Abstracts – INCO Members INCO partner: 1 PRESENT STUDIES ON BIODIVERSITY OF ARTEMIA POPULATIONS IN NORTHERM PRESENT STUDIES ON BIODIVERSITY OF ARTEMIA POPULATIONS IN NORTHERM ASIA ASIA G. Van Stappen, B. Marden, L. Litvinenko, I. Mirabdullayev, I. Zholdasova, Xin Naihong, P. Bossier, P. Sorgeloos Laboratory of Aquaculture & Artemia Reference Center (ARC), Ghent University, Rozier 44, B-9000 Gent, Belgium, tel. 32-9-264 37 54; fax 32-9-264 41 93 1. Distribution of Artemia in lakes of South-Siberia Topography and climatological aspects A survey of Artemia habitats in south Siberia resulted in the registration of 90 sites, distributed over the Altai region (total area of Artemia lakes 1280 km 2 ), Kurgan region (123 km 2 ), Omsk region (92 km 2 ) and Novosibirsk region (92 km 2 ). Most of these lakes are small to medium sized, generally ranging between 1 and 10 km² surface area. All sites are located in a relatively flat arid or semi-arid zone, merging into the semi-deserts of Kazakhstan in the south. The climate is typically continental with low winter temperatures (water temperature may drop below –19°С; ice layer may be 15-20 cm thick in winter, only above 250 ppt no ice formation occurs) and relatively warm summer temperatures (water temperatures up to 25-30°C and even exceeding 40°C in shallow lakes). Because of the shallowness of most lakes, generally no temperature stratification occurs. Over a 6-year observation period (1995-2001), considerable fluctuations in salinity have been observed as a consequence of fluctuations in precipitation (e.g. doubling of salinity in Medvezhye Lake in the period 1995-2000, followed by a very rainy 2001). Some lakes show total desiccation every summer; others periodically dry out, e.g. in years with reduced rainfall and/or increased evaporation. Precipitation is limited (250-400 mm annually) and generally occurs as snowfall. Increase of salinity and/or elevated temperatures in summer may result in total mortality of brine shrimp

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Abstracts – INCO Members
INCO partner: 1
PRESENT STUDIES ON BIODIVERSITY OF ARTEMIA POPULATIONS IN NORTHERMPRESENT STUDIES ON BIODIVERSITY OF ARTEMIA POPULATIONS IN NORTHERM ASIAASIA
G. Van Stappen, B. Marden, L. Litvinenko, I. Mirabdullayev, I. Zholdasova, Xin Naihong, P. Bossier, P. Sorgeloos
Laboratory of Aquaculture & Artemia Reference Center (ARC), Ghent University, Rozier 44, B-9000 Gent, Belgium, tel. 32-9-264 37 54; fax 32-9-264 41 93
1. Distribution of Artemia in lakes of South-Siberia
Topography and climatological aspectsA survey of Artemia habitats in south Siberia resulted in the registration of 90 sites, distributed over the Altai region (total area of Artemia lakes 1280 km2), Kurgan region (123 km2), Omsk region (92 km2) and Novosibirsk region (92 km2). Most of these lakes are small to medium sized, generally ranging between 1 and 10 km² surface area. All sites are located in a relatively flat arid or semi-arid zone, merging into the semi-deserts of Kazakhstan in the south. The climate is typically continental with low winter temperatures (water temperature may drop below –19°С; ice layer may be 15-20 cm thick in winter, only above 250 ppt no ice formation occurs) and relatively warm summer temperatures (water temperatures up to 25-30°C and even exceeding 40°C in shallow lakes). Because of the shallowness of most lakes, generally no temperature stratification occurs.
Over a 6-year observation period (1995-2001), considerable fluctuations in salinity have been observed as a consequence of fluctuations in precipitation (e.g. doubling of salinity in Medvezhye Lake in the period 1995-2000, followed by a very rainy 2001). Some lakes show total desiccation every summer; others periodically dry out, e.g. in years with reduced rainfall and/or increased evaporation. Precipitation is limited (250-400 mm annually) and generally occurs as snowfall. Increase of salinity and/or elevated temperatures in summer may result in total mortality of brine shrimp in some lakes. There are a few low-salinity lakes where Artemia occurred in the past, but due to increased precipitation it is presently not found. Furthermore in several lakes with relatively low salinity Artemia faces competition with other zooplanktonts; lakes with Artemia as monoculture are a minority.
Hydrology and ionic compositionDepth of the lakes is generally in the order of 2-3 m at most; most lakes are thalassohaline, though carbonate and sulfate lakes are found; about 2/3 of the lakes have an average salinity below 150 ppt.
PhytoplanktonPhytoplankton species diversity is limited, and cell densities are overall low, reflected in high water transparencies. All species identified sofar are ubiquitous species of brackish water and salt lakes. In terms of abundance, green (and to a lower extent blue-green) algae prevail, generally making up 70-90 % of total phytoplankton biomass. The seasonal dynamics of phytoplankton biomass was monitored in a number of lakes. Generally phytoplankton densities peak in spring (April-May) before maximal Artemia densities are reached, and a second,

smaller, peak is observed just before winter (October-November) when the Artemia population has declined.
Artemia population dynamicsThe growth and reproduction period for Artemia (> 4-5°C water temperature) is limited to the period end of April-early October. According to present data, Artemia produces 3 to 4 generations within this period; in small and shallow lakes, subject to summer desiccation, only 1 or 2 generations are produced.
Parthenogenetic populations predominate; until now bisexual populations have only been found in 3 lakes. Depending on the available field samples, cyst biometrics and adult morphometrics have been determined, as well as cytogenetics for a number of populations.
Detailed Artemia population dynamics have been monitored for two consecutive years in Medvezhye lake and for one year in Nevidim and Vishnakovskoye lakes. Maximal biomass densities are observed late spring, early summer. Later on densities decrease, presumably as a consequence of food depletion. Brood sizes are generally relatively low, reflecting the low productivity of the lakes. In small shallow lakes Artemia distribution is very much dependent on weather conditions. Small commercial quantities of cysts are harvested in several lakes
2. NATO Collaborative Linkage Grant “Artemia colonization of the Aral Sea: hope for a dying ecosystem”ARC cooperation with 1) Laboratory of Ichthyology and Hydrobiology, Institute of Zoology, Tashkent, Uzbekistan; 2) Institute of Bioecology of the Karakalpak Branch of the Uzbek Academy of Sciences, Nukus, Uzbekistan; 3) Inve Aquaculture, Inc.
Background The Aral Sea has been claimed to be one of the worst human-induced ecological disasters of this century. Water management alternatives, the introduction of drought tolerant crop strains, less wasteful irrigation methods and other means of mitigating the economic losses and ecological catastrophe for the Aral Basin have been proposed and water conservation measures for the entire Aral Basin have been initiated. But these efforts to increase the Aral Sea elevation, to reduce the level of salinity, and reverse the ecological demise of the region have been largely unsuccessful. The contemporary Aral Sea is essentially a hypersaline lake with near-total elimination of species of freshwater origin.
The economic consequences of the Aral crisis have had devastating financial and health impacts on local economies and communities. Any means of economic recovery for these communities would be a most welcome and much needed benefit. Prudent commercial exploitation of an emerging Artemia population could provide income and employment. It is unknown, however, whether the current hydrobiological and hydrochemical status and primary productivity of the Aral Sea is sufficient to support a stable Artemia population. Planning for immediate cyst and/or biomass harvesting at this stage is premature as this could jeopardize the future potential for a viable industry. Detailed ecological information is clearly needed in order to evaluate the feasibility and potential benefits of commercial exploitation of Artemia.
Scope and research programme The scope of this grant is to establish a team of biologists, limnologists, chemists, and natural resource experts from NATO countries and from Uzbekistan to document the ecological characteristics of the Aral Sea, more specifically the hydrobiological and hydrochemical status of the Aral Sea as it relates to the successful colonization by an Artemia population. A monitoring programme is taking place, focusing on Aral Sea developments in hydrochemistry, phytoplankton and its ongoing gradual colonization by a parthenogenetic population.
3. Contamination of Bohai Bay (PR China) samples with A. franciscana

Standard culture experiments with Artemia samples from the Bohai Bay area in China, harvested as early as 1980 until now, have been performed in order to perform a preliminary assessment of the gradual contamination of the autochthonous A. parthenogenetica populations by introduced A. franciscana. Provisional results show that from ± 1991 onwards, all available samples are at least predominantly bisexual, with percentages of males fluctuating between 31 and 50 %. Before 1991, ‘typical’ male percentages in the order 0.1-0.3 % are observed. As especially older samples (1980’s and early 90’s) often show no hatchability anymore, these data will be complemented and refined by identification of the species status of individual cysts, according to the authentication method developed by Bossier et al.
INCO partner: 1 & 2INCO partner: 1 & 2
POLYMORPHISM IN ARTEMIA SPECIES AS REVEALED WITH MITOCHONDRIAL ANDPOLYMORPHISM IN ARTEMIA SPECIES AS REVEALED WITH MITOCHONDRIAL AND NUCLEAR MARKERSNUCLEAR MARKERS
Daan Delbare1, Peter Bossier1, Stefania Dooms 1,2, Gilbert Van Stappen2, Patrick Sorgeloos2, Zhijun Qiu3, Thomas H. MacRae3
1 Agricultural Research Center-Ghent/Department of Sea Fisheries (CLO-DVZ), Ankerstraat 1, 8400 Oostende, Belgium
2 Laboratory of Aquaculture & Artemia Reference Center (ARC), Ghent University, Rozier 44, B-9000 Gent, Belgium
3 Department of Biology, Dalhousie University, Halifax, N.S., Canada
As reported in previous meetings a database of RFLP patterns of a 1500 bp mitochondrial rDNA fragment has been constructed. This database contains now RFLP patterns (using 8 restriction enzymes)on more than 100 samples. A subset of these samples is now used as reference database for authenticating unknown samples at the species level. The 1500 bp mitochondrial rDNA fragment can be amplified from either a single cyst or a small amount of cysts. Detailed analysis of this reference database revealed that the RFLP patterns of one single restriction enzyme (HpaII) is sufficient to differentiate between the species present in the reference database. Samples grouping to certain species were characterised by the presence of unique marker fragments. The HpaII restriction patterns displayed two fragments, namely 1200 and 240 bp, that were present in all strains from the “franciscana” group and absent in all other samples, making them putative markers for that group. In addition all Artemia samples originating from North America displayed an unique 541 bp HinfI fragment. Other groups also displayed unique restriction fragments: samples from the “sinica” group originating from China showed unique 1486 bp TaqI and 464 bp Hinf I fragments. The 317 bp HinfI, the 273 DdeI and the 205 bp HaeIII fragments were characteristic for the “salina” group. Finally the “parthenogenetic” group was characterised by the presence of a typical 297 bp HpaII fragment. A single A. persimilis sample is characterised by two HpaII (656 and 611 bp) and two DdeI fragments (453 and 421 bp) that were unique for this sample. It remains to be established if these fragments are real markers for this group, through the analysis of other A. persimilis populations (collaboration with partner 14, Gonzalo Gajardo, Chili seems to confirm this). These markers and the typical HpaII restriction patterns can now be used as an easy tool to identify Artemia at the species level. As it is possible to perform the analysis on a single cyst, it becomes feasible to study the populations dynamics in cases of syntopic occurrence of bisexual and parthenogenetic species.

The use of mitochondrial markers is in a considerable amount of samples hampered by the occurrence of “double” restriction patterns. These double restriction patterns are characterised by a restriction banding pattern of which the total sum of the visualised bands sizes is bigger than the 1500bp. Sufficient evidence has been gathered that shows that the observed phenomenon is not the consequence of a partial restriction digest. The phenomenon even occurs in mitochondrial DNA amplified from a single individual. The 1500 bp rDNA fragment as well as a smaller cyt B fragment is affected by this phenomenon. Recent analysis on parthenogenetic individuals from Urmia lake and surroundings (collaboration with partner 11) has also in these samples confirmed this phenomenon. It is suspected that this is the result of either heteroplasmy or pseudogenes.
The Hsp26 gene can be amplified from cysts by RT-PCR using a primer couple developed in the lab of Tom MacRae. Polymorphism was detected by RFLP (see previous report). The PCR fragments have now been sequenced at the lab of Tom MacRae. Part of that sequence is shown below. The genetic distance between these sequences will be shown at the meeting. Of particular interest is the polymorphism that was found at one particular nucleotide (marked in yellow in the figure), differentiating Artemia franciscana sampled in North America and in Vietnam (Vinh Chau). As the latter are more thermotolerant it is now investigated whether this difference is a marker for the Vinh Chau strain, by verifying much more individuals. In addition at the lab of Tom MacRae the genomic HSP26 sequence has been established. This opens the possibility to amplify a fragment containing the putative polymorphic nucleotide starting from a single individual or cyst (a protocol for doing RT-PCR on a single cyst has not been developed so far)
INCO partner: 3 (I)INCO partner: 3 (I)
PRELIMINARY DATA ON INTRASPECIFIC GENETIC DIVERGENCE BETWEEN PRELIMINARY DATA ON INTRASPECIFIC GENETIC DIVERGENCE BETWEEN ARTEMIAARTEMIA FRANCISCANAFRANCISCANA, SAN FRANCISCO BAY AND INOCULATED POPULATIONS IN VIETNAM, SAN FRANCISCO BAY AND INOCULATED POPULATIONS IN VIETNAM
Ilias Kappas1, Theodore J. Abatzopoulos1, Nguyen Van Hoa2, Patrick Sorgeloos3 & John A. Beardmore4
1 Department of Genetics, Development & Molecular Biology, Aristotle University of Thessaloniki, 541 24 Thessaloniki, Greece
2 Institute of Marine Aquaculture, University of Can Tho,Vietnam
3 Laboratory of Aquaculture and Artemia Reference Center, Ghent University, Rozier 44, Ghent B-9000, Belgium
4 School of Biological Sciences, University of Wales Swansea, Singleton Park, Swansea SA2 8PP, Wales, UK
The main objective in this study was to investigate the microevolutionary changes that have taken place since the introduction of Artemia franciscana (SFB) into Vietnam. Temperature is a key factor, expected to have a bearing on the genetic architecture of Artemia populations in the area of Vinh Chau saltworks, Vietnam. Therefore, the hypothesis in question dealt with the response of the source population (A. franciscana, SFB) to a novel environment. In addition, the speed of this response was tested through comparisons of early and late Vietnamese Artemia samples.

The methodological approach consisted of the examination of genetic differences at three different levels. A number of reproductive characters were tested for intrapopulation differentiation at 26°C and 30°C. Also, a battery of allozyme loci, routinely employed in Artemia population genetics, were screened for variation within, and differentiation between, Artemia populations from Vietnam. Finally, the same populations were assayed by way of Restriction Fragment Length Polymorphism (RFLP) analysis of a region of the mitochondrial (mt) DNA.
Materials and MethodsThe samples of A. franciscana used in this study consisted of A. franciscana, San Francisco Bay (SFB) considered as the inoculum (source) population, the Vinh Chau (VC) strain, present in Vinh Chau saltfields since inoculation (1986) and strains of Year 1 (Y1), Year 3 (Y3) and Year 4 (Y4), originated after one, three and four culture seasons in Vinh Chau, respectively.Strain Y2 (after two culture seasons in Vinh Chau) was also used initially but it was subsequently dropped from the study due to crash.Reproductive charactersCyst samples of each strain were hatched (according to Sorgeloos et al., 1986) and young nauplii were transferred to cylindroconical tubes until they reached maturity. Mating pairs (in Falcon tubes) for each strain were set up in a salinity of 80ppt and two temperatures (26°C and 30°C). Five reproductive characters were daily recorded: number of cysts per female, number of nauplii per female, total number of offspring per female, number of broods per female and number of encycted broods per female. Data for each variable were analysed through a two-way ANOVA.AllozymesTwenty enzyme-coding loci were scored in all samples. The TFPGA-1.3 (Miller, 1997) software was used to calculate common measures of genetic variability and differentiation as well as to construct a UPGMA dendrogram of populations.MtDNAFollowing DNA extraction (Bardakci & Skibinski, 1994) samples were loaded on an automatic thermocycler for amplification of a 2963 bp long mtDNA target sequence. Eight restriction endonucleases were employed to assess variation in the amplified region. Two computer software, REAP, version 4.0 (McElroy et al., 1992) and NTSYS®, version 1.2 (Applied Biostatistics, Inc.) were used to obtain mtDNA variability indices and dendrograms.
Results and DiscussionThere is ample evidence of divergence in reproductive traits between the source A. franciscana, SFB population and the founded ones in Vietnam as well as between the latter. Temperature appears to be invariably involved in the observed differentiation between samples, either as a single factor or interactively with respect to the particular strain. At the temperature of 30°C the VC strain showed significantly higher reproductive output compared to all other strains, which is indicative of its thermal adaptation.Similar differentiation to that observed with reproductive characters was evident in the allozymic survey. The source population and the Vietnamese strains showed detectable genetic differences and patterns of genetic differentiation comparable to those found at an initial stage of divergence between geographic populations in nature. In addition, there was no evidence of genetic impoverishment through the successive generations of Vietnamese Artemia, a fact particularly important for the success of inoculation schemes.Unlike allozymes, strong indications of reduction in mtDNA gene diversity were obtained in the Vinh Chau populations. In particular, the VC strain displayed the smallest number of haplotypes and the lowest level of haplotype diversity compared to the rest. It seems that a global composite haplotype is driven through time (year classes) to near fixation (VC). The application of new molecular markers and tools (see Abatzopoulos et al., 2002) has revealed patterns of evolution previously undetected and has boosted considerably fine-scale genetic investigations.Overall, the “sequential” culture scheme used in Vinh Chau ponds favours the gradual accumulation of genetic adaptations. The different strains show considerable differentiation brought about by temperature. This is also supported by Clegg et al. (2000) who found a similar pattern in the thermal adaptation of cysts samples derived form the same strains.

ReferencesAbatzopoulos Th.J, Beardmore JA, Clegg JS, & Sorgeloos P. 2002. Artemia: Basic and Applied Biology. Kluwer Academic Publishers, Dordrecht.Bardakci F, & Skibinski DOF. 1994. Application of the RAPD technique in tilapia fish – species and subspecies identification. Heredity 73 (2): 117-123.Clegg JS, Jackson SA, Hoa NV, & Sorgeloos P. 2000. Thermal resistance, developmental rate and heat shock proteins in Artemia franciscana, from San Francisco Bay and southern Vietnam. Journal of Experimental Marine Biology and Ecology 252: 85-96.McElroy D, Moran P, Bermingham E, & Kornfield I. 1992. REAP – the restriction enzyme analysis package. Journal of Heredity 83: 157-158.Miller MP. 1997. TFPGA version 1.3, Department of Biological Sciences, Northern Arizona University, Box 5640, Flagstaff, AZ 86011-5640, USA.Sorgeloos P, Lavens P, Léger P, Tackaert W, & Versichele D. 1986. Manual for the culture and use of brine shrimp Artemia in Aquaculture. State University of Ghent, Belgium.
INCO partner 3: (II)INCO partner 3: (II)
HOW DO MITOCHONDRIALLY IDENTICAL HOW DO MITOCHONDRIALLY IDENTICAL ARTEMIAARTEMIA CLONES RESPOND TO CLONES RESPOND TO DIFFERENT SALINITIES?DIFFERENT SALINITIES?
G. Deliopoulos, A.D. Baxevanis & T.J. Abatzopoulos
Department of Genetics, Development and Molecular Biology, School of Biology, Aristotle University of Thessaloniki, 541 24 Thessaloniki, Greece
The brine shrimp Artemia consists of a number of sexual species and a large number of obligatory parthenogenetic strains. Polyploid parthenogens are characterized by apomixis (i.e. meiosis is totally suppressed), while diploid Artemia populations are characterized by automixis (i.e. capable of limited meiotic recombination); therefore, the former are considered monoclonal and the latter polyclonal (Barigozzi, 1974; Abatzopoulos et al. 2002, 2003).The majority of the studies, that have estimated environmental and genetic components of variance for life span and reproductive traits of Artemia, have focused on temperature and salinity; these are the most important abiotic factors affecting the life history of hypersaline organisms (Barata et al. 1996; Browne and Wanigasekera, 2000; Abatzopoulos et al. 2003).Μonoclonal populations produced by apomictic parthenogenetic Artemia (i.e. lineages derived from the same mother) could be used successfully for studying phenotypic traits (such as life span and reproductive characteristics), although this approach has been poorly utilized until very recently (Browne et al. 2002; Abatzopoulos et al. 2003). In this study, we tried to document the phenotypic expressions (ten life history traits) of two genotypic lineages exposed to variable levels of salinity; these lineages came from the same parthenogenetic population (but from different females) and they were identical according to RFLP-mtDNA analysis.
Materials and MethodsTwo Artemia clones were isolated from the parthenogenetic tetraploid population of Polychnitos saltworks (Lesbos island, Aegean Sea, Greece – Triantaphyllidis et al. 1993).

Culture conditionsStock culture of Polychnitos population (1 individual / 4ml) was kept in 1L cylindroconical glass jars maintained at 24oC. The salinity of the culture medium was 60 g l-1. Animals were fed with 75% of yeast-based diet LANSY-PZ (INVE Aquaculture NV, Belgium) and 25% Dunaliella tertiolecta according to Abatzopoulos et al. (2003). Experimental designExperimental designTwenty five females were isolated in equal of 50 ml cylindroconical tubes before they reached adulthood. After their first brood, five nauplii from each individual were sacrificed for DNA extraction (see mtDNA-RFLP analysis section). Extracted DNA was used for RFLP-mtDNA analysis; this technique revealed that there were two different clades of mtDNA in the same population (clade 1 and clade 2, respectively). Two individuals were chosen from clade 1 (A and B, respectively) for the continuation of the experiment. These two individuals produced clones which were used for recording ten life span and reproductive characteristics (total lifespan, pre-reproductive, reproductive and post-reproductive period, number of broods, days between broods, number of nauplii, number of encysted embryos, offspring per brood and offspring per reproductive day) under three different salinities (50, 100 and 150 g l -1; temperature was constant at 24±1oC). Reproductive and life span characteristics were analyzed by using non-parametric tests (Kruskal-Wallis), since the ANOVA assumptions were not fulfilled. The significance level was set to 0.05.
mmtDNAtDNA--RFLP analysisRFLP analysisDNA was extracted using the CTAB protocol described in Hillis et al. (1996). Part of the 16S rRNA gene was amplified using the universal primers L2510 and H3080, described by Palumbi et al. (1996). PCR products were digested with 9 restriction endonucleases, electrophoretically separated in 1.5% agarose gel, stained with ethidium bromide, visualized and photographed under UV light.
Results and DiscussionSix reproductive and four life span characters from the parthenogenetic clones A and B at three different salinities are summarized in Table 1. Statistical analysis using Kruskal-Wallis test indicated that significant differences exist between clones in each salinity treatment for the majority of the characters studied (see Table 1). More specifically, at 50 g l-1 clone A and B were statistically different in eight out of ten characters studied; clone A presented statistically lower values than clone B in total life span, reproductive period, post reproductive period, number of broods, days between broods, number of nauplii, offspring per brood and offspring per reproductive day. It is worth noting that at 50 g l-1 both clones have produced no encysted embryos. At 100 g l-1, eight out of the ten scored characters were statistically different; clone A and B appeared to have similar values only in post-reproductive period and in number of encysted embryos; at the same salinity, clone A outperformed clone B. At 150 g l-1, the same characteristics were statistically different between the two clones but in this case clone B seemed to be the fittest. In conclusion, the two investigated clones (A and B) appear to have different response to the elevation of salinity. There is no supporting evidence that clones, which appear to be identical in terms of mtDNA, should exhibit similar reproductive performance. However, mtDNA can be a useful tool when assessing verified clonal lineages (i.e. offspring produced by a single parthenogenetic apomictic female).
References Abatzopoulos T.J. et al., 2002. Artemia: Basic and Applied Biology, Kluwer Academic
Publishers. Abatzopoulos T.J. et al., 2003. Hydrobiologia 492: 191-199. Barata C. et al., 1996. J. Exp. Mar. Biol. Ecol. 196: 329-340. Barigozzi C., 1974. Evol. Biol. 7: 221-252. Browne R.A. & G. Wanigasekera, 2000. J. Exp. Mar. Biol. Ecol. 244: 29-44. Browne R.A. et al., 2002. J. Exp. Mar. Biol. Ecol. 267: 107-119. Hillis D.M. et al., 1996. Molecular Systematics, Sinauer Associates.

Palumbi S.R. et al., 1996. Simple Fool’s Guide to PCR, University of Hawai. Triantaphyllidis G.V. et al., 1993. Int. J. Salt Lake Res. 2: 59-68.
Table 1. Mean values (±S.D.) of various reproductive and life span characteristics for the two parthenogenetic clones A and B reared at 3 different salinities (temperature was 24±1oC). Significant differences were determined by Kruskal-Wallis test (p<0.05). Values in each characteristic and each salinity that share the same letter are not significantly different. Life span, days between broods, pre-, post-, and reproductive period are expressed in days
INCO partner: 3(III) & 4
THE USE OF RFLP 16S RDNA ANALYSIS FOR DETECTING OF THE USE OF RFLP 16S RDNA ANALYSIS FOR DETECTING OF ARTEMIAARTEMIA FRANCISCANA FRANCISCANA IN WESTERN MEDITERRANEAN REGIONIN WESTERN MEDITERRANEAN REGION
F. Amat1, A.D. Baxevanis2, A. Triantafyllidis2, A Tzika2, G. Mura3 & T.J. Abatzopoulos2
1Instituto de Acuicultura de Torre de la Sal (CSIC), Ribera de Cabanes (Castellón), Spain
2Department of Genetics, Development & Molecular Biology, School of Biology, Aristotle University of Thessaloniki, 541 24 Thessaloniki, Greece
3Dipartamento d Biologia Animale é dell’Uomo, Laboratorio di Zoologia Applicata, Universitá La Sapienza, Rome, Italy
There is an informative cyst bank and database on Artemia populations collected in the
Western Mediterranean region dating from early ‘80s until today. This fact together with the acquisition of brine shrimp cysts from waterbirds (i.e. dunlin-Calidris alpina and godwit-Limosa limosa) faeces and pellets in two areas of SW Iberian Peninsula (Cadiz Bay, Spain and Castro Marim salterns, Portugal), enabled a research effort for studying the present distribution of autochthonous Artemia populations considering, at the same time, the threatening expansion of A. franciscana which behaves as an exotic invasive species.

The results obtained so far show the exclusive presence of A. franciscana populations in Portuguese salterns, in some old salterns in Cadiz bay (Spain) and in the French Mediterranean shore. Co-occurrence of autochthonous (parthenogenetic) and American brine shrimp populations was, also, reported for Morocco (Mar Chica) and France (Aigues Mortes), while A. franciscana has not been found so far in Italian cyst samples. Furthermore, this attempt is expected to initiate a thorough search on intentional or non-intentional A. franciscana inoculations through anthropogenic activities (i.e. saltworks management and hatchery effluents), and to track down the dispersal of A. franciscana cysts by shorebirds in the SW Iberian peninsula. (Results submitted to the EEI 2003, 1rst National Congress on Exotic Invasive Species, Leon, Spain – June, 2003).
In this preliminary study, autochthonous and invasive species present in SW Spain were genetically characterised by using PCR (Polymerase Chain Reaction) – RFLP (Restriction Fragment Length Polymorphism) analyses on part of the mitochondrial genome through a collaborative research with INCO partner 3 (AUTH). These analyses were a continuation of the previous work done by CSIC in terms of morphometry, reproductive characteristics etc. The main goals of the current work were (i) to confirm the species status (assigned as A. franciscana) of El Pilar and El Estanquillo populations, and (ii) to find out if there are any differences in the genetic make up of diploid and tetraploid parthenogenetic Artemia populations in this region.
Artemia individuals from El Pilar and El Estanquillo populations already assigned as A. franciscana by CSIC (based on morphology), were scored for mtDNA analysis. Following DNA extraction, a 535 bp region of the mitochondrial 16S gene was successfully amplified by the PCR technique. Four restriction endonucleases were subsequently employed in order to digest the PCR product. These enzymes gave specific restriction profiles for the discrimination of A. franciscana, A. persimilis and A. salina. The RFLP analyses of both populations produced patterns typical of A. franciscana, which is in accordance with the CSIC chatacterisation.
The Odiel parthenogenetic Artemia population is a mixture of diploid and tetraploid individuals (based on CSIC results). Twenty individuals were scored for three enzymes and polymorphism was revealed for the same mitochondrial region. Individuals ascribed as diploid produced patterns different from those produced by individuals ascribed as tetraploid. This is a strong indication that the two groups belong to different strains which reinforces the initial characterisation by CSIC.
The Rocio population is comprised of bisexual A. salina and a diploid parthenogenetic strain. Twenty individuals (A. salina males and females as well as parthenogenetic females) were similarly scored for three restriction enzymes digesting the same mitochondrial region. These enzymes can discriminate A. salina from parthenogenetic strains. The individuals ascribed as A. salina gave the same patterns to those produced by a reference population of A. salina (Sfax, Tunisia). The rest individuals produced different patterns but identical to those of parthenogenetic diploid individuals from Odiel. The apparent similarity of Odiel and Rocio populations cannot be safely deduced before a greater number of restriction enzymes are employed.
As a conclusion, the RFLP analyses on the 16S mitochondrial DNA region support the characterisation (based on morphological and cross-breeding data) performed by CSIC and it is the first time that the presence of an A. franciscana population is recorded in the Mediterranean Sea. The combination of traditional (e.g. biometrics, morphometry, etc.) and molecular (e.g. RFLP analysis) techniques seems to be a useful tool for studying invasion patterns and species status in the genus Artemia.

INCO partner: 4INCO partner: 4
ARTEMIAARTEMIA BIODIVERSITY: CURRENT GLOBAL RESOURCES AND THEIR SUSTAINABLE BIODIVERSITY: CURRENT GLOBAL RESOURCES AND THEIR SUSTAINABLE EXPLOITATIONEXPLOITATION
Francisco Amat
Instituto de Acuicultura de Torre de la Sal (IATS-CSIC)(Consejo Superior de Investigaciones Científicas de España)
12595 Ribera de Cabanes (Castellón) SPAIN.
In accordance to the commitments established during the last workshop held in Beijing (China), and the results obtained in the research lines developed and bound to the current projects, during the INCO workshop in Puerto Varas (Chile), the IATS-CSIC group is going to submit information dealing with:
1) Brine shrimp Artemia franciscana as exotic invasive species in the Western Mediterranean and its dispersal by waterbirdsThe updating of cyst bank and database on Artemia cyst samples performed by this group showed the availability of cyst samples collected in the Western Mediterranean area from the beginning of the 80s until today. This availability, together with the prospection and recovering of brine shrimp cysts from waterbirds (Dunlin Calidris alpina and Godwit Limosa limosa) faeces and pellets in two areas in the Southwest of the Iberian peninsula: Cadiz Bay (Spain) and Castro Marim salterns (Portugal), enabled a research led to establish the present distribution of autochthonous brine shrimp populations facing the threatening development of A. franciscana populations behaving as an exotic invasive species.
The results obtained show the exclusive presence of A. franciscana populations in Portuguese salterns and in some old salterns in Cadiz bay (Spain) and in the French Mediterranean shore. Cooccurrence of autochthonous (parthenogenetic) and American brine shrimp populations was found in Morocco (Mar Chica) and France (Aigues Mortes), while A. franciscana was not found in Italian cyst samples. It is also expected to explain the origin of these foraneous A. franciscana populations as intentional or non-intentional inoculations through salterns management, aquacultural (hatchery effluents) and pet market activities, and to assess their ability to be dispersed by shorebirds within and between salterns in the Southwestern Iberian peninsula. (Results submitted to the EEI 2003, 1rst National Congress on Exotic Invasive Species, Leon, Spain , 4 - 7 June, 2003).Autochthonous and invasive species present in the Southwest of Spain were also genetically characterised (PCR-RFLP) through a collaborative research with INCO partner 3 (AUTH).
2) Brine shrimp in ArgentinaIn concertation with INCO partner 6 (UBA) a scientist was invited to develop an experiment designed to compare some bio-ecological parameters of the Argentinean brine shrimp species: A .persimilis (Salinas Grandes de Hidalgo) and A. franciscana (Mar Chiquita). The experience consisted in the exposure of nauplii of both populations to three different temperatures (12º, 21º and 28ºC) and four different salinities (30, 60, 90, 120 g. L -1) during 12-60 days, up to attaining reproductive adulthood. The most important results were:
A. persimilis always showed high mortality at low salinities (30 g. L -1). A. franciscana always showed low survival at the low temperature (12ºC), but survived
better than A. persimilis at 21º and 28ºC A. franciscana always showed higher fecundity rates than A. persimilis. A. persimilis showed higher growth rates than A. franciscana at all temperatures. A. persimilis showed sexual maturity earlier than A. franciscana at 12ºC. For the other
temperatures the results were similar.

In both species the increase of temperature is associated to an increase of survival and growth rates, and to a decrease of pre-reproductive period and maturity fulfilment.
This cooperation with Argentina (IATS – Universidad de Buenos Aires), supported by the Spanish Government (AECI-ICI Programmes), allowed to develop a third prospecting of Artemia populations mission in the Argentinean provinces of Chubut and Santa Cruz. More than a dozen hypersaline ecosystems, located between 45º 55´and 49º 18´ S, were visited, and Artemia presence, as living populations and/or cysts, was registered in nine of them. Collected cysts allowed to hatch living nauplii which were grown till adulthood under standard culture conditions. The morphometrics of adult males and females studied by multivariate discriminant analysis, provided evidence that all these populations belonged to the species A. persimils. In this moment the available information from Artemia distribution in Argentina allows to venture the presence of A. franciscana in 7 localities north to 36º S, and of A. persimilis in 25 localities south to 36º S. This A. persimilis distribution in Argentina can be linked to the presence of the same species in the localities of Laguna Amarga (Torres del Paine National Park) and Laguna de los Cisnes (Isla Grande) in the Chilean Patagonia and Tierra del Fuego, respectively.This material is presently under study to establish cysts (hydrated and decapsulated) biometry ; PUFA content ; reproductive output in terms of ovoviviparism and oviparism bound to environmental conditions under laboratory mass culture. 3) Brine shrimp in TunisiaIn concertation with INCO partner 8 (INAT) a scientist was invited to develop the characterisation of three autochthonous Artemia populations from Tunisia.: El Adhibet, Sijoumi and Sahline. They showed their taxonomical adscription to the Mediterranean bisexual Artemia salina.
4) Brine shrimp in IndiaIn concertation with INCO partner 15 (IART) a scientist was invited to apply the morphometric characterisation of Artemia populations under standard culture conditions, according to the method applied and the data base developed by the group INCO partner 4. Six samples were afforded from India: PUTHALAM cysts showed the exclusive presence of an autochthonous diploid (?) parthenogenetic strain. THAMARIKULAM cysts did not hatch (suspected parthenogenetic). VEMBAR, VEPPELODAI, ARASARADI and THERASPURAN cysts, presumably a mixture of autochthonous diploid parthenogenetic strain and introduced American brine shrimp, showed the exclusive presence of A. franciscana. This is a new example of complete outcompeting exclusion of autochthonous strains by the American brine shrimp, behaving as an exotic invasive species
INCO partner: 6INCO partner: 6
RECORD OF NEW ARTEMIA POPULATIONS AND CONTRIBUTION TO THERECORD OF NEW ARTEMIA POPULATIONS AND CONTRIBUTION TO THE MORPHOLOGICAL AND ECOLOGICAL CHARACTERIZATION OF SOMEMORPHOLOGICAL AND ECOLOGICAL CHARACTERIZATION OF SOME ARGENTINEAN POPULATIONSARGENTINEAN POPULATIONS
Rosa Graciela Cohen1, Carolina Arbasetti1, Ximena Inés Pastorino1, Julieta Goenaga1, Francisco Amat2.
1.Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Ciudad Universitaria, Núñez, Pab. II, 4° piso,
C1428EHA, Buenos Aires, Argentina. TELEFAX: 54 11 4 576-3384; TE: 54 11 4 576-3349
2. Instituto de Acuicultura de Torre de la Sal (IATS-CSIC)

(Consejo Superior de Investigaciones Científicas de España)12595 Ribera de Cabanes (Castellón) Spain.
From the last workshop in China, we have developed diverse activities. Firstly, we have prospected several new saline waterbodies along the coast of Patagonia (Argentina). Here, we have found and sampled seven new Artemia locations in South Chubut and North Santa Cruz provinces.Also, looking for morphological differences between species, we have performed microscopic studies on the amplexial zone in specific pairs of both A. persimilis, A. franciscana and interspecific (hybrid) pairs, with the description of male and females specific structures involved in the amplexus.On the other hand, aiming to help in the understanding of the biogeographic distribution of the species in our country, we have evaluated at the laboratory, the effect of diverse saline treatments in two Argentinean populations belonging to A. persimilis (from La Pampa province) and A. franciscana (from Córdoba province), by means of survival, longevity, sexual maturity and size of adults. In order to complete these results performing a more elaborated experimental design we have sent one person for training by the Spanish partner in the IATS laboratories. The experiments developed here included the simultaneous variation of two environmental variables conforming a complex grid of combined conditions (four salinities per three temperatures), plus their corresponding replicates. The effects of each condition were evaluated by means of survival, sexual maturity, growing rate and fecundity. Our previous results together with these resulting from the training stay, allowed us now to have a reasonable explanation for the clearly delimited latitudinal distribution of Artemia species in Argentina.
INCO partner: 8
STRAIN CHARACTERISATION AND CLIMATE EFFECT ON ARTEMIA FROM TUNISIASTRAIN CHARACTERISATION AND CLIMATE EFFECT ON ARTEMIA FROM TUNISIA
Romdhane Mohamed Salah
Institut National Agronomique de Tunisie, 43 av. Charles Nicolle 1082 Tunis Tunisiatel :++21697 325 090 fax :++216 71 799391
During last for years Tunisia had assumed successive dry seasons, were many wetlands are dry and no more artemia or cysts appears, the only areas where cysts could be collect are those linked to salt works or permanent waters. Among sites periodically investigated by the INAT team six localities reveal the permanent presence of cysts, nauplii or adult: Saltwork of Sahline Centre East (SAH) and Sfax centre east (SFX), sebkhas Ahdhibet South East(ADH), Moknine Centre East(MOK), Sijoumi North East(SIJ) and Korsia North West (KOR); Non representative sample (very low quantity or bad quality of cysts) are collected at 3 other sites : Sebkha Ariana, Sebkha Sidi El Hani and Chott Jerid.
Effect of some physico-chemical parameter on artemia characteristic have been realised (reproduction) compared with natural conditions.Morphological characteristic of 6 strain are studied according to standard methodology, measurement are taken after sexual differentiations on both male and female of each strain
With collaboration with the Spanich team (INCO partner 4) The taxonomical characterisations have been performed and compared with other strain, morphometry,

biometry and cross breeding experiment are conduct with 15 artemia strains from which 4 Tunisian samples.
INCO partner: 9
ARTEMIA RESEARCH IN SOUTHERN AFRICA: A REPORT ON A SAMPLING EXPEDITION THROUGH PARTS OF SOUTH AFRICA AND NAMIBIA
Horst Kaiser and Tom HechtHorst Kaiser and Tom Hecht
Department of Ichthyology and Fisheries Science, Rhodes University, Grahamstown 6140,Department of Ichthyology and Fisheries Science, Rhodes University, Grahamstown 6140, South AfricaSouth Africa
During the first year of the INCO Artemia project (2002) we investigated the genetics of a previously untested Artemia population from a salt works located in the Eastern Cape Province of South Africa. It was shown for the first time that this site comprises two Artemia populations, a bisexual and a parthenogenetic species.As southern Africa has a very poor Artemia sampling record, these interesting preliminary findings provided the justification for a sampling trip through South Africa and Namibia. In July 2003 we were able to sample cysts and adults at eight locations and plan to conduct genetic analyses on some of these populations in 2004.We present a map of sampling locations and their basic characteristics and discuss the need for future sampling trips. In addition, based on discussions with salt works managers and salinity records, we list potential sites for sampling that may contain cysts or adults depending on the season.Currently, there are records of 43 sites in South Africa and 5 locations in Namibia. Two Namibian sites had been mentioned in previous reviews on the biogeography of Artemia, while the other three were first sampled during this trip. In South Africa six sites had been sampled or reported on before, with three locations recorded as part of the INCO action and three others mentioned by other authors while 37 new sites were visited, and cysts and nauplii could be obtained from eight locations. However, most sites are potential Artemia habitat and should be visited again, hopefully next year during spring season. Most sites belong to operational salt works.Salt works visited were categorised into two sizes, either below or above approximately 2 ha.Of 37 South African sites visited during this collection trip at which Artemia are known to occur 19 pans (51%) were larger than 2 ha. There appear to be at least three commercial operations in South Africa selling Artemia cysts. These are located at Velddrift (32°47'S-18°10'E), Reynekerspan (33°41'S-25°47'E), and Missionvale (33°46'S-25°40'E). Discussions with managers on-site revealed that Artemia are frequently translocated between salt works.
A review of the literature and ARC records for Africa shows some, but often very limited, information on Artemia habitats in Algeria, Egypt, Kenya, Libya, Madagascar, Morocco, Mozambique, Namibia, Niger, Senegal, Tunisia, and South Africa. However, few sites have been analysed and species identifications are tentative or have not been completed.
INCO partner: 10INCO partner: 10
Distribution and characterization of Artemia in Bohai BayDistribution and characterization of Artemia in Bohai Bay

Xin Naihong, Li Yanan
Salt Research Institute, Yingkou Road 831, 300450 Tanggu, Tianjin, PR China
The Bohai bay is an important solar salt production area for China, more than 200 saltworks are over there. The saltworks in Bohai Bay provide unique environment for the production of Artemia, they are characterized by a high primary productivity providing abundant food to sustain large population of Artemia. 600 tons of Artemia cysts and several ten thousand tons of Artemia biomass are produced every year. Before early 90’s local Artemia species in the saltworks of Bohai bay were parthenogenetic. Since 1992 inoculation of A. franciscana were firstly taken in 4 saltworks of Bohai bay (Tanggu, Hangu, Luannan and Nanpu), A. franciscana strain were dominant in those saltworks in several years after inoculation, later on due to the using of exotic Artemia strains in the aquaculture around Bohai bay, the bisexual Artemia strains were further expanded to other saltworks of Bohai bay, field observation indicated that the dominant Artemia population in salt works of Bohai bay are bisexual strains. In this study we investigated the distribution of the Artemia resources in Bohai bay, 10 Artemia samples were collected from different saltworks. The hatching ability for original and treated cysts, hatching rate, cyst diameter, length of the nauplii and fatty acid profile were analyzed.
Figure 1. Distribution and sampling sites of Artemia in Bohai Bay, PR China

Table 1. The hatching ability and hatching rate of the Artemia cysts from different saltworks.
Name Abbreviation
Hatching ability (%) Hatching rate (hrs) at 25oC
Original Treated T0 T10 T90 Ts
Daqinghe DQH 23.563.78
85.783.68
17 19 28 9
Nanpu NP 32.345.67
75.642.24
16 18 28 10
Hangu HG 47.663.89
75.644.37
17 19 27 8
Tanggu TG 36.874.12
78.563.42
15 18 27 9
Dagang DG 42.982.46
86.652.78
15 17 26 9
Zhongjie ZJ 57.672.79
82.783.46
14 16 26 10
Haixing HX 45.783.49
83.562.65
16 19 29 10
Chengkou
CK 56.355.64
82.434.23
16 18 29 11
Lubei LB 67.655.60
86.752.34
17 19 29 10
Zhanhua ZH 58.603.56
88.603.20
17 19 27 8
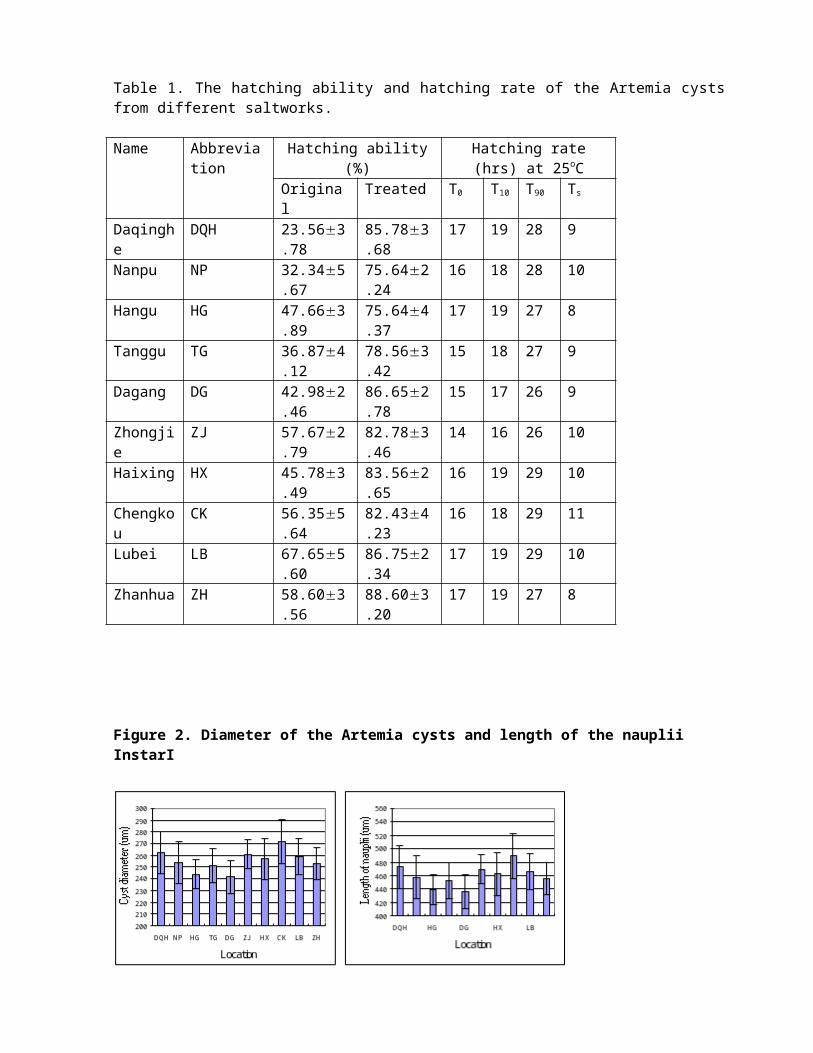
Figure 2. Diameter of the Artemia cysts and length of the nauplii InstarI

Figure 3. HUFA content of Artemia cysts from different saltworks
INCO partner: 11
DIVERSITY OF ARTEMIA POPULATIONS AT LAKE URMIA
Agh Naser1, Sorgeloos P.2, Abatzopoulos T.3, Van Stappen G.2, Sanders, L.4, Belbare, D.4,
Bossier P.2
1 Artemia & Aquatic Animals Research Center, Urmia University, Urmia, Iran 2 Laboratory of Aquaculture & Artemia Reference Center, Ghent University, Belgium3 Department of Genetics, Development & Molecular Biology, Faculty of Sciences, Aristotle
University of Thessaloniki, 541 24 Thessaloniki, Greece
4CLO Sea Fisheries Department, Ankerstraat 1, 8400 Oostende, Belgium
Existence of Artemia was first reported from the Lake Urmia by Günther in 1890. Clark and Bowen (1976) demonstrated the reproductive isolation of this species through laboratory crosses with A. franciscana and A. tunisiana and characterized it as a separate bisexual species and thereafter it was named as Artemia urmiana. But later Barigozzi et al. (1987a , b) and Badaracco et al. (1987) expressed Artemia urmiana as an exclusively parthenogenetic population, showing various ploidy levels (di-, tetra- and pentaploid). As a result of these findings Barigozzi (1988) proposed to cancel the species designation of Artemia urmiana. Nevertheless others (Azari Takami, 1989 and Browne, 1991) proposed probability of coexisting bisexual and parthenogenetic populations of Artemia in the Lake Urmia. So the sexual status of Artemia urmiana and possibility of a coexisting parthenogenetic Artemia in the Lake Urmia remained a mystery. In more recent years a number of studies have once again demonstrated presence of bisexual Artemia from the Lake Urmia. Regular observations of the living biomass in the lake by the author for a period of over 10 years (1992-2003) and the experiments performed by Sorgeloos (1997), Agh et al. (2002), Noori (2002), VanStappen (2002) showed that Artemia from Lake Urmia reproduces bisexually. Our experiments based on competition tests at different salinities ranging from 15-80 g/l proved presence of both bisexual and parthenogenetic populations of Artemia in the Lake Urmia. Salinity

based competition test was found to be a suitable and easy method for isolating parthenogenetic Artemia from bisexual species in a mixed cyst sample. At the same time presence of parthenogenetic Artemia at closely located lagoons was proven once again, supporting earlier findings by Agh and Noori (1997).
In another experiments the RFLP fingerprinting technique, applied on a 1500 bp mitochondrial rDNA fragment, was used to study the origin and similarity of Artemia populations from the Lake Urmia region. During this work adult bisexual and parthenogenetic females from the Lake Urmia and the lagoons at peripheries of the lake, were analysed. DNA was extracted from the whole body using Wizard® Genomic Purification Kit (Promegatm, mouse tail protocol). The PCR reactions of extracted DNA were performed using one pair of primers, namely 12S-SP (5’-CTAGGATTAGATACCCTA-3’) and 16S-SP (5’CCGGTCTGAACTCAGATC-3’) in a Hybrid PCR express (Labsystem TM, Belgium). The PCR products were then purified using and Wizard! ® PCR Preps Purification system (Promegatm ), followed by a confirmation of the presence of the PCR products, using agarose electrophoresis. In order to display the polymorphism in the mitochondrial rDNA gene, each sample was digested with 4 restriction enzymes (Hpa II, Taq I, Dde I & Nde II). The digested products were separated through an agarose gel (2.5%), stained with ethidium bromide or syber gold and photographed with a Polaroid film under an Ultra Violet transilluminator. The results indicate that the bisexual and parthenogenetic Artemia individuals from Urmia lake display identical restriction patterns, which correspond with the typical restriction pattern of parthenogenetic strains. (Bossier et al., 2003, in publication), According to Abreu-Grobois & Beardmore (1991) the phylogenetic relationship of Artemia urmiana with parthenogenetic forms is suggestive of a recent single common ancestral lineage branching to both A. urmiana and the asexual forms. Browne et al (1991), on the other hand, suggested that parthenogenetic Artemia has evolved from Artemia urmiana. These findings corroborate with the mitochondrial DNA analysis. In the future it will be interesting to use, in addition to mitochondrial markers, nuclear markers to analyse individuals from lake Urmia and its neighbourhood. The presence of parthenogenetic Artemia within the Lake Urmia and at close vicinity to it is a valuable indication to show the probable link between the different populations. The emergence of a dominating parthenogenetic population of Artemia in several ponds which were inoculated with nauplii hatched from Urmia Lake cysts collected in the previous season (Naser Agh, unpublished data) is another interesting phenomenon.These findings and the close genetic relationships between the bisexual Artemia urmiana and the parthenogenetic populations from the Old World, strongly supports the idea of Lake Urmia could be the probable source for expansion of parthenogenetic Artemia on one hand and that these asexual forms are derived from A. urmiana on the other. The results obtained through our field and laboratory experiments indicate that the historical evolution of parthenogenetic Artemia from the bisexual species (Artemia urmiana) could have occurred as a result of great reductions in water salinity rather then increased salinity levels as proposed by Hsu, et al. (1972) and Beardmore & Abreu-Grobois (1983). The existence of thriving parthenogenetic populations in temporary brackish water bodies with salinities as low as 10 g/l, is another strong evidence that Artemia can survive, grow and get localized at such low saline waters in absence of predators. Literature cited- Abreu-Grobois, F. A. & Beardmore, J. A. (1991). Genetic characterization and intra-genetic relationships of Artemia monica and Artemia urmiana Günther. Hydrobiologia, 212: 151-168. - Agh, N. and Noori, F. (1997). Introduction of a parthenogenetic population of Artemia from lagoons around Urmia Lake and its morphological comparison with Artemia urmiana. First Iranian Congress of Zoology, University of Teacher Education (Tarbiat Moellem), 17-18 Sep. 1997, Tehran, Iran. - Agh, N. (2002). Co-existence of bisexual and parthenogenetic populations of Artemia at Urmia Lake region. China regional workshop on Artemia. China National Institute of Salt Research. 23-26 Sep. 2002, Beijing, China. - Azari Takami, G., 1989. Two strains of Artemia in Urmia Lake (Iran). Artemia Newsletter. 13:5.

- Badaracco, G., Baratelli, L., Ginelli, E., Meneveri, R., Plevani, P., Valsasnini, P. and Barigozzi, C. (1987). Variations in repetitive DNA and heterochromatin in genus Artemia. Chromosoma, 95: 71-75. - Barigozzi, C., Varotto, V., Baratelli, L. and Giarrizzo, R. (1987a). The Artemia of Urmia Lake (Iran): mode of reproduction and chromosome numbers. Atti Acc. Lincei Rend. Fis. (8), LXXXI, pp. 87-90. - Barigozzi, C., Valsasnini, P., Ginelli, E., Badaracco, G., Pelvani, P. and Baratelli, L. (1987b). Further data on repetitive DNA and speciation in Artemia. In: Artemia Research and its applications. Vol. 1. Morphology, Genetics, Strain characterization, Toxicology. P. Sorgeloos, D. A. Bengtson, W. Decleir, and E. Jaspers (Eds). Universa Press, Wetteren, Belgium. pp. 103-105. - Barigozzi, C. (1988). The Artemia of Urmia Lake in Iran. Artemia Newsletter. 11:9. - Beardmore, J. A. and Abreu-Grobois, F. A. (1983). Taxonomy and Evolution in the Brine Shrimp Artemia. In: G. S. Oxford, G. S. and Rollinson (Eds), Protein Polymorphism: Adaptive and Taxonomic Significance. The Systematics Association Special Volume No. 24,. Academic Press, London and New York. pp. 153-164. - Browne, R. A., Li, M., Wanigasekara, G., Simonek, S., Brownlee, D., Eiband, E. and Cowan, J. (1991). Ecological and genetic divergence of sexual and asexual (diploid and polyploidy) brine shrimp (Artemia). Advances in ecology,1, pp. 41-52. - Clark, L.S. & Bowen, S.T. (1976). The genetics of Artemia salina. VII Reproductive isolation. J. Heredity, 67, pp. 385-388. - Gunther, R. T., 1890. Contributions to natural history of Lake Urmi, N. W. Persia and its neighbourhood. J. Linn. Soc. London 27: 394-398. - Hsu, K. J., Montadert, L., Bernoulli, D., Cita, M. B., Garrison, R. E., Kidd, R. B., Melieres, F., Meller, C. & Wright , R. (1972). Nature, 78:399-401. - Sorgeloos, P., (1997) Lake Urmia cooperation project – contract item A, Report on the determination and identification of biological characteristics of Artemia urmiana for application in aquaculture. Faculty of agriculture and applied biological science, Laboratory of aquaculture And Artemia reference center, Gent University, Belgium, p. 6–16 . - Van Stappen G. (2002). Zoogeography. In: Artemia: Basic and Applied Biology. Chapter IV.
Th. J. Abatzopoulos et al. (Eds). Kluwer Academic Publisher. pp. 171-224.
INCO partner: 12
MORPHOLOGICAL AND BIOCHEMISTRY CHARACTERIZATION OF MEXICANMORPHOLOGICAL AND BIOCHEMISTRY CHARACTERIZATION OF MEXICAN POPULATIONS OF POPULATIONS OF ARTEMIA FRANCISCANAARTEMIA FRANCISCANA AS WELL AS REPRODUCTIVE ISOLATION AS WELL AS REPRODUCTIVE ISOLATION AND BIOENCAPSULACIÓN OF MEDICATIONS AND BIOENCAPSULACIÓN OF MEDICATIONS
Jorge Castro, Aída Malpica, Germán Castro, Thalía Castro y Ramón De Lara
Departamento El Hombre y su Ambiente. División de Ciencias Biológicas y de la Salud. Universidad Autónoma Metropolitana-Xochimilco. Distrito Federal, México.
Tel(52) (5) 5483-7151; Fax (52) (5) 5483-7469Tel(52) (5) 5483-7151; Fax (52) (5) 5483-7469Email:[email protected]

This morphological study was carried out for seven Artemia populations: four from coastal waters: Yavaros, Sonora (YAV); Ohuira, Sinaloa (OHUI); Juchitán, Oaxaca (JUCH); Real de las Salinas, Campeche (RSAL); and three from inland waters: Cuatro Ciénegas, Coahuila (CCIEN); Las Salinas de Hidalgo, San Luis Potosí (SLP) and Texcoco, Estado de Mexico (TEX). Hydrated cysts; decapsulated embryos; nauplii and male and female adults were studied. Some 100 data were taken for each developmental stage and were compared to each other to determine significant differences (P < 0.5). If there was, a significant difference, Bonferroni and/or Tukey tests were applied to determine exactly how similar or different the populations were. A discriminating analysis for all the stages, using SYSTAT 9.0 was also carried out.
For reproductive isolation, 25 repetitions were made for each cross breeding. The produced nauplii (F1) were cultivated until reached the adult stage and then were submitted to a second cross (F2) with five repetitions.
Results: the population that presented the largest diameter in the hydrated cyst was JUCH (275. 52 µm) and the smallest YAV (229.20 µm + 8.97). The smallest value of chorion thickness was found in the population from SLP (6.43 µm) and the largest CCIEN (9.15 µm) as well.The smallest nauplius was OHUI (379.70 µm + 19.47µm).and the largest CCIEN (472.41 26.96 Nm)
The morphology of the adults (male/female) shows that the JUCH population presented the largest values in all the morphological structures. The discriminating analysis shows that the structures that mainly discriminate for males are: the left and right knobs; head and width of abdomen ; for females: the length of furcae and antennule; width of the abdomen and separation between the eyes.
The success of cross breeding was: YAV-OHUI (for males as well as for females); male CCIEN-female YAV; male CCIEN-female OHUI; male TEX-female OHUI; male CCIEN-female JUCH and male JUCH-female TEX.
We can conclude that Mexican strains are starting to separate since they present specific morphological characteristics, and also that Mexican strains present pre-reproductive problems such as morphological or sex attraction incompatibility, and post-reproductive problems (poor viability of hybrids).
Another conclusion is that Mexican strains present the beginning of reproductive isolation and consequently for some sub-specie formation.
The external morphology of the cyst and the membrane that covers the embryo from six Mexican populations of Artemia franciscana (SLP, CCIEN, TEX, YAV, RSAL, La Colorada, Oaxaca (COLOAX)),were examined under an electron scanning microscope. The photographic images showed that the external appearance of the embryonic membrane of the cysts in the six populations was similar; this membrane is formed by pentagonal and hexagonal structures, similar to ones found in other arthropods.
Also we carried out biochemical and molecular studies to determine the percentage of crude protein (Kjeldah method), total lipids (Sohxlet method) and total carbohydrates (Dubois method) in cyst and embryos from six Mexican populations. The fatty acids profile was determined by gas chromatography and amino acids by liquid and cationic interchange chromatography; the presence of cations in the chorion were take by dispersive energy spectrometry and X rays. The results showed that there were significant differences (P < 0.05) in the concentration of the valued compounds, even though it is not possible to establish a

biochemical pattern by geographical area with the Mexican Artemia strains. The cysts and embryos natives to TEX, and RSAL, presented the largest values in all the biochemical compounds. The composition of cations in the chorion indicates that there is direct relationship with the composition of the water.
At the present we are working with cysts, embryos and nauplii, to extract and to determine the protein content through electrophoretic profiles, in policrilamida gels under desnaturalized conditions. The molecular aspects were studied by using the amplification of specific fragments of DNA by polymer chain reaction (PCR), and determining the restriction fragments of polimorphic lenghts (RFPL). The biochemical and molecular results will be analyzed by numeric taxonomic and phylogenetic study.
Another aspect we have studied is the antibiotic incorporation in different Artemia stages to inhibit the growth of Aeromonas hidrophyla. Until know the following antibiotics in nauplii and metanauplii have been incorporated: Chloramphenicol, Nitrofurantoina (Macrodantina (50 mg capsules) and Ciprofloxacina (50 mg capsules mg). Several experiments have been carried out to determine the incorporation rate of each one of the antibiotics in the organisms. We obtained the calibration curves for each antibiotic to be able to determine the relationship that exists between the diameter of the halo inhibition and the concentration of the antibiotic. Bioassays were carried out using the plate diffusion technique. The results indicate that the halo inhibition in nauplii was: with Chloramphenicol 17.56 mm; with Ciprofloxacina; 10.33 mm and with Furantoina, 22.26 mm. In metanauplii, the diameter of halo inhibition was 15.40 mm in Chloramphenicol; 22.56mm in Ciprofloxacina; and with Nitrofurantoina the metanauplii did not show any halo inhibition. At present we are working with juvenile and adult stages.
INCO partner: 13INCO partner: 13
PRELIMINARY CHARACTERIZATION OF BRAZILIAN PRELIMINARY CHARACTERIZATION OF BRAZILIAN ARTEMIA FRANCISCANAARTEMIA FRANCISCANA POPULATIONS BY MEANS OF MTDNA RFLP ANALYSIS POPULATIONS BY MEANS OF MTDNA RFLP ANALYSIS
Marcos R. Camara1, Alexandros Triantafyllidis2, Athanasios D. Baxevanis2 & Theodore J. Abatzopoulos2
1Department of Oceanography and Limnology, Universidade Federal do Rio Grande do Norte, Natal, RN, 59072-970, Brazil
2Department of Genetics, Development and Molecular Biology, Aristotle University of Thessaloniki, 54124, Thessaloniki, Greece
Introduction
In this paper, a preliminary report on the molecular genetic diversity of three Artemia franciscana populations (Macau, Galinhos and Areia Branca/Grossos) from northeastern Brazil, by means of RFLP (Restriction Fragment Length Polymorphism) analysis of their mtDNA, is made. Data were achieved through several steps. First, these feral populations were screened in order to confirm if they all belonged to the Artemia franciscana superspecies. Following, their northern (San Francisco bay, California, USA) or southern (South America) origin was determined. And finally, the resulting composite genotypes were established. All experimental work was carried out during a three-month stay at the Laboratory of Genetics, Department of Genetics, Development and Molecular Biology of the Aristotle University of Thessaloniki, Greece.
Materials and methods
Individuals from three Artemia franciscana populations from the State of Rio Grande do Norte (RN) in northeastern Brazil were collected in January, 2003 from saltworks located in the municipalities of Macau (5º 06' S; 36º 38' W), Galinhos (5º 05' S; 36º 16' W), and Areia Branca/Grossos (4º 58' S; 37º 09' W), respectively. Approximately, 50-70 mature animals from each population were collected and kept in ethanol (> 90 %) prior to analysis.
Macau (MAC) was chosen as the site where Artemia franciscana was originally inoculated in RN in 1977 (Camara, 2001). Galinhos (GAL) and Areia Branca/Grossos (ABG) are equidistant (about 100 km) from Macau. In addition, Galinhos saltworks act as wintering grounds or passage areas for a diverse and abundant avifauna. Thus the possible role of migratory birds as contributors to gene flow within and between populations of Artemia in RN would be better estimated in Galinhos.
DNA was successfully extracted from 70 collected specimens using modified protocols for CTAB (Estoup et al., 1996) (65 extractions) and a simplified Chelex method (Tagart et al., 1992) (5 extractions) for total DNA isolation from Artemia. PCR amplification was carried out using an automatic thermocycler (Mastercycler EppendorfTM). The mtDNA target sequence was a segment of ca. 550 bp in length, which included a partial portion of the large subunit ribosomal RNA genes (16S rDNA) (Valverde et al., 1994). Genetic diversity (variability) of experimental populations was assessed by Restriction Fragment Length Polymorphism (RFLP) analysis of PCR amplified mitochondrial DNA (mtDNA). Four restriction enzymes (BioLabs Inc.) were used. These enzymes have already been found to give species-specific haplotypes for Artemia franciscana, differentiate its southern or northern origin and indicate variation within populations of Artemia franciscana (unpublished data).
Results and discussion
Fragment patterns observed among Brazilian Artemia populations in part of the 16S RNA region showed a consistent homogeneity as only one single composite haplotype (AAAA) occurred in all individuals scored.
Unpublished data have already shown that Artemia franciscana presents the same composite haplotype (AAAA) found in the present study. Thus, in spite of the relatively conservative region of the mtDNA studied and the limited number of restriction enzymes used, the RFLP data obtained in this preliminary characterization confirmed that the feral populations of Artemia franciscana found in the state of Rio Grande do Norte, northeastern Brazil, belong to the Artemia franciscana superspecies. In addition, their proposed origin, from San Francisco Bay cysts, was clearly demonstrated. This second information corroborates a previous report by Gajardo et al. (1995) based on allozyme evidence derived from the Macau population.
References
Camara, M. R., 2001. Dispersal of Artemia franciscana Kellogg (Crustacea; Anostraca) populations in the coastal saltworks of Rio Grande do Norte, northeastern Brazil. Hydrobiologia 466: 145-148.
Estoup, A. C. R. Largiadièr, E. Perrot & D. Chourrout, 1996. Rapid one-tube DNA extraction for reliable PCR detection of fish polymorphic markers and transgenes. Molecular Marine Biology and Biotechnology 5(4): 295-298.
Gajardo, G., M. Da Conceicao, L. Weber & J. A. Beardmore, 1995. Genetic variability and interpopulational differences in Artemia strains from South America. Hydrobiologia 302: 21-29.

Taggart, J. B., R. A. Hynes, P. A. Prodöhl & A. Ferguson, 1992. A simplified protocol for routine total DNA isolation from salmonid fishes. Journal of Fish Biology 40: 963-965.
Valverde, J.R., B. Batuecas, C. Moratilla, R. Marco & R. Garesse, 1994. The complete mitochondrial DNA sequence of the crustacean Artemia franciscana. Journal of Molecular Evolution 39, 400-408.
INCO partner: 14INCO partner: 14
PRELIMINARY ANALYSIS OF DNA SEQUENCES OF THE PRELIMINARY ANALYSIS OF DNA SEQUENCES OF THE CYTOCHROME C OXIDASECYTOCHROME C OXIDASE SUBUNIT I (COI) IN CHILEAN ARTEMIA POPULATIONS.SUBUNIT I (COI) IN CHILEAN ARTEMIA POPULATIONS.
Patricia Beristain 1,2, Stephan M. Funk 2 & Gonzalo Gajardo 1
1. Laboratory of Genetics & Aquaculture, Universidad de Los Lagos, Osorno, Chile.2. Zoological Society of London, Institute of Zoology, London.
According to possibilities we have considered different levels of analysis, from morphology to DNA, to understand the origin and evolutionary pattern of Artemia species from Chile and adjacent countries in the South American continent (Gajardo & Beardmore, 2001; Gajardo et al, 1995, 1998, 1999, 2000, 2001, 2002). Focus has been placed in the two New World sibling species, A. franciscana and A. persimilis, found in Chile and Argentina, both representing interesting models to evaluate the pattern of intraspecific genetic variation and differentiation as well as genetic differences at the species level. While these kind of multi-level studies are good to documenting Artemia biodiversity in the continent, the availability of a wide range of genetic markers facilitates the characterization, exploitation, management and conservation of valuable local genetic resources.The finding of A. persimilis in southern Chile, geographically segregated from A. franciscana, which is the dominant species in both the country and the continent, has raised interesting questions that we have investigated by using different approaches. Particularly, the genetic structure of A. franciscana has been studied throughout the species range, confronting data produced at different levels of analysis. This report considers preliminary findings of a study aimed at sequencing and comparing a 680bp fragment of the cytochrome C Oxydase subunit I (COI) in the Chilean Artemia populations.
Six Artemia samples from La Rinconada (RIN), Salar Llamara (LLA), Convento (CON), Pichilemu (PIC), Laguna Amarga (LAM) and Laguna de Los Cisnes (CIS), were considered as well as reference samples of A. franciscana (San Francisco Bay, SFB, USA; Macau, MAC, Brazil) and A. persimilis (ARC code 1321) (kindly provided as cysts by the Artemia Reference Center, ARC, Ghent). Adult female of each population (n=20) were fixed in absolute ethanol (99.9 %) until required for analysis.
DNA extractionDNA extractionDNA extraction from each individual followed the protocol of QIAampTM tissue extraction kit (Qiagen), whilst amplification of the 680-bp fragment of the mitochondrial (mt) cytochrome c oxidase subunit I was carried out in a reaction volume of 25 containing 2.5 mM Mg+2, 0.2 M of each dNTP’s, 0.2g of Bovine Serum Albumin (BSA), 0.25 unit of BiotaqTM polymerase, 1X of 10X PCR Buffer (QIAgen) and 0.25M of each primer: LCOI490 (5’-GGT-CAA-CCA-ATC-ATA-AAG-ATA-TTG-G-3’) and HCO2198 (5’TAA-ACT-TCA-GGG-TGA-CCA-AAA-AAT-CA-3’) (Folmer et al., 1994).
Sequencing the mtDNA cytochrome c oxidase subunit I of Artemia

The purified PCR products of each population (n=10) were subjected to sequencing reaction containing 0.67l ABI BigDye terminator v3.1, 3.3l Better Buffer, 2 l of LCOI490 or HCO2198, 2l of template adjusting the sample with double distillated water to a final volume to 10l. The thermal regime for sequencing consisted of 1cycle of 3 min at 96C and 30 cycles of 15 sec at 96C, 10 sec at 50C and 4 min at 60C. The DNA was precipitated with ethanol and the pellet were diluted in deionized formamide and loading buffer. The diluted samples were denaturated at 95C during 1.5 min and loaded on a polyacrylamida gel. The gel matrix was prepared using 12.6 gr urea, 3.5l longer ranger, 3.5l 1XTBE and 18.2l double distillated water. To gel solution, 180l of APS and 18l of TEMED were added. The products were separated electrophoretically using an ABI PRISM ™377 DNA sequencer (Perkin Elmer).
Results and discussion
Fig. 1 shows gels with the identification of the 680 pb COI fragment in the Chilean and the reference Artemia samples considered, whilst Fig. 2 shows the sequence of a partial fragment in an individual from Convento, a coastal lagoone in Central Chile. The statistical analysis of the full sequence in all samples will be produced shortly, and these results will complement those done in collaboration with the ARC (Dr. Peter Bossier).
Figure 1: PCR of Artemia samples
Cloning of Artemia samples from Pichilemu
Since not all individuals from Pichilemu showed a clear sequence pattern on the 680 bp fragment, PCR products of this sample sample were inserted into the bacterial cells by using TA Cloning® kit (Invitogen) and replicated. After several cycles of ligation/transformation the transformed cells containing the inserted DNA sequence were selected (Fig. 2)
Mitochondrial DNA is a valuable marker to indicate maternal gene flow since the somewhatMitochondrial DNA is a valuable marker to indicate maternal gene flow since the somewhat confounding effect of recombination is avoided (Avise, 1994). Hence, the level of variabilityconfounding effect of recombination is avoided (Avise, 1994). Hence, the level of variability between samples depends on their degree of genetic isolation, and this turns out to be abetween samples depends on their degree of genetic isolation, and this turns out to be a powerful tool when closely related (sibling) species like powerful tool when closely related (sibling) species like A. franciscanaA. franciscana and and A. persimilis A. persimilis areare compared. As noted by Bossier (partner 1, 3) and also in aphids (Sunnucks compared. As noted by Bossier (partner 1, 3) and also in aphids (Sunnucks et al., et al., 1996), a1996), a confounding factor in the use of mt DNA is the presence of multiple copies of certain mtDNAconfounding factor in the use of mt DNA is the presence of multiple copies of certain mtDNA genes, which is reflected in a double-banding pattern, or as an unexpected sequence pattern.genes, which is reflected in a double-banding pattern, or as an unexpected sequence pattern. The fact that the sample from Pichilemu exhibited a weird pattern needs further attention, butThe fact that the sample from Pichilemu exhibited a weird pattern needs further attention, but also confirms our previous findings showing this is a key location that seems to set be aalso confirms our previous findings showing this is a key location that seems to set be a
Pichilemu 1-8 SFB 1-8Pichilemu 1-8 SFB 1-8

natural barrier for the southward distribution natural barrier for the southward distribution of A, franciscanaof A, franciscana and and A. persimilisA. persimilis nortward. The nortward. The use of new genetic markers will contribute to further investigate this transitional zone, whichuse of new genetic markers will contribute to further investigate this transitional zone, which was among the problems highlighted in the China workshop.was among the problems highlighted in the China workshop.
Figure 2: COI Sequence fragment of 500bp of Artemia from Convento
References Avise, J.C. (1994) Molecular markers, natural history and evolution. New York, Chapman &
Hall. Cox, A. and Hebert, P. 2001. Colonization, extinction and phylogeographic patterning in a
freshwater crustacean. Molecular ecology 10: 371-386. Folmer, O., Black, M., Hoeh, W., Lutz, R. and Vrijenhoek, R. 1994. DNA primers for
amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3: 294-299.
Lavens, P. and Sorgeloos, P. 1996. Manual on the Production and Use of Live Food for aquaculture. FAO Fisheries Technical Paper 361.
Sunnucks, P., England, P.E., Taylor, A.C. & Hales, D.F. (1996) Microsatellite and chromosome evolution of parthenogenetic Sitobion aphids in Australia. Genetics 144, 747–756.
Related Documents











