T Cell Activation Jennifer E. Smith-Garvin, 1 Gary A. Koretzky, 1,2,3 and Martha S. Jordan 1,2 1 Abramson Family Cancer Research Institute, 2 Department of Pathology and Laboratory Medicine, 3 Department of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania 19104; email: [email protected], [email protected], [email protected] Annu. Rev. Immunol. 2009. 27:591–619 First published online as a Review in Advance on January 8, 2009 The Annual Review of Immunology is online at immunol.annualreviews.org This article’s doi: 10.1146/annurev.immunol.021908.132706 Copyright c 2009 by Annual Reviews. All rights reserved 0732-0582/09/0423-0591$20.00 Key Words signal transduction, immunoreceptor, integrin Abstract This year marks the 25th anniversary of the first Annual Review of Immunology article to describe features of the T cell antigen receptor (TCR). In celebration of this anniversary, we begin with a brief intro- duction outlining the chronology of the earliest studies that established the basic paradigm for how the engaged TCR transduces its signals. This review continues with a description of the current state of our un- derstanding of TCR signaling, as well as a summary of recent findings examining other key aspects of T cell activation, including cross talk between the TCR and integrins, the role of costimulatory molecules, and how signals may negatively regulate T cell function. 591 Annu. Rev. Immunol. 2009.27:591-619. Downloaded from www.annualreviews.org by Universidade Federal de Uberlandia on 04/26/12. For personal use only.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ANRV371-IY27-21 ARI 16 February 2009 13:31
T Cell ActivationJennifer E. Smith-Garvin,1 Gary A. Koretzky,1,2,3
and Martha S. Jordan1,2
1Abramson Family Cancer Research Institute, 2Department of Pathology and LaboratoryMedicine, 3Department of Medicine, University of Pennsylvania, Philadelphia,Pennsylvania 19104; email: [email protected], [email protected],[email protected]
Annu. Rev. Immunol. 2009. 27:591–619
First published online as a Review in Advance onJanuary 8, 2009
The Annual Review of Immunology is online atimmunol.annualreviews.org
This article’s doi:10.1146/annurev.immunol.021908.132706
Copyright c© 2009 by Annual Reviews.All rights reserved
0732-0582/09/0423-0591$20.00
Key Words
signal transduction, immunoreceptor, integrin
AbstractThis year marks the 25th anniversary of the first Annual Review ofImmunology article to describe features of the T cell antigen receptor(TCR). In celebration of this anniversary, we begin with a brief intro-duction outlining the chronology of the earliest studies that establishedthe basic paradigm for how the engaged TCR transduces its signals.This review continues with a description of the current state of our un-derstanding of TCR signaling, as well as a summary of recent findingsexamining other key aspects of T cell activation, including cross talkbetween the TCR and integrins, the role of costimulatory molecules,and how signals may negatively regulate T cell function.
591
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
TCR: T cell antigenreceptor
Signal transduction:biochemical eventslinking surfacereceptor engagementto cellular responses
INTRODUCTION
Twenty-five years ago, Annual Review of Im-munology published its first review describingfeatures of the structure we now know as theT cell antigen receptor (TCR) (1). In recog-nition of this anniversary, this article begins byhighlighting a sampling of seminal observationsmade in the decade following the initial descrip-tion of the TCR. The discoveries made dur-ing this period established the basic paradigmfor how TCR engagement initiates the earli-est biochemical events leading to cellular ac-tivation, described nicely in an Annual Reviewof Immunology article in 1996 (2) (Figure 1a).This historical perspective sets the stage for adiscussion of our current state of understandingof the molecular and biochemical events criti-cal for T cell activation that have emerged fromthe work of multiple laboratories.
DESCRIBING THETCR COMPLEX
In the early 1980s, several groups began ex-periments to identify and characterize the anti-gen receptor on T cells. One approach usednewly developed molecular techniques, ulti-mately leading to the identification of the genesresponsible for the antigen-binding proteins(3–5). An even earlier approach relied on im-
−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−→Figure 1TCR proximal signaling then and now: filling in the gaps. Then (a): TCR signaling mid-1990s (adapted from Reference 2). Ligation ofthe TCR/CD3 results in activation of Src and Syk family PTKs associated with the intracellular CD3 domains that then activatePLCγ1 and Ras-dependent pathways. PLCγ1 hydrolizes PIP2 to form IP3 and DAG. Now (b–d ): Current understanding of how theTCR couples to downstream pathways (b), the molecular basis for Ca2+ influx (c), and the positive feedback loop responsible for Rasactivation (d ). (b) The link between PTKs and downstream pathways is a multimolecular signaling complex nucleated by the adapterproteins LAT, Gads, and SLP-76. Lck activates ZAP-70 to phosphorylate (p) tyrosine residues on LAT, which then recruits Gads andits constitutive binding partner SLP-76. ZAP-70-mediated phosphorylation of SLP-76 results in the recruitment of multiple SH2domain–containing effector molecules (circles) and adapter proteins (octogons). SH3 domains (shaded overlapping areas) also link effectorsto adapters and contribute to stabilization of the complex. (c) The link between depletion of ER Ca2+ stores and CRAC channelactivation is STIM. TCR-induced IP3 production (see b) results in the activation of IP3 receptors (IP3R), which release Ca2+ from theER into the cytoplasm. STIM contains paired EF hands within the ER lumen, which bind a single Ca2+ molecule. Upon depletion ofER stores, Ca2+-free STIM molecules oligomerize and move to areas of ER/plasma membrane proximity, where they colocalize withand induce dimerization of Orai1 dimers, resulting in a functional CRAC channel and subsequent influx of Ca2+. (d ) Activation of Rasinvolves two Ras GEFs (SOS and RasGRP) in a positive feedback loop. TCR-induced production of DAG (see b) results in themembrane recruitment of RasGRP, where it is phosphorylated and activated by PKCθ. RasGRP then facilitates the removal of GDPfrom Ras, which can then bind GTP and become activated. GTP-bound Ras then binds SOS, which is bound constitutively to GRB2and is inducibly recruited to LAT, increasing its GEF activity, resulting in a positive feedback loop and robust Ras activity.
munization of mice with T cell clones of a de-fined specificity, hybridomas, or clonal T celltumors in the hope that antibodies would begenerated that would react with the receptorresponsible for binding to antigen (6–8). Suchantibodies were produced and were then usedfirst to demonstrate interference with antigenicresponses and later to perform the initial bio-chemical characterization of the receptor it-self. These studies revealed a complicated cellsurface structure that included proteins reac-tive with antibodies against the nonpolymor-phic CD3 proteins (initially thought of as threepolypeptides, γ, δ, and ε) (9), as well as variableproteins designated α and β.
The antigen-binding function of αβ wasobvious early on, by inference owing to theirhighly polymorphic nature and similarity toimmunoglobulin and experimentally owing toearly gene transfer experiments that demon-strated that antigen/major histocompatibilitycomplex (MHC) reactivity tracked with expres-sion of these receptor components (10, 11).However, it was less obvious what role the CD3proteins played in TCR function. Several linesof evidence suggested that the CD3 moleculeswere critical for signal transduction: Unlike αβ,the CD3 molecules had long cytoplasmic tails,and anti-CD3 antibodies resulted in T cell ac-tivation, although it was difficult to demon-strate signaling function conclusively. One early
592 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
www.annualreviews.org • T Cell Activation 593
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
PKC: protein kinaseC
PLC: phospholipaseC
PTK: proteintyrosine kinase
Immunoreceptortyrosine-basedactivation motif(ITAM): a shortpeptide sequence inthe cytoplasmic tails ofkey surface receptorson hematopoietic cellsthat is characterized bytyrosine residues thatare phosphorylated bySrc family PTKs,enabling the ITAM torecruit activated Sykfamily kinases
effort involved an attempt to create cell linesexpressing αβ without CD3 or CD3 withoutαβ, thereby creating reagents to investigate theindependent role of the receptor components.These experiments were not successful, as itbecame clear that there is an obligatory coex-pression of αβ with CD3 (12). These studiesdid result, however, in the creation of the firstin a long line of genetically altered Jurkat Tcells that have been instrumental in our under-standing of how the TCR complex couples to itssignaling machinery (13).
EARLY SIGNALING STUDIES
Initial investigations into how the TCR trans-duces its signals began with the observation thatTCR-deficient Jurkat T cells could be stimu-lated pharmacologically with the combinationof phorbol esters and Ca2+ ionophores (14).This observation led to speculation that en-gagement of the TCR might stimulate the samesignals that these reagents induced. This no-tion was tested directly in experiments demon-strating increases in intracellular free Ca2+
following CD3 or TCR stimulation in bothJurkat cells and primary T cells. The sourceof the Ca2+ increase was shown to be a com-bination of Ca2+ released from an intracellularpool in response to inositol trisphosphate (IP3)production and influx of Ca2+ from outside ofthe cell (15). It was presumed that phorbol es-ters were important because of their ability toactivate protein kinase C (PKC, now known tobe a family of enzymes), whose activity is regu-lated physiologically by diacylglycerol (DAG).Concurrent studies in other systems haddemonstrated that a single enzymatic reaction,hydrolysis of the membrane PI(4,5)P2 (phos-pholipid phosphatidylinositol 4,5 bisphos-phate) by phospholipase C (PLC), generatesboth IP3 and DAG, suggesting that the TCRmay function to regulate PLC activity.
Testing this notion proved somewhatdifficult at first, as PLC regulation was initiallydescribed downstream of heterotrimericguanosine triphosphate (GTP)-binding pro-teins associated with seven-transmembrane
domain receptors. An exhaustive but unsuc-cessful search ensued for the GTP-bindingprotein critical for coupling the TCR toPLC activation. Insight into how the TCRmay initiate PLC activation emerged from aconfluence of discoveries in immune cells andother lineages. For example, stimulation of theTCR results in changes in protein phospho-rylation, including inducible phosphorylationof the newly described ζ chain of the CD3complex (16). Furthermore, growth factorreceptors with intrinsic protein tyrosine kinase(PTK) activity also stimulate PLC function.In contrast to seven-transmembrane domainreceptors, however, receptor PTKs stimulateanother PLC isoform, PLCγ. Although noneof the TCR components possessed enzymaticactivity themselves, cytosolic PTKs of theSrc family (in particular Lck and Fyn) werebeing described in T cells. These PTKs wereassociated either with the TCR (17) or withthe CD4 and CD8 coreceptors (18, 19),both important for TCR signaling. Theseobservations, coupled with the new knowledgethat T cell activation requires PTK function(20), led to a new TCR signaling paradigmin which the TCR recruits cytosolic PTKs toactivate key second messengers.
HOW CD3 TRANSDUCESITS SIGNALS
Testing this model led to a search for addi-tional substrates of the TCR-stimulated PTKs.Among the most attractive candidates were ty-rosines within the CD3 molecules themselvesthat fell within a motif present once withinCD3 ε, γ, and δ and in triplicate within each ζ
chain and also in key immunoreceptors on otherimmune cell lineages. The signature ofthese motifs (21), eventually designated im-munoreceptor tyrosine-based activation motifs(ITAMs), is two tyrosines flanking a series ofamino acids, including key leucine/isoleucineswith stereotypic spacing. Numerous labora-tories demonstrated that the ITAM tyrosinesare in fact phosphorylated upon TCR lig-ation, but defining the importance of this
594 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
posttranslational modification for TCR func-tion was not trivial. Because the CD3 moleculescould not be expressed without the αβ chains,several groups created chimeric molecules fus-ing the cytoplasmic domains of individual CD3chains to extracellular and transmembrane do-mains from other proteins (22–25). ThesecDNAs in chimeric proteins were then trans-fected into T cell lines selected for TCR loss.Antibody crosslinking of the extracellular do-main of the chimeras recapitulated all theknown TCR-mediated signaling events lead-ing to cellular activation. Mutation of the keyITAM tyrosines (or altering their spacing) abro-gated the ability of the chimeras to activate thecells, thus demonstrating that tyrosine phos-phorylation of the CD3 ITAMs is an earlyand requisite step for TCR-mediated T cellactivation.
Studies of the CD3 chimeras led naturally tothe question of the purpose for ITAM phospho-rylation. Unlike growth factor receptors whoseintrinsic enzymatic activity is enhanced by tyro-sine phosphorylation, the CD3 molecules haveno such effector function on their own. Investi-gators speculated, therefore, that tyrosine phos-phorylation of the ITAMs might serve as dock-ing sites for interactions with other proteins.Indeed, it was soon shown that phosphory-lated CD3ζ (and later other ITAM-containingproteins) is a recruitment site of a 70-kDaphosphoprotein, the Syk kinase family memberZAP-70 (ζ-associated protein of 70 kDa) PTK(26). A model therefore emerged that engage-ment of the TCR leads to Src family PTK ac-tivity resulting in ITAM phosphorylation andrecruitment of ZAP-70. This converted theTCR with no intrinsic enzymatic function to anactive PTK able to phosphorylate a spectrumof substrates leading to a myriad of down-stream signals that, when integrated appropri-ately (along with signals from other corecep-tors), lead to T cell activation (27).
The basic tenets of this model have stood thetest of intensive investigation. In the 15 yearssince ZAP-70 was cloned, investigators havefilled in many of the gaps between the TCRand initiation of effector functions. Much has
ARGUING BY ANALOGY: COMPARINGAND CONTRASTING SIGNALING PATHWAYSBY MULTIPLE RECEPTORS AND LINEAGES
Key insights important for understanding T cell activation havecome from studies in nonimmune mammalian cells and cells oflower organisms, including the observation that PTKs could linkto the phosphatidylinositol pathway and the paradigm describingadapter proteins as integrators of signal transduction. T cell biol-ogists have also provided unique insights to their nonimmunol-ogist colleagues. Examples include a mechanism for recruitmentof nonreceptor PTKs to enzymatically inactive surface receptorsand the notion that protein tyrosine phosphatases may be positiveas well as negative regulators of PTK pathways.
Within the immune system, biologists studying TCR signal-ing have been the donors and recipients of information that hasbeen useful for investigators examining the signaling pathways ofother cell surface receptors (e.g., integrins) and other hematopoi-etic lineages. It is clear that, although basic paradigms may besimilar, each cell type and receptor utilizes unique ways to regu-late signal transduction cascades. Additionally, recent studies havedemonstrated that pathways thought to be distinct (e.g., integrinsand immunoreceptors) instead intersect at multiple levels. Fu-ture insights into how diverse signaling pathways are integratedto result in the appropriate biologic response will undoubtedlycontinue to benefit from comparing and contrasting activationevents downstream of multiple receptors in different immuneand nonimmune cell lineages.
Adapter protein:cellular protein thatfunctions to bridgemolecular interactionsvia characteristicdomains able tomediateprotein/protein orprotein/lipidinteractions
been learned about substrates of the PTKs (in-cluding Src, Syk, and more recently Tec fam-ily members) activated by the TCR and howthese molecules participate in T cell activation,about how signaling complexes are organizedby adapter proteins to bring effector proteinstogether, and about the unexpected intersec-tion of particular signaling pathways (see alsothe side bar, Arguing by Analogy: Comparingand Contrasting Signaling Pathways by Mul-tiple Receptors and Lineages). With the ac-cumulation of data, it has also become clearthat signaling via the TCR complex is not alinear event starting at the receptor and end-ing in the nucleus. Instead, there appears to becomplex feedback and feedforward regulationat each step. Ironically, one of the most central
www.annualreviews.org • T Cell Activation 595
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
pMHC: peptidemajorhistocompatibilitycomplex (MHC)complex
signaling questions that remains is how receptorbinding translates most proximally into an ac-tivating signal. Many models have been pro-posed, but none has yet withstood the rigorof subsequent investigation. This review sum-marizes our current understanding of many ofthese issues and poses some of the intriguingquestions that remain.
INITIATING TCR SIGNALTRANSDUCTION
Since identification of the TCR as a complexconsisting of the variable αβ chains nonco-valently associated with the nonpolymorphicCD3 proteins, considerable work has gone intodefining the stoichiometry of these interactingmolecules. We now know that the CD3 proteinsexist as a series of dimers including γε, δε, andζζ associated with a single αβ heterodimer. Al-though it has been clear for more than 15 yearsthat CD3 transduces signals from the engagedreceptor via its ITAMs, exactly how ligationof the TCR is translated into the first signalremains controversial. Current models suggestthat both TCR aggregation and conformationalchanges may play roles in signal initiation.
Two separate but not mutually exclusiveconformational changes within the CD3 cy-toplasmic tails have been proposed as mech-anisms for TCR-inducible ITAM phosphory-lation. The first mechanism is based on theobservation that, in the presence of acidiclipid designed to mimic the inner leaflet ofthe plasma membrane, the cytoplasmic tails ofCD3ε, and to a lesser extent of CD3ζ, fold orinteract with the lipid in such a way as to pre-vent phosphorylation. Conversely, in aqueoussolution or in the presence of zwitterionic lipid,the CD3 tails are more readily able to be phos-phorylated by Lck (28, 29). These findings haveled to the hypothesis that in resting T cells theCD3ζ and/or CD3ε tails are tightly associatedwith the lipid-rich inner leaflet of the plasmamembrane, rendering them inaccessible to Lckphosphorylation, but following TCR ligation,they are released from the membrane and phos-phorylated. Investigations have begun to test
this hypothesis in T cells, and recently the in-teraction of the CD3ε cytoplasmic domain withthe plasma membrane has been confirmed inresting Jurkat T cells by fluorescence resonanceenergy transfer (FRET) (29). A second possibleconformational change in CD3ε has garneredmuch attention following the observation thatupon TCR ligation a proline-rich region (PRR)in CD3ε is exposed and available to recruit anSH3 domain of the adapter protein noncat-alytic tyrosine kinase (Nck). Importantly, thisoccurs prior to CD3ζ and CD3ε ITAM phos-phorylation (30). Investigators speculated thatNck could then recruit and activate effectormolecules required for subsequent ITAM phos-phorylation. However, recent data using micewith retrogenic or knockin mutations of theCD3ε PRR have failed to show a requirementfor the CD3ε/Nck interaction in the activationof peripheral T cells (31, 32). This observationmust be considered in light of the T cell devel-opmental abnormalities in the knockin mice.Resolving the importance of the PRR for ma-ture T cell function requires temporal controlof expression of the mutant.
Although the seminal finding that there isa non-ITAM region in CD3 that is critical forTCR signaling appears firm, several controver-sies remain regarding the CD3ε PRR. Thereis disagreement regarding whether the inter-action between Nck and CD3ε is constitutiverather than inducible (31, 33–35). The role ofthe CD3ε/Nck interaction in mature T cells isalso controversial, as it has been suggested thatthe interaction (a) is not required for mature Tcell activation (32), (b) is required for signal am-plification and ITAM phosphorylation follow-ing weak TCR ligation (34), or (c) is required forregulation of TCR activity by inhibiting ITAMphosphorylation and promoting TCR degrada-tion (35). Ongoing work will provide insightsinto these controversies.
Although CD3 conformational changes mayexplain the initiation of the intracellular ki-nase cascade, we do not know how pep-tide/MHC (pMHC)/TCR interactions resultin conformational changes in the intracellu-lar regions of the CD3 complex. Models based
596 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
Table 1 Recent models for initial TCR triggering
Model DescriptionPiston-like movement An external force from the APC/T cell interaction pushes the TCR/CD3 complex in a piston-like
fashion perpendicular to the plasma membrane, exposing activating motifs on the intracellular regionsof CD3 chains
Receptor deformation The force of the motility of the T cell as it moves over an APC induces TCR triggering only whenpMHC/TCR interactions are of a high enough affinity to withstand the force long enough to undergoa conformational change
Permissive geometry pMHC dimers bind TCR/CD3 dimers inducing a rotational scissor-like conformational change in CD3chain(s) that reveal previously hidden intracellular activation motif(s)
Kinetic segregation Signal initiation occurs because of exclusion of inhibitory molecules in the tight contact zone betweenthe T cell and the APC, thereby shifting the enzymatic steady state toward an activating state
Diffusion trapping Expanded kinetic segregation model in which the affinity/diffusion coefficient of the pMHC/TCRdictates the valency of TCRs required to trap the complex in the tight contact zone and thereforeinitiate a productive signal
Pseudodimer Agonist pMHC/TCR recruits a second TCR via interaction with its associated CD4, the second TCRthen binds a coagonist endogenous pMHC forming a stable pseudodimer that triggers signaling viaproximity of ITAMs to CD4-associated Lck and/or by conformational change
on ligand-induced conformational changes inthe TCR and/or TCR aggregation have beenproposed (Table 1). The crystal structuresof several pMHC/TCR complexes have re-vealed little conformational changes in theTCR heterodimer outside of the binding do-main (reviewed in 36). However, owing to thecurrent technical limitations of crystallogra-phy and protein nuclear magnetic resonancespectroscopy, only pMHC/TCR structures inthe absence of transmembrane domains andCD3 components have been elucidated. There-fore, changes between the components of theTCR/CD3 complex in relation to one anotheror to the plasma membrane cannot be ruled out.The rigid, rod-like transmembrane domain ofthe CD3εγ could mediate a piston-like move-ment of the TCR complex perpendicular tothe plasma membrane after ligand binding, ina manner similar to that reported for the as-partate receptor (37). Such a model would re-quire external forces that could be provided bycell-to-cell contact, but this possibility is dif-ficult to reconcile with the ability of solubleligands to activate the TCR. Similarly, an ex-ternal force is required for the recently pro-posed receptor deformation model, in whichthe movement of the T cell as it travels acrossthe antigen-presenting cell (APC) exerts a force
on transient pMHC/TCR interactions. Onlywhen the force required to induce conforma-tional changes in the TCR is less than the forcerequired to break the pMHC/TCR interactionwill TCR triggering occur (38). Alternatively,the permissive geometry model for TCR trig-gering predicts that dimeric pMHCs bind twoTCR/CD3 complexes (39). This interaction re-sults in a rotation of the TCR heterodimersaround one another, displacing the extracellulardomains of the associated CD3 molecules. Thetransmembrane interactions between the CD3and TCR dimers serve as pivot points, and theCD3 movement is scissor-like, exposing previ-ously shielded ITAMs and/or other functionallyimportant domains. This model is consistentwith studies that suggest that TCR aggregationis required for TCR triggering.
TCR aggregation has long been impli-cated as a mechanism for TCR triggering. Inthe classic aggregation model, crosslinking ofTCR/CD3 complexes with multimeric pMHCenables close contact and transphosphorylationbetween the CD3 tails and associated PTKs.Clustering of several TCR complexes also mayresult in competition for membrane lipid be-tween the CD3 chains, resulting in dissocia-tion of the cytoplasmic domains from the mem-brane and subsequent ITAM phosphorylation
www.annualreviews.org • T Cell Activation 597
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
PI3K:phosphoinositide3-kinase
(28). Aggregation is supported strongly byobservations that soluble multimeric but notmonomeric pMHC can trigger TCR activa-tion. However, recent biochemical and micro-scopic studies suggest that preformed TCRaggregates are present on nonactivated T cells(reviewed in 40). In the kinetic segregationmodel, adhesion and other accessory moleculeswith short extracellular domains such as CD2initiate close contact zones between an APCand a T cell (41). Inhibitory phosphatases withlong extracellular domains such as CD45 areexcluded because of their size. TCR complexesthat engage pMHC on the APC surface remainin the close contact zone, where they are seg-regated from phosphatases and are able to ini-tiate signaling. TCR complexes that do not en-gage pMHC are free to diffuse outside of theclose contact zone. Shortening the extracellulardomains of inhibitory molecules or lengthen-ing the extracellular domains of adhesion or ac-cessory molecules can abrogate TCR signaling,suggesting that kinetic segregation is an impor-tant aspect of TCR signal initiation (reviewedin 41). This notion was expanded with the dif-fusion trapping model in which immobilizationor trapping of pMHC/TCRs in the close con-tact zones is responsible for TCR signal initi-ation (42). The valency, or degree of aggrega-tion, of the pMHC/TCR required for trappingand therefore triggering depends on the affin-ity and diffusion coefficient of the pMHC/TCRinteraction. Another model has been proposedthat suggests that endogenous pMHCs amplifysignals produced by the rare agonist pMHCsby promoting TCR aggregation and a subse-quent phosphorylation cascade (40). Similarly,the pseudodimer model proposes that an ago-nist pMHC/TCR can recruit a second TCR in aCD4-dependent manner, and this second TCRbinds a coagonist endogenous pMHC, forminga stable pseudodimer that could trigger signal-ing through ITAM/Lck proximity or throughCD3 conformational changes (43). Many ofthese models are not mutually exclusive, and itis likely that TCR aggregation, conformationalchanges within the TCR complex, and exclu-sion of inhibitory molecules are all required
for TCR triggering, perhaps in a stepwisefashion.
PROXIMAL SIGNALINGCOMPLEX
The earliest step in intracellular signaling fol-lowing TCR ligation is the activation of Src(Lck and Fyn) PTKs, leading to phosphoryla-tion of the CD3 ITAMs. Recruitment of ZAP-70 follows, leading to a cascade of phospho-rylation events. The past decade has seen thedescription of a subcellular assembly and acti-vation of an adapter protein nucleated multi-molecular signaling complex (Figure 1b). Thiscomplex is responsible for propagating theTCR/PTK signal into multiple and diverse dis-tal signaling pathways.
Among the most important of the ZAP-70targets are the transmembrane adapter proteinlinker for the activation of T cells (LAT) andthe cytosolic adapter protein Src homology 2(SH2) domain–containing leukocyte phospho-protein of 76 kDa (SLP-76) (44, 45). Thesetwo adapters form the backbone of the com-plex that organizes effector molecules in thecorrect spatiotemporal manner to allow for theactivation of multiple signaling pathways. Theimportance of these adapters is underscored bystudies showing that the loss of either LAT orSLP-76 results in a near complete loss of TCRsignal transduction reminiscent of Syk/ZAP-70or Lck/Fyn double-deficient T cells (46–48).
LAT contains nine tyrosines that are phos-phorylated upon TCR engagement, which bindthe C-terminal SH2 domain of PLCγ1, the p85subunit of phosphoinositide 3-kinase (PI3K),and the adapters growth factor receptor-boundprotein 2 (GRB2) and GRB2-related adapterdownstream of Shc (Gads) (reviewed in 47).SLP-76 is then recruited to phosphorylatedLAT via their mutual binding partner Gads(49). SLP-76 itself contains three modulardomains: an N-terminal acidic domain withthree phosphorylatable tyrosines that interactwith the SH2 domains of Vav1, Nck, and IL-2-induced tyrosine kinase (Itk); a PRR thatbinds constitutively Gads and PLCγ1; and a
598 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
C-terminal SH2 region that can bind adhesionand degranulation–promoting adapter protein(ADAP) and hematopoietic progenitor kinase1 (HPK1) (reviewed in 46). Although LAT andSLP-76 serve to nucleate this large signalingcomplex, the effector molecules themselves alsoare important for stabilizing the complex. Forexample, the Tec family kinase Itk is requiredin a kinase-independent manner for the re-cruitment of the guanine nucleotide exchangefactor (GEF) Vav1 to the APC contact site,whereas Vav1 is required for optimal SLP-76phosphorylation and recruitment to LAT aswell as for Itk activation (50–52). These andother data suggest that the formation of thecomplex is more complicated than the linearmodel most often invoked for simplicity. For ex-ample, PLCγ1 directly binds to SLP-76, LAT,and Vav1 as well as to its activating kinaseItk (reviewed in 53). It is thought that theseinteractions collectively are required to stabi-lize PLCγ1 in the correct conformation withinthe complex to allow for its optimal activity(54). Advances in biochemical and structuraltechniques are needed to elucidate the preciseallosteric and perhaps stoichiometric changeswithin the multimolecular complex that allowfor signal transduction.
To investigate more precisely the impor-tance of these complex interactions in primaryT cells, several laboratories have generatedmice expressing transgenic or knockin muta-tions in specific binding regions in variousmolecules involved in proximal signaling (55–57). Tyrosine to phenylalanine mutations inSLP-76 at residues 112 and 128 or 145 in pri-mary thymocytes and T cells do not result ina loss of SLP-76/Vav1/Nck/Itk interactions, aswould be expected from earlier phosphopeptidemapping studies and studies in cell lines (57).However, these tyrosine mutations still resultin severe defects in downstream signaling path-ways consistent with defective Vav1 or Itk activ-ity. Similarly, mutation of tyrosines of Vav1 doesnot result in a loss of interaction with their pro-posed binding partners, although it does resultin dysregulated Vav1-dependent signaling (55).Although the continued interactions of these
proteins seen by immunoprecipitation exper-iments are likely due to tertiary interactionswith other domains or other molecules, thesestudies suggest that SH2/phosphotyrosine in-teractions may play important regulatory rolesfor the activation of effector molecules. Indeed,structural studies have suggested that the in-teraction between the SH2 domain of Itk anda phosphotyrosine results in a conformationalswitch allowing kinase activity (58). Consistentwith these data, investigators recently showedin Jurkat T cell lines that an Itk/SLP-76 in-teraction is required for Itk kinase activity, al-though we do not yet know if it is specificallythe SH2/phosphotyrosine interaction that me-diates this kinase activity (59). Further stud-ies are required to determine how the activ-ities of molecules beyond Itk are affected byspecific domain/domain interactions within thecomplex.
The proximal signaling complex resultsin the activation of PLCγ1-dependent path-ways including Ca2+- and DAG-inducedresponses, cytoskeletal rearrangements, andintegrin activation pathways. Ligation of co-stimulatory receptors such as CD28 augmentsthese pathways. Below, we discuss the mecha-nisms by which these pathways are activated andregulated.
PLCγ1 ACTIVATIONAND SIGNAL TRANSDUCTION
Following TCR ligation, PLCγ1 is found inthe proximal signaling complex bound to SLP-76, Vav1, and LAT, where it is phosphorylatedand activated by Itk. Activated PLCγ1 then hy-drolyzes the membrane lipid PI(4,5)P2, produc-ing the second messengers IP3 and DAG. Thesetwo messengers are essential for T cell function,and therefore the regulation of PLCγ1 acti-vation has been the subject of intensive stud-ies. Localization of PLCγ1 to the proximalsignaling complex is dependent on LAT andthe Gads-binding region of SLP-76 (54). Ac-tivation of PLCγ1 is dependent on Itk kinaseactivity that, in turn, is dependent on Vav1,Lck, ZAP-70, LAT, and SLP-76 (52, 60, 61).
www.annualreviews.org • T Cell Activation 599
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
Following TCR ligation, Itk is recruited to themembrane through PH domain interactionswith PIP3, which has been locally generatedby Lck-induced PI3K activity (reviewed in 61).At the membrane, Lck phosphorylates Itk, andthe SH2 and SH3 domains of Itk interact withphosphorylated tyrosine 145 and the PRR ofSLP-76, respectively (62–64). The role of Vav1and ZAP-70 in Itk activation is not understoodbut may relate to their involvement in the phos-phorylation of SLP-76 or PI3K activation (45,51, 52). A second Tec family kinase, Rlk, can alsophosphorylate PLCγ1, resulting in a relativelymild defect in Itk-deficient mice and requiringthe study of Rlk/Itk double-deficient mice tobetter understand the role of Tec kinases in Tcell activation (reviewed in 61).
DAG-MEDIATED SIGNALINGPATHWAYS
TCR-induced production of DAG results inthe activation of two major pathways involvingRas and PKCθ. Ras is a guanine nucleotide–binding protein and is required for the acti-vation of the serine-threonine kinase Raf-1,which initiates a mitogen-associated proteinkinase (MAPK) phosphorylation and activa-tion cascade. Raf-1 is a MAPK kinase kinase(MAPKKK) that phosphorylates and activatesMAPK kinases (MAPKKs), which in turn phos-phorylate and activate the MAPK’s extracellularsignal-regulated kinase 1 (Erk1) and Erk2. Erkkinase activity results in the activation of thetranscription factor Elk1, which contributes tothe activation of the activator protein-1 (AP-1)( Jun/Fos) transcription complex via regulationof Fos expression. Additionally, Erk activity canresult in the transcriptional activation of sig-nal transducer and activator of transcription 3(STAT3) and in the serine phosphorylation ofLck (reviewed in 65).
Ras is only active in the GTP-boundstate, and its activation is facilitated by GEFsand is suppressed by GTPase-activating pro-teins (GAPs). Two Ras GEFs are presentin T cells, son of sevenless (SOS) and Rasguanyl nucleotide-releasing protein (RasGRP).
RasGRP appears more dominant for early ac-tivation of Ras, as SOS cannot compensatefor RasGRP deficiency (66–68). RasGRP is in-ducibly recruited to the membrane through aDAG-binding domain (69), where it is phos-phorylated by PKCθ (70). SOS is constitu-tively bound to the adapter protein GRB2,and upon TCR stimulation, the GRB2 SH2domain is recruited to and binds phospho-rylated tyrosines on LAT, thereby bringingSOS into the proximal signaling complex,where it can facilitate the localized activa-tion of Ras (71). The significance of thesetwo modes of Ras activation was unclear un-til recently, when it was shown that RasGRP-dependent RasGTP production catalyzes SOSactivity, resulting in a positive feedback loopand robust TCR-induced Ras activation (72)(Figure 1c). This unified model explains theRasGRP dominance, as RasGRP would be re-quired to initiate the production of RasGTP,which upon reaching threshold levels wouldcatalyze SOS activity and further amplify thesignal.
The second major signaling pathway regu-lated by DAG is mediated by PKCθ, a PKCfamily member that contains a lipid-bindingdomain specific for DAG, which is importantfor recruiting PKCθ to the plasma membranefollowing T cell activation. However, phospho-rylation of PKCθ by Lck (reviewed in 73) maybe required to induce a conformational changethat enables binding to the lipid phosphatidylserine (PS), which in turn enhances binding toDAG, resulting in PKCθ activation (74). Otherproximal signaling molecules, including Vav1,PI3K, and 3-phosphoinositide-dependent ki-nase 1 (PDK1), also play roles in PKCθ local-ization, but details of their contributions are notcompletely defined (73).
One critical pathway that PKCθ regulatesis NF-κB activation. Because both the canoni-cal and noncanonical activation of this pathwayin T cells is described in detail in this volumeof Annual Review of Immunology [see review byKarin and colleagues (75)], we highlight onlythe major steps in the classical activation ofNF-κB downstream of the TCR. The NF-κB
600 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
family of transcription factors consists of fivemembers. In resting cells, NF-κB is found inthe cytosol associated with inhibitor of NF-κB(IκB) family members that keep NF-κB frommoving into the nucleus. Upon T cell activa-tion, IκB is phosphorylated by the IκB kinase(IKK) complex, ubiquitinylated, and degraded,allowing NF-κB to translocate into the nucleus,where it activates genes involved in the func-tion, survival, and homeostasis of T cells (re-viewed in 76). Although we have known thegeneral pathway of NF-κB activation for sometime, the specifics of how PKCθ activation leadsto nuclear import of NF-κB in T cells are stillbeing elucidated.
Over the past several years, the identifi-cation and characterization of a lymphocyte-specific activation complex have provided someinsight into this question. Following TCRstimulation, a trimolecular complex forms be-tween CARMA1 [caspase recruitment domain(CARD) and membrane-associated guanylatekinase (MAGUK)-containing scaffold pro-tein], the CARD-containing adapter proteinBcl10, and mucosa-associated lymphoid tis-sue lymphoma translocation gene 1 (MALT1)(73, 76). The assembly of this CBM com-plex (CARMA1/Bcl10/MALT1) is regulatedby PKCθ through its phosphorylation ofCARMA1, which is required for CARMA1oligomerization and association with Bcl10 (77,78). MALT1 binds to Bcl10 and contributesto the degradation of the regulatory subunitof the IKK complex (IKKγ) by facilitating itspolyubiquitination, possibly via activation ofthe E3 ubiquitin ligase tumor necrosis factorreceptor-associated factor 6 (TRAF6) (79, 80).Degradation of this regulatory subunit allowsfor phosphorylation of IκB by the IKK cat-alytic subunits, subsequent IκB degradation,and release of NF-κB, resulting in NF-κB nu-clear localization and gene activation. Recently,MALT1 was shown to enhance NF-κB signal-ing through its ability to degrade the deubiq-uitinating enzyme A20, a negative regulator ofNF-κB activation (81, 82).
One additional component of the complexthat was recently identified as a CARMA1-
associating protein is ADAP (83). ADAP,originally defined as a SLP-76-binding part-ner, associates with the MAGUK domain ofCARMA1 following T cell stimulation. Inves-tigators proposed that this interaction might al-ter the conformation of CARMA1 and enhanceits association with Bcl10 and MALT1 and/orstabilize the CBM complex.
Ca2+-MEDIATED SIGNALINGPATHWAYS
Ca2+ ions are universal second messengers ineukaryotic cells. The IP3 generated by TCR-stimulated PLCγ1 activity stimulates Ca2+-permeable ion channel receptors (IP3R) on theendoplasmic reticulum (ER) membrane, lead-ing to the release of ER Ca2+ stores intothe cytoplasm. Depletion of ER Ca2+ trig-gers a sustained influx of extracellular Ca2+
through the activation of plasma membraneCa2+ release-activated Ca2+ (CRAC) channelsin a process known as store-operated Ca2+ en-try (SOCE) (reviewed in 84) (Figure 1d ). Fordecades the CRAC channels had only beenidentified by their biophysical properties, andit has just been in the past few years thatthe pore-forming subunit of the channels wasidentified as the four-transmembrane domain–containing molecule Orai1 (85–87). Addition-ally, studies in the past few years have revealedthe sensor for depleted ER Ca2+ stores andthe activator of CRAC channels as stromal in-teraction molecule (STIM) (88, 89). STIM isan ER-resident transmembrane protein with aC-terminal cytoplasmic coiled-coil motif and,within the ER lumen, an N-terminal sterileα motif (SAM) and paired EF hands, whereone hand binds a single Ca2+ ion with lowaffinity (88, 89). Two STIM proteins, STIM1and STIM2, are found in mammals, and re-cent work has shown that STIM1 is importantfor the initial robust phase of SOCE, whereasSTIM2 is important for maintaining basal Ca2+
levels and sustaining the late phase of SOCE(90, 91). Following ER Ca2+ depletion, STIM1molecules aggregate in clusters that preferen-tially localize to sites of ER plasma membrane
www.annualreviews.org • T Cell Activation 601
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
NFAT: nuclear factorof activated T cells
apposition, where they colocalize with clustersof Orai1, forming punctae (92–94). Thebiochemical mechanism by which STIM1 cou-ples Ca2+ depletion to CRAC activation isnot yet fully understood and is an area of in-tense investigation. Recent work has shown thatSTIM1 oligomerization is sufficient to inducepunctae formation and CRAC channel activa-tion independent of Ca2+ store depletion (95).Further studies showed that the C-terminalcoiled-coil domain of STIM1 alone can inducedimerization of Orai1 dimers, which is suffi-cient for CRAC channel activation independentof punctae formation and store depletion (96).How STIM1 oligomers translocate to areas ofER plasma membrane apposition and how, oncethere, they induce the Orai1 tetramerization re-main unknown. Interestingly, the WASp familyverprolin homologous protein (WAVE2) com-plex is also required for SOCE activity, througha mechanism that remains to be fully elucidated,although independent of its function in actinremodeling (97, 98). CRAC channels appear tobe the dominant mode of Ca2+ entry in T cells,but other Ca2+ channels exist. Their relevanceremains unclear (reviewed in 84).
TCR-induced increases in intracellularCa2+ levels result in the activation of Ca2+
and calmodulin-dependent transcription fac-tors, including myocyte-enhancing factor 2(MEF2) and downstream regulatory elementantagonist modulator (DREAM), as well as sig-naling proteins, including the phosphatase cal-cineurin and the Ca2+-calmodulin-dependentkinase (CaMK), that in turn activate a vari-ety of transcription programs (reviewed in 99).Activated calcineurin dephosphorylates mem-bers of the nuclear factor of activated T cells(NFAT) family, leading to their translocationto the nucleus. In the nucleus, NFAT iso-forms can form cooperative complexes with avariety of other transcription factors, therebyintegrating signaling pathways, resulting in dif-ferential gene expression patterns and func-tional outcomes, depending on the context ofthe TCR signal. The most well-studied inter-action is NFAT/AP-1, which integrates Ca2+
and Ras signals and results in the expression of
genes important for T cell activation includ-ing IL-2. In contrast, NFAT activity in the ab-sence of AP-1 activation induces a pattern ofgene expression that ultimately results in T cellanergy and a characteristic lack of IL-2 pro-duction (100). It is still unclear whether NFATisoforms are cooperating with other transcrip-tion factors or are functioning as dimers toinduce the anergic transcriptional pattern (re-viewed in 101). The regulatory T cell lineage–specific transcription factor forkhead boxprotein 3 (FOXP3) also cooperates with NFATand antagonizes NFAT/AP-1 gene transcrip-tion, resulting in Treg functional gene expres-sion and a lack of IL-2 production (102). Finally,NFAT family members can also cooperate withSTAT proteins to induce either Th1 or Th2differentiation through expression of T-bet orGATA3, respectively (99).
ACTIN AND CYTOSKELETALRESPONSES
When a T cell is presented with cognate anti-gen by an APC, signals from the TCR initiatea program of actin cytoskeletal rearrangementsthat results in polarization and activation of theT cell (reviewed in 104). Actin reorganization isessential for T cell function, as actin polymer-ization inhibitors impede T cell/APC interac-tions (105) and abolish proximal TCR signals(106).
T cell/APC conjugation results in morpho-logical changes, as the stimulated T cell roundsup and accumulates filamentous actin (F-actin)at the stimulatory interface. These changes arethought to depend on a TCR-induced increasein plasma membrane fluidity and a decreasein cellular motility. Cessation of motility isassociated with TCR-induced Ca2+-dependentphosphorylation and deactivation of the myosinmotor protein MyH9; however, the signalingpathway linking the TCR to this event has notyet been defined fully (107). Plasma membranefluidity is increased, in part, by the TCR- andVav1-dependent transient dephosphorylationof ERM (ezrin, radixin, and moesin) proteins,resulting in the loss of their ability to link the
602 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
plasma membrane to the actin cytoskeleton(108). Ca2+ signaling and integrin activationdownstream of the TCR result in additionalmodifications of actin-associated proteins thatmay play roles in altering plasma membranerigidity (104).
Accumulation of F-actin at the T cell/APCinterface is the result of TCR-induced lo-calized activation of multiple actin regulatoryand polymerizing pathways, the best stud-ied of which involves the actin-related pro-teins 2/3 (Arp2/3) complex, although Arp2/3-independent pathways also contribute to thisprocess (109). Activation of Arp2/3 requires itsinteraction with nucleation-promoting factors(NPF) including Wiskott-Aldrich syndromeprotein (WASp), WAVE2, and hematopoieticcell lineage–specific protein 1 (HS1). WASp isrecruited to the site of TCR activation throughits interaction with the SLP-76-associatedadapter protein Nck, where it is activated viaVav1-dependent stimulation of the Rho fam-ily GTPase Cdc42 (110). Vav1-mediated acti-vation of a second Rho family GTPase, Rac1,results in the activation of WAVE2 (104). Giventhe proposed reliance of WASp and WAVE2on Vav1-mediated GTPase activation, it is in-teresting that actin-dependent processes thatare defective in Vav1-deficient T cells canbe rescued with the expression of a GEF-inactive Vav1 mutant, suggesting that otherRac and Cdc42 GEFs may be able to sup-port TCR-induced actin changes (55). Thisresult also suggests that other Vav1 functionsare important for TCR-induced actin changes.Consistent with this is the observation that,through its protein interaction domains, Vav1may contribute to WAVE2 and WASp activa-tion through the recruitment of Dynamin2, aGTPase that is important for TCR-inducedactin dynamics (111).
Activation of the T cell in response to anAPC also results in the polarization of the Tcell, whereby the microtubular organizing cen-ter (MTOC) moves toward the T cell/APCcontact site (112). Although polarization of theMTOC has long been observed as a hallmark ofproductive T cell/APC conjugation, the signal-
cSMAC: centralsupramolecularactivation cluster
pSMAC: peripheralsupramolecularactivation cluster
ing mechanisms responsible for this movementremain undefined. Recent data suggest, how-ever, that the adapter protein ADAP (a com-ponent of the SLP-76-nucleated complex) mayplay a role through its interaction with the mi-crotubule motor protein dynein (113). Move-ment of the MTOC appears essential for theformation of the immunological synapse (IS).
The IS is an organized structure that devel-ops at the contact site between the T cell and theAPC. It is composed of two concentric regionsbased on molecular composition: the TCR-rich central supramolecular activation cluster(cSMAC), surrounded by the integrin-rich pe-ripheral SMAC (pSMAC) (114, 115). Althoughthe IS was described approximately 10 yearsago, its precise role in T cell activation re-mains unclear. Initially, the concentration ofreceptors in the cSMAC led to the proposalthat the cSMAC is the site of enhanced recep-tor engagement and prolonged signaling (115).However, later studies showed that TCR signalspeak prior to cSMAC formation and suggestedthat the cSMAC is primarily the site of TCRdegradation (116, 121, 122). More recently, ithas been proposed that the cSMAC is the siteof both TCR signal enhancement and TCRdegradation, and the balance between these twoprocesses is determined by the quality of theantigen such that the cSMAC can serve to am-plify weak agonist signals (117). Most studies ofthe cSMAC have focused on the TCR and itsassociated molecules. However, a newly definedsubregion of the cSMAC that is rich in CD28, acostimulatory molecule (see below), and PKCθ
but relatively devoid of the TCR points to thepotential importance of the cSMAC for costim-ulatory molecule signal transduction (118).
Although the precise roles of the cSMACremain controversial, it is now well acceptedthat the initiation of TCR signals occurs in pe-ripheral microclusters that begin to form priorto IS formation. pMHC/TCR ligation resultsin the formation of intracellular microclustersthat contain the TCR complex and associatedsignaling molecules, including LAT and SLP-76 (120). These clusters initiate and can sus-tain Ca2+ signals (121). The clusters persist
www.annualreviews.org • T Cell Activation 603
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
Inside-out signaling:signals initiated byengagement ofimmunoreceptors thatlead to conformationalchanges and clusteringof integrins, therebyincreasing the affinityand avidity of theintegrins for theirligands
for a short time, after which they convergetoward the cSMAC (121, 122). Formation andtranslocation of the clusters are dependent onF-actin dynamics, and new clusters continue toform even after the mature IS is established(121–123). Integrins play a key role in sus-taining microclusters, emphasizing their im-portance for T cell activation (124).
Opposite the MTOC and IS, another or-dered structure forms, known as the distal polecomplex (DPC). Although the role of the DPCis not known for certain, investigators specu-late that the DPC is critical for sequesteringnegative regulators away from the TCR acti-vation complex (104). Additionally, the DPCmay contribute to the polarization of keysignaling molecules that may be required fordistinguishing memory versus effector fate de-cisions in recently divided cells (125). Forma-tion of this complex is dependent on F-actinand the rephosphorylation of ERM proteinsthat migrate to the distal pole linking signal-ing molecules to the cytoskeleton (126).
TCR signaling cascades and pathwaysdownstream of actin reorganization are inter-twined and difficult to tease apart, as manyof the effector molecules involved have mul-tiple enzymatic and adapter functions. WASp,WAVE2, and Vav1 signals play roles in TCR-induced signaling that appear to be independentof their roles in actin responses (52, 97, 127).Therefore, loss of different actin regulators mayresult in complex TCR signaling defects (128).Future studies are required to understand fullythe feedforward and feedback mechanisms thatdefine the interdependence between cytoskele-tal dynamics and T cell activation.
TCR INSIDE-OUT SIGNALINGTO INTEGRINS
Integrins are αβ heterodimeric receptors re-sponsible for mediating cell/cell or cell/matrixadhesions. Key T cell integrins include leuko-cyte function–associated antigen-1 (LFA-1)and very late antigen-4 (VLA-4), which bindtheir respective ligands intercellular adhe-sion molecule (ICAM) and vascular cell ad-
hesion molecule (VCAM) and fibronectin onother immune cells, endothelial cells, fibrob-lasts, and extracellular matrix proteins. Activa-tion of integrins (increasing their affinity andavidity for ligand) is critically dependent onbiochemical events initiated by the TCR, aprocess designated inside-out signaling (129).Although the pathway from the TCR to inte-grin activation has not been completely eluci-dated, TCR-mediated activation of several keysignaling molecules as well as TCR-inducedactin/cytoskeletal changes have been implicatedin this process.
A central regulator of inside-out signalingis the small GTPase Ras-proximity-1 (Rap1).Rap1 enhances T cell activation by mediatingTCR-induced adhesion to ICAM-1. This con-clusion is based on studies utilizing overexpres-sion of dominant-negative and constitutivelyactive forms of Rap1 (130, 131) and more re-cently by analysis of mice deficient in Rap1A,in which TCR-induced adhesion to ICAM-1is markedly reduced (132). The importance ofunderstanding the role of Rap1 is clear, as muta-tions in Rap1-mediated integrin activation havebeen linked to leukocyte adhesion–deficiencysyndrome, a disease that can lead to severe bac-terial infections (reviewed in 129).
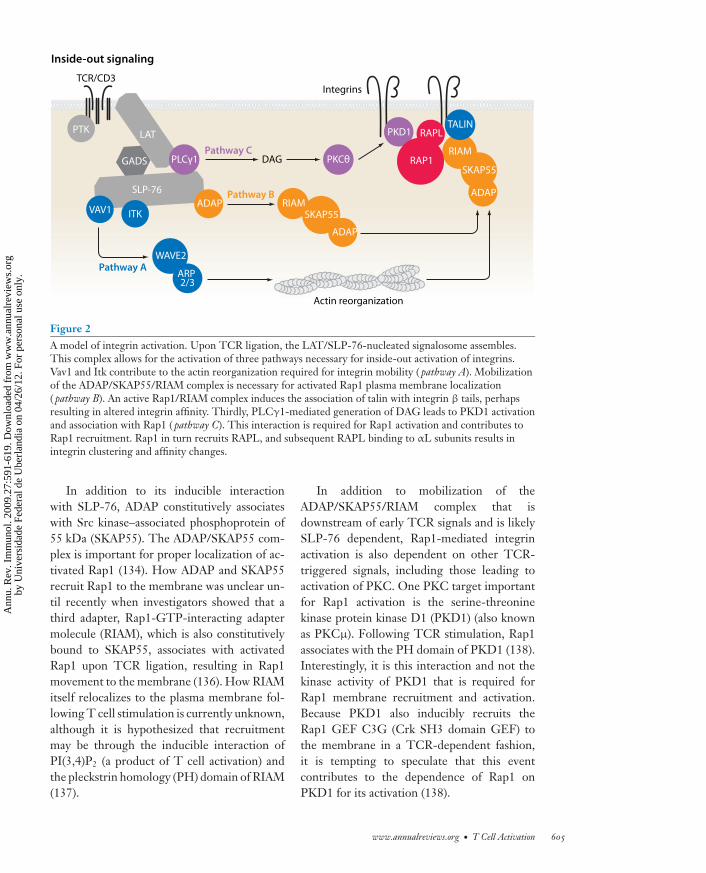
Many proximal signaling molecules thatcomprise the early TCR signalosome, includ-ing LAT, SLP-76, and PLCγ1, are required forintegrin and Rap1 activation (reviewed in 129)(Figure 2). However, there are downstream ef-fectors that have a more selective role in in-tegrin activation, including the adapter pro-tein ADAP. T cells from ADAP-deficient miceare defective in TCR-induced LFA-1 cluster-ing and adhesion to ICAM (133). The associ-ation of ADAP with SLP-76 appears to be re-quired for integrin activation, as overexpressionof ADAP but not a mutant of ADAP that can-not bind to SLP-76 enhances integrin functionfollowing TCR ligation (134, 135). Althoughmice expressing the reciprocal mutation in theSH2 domain of SLP-76 have been generatedand share several characteristics with ADAP-deficient mice, TCR-induced integrin activa-tion in these mice has not yet been reported.
604 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.
ANRV371-IY27-21 ARI 16 February 2009 13:31
Pathway A
Pathway B
Pathway C
Actin reorganization
TCR/CD3Integrins
Inside-out signaling
DAG
PTK
SLP-76
LAT
ADAP RIAMSKAP55
ADAP
ADAP
RIAM
SKAP55
VAV1
WAVE2
TALIN
ARP 2/3
ITK
PLCγ1 PKCθ
PKD1 RAPL
RAP1GADS
Figure 2A model of integrin activation. Upon TCR ligation, the LAT/SLP-76-nucleated signalosome assembles.This complex allows for the activation of three pathways necessary for inside-out activation of integrins.Vav1 and Itk contribute to the actin reorganization required for integrin mobility ( pathway A). Mobilizationof the ADAP/SKAP55/RIAM complex is necessary for activated Rap1 plasma membrane localization( pathway B). An active Rap1/RIAM complex induces the association of talin with integrin β tails, perhapsresulting in altered integrin affinity. Thirdly, PLCγ1-mediated generation of DAG leads to PKD1 activationand association with Rap1 ( pathway C). This interaction is required for Rap1 activation and contributes toRap1 recruitment. Rap1 in turn recruits RAPL, and subsequent RAPL binding to αL subunits results inintegrin clustering and affinity changes.
In addition to its inducible interactionwith SLP-76, ADAP constitutively associateswith Src kinase–associated phosphoprotein of55 kDa (SKAP55). The ADAP/SKAP55 com-plex is important for proper localization of ac-tivated Rap1 (134). How ADAP and SKAP55recruit Rap1 to the membrane was unclear un-til recently when investigators showed that athird adapter, Rap1-GTP-interacting adaptermolecule (RIAM), which is also constitutivelybound to SKAP55, associates with activatedRap1 upon TCR ligation, resulting in Rap1movement to the membrane (136). How RIAMitself relocalizes to the plasma membrane fol-lowing T cell stimulation is currently unknown,although it is hypothesized that recruitmentmay be through the inducible interaction ofPI(3,4)P2 (a product of T cell activation) andthe pleckstrin homology (PH) domain of RIAM(137).
In addition to mobilization of theADAP/SKAP55/RIAM complex that isdownstream of early TCR signals and is likelySLP-76 dependent, Rap1-mediated integrinactivation is also dependent on other TCR-triggered signals, including those leading toactivation of PKC. One PKC target importantfor Rap1 activation is the serine-threoninekinase protein kinase D1 (PKD1) (also knownas PKCμ). Following TCR stimulation, Rap1associates with the PH domain of PKD1 (138).Interestingly, it is this interaction and not thekinase activity of PKD1 that is required forRap1 membrane recruitment and activation.Because PKD1 also inducibly recruits theRap1 GEF C3G (Crk SH3 domain GEF) tothe membrane in a TCR-dependent fashion,it is tempting to speculate that this eventcontributes to the dependence of Rap1 onPKD1 for its activation (138).
www.annualreviews.org • T Cell Activation 605
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
Costimulation:signals delivered to Tcells by cell surfacereceptors other thanthe TCR itself thatpotentiate T cellactivation
Activated Rap1 can also associate with theeffector regulator of cell adhesion and polariza-tion enriched in lymphoid tissues (RAPL). Thisinteraction is coincident with RAPL membranelocalization and necessary for binding of RAPLto the αL subunit of LFA-1. In cell line models,this association is important for LFA-1 cluster-ing as well as affinity modulation (139).
In addition to regulating the activationof Rap1, signals from the TCR also reg-ulate cytoskeletal attachments to integrins.One cytoskeletal binding protein important forintegrin activation is talin. Recent studies inplatelets have demonstrated that Rap1 ac-tivity enhances the association of talin withβ-integrin subunits through association withRIAM (140). It is possible that formation ofa talin/RIAM/Rap1 complex may enable talinto bind integrins, which may induce the high-affinity ligand-binding state. Talin is not theonly actin-binding protein implicated in TCR-induced integrin activation. Vinculin, WAVE2(98), and the Arp2/3 complex also play roles inthis process, and defining the precise steps thatlink early TCR signals to activation of thesemolecules is an area of active investigation.
COSTIMULATION
One central tenet of T cell activation is that sig-naling solely through the TCR results in a non-responsive state (anergy) in which T cells arerefractory to restimulation. Coligation of othercell surface receptors provides additional sig-nals required for anergy avoidance and produc-tive T cell activation. Although many cell sur-face receptors can enhance signaling throughthe TCR, CD28 does so more robustly thanother costimulatory molecules.
Numerous studies have shown that CD28promotes T cell proliferation, cytokine produc-tion (via gene transcription and mRNA stabil-ity), cell survival, and cellular metabolism (re-viewed in 141). One key effector downstreamof CD28 is PI3K. Following binding of CD28to its ligands CD80 or CD86 on APCs, thep85 regulatory subunit of PI3K associates witha pYMNM motif located in the cytoplasmic
tail of CD28 (142). This regulatory subunit re-cruits the p110 catalytic subunit of PI3K, whichconverts PIP2 to phosphatidylinositol (3,4,5)trisphosphate (PIP3) at the membrane. Local-ized PIP3 generation serves as a docking site forthe PH domains of PDK1 (3-phosphoinositide-dependent protein kinase 1) and its targetAkt.
Akt phosphorylates multiple proteins, en-abling it to affect numerous cellular responses.Activation of Akt enhances the nuclear translo-cation of NF-κB, which has positive effectson the expression of prosurvival genes includ-ing Bcl-xl. Emerging data suggest that Akt ac-complishes this function by associating withCARMA1 and facilitating the assembly of theCBM complex (143, 144), a step critical forNF-κB activation (see above). The ability ofAkt to promote prosurvival gene expression,coupled with the ability of Akt to inhibit tran-scription factors that promote cell cycle arrest,results in Akt-driven cell survival and prolifera-tion (141). Akt also affects optimal transcriptionof NFAT-regulated genes, such as IL-2. Onewell-known target of Akt is GSK-3 (glycogen-synthase kinase 3), a serine-threonine kinasethat promotes nuclear export of NFAT (145).Thus, inactivation of GSK-3 by Akt might beone pathway responsible for prolonged NFATnuclear localization and thus IL-2 transcrip-tion following CD28 costimulation. Recently,a GSK-3-independent mechanism by whichAkt may regulate NFAT activity was suggested.This model posits that phosphorylated NFATis bound by the scaffolding protein Homer,thus inhibiting access of calcineurin to NFAT(146). Investigators proposed that CD28 lig-ation induces Akt-mediated phosphorylationof Homer, resulting in its dissociation frompNFAT. Unbound pNFAT would then be sus-ceptible to calcineurin phosphatase activity andnuclear entry. Whether this pathway is Akt de-pendent remains to be rigorously tested. Lastly,TCR/CD28 coligation increases the cell sur-face expression of the insulin transporter Glut1,leading to increased glucose uptake and glycol-ysis (147, 148), which is also mediated by Akt.Together, these data provide a framework for
606 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
how Akt mediates T cell growth and survivaldownstream of CD28.
The CD28-mediated generation of PIP3
also serves as a docking site for the PHdomain of Itk. Although Itk inducibly associateswith the LAT/SLP-76/Gads/PLCγ1 complexthat forms following TCR ligation, its local-ization and activation also depends on PI3K-generated PIP3 (61). Itk can also associate di-rectly with CD28 via the CD28 proximal PxxPmotif (61). It is possible that this interactionkeeps Itk close to Lck (which binds to the dis-tal PxxP motif of CD28), allowing for Lck-mediated phosphorylation and activation of Itk(149) and enhanced Ca2+ flux, another charac-teristic of CD28-mediated signaling. Despitethe fact that Itk and CD28 associate with oneanother, studies in Itk-deficient T cells demon-strate that some CD28 signaling is still intact inthe absence of Itk, suggesting that this interac-tion may not be that critical for CD28 signal-ing (150). Although the proline motifs in thetail of CD28 are required for CD28-mediatedproliferation and IL-2 production, these motifsare dispensable for Bcl-xl upregulation (151).This function appears to be more reliant on theproximal YMNM p85 binding site (151). Thus,CD28 can differentially regulate proliferationand survival in activated T cells.
Another more recently described functionof CD28 is induction of arginine methyla-tion. Following CD28 ligation, protein argi-nine methyltransferase activity increases, andarginine methylation of multiple proteins, in-cluding Vav1, is induced. Vav1 arginine methy-lation appears to occur in the nucleus and corre-lates with IL-2 production (152). Although theprecise biologic significance of this posttrans-lational modification is unknown, this pathwaymay provide yet another mechanism by whichCD28 regulates TCR signaling.
Many of the pathways described above areactivated by TCR ligation alone; however, themagnitude of the response is considerably aug-mented with CD28 coligation. This observa-tion has led to speculation that CD28 engage-ment results primarily in a quantitative ratherthan a qualitative change in T cell activation
parameters (141). Although this appears to betrue, it is also true that quantitative differencesin signaling can result in qualitatively distinctfunctional outcomes.
CD28-deficient mice exhibit dampened im-mune responses to a variety of infectious agents(reviewed in 141). Although these studiesdemonstrate the importance of costimulationby CD28, not all immune responses are severelyimpacted by its loss (153). Such observations in-dicate that molecules other than CD28 can pro-vide costimulation for T cells. Indeed, multiplesurface receptors have been described as havingcostimulatory functions. Included among theseare CD2, CD5, CD30, 4-1BB, OX40, induciblecostimulator (ICOS), and LFA-1. For this re-view, we focus on the CD28-related proteinICOS and two members of the tumor necro-sis factor receptor (TNFR) family to provideexamples of how costimulatory molecules canlink to downstream effectors, either directly orthrough adapter proteins.
Unlike CD28, which is expressed at con-stant levels on both resting and activated T cells,ICOS is inducibly expressed on activated T cells(154). ICOS deficiency results in impaired im-mune responses, similar to yet not as severeas those observed in CD28 knockout models,suggesting that these two molecules may func-tion in similar pathways (155). Indeed, ICOSshares several structural features with CD28,including a YMXM motif in its cytoplasmictail that associates with p85 (155). This siteis presumed to be responsible for PI3K-drivenAkt and/or Itk activation observed downstreamof CD3/ICOS stimulation, which likely con-tributes to the similarities seen in CD3/CD28and CD3/ICOS-stimulated cells (156). ICOSdoes not induce IL-2 gene transcription asCD28 does. Failure to induce IL-2 has beenattributed, at least in part, to the inability of theYxxM motif of ICOS to associate with Grb2,an association that is present via this motif inCD28 (157). Therefore, although ICOS acti-vates genes similar to those induced by CD28,there are differences in the degree to which par-ticular genes are expressed. These differenceshave in vivo relevance, as mice doubly deficient
www.annualreviews.org • T Cell Activation 607
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
in CD28 and ICOS are severely defective ingenerating immune responses (158).
Outside of the CD28 family of costimu-latory molecules, the OX40 (CD134) and 4-1BB (CD137) members of the TNFR familyprovide costimulation upon engagement withtheir ligands OX40L and 4-1BBL. Like CD28,OX40 or 4-1BB ligation induces activation ofPI3K/Akt, NF-κB, JNK, and p38 MAPK (159).However, unlike CD28 and ICOS, OX40 and4-1BB do not directly associate with proteinkinases but rather link to downstream sig-naling through the TRAF (TNFR-associatedfactor) family of adapter proteins. The pro-teins involved in linking TRAF signaling toNF-κB and the JNK/p38 pathways have notbeen completely elucidated in primary T cells.However, studies in other systems implicateTRAFs themselves in the direct recruitment ofthe IKK complex as well as of serine/threoninekinases that initiate signaling to JNK and p38(reviewed in 160).
Many of the same genes are regulated down-stream of CD28 and TNFR family members.One reason for such overlap may be due tothe timing of receptor expression. CD28 is ex-pressed early and is critical for induction ofan immune response. It promotes expressionof several other costimulatory molecules in-cluding ICOS, OX40, and 4-1BB. Once ex-pressed, these receptors prolong or sustain animmune response and, in the case of OX40and 4-1BB, are important for memory T cellformation (159). The use of alternative meansby which to activate these similar pathways,e.g., direct binding to kinases versus use ofadapter proteins, may also allow for differentialnegative regulation of these pathways. In fact,some TRAF proteins negatively regulate NF-κB; thus, association of TNFRs with differentTRAF family members can modulate the im-mune response (160). Lastly, a role for manycostimulatory molecules, including ICOS and4-1BB, in the development and/or function ofregulatory T cells is becoming increasingly ap-parent (161). This layer of complexity will haveto be taken into account when deciphering theroles of costimulatory molecules in vivo.
NEGATIVE REGULATIONOF TCR SIGNALING
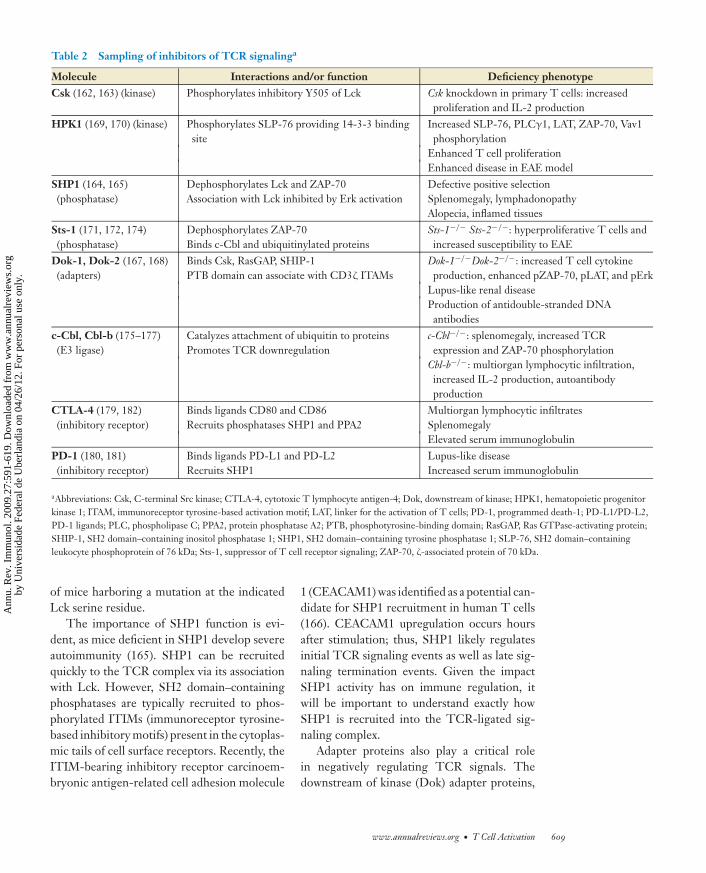
As outlined above, signaling through the TCRtriggers an array of signals that activate multipleeffector pathways. Activation of these pathwaysis regulated to ensure that T cells respond toappropriate ligands and for the proper dura-tion. As with positive regulation of T cell sig-naling, negative regulation is mediated throughboth TCR-generated signals and those emanat-ing from other cell surface receptors (Table 2).
Even the most proximal TCR signalingevents are actively regulated. For example, mul-tiple proteins contribute to the regulation ofLck activity. C-terminal Src kinase (Csk) is re-sponsible for phosphorylating Lck on its in-hibitory tyrosine residue (Y505) and maintain-ing Lck in an inactive state (162, reviewed in163). Countering this is the phosphatase CD45that dephosphorylates the inhibitory site allow-ing for Lck autophosphorylation and activa-tion. Interestingly, CD45 can also limit Lckactivity by dephosphorylating its active site(163). Whether CD45 negatively or positivelyimpacts TCR signaling is likely to be con-trolled by its proximity to TCR-stimulated ef-fector molecules during TCR engagement andwhether CD45 itself is in an enzymatically fa-vorable conformation.
An additional layer of Lck regulation initi-ated by TCR signals has been proposed. As ameans to explain how the TCR can distinguishbetween strong and weak ligands, investigatorsshowed that weak or antagonistic TCR ligationresults in rapid Lck-mediated phosphorylationof SHP1 (SH2 domain–containing protein-tyrosine phosphatase) (164). SHP1 then de-phosphorylates the active site of Lck, resultingin cessation of the TCR signal. Conversely, inthe presence of strong or agonistic TCR lig-ation, Erk is rapidly activated and phosphory-lates Lck on Ser59. This activity is thought toprevent SHP1 binding, thus keeping Lck ac-tive to sustain TCR signals and further amplifyErk activity. The extent to which this regulatoryloop operates in vivo in the context of agoniststimulation awaits the generation and analysis
608 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.
ANRV371-IY27-21 ARI 16 February 2009 13:31
Table 2 Sampling of inhibitors of TCR signalinga
Molecule Interactions and/or function Deficiency phenotypeCsk (162, 163) (kinase) Phosphorylates inhibitory Y505 of Lck Csk knockdown in primary T cells: increased
proliferation and IL-2 productionHPK1 (169, 170) (kinase) Phosphorylates SLP-76 providing 14-3-3 binding
siteIncreased SLP-76, PLCγ1, LAT, ZAP-70, Vav1phosphorylation
Enhanced T cell proliferationEnhanced disease in EAE model
SHP1 (164, 165)(phosphatase)
Dephosphorylates Lck and ZAP-70Association with Lck inhibited by Erk activation
Defective positive selectionSplenomegaly, lymphadonopathyAlopecia, inflamed tissues
Sts-1 (171, 172, 174)(phosphatase)
Dephosphorylates ZAP-70Binds c-Cbl and ubiquitinylated proteins
Sts-1−/− Sts-2−/−: hyperproliferative T cells andincreased susceptibility to EAE
Dok-1, Dok-2 (167, 168)(adapters)
Binds Csk, RasGAP, SHIP-1PTB domain can associate with CD3ζ ITAMs
Dok-1−/−Dok-2−/−: increased T cell cytokineproduction, enhanced pZAP-70, pLAT, and pErk
Lupus-like renal diseaseProduction of antidouble-stranded DNAantibodies
c-Cbl, Cbl-b (175–177)(E3 ligase)
Catalyzes attachment of ubiquitin to proteinsPromotes TCR downregulation
c-Cbl−/−: splenomegaly, increased TCRexpression and ZAP-70 phosphorylation
Cbl-b−/−: multiorgan lymphocytic infiltration,increased IL-2 production, autoantibodyproduction
CTLA-4 (179, 182)(inhibitory receptor)
Binds ligands CD80 and CD86Recruits phosphatases SHP1 and PPA2
Multiorgan lymphocytic infiltratesSplenomegalyElevated serum immunoglobulin
PD-1 (180, 181)(inhibitory receptor)
Binds ligands PD-L1 and PD-L2Recruits SHP1
Lupus-like diseaseIncreased serum immunoglobulin
aAbbreviations: Csk, C-terminal Src kinase; CTLA-4, cytotoxic T lymphocyte antigen-4; Dok, downstream of kinase; HPK1, hematopoietic progenitorkinase 1; ITAM, immunoreceptor tyrosine-based activation motif; LAT, linker for the activation of T cells; PD-1, programmed death-1; PD-L1/PD-L2,PD-1 ligands; PLC, phospholipase C; PPA2, protein phosphatase A2; PTB, phosphotyrosine-binding domain; RasGAP, Ras GTPase-activating protein;SHIP-1, SH2 domain–containing inositol phosphatase 1; SHP1, SH2 domain–containing tyrosine phosphatase 1; SLP-76, SH2 domain–containingleukocyte phosphoprotein of 76 kDa; Sts-1, suppressor of T cell receptor signaling; ZAP-70, ζ-associated protein of 70 kDa.
of mice harboring a mutation at the indicatedLck serine residue.
The importance of SHP1 function is evi-dent, as mice deficient in SHP1 develop severeautoimmunity (165). SHP1 can be recruitedquickly to the TCR complex via its associationwith Lck. However, SH2 domain–containingphosphatases are typically recruited to phos-phorylated ITIMs (immunoreceptor tyrosine-based inhibitory motifs) present in the cytoplas-mic tails of cell surface receptors. Recently, theITIM-bearing inhibitory receptor carcinoem-bryonic antigen-related cell adhesion molecule
1 (CEACAM1) was identified as a potential can-didate for SHP1 recruitment in human T cells(166). CEACAM1 upregulation occurs hoursafter stimulation; thus, SHP1 likely regulatesinitial TCR signaling events as well as late sig-naling termination events. Given the impactSHP1 activity has on immune regulation, itwill be important to understand exactly howSHP1 is recruited into the TCR-ligated sig-naling complex.
Adapter proteins also play a critical rolein negatively regulating TCR signals. Thedownstream of kinase (Dok) adapter proteins,
www.annualreviews.org • T Cell Activation 609
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
Dok-1 and -2, are expressed in T cells. Co-ordinated deletion or knockdown of theseproteins results in increased TCR cytokineproduction and proliferation; prolonged phos-phorylation of ZAP-70, LAT, SLP-76, andAkt; and the development of a lupus-like re-nal disease with high antidouble-stranded DNAantibody titers (167, 168). Dok can asso-ciate with several negative regulators includ-ing SHIP-1 (SH2 domain–containing inositolphosphatase), Csk, and Ras GTPase-activatingprotein (RasGAP); however, how these associ-ations mediate the negative regulatory role ofDok proteins remains to fully elucidated. Re-cent structure/function studies in T cell linesshowed that the phosphotyrosine-binding do-main (PTB) of Dok-1 and -2 can bind the ITAMmotif in CD3ζ, making it tempting to specu-late that Dok may compete with ZAP-70 forITAM binding (167). Whether this interactionis relevant in vivo remains to be shown. Dokproteins can also be recruited inducibly to aLAT-, Grb2-, and SHIP-1-containing molecu-lar complex (168). This finding is intriguing andbegs the question as to how LAT participates inboth positive and negative signaling pathways.Determining the domains necessary for the as-sembly of this negative regulatory complex andwhether it is separate from or a part of the stim-ulatory LAT-containing complex may provideinsight into how early phosphorylation eventsare regulated.
It has been suggested that SLP-76 can alsobe a target of negative regulation. The serine/threonine kinase HPK1 inducibly binds to theSH2 domain of SLP-76. This kinase has bothpositive and negative effects on TCR signaltransduction; however, its role as a promi-nent negative regulator was confirmed re-cently by the description of HPK1-deficientmice. HPK1-deficient T cells exhibit en-hanced phosphorylation of several early sig-naling molecules, and HPK1-deficient miceare more susceptible to experimental autoim-mune encephalomyelitis (EAE) than are wild-type mice (169). As a possible explanation forthese phenotypes, two groups have shown that
HPK1 can phosphorylate a serine residue inSLP-76 that mediates recruitment of 14-3-3family members (169, 170). 14-3-3 proteinshave been implicated in the regulation of severalsignaling pathways. Thus, although the pre-cise mechanism by which HPK1 disrupts TCRsignal transduction remains unclear, identifica-tion of a SLP-76/14-3-3 interaction may indi-cate that this mechanism involves a conservedmethod of signal transduction regulation.
The role of a novel family of proteins in reg-ulating TCR signaling has become appreciatedmore recently. The suppressor of T cell recep-tor signaling (Sts) family of proteins containstwo members, Sts-1 (TULA2) and -2 (TULA).Combined deficiency in these proteins leads tohyperproliferative T cells and an increased sus-ceptibility to autoimmunity (171). When thephenotype of Sts-1 and -2 knockout mice wasdescribed, the mechanism of negative regula-tion was unclear. Although still not understoodfully, the C-terminal phosphoglycerate mutasedomain of Sts-1 acts as a phosphatase withspecificity for Syk and, to a lesser degree, Srcfamily members, thus providing a model forhow Sts-1 may negatively regulate TCR sig-nal transduction (172). Interestingly, Sts-2 hasvery little phosphatase activity and, in cell linemodels, can enhance ZAP-70 activation (173).Recent studies demonstrating that Sts-2 canbind to and induce the degradation of c-Cbl(a negative regulator of TCR signaling; see be-low) have provided a mechanism by which Sts-2may act as a positive regulator of T cell activa-tion (174). Whether net ZAP-70 activity is ulti-mately increased or decreased in response to anin vivo challenge likely depends on the relativeexpression of these two family members.
Cbl proteins play an important role as neg-ative regulators of T cell signaling. Two familymembers, c-Cbl and Cbl-b, are immune mod-ulators, as deletion of each gene in vivo leads tohypercellularity and, in the case of Cbl-b, spon-taneous mulitorgan infiltration (175, 176). Bothc-Cbl and Cbl-b facilitate the ubiquitination ofproteins, targeting them for degradation. Theyalso mediate the downregulation of the TCR
610 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
following stimulation with antigenic peptidesand have been recently shown to play a role inthe dissipation of the early signaling complex(176–178). Thus, Cbl proteins, by way of theirability to regulate protein degradation, providean example of yet another means by which TCRsignaling is terminated.
Similar to the necessity of signals providedby the TCR to be complemented by costim-ulatory molecules, TCR-generated regulatorysignals are also aided by coreceptor signals.Cytotoxic T lymphocyte antigen-4 (CTLA-4)and programmed death-1 (PD-1) are two ex-amples of such receptors that limit the ex-pansion and activation of TCR-triggered Tcells. These molecules are found on activatedT cells with peak expression 24–48 h afterstimulation. Genetic studies have documentedthe importance of both for maintaining self-tolerance. By 5–6 days after birth, CTLA-4-deficient mice exhibit activated peripheral Tcells, splenomegaly, and lymphocytic infiltratesinto nonlymphoid organs (179). Although lessstriking, PD-1-deficient mice also develop au-toimmune features, including the developmentof a lupus-like disease by 6 months of age (180).Like costimulatory receptors, inhibitory recep-tors utilize motifs and molecular pathways simi-lar to those used by the TCR when propagatinga negative signal; they may even provide dock-ing sites for these shared molecules (181, 182).Another mode by which inhibitory receptorshave been proposed to function is to utilize thesame ligand and/or signaling molecules as cos-timulatory receptors, thus setting up the poten-tial for competition or sequestration of ligandsor key substrates. This means is best illustratedby CTLA-4 and CD28, which share the lig-ands CD80 and CD86 and counter each otherin the regulation of cell cycle proteins, cytokineexpression, and Cbl-b expression.
CONCLUSIONS
As in the first 10 years after identification ofthe TCR, the past 15 years have seen a dra-matic expansion in our understanding of thebiochemical pathways triggered downstream ofthe TCR. We now appreciate more fully howPTKs couple to effectors and second messen-gers via the utilization of adapter proteins, howRas and NF-κB pathways are activated, howCa2+ ions are sensed, that TCR signaling can berequired for the activation of other cell surfacereceptors, and that the actin cytoskeleton doesmore than just dictate cell shape. Studies inves-tigating signal transduction by costimulatorymolecules have highlighted the importance ofsignal amplification, and discoveries of spon-taneous or induced models of autoimmunityhave brought negative regulation of these sig-naling pathways to the forefront. However, asoften noted in this review, links between manysignaling molecules and cellular outcomes re-main poorly defined. Future studies will berequired to fill gaps in our knowledge andlikely will reveal new and unexpected interac-tions between currently known and unknownmolecules.
This review primarily focused on the signaltransduction pathways in naive T cells. Overthe next 15 years, it will be important to ap-ply current and new knowledge to other T celllineages, including T regulatory cells, naturalkiller T cells, and memory T cells, to estab-lish whether these paradigms are universal. Inthe coming years, the population-based analy-ses that established the field of signal transduc-tion will be driven toward single cell analyses.We can anticipate that this technological shiftwill allow for better analysis of human T cellsand the application of basic science to humandisease.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
www.annualreviews.org • T Cell Activation 611
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
ACKNOWLEDGMENTS
We thank Christopher P. Garvin for figure design, Dr. Art Weiss for helpful comments, and JustinaStadanlick for editorial assistance.
LITERATURE CITED
1. Haskins K, Kappler J, Marrack P. 1984. The major histocompatibility complex-restricted antigen re-ceptor on T cells. Annu. Rev. Immunol. 2:51–66
2. Cantrell D. 1996. T cell antigen receptor signal transduction pathways. Annu. Rev. Immunol. 14:259–743. Yanagi Y, Yoshikai Y, Leggett K, Clark SP, Aleksander I, Mak TW. 1984. A human T cell-specific cDNA
clone encodes a protein having extensive homology to immunoglobulin chains. Nature 308:145–494. Hedrick SM, Cohen DI, Nielsen EA, Davis MM. 1984. Isolation of cDNA clones encoding T cell-specific
membrane-associated proteins. Nature 308:149–535. Malissen M, Minard K, Mjolsness S, Kronenberg M, Goverman J, et al. 1984. Mouse T cell antigen re-
ceptor: structure and organization of constant and joining gene segments encoding the beta polypeptide.Cell 37:1101–10
6. Allison JP, McIntyre BW, Bloch D. 1982. Tumor-specific antigen of murine T-lymphoma defined withmonoclonal antibody. J. Immunol. 129:2293–300
7. Haskins K, Kubo R, White J, Pigeon M, Kappler J, Marrack P. 1983. The major histocompatibilitycomplex-restricted antigen receptor on T cells. I. Isolation with a monoclonal antibody. J. Exp. Med.157:1149–69
8. Meuer SC, Fitzgerald KA, Hussey RE, Hodgdon JC, Schlossman SF, Reinherz EL. 1983. Clonotypicstructures involved in antigen-specific human T cell function. Relationship to the T3 molecular complex.J. Exp. Med. 157:705–19
9. Borst J, Coligan JE, Oettgen H, Pessano S, Malin R, Terhorst C. 1984. The delta- and epsilon-chainsof the human T3/T-cell receptor complex are distinct polypeptides. Nature 312:455–58
10. Dembic Z, Haas W, Weiss S, McCubrey J, Kiefer H, et al. 1986. Transfer of specificity by murine α andβ T-cell receptor genes. Nature 320:232–38
11. Saito T, Germain RN. 1987. Predictable acquisition of a new MHC recognition specificity followingexpression of a transfected T-cell receptor β-chain gene. Nature 329:256–59
12. Weiss A, Stobo JD. 1984. Requirement for the coexpression of T3 and the T cell antigen receptor on amalignant human T cell line. J. Exp. Med. 160:1284–99
13. Abraham RT, Weiss A. 2004. Jurkat T cells and development of the T-cell receptor signalling paradigm.Nat. Rev. Immunol. 4:301–8
14. Weiss A, Imboden JB. 1987. Cell surface molecules and early events involved in human T lymphocyteactivation. Adv. Immunol. 41:1–38
15. Imboden JB, Stobo JD. 1985. Transmembrane signalling by the T cell antigen receptor. Perturbation ofthe T3-antigen receptor complex generates inositol phosphates and releases calcium ions from intracel-lular stores. J. Exp. Med. 161:446–56
16. Samelson LE, Patel MD, Weissman AM, Harford JB, Klausner RD. 1986. Antigen activation of murineT cells induces tyrosine phosphorylation of a polypeptide associated with the T cell antigen receptor.Cell 46:1083–90
17. Samelson LE, Phillips AF, Luong ET, Klausner RD. 1990. Association of the fyn protein-tyrosine kinasewith the T-cell antigen receptor. Proc. Natl. Acad. Sci. USA 87:4358–62
18. Veillette A, Bookman MA, Horak EM, Bolen JB. 1988. The CD4 and CD8 T cell surface antigens areassociated with the internal membrane tyrosine-protein kinase p56lck. Cell 55:301–8
19. Barber EK, Dasgupta JD, Schlossman SF, Trevillyan JM, Rudd CE. 1989. The CD4 and CD8 antigensare coupled to a protein-tyrosine kinase (p56lck) that phosphorylates the CD3 complex. Proc. Natl. Acad.Sci. USA 86:3277–81
20. June CH, Fletcher MC, Ledbetter JA, Schieven GL, Siegel JN, et al. 1990. Inhibition of tyrosine phos-phorylation prevents T-cell receptor-mediated signal transduction. Proc. Natl. Acad. Sci. USA 87:7722–26
612 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
21. Reth M. 1989. Antigen receptor tail clue. Nature 338:383–8422–25. Four papersdescribing chimericproteins thatdemonstrate thesignaling capability ofCD3ζ, supporting thenotion that CD3 is thesignaling component ofthe TCR complex.
22. Irving BA, Weiss A. 1991. The cytoplasmic domain of the T cell receptor ζ chain is sufficient tocouple to receptor-associated signal transduction pathways. Cell 64:891–901
23. Romeo C, Amiot M, Seed B. 1992. Sequence requirements for induction of cytolysis by the Tcell antigen/Fc receptor ζ chain. Cell 68:889–97
24. Wegener AM, Letourneur F, Hoeveler A, Brocker T, Luton F, Malissen B. 1992. The T cellreceptor/CD3 complex is composed of at least two autonomous transduction modules. Cell
68:83–9525. Letourneur F, Klausner RD. 1992. Activation of T cells by a tyrosine kinase activation domain
in the cytoplasmic tail of CD3ε. Science 255:79–82
26. Describes thecloning and initialcharacterization ofZAP-70, the Syk familyPTK essential forcoupling the TCR to itsdownstream signalingmachinery.
26. Chan AC, Iwashima M, Turck CW, Weiss A. 1992. ZAP-70: a 70 kd protein-tyrosine kinase thatassociates with the TCR ζ chain. Cell 71:649–62
27. Iwashima M, Irving BA, van Oers NS, Chan AC, Weiss A. 1994. Sequential interactions of the TCRwith two distinct cytoplasmic tyrosine kinases. Science 263:1136–39
28. Aivazian D, Stern LJ. 2000. Phosphorylation of T cell receptor ζ is regulated by a lipid dependent foldingtransition. Nat. Struct. Biol. 7:1023–26
29. Xu C, Gagnon E, Call ME, Schnell JR, Schwieters CD, et al. 2008. Regulation of T cell receptoractivation by dynamic membrane binding of the CD3epsilon cytoplasmic tyrosine-based motif. Cell135:702–13
30. The first ITAM-independent functionfor a CD3 molecule isidentified in the PRR ofCD3ε as a binding sitefor Nck.
30. Gil D, Schamel WW, Montoya M, Sanchez-Madrid F, Alarcon B. 2002. Recruitment of Nck byCD3ε reveals a ligand-induced conformational change essential for T cell receptor signaling andsynapse formation. Cell 109:901–12
31. Mingueneau M, Sansoni A, Gregoire C, Roncagalli R, Aguado E, et al. 2008. The proline-rich sequence ofCD3ε controls T cell antigen receptor expression on and signaling potency in preselection CD4+CD8+
thymocytes. Nat. Immunol. 9:522–3232. Szymczak AL, Workman CJ, Gil D, Dilioglou S, Vignali KM, et al. 2005. The CD3ε proline-rich
sequence, and its interaction with Nck, is not required for T cell development and function. J. Immunol.175:270–75
33. Gil D, Schrum AG, Daniels MA, Palmer E. 2008. A role for CD8 in the developmental tuning of antigenrecognition and CD3 conformational change. J. Immunol. 180:3900–9
34. Tailor P, Tsai S, Shameli A, Serra P, Wang J, et al. 2008. The proline-rich sequence of CD3ε as anamplifier of low-avidity TCR signaling. J. Immunol. 181:243–55
35. Takeuchi K, Yang H, Ng E, Park SY, Sun ZY, et al. 2008. Structural and functional evidence that Nckinteraction with CD3ε regulates T-cell receptor activity. J. Mol. Biol. 380:704–16
36. Rudolph MG, Stanfield RL, Wilson IA. 2006. How TCRs bind MHCs, peptides, and coreceptors. Annu.Rev. Immunol. 24:419–66
37. Sun ZJ, Kim KS, Wagner G, Reinherz EL. 2001. Mechanisms contributing to T cell receptor signalingand assembly revealed by the solution structure of an ectodomain fragment of the CD3εγ heterodimer.Cell 105:913–23
38. Ma Z, Janmey PA, Finkel TH. 2008. The receptor deformation model of TCR triggering. FASEB J.22:1002–8
39. Minguet S, Schamel WW. 2008. A permissive geometry model for TCR-CD3 activation. Trends Biochem.Sci. 33:51–57
40. Alarcon B, Swamy M, van Santen HM, Schamel WW. 2006. T-cell antigen-receptor stoichiometry:preclustering for sensitivity. EMBO Rep. 7:490–95
41. Davis SJ, Van Der Merwe PA. 2006. The kinetic-segregation model: TCR triggering and beyond. Nat.Immunol. 7:803–9
42. Varma R. 2008. TCR triggering by the pMHC complex: valency, affinity, and dynamics. Sci. Signal 1:pe2143. Krogsgaard M, Li QJ, Sumen C, Huppa JB, Huse M, Davis MM. 2005. Agonist/endogenous peptide-
MHC heterodimers drive T cell activation and sensitivity. Nature 434:238–4344. Zhang W, Sloan-Lancaster J, Kitchen J, Trible RP, Samelson LE. 1998. LAT: the ZAP-70 tyrosine
kinase substrate that links T cell receptor to cellular activation. Cell 92:83–92
www.annualreviews.org • T Cell Activation 613
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
45. Bubeck Wardenburg J, Fu C, Jackman JK, Flotow H, Wilkinson SE, et al. 1996. Phosphorylation ofSLP-76 by the ZAP-70 protein-tyrosine kinase is required for T-cell receptor function. J. Biol. Chem.271:19641–44
46. Koretzky GA, Abtahian F, Silverman MA. 2006. SLP76 and SLP65: complex regulation of signalling inlymphocytes and beyond. Nat. Rev. Immunol. 6:67–78
47. Sommers CL, Samelson LE, Love PE. 2004. LAT: a T lymphocyte adapter protein that couples theantigen receptor to downstream signaling pathways. Bioessays 26:61–67
48. LAT−/− mice reveala complete block in Tcell development that,along with studiesdescribing SLP-76−/−
mice, exemplifies in vivothe essential role ofadapter proteins forsignal transduction.
48. Zhang W, Sommers CL, Burshtyn DN, Stebbins CC, DeJarnette JB, et al. 1999. Essential roleof LAT in T cell development. Immunity 10:323–32
49. Liu SK, Fang N, Koretzky GA, McGlade CJ. 1999. The hematopoietic-specific adaptor protein Gadsfunctions in T-cell signaling via interactions with the SLP-76 and LAT adaptors. Curr. Biol. 9:67–75
50. Dombroski D, Houghtling RA, Labno CM, Precht P, Takesono A, et al. 2005. Kinase-independentfunctions for Itk in TCR-induced regulation of Vav and the actin cytoskeleton. J. Immunol. 174:1385–92
51. Reynolds LF, de Bettignies C, Norton T, Beeser A, Chernoff J, Tybulewicz VL. 2004. Vav1 transducesT cell receptor signals to the activation of the Ras/ERK pathway via LAT, Sos, and RasGRP1. J. Biol.Chem. 279:18239–46
52. Reynolds LF, Smyth LA, Norton T, Freshney N, Downward J, et al. 2002. Vav1 transduces T cellreceptor signals to the activation of phospholipase C-γ1 via phosphoinositide 3-kinase-dependent and-independent pathways. J. Exp. Med. 195:1103–14
53. Qi Q, August A. 2007. Keeping the (kinase) party going: SLP-76 and ITK dance to the beat. Sci. STKE2007:pe39
54. Beach D, Gonen R, Bogin Y, Reischl IG, Yablonski D. 2007. Dual role of SLP-76 in mediating T cellreceptor-induced activation of phospholipase C-γ1. J. Biol. Chem. 282:2937–46
55. Miletic AV, Sakata-Sogawa K, Hiroshima M, Hamann MJ, Gomez TS, et al. 2006. Vav1 acidic regiontyrosine 174 is required for the formation of T cell receptor-induced microclusters and is essential in Tcell development and activation. J. Biol. Chem. 281:38257–65
56. Sommers CL, Lee J, Steiner KL, Gurson JM, Depersis CL, et al. 2005. Mutation of the phospholipaseC-γ1-binding site of LAT affects both positive and negative thymocyte selection. J. Exp. Med. 201:1125–34
57. Jordan MS, Smith JE, Burns JC, Austin JE, Nichols KE, et al. 2008. Complementation in trans of alteredthymocyte development in mice expressing mutant forms of the adaptor molecule SLP76. Immunity28:359–69
58. Pletneva EV, Sundd M, Fulton DB, Andreotti AH. 2006. Molecular details of Itk activation by pro-lyl isomerization and phospholigand binding: the NMR structure of the Itk SH2 domain bound to aphosphopeptide. J. Mol. Biol. 357:550–61
59. Bogin Y, Ainey C, Beach D, Yablonski D. 2007. SLP-76 mediates and maintains activation of theTec family kinase ITK via the T cell antigen receptor-induced association between SLP-76 and ITK.Proc. Natl. Acad. Sci. USA 104:6638–43
60. Liu KQ, Bunnell SC, Gurniak CB, Berg LJ. 1998. T cell receptor-initiated calcium release is uncoupledfrom capacitative calcium entry in Itk-deficient T cells. J. Exp. Med. 187:1721–27
61. Berg LJ, Finkelstein LD, Lucas JA, Schwartzberg PL. 2005. Tec family kinases in T lymphocyte devel-opment and function. Annu. Rev. Immunol. 23:549–600
62. Shan X, Wange RL. 1999. Itk/Emt/Tsk activation in response to CD3 cross-linking in Jurkat T cellsrequires ZAP-70 and Lat and is independent of membrane recruitment. J. Biol. Chem. 274:29323–30
63. Bunnell SC, Diehn M, Yaffe MB, Findell PR, Cantley LC, Berg LJ. 2000. Biochemical interactionsintegrating Itk with the T cell receptor-initiated signaling cascade. J. Biol. Chem. 275:2219–30
64. Su YW, Zhang Y, Schweikert J, Koretzky GA, Reth M, Wienands J. 1999. Interaction of SLP adaptorswith the SH2 domain of Tec family kinases. Eur. J. Immunol. 29:3702–11
65. Genot E, Cantrell DA. 2000. Ras regulation and function in lymphocytes. Curr. Opin. Immunol. 12:289–94
66. Ebinu JO, Stang SL, Teixeira C, Bottorff DA, Hooton J, et al. 2000. RasGRP links T-cell receptorsignaling to Ras. Blood 95:3199–203
614 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
67. Egan SE, Giddings BW, Brooks MW, Buday L, Sizeland AM, Weinberg RA. 1993. Association of SosRas exchange protein with Grb2 is implicated in tyrosine kinase signal transduction and transformation.Nature 363:45–51
68. Dower NA, Stang SL, Bottorff DA, Ebinu JO, Dickie P, et al. 2000. RasGRP is essential for mousethymocyte differentiation and TCR signaling. Nat. Immunol. 1:317–21
69. Ebinu JO, Bottorff DA, Chan EY, Stang SL, Dunn RJ, Stone JC. 1998. RasGRP, a Ras guanyl nucleotide-releasing protein with calcium- and diacylglycerol-binding motifs. Science 280:1082–86
70. Roose JP, Mollenauer M, Gupta VA, Stone J, Weiss A. 2005. A diacylglycerol-protein kinase C-RasGRP1pathway directs Ras activation upon antigen receptor stimulation of T cells. Mol. Cell. Biol. 25:4426–41
71. Finco TS, Kadlecek T, Zhang W, Samelson LE, Weiss A. 1998. LAT is required for TCR-mediatedactivation of PLC-γ1 and the Ras pathway. Immunity 9:617–26
72. Roose JP, Mollenauer M, Ho M, Kurosaki T, Weiss A. 2007. Unusual interplay of two types of Rasactivators, RasGRP and SOS, establishes sensitive and robust Ras activation in lymphocytes. Mol. Cell.Biol. 27:2732–45
73. Hayashi K, Altman A. 2007. Protein kinase C theta (PKCθ): a key player in T cell life and death.Pharmacol. Res. 55:537–44
74. Melowic HR, Stahelin RV, Blatner NR, Tian W, Hayashi K, et al. 2007. Mechanism of diacylglycerol-induced membrane targeting and activation of protein kinase Cθ. J. Biol. Chem. 282:21467–76
75. Vallabhapurapu S, Karin M. 2009. Regulation and function of NFκB transcription factors in the immunesystem. Annu. Rev. Immunol. 27:693–733
76. Schulze-Luehrmann J, Ghosh S. 2006. Antigen-receptor signaling to nuclear factor κB. Immunity 25:701–15
77. Matsumoto R, Wang D, Blonska M, Li H, Kobayashi M, et al. 2005. Phosphorylation of CARMA1 playsa critical role in T cell receptor-mediated NFκB activation. Immunity 23:575–85
78. Sommer K, Guo B, Pomerantz JL, Bandaranayake AD, Moreno-Garcia ME, et al. 2005. Phosphorylationof the CARMA1 linker controls NFκB activation. Immunity 23:561–74
79. Sun L, Deng L, Ea CK, Xia ZP, Chen ZJ. 2004. The TRAF6 ubiquitin ligase and TAK1 kinase mediateIKK activation by BCL10 and MALT1 in T lymphocytes. Mol. Cell 14:289–301
80. Zhou H, Wertz I, O’Rourke K, Ultsch M, Seshagiri S, et al. 2004. Bcl10 activates the NF-κB pathwaythrough ubiquitination of NEMO. Nature 427:167–71
81. Coornaert B, Baens M, Heyninck K, Bekaert T, Haegman M, et al. 2008. T cell antigen receptorstimulation induces MALT1 paracaspase-mediated cleavage of the NF-κB inhibitor A20. Nat. Immunol.9:263–71
82. Rebeaud F, Hailfinger S, Posevitz-Fejfar A, Tapernoux M, Moser R, et al. 2008. The proteolytic activityof the paracaspase MALT1 is key in T cell activation. Nat. Immunol. 9:272–81
83. Medeiros RB, Burbach BJ, Mueller KL, Srivastava R, Moon JJ, et al. 2007. Regulation of NFκB activationin T cells via association of the adapter proteins ADAP and CARMA1. Science 316:754–58
84. Oh-Hora M, Rao A. 2008. Calcium signaling in lymphocytes. Curr. Opin. Immunol. 20:250–5885. Zhang SL, Yeromin AV, Zhang XH, Yu Y, Safrina O, et al. 2006. Genome-wide RNAi screen of Ca2+
influx identifies genes that regulate Ca2+ release-activated Ca2+ channel activity. Proc. Natl. Acad. Sci.USA 103:9357–62
86. Feske S, Gwack Y, Prakriya M, Srikanth S, Puppel SH, et al. 2006. A mutation in Orai1 causes immunedeficiency by abrogating CRAC channel function. Nature 441:179–85
87. Vig M, Peinelt C, Beck A, Koomoa DL, Rabah D, et al. 2006. CRACM1 is a plasma membrane proteinessential for store-operated Ca2+ entry. Science 312:1220–23
88. Roos J, DiGregorio PJ, Yeromin AV, Ohlsen K, Lioudyno M, et al. 2005. STIM1, an essential andconserved component of store-operated Ca2+ channel function. J. Cell Biol. 169:435–45
89. Liou J, Kim ML, Heo WD, Jones JT, Myers JW, et al. 2005. STIM is a Ca2+ sensor essential forCa2+-store-depletion-triggered Ca2+ influx. Curr. Biol. 15:1235–41
90. Oh-Hora M, Yamashita M, Hogan PG, Sharma S, Lamperti E, et al. 2008. Dual functions for theendoplasmic reticulum calcium sensors STIM1 and STIM2 in T cell activation and tolerance. Nat.Immunol. 9:432–43
www.annualreviews.org • T Cell Activation 615
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
91. Brandman O, Liou J, Park WS, Meyer T. 2007. STIM2 is a feedback regulator that stabilizes basalcytosolic and endoplasmic reticulum Ca2+ levels. Cell 131:1327–39
92. Liou J, Fivaz M, Inoue T, Meyer T. 2007. Live-cell imaging reveals sequential oligomerization and localplasma membrane targeting of stromal interaction molecule 1 after Ca2+ store depletion. Proc. Natl.Acad. Sci. USA 104:9301–6
93. Wu MM, Buchanan J, Luik RM, Lewis RS. 2006. Ca2+ store depletion causes STIM1 to accumulate inER regions closely associated with the plasma membrane. J. Cell Biol. 174:803–13
94. Luik RM, Wu MM, Buchanan J, Lewis RS. 2006. The elementary unit of store-operated Ca2+ entry:local activation of CRAC channels by STIM1 at ER-plasma membrane junctions. J. Cell Biol. 174:815–25
95. Luik RM, Wang B, Prakriya M, Wu MM, Lewis RS. 2008. Oligomerization of STIM1 couples ERcalcium depletion to CRAC channel activation. Nature 454:538–42
96. Penna A, Demuro A, Yeromin AV, Zhang SL, Safrina O, et al. 2008. The CRAC channel consists of atetramer formed by Stim-induced dimerization of Orai dimers. Nature 456:116–20
97. Nolz JC, Gomez TS, Zhu P, Li S, Medeiros RB, et al. 2006. The WAVE2 complex regulates actincytoskeletal reorganization and CRAC-mediated calcium entry during T cell activation. Curr. Biol. 16:24–34
98. Nolz JC, Medeiros RB, Mitchell JS, Zhu P, Freedman BD, et al. 2007. WAVE2 regulates high-affinityintegrin binding by recruiting vinculin and talin to the immunological synapse. Mol. Cell. Biol. 27:5986–6000
99. Savignac M, Mellstrom B, Naranjo JR. 2007. Calcium-dependent transcription of cytokine genes in Tlymphocytes. Pflugers Arch. 454:523–33
100. Macian F, Garcia-Cozar F, Im SH, Horton HF, Byrne MC, Rao A. 2002. Transcriptional mechanismsunderlying lymphocyte tolerance. Cell 109:719–31
101. Bandyopadhyay S, Soto-Nieves N, Macian F. 2007. Transcriptional regulation of T cell tolerance. Semin.Immunol. 19:180–87
102. Wu Y, Borde M, Heissmeyer V, Feuerer M, Lapan AD, et al. 2006. FOXP3 controls regulatory T cellfunction through cooperation with NFAT. Cell 126:375–87
103. Dolmetsch RE, Lewis RS, Goodnow CC, Healy JI. 1997. Differential activation of transcription factorsinduced by Ca2+ response amplitude and duration. Nature 386:855–58
104. Burkhardt JK, Carrizosa E, Shaffer MH. 2008. The actin cytoskeleton in T cell activation. Annu. Rev.Immunol. 26:233–59
105. Henney CS, Bubbers JE. 1973. Antigen-T lymphocyte interactions: inhibition by cytochalasin B.J. Immunol. 111:85–90
106. Holsinger LJ, Graef IA, Swat W, Chi T, Bautista DM, et al. 1998. Defects in actin-cap formation inVav-deficient mice implicate an actin requirement for lymphocyte signal transduction. Curr. Biol. 8:563–72
107. Jacobelli J, Chmura SA, Buxton DB, Davis MM, Krummel MF. 2004. A single class II myosin modulatesT cell motility and stopping, but not synapse formation. Nat. Immunol. 5:531–38
108. Faure S, Salazar-Fontana LI, Semichon M, Tybulewicz VL, Bismuth G, et al. 2004. ERM proteinsregulate cytoskeleton relaxation promoting T cell-APC conjugation. Nat. Immunol. 5:272–79
109. Gomez TS, Kumar K, Medeiros RB, Shimizu Y, Leibson PJ, Billadeau DD. 2007. Formins regulatethe actin-related protein 2/3 complex-independent polarization of the centrosome to the immunologicalsynapse. Immunity 26:177–90
110. Zeng R, Cannon JL, Abraham RT, Way M, Billadeau DD, et al. 2003. SLP-76 coordinates Nck-dependent Wiskott-Aldrich syndrome protein recruitment with Vav-1/Cdc42-dependent Wiskott-Aldrich syndrome protein activation at the T cell-APC contact site. J. Immunol. 171:1360–68
111. Gomez TS, Hamann MJ, McCarney S, Savoy DN, Lubking CM, et al. 2005. Dynamin 2 regulates T cellactivation by controlling actin polymerization at the immunological synapse. Nat. Immunol. 6:261–70
112. Kupfer A, Swain SL, Singer SJ. 1987. The specific direct interaction of helper T cells and antigen-presenting B cells. II. Reorientation of the microtubule organizing center and reorganization of themembrane-associated cytoskeleton inside the bound helper T cells. J. Exp. Med. 165:1565–80
113. Combs J, Kim SJ, Tan S, Ligon LA, Holzbaur EL, et al. 2006. Recruitment of dynein to the Jurkatimmunological synapse. Proc. Natl. Acad. Sci. USA 103:14883–88
616 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
114. Monks CR, Freiberg BA, Kupfer H, Sciaky N, Kupfer A. 1998. Three-dimensional segregation ofsupramolecular activation clusters in T cells. Nature 395:82–86
115. Grakoui A, Bromley SK, Sumen C, Davis MM, Shaw AS, et al. 1999. The immunological synapse: amolecular machine controlling T cell activation. Science 285:221–27
116. Lee KH, Holdorf AD, Dustin ML, Chan AC, Allen PM, Shaw AS. 2002. T cell receptor signalingprecedes immunological synapse formation. Science 295:1539–42
117. Cemerski S, Das J, Giurisato E, Markiewicz MA, Allen PM, Chakraborty AK, Shaw AS. 2008. Thebalance between T cell receptor signaling and degradation at the center of the immunological synapseis determined by antigen quality. Immunity 29:414–22
118. Yokosuka T, Kobayashi W, Sakata-Sogawa K, Takamatsu M, Hashimoto-Tane A, et al. 2008. Spa-tiotemporal regulation of T cell costimulation by TCR-CD28 microclusters and protein kinase Cθ
translocation. Immunity 29:589–601119. Dustin ML. 2008. T-cell activation through immunological synapses and kinapses. Immunol. Rev. 221:77–
89120. Bunnell SC, Hong DI, Kardon JR, Yamazaki T, McGlade CJ, et al. 2002. T cell receptor ligation induces
the formation of dynamically regulated signaling assemblies. J. Cell Biol. 158:1263–75121. Demonstrated thatperipheralmicroclusters, notcSMAC, are the sites ofboth TCR signalinitiation andmaintenance.
121. Yokosuka T, Sakata-Sogawa K, Kobayashi W, Hiroshima M, Hashimoto-Tane A, et al. 2005.Newly generated T cell receptor microclusters initiate and sustain T cell activation by recruit-ment of Zap70 and SLP-76. Nat. Immunol. 6:1253–62
122. Varma R, Campi G, Yokosuka T, Saito T, Dustin ML. 2006. T cell receptor-proximal signals are sustainedin peripheral microclusters and terminated in the central supramolecular activation cluster. Immunity25:117–27
123. Campi G, Varma R, Dustin ML. 2005. Actin and agonist MHC-peptide complex-dependent T cellreceptor microclusters as scaffolds for signaling. J. Exp. Med. 202:1031–36
124. Nguyen K, Sylvain NR, Bunnell SC. 2008. T cell costimulation via the integrin VLA-4 inhibits the actin-dependent centralization of signaling microclusters containing the adaptor SLP-76. Immunity 28:810–21
125. Chang JT, Palanivel VR, Kinjyo I, Schambach F, Intlekofer AM, et al. 2007. Asymmetric T lymphocytedivision in the initiation of adaptive immune responses. Science 315:1687–91
126. Allenspach EJ, Cullinan P, Tong J, Tang Q, Tesciuba AG, et al. 2001. ERM-dependent movement ofCD43 defines a novel protein complex distal to the immunological synapse. Immunity 15:739–50
127. Zhang W, Irvin BJ, Trible RP, Abraham RT, Samelson LE. 1999. Functional analysis of LAT in TCR-mediated signaling pathways using a LAT-deficient Jurkat cell line. Int. Immunol. 11:943–50
128. Billadeau DD, Nolz JC, Gomez TS. 2007. Regulation of T-cell activation by the cytoskeleton. Nat. Rev.Immunol. 7:131–43
129. Menasche G, Kliche S, Bezman N, Schraven B. 2007. Regulation of T-cell antigen receptor-mediatedinside-out signaling by cytosolic adapter proteins and Rap1 effector molecules. Immunol. Rev. 218:82–91
130. Katagiri K, Hattori M, Minato N, Irie S, Takatsu K, Kinashi T. 2000. Rap1 is a potent activation signal forleukocyte function-associated antigen 1 distinct from protein kinase C and phosphatidylinositol-3-OHkinase. Mol. Cell. Biol. 20:1956–69
131. Sebzda E, Bracke M, Tugal T, Hogg N, Cantrell DA. 2002. Rap1A positively regulates T cells via integrinactivation rather than inhibiting lymphocyte signaling. Nat. Immunol. 3:251–58
132. Duchniewicz M, Zemojtel T, Kolanczyk M, Grossmann S, Scheele JS, Zwartkruis FJ. 2006. Rap1A-deficient T and B cells show impaired integrin-mediated cell adhesion. Mol. Cell. Biol. 26:643–53
133. Peterson EJ, Woods ML, Dmowski SA, Derimanov G, Jordan MS, et al. 2001. Coupling of the TCR tointegrin activation by Slap-130/Fyb. Science 293:2263–65
134. Kliche S, Breitling D, Togni M, Pusch R, Heuer K, et al. 2006. The ADAP/SKAP55 signaling moduleregulates T-cell receptor-mediated integrin activation through plasma membrane targeting of Rap1. Mol.Cell. Biol. 26:7130–44
135. Wang H, McCann FE, Gordan JD, Wu X, Raab M, et al. 2004. ADAP-SLP-76 binding differen-tially regulates supramolecular activation cluster (SMAC) formation relative to T cell-APC conjugation.J. Exp. Med. 200:1063–74
www.annualreviews.org • T Cell Activation 617
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
136. Menasche G, Kliche S, Chen EJ, Stradal TE, Schraven B, Koretzky G. 2007. RIAM links theADAP/SKAP-55 signaling module to Rap1, facilitating T-cell-receptor-mediated integrin activation.Mol. Cell Biol. 27:4070–81
137. Identified RIAM asa key adapter in Rap1plasma membranelocalization and integrinactivation in T cells.
137. Lafuente EM, van Puijenbroek AA, Krause M, Carman CV, Freeman GJ, et al. 2004. RIAM, anEna/VASP and profilin ligand, interacts with Rap1-GTP and mediates Rap1-induced adhesion.Dev. Cell 7:585–95
138. Medeiros RB, Dickey DM, Chung H, Quale AC, Nagarajan LR, et al. 2005. Protein kinase D1 and theβ1 integrin cytoplasmic domain control β1 integrin function via regulation of Rap1 activation. Immunity23:213–26
139. Katagiri K, Maeda A, Shimonaka M, Kinashi T. 2003. RAPL, a Rap1-binding molecule that mediatesRap1-induced adhesion through spatial regulation of LFA-1. Nat. Immunol. 4:741–48
140. Han J, Lim CJ, Watanabe N, Soriani A, Ratnikov B, et al. 2006. Reconstructing and deconstructingagonist-induced activation of integrin αIIbβ3. Curr. Biol. 16:1796–806
141. Acuto O, Michel F. 2003. CD28-mediated co-stimulation: a quantitative support for TCR signalling.Nat. Rev. Immunol. 3:939–51
142. Pages F, Ragueneau M, Rottapel R, Truneh A, Nunes J, et al. 1994. Binding of phosphatidylinositol-3-OH kinase to CD28 is required for T-cell signalling. Nature 369:327–29
143. Narayan P, Holt B, Tosti R, Kane LP. 2006. CARMA1 is required for Akt-mediated NF-κB activationin T cells. Mol. Cell. Biol. 26:2327–36
144. Qiao G, Li Z, Molinero L, Alegre ML, Ying H, et al. 2008. T-cell receptor-induced NFκB activation isnegatively regulated by E3 ubiquitin ligase Cbl-b. Mol. Cell. Biol. 28:2470–80
145. Beals CR, Sheridan CM, Turck CW, Gardner P, Crabtree GR. 1997. Nuclear export of NF-ATc enhancedby glycogen synthase kinase-3. Science 275:1930–34
146. Huang GN, Huso DL, Bouyain S, Tu J, McCorkell KA, et al. 2008. NFAT binding and regulation of Tcell activation by the cytoplasmic scaffolding Homer proteins. Science 319:476–81
147. CD28costimulation increasesglucose uptake andglycolysis, via aPI3K/Akt pathway, tolevels exceeding thatrequired for immediateuse.
147. Frauwirth KA, Riley JL, Harris MH, Parry RV, Rathmell JC, et al. 2002. The CD28 signalingpathway regulates glucose metabolism. Immunity 16:769–77
148. Jacobs SR, Herman CE, Maciver NJ, Wofford JA, Wieman HL, et al. 2008. Glucose uptake is limitingin T cell activation and requires CD28-mediated Akt-dependent and independent pathways. J. Immunol.180:4476–86
149. Heyeck SD, Wilcox HM, Bunnell SC, Berg LJ. 1997. Lck phosphorylates the activation loop tyrosineof the Itk kinase domain and activates Itk kinase activity. J. Biol. Chem. 272:25401–8
150. Li CR, Berg LJ. 2005. Itk is not essential for CD28 signaling in naive T cells. J. Immunol. 174:4475–79151. Burr JS, Savage ND, Messah GE, Kimzey SL, Shaw AS, et al. 2001. Cutting edge: distinct motifs within
CD28 regulate T cell proliferation and induction of Bcl-XL. J. Immunol. 166:5331–35152. Blanchet F, Cardona A, Letimier FA, Hershfield MS, Acuto O. 2005. CD28 costimulatory signal induces
protein arginine methylation in T cells. J. Exp. Med. 202:371–77153. Bachmaier K, Pummerer C, Shahinian A, Ionescu J, Neu N, et al. 1996. Induction of autoimmunity in
the absence of CD28 costimulation. J. Immunol. 157:1752–57154. Hutloff A, Dittrich AM, Beier KC, Eljaschewitsch B, Kraft R, et al. 1999. ICOS is an inducible T-cell
co-stimulator structurally and functionally related to CD28. Nature 397:263–66155. Coyle AJ, Lehar S, Lloyd C, Tian J, Delaney T, et al. 2000. The CD28-related molecule ICOS is required
for effective T cell-dependent immune responses. Immunity 13:95–105156. Arimura Y, Kato H, Dianzani U, Okamoto T, Kamekura S, et al. 2002. A co-stimulatory molecule
on activated T cells, H4/ICOS, delivers specific signals in Th cells and regulates their responses. Int.Immunol. 14:555–66
157. Harada Y, Ohgai D, Watanabe R, Okano K, Koiwai O, et al. 2003. A single amino acid alterationin cytoplasmic domain determines IL-2 promoter activation by ligation of CD28 but not induciblecostimulator (ICOS). J. Exp. Med. 197:257–62
158. Suh WK, Tafuri A, Berg-Brown NN, Shahinian A, Plyte S, et al. 2004. The inducible costimulatorplays the major costimulatory role in humoral immune responses in the absence of CD28. J. Immunol.172:5917–23
618 Smith-Garvin · Koretzky · Jordan
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

ANRV371-IY27-21 ARI 16 February 2009 13:31
159. Watts TH. 2005. TNF/TNFR family members in costimulation of T cell responses. Annu. Rev. Immunol.23:23–68
160. Chung JY, Park YC, Ye H, Wu H. 2002. All TRAFs are not created equal: common and distinct molecularmechanisms of TRAF-mediated signal transduction. J. Cell Sci. 115:679–88
161. So T, Lee SW, Croft M. 2008. Immune regulation and control of regulatory T cells by OX40 and 4-1BB.Cytokine Growth Factor Rev. 19:253–62
162. Vang T, Abrahamsen H, Myklebust S, Enserink J, Prydz H, et al. 2004. Knockdown of C-terminal Srckinase by siRNA-mediated RNA interference augments T cell receptor signaling in mature T cells.Eur. J. Immunol. 34:2191–99
163. Hermiston ML, Xu Z, Weiss A. 2003. CD45: a critical regulator of signaling thresholds in immune cells.Annu. Rev. Immunol. 21:107–37
164. Stefanova I, Hemmer B, Vergelli M, Martin R, Biddison WE, Germain RN. 2003. TCR ligand discrim-ination is enforced by competing ERK positive and SHP-1 negative feedback pathways. Nat. Immunol.4:248–54
165. Tsui HW, Siminovitch KA, de Souza L, Tsui FW. 1993. Motheaten and viable motheaten mice havemutations in the haematopoietic cell phosphatase gene. Nat. Genet. 4:124–29
166. Nagaishi T, Pao L, Lin SH, Iijima H, Kaser A, et al. 2006. SHP1 phosphatase-dependent T cell inhibitionby CEACAM1 adhesion molecule isoforms. Immunity 25:769–81
167. Yasuda T, Bundo K, Hino A, Honda K, Inoue A, et al. 2007. Dok-1 and Dok-2 are negative regulatorsof T cell receptor signaling. Int. Immunol. 19:487–95
168. Dong S, Corre B, Foulon E, Dufour E, Veillette A, et al. 2006. T cell receptor for antigen induces linkerfor activation of T cell-dependent activation of a negative signaling complex involving Dok-2, SHIP-1,and Grb-2. J. Exp. Med. 203:2509–18
169. Shui JW, Boomer JS, Han J, Xu J, Dement GA, et al. 2007. Hematopoietic progenitor kinase 1 negativelyregulates T cell receptor signaling and T cell-mediated immune responses. Nat. Immunol. 8:84–91
170. Di Bartolo V, Montagne B, Salek M, Jungwirth B, Carrette F, et al. 2007. A novel pathway down-modulating T cell activation involves HPK-1-dependent recruitment of 14-3-3 proteins on SLP-76.J. Exp. Med. 204:681–91
171. Carpino N, Turner S, Mekala D, Takahashi Y, Zang H, et al. 2004. Regulation of ZAP-70 activation andTCR signaling by two related proteins, Sts-1 and Sts-2. Immunity 20:37–46
172. Mikhailik A, Ford B, Keller J, Chen Y, Nassar N, Carpino N. 2007. A phosphatase activity of Sts-1contributes to the suppression of TCR signaling. Mol. Cell 27:486–97
173. Agrawal R, Carpino N, Tsygankov A. 2008. TULA proteins regulate activity of the protein tyrosinekinase Syk. J. Cell. Biochem. 104:953–64
174. Feshchenko EA, Smirnova EV, Swaminathan G, Teckchandani AM, Agrawal R, et al. 2004. TULA: anSH3- and UBA-containing protein that binds to c-Cbl and ubiquitin. Oncogene 23:4690–706
175–177. Three papersdemonstratingautoimmunity inCbl−/− mice andidentifying Cbl familymembers as criticalnegative regulators of Tcell activation.
175. Bachmaier K, Krawczyk C, Kozieradzki I, Kong YY, Sasaki T, et al. 2000. Negative regulationof lymphocyte activation and autoimmunity by the molecular adaptor Cbl-b. Nature 403:211–16
176. Naramura M, Jang IK, Kole H, Huang F, Haines D, Gu H. 2002. c-Cbl and Cbl-b regulate T cellresponsiveness by promoting ligand-induced TCR down-modulation. Nat. Immunol. 3:1192–99
177. Murphy MA, Schnall RG, Venter DJ, Barnett L, Bertoncello I, et al. 1998. Tissue hyperplasiaand enhanced T-cell signalling via ZAP-70 in c-Cbl-deficient mice. Mol. Cell. Biol. 18:4872–82
178. Balagopalan L, Barr VA, Sommers CL, Barda-Saad M, Goyal A, et al. 2007. c-Cbl-mediated regulationof LAT-nucleated signaling complexes. Mol. Cell. Biol. 27:8622–36
179. Waterhouse P, Penninger JM, Timms E, Wakeham A, Shahinian A, et al. 1995. Lymphoproliferativedisorders with early lethality in mice deficient in Ctla-4. Science 270:985–88
180. Nishimura H, Okazaki T, Tanaka Y, Nakatani K, Hara M, et al. 2001. Autoimmune dilated cardiomy-opathy in PD-1 receptor-deficient mice. Science 291:319–22
181. Chemnitz JM, Parry RV, Nichols KE, June CH, Riley JL. 2004. SHP-1 and SHP-2 associate withimmunoreceptor tyrosine-based switch motif of programmed death 1 upon primary human T cell stim-ulation, but only receptor ligation prevents T cell activation. J. Immunol. 173:945–54
182. Teft WA, Kirchhof MG, Madrenas J. 2006. A molecular perspective of CTLA-4 function. Annu. Rev.Immunol. 24:65–97
www.annualreviews.org • T Cell Activation 619
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

AR371-FM ARI 16 February 2009 15:37
Annual Review ofImmunology
Volume 27, 2009Contents
FrontispieceMarc Feldmann � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � x
Translating Molecular Insights in Autoimmunity into EffectiveTherapyMarc Feldmann � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �1
Structural Biology of Shared Cytokine ReceptorsXinquan Wang, Patrick Lupardus, Sherry L. LaPorte,and K. Christopher Garcia � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 29
Immunity to Respiratory VirusesJacob E. Kohlmeier and David L. Woodland � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 61
Immune Therapy for CancerMichael Dougan and Glenn Dranoff � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 83
Microglial Physiology: Unique Stimuli, Specialized ResponsesRichard M. Ransohoff and V. Hugh Perry � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �119
The Liver as a Lymphoid OrganIan Nicholas Crispe � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �147
Immune and Inflammatory Mechanisms of AtherosclerosisElena Galkina and Klaus Ley � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �165
Primary B Cell Immunodeficiencies: Comparisons and ContrastsMary Ellen Conley, A. Kerry Dobbs, Dana M. Farmer, Sebnem Kilic,Kenneth Paris, Sofia Grigoriadou, Elaine Coustan-Smith, Vanessa Howard,and Dario Campana � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �199
The Inflammasomes: Guardians of the BodyFabio Martinon, Annick Mayor, and Jürg Tschopp � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �229
Human Marginal Zone B CellsJean-Claude Weill, Sandra Weller, and Claude-Agnes Reynaud � � � � � � � � � � � � � � � � � � � � � �267
v
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.

AR371-FM ARI 16 February 2009 15:37
AireDiane Mathis and Christophe Benoist � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �287
Regulatory Lymphocytes and Intestinal InflammationAna Izcue, Janine L. Coombes, and Fiona Powrie � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �313
The Ins and Outs of Leukocyte Integrin SignalingClare L. Abram and Clifford A. Lowell � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �339
Recent Advances in the Genetics of Autoimmune DiseasePeter K. Gregersen and Lina M. Olsson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �363
Cell-Mediated Immune Responses in TuberculosisAndrea M. Cooper � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �393
Enhancing Immunity Through AutophagyChristian Munz � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �423
Alternative Activation of Macrophages: An Immunologic FunctionalPerspectiveFernando O. Martinez, Laura Helming, and Siamon Gordon � � � � � � � � � � � � � � � � � � � � � � � �451
IL-17 and Th17 CellsThomas Korn, Estelle Bettelli, Mohamed Oukka, and Vijay K. Kuchroo � � � � � � � � � � � � � �485
Immunological and Inflammatory Functions of the Interleukin-1FamilyCharles A. Dinarello � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �519
Regulatory T Cells in the Control of Host-Microorganism InteractionsYasmine Belkaid and Kristin Tarbell � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �551
T Cell ActivationJennifer E. Smith-Garvin, Gary A. Koretzky, and Martha S. Jordan � � � � � � � � � � � � � � �591
Horror Autoinflammaticus: The Molecular Pathophysiology ofAutoinflammatory DiseaseSeth L. Masters, Anna Simon, Ivona Aksentijevich, and Daniel L. Kastner � � � � � � � � �621
Blood Monocytes: Development, Heterogeneity, and Relationshipwith Dendritic CellsCedric Auffray, Michael H. Sieweke, and Frederic Geissmann � � � � � � � � � � � � � � � � � � � � � � � �669
Regulation and Function of NF-κB Transcription Factors in theImmune SystemSivakumar Vallabhapurapu and Michael Karin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �693
vi Contents
Ann
u. R
ev. I
mm
unol
. 200
9.27
:591
-619
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Ube
rlan
dia
on 0
4/26
/12.
For
per
sona
l use
onl
y.
Related Documents




![Artigo Limoeiro[1]](https://static.cupdf.com/doc/110x72/54938845b479595b4d8b4881/artigo-limoeiro1.jpg)






