2 species 4 species 8 species
Basal respiration (
g CO
2 - C/g/h)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
Maximal evenness Low evenness
2 species 4 species 8 species
Mic
robi
al b
iom
ass
(g
CO
2 -
C/g
/h)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
2 species 4 species 8 species
Microbial biom
ass (g C
O2 - C
/g/h)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
2 species 4 species 8 species
qCO
2
0,0
0,5
1,0
1,5
2,0
2,5
3,0Maximal evenness Low evenness
2 species 4 species 8 species
q CO
2
0,0
0,5
1,0
1,5
2,0
2,5
3,0Maximal evenness Low evenness
2 species 4 species 8 species
H'
0
1
2
3
Species richness
2 species 4 species 8 species
H'
0
1
2
3
Fig. 2 Soil variables measured in the Composition experiment. Data represent means ± SE (n=6). qCO2 = metabolic quotient,H′ = Shannon-Weaver index. A to D denote the four composition levels evaluated
308 Plant Soil (2015) 386:303–316
Clumped spatial pattern
4 species 8 species
Bas
al r
espi
ratio
n (
g C
O2
- C
/g/h
)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
4 species 8 species
Basal respiration (
g CO
2 - C/g/h)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
A B C D
Random spatial pattern
4 species 8 species
Mic
robi
al b
iom
ass
(g
CO
2 -
C/g
/h)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
4 species 8 species
Microbial biom
ass (g C
O2 - C
/g/h)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
A B C D
4 species4 species 8 species
qCO
2
0
1
2
3
4
5
6
q CO
2
0
1
2
3
4
5
6
A B C D
4 species 8 species
H'
0,0
0,5
1,0
1,5
2,0
2,5
3,0
Species richness
4 species 8 species
H'
0,0
0,5
1,0
1,5
2,0
2,5
3,0
A B C D
4 species8 species
Fig. 3 Soil variables measured in the Evenness experiment. Data represent means ± SE, n=6 in all cases. q CO2 = metabolic quotient,H′ = Shannon-Weaver index
Plant Soil (2015) 386:303–316 309

and metabolic quotient from the Evenness experiment(Appendix D, F).
None of the biocrust community factors evaluatedaffected the H′, in either experiment (Figs. 2 and 3;Appendices E, F). However when we analyzed H′ forthe five carbon groups independently, we found thatmicrocosms with a random Pa had higher H′ values ofamino acid than those with a clumped Pa (F1,60 = 5.021,P=0.028; Fig. 4). Among the five carbon groups, car-bohydrates had the highest H′ values followed by car-boxylic, amino acid and then fatty and phenolic acids,regardless the factor evaluated (Fig. 4).
Our SEMs provided estimates on the relative effectsof the attributes of biocrust upon the microbial catabolic
profile (Table 1). We did not find significant main orinteractive effects in the Composition experiment, butwere most successful modeling the Evenness experi-ment, explaining nearly 20 % of the variance found inthis response variable (Fig. 5). In this experiment, the Ri× Ev x Pa interaction (r=0.35, P<0.002) was the mostinformative predictor of variation in microbial catabolicprofile. However, we did not find a significant effect ofthe Ri x Ev interaction on this variable (Fig. 5), despitethat this interaction was significant when we analyzedthe data using PERMANOVA (F1,60 = 3.114, P=0.021).No direct effects of biocrust attributes were detected ineither experiment (Fig. 5). Together, the SEM andPERMANOVA results suggest that the most important
Cl_2sp Cl_4sp Cl_8sp Ra_2sp Ra_4sp Ra_8sp
H'
0,10
0,15
0,20
0,25
0,30
0,35
0,40
CarbohydratesCarboxylic acid Amino acid Fatty acid Phenoloc acid
(A) Maximal evenness
(B) Geometic distribution
Cl_2sp Cl_4sp Cl_8sp Ra_2sp Ra_4sp Ra_8sp
H'
0,10
0,15
0,20
0,25
0,30
0,35
0,40
Fig. 4 Shannon-Weaver index(H′) of five carbon groupsobtained in the Evennessexperiment. Cl_2sp, Cl_4sp andCl_8sp refer to microcosms withclumped spatial pattern and two,four and eight species,respectively. Ra_2sp, Ra_4sp andRa_8sp refer to microcosms withrandom spatial pattern with two,four and eight species,respectively. Microcosms withmaximal evenness (a),Microcosms with a geometricdistribution of abundances amongspecies (b). Data are means ± SE(n=6)
310 Plant Soil (2015) 386:303–316
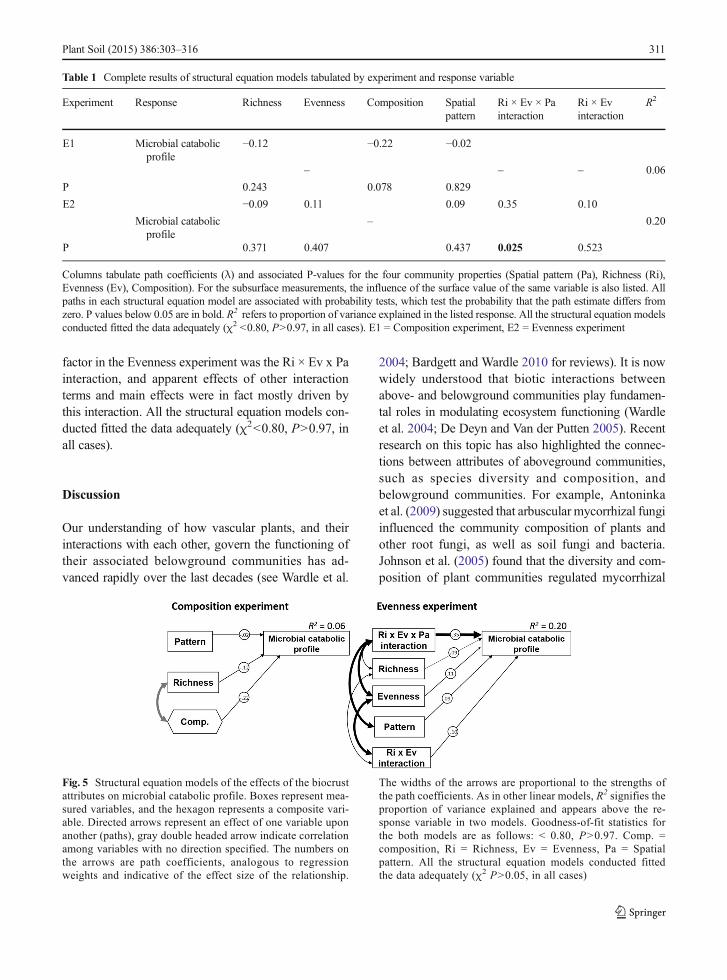
factor in the Evenness experiment was the Ri × Ev x Painteraction, and apparent effects of other interactionterms and main effects were in fact mostly driven bythis interaction. All the structural equation models con-ducted fitted the data adequately (χ2<0.80, P>0.97, inall cases).
Discussion
Our understanding of how vascular plants, and theirinteractions with each other, govern the functioning oftheir associated belowground communities has ad-vanced rapidly over the last decades (see Wardle et al.
2004; Bardgett and Wardle 2010 for reviews). It is nowwidely understood that biotic interactions betweenabove- and belowground communities play fundamen-tal roles in modulating ecosystem functioning (Wardleet al. 2004; De Deyn and Van der Putten 2005). Recentresearch on this topic has also highlighted the connec-tions between attributes of aboveground communities,such as species diversity and composition, andbelowground communities. For example, Antoninkaet al. (2009) suggested that arbuscular mycorrhizal fungiinfluenced the community composition of plants andother root fungi, as well as soil fungi and bacteria.Johnson et al. (2005) found that the diversity and com-position of plant communities regulated mycorrhizal
Table 1 Complete results of structural equation models tabulated by experiment and response variable
Experiment Response Richness Evenness Composition Spatialpattern
Ri × Ev × Painteraction
Ri × Evinteraction
R2
E1 Microbial catabolicprofile
−0.12 −0.22 −0.02
– – – 0.06
P 0.243 0.078 0.829
E2 −0.09 0.11 0.09 0.35 0.10
Microbial catabolicprofile
– 0.20
P 0.371 0.407 0.437 0.025 0.523
Columns tabulate path coefficients (λ) and associated P-values for the four community properties (Spatial pattern (Pa), Richness (Ri),Evenness (Ev), Composition). For the subsurface measurements, the influence of the surface value of the same variable is also listed. Allpaths in each structural equation model are associated with probability tests, which test the probability that the path estimate differs fromzero. P values below 0.05 are in bold. R2 refers to proportion of variance explained in the listed response. All the structural equation modelsconducted fitted the data adequately (χ2 <0.80, P>0.97, in all cases). E1 = Composition experiment, E2 = Evenness experiment
Fig. 5 Structural equation models of the effects of the biocrustattributes on microbial catabolic profile. Boxes represent mea-sured variables, and the hexagon represents a composite vari-able. Directed arrows represent an effect of one variable uponanother (paths), gray double headed arrow indicate correlationamong variables with no direction specified. The numbers onthe arrows are path coefficients, analogous to regressionweights and indicative of the effect size of the relationship.
The widths of the arrows are proportional to the strengths ofthe path coefficients. As in other linear models, R2 signifies theproportion of variance explained and appears above the re-sponse variable in two models. Goodness-of-fit statistics forthe both models are as follows: < 0.80, P>0.97. Comp. =composition, Ri = Richness, Ev = Evenness, Pa = Spatialpattern. All the structural equation models conducted fittedthe data adequately (χ2 P>0.05, in all cases)
Plant Soil (2015) 386:303–316 311

communities in forests and grasslands. However, al-though the biocrusts represent the dominant soil surfacesystem in drylands, covering up to 70% of their surfaces(Belnap and Lange 2003), very few studies so far haveevaluated how attributes of biocrusts affect soil micro-bial communities (Castillo-Monroy et al. 2011b, García-Palacios et al. 2011, Yu et al. 2012). The results of ourexperiments indicate that soil microorganisms are influ-enced by biocrusts, and highlight the importance ofhigher order interactions among species richness, even-ness, and spatial pattern as drivers of the functioning ofmicrobial communities.
Most of the research on the relationships betweenbiodiversity and ecosystem functioning conducted todate has focused on only one community attribute at atime, such as species richness (Hooper et al. 2005;Cardinale et al. 2011), species composition (Petersenet al., 2012), evenness (Wilsey and Polley 2004;Downing 2005) or spatial pattern (Maestre et al.2005). While the need of understanding the relation-ships among different biodiversity attributes as determi-nants of ecosystem functioning has been alreadyhighlighted (Loreau et al. 2001; Maestre et al., 2012a),few studies so far have focused on studying how thesefactors determine associated soil microorganisms (butsee, Johnson et al. 2005; Antoninka et al. 2009).Furthermore, and to our knowledge, only one previousstudy has evaluated how multiple attributes of biocrustssimultaneously influence microbial communities(Castillo-Monroy et al. 2011b).
We did not find significant effects of species richnesson any of the microbial attributes measured. Theseresults were unexpected because autotrophic richnesswould be expected to influence heterotrophic functionaldiversity through its effects on substrate type and quan-tity. Further, autotrophic richness was previously foundto be related to different soil functions under biocrusts,(e.g. Bowker et al. 2010, 2013; Maestre et al. 2012b),and one might hypothesize this effect to be partiallymediated by the catabolic profile of heterotrophs. Ourresults might suggest that autotrophic richness does notstrongly influence community composition of microbesin the biocrust system. In agreement with our findings,Castillo-Monroy et al. (2011b) did not find a significanteffect of biocrust richness on the abundance and/orrichness of soil bacteria. Results from that study indicatethat the link between richness of biocrusts-forming li-chens and their associated belowground communitieswas not very strong, but that particular lichen species,
such as Collema crispum and Toninia sedifolia, exertedimportant effects on particular bacterial taxa. On theother hand, it is also possible that autotrophic richnessdoes affect the heterotrophic species composition, butnot enough to change the microbial catabolic profile,possibly due to functionally redundancy. It is wellknown that soil microorganisms comprise an incrediblylarge number of species, and because there are largenumbers of trophically equivalent organisms, many spe-cies may be functionally redundant (Setälä et al. 2005).In the same set of experiments used here, Maestre et al.(2012b) reported that biocrust richness had higher rela-tive importance compared to other attributes evaluatedin determining soil nutrient stocks and cycling. It ap-pears from the current study that this effect of autotro-phic richness was not attributable to the catabolic activ-ity of soil microbes. The results of Castillo-Monroyet al. (2011b) also suggested that functional profile ofbiocrusts were more clearly attributable to the richnessof autotrophic biocrust components rather than that ofthe heterotrophic ones.
In the Evenness experiment, microcosms with a ran-dom spatial pattern had higher microbial catabolic pro-file than those with a clumped spatial pattern. Eachlichen species is expected to exert a zone of influencein the vicinity of the thallus, and the microbial commu-nity would be likely to respond differently to the influ-ences of different species due to the variation in chem-ical constituents of the lichens, something that has al-ready been observed with rock-colonizing lichens(Bates et al. 2010). A random, rather than aggregatedspatial pattern would tend to isolate different zones ofinfluence, where a single species is able to exert its ownunique set of influences upon the microbial community.This assortment of unique influences might increasemicrobial niche space, leading to a higher catabolicactivity across the entire sample. Also, it is likely thatthe distribution of resources under the random spatialpattern of lichens would be heterogeneous, with isolatedareas being resource rich; thus, enabling the acquisitionof resources for soil microorganisms without intensecompetition. Our results highlight the importance thatthe spatial pattern of biocrusts-forming lichens mayhave for the associated microbial communities, andadd to recent observational and experimental studiesshowing how the spatial pattern of primary producerscan affect that of consumers and the ecosystem process-es that depend on them (e.g. Maestre et al. 2005, 2012b;Pringle et al. 2010).
312 Plant Soil (2015) 386:303–316

The H′ index (our surrogate of microbial functionaldiversity) of amino acids were higher in microcosmswith random patterns. This was not a surprising result,as certain biocrust constituents, such as cyanobacteriaand some bacterial species, have been reported to beable to liberate quantities of free amino acids into theexternal environment (Antarikanonda 1984;Gundlapally and Garcia-Pichel 2006). Although wedid not measure cyanobacteria in our microcosms,free-living cyanobacteria are commonly associated withbiocrusts-forming lichens such as those used in ourexperiment (Maestre et al. 2006). Phenolic acids areproduced by lichens (Lawrey 1995), and thus the addi-tion of them to soil can stimulate those microbial pop-ulations using them as a carbon source (Blum 2011).However, the H′ values of phenolic acids were lowerthan those of other carbon sources (Fig. 4), suggestingthat microbial communities were not able to efficientlycatabolize the phenolic compounds added in our exper-iment as well as we expected. This could be due to thefact that most of the lichen substances are insoluble orpoorly soluble in water (Huneck and Yoshimura 1996).Additionally, only some microorganisms have the ca-pacity to utilize phenolic compounds as a carbon source(Kefeli et al. 2013), and they could be absent in ourmicrocosms.
Microbial communities in our microcosms were ableto metabolize carbohydrates more efficiently than othercarbon sources, which meant a higher respiration ratecompared to other carbon sources (Fig. 4). Thus, micro-bial communities underneath lichens could be usingsubstances produced by those lichens, since they,through their photobiont (i.e. alga or cyanobacterium),produce carbohydrates via photosynthesis which thenserve as food for their fungus (Nash 1996). Besides,carbohydrates serve as major source of carbon for thegrowth of many microorganisms (Wright 1984;Goldman et al. 1987). These results were fully expectedsince carbohydrates are the most generally utilized car-bon source by soil microbes (Stevenson 1993).
Species evenness did not significantly affect any ofthe variables evaluated. This result contrasts withBowker et al. (2010) and Wilsey and Polley (2004),who found significant effects of species evenness onindividual ecosystem functions driven by biocrusts andplants, respectively. It is not surprising that our result aredifferent from those above mentioned since it is usual tofind discrepancies in experimental and observationalstudies when evaluating the same variables or
treatments. However, we can interpret our result as thatin uneven communities where one species attains dom-inance, many individual zones of influence of a lichenthallus may be essentially interchangeable because theyare associated with the same species, and thus the influ-ence of rare lichens species on microbial niche space inthe whole sample may be harder to discern. In an evencommunity, each lichen exerts its unique set of influ-ences equally strongly across the whole sample.Another hypothesis is that natural communities are ei-ther even or not due to specific reasons which shapecommunity structure, such as a legacy of species inter-actions. On contrast, an artificially even community isnot shaped by these forces. Thus experimentally-induced evenness may not have the same effects asnaturally-occurring evenness as studied in Bowkeret al. (2010).
In the Evenness experiment, when the effects of thedifferent factors were evaluated using a semiparametricapproach, it was found that significant species richness× evenness interactions modulated microbial catabolicprofile and biomass. The same interaction was foundwhen analyzing soil organic C in this experiment(Maestre et al. 2012b). Thus, our results suggest thatthe higher the active microbial biomass, the higheravailability of carbon on soils provided by the fixationof atmospheric CO2 by lichens, at least within the rangeexplored in our experiments.
A third-order interaction among species richness,evenness and spatial pattern was a major determinantof variations in microbial catabolic profile in ourEvenness experiment (Fig. 5). This result highlightsthe importance of studying several biotic attributes si-multaneously to create a realistic image of natural con-ditions, as community properties do not exert their in-fluence in isolation in the field. However, understandingwhy intermediate richness might, in concert with max-imal evenness and random spatial patterning, promotemicrobial catabolic activity is not easy. Using a minia-ture model of plant-soil interactions in constructedbiocrusts, we asked the question: What biotic attributesof autotrophic communities structure the communitiesof soil heterotrophic microbes? A major finding of ourstudy is that interactive effects of multiple biotic attri-butes may dictate the structure of soil microbial com-munities. Therefore, there is a clear need for multi-factorcrossed experiments to arrive at generalities regardinghow aboveground communities influence microbialcommunities and the soil functions that depend on them.
Plant Soil (2015) 386:303–316 313

Biocrusts are an ideal system to evaluate such questions,as we can experimentally manipulate multiple bioticattributes within a community at low cost and withlow space requirements, and their increased use withthis aim will undoubtedly increase our understanding oftheir role as drivers of soil functioning.
Acknowledgments We thank David Elliott and two anonymousreviewers for comments on a previous version of this manuscript.We also thank A. Escudero, I. Martínez, P. Alonso, E. Polaina, S.Soliveres, M. D. Puche, Y. Valiñani, C. Escolar, E. Barahona, S.Beltran de Guevara, C. Iriarte, J. Margalet, C. Díaz, R. Sendra, B.Paredes, L. Giménez-Benavides, Y. Cabrea, I. Conde and I. Pardofor their invaluable help during the development of this work.APCM was supported by a Studentship from the FundaciónBBVA (BIOCON06/105 grant). This research was funded by theBritish Ecological Society (Early Career Project Grant 231/607)and by the European Research Council under the EuropeanCommunity’s Seventh Framework Programme (FP7/2007-2013)/ERC Grant agreement n° 242658 (BIOCOM).
References
Anderson MJ (2001) A new method for non-parametric multivar-iate analysis of variance. Aust Ecol 26:32–46
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA +for PRIMER: guide to software and statistical methods.PRIMER-E, PlymouthUK
Antarikanonda P (1984) Production of extracellular free aminoacids by cyanobacterium Anabaena siamensis. CurrMicrobiol 11:191–195
Antoninka A, Wolf JE, Bowker MA, Classen AT, Johnson NC(2009) Linking above- and belowground responses to globalchange at community and ecosystem scales. Global ChangeBiol 15:914–929
Bardgett R, Wardle D (2010) Aboveground-belowground link-ages: biotic interactions, ecosystem processes, and globalchange. Oxford University Press, Oxford
Bates ST, Nash TH, Sweat KG, Garcia-pichel F (2010) Fungalcommunities of lichen-dominated biological soil crusts:Diversity, relative microbial biomass, and their relationshipto disturbance and crust cover. J Arid Environ 74:1192–1199
Belnap J (2002) Nitrogen fixation in biological soil crusts fromsoutheast Utah, USA. Biol Fertil Soils 35:128–135
Belnap J, Lange OL (2003) Biological soil crusts: structure func-tion and management. Springer, Berlin
Ben Sassi M, Dollinger J, Renault P, Tlili A, Bérard A (2012) TheFungiResp method: an application of the MicroResp™ meth-od to assess fungi in microbial communities as soil biologicalindicators. Ecol Indic 23:482–490
Bever JD (2002) Soil community feedback and the coexistence ofcompetitors: conceptual frameworks and empirical tests.New Phytol 157:465–473
Billings SA, Schaeffer SM, Evans RD (2003) Nitrogen fixation bybiological soil crusts and heterotrophic bacteria in an intactMojave Desert ecosystem with elevated CO2 and added soilcarbon. Soil Biol Biochem 35:643–649
Bowker MA, Maestre FT, Escolar C (2010) Biological crusts as amodel system for examining the biodiversity-ecosystemfunction relationship in soils. Soil Biol Biochem 42:405–417
Bowker MA, Maestre FT, Mau R (2013) Diversity and patch-sizedistributions of biological soil crusts regulate dryland eco-system multifunctionality. Ecosystems 16:923–933
Bowker MA, Maestre FT, Eldridge D, Belnap J, Castillo-MonroyA, Escolar C, Soliveres S (2014) Biological soil crusts as amodel system in community, landscape and ecosystem ecol-ogy. Biodivers Conserv. doi:10.1007/s10531-014-0658-x
Bowker MA, Mau RL, Maestre FT, Escolar C, Castillo-MonroyAP (2011) Functional profiles reveal unique ecological rolesof various biological soil crust organisms. Funct Ecol 25:787–795
Blum U (2011) Plant-plat allelopathic interactions: phenolic acids,cover crops and weed emergence. Springer science, NY
Campbell CD, Chapman SJ, Cameron CM, Davidson MS, PottsJM (2003) A rapidmicrotiter plate method tomeasure carbondioxide evolved from carbon substrate amendments so as todetermine the physiological profiles of soil microbial com-munities by using whole soil. Appl Environ Microbiol 69:3593–3599
Cardinale BJ, Matulich KL, Hooper DU, Byrnes JE, Duffy E,Gamfeldt L, Balvanera P, O’Connor MI, González A (2011)The functional role of producer diversity in ecosystems. Am JBot 98:572–592
Castillo-Monroy AP, Maestre FT (2014) Data from “Aspects ofsoil lichen biodiversity and aggregation interact to influencesubsurface microbial function”. figshare. doi:10.6084/m9.figshare.1157787
Castillo-MonroyAP,Maestre FT, Delgado-BaquerizoM, GallardoA (2010) Biological soil crust modulate nitrogen availabilityin semi-arid ecosystem: insights froma Mediterranean grass-land. Plant Soil 333:21–34
Castillo-Monroy AP, Maestre FT, Rey A, Soliveres S, García-Palacios P (2011a) Biological soil crust microsites are themain contributor to soil respiration in a semiarid ecosystem.Ecosystems 14:835–847
Castillo-Monroy AP, Bowker MA, Maestre FT, Rodríguez-Echeverría S, Martínez I, Barraza-Zepeda CE, Escolar C(2011b) The relative importance of biological soil crust andsoil bacterial diversity and abundance as drivers of ecosys-tem functioning in a semi-arid environment. J Veg Sci 22:165–174
Chaudhary VB, Bowker MA, O’Dell TE, Grace JB, Redman AE,Rillig MC, Johnson NC (2009) Untangling the biologicalcontributions to soil stability in semiarid shrublands. EcolAppl 19:110–122
Donoso DA, JohnstonMK, Clay N, Kaspari ME (2013) Trees andseasonality as templates for trophic structure of tropical litterarthropod communities. Soil Biol Biochem 61:45–61
Downing AL (2005) Relative effects of species composition andrichness on ecosystem properties in ponds. Ecology 86:701–715
De Deyn GB, Van der Putten WH (2005) Linking abovegroundand belowground diversity. Trends Ecol Evol 20:625–633
Doncaster CP, Davey AJH (2007) Analysis of variance and co-variance. Cambridge University Press, Cambridge
Eldridge DJ, Bowker MA, Maestre FT, Alonso P, Mau RL,Papadopolous J, Escudero A (2010) Interactive effects of
314 Plant Soil (2015) 386:303–316

three ecosystem engineers on infiltration in a semi-arid grass-land. Ecosystems 13:499–510
García-Palacios P, Bowker MA, Maestre FT, Soliveres S,Valladares F, Papadopoulos J, Escudero A (2011)Ecosystem development in roadside grasslands: biotic con-trol, plant–soil interactions, and dispersal limitations. EcolAppl 21:2806–2821
Goldman JC, Caron DA, Dennett MR (1987) Regulation ofgross growth efficiency and ammonium regeneration inbacteria by substrate C:N ratio. Limnol Oceanogr 32:1239–1252
Grace JB (2006) Structural equation modeling and natural sys-tems. Cambridge University Press, Cambridge
Green LE, Porras-Alfaro A, Sinsabaugh RL (2008) Translocationof nitrogen and carbon integrates biotic crust and grass pro-duction in desert grassland. J Ecol 96:1076–1085
Gundlapally SR, Garcia-Pichel F (2006) The community andphylogenetic diversity of biological soil crusts in theColorado Plateau Studied by molecular fingerprinting andintensive cultivation. Microb Ecol 52:345–357
Huneck S, Yoshimura I (1996) Identification of lichen substances.Springer, Berlin
Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, Lavorel S,Lawton HD, Lodge M, Loreau M, Naeem V, Schmid B,Setälä H, Symstad AJ, Vandermeer J, Wardle DA (2005)Effects of biodiversity on ecosystem functioning: a consen-sus of current knowledge. Ecol Monogr 75:3–35
Johnson D, Ijdo M, Genney DR, Anderson IC, Alexander IJ(2005) How do plants regulate the function, communitystructure, and diversity of mycorrhizal fungi? J Exp Bot 56:1751–1760
Johnson SL, Kuske CL, Carney TD, Housman DC, Gallegos-Graves L, Belnap J (2012) Increased temperature and alteredsummer precipitation have differential effects on biologicalsoil crusts in a dryland ecosystem. Global Change Biol 18:2583–2593
Kardol P, Bezemer TM, van der Putten WH (2006) Temporalvariation in plant–soil feedback controls succession. EcolLett 9:1080–1088
Kefeli VI, Kalevitch MV, Borsari B (2013) Phenolic cycle inplants and environment. J Cell Mol Biol 2:13–18
Lange OL, Kidron G, Büdel B, Meyer A, Kilian E, Abeliovich A(1992) Taxonomic composition and photosynthetic char-acteristics of the “biological soil crusts” covering sanddunes in the western Negev Desert. Funct Ecol 6:519–527
Lawrey JD (1995) The chemical ecology of lichen mycoparasites:a review. Can J Bot 73:603–608
Loreau M, Naeem S, Inchausti O, Bengtsson J, Grime JP, HectorA, Hooper DU, Huston MA, Raffaelli V, Schmid B, TilmanD, Wardle DA (2001) Biodiversity and ecosystem function-ing: current knowledge and future challenges. Science 294:804–808
McHugh TA, Gehring CA (2006) Below-ground interactions witharbuscular mycorrhizal shrubs decrease the performance ofpinyon pine and the abundance of its ectomycorrhizas. NewPhytol 171:171–178
Maestre FT, Castillo-Monroy AP, Bowker MA, Ochoa-Hueso R(2012a) Species r ichness effects on ecosystemmultifunctionality depend on evenness, composition and spa-tial pattern. J Ecol 100:317–330
Maestre FT, Martín N, Díez B, López-Poma R, Santos F, Luque I,Cortina J (2006) Watering, fertilization, and slurry inocula-tion promote recovery of biological crust function in degrad-ed soils. Microb Ecol 52:365–377
Maestre FT, Salguero-Gómez R, Quero JL (2012b) It’s gettinghotter in here: determining and projecting the impacts ofglobal change on drylands. Philos Trans R Soc B 367:3062–3075
Maestre FT, Escolar C, Martínez I, Escudero A (2008) Are soillichen communities structured by biotic interactions? A nullmodel analysis. J Veg Sci 19:261–266
Maestre FT, Escudero A, Martinez I, Guerrero C, Rubio A (2005)Does spatial pattern matter to ecosystem functioning?Insights from biological soil crusts. Funct Ecol 19:566–573
Nash TH (1996) Lichen biology. Cambridge University Press,Cambridge
Neher DA, Lewins SA, Weicht TR, Darby BJ (2009)Microarthropod communities associated with biological soilcrusts in the Colorado Plateau and Chihuahuan deserts. J AridEnviron 73:672–677
Oren A, Steinberger Y (2008a) Catabolic profiles of soil fungalcommunities along a geographic climatic gradient in Israel.Soil Biol Biochem 40:2578–2587
Oren A, Steinberger Y (2008b) Coping with artifacts inducedby CaCO3–CO2–H2O equilibria in substrate utilizationprofiling of calcareous soils. Soil Biol Biochem 40:2569–2577
Øvreås L (2000) Population and community level approaches foranalyzing microbial diversity in natural environments a re-view. Ecol Lett 3:236–251
Petersen U, Wrage N, Köhle L, Leuschner C, Isselstein J (2012)Manipulating the species composition of permanent grass-lands - A new approach to biodiversity experiments. BasicAppl Ecol 13:1–9
Pringle RM, Doak DF, Brody AK, Jocqué R, Palmer TM (2010)Spatial pattern enhances ecosystem functioning in an Africansavanna. PLoS Biol 8:e1000377
Setälä H, Berg MP, Jones TH (2005) Trophic structure and func-tional redundancy in soil communities. In: Bardgett RD,UsherMB, Hopkind DW (eds) Biological diversity and function insoil. Cambridge University Press, Cambridge, pp 236–249
Schwintzer CR, Tjepkema JD (2001) Effect of elevated carbondioxide in the root atmosphere on nitrogenase activity inthree actinorhizal plants. Can J Bot 79:1010–1018
Shipley B (2000) Cause and correlation in Biology. CambridgeUniversity Press, UK
Steven B, Gallegos-Graves L, Belnap J, Kuske C (2013) Drylandsoil microbial communities display spatial biogeographicpatterns associated with soil depth and soil parent material.FEMS Microbiol Ecol 86:101–113
Stevenson FJ (1993) Hummus chemistry: genesis, compositionreactions. Wiley, New York
Tjepkema JD, Schwintzer CR, Burris RH, Johnson GV, SilvesterWB (2000) Natural abundance of 15N in actinorhizal plantsand nodules. Plant Soil 219:285–289
Van der Heijden MGA, Klironomos JN, Ursic M, Moutoglis P,Streitwolf-Engel R, Boller T, Wiemken A, Sanders IR (1998)Mycorrhizal fungal diversity determines plant diodiversity,ecosystem variability and productivity. Nature 396:69–72
Plant Soil (2015) 386:303–316 315

Vivanco L, Austin AT (2008) Tree species identity alters forestlitter decomposition through long-term plant and soil inter-actions in Patagonia, Argentina. J Ecol 96:l727–l736
Wardle DA,Bardgett RD,Klironomos JN, Setälä H, van der PuttenWH, Wall DH (2004) Ecological linkages between above-ground and belowground biota. Science 304:1629–1633
Wilsey BJ, Polley HW (2004) Realistically low species evennessdoes not alter grassland species-richness-productivity rela-tionship. Ecology 85:2693–2700
Wright RT (1984) Dynamic pools of dissolved organic carbon. In:Hobbie JE, Williams PJ (Eds) Heterotrophic activity in thesea. pp. 121–154. Plenum
Yeager CM, Kornosky JL, Housman DC, Grote EE, Belnap J,Kuske CR (2004) Diazotrophic community structure andfunction in two successional stages of biological soil crustsfrom the Colorado Plateau and Chihuahuan Desert. ApplEnviron Microbiol 70:973–983
Yu J, Kidron GJ, Pen-Mouratov S, Wasserstrom H, Barness G(2012) Do development stages of biological soil crusts deter-mine activity and functional diversity in a sand-dune ecosys-tem? Soil Biol Biochem 51:66–72
Zak JC, Willig MR, Moorhead DL, Wildman HG (1994)Functional diversity of microbial communities: a quantitativeapproach. Soil Biol Biochem 26:1101–1108
316 Plant Soil (2015) 386:303–316
Related Documents











