european journal of pharmaceutical sciences 33 ( 2 0 0 8 ) 399–408 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/ejps Articaine interaction with DSPC bilayer: A 13 C and 31 P solid-state NMR study Chen Song a , Henning Lygre b , Willy Nerdal a,∗ a Department of Chemistry, University of Bergen, All´ egaten 41, N-5007 Bergen, Norway b Department of Dental Science–Dental Pharmacology, University of Bergen, Armauer Hansens hus, Haukelandsv. 28, N-5021 Bergen, Norway article info Article history: Received 5 July 2007 Received in revised form 4 January 2008 Accepted 22 January 2008 Published on line 2 February 2008 Keywords: 13 C MASNMR 31 P NMR Bilayers Articaine bilayer interaction abstract Articaine hydrochloride, 4-methyl-3-(2-[propylamino]propionamido)-2-thiophenecarboxy- lic acid, methyl ester hydrochloride, is a local anaesthetic commonly used in dentistry, and is classified as an amide local anaesthetic. Solid-state 13 C and 31 P NMR were used to investigate the uncharged articaine species (sample pH 10.0) when interacting with distearoylphos- phatidylcholine (DSPC) model membranes. The DSPC phospholipid bilayer was studied at four different molar ratios of articaine, 10, 25, 40, and 55mol%, respectively. The articaine concentration-dependent decrease in the DSPC bilayer gel-to-liquid-crystalline phase- transition temperature demonstrates substantial articaine interaction with this bilayer. A DSPC bilayer contains a large hydrophobic core and the 13 C and 31 P NMR spectra of the 40 mol% articaine-containing sample demonstrate a disturbance in the molecular packing of the polar bilayer region that extends into the hydrophobic region, evidenced by carbon 2 and 3 of the stearoyl acyl chains. Observed 31 P and 13 C NMR spectral changes when articaine is increased from 40 to 55 mol%, suggest formation of articaine aggregates and decrease in DSPC bilayer perturbation at the latter articaine level. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Local anesthetics are used to prevent generation of the nerve impulse by their binding to the voltage-gated sodium chan- nel and blocking the inward sodium transport that creates the action potential of axons (Butterworth and Strichartz, 1990). Other membrane proteins are also targets of local anes- thetics (Papahadjoupoulos et al., 1975; Tanaka and Hidaka, 1981; Vanderkooi and Adade, 1986; Arias et al., 1990; Br¨ au et al., 1995; Hollman et al., 2001) and the packing of phospho- lipids in biological bilayers is perturbed by these compounds This work was supported by a grant from the Norwegian Research Council (NFR). ∗ Corresponding author. Tel.: +47 55 583353; fax: +47 55 589400. E-mail addresses: [email protected] (C. Song), [email protected] (H. Lygre), [email protected] (W. Nerdal). Abbreviations: DMPC, 1,2-dimyristoyl-sn-glycerol-3-phosphatidylcholine; DSPC, 1,2-distearoyl-sn-glycerol-3-phosphatidylcholine; PC, phosphatidylcholine; PS, phosphatidylserine; MAS, magic-angle spinning; NMR, nuclear magnetic resonance; CSA, chemical shift anisotropy; NOE, nuclear overhauser effect. (Boulanger et al., 1981; Auger et al., 1990; de Paula and Schreier, 1995; Pinto et al., 2000) with a possible change in membrane protein structure and function. Thus, perturba- tion of phospholipid packing in bilayers is intimately tied to a compound’s anesthetic effect. In general, depression of bilayer gel-to-liquid-crystalline phase-transition temperature by local anesthetics has been found to correlate with the hydrophobicity of the anesthetic (Gupta, 1991; Hata et al., 2000a). The hemolytic effect of several local anesthetics on rat erythrocytes has been found to be greatly influenced by the uncharged species, even at a pH of 7.4, where there is a pre- 0928-0987/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.ejps.2008.01.010

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AA
Ca
b
H
a
A
R
R
4
A
P
K1
3
B
A
1
Lint1t1al
pa0d
e u r o p e a n j o u r n a l o f p h a r m a c e u t i c a l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408
avai lab le at www.sc iencedi rec t .com
journa l homepage: www.e lsev ier .com/ locate /e jps
rticaine interaction with DSPC bilayer:13C and 31P solid-state NMR study�
hen Songa, Henning Lygreb, Willy Nerdala,∗
Department of Chemistry, University of Bergen, Allegaten 41, N-5007 Bergen, NorwayDepartment of Dental Science–Dental Pharmacology, University of Bergen, Armauer Hansens hus,aukelandsv. 28, N-5021 Bergen, Norway
r t i c l e i n f o
rticle history:
eceived 5 July 2007
eceived in revised form
January 2008
ccepted 22 January 2008
ublished on line 2 February 2008
eywords:3C MASNMR
a b s t r a c t
Articaine hydrochloride, 4-methyl-3-(2-[propylamino]propionamido)-2-thiophenecarboxy-
lic acid, methyl ester hydrochloride, is a local anaesthetic commonly used in dentistry, and is
classified as an amide local anaesthetic. Solid-state 13C and 31P NMR were used to investigate
the uncharged articaine species (sample pH 10.0) when interacting with distearoylphos-
phatidylcholine (DSPC) model membranes. The DSPC phospholipid bilayer was studied at
four different molar ratios of articaine, 10, 25, 40, and 55 mol%, respectively. The articaine
concentration-dependent decrease in the DSPC bilayer gel-to-liquid-crystalline phase-
transition temperature demonstrates substantial articaine interaction with this bilayer.
A DSPC bilayer contains a large hydrophobic core and the 13C and 31P NMR spectra of the
1P NMRilayers
rticaine bilayer interaction
40 mol% articaine-containing sample demonstrate a disturbance in the molecular packing
of the polar bilayer region that extends into the hydrophobic region, evidenced by carbon 2
and 3 of the stearoyl acyl chains. Observed 31P and 13C NMR spectral changes when articaine
is increased from 40 to 55 mol%, suggest formation of articaine aggregates and decrease in
ation
hydrophobicity of the anesthetic (Gupta, 1991; Hata et al.,
DSPC bilayer perturb
. Introduction
ocal anesthetics are used to prevent generation of the nervempulse by their binding to the voltage-gated sodium chan-el and blocking the inward sodium transport that createshe action potential of axons (Butterworth and Strichartz,990). Other membrane proteins are also targets of local anes-hetics (Papahadjoupoulos et al., 1975; Tanaka and Hidaka,
981; Vanderkooi and Adade, 1986; Arias et al., 1990; Brau etl., 1995; Hollman et al., 2001) and the packing of phospho-ipids in biological bilayers is perturbed by these compounds� This work was supported by a grant from the Norwegian Research C∗ Corresponding author. Tel.: +47 55 583353; fax: +47 55 589400.
E-mail addresses: [email protected] (C. Song), Henning.Lygre@oAbbreviations: DMPC, 1,2-dimyristoyl-sn-glycerol-3-phosphatidylcho
hosphatidylcholine; PS, phosphatidylserine; MAS, magic-angle spinnisotropy; NOE, nuclear overhauser effect.928-0987/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.ejps.2008.01.010
at the latter articaine level.
© 2008 Elsevier B.V. All rights reserved.
(Boulanger et al., 1981; Auger et al., 1990; de Paula andSchreier, 1995; Pinto et al., 2000) with a possible change inmembrane protein structure and function. Thus, perturba-tion of phospholipid packing in bilayers is intimately tiedto a compound’s anesthetic effect. In general, depression ofbilayer gel-to-liquid-crystalline phase-transition temperatureby local anesthetics has been found to correlate with the
ouncil (NFR).
dont.uib.no (H. Lygre), [email protected] (W. Nerdal).line; DSPC, 1,2-distearoyl-sn-glycerol-3-phosphatidylcholine; PC,ning; NMR, nuclear magnetic resonance; CSA, chemical shift
2000a). The hemolytic effect of several local anesthetics on raterythrocytes has been found to be greatly influenced by theuncharged species, even at a pH of 7.4, where there is a pre-

400 e u r o p e a n j o u r n a l o f p h a r m a c e u t i c
Scheme 1 – Positively charged articaine hydrochloride.
dominance of the positively charged species, indicating thatthe disrupting effect on the erythrocyte membranes is mainlydue to hydrophobic interactions (Malheiros et al., 2004).
Articaine hydrochloride, 4-methyl-3-(2-[propylamino]propionamido)-2-thiophenecarboxylic acid, methyl esterhydrochloride (Scheme 1), is a local anesthetic commonlyused in dentistry (Malamed et al., 2000a, 2001), and is classi-fied as an amide local anesthetic. However, unlike many otherlocal anesthetics, a thiophene group increases articaine’sliposolubility (Malamed et al., 2000b), and ability to penetratethe nerve sheath. In contrast to most local anesthetics,articaine contains an ester linkage that causes articaine to behydrolyzed by plasma esterase (Vree et al., 1988; Grossmannet al., 2004) to articainic acid. In studies of local anestheticsinteracting with model membranes, several methods havebeen employed, such as fluorescence spectroscopy (Westmanet al., 1982; Fraceto et al., 2002), infrared spectroscopy (Fracetoet al., 2002; Suwalsky et al., 2002), X-ray diffraction (Costeret al., 1981; Suwalsky et al., 2002, 2005), electron spin res-onance (Neal et al., 1976; Pinto et al., 2000; Fraceto et al.,2002), ultraviolet light absorption spectroscopy (Westmanet al., 1982) and nuclear magnetic resonance (Boulangeret al., 1981; Kelusky and Smith, 1984; Yoshino et al., 1998;Fraceto et al., 2002, 2005). Here, we employ solid-state 13C(Wu and Chi, 1990) and 31P NMR (Browning and Seelig, 1980)to study the uncharged articaine species interacting withdistearoylphosphatidylcholine (DSPC) model membranes.The DSPC bilayer samples subjected to NMR experimentswere adjusted to a pH of 10.0 where more than 99% of thearticaine molecules are in the uncharged form (pKa of 7.8(Malamed et al., 2000a,b)). In the uncharged form, articaine ismore hydrophobic (Butterworth and Strichartz, 1990; Pinto etal., 2000) and more potent (Kaneshina et al., 1997; Hata et al.,2000b) for phospholipid bilayer interaction than the chargedspecies.
The hydrophobicity of articaine, and thereby its potentialfor interaction with the hydrophobic part of the phospho-lipid bilayer (articaine’s lipophilicity) can be evaluated bythe corresponding octanol/water partition coefficient. Anoctanol/water partition coefficient of 257 has been found for
articaine and this high articaine preference for octanol overwater was furthermore found to correlate well with arti-caine binding to isolated guinea pig atria tissue (Lullmanet al., 1980). Thus, more than 99% of the articaine addeda l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408
to the DSPC phospholipid samples used in this study willprefer the hydrophobic environment provided by the phos-pholipid bilayer and only a negligible fraction will prefer thewater phase. An amphiphilic compound like articaine will,depending on pH, have an ionic or nonionic polar moietyand a hydrophobic part. In an aqueous medium, amphiphilicmolecules are able to organize themselves as micelles, bilay-ers, monolayers, hexagonal or cubic phases. Thus, it is likelythat at certain conditions, e.g. concentration, an amphiphilelike articaine can form self-aggregated structures. This hasbeen observed for other local anesthetics like dibucaine andtetracaine (Schreier et al., 2000), where the critical micelleconcentrations (CMC) were found to be 0.066 and 0.130 M,respectively.
In the present study, the DSPC phospholipid bilayer wasprepared at four different molar ratios of articaine, 10, 25, 40,and 55 mol%, respectively. In the clinic, a 4% articaine solu-tion is used (Malamed et al., 2000a,b), and both the water/lipidpartitioning coefficient of local anesthetics (Malheiros et al.,2004) and the amount of bilayers in myelinized nervous fibersensure that the injected anesthetic will reside in the bilayer.In addition to study the interaction of articaine with the DSPCbilayer, the experiments can also provide information aboutuncharged articaine self-association when interacting withthe DSPC bilayer (de Verteuil et al., 1981; Garcia-Soto andFernandez, 1983).
Static 31P NMR of phospholipid bilayers (lamellar) gives thechemical shielding anisotropy as the difference between thehigh ppm and the low ppm edges of the resonance due torapid phospholipid motion around an axis perpendicular tothe bilayer surface (Seelig, 1978). In comparison, the powderpattern of an inverted hexagonal lipid phase has only halfthe width and the opposite sign than the powder pattern ofthe corresponding bilayer phase (Wenk and Seelig, 1998). Thisdemonstrates that the 31P static chemical shielding tensor issensitive to the charge distribution at the phosphorous atom.For instance, unsaturated 3-sn-phosphatidylethanolaminesare likely to undergo the lamellar to hexagonal phase tran-sition at higher temperatures and this can be followed by31P NMR. Biological membranes contain regions of liquid-crystalline phospholipids in a bilayer environment and the 31Pchemical shielding anisotropy suggests that the motions ofthe phosphate segments are probably similar in most mem-branes (Cullis and McLaughlin, 1977). Furthermore, the 31P T1
relaxation time in phospholipid vesicles has been found to bedominated by dipolar interactions (Yeagle et al., 1977).
2. Experimental
2.1. Sample preparation: pure DSPC and DSPC with(10, 25, 40, and 55 mol%) articaine
Articaine hydrochloride was obtained from Septodont (Saint-Maur-des-Fosses, France), and synthetic 1,2-distearoyl-sn-glycerol-3-phosphatidylcholine (DSPC, powder) was pur-
chased from Sigma Chemical Co. (St. Louis, MO, USA) andused without further purification. The desired amount of DSPCwas dissolved in spectroscopic grade chloroform and thenlyophilized to dryness, and the DSPC bilayer was kept under an
t i c a
apwiawmpssswsctopbse(a
patt4wDso7f2a(tmwpp
2
TowwaEswtoceamo
e u r o p e a n j o u r n a l o f p h a r m a c e u
rgon atmosphere and not exposed to air and light. The sam-le of dry powder was then suspended in degassed distilledater. Subsequently, the lipid suspension of DSPC was divided
nto eight equi-amount samples and to four, the appropriatemounts of articaine (dissolved in degassed distilled water)ere added to obtain four bilayer samples with an articaineolar ratio of 10, 25, 40 and 55 mol%, respectively. The sus-
ensions contained multilamellar liposomes and unilamellarystems were obtained by freeze–thawing nine times. Thisample preparation procedure gives large unilamellar lipo-omes of unequal size. However, in the final samples, bulkater is eliminated and the bilayer produced is found to both
table and without artifacts (Song et al., 2006) such as highurvature liposomes, i.e. small diameter liposomes that tendo change to larger diameter liposomes of higher stability. Inur experience, employing an additional step in the samplereparation procedure of extrusion through a (1000 nm) car-onate filter does not change or improve the subsequent NMRpectra, i.e. the sample at the time of data acquisition (Nerdalt al., 2000; Gjerde et al., 2004). In fact, some phospholipidsphosphatidylserines) tend to adhere to the carbonate filternd not pass the filter quantitatively (Gjerde et al., 2004).
At the freeze–thawing stage, one of the pure DSPC sam-les and all articaine-containing samples were adjusted topH of 10.0 by adding a small amount of 0.05 M NaOH, and
he remaining sample of the pure DSPC bilayer was adjustedo a pH of 7.4. Furthermore, two DSPC samples containing0 mol% articaine at sample pH of 5.0 and 7.4, respectively,ere prepared in a similar manner. Thus, four samples ofSPC and articaine HCl at a sample pH of 10.0, two DSPCamples with 40 mol% articaine at pH of 5.0 and at 7.4, werebtained as well as two pure DSPC samples of pH values of.4 and 10.0. All samples were equilibrated on an oil bathor 24 h at 60 ◦C. Subsequently, the samples were subjected to4 h of lyophilization giving partially hydrated liposomes withhydration level of ∼12 water molecules per lipid molecule
determined by 1H MASNMR). Water was added to each sampleo obtain fully hydrated bilayers (∼30 water molecules per lipid
olecule) (Small, 1967; Janiak et al., 1979) and the samplesere equilibrated at 60 ◦C for another 48 h (above the sam-les gel-to-liquid-crystalline transition temperature(s)) andacked into NMR rotors.
.2. 13C MASNMR spectroscopy
he 13C MASNMR experiments were obtained at 125.76 MHzn a Bruker 500 MHz Ultrashield Plus instrument equippedith magic-angle spinning hardware and using ZrO2 rotorsith a diameter of 4 mm. These rotors (and lids) are guar-
nteed by the manufacturer (Bruker) for use up to 50 ◦C.xperiments were done at a sample temperature of 44 ◦C withample spinning rate of 2 kHz. The experiments of this studyere carried out with high-power proton decoupling during
he acquisition, i.e. without NOE and with a relaxation delayf 6 s between transients. Acquisition of 16,384 transients wasarried out in each experiment, with the exception of the
xperiments on pure articaine, where 12,288 transients werecquired in each. The fids were subjected to Fourier transfor-ation without the use of an exponential window function inrder to optimize spectral resolution.
l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408 401
2.3. 31P NMR spectroscopy
31P experiments were carried out on a Bruker 500 MHz Ultra-shield Plus instrument and on a Bruker AVANCE DMX 400instrument. Magic-angle spinning 31P experiments were car-ried out at 161.98 MHz with a rotor spinning rate of 1.5, 2,and 3 kHz, respectively. Typically, 1024 transients with a relax-ation delay of 4 s between transients were accumulated at 44and 57 ◦C, and Fourier transformed without apodization inorder to optimize spectral resolution. The majority of the static31P spectra were carried out at 161.98 MHz with 10,240 tran-sients and a relaxation delay of 5 s between transients. Theseexperiments were acquired on samples with a pH of 10.0 attemperatures 42–57 ◦C. Static 31P experiments were acquiredat 202.46 MHz on DSPC bilayer with 40 mol% articaine at sam-ple temperatures of 42 and 52 ◦C, respectively. In these twolatter experiments the amount of sample was about 1/3 of theother samples, 12,288 transients were collected with a relax-ation delay of 5 s between transients and the probe-head wasa flat-coil (shown in Fig. 3, panel B). All 31P experiments wereacquired with proton high-power decoupling during acqui-sition, i.e. without NOE. The static 31P fids were multipliedwith an exponential window function increasing the linewidthby 50 Hz to reduce noise prior to Fourier transformation. The31P chemical shifts were referenced to 85% phosphoric acid(H3PO4, 0 ppm).
3. Results
Articaine has a pKa of 7.8, and consequently, at a pH of 7.4,two articaine species will be present, the positively chargedand the neutral species, at about 70 and 30%, respectively.Fig. 1a shows the 120–180 ppm region of a 13C MASNMR spec-trum of pure articaine (with 15 wt% D2O). Displayed in Fig. 1band c are the 120–180 ppm regions of 13C MASNMR spectra offully hydrated DSPC bilayer with 40 mol% articaine added, atsample pH of 7.4 and 5.0, respectively. All spectra of Fig. 1 areacquired at 44 ◦C, where the DSPC bilayer is in the gel state(DSPC has a Tm of 54 ◦C and will depend on pH). The aster-isks in Fig. 1c label articaine resonances have a pronouncedchange in chemical shifts upon increased sample acidity. Ina 13C MASNMR spectrum of dipalmitoylphosphatidylcholine(DMPC) bilayer at pH of 7.4 (data not shown) acquired at44 ◦C (above DMPCs Tm of 34 ◦C), the 70%/30% distributionwas demonstrated by two articaine resonances (with this peakintensity ratio) at 163.1 ppm. The articaine carbonyl (Scheme 1)will most likely be influenced by protonation of the amidenitrogen three covalent bonds away in the molecule andthis identifies the articaine carbonyl resonance (see Fig. 1a).Furthermore, the articaine carboxyl resonance can now beidentified to be the resonance at 170.2 ppm, see top spectrum.The left-most resonance in Fig. 1b and c, is the carbonyl reso-nance of DSPC. In order to study the interaction of the neutraland most potent articaine species with the DSPC bilayer, and atthe same time avoid drawbacks due to the presence of two arti-
caine species in the sample, a sample pH of 10.0 was chosenfor the articaine interaction study with the DSPC bilayer. Fig. 2(top), shows a 13C MASNMR spectrum of DSPC at pH of 7.4,note that the spectral region without resonances, 76–170 ppm,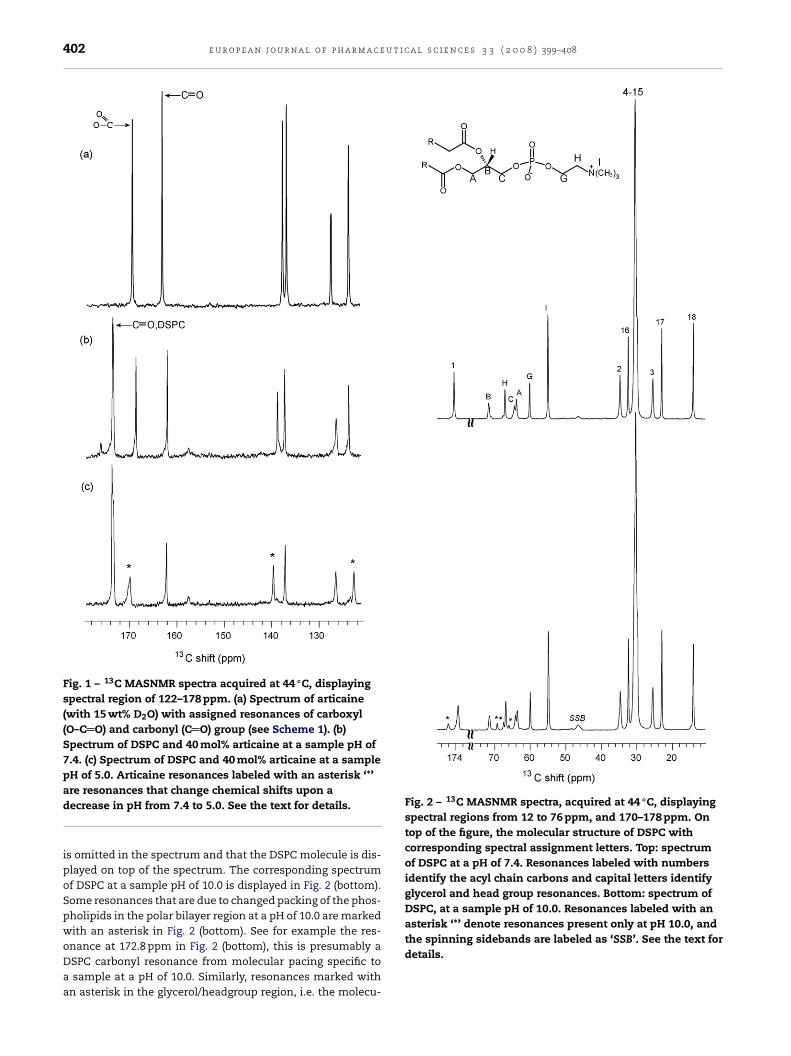
402 e u r o p e a n j o u r n a l o f p h a r m a c e u t i c a l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408
Fig. 1 – 13C MASNMR spectra acquired at 44 ◦C, displayingspectral region of 122–178 ppm. (a) Spectrum of articaine(with 15 wt% D2O) with assigned resonances of carboxyl(O–C O) and carbonyl (C O) group (see Scheme 1). (b)Spectrum of DSPC and 40 mol% articaine at a sample pH of7.4. (c) Spectrum of DSPC and 40 mol% articaine at a samplepH of 5.0. Articaine resonances labeled with an asterisk ‘*’
Fig. 2 – 13C MASNMR spectra, acquired at 44 ◦C, displayingspectral regions from 12 to 76 ppm, and 170–178 ppm. Ontop of the figure, the molecular structure of DSPC withcorresponding spectral assignment letters. Top: spectrumof DSPC at a pH of 7.4. Resonances labeled with numbersidentify the acyl chain carbons and capital letters identifyglycerol and head group resonances. Bottom: spectrum ofDSPC, at a sample pH of 10.0. Resonances labeled with anasterisk ‘*’ denote resonances present only at pH 10.0, and
are resonances that change chemical shifts upon adecrease in pH from 7.4 to 5.0. See the text for details.
is omitted in the spectrum and that the DSPC molecule is dis-played on top of the spectrum. The corresponding spectrumof DSPC at a sample pH of 10.0 is displayed in Fig. 2 (bottom).Some resonances that are due to changed packing of the phos-pholipids in the polar bilayer region at a pH of 10.0 are markedwith an asterisk in Fig. 2 (bottom). See for example the res-
onance at 172.8 ppm in Fig. 2 (bottom), this is presumably aDSPC carbonyl resonance from molecular pacing specific toa sample at a pH of 10.0. Similarly, resonances marked withan asterisk in the glycerol/headgroup region, i.e. the molecu-the spinning sidebands are labeled as ‘SSB’. See the text fordetails.

t i c a l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408 403
lprbdiNattoltstsa4ptDaaaawietemddpiaicaegtttapi(pm(lFTapaata
Fig. 3 – (Panel A) Static 31P NMR spectra acquired at asample pH of 10.0 at 42 ◦C (top row) and at 57 ◦C (bottomrow). The two spectra of the left column are of pure DSPC.The corresponding two spectra of the right column areacquired on DSPC with 40 mol% articaine. The asterisk ‘*’ inright column spectra denotes an isotropic phase resonance.(Panel B) Static 31P NMR spectra acquired at a sample pH of7.4 at 42 ◦C (top row) and at 57 ◦C (bottom row). The twospectra of the left column are of pure DSPC. The
e u r o p e a n j o u r n a l o f p h a r m a c e u
ar packing of a minor part of the DSPC molecules that takeslace at a pH of 10.0 are visible in the glycerol/headgroupegion. Whereas, the hydrophobic part of the bilayer, reportedy the resonances in the 14–36 ppm region displays no changesue to the high pH. A further documentation of alterations
n the polar region of the bilayer is found in the 31P staticMR spectra displayed in Fig. 3. In Fig. 3A, all four spectrare acquired on samples with a pH of 10.0. The two spec-ra in the top row are acquired at 42 ◦C (below Tm) and thewo spectra of the bottom row at 57 ◦C (above Tm). The CSAf the static 31P spectra shown in Fig. 3A are: 97 ppm in top
eft spectrum (pure DSPC at 42 ◦C), 80 ppm in top right spec-rum (DSPC/40 mol% articaine at 42 ◦C), 68 ppm in lower leftpectrum (pure DSPC at 57 ◦C) and 60 ppm in lower right spec-rum (DSPC/40 mol% articaine at 57 ◦C). Furthermore, the twopectra in the left column are acquired on pure DSPC bilayernd the two spectra in the right column on DSPC bilayer with0 mol% articaine. The corresponding spectra of samples at aH of 7.4 are displayed in Fig. 3B where the observed CSA ofhese static 31P spectra are: 97 ppm in top left spectrum (pureSPC at 42 ◦C), 95 ppm in top right spectrum (DSPC/40 mol%rticaine at 42 ◦C), 62 ppm in lower left spectrum (pure DSPCt 57 ◦C) and 60 ppm in lower right spectrum (DSPC/40 mol%rticaine at 57 ◦C). In Fig. 3B, the two spectra with 40 mol%rticaine are obtained using a flat-coil probe-head. The some-hat low signal intensity of the peak shoulder at a high ppm
n DSPC spectrum at 42 ◦C and with 40 mol% articaine can bexplained by lower signal to noise ratio (less sample, see Sec-ion 2) than the pure DSPC spectrum at 42 ◦C and possiblyffects of chemical shielding anisotropy being only approxi-ate below the phase transition because the applied proton
ecoupling power was insufficient to give a perfect protonecoupled spectrum (Browning and Seelig, 1980). The mostronounced reduction of the phospholipid phosphorous CSA
n presence of 40 mol% articaine is found at a sample pH of 10.0t 42 ◦C (below Tm) (Fig. 3A, top row). Thus, the correspondingncrease in mobility of the phospholipid phosphorous atomausing the reduction in the observed CSA is most pronouncedt a pH of 10.0 and 42 ◦C for a DSPC sample with 40 mol%lectrically neutral articaine. At a pH of 7.4, the bilayer headroup interaction of 40 mol% partially charged (∼70% posi-ively charged) articaine is not significant, as demonstrated byhe fairly similar CSA values of spectra shown in Fig. 3B. Fur-hermore, at a sample pH of 10.0 and in presence of 40 mol%rticaine a small fraction of the phospholipids will be in ahase (inverted micellar, rhombic or cubic) where effectively
sotropic motional averaging occurs (see Fig. 3A, right column)Cullis et al., 1978). The isotropic peak in the two spectra atH of 10.0 (Fig. 3A, left column) can be corroborated with theinor resonances in the 13C MASNMR spectra shown in Fig. 2
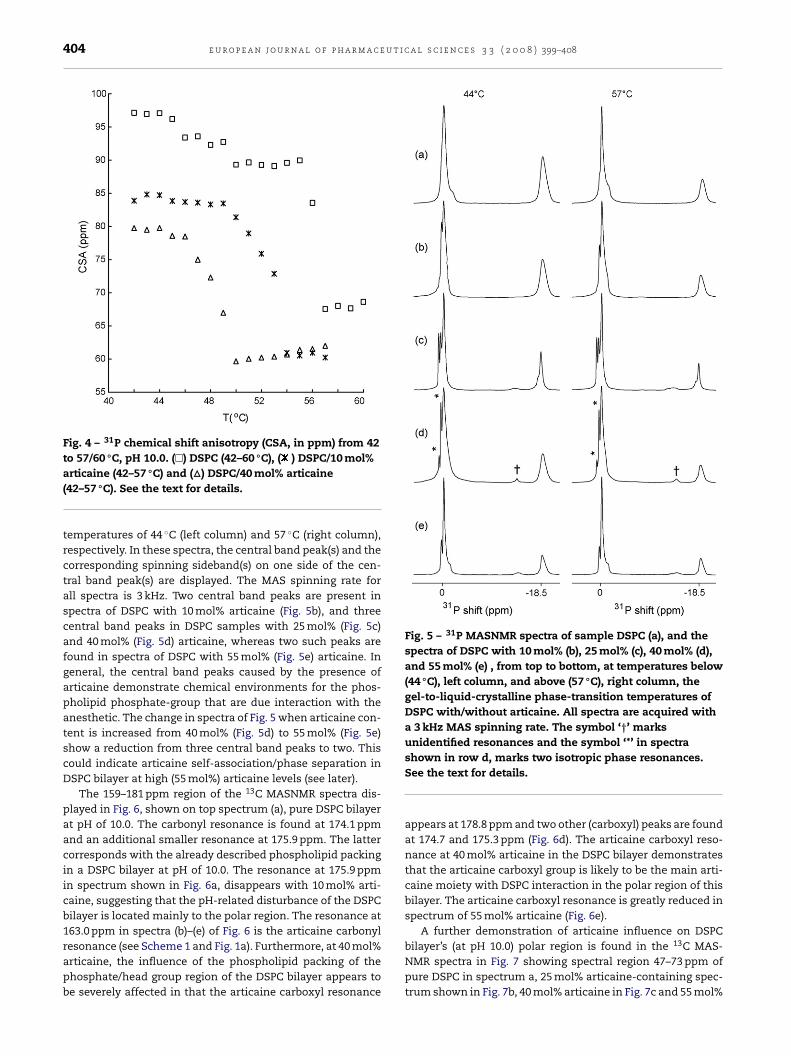
bottom), in that a pH of 10.0 has some effect on the molecu-ar packing in the glycerol/headgroup part of the DSPC bilayer.ig. 4 shows the 31P CSA as a function of sample temperature.he CSA of pure DSPC (� in Fig. 4) displays a sudden dropt the DSPCs gel-to-liquid-crystalline phase-transition tem-erature of 56 ◦C. CSA values of DSPC bilayer with 10 mol%
rticaine (* in Fig. 4) and 40 mol% articaine (� in Fig. 4) displaybilayer phase transition temperature of 52 and 48 ◦C, respec-ively. The DSPC bilayer samples with articaine contents of 25nd 55 mol%, show gel-to-liquid-crystalline phase-transition
corresponding two spectra of the right column are acquiredon DSPC with 40 mol% articaine. See the text for details.
temperatures of 48 and 46 ◦C, respectively (data not shown).Thus, the observed concentration-dependent reduction in
bilayer gel-to-liquid-crystalline phase-transition temperaturedemonstrates articaine interaction with the DSPC bilayer.Fig. 5 shows 31P MASNMR spectra of DSPC bilayer without(Fig. 5a) and with 10–55 mol% articaine (Fig. 5b–e) at sample
404 e u r o p e a n j o u r n a l o f p h a r m a c e u t i c a l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408
Fig. 4 – 31P chemical shift anisotropy (CSA, in ppm) from 42to 57/60 ◦C, pH 10.0. (�) DSPC (42–60 ◦C), ( ) DSPC/10 mol%
Fig. 5 – 31P MASNMR spectra of sample DSPC (a), and thespectra of DSPC with 10 mol% (b), 25 mol% (c), 40 mol% (d),and 55 mol% (e) , from top to bottom, at temperatures below(44 ◦C), left column, and above (57 ◦C), right column, thegel-to-liquid-crystalline phase-transition temperatures ofDSPC with/without articaine. All spectra are acquired witha 3 kHz MAS spinning rate. The symbol ‘†’ marksunidentified resonances and the symbol ‘*’ in spectra
articaine (42–57 ◦C) and (�) DSPC/40 mol% articaine(42–57 ◦C). See the text for details.
temperatures of 44 ◦C (left column) and 57 ◦C (right column),respectively. In these spectra, the central band peak(s) and thecorresponding spinning sideband(s) on one side of the cen-tral band peak(s) are displayed. The MAS spinning rate forall spectra is 3 kHz. Two central band peaks are present inspectra of DSPC with 10 mol% articaine (Fig. 5b), and threecentral band peaks in DSPC samples with 25 mol% (Fig. 5c)and 40 mol% (Fig. 5d) articaine, whereas two such peaks arefound in spectra of DSPC with 55 mol% (Fig. 5e) articaine. Ingeneral, the central band peaks caused by the presence ofarticaine demonstrate chemical environments for the phos-pholipid phosphate-group that are due interaction with theanesthetic. The change in spectra of Fig. 5 when articaine con-tent is increased from 40 mol% (Fig. 5d) to 55 mol% (Fig. 5e)show a reduction from three central band peaks to two. Thiscould indicate articaine self-association/phase separation inDSPC bilayer at high (55 mol%) articaine levels (see later).
The 159–181 ppm region of the 13C MASNMR spectra dis-played in Fig. 6, shown on top spectrum (a), pure DSPC bilayerat pH of 10.0. The carbonyl resonance is found at 174.1 ppmand an additional smaller resonance at 175.9 ppm. The lattercorresponds with the already described phospholipid packingin a DSPC bilayer at pH of 10.0. The resonance at 175.9 ppmin spectrum shown in Fig. 6a, disappears with 10 mol% arti-caine, suggesting that the pH-related disturbance of the DSPCbilayer is located mainly to the polar region. The resonance at163.0 ppm in spectra (b)–(e) of Fig. 6 is the articaine carbonyl
resonance (see Scheme 1 and Fig. 1a). Furthermore, at 40 mol%articaine, the influence of the phospholipid packing of thephosphate/head group region of the DSPC bilayer appears tobe severely affected in that the articaine carboxyl resonanceshown in row d, marks two isotropic phase resonances.See the text for details.
appears at 178.8 ppm and two other (carboxyl) peaks are foundat 174.7 and 175.3 ppm (Fig. 6d). The articaine carboxyl reso-nance at 40 mol% articaine in the DSPC bilayer demonstratesthat the articaine carboxyl group is likely to be the main arti-caine moiety with DSPC interaction in the polar region of thisbilayer. The articaine carboxyl resonance is greatly reduced inspectrum of 55 mol% articaine (Fig. 6e).
A further demonstration of articaine influence on DSPC
bilayer’s (at pH 10.0) polar region is found in the 13C MAS-NMR spectra in Fig. 7 showing spectral region 47–73 ppm ofpure DSPC in spectrum a, 25 mol% articaine-containing spec-trum shown in Fig. 7b, 40 mol% articaine in Fig. 7c and 55 mol%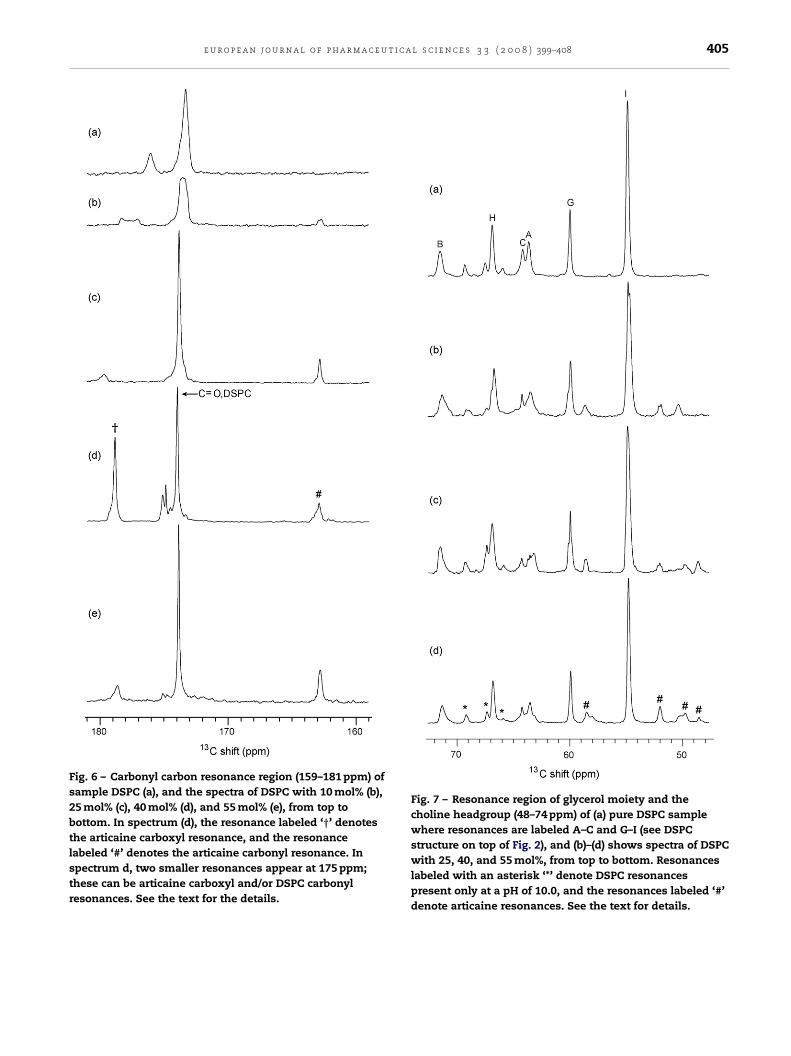
e u r o p e a n j o u r n a l o f p h a r m a c e u t i c a l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408 405
Fig. 6 – Carbonyl carbon resonance region (159–181 ppm) ofsample DSPC (a), and the spectra of DSPC with 10 mol% (b),25 mol% (c), 40 mol% (d), and 55 mol% (e), from top tobottom. In spectrum (d), the resonance labeled ‘†’ denotesthe articaine carboxyl resonance, and the resonancelabeled ‘#’ denotes the articaine carbonyl resonance. Inspectrum d, two smaller resonances appear at 175 ppm;these can be articaine carboxyl and/or DSPC carbonylresonances. See the text for the details.
Fig. 7 – Resonance region of glycerol moiety and thecholine headgroup (48–74 ppm) of (a) pure DSPC samplewhere resonances are labeled A–C and G–I (see DSPCstructure on top of Fig. 2), and (b)–(d) shows spectra of DSPCwith 25, 40, and 55 mol%, from top to bottom. Resonanceslabeled with an asterisk ‘*’ denote DSPC resonancespresent only at a pH of 10.0, and the resonances labeled ‘#’denote articaine resonances. See the text for details.

u t i c a l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408
Fig. 8 – Acyl chain carbon resonance in regions, 9–27 ppm,and 34–37 ppm (main acyl chain peak not shown), of (a)pure DSPC sample, (b) DSPC with 40 mol% articaine, and (c)DSPC with 55 mol% articaine. Resonances labeled with ‘†’denote new DSPC resonances when articaine is present,and resonances labeled with ‘#’ denote articaineresonances. Arrows point at broad articaine resonancesthat appear at high articaine ratios (55 mol%). These
406 e u r o p e a n j o u r n a l o f p h a r m a c e
articaine in Fig. 7d. In Fig. 7a the glycerol and choline headgroup carbon resonances are labeled A–C, G–I, and smallerresonances in this spectral region are due to some pertur-bation of phospholipids packing at a pH of 10.0. Articaineresonances are present in spectra of Fig. 7b–d at 58.5, 52.2,49.9 and 49.5 ppm, respectively.
Evidence for articaine influence on phospholipid pack-ing in DSPC bilayer (at pH 10.0) hydrophobic region is foundwhen comparing 13C MASNMR spectra shown in Fig. 8. Spec-trum of pure DSPC is shown in Fig. 8a, DSPC with 40 mol%articaine in Fig. 8b, DSPC with 55 mol% articaine in Fig. 8cand pure articaine (with 15 wt% D2O) in Fig. 8d. These spec-tra of Fig. 8 show spectral regions 9–27 ppm and 34–37 ppm.Major spectral changes are evident for acyl chain carbon res-onances 2 and 3, in Fig. 8b, where the two resonances thatcan be assigned to carbon 2 of the stearoyl acyl chain, andsimilarly the two resonances assigned to carbon 3, suggestconversion between two mesomorphic lipid states at a rateslower than the NMR timescale. Furthermore, smaller arti-caine resonances are present at 20.0, 16.8, 11.5 and 11.0 ppm.As demonstrated in Fig. 8c, the large molecular packing effecton the phospholipids by the 40 mol% articaine (Fig. 8b) isreduced in 55 mol% articaine-containing sample (Fig. 8c). Thearrows in Fig. 8c point at broad resonances that are not presentin spectrum 40 mol% articaine-containing sample (Fig. 8b).Accompanying these spectral changes is a marked reductionin articaine resonances (marked with ‘#’ in Fig. 8c) even thoughthe total articaine molar ratio increases from 40 mol% (Fig. 8b)to 55 mol% (Fig. 8c). The broad resonances only present inspectrum of the 55 mol% DSPC sample (indicated by arrowsin Fig. 8c) could very well come from self-aggregated arti-caine molecules, such an interpretation is supported by theobserved reduction in signal intensity of articaine resonances(marked with ‘#’ in Fig. 8c) as articaine mol% is increased from40 to 55.
4. Discussion
Our samples contain phospholipids hydrated with ∼32 watermolecules per phospholipid molecule, i.e. without bulk waterpresent the amount of articaine that can be accommodated inthe bilayer can be followed in the NMR spectra of our sampleswith 10, 25, 40 and 55 mol% articaine. Thus, the absence ofbulk water in our samples makes a lipid (or octanol)/waterpartition coefficient not directly applicable. Apparently, anarticaine molar ratio of 55 is above the amount that can beaccommodated in the DSPC bilayer and some form of articaineaggregates coexists with the articaine saturated bilayer. Thiscomes into effect when the articaine molar ratio is increasedfrom 40 to 55. In case of hydrolysis of the articaine at a pHof 10.0, formation of free carboxylic acid will follow. Evidenceof free carboxylic acid is not found in our 13C NMR spec-tra in that an eventual free carboxylic acid carbon resonancewould appear in the carbonyl chemical shift region (Fig. 6), andwould increase with increasing articaine molar ratio as well
as show a narrow line-shape compared with the other reso-nances due to the high mobility of a small molecule. No carbonresonance with these characteristics can be found in our spec-tra (Fig. 6). The uncharged articaine concentration-dependentresonances could be due to presence of articaineaggregates. (d) Spectrum of pure articaine showing regions,9–27 ppm, and 34–37 ppm. See the text for the details.

t i c a
rtitahbaeecDDcbcbtabd1
4dl
r
A
A
B
B
B
B
C
C
C
d
e u r o p e a n j o u r n a l o f p h a r m a c e u
eduction in the DSPC bilayer gel-to-liquid-crystalline phase-ransition temperature demonstrates substantial articainenteraction with this bilayer. In general, depression of gel-o-liquid-crystalline phase-transition temperature by localnesthetics has been found to correlate with the anestheticydrophobicity (Malheiros et al., 2004). However, hydropho-icity of the anesthetic as well as steric effects in thenesthetic–phospholipid interaction will determine the pref-rential positioning of the anesthetic in the bilayer (Fracetot al., 2005). The importance of articaine hydrophobicityan be seen when comparing the 13C MASNMR spectra ofSPC/40 mol% articaine at a pH of 7.4 (Fig. 1b) with that ofSPC/40 mol% articaine at a pH of 10.0 (Fig. 6d). Articainearboxyl group interaction with the PC bilayer appears toe substantial at a sample pH of 10.0, with uncharged arti-aine, and not present/minor at a sample pH of 7.4. A DSPCilayer contains a large hydrophobic core and the NMR spec-ra of the 40 mol% articaine-containing sample demonstrate
large disturbance in the molecular packing of the polarilayer region that extends into the hydrophobic region, evi-enced by carbon 2 and 3 of the acyl chains. Observed 31P and
3C NMR spectral changes when articaine is increased from0 to 55 mol%, suggest formation of articaine aggregates andecrease in DSPC bilayer perturbation at the latter articaine
evel.
e f e r e n c e s
rias, H.R., Sankaram, M.B., Marsh, D., Barrantes, F.J., 1990. Effectof local anaesthetics on steroid–nicotinic acetylcholinereceptor interactions in native membranes of Torpedomarmorata electric organ. Biochim. Biophys. Acta 1027 (3),287–294.
uger, M., Smith, I.C.P., Mantsch, H.H., Wong, P.T.T., 1990.High-pressure infrared study of phosphatidylserine bilayersand their interactions with the local anesthetic tetracaine.Biochemistry 29, 2008–2015.
oulanger, Y., Schreier, S., Smith, I.C.P., 1981. Molecular details ofanesthetic–lipid interaction as seen by deuterium andphosphorus-31 nuclear magnetic resonance. Biochemistry 20,6824–6830.
rau, M.E., Nau, C., Hempelmann, G., Vogel, W., 1995. Localanesthetics potently block a potential insensitive potassiumchannel in myelinated nerve. J. Gen. Physiol. 105, 485–505.
rowning, J.L., Seelig, J., 1980. Bilayers of phosphatidylserine: adeuterium and phosphorus nuclear magnetic resonancestudy. Biochemistry 19, 1262–1270.
utterworth, J.F.I.V., Strichartz, G.R., 1990. Molecular mechanismsof local anesthesia: a review. Anesthesiology 72, 711–734.
oster, H.G.L., James, V.J., Berthet, C., Miller, A., 1981. Locationand effect of procaine on lecithin/cholesterol membranesusing X-ray diffraction methods. Biochim. Biophys. Acta 641,281–285.
ullis, P.R., McLaughlin, A.C., 1977. Phosphorus nuclear magneticresonance studies of model and biological membranes.Trends Biol. Sci. 2, 213–226.
ullis, P.R., van Dijck, P.W.M., de Kruijff, B., de Gier, J., 1978. Effectsof cholesterol on the properties of equimolar mixtures of
synthetic phosphatidylethanolamine andphosphatidylcholine A 31P NMR and differential scanningcalorimetry study. Biochim. Biophys. Acta 513, 21–30.e Paula, E., Schreier, S., 1995. Use of a novel method fordetermination of partition coefficients to compare the effect
l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408 407
of local anesthetics on membrane structure. Biochim.Biophys. Acta 1240, 25–33.
de Verteuil, F., Pink, D.A., Vadas, E.B., Zuckermann, M.J., 1981.Phase diagrams for impure lipid systems application tolipid/anaesthetic mixtures. Biochim. Biophys. Acta 640,207–222.
Fraceto, L.F., Pinto, L.de.M.A., Franzoni, L., Braga, A.A.C., Spisni,A., Schreier, S., de Paula, E., 2002. Spectroscopic evidence for apreferential location of lidocaine inside phospholipid bilayers.Biophys. Chem. 99, 229–243.
Fraceto, L.F., Spisni, A., Schreier, S., de Paula, E., 2005. Differentialeffects of uncharged aminoamide local anesthetics onphospholipid bilayers, as monitored by 1H-NMRmeasurements. Biophys. Chem. 115, 11–18.
Garcia-Soto, J., Fernandez, M.S., 1983. The effect of neutral andcharged micelles on the acid–base dissociation of the localanesthetic tetracaine. Biochim. Biophys. Acta 731, 275–281.
Gjerde, A.U., Holmsen, H., Nerdal, W., 2004. Chlorpromazineinteraction with phosphatidylserines: a 13C and 31P solid-stateNMR study. Biochim. Biophys. Acta 1682, 28–37.
Grossmann, M., Sattler, G., Pistner, H., Oertel, R., Richter, K.,Schinzel, S., Jacobs, L.-D., 2004. Pharmacokinetics of articainehydrochloride in tumescent local anesthesia for liposuction. J.Clin. Pharmacol. 44, 1282–1289.
Gupta, S.P., 1991. QSAR (quantitative structure–activityrelationship) studies on local anesthetics. Chem. Rev. 91,1109–1119.
Hata, T., Matsuki, H., Kaneshina, S., 2000a. Effect of localanesthetics on the phase transition temperatures of ether-and ester-linked phospholipid bilayer membranes. ColloidsSurf. B 18, 41–50.
Hata, T., Sakamato, T., Matsuki, H., Kaneshina, S., 2000b. Partitioncoefficients of charged and uncharged local anesthetics intodipalmitoylphosphatidylcholine bilayer membrane:estimation from pH dependence on the depression of phasetransition temperatures. Colloids Surf. B 22, 77–84.
Hollman, M.W., Wieczorek, K.S., Berger, A., Durieux, M.E., 2001.Local anesthetic inhibition of G protein-coupled receptorsignaling by interference with G˛q protein function. Mol.Pharmacol. 59, 294–301.
Janiak, M.J., Small, D.M., Shipley, G.G., 1979. Temperature andcompositional dependence of the structure of hydrateddimyristoyl lecithin. J. Biol. Chem. 254, 6068–6078.
Kaneshina, S., Satake, H., Yamamoto, T., Kume, Y., Matsuki, H.,1997. Partitioning of local anesthetic dibucaine into bilayermembranes of dimyristoylphosphatidylcholine. Colloids Surf.B 10, 51–57.
Kelusky, E.C., Smith, I.C.P., 1984. The influence of local anestheticson molecular organization in phosphatidylethanolaminemembranes. Mol. Pharmacol. 26, 314–321.
Lullman, H., Timmermans, P.B.M.W.M., Weikert, G.M., Ziegler, A.,1980. Accumulation of drugs by guinea pig isolated atriaquantitative correlations. J. Med. Chem. 23, 560–565.
Malamed, S.F., Gagnon, S., Leblanc, D., 2000a. Efficacy of articaine:a new amide local anesthetic. J. Am. Dent. Assoc. 131, 635–642.
Malamed, S.F., Gagnon, S., Leblanc, D., 2000b. A comparisonbetween articaine HCl and lidocaine HCl in pediatric dentalpatients. Pediatr. Dent. 22, 307–311.
Malamed, S.F., Gagnon, S., Leblanc, D., 2001. Articainehydrochloride: a study of the safety of a new amide localanesthetic. J. Am. Dent. Assoc. 132, 177–185.
Malheiros, S.V.P., Pinto, L.M.A., Gottardo, L., Yokaichiya, D.K.,Fraceto, L.F., Meirelles, N.C., de Paula, E., 2004. A new look atthe hemolytic effect of local anesthetics, considering their
real membrane/water partitioning at pH 7.4. Biophys. Chem.110, 213–221.Neal, M.J., Butler, K.W., Polnaszek, C.F., Smith, I.C.P., 1976. Theinfluence of anesthetics and cholesterol on the degree ofmolecular organization and mobility of ox brain white matter

u t i c
4344–4349.
408 e u r o p e a n j o u r n a l o f p h a r m a c e
Lipids in multibilayer membranes: a spin probe study usingspectral simulation by the stochastic method. Mol.Pharmacol. 12, 144–155.
Nerdal, W., Gundersen, S.A., Thorsen, V., Høiland, H., Holmsen,H., 2000. Chlorpromazine interaction withglycerophospholipid liposomes studied by magic anglespinning solid-state 13C-NMR and differential scanningcalorimetry. Biochim. Biophys. Acta 1464, 165–175.
Papahadjoupoulos, D., Jacobson, K., Poste, G., Shepherd, G., 1975.Effects of local anesthetics on membrane properties. I.Changes in the fluidity of phospholipid bilayers. Biochim.Biophys. Acta 394, 504–519.
Pinto, L., de, M.A., Yokaichiya, D.K., Fraceto, L., de Paula, F.E., 2000.Interaction of benzocaine with model membranes. Biophys.Chem. 87, 213–223.
Schreier, S., Malheiros, S.V.P., de Paula, E., 2000. Surface activedrugs: self-association and interaction with membranes andsurfactants physicochemical and biological aspects. Biochim.Biophys. Acta 1508, 210–234.
Seelig, J., 1978. 31P nuclear magnetic resonance and the headgroup structure of phospholipids in membranes. Biochim.Biophys. Acta 515, 105–140.
Small, D.M., 1967. Phase equilibria and structure of dry andhydrated egg lecithin. J. Lipid Res. 8, 551–557.
Song, C., Holmsen, H., Nerdal, W., 2006. Existence of lipidmicrodomains in bilayer of dipalmitoylphosphatidylcholine(DPPC) and 1-stearoyl-2-docosahexenoyl phosphatidylserine(SDPS) and their perturbation by chlorpromazine: a 13C and31P solid-state NMR study. Biophys. Chem. 120, 178–187.
Suwalsky, M., Schneider, C., Villena, F., Norris, B., Cardenas, H.,Cuevas, F., Sotomayor, C.P., 2002. Structural effects of the local
anesthetic bupivacaine hydrochloride on the humanerythrocyte membrane and molecular models. Blood CellsMol. Dis. 29, 14–23.Suwalsky, M., Schneider, C., Villena, F., Norris, B., Cardenas, H.,Cuevas, F., Sotomayor, C.P., 2005. A study of the perturbation
a l s c i e n c e s 3 3 ( 2 0 0 8 ) 399–408
effects of the local anesthetic procaine on human erythrocyteand model membranes and of modifications of the sodiumtransport in toad skin. Biophys. Chem. 116, 227–235.
Tanaka, T., Hidaka, H., 1981. Interaction of local anesthetics withcalmodulin. Biochem. Biophys. Res. Commun. 101, 447–453.
Vanderkooi, G., Adade, A.B., 1986. Stoichiometry and dissociationconstants for interaction of tetracaine with mitochondrialadenosinetriphosphatase as determined by fluorescence.Biochemistry 25, 7118–7124.
Vree, T.B., Baars, A.M., Van Oss, G.E.C.J.M., Booij, L.H.D.J., 1988.High-performance liquid chromatography and preliminarypharmacokinetics of articaine and its 2-carboxy metabolite inhuman serum and urine. J. Chromatogr. 424, 440–444.
Wenk, M.R., Seelig, J., 1998. Proton induced vesicle fusion and theisothermal L�→HII phase transition of lipid bilayers A31P-NMR and titration calorimetry study. Biochim. Biophys.Acta 1372, 227–236.
Westman, J., Boulanger, Y., Ehrenberg, A., Smith, I.C.P., 1982.Charge and pH dependent drub binding to model membranesA 2H-NMR and light absorption study. Biochim. Biophys. Acta685, 315–328.
Wu, W.-G., Chi, L.-M., 1990. Comparisons of lipid dynamics andpacking in fully interdigitatedmonoarachidoylphosphatidylcholine and non-interdigitateddipalmitoylphosphatidylcholine bilayers: crosspolarization/magic angle spinning 13C-NMR studies. Biochim.Biophys. Acta 1026, 225–235.
Yeagle, P.L., Hutton, W.C., Huang, C., Martin, R.B., 1977.Phospholipid head-group conformations; intermolecularinteractions and cholesterol effects. Biochemistry 16,
Yoshino, A., Yoshida, T., Okabayashi, H., Kamaya, H., Ueda, I.,1998. 19F and 1H NMR and NOE study on halothane–micelleinteraction: residence location of anesthetic molecules. J.Colloid Interf. Sci. 198, 319–322.
Related Documents










![Modos De Aprendizaje [ABP y otros, 31p]](https://static.cupdf.com/doc/110x72/5695d17c1a28ab9b0296b8d1/modos-de-aprendizaje-abp-y-otros-31p.jpg)
