Arsenic programmes cellular genomic-immunity through miR-2909 RNomics Deepak Kaul ⁎, S. Sharma, M. Sharma, M. Arora, Mansi Arora Department of Experimental Medicine & Biotechnology, Post-graduate Institute of Medical Education & Research, Chandigarh 160012, India abstract article info Article history: Accepted 2 December 2013 Available online 18 December 2013 Keywords: miR-2909 Gene regulation APOBEC3G IFNβ Viral-RNA sensing It is widely recognized that human cells are equipped with innate antiviral-RNA armour involving the production of type I interferons and APOBEC3G (apolipoprotein B mRNA-editing, enzyme-catalytic, polypeptide-like 3G) gene-product. Although arsenic has been shown to have paradoxical effect on one arm of this armour involving APOBEC3G, the exact molecular mechanism of its action in this regard is far from clear. The present study, ad- dressed to explore as to how arsenic programmes this innate antiviral-RNA cellular-sensing pathway, clearly re- vealed that arsenic programmes this innate cellular antiviral genomic response through its inherent capacity to initiate cellular miR-2909 RNomics pathway, involving not only the modulation of APOBEC3G gene but also KLF4 (Kruppel-like factor 4) dependent regulation of gene coding for IKBKε (Inhibitor of nuclear factor kappa-B kinase subunit epsilon) which in turn modulates RIG-I (retinoic acid-inducible gene 1) pathway responsible for the pro- duction of IFNβ (interferon beta) through restriction of CYLD (Cylindromatosis) deubiqutinating activity. This re- stricted inhibitory enzyme activity of CYLD upon NFkB (nuclear factor kappa-light-chain-enhancer of activated B cells) also ensures sustained expression of miR-2909. Our results for the first time show that cellular miR-2909 RNomics may constitute an innate genomic armour to promote as well as restrict retroviral infection. © 2013 Elsevier B.V. All rights reserved. 1. Introduction Extensive studies have shown that host microRNAs (miRNAs) regu- late fundamental genetic programmes responsible for providing cells with sensing mechanism against invading RNA viruses (tenOever, 2013). This cellular sensing mechanism has been widely recognized to be provided by both the RIG-I/MAVS pathway involving type-1 inter- ferons (Kato et al., 2005; Rehwinkel et al., 2010) and the induction of APOBEC3G gene product that lethally hyper mutates viral RNA (Russell et al., 2009). In this context, it is interesting to note that the tu- mour suppressor ‘CYLD’, a deubiqutinating enzyme has not only been shown to inhibit the IRF3 (Interferon regulatory factor 3) signalling pathway and type-1 interferon production triggered by RIG-I (Friedman et al., 2008) but its expression was also found to be down- regulated upon viral infection. Interestingly, ability of CYLD to deubiquitinate and consequently down-regulate RIG-I is restricted by its own phosphorylation via IKBKε (Zhang et al., 2008). Recently arsenic trioxide, used to treat acute promyelocytic leukaemia, was shown to counteract the APOBEC3G mediated restriction of retroviral (HIV-1, in particular) infection in dendritic cells but not in the more permissive CD4 + T cells (Chou et al., 2005; Stalder et al., 2010). Although CYLD is documented to inhibit NFkB binding activity (Kovalenko et al., 2003), arsenic is known to promote such an activity in order to regulate the ex- pression of its various target proto-oncogenes (Liao et al., 2004). A re- cent study from our lab suggests a novel epigenetic mechanism of oncogenesis involving arsenic trioxide dependent miR-2909 RNomics (Sharma et al., 2013). Also, it was shown that chronic exposure to low dose arsenic significantly altered the gene and protein expression of many regulators of innate immunity and led to a compromised re- sponse to a subsequent immune challenge (Kozul et al., 2009). Keeping in view all the above mentioned features of arsenic biology, the present study was addressed to understand the detailed mechanism through which arsenic could programme the genomic immunity in- volving the interplay between arsenic-induced cellular miR-2909 expression and the genes involved in the regulatory pathway re- sponsible for the generation of type I interferons and APOBEC3G expression. 2. Materials & methods 2.1. Bioinformatics analysis Promoter sequences for relevant genes were obtained from Mamma- lian Promoter sequence database (http://rulai.cshl.edu/CSHLmpd2/). Putative binding sites for various transcription factors were retrieved from JASPAR database (http://jaspar.cgb.ki.se/) at a default threshold Gene 536 (2014) 326–331 Abbreviations: miRNA, MicroRNA; PBMCs, Peripheral blood mononuclear cells; CYLD, Cylindromatosis; KLF4, Kruppel-like factor4; NFκB, Nuclear factor kappa B; FCS, Foetal Calf Serum; As, Arsenic; IFNβ, Interferon beta; APOBEC3G, Apolipoprotein B mRNA-editing, enzyme-catalytic, polypeptide-like 3G; IKBKε, Inhibitor of nuclear factor kappa-B kinase subunit epsilon; RIG1, Retinoic acid-inducible gene 1; IRF3, Interferon regulatory factor 3. ⁎ Corresponding author. Fax: +91 172 2744401. E-mail address: [email protected] (D. Kaul). 0378-1119/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.gene.2013.12.004 Contents lists available at ScienceDirect Gene journal homepage: www.elsevier.com/locate/gene

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Gene 536 (2014) 326–331
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
Arsenic programmes cellular genomic-immunity throughmiR-2909 RNomics
Deepak Kaul ⁎, S. Sharma, M. Sharma, M. Arora, Mansi AroraDepartment of Experimental Medicine & Biotechnology, Post-graduate Institute of Medical Education & Research, Chandigarh 160012, India
Abbreviations:miRNA, MicroRNA; PBMCs, PeripheralCylindromatosis; KLF4, Kruppel-like factor4; NFκB, NucleaSerum; As, Arsenic; IFNβ, Interferon beta; APOBEC3G, Aenzyme-catalytic, polypeptide-like 3G; IKBKε, Inhibitor osubunit epsilon; RIG1, Retinoic acid-inducible gene 1; IRF3⁎ Corresponding author. Fax: +91 172 2744401.
E-mail address: [email protected] (D. Kaul).
0378-1119/$ – see front matter © 2013 Elsevier B.V. All rhttp://dx.doi.org/10.1016/j.gene.2013.12.004
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 2 December 2013Available online 18 December 2013
Keywords:miR-2909Gene regulationAPOBEC3GIFNβViral-RNA sensing
It iswidely recognized that human cells are equippedwith innate antiviral-RNA armour involving the productionof type I interferons and APOBEC3G (apolipoprotein B mRNA-editing, enzyme-catalytic, polypeptide-like 3G)gene-product. Although arsenic has been shown to have paradoxical effect on one arm of this armour involvingAPOBEC3G, the exact molecular mechanism of its action in this regard is far from clear. The present study, ad-dressed to explore as to how arsenic programmes this innate antiviral-RNA cellular-sensing pathway, clearly re-vealed that arsenic programmes this innate cellular antiviral genomic response through its inherent capacity toinitiate cellularmiR-2909 RNomics pathway, involving not only themodulation of APOBEC3G gene but also KLF4(Kruppel-like factor 4) dependent regulation of gene coding for IKBKε (Inhibitor of nuclear factor kappa-B kinasesubunit epsilon)which in turnmodulates RIG-I (retinoic acid-inducible gene 1) pathway responsible for the pro-duction of IFNβ (interferon beta) through restriction of CYLD (Cylindromatosis) deubiqutinating activity. This re-stricted inhibitory enzyme activity of CYLD upon NFkB (nuclear factor kappa-light-chain-enhancer of activated Bcells) also ensures sustained expression of miR-2909. Our results for the first time show that cellular miR-2909RNomics may constitute an innate genomic armour to promote as well as restrict retroviral infection.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Extensive studies have shown that host microRNAs (miRNAs) regu-late fundamental genetic programmes responsible for providing cellswith sensing mechanism against invading RNA viruses (tenOever,2013). This cellular sensing mechanism has been widely recognized tobe provided by both the RIG-I/MAVS pathway involving type-1 inter-ferons (Kato et al., 2005; Rehwinkel et al., 2010) and the induction ofAPOBEC3G gene product that lethally hyper mutates viral RNA(Russell et al., 2009). In this context, it is interesting to note that the tu-mour suppressor ‘CYLD’, a deubiqutinating enzyme has not only beenshown to inhibit the IRF3 (Interferon regulatory factor 3) signallingpathway and type-1 interferon production triggered by RIG-I(Friedman et al., 2008) but its expression was also found to be down-regulated upon viral infection. Interestingly, ability of CYLD todeubiquitinate and consequently down-regulate RIG-I is restricted byits own phosphorylation via IKBKε (Zhang et al., 2008). Recently arsenictrioxide, used to treat acute promyelocytic leukaemia, was shown tocounteract the APOBEC3G mediated restriction of retroviral (HIV-1, in
blood mononuclear cells; CYLD,r factor kappa B; FCS, Foetal Calfpolipoprotein B mRNA-editing,f nuclear factor kappa-B kinase, Interferon regulatory factor 3.
ights reserved.
particular) infection in dendritic cells but not in the more permissiveCD4+ T cells (Chou et al., 2005; Stalder et al., 2010). Although CYLD isdocumented to inhibit NFkB binding activity (Kovalenko et al., 2003),arsenic is known to promote such an activity in order to regulate the ex-pression of its various target proto-oncogenes (Liao et al., 2004). A re-cent study from our lab suggests a novel epigenetic mechanism ofoncogenesis involving arsenic trioxide dependent miR-2909 RNomics(Sharma et al., 2013). Also, it was shown that chronic exposure to lowdose arsenic significantly altered the gene and protein expression ofmany regulators of innate immunity and led to a compromised re-sponse to a subsequent immune challenge (Kozul et al., 2009).
Keeping in view all the above mentioned features of arsenic biology,the present studywas addressed to understand the detailedmechanismthrough which arsenic could programme the genomic immunity in-volving the interplay between arsenic-induced cellular miR-2909expression and the genes involved in the regulatory pathway re-sponsible for the generation of type I interferons and APOBEC3Gexpression.
2. Materials & methods
2.1. Bioinformatics analysis
Promoter sequences for relevant geneswere obtained fromMamma-lian Promoter sequence database (http://rulai.cshl.edu/CSHLmpd2/).Putative binding sites for various transcription factors were retrievedfrom JASPAR database (http://jaspar.cgb.ki.se/) at a default threshold
327D. Kaul et al. / Gene 536 (2014) 326–331
score of 85.0. The presence of putative target site of miR-2909 on 3′UTRof KLF-4 gene was explored bioinformatically using RNA Hybrid tool(http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/).
2.2. Cellular model employed
Human peripheral blood mononuclear cells (PBMCs) were obtainedfrom 25 normal healthy volunteers (with their prior informed consent),who were fasting for 12 h or had abstained from any medication for2 weeks before blood donation and subsequently these cells were cul-tured in vitro as per the standard procedure reported by us earlier(Sharma et al., 2013).
2.3. Cell transfection studies
miR-2909 knock-down PBMCs were generated by transfecting mer-cury LNA™miR-2909 inhibitor (EXIQON) using lipofectamine transfec-tion reagent (Invitrogen) and subsequently these PBMCs were exposedto medium enriched with or without sodium arsenite (2 μM) and incu-bated for 36 h at 37 °C in 5% CO2 atmosphere. KLF4 expression plasmid(Addgene plasmid 17967) (Lowry et al., 2008) was transfected intoPBMCs using escort transfection reagent (Sigma) and incubated for48 h at 37 °C in 5% CO2 atmosphere. At the end of incubation period,the cells from each well were processed for RNA and protein isolation.
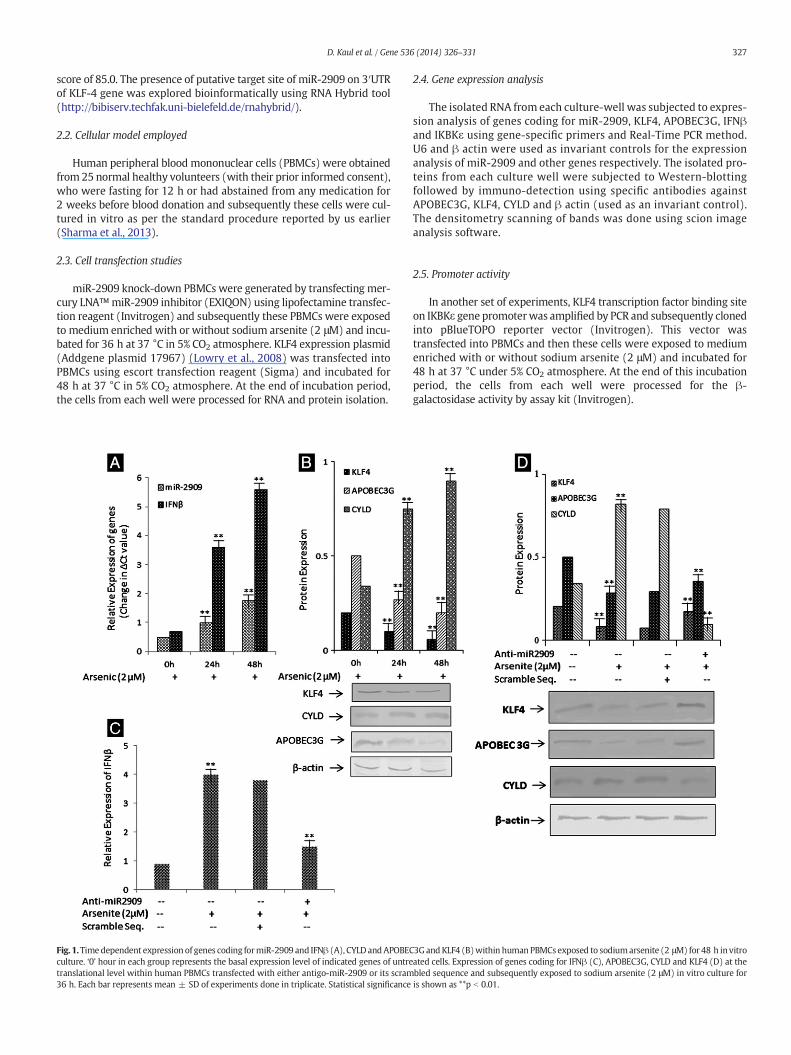
Fig. 1.Timedependent expression of genes coding formiR-2909 and IFNβ (A), CYLD andAPOBECculture. ‘0’ hour in each group represents the basal expression level of indicated genes of untretranslational level within human PBMCs transfected with either antigo-miR-2909 or its scram36 h. Each bar represents mean ± SD of experiments done in triplicate. Statistical significance
2.4. Gene expression analysis
The isolated RNA from each culture-well was subjected to expres-sion analysis of genes coding for miR-2909, KLF4, APOBEC3G, IFNβand IKBKε using gene-specific primers and Real-Time PCR method.U6 and β actin were used as invariant controls for the expressionanalysis of miR-2909 and other genes respectively. The isolated pro-teins from each culture well were subjected to Western-blottingfollowed by immuno-detection using specific antibodies againstAPOBEC3G, KLF4, CYLD and β actin (used as an invariant control).The densitometry scanning of bands was done using scion imageanalysis software.
2.5. Promoter activity
In another set of experiments, KLF4 transcription factor binding siteon IKBKε gene promoterwas amplified by PCR and subsequently clonedinto pBlueTOPO reporter vector (Invitrogen). This vector wastransfected into PBMCs and then these cells were exposed to mediumenriched with or without sodium arsenite (2 μM) and incubated for48 h at 37 °C under 5% CO2 atmosphere. At the end of this incubationperiod, the cells from each well were processed for the β-galactosidase activity by assay kit (Invitrogen).
3GandKLF4 (B)within human PBMCs exposed to sodiumarsenite (2 μM) for 48 h invitroated cells. Expression of genes coding for IFNβ (C), APOBEC3G, CYLD and KLF4 (D) at thebled sequence and subsequently exposed to sodium arsenite (2 μM) in vitro culture foris shown as **p b 0.01.

328 D. Kaul et al. / Gene 536 (2014) 326–331
2.6. In vitro miRNA target analysis
Amplicons corresponding to the target site ofmiR-2909 on 3′UTR se-quence of KLF4 gene were amplified and cloned in the miRNASelect™pMIR-GFP Reporter System (Cell BioLabs). The plasmid containing theinserted target site was then transfected into HEK293 cells (Havinghigh intrinsic expression of miR-2909) and these cells were maintainedup to 36 h at 37 °C under 5% CO2 atmosphere. At the end of incubationperiod, the cells were analysed for GFP expression, through microscopyas well as by quantification using flow cytometer.
2.7. Co-immunoprecipitation (Co-IP)
Human PBMCswere exposed to fixed concentration of 2 μMof sodi-um arsenite up to 48 h at 37 °C. The cells were harvested and processedfor CYLD protein immuno-precipitation using ice cold non denaturingcell lysis buffer. Following IP, protein was immuno-detected withPhospho-CYLD antibody (cell signalling technology) and β-actin wasused as an invariant control.
2.8. Statistical analysis
Spearman rank correlation coefficient was calculated for time de-pendent arsenic effect on expression of all genes. Analysis was per-formed with the use of SPSS windows version 19. p value less than0.01 was taken to be significant.
3. Results
3.1. Gene regulation by arsenic
Exposure of human blood mononuclear cells (PBMCs) maintainedin vitro culture to sodium arsenite (2 μM) for 48 h resulted in time de-pendent increase inmiR-2909 expression and this phenomenonwas ac-companied by time dependent increase in the expression of genescoding for IFNβ and CYLD (Fig. 1A, B) as well as down-regulation of
Fig. 2. Inter-gene relationship between miR-2909 expression and the express
genes coding for KLF4 and APOBEC3G (Fig. 1B). In order to ascertainthe contribution of miR-2909 in arsenic modulated gene expression,we repeated the above mentioned experiments in the presence ofantigo-miR-2909 and the results of such experiments indicated thatthe arsenic dependent regulation of genes coding for IFNβ (Fig. 1C),KLF4, APOBEC3G and CYLD (Fig. 1D) was indeed mediated through itscapacity to induce miR-2909 expression. This view was strengthenedby the inter-gene correlation betweenmiR-2909 expression and the ex-pression of genes coding for IFNβ, KLF4, APOBEC3G and CYLD (Fig. 2).
3.2. miR-2909 targets KLF4 gene
In order to understand as to how miR-2909 regulates the cellulargenes, we carried out the target analysis of miR-2909 upon human ge-nome using bioinformatic tools. It was interesting to note that miR-2909 could conspicuously target the 3′UTR region of KLF4 gene(Fig. 3A) and hence such a possibility was experimentally verified bytransfecting the reporter plasmid (carrying the 3′UTR target sequenceof KLF4 gene) into HEK293 cells, which have intrinsically high miR-2909 expression (Fig. 3C). As compared to control cells, we observed re-duced expression of green fluorescent gene marker in the cellstransfected with this reporter plasmid (carrying the 3′UTR of KLF4gene) as determined by fluorescence microscopy (Fig. 3B) and quanti-tated byflow cytometry (Fig. 3B, D). The results of such a study revealedthat miR-2909 has the inherent capacity to down regulate KLF4 gene.Hence, it became pertinent to explore the contribution of KLF4 in miR-2909 mediated regulation of cellular genes widely recognized to be in-volved in viral RNA sensing pathway, i.e. IFNβ, APOBEC3G and CYLD.
3.3. KLF4 dependent gene regulation
In order to verify if KLF4 gene contributes to the regulation of genesinvolved in viral sensing pathway (i.e. APOBEC3G, IFNβ and CYLD), wetransfected human PBMCs with KLF4 expression plasmid and checkedthe expression of these genes. We found significant decrease in the ex-pression of IFNβ but insignificant change in the levels of CYLD and
ion of genes coding for APOBEC3G (A), KLF4 (B), IFNβ (C) and CYLD (D).
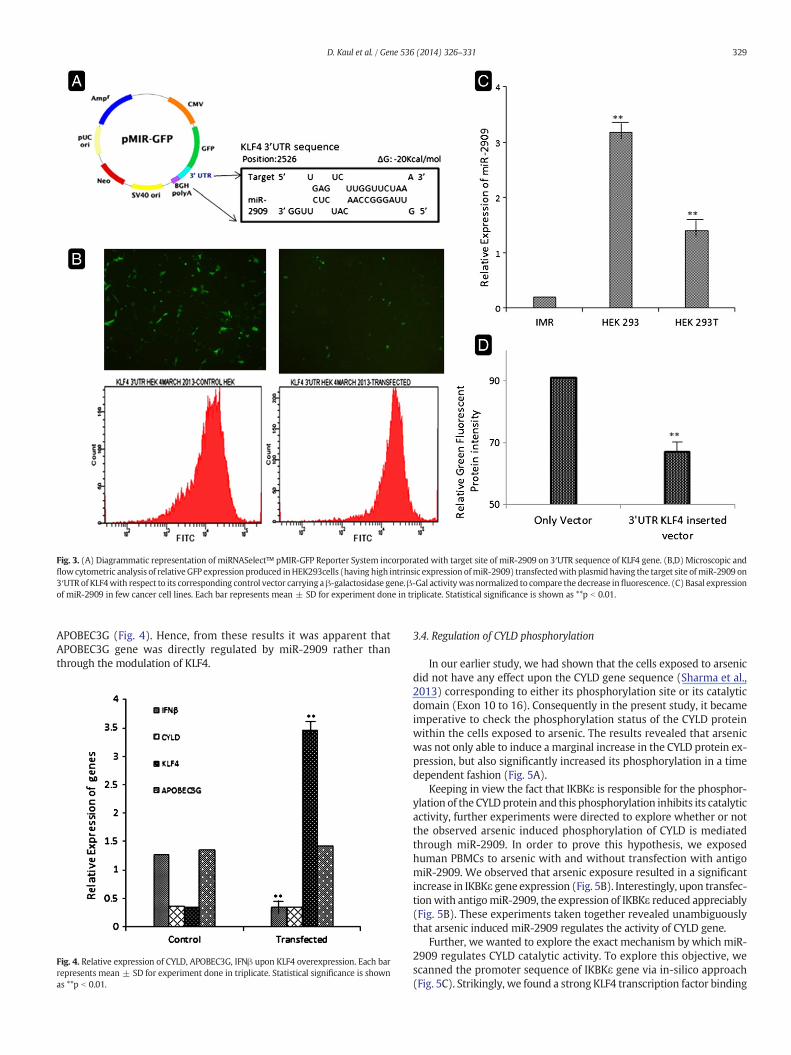
Fig. 3. (A) Diagrammatic representation of miRNASelect™ pMIR-GFP Reporter System incorporated with target site of miR-2909 on 3′UTR sequence of KLF4 gene. (B,D) Microscopic andflowcytometric analysis of relativeGFP expression produced inHEK293cells (having high intrinsic expression ofmiR-2909) transfectedwith plasmidhaving the target site ofmiR-2909 on3′UTR of KLF4with respect to its corresponding control vector carrying aβ-galactosidase gene.β-Gal activitywas normalized to compare the decrease influorescence. (C) Basal expressionof miR-2909 in few cancer cell lines. Each bar represents mean ± SD for experiment done in triplicate. Statistical significance is shown as **p b 0.01.
329D. Kaul et al. / Gene 536 (2014) 326–331
APOBEC3G (Fig. 4). Hence, from these results it was apparent thatAPOBEC3G gene was directly regulated by miR-2909 rather thanthrough the modulation of KLF4.
Fig. 4. Relative expression of CYLD, APOBEC3G, IFNβ upon KLF4 overexpression. Each barrepresents mean ± SD for experiment done in triplicate. Statistical significance is shownas **p b 0.01.
3.4. Regulation of CYLD phosphorylation
In our earlier study, we had shown that the cells exposed to arsenicdid not have any effect upon the CYLD gene sequence (Sharma et al.,2013) corresponding to either its phosphorylation site or its catalyticdomain (Exon 10 to 16). Consequently in the present study, it becameimperative to check the phosphorylation status of the CYLD proteinwithin the cells exposed to arsenic. The results revealed that arsenicwas not only able to induce a marginal increase in the CYLD protein ex-pression, but also significantly increased its phosphorylation in a timedependent fashion (Fig. 5A).
Keeping in view the fact that IKBKε is responsible for the phosphor-ylation of the CYLDprotein and this phosphorylation inhibits its catalyticactivity, further experiments were directed to explore whether or notthe observed arsenic induced phosphorylation of CYLD is mediatedthrough miR-2909. In order to prove this hypothesis, we exposedhuman PBMCs to arsenic with and without transfection with antigomiR-2909. We observed that arsenic exposure resulted in a significantincrease in IKBKε gene expression (Fig. 5B). Interestingly, upon transfec-tionwith antigomiR-2909, the expression of IKBKε reduced appreciably(Fig. 5B). These experiments taken together revealed unambiguouslythat arsenic induced miR-2909 regulates the activity of CYLD gene.
Further, we wanted to explore the exact mechanism by which miR-2909 regulates CYLD catalytic activity. To explore this objective, wescanned the promoter sequence of IKBKε gene via in-silico approach(Fig. 5C). Strikingly, we found a strong KLF4 transcription factor binding
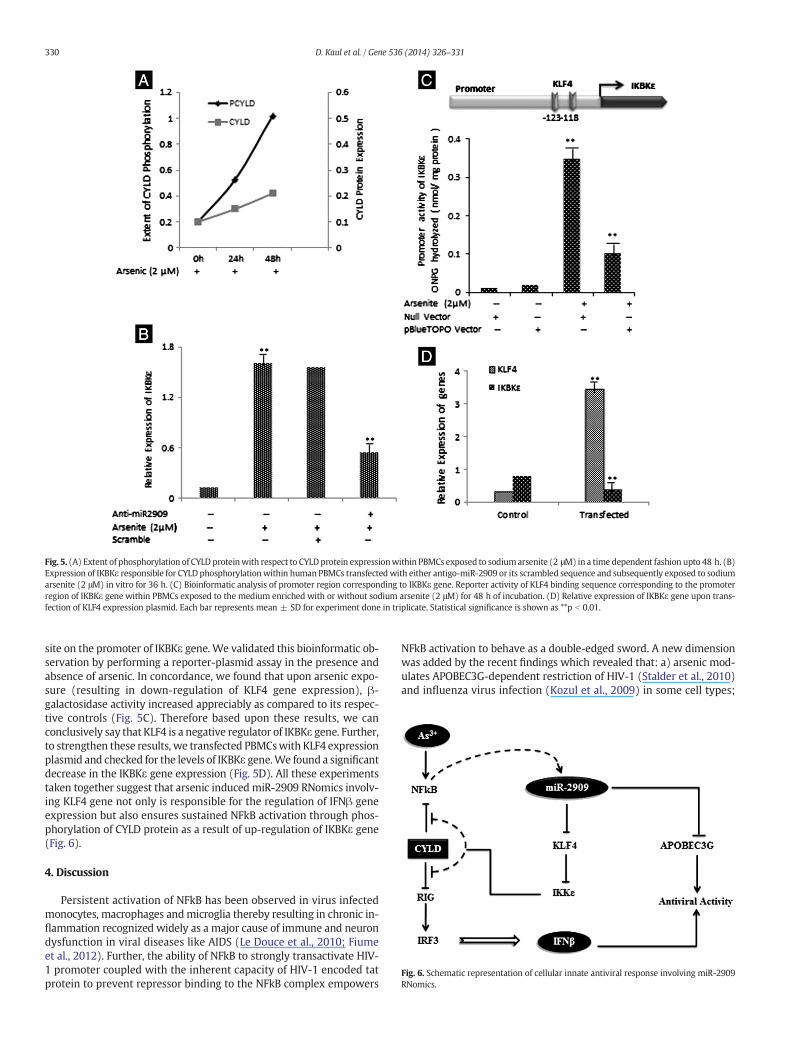
Fig. 5. (A) Extent of phosphorylation of CYLD proteinwith respect to CYLDprotein expressionwithin PBMCs exposed to sodiumarsenite (2 μM) in a time dependent fashion upto 48 h. (B)Expression of IKBKε responsible for CYLD phosphorylationwithin human PBMCs transfectedwith either antigo-miR-2909 or its scrambled sequence and subsequently exposed to sodiumarsenite (2 μM) in vitro for 36 h. (C) Bioinformatic analysis of promoter region corresponding to IKBKε gene. Reporter activity of KLF4 binding sequence corresponding to the promoterregion of IKBKε gene within PBMCs exposed to the medium enriched with or without sodium arsenite (2 μM) for 48 h of incubation. (D) Relative expression of IKBKε gene upon trans-fection of KLF4 expression plasmid. Each bar represents mean ± SD for experiment done in triplicate. Statistical significance is shown as **p b 0.01.
Fig. 6. Schematic representation of cellular innate antiviral response involving miR-2909RNomics.
330 D. Kaul et al. / Gene 536 (2014) 326–331
site on the promoter of IKBKε gene. We validated this bioinformatic ob-servation by performing a reporter-plasmid assay in the presence andabsence of arsenic. In concordance, we found that upon arsenic expo-sure (resulting in down-regulation of KLF4 gene expression), β-galactosidase activity increased appreciably as compared to its respec-tive controls (Fig. 5C). Therefore based upon these results, we canconclusively say that KLF4 is a negative regulator of IKBKε gene. Further,to strengthen these results, we transfected PBMCswith KLF4 expressionplasmid and checked for the levels of IKBKε gene.We found a significantdecrease in the IKBKε gene expression (Fig. 5D). All these experimentstaken together suggest that arsenic induced miR-2909 RNomics involv-ing KLF4 gene not only is responsible for the regulation of IFNβ geneexpression but also ensures sustained NFkB activation through phos-phorylation of CYLD protein as a result of up-regulation of IKBKε gene(Fig. 6).
4. Discussion
Persistent activation of NFkB has been observed in virus infectedmonocytes, macrophages andmicroglia thereby resulting in chronic in-flammation recognized widely as a major cause of immune and neurondysfunction in viral diseases like AIDS (Le Douce et al., 2010; Fiumeet al., 2012). Further, the ability of NFkB to strongly transactivate HIV-1 promoter coupled with the inherent capacity of HIV-1 encoded tatprotein to prevent repressor binding to the NFkB complex empowers
NFkB activation to behave as a double-edged sword. A new dimensionwas added by the recent findings which revealed that: a) arsenic mod-ulates APOBEC3G-dependent restriction of HIV-1 (Stalder et al., 2010)and influenza virus infection (Kozul et al., 2009) in some cell types;

331D. Kaul et al. / Gene 536 (2014) 326–331
b) arsenic has biphasic effect on NFkB binding activity therebyactivating its binding at low concentrationswhile inhibiting at high con-centrations; c) the deubiquitinating enzyme “CYLD” protein level is re-duced in the presence of viral infection and this enzyme inhibits NFkBactivity which, in turn, is known to upregulate the expression of CYLDgene (Kovalenko et al., 2003;Massoumi et al., 2006). Also, CYLD activityis down-regulated by its phosphorylation at Serine 418 by IKBKε (Zhanget al., 2008). It is in this context, the results reported here assume im-portance especially the novel observation that arsenic exerts its influ-ence upon the cellular ability to evoke an effective antiviral responseby regulating the genes coding for APOBEC3G and IFNβ through miR-2909 (Fig. 1A, B). This view was further strengthened by the ability ofantigo-miR-2909 to abolish themodulatory role of arsenic upon the cel-lular genes coding for APOBEC3G and IFNβ (Figure-1C, D). Further, theability of miR-2909 to repress KLF4 translational expression resultedin not only sustained activation of cellular NFkB and RIG-I responsiblefor IFNβ production but also sustained inhibition of CYLD activitythrough IKBKε gene up-regulation. Hence, arsenic induced cellularmiR-2909 expression may provide innate armour for host to combatviral infection in general and HIV-1 in particular. Our finding is in con-formity with the known inhibitory activity of KLF4 on the productionof CD8+ T cells (Wen et al., 2011). Hence, regulation of KLF4 by miR-2909 may provide a crucial pathway for HIV infection and its control.
5. Conclusion
The ability of arsenic to initiate cellular miR-2909 RNomics has re-vealed a novel pathway throughwhich the genomic immunity providedby IFNβ and APOBEC3G is programmed. Hence, such a pathway can beexploited for devising various futuristic strategies to combat viral infec-tion and HIV in particular. However, more elaborate investigation is re-quired before such a possibility could be widely recognized.
Conflict of interest statement
This is to certify that there is no conflict of interest includingany financial, personal or other relationships with other people or
organizations within three years from the date of submitted workin the form of present manuscript.
References
Chou, W.C., Chen, H.Y., Yu, S.L., Cheng, L., Yang, P.C., Dang, C.V., 2005. Arsenic suppressesgene expression in promyelocytic leukemia cells partly through Sp1 oxidation. Blood106, 304–310.
Fiume, G., et al., Apr 2012. Human immunodeficiency virus-1 Tat activates NF-κB viaphysical interaction with IκB-α and p65. Nucleic Acids Res. 40 (8), 3548–3562.
Friedman, C.S., et al., 2008. The tumour suppressor CYLD is a negative regulator ofRIG-I-mediated antiviral response. EMBO Rep. 9, 930–936.
Kato, H., et al., 2005. Cell type-specific involvement of RIG-I in antiviral response. Immu-nity 23, 19–28.
Kovalenko, A., Chable-Bessia, C., Cantarella, G., Israël, A., Wallach, D., Courtois, G., 2003.The tumour suppressor CYLD negatively regulates NF-kappaB signalling bydeubiquitination. Nature 424, 801–805.
Kozul, C.D., Ely, K.H., Enelow, R.I., Hamilton, J.W., 2009. Low-dose arsenic compromisesthe immune response to influenza A infection in vivo. Environ. Health Perspect.117, 1441–1447.
Le Douce, V., Herbein, G., Rohr, O., Schwartz, C., 2010. Molecular mechanisms of HIV-1persistence in the monocyte-macrophage lineage. Retrovirology. 7, 32.
Liao, W.T., Chang, K.L., Yu, C.L., Chen, G.S., Chang, L.W., Yu, H.S., 2004. Arsenic induceshuman keratinocyte apoptosis by the FAS/FAS ligand pathway, which correlateswith alterations in nuclear factor-kappa B and activator protein-1 activity. J. Invest.Dermatol. 122, 125–129.
Lowry, W.E., et al., 2008. Generation of human induced pluripotent stem cells from der-mal fibroblasts. Proc. Natl. Acad. Sci. U. S. A. 105, 2883–2888.
Massoumi, R., Chmielarska, K., Hennecke, K., Pfeifer, A., Fässler, R., 2006. Cyld inhibitstumor cell proliferation by blocking Bcl-3-dependent NF-kappaB signaling. Cell 125,665–677.
Rehwinkel, J., et al., 2010. RIG-I detects viral genomic RNA during negative-strand RNAvirus infection. Cell 140, 397–408.
Russell, R.A., Moore, M.D., Hu, W.S., Pathak, V.K., 2009. APOBEC3G induces ahypermutation gradient: purifying selection at multiple steps during HIV-1 replica-tion results in levels of G-to-A mutations that are high in DNA, intermediate in cellu-lar viral RNA, and low in virion RNA. Retrovirology 6, 16.
Sharma, M., Sharma, S., Arora, M., Kaul, D., 2013. Regulation of cellular Cyclin D1 gene byarsenic is mediated through miR-2909. Gene 522, 60–64.
Stalder, R., Blanchet, F., Mangeat, B., Piguet, V., 2010. Arsenic modulates APOBEC3G-mediated restriction to HIV-1 infection in myeloid dendritic cells. J. Leukoc. Biol. 88,1251–1258.
tenOever, B.R., 2013. RNA viruses and the host microRNA machinery. Nat. Rev. Microbiol.11, 169–180.
Wen, X., Liu, H., Xiao, G., Liu, X., 2011. Downregulation of the transcription factor KLF4 isrequired for the lineage commitment of T cells. Cell Res. 21, 1701–1710.
Zhang, M., et al., 2008. Regulation of IkappaB kinase-related kinases and antiviral re-sponses by tumor suppressor CYLD. J. Biol. Chem. 283, 18621–18626.
Related Documents











