Experimental Physiology/2008/043653-R1 Are L-glutamate and ATP cotransmitters of the peripheral chemoreflex in the nucleus tractus solitarius? Daniela Accorsi-Mendonça, Leni G.H. Bonagamba, Ricardo M. Leão and Benedito H. Machado* Department of Physiology, School of Medicine of Ribeirão Preto, University of São Paulo, 14049-900, Ribeirão Preto, SP, Brazil Running title: Glutamatergic and purinergic transmission in the NTS output neurons. Key words: glutamatergic, purinergic, NTS, sympathoexcitation, chemoreflex, RVLM * Corresponding author: Department of Physiology School of Medicine of Ribeirão Preto University of São Paulo 14049-900, Ribeirão Preto, SP, Brazil e-mail: [email protected] Phone: 55-16-3602-3015 FAX: 55-16-3633-0017 Physiology in Press; published online on October 22, 2008 as 10.1113/expphysiol.2008.043653 Copyright 2008 by The Physiological Society ) at CAPES - Usage on December 19, 2008 ep.physoc.org Downloaded from Exp Physiol (

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Experimental Physiology/2008/043653-R1
Are L-glutamate and ATP cotransmitters of the peripheral chemoreflex in the nucleus tractus solitarius?
Daniela Accorsi-Mendonça, Leni G.H. Bonagamba, Ricardo M. Leão and Benedito H. Machado*
Department of Physiology, School of Medicine of Ribeirão Preto, University of São Paulo, 14049-900, Ribeirão Preto, SP, Brazil
Running title: Glutamatergic and purinergic transmission in the NTS output neurons. Key words: glutamatergic, purinergic, NTS, sympathoexcitation, chemoreflex, RVLM * Corresponding author: Department of Physiology School of Medicine of Ribeirão Preto University of São Paulo 14049-900, Ribeirão Preto, SP, Brazil e-mail: [email protected] Phone: 55-16-3602-3015 FAX: 55-16-3633-0017
Physiology in Press; published online on October 22, 2008 as 10.1113/expphysiol.2008.043653
Copyright 2008 by The Physiological Society) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

2
ABSTRACT
Peripheral chemoreflex activation in awake rats or in the working heart brainstem
preparation (WHBP) produces sympathoexcitation, bradycardia and increase in the
frequency of phrenic nerve activity (PNA). Our focus is the neurotransmission of the
sympathoexcitatory component of the chemoreflex within the nucleus of tractus
solitarius (NTS) and recently we verified that the simultaneous antagonism of
ionotropic glutamate and purinergic P2 receptors in the NTS blocked pressor response
and increased thoracic sympathetic activity in awake rats and WHBP, respectively, in
response to peripheral chemoreflex activation. These previous data suggested the
involvement of ATP and L-glutamate in the NTS in the processing of the
sympathoexcitatory component of the chemoreflex by unknown mechanisms. For a
better understanding of these mechanisms, here we used a patch-clamp approach in
brainstem slices to evaluate the characteristics of the synaptic transmission of NTS
neurons sending projections to the ventral medulla, which include the pre-motor
neurons involved in the generation of the sympathetic outflow. The NTS neurons
sending projections to ventral medulla were identified by previous microinjection of the
membrane DiI tracer in the ventral medulla and the spontaneous (sEPSCs) and tractus
solitarius (TS)-evoked excitatory post-synaptic current (TS-eEPSCs) were recorded
using patch clamp. With this approach we verified that in NTS neurons projecting to the
ventral medulla: i) the sEPSCs and TS-eEPSCs of DiI-labeled NTS neurons were
completely abolished by DNQX, an antagonist of ionotropic non-NMDA glutamatergic
receptors, showing that they are mediated by L-glutamate; ii) application of ATP
increases the frequency of appearance of spontaneous glutamatergic currents reflecting
an increased exocytosis of glutamatergic vesicles; and iii) ATP decreases the peak of
TS-evoked glutamatergic currents. We conclude that L-glutamate is the main
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

3
neurotransmitter of spontaneous and TS-evoked synaptic activities in the NTS neurons
projecting to ventral medulla and that ATP has a dual modulatory role on this excitatory
transmission facilitating the spontaneous glutamatergic transmission and inhibiting the
TS-evoked glutamatergic transmission. These data also suggest that ATP is not acting
as a cotransmitter with L-glutamate, at least at the level of this sub-population of NTS
neurons studied.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

4
The NTS is the site of integration of the viscerosensory information in the
brainstem including cardiovascular and respiratory afferents (Mifflin, 1992; Ciriello et
al., 1994; Chitravanshi & Sapru, 1995; Machado, 2001, 2004) and there is anatomical
and functional evidence that the first synapse of the peripheral chemoreceptor afferents
is located in the commissural NTS (Donoghue et al. 1984; Finley & Katz, 1992; Mifflin,
1992; Ciriello et al. 1994; Chitravanshi & Sapru, 1995; Paton et al. 2001). Activation of
the peripheral chemoreceptors by intravenous injection of potassium cyanide (KCN) in
awake rats or the working heart brainstem preparation (WHBP) produces bradycardia,
tachypnoea and increases in the sympathetic outflow and arterial pressure, (Franchini &
Krieger, 1992; Haibara et al. 1995; Boscan & Paton, 2001; Paton et al. 2002; Machado
& Bonagamba, 2005; Antunes et al. 2005b; Braga & Machado, 2006; Braga et al., 2007;
Zoccal et al., 2008). There is also evidence that the sympathoexcitatory component of
chemoreflex involves NTS neurons sending projections to the rostral ventrolateral
medulla [RVLM, (Ross, 1985; Urbanski & Sapru, 1988; Seller et al, 1990; Koshiya et
al., 1993; Granata, 1994; Aicher, et al., 1996)].
The increase in sympathetic activity during hypoxia condition is a physiological
response to increase the peripheral vascular resistance and the arterial perfusion
pressure. These autonomic and circulatory events occur in order to keep the cerebral
blood flow in the autoregulatory range. However, abnormal and frequent increase in the
sympathetic activity due to chemoreflex activation may contribute to the development
of hypertension, as observed in pathophysiological conditions (Machado, 2004). Our
laboratories are deeply involved with studies on the neurotransmission of the
sympathoexcitatory component of the chemoreflex at the NTS level, with special
attention to the role of L-glutamate and ATP as putative neurotransmitters or
neuromodulators (Haibara et al., 1995, 1999; de Paula et al., 2004; Machado &
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

5
Bonagamba, 2005; Antunes et al., 2005; Braga & Machado, 2006; Braga et al., 2007).
In a series of recent studies we verified that the antagonism of L-glutamate ionotropic
and metabotropic receptors in the NTS was not effective in blocking the pressor
response in awake rats to chemoreflex activation and produced no effect on the
sympathoexcitatory response in the WHBP to chemoreflex activation (Haibara et al.,
1999; Machado & Bonagamba, 2005; Braga & Machado, 2006). The antagonism of P2
receptors in the NTS of awake rats was also not effective in blocking the pressor
response in awake rats or the sympathoexcitatory response of the chemoreflex in the
WHBP (Braga et al., 2007). Therefore, the possibility that both L-glutamate and ATP
might be acting as cotransmitters in the processing of the sympathoexcitatory
component in the NTS was considered and simultaneous blockade of glutamatergic and
purinergic receptors were performed. We verified that the simultaneous antagonism of
these receptors was effective in blocking the pressor response in awake rats and the
sympathoexcitatory response to chemoreflex activation in the WHBP (Braga et al.,
2007).
These findings, indicating that both L-glutamate and ATP are part of the
complex neurotransmission system of the sympathoexcitatory component of the
chemoreflex in the NTS, opened interesting possibilities to explore a possible
cotransmission of both neurotransmitter in the NTS. In order to test this hypothesis, we
combined in vivo retrograde labeling and in vitro whole cell patch clamp recordings in
brainstem slices containing the NTS to perform a characterization of glutamatergic and
purinergic mechanisms on the spontaneous and evoked excitatory neurotransmission of
NTS neurons sending projections to the ventral medulla, which includes the pre-motor
neurons involved in the generation of the sympathetic activity.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

6
Application of DiI into the ventral medulla and brainstem slices preparation
Wistar male rats (270-290 g) under tribromoethanol anesthesia (250 mg kg–1, ip,
Aldrich Chemical, St Louis, USA) were placed in a stereotaxic apparatus (David Kopf,
Tujunga, USA) for unilateral microinjection of the membrane tracer dye 1,1'-
dioctadecyl-3,3,3',3'- tetramethylindocarbocyanine perchlorate (DiI, 0.5% in ethanol,
Invitrogen, Carlsbad, USA) into the ventral medulla. The stereotaxic coordinates
correspondent to the site of microinjections in the ventral medulla of DiI were 1.8 mm
lateral to the midline, 4.1 mm caudal to the bregma and 10.2 mm below the skull
surface.
Brainstem slices (250 µm thick) were sectioned at least 24 hours after DiI
microinjection in modified ice-cold (4C) artificial cerebrospinal fluid (aCSF)
containing the following (in mM): 75 sucrose, 87 NaCl, 2.5 KCl, 7 MgCl2, 1.25
NaH2PO4, 25 NaHCO3, 25 glucose and 0.2 CaCl2, (330-350 mOsm/Kg.H2O and pH
7.4) using an oscillating slicer (Vibratome 1000 plus, Vibratome, St. Louis, USA) and
then the slices were kept at 37C for 45 min. The site of DiI microinjection was
confirmed in fresh brainstem slices in a microscope equipped with fluorescence
(Olympus BX51WI, Olympus, Tokyo, Japan) using the nucleus ambiguus (NA) as an
anatomical reference. Thereafter, the slices were kept at room temperature (RT, 23-
25C) in normal aCSF containing (mM): 125 NaCl, 2.5 KCl, 1 MgCl2, 1.25 NaH2PO4,
25 NaHCO3, 25 glucose and 2 CaCl2, with osmolality of 310-330 mOsm/Kg.H2O, a rate
flow of approximately 3 ml/min and continuously bubbled with 95% O2 and 5% CO2.
Neurons in brainstem slice were visualized by infrared differential interference contrast
(IR-DIC; Figure 1, panel A) and fluorescence microscopy using a 40X water immersion
objective (LUMPlain F1-IR, Olympus, Tokyo, Japan) and a CCD camera (C7500-50,
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

7
Hamamatsu, Iwata-City, Japan) to identify DiI-labeled cell in the NTS slice (Figure 1,
panel B).
Whole cell patch-clamp electrophysiological recordings
Whole cell recordings (voltage clamp configuration) were made with patch
pipettes pulled from thick-walled borosilicate glass capillaries (Sutter Instruments,
Novato, USA) filled with an internal solution containing (mM): 150 KCl, 5 ethylene
glycol-bis (-aminoethylether)-N,N,N’,N’-tetraacetic acid (EGTA) and 10 N-2-
hydroxyethyl-piperazine-N’-2-ethanesulfonic acid (HEPES)(320-330 mOsm/Kg.H2O,
pH 7.4, final resistance: 4 to 8 M). The signals were acquired using an Axopatch 200B
amplifier (Axon Instruments, Sunnyvalley, USA) connected to a microcomputer via a
Digidata 1440A board (Axon Instruments, Sunnyvalley, USA). Data was low-pass
filtered at 2 kHz and acquired at 10 kHz. The holding potential was kept at -70 mV.
eEPSCs were generated by electrical stimulation of the TS using a bipolar
tungsten electrode (FHC, Bowdoin, USA) connected to an isolated stimulator (S48
Stimulator, Grass Instrument, West Warwick, USA). sEPSCs were recorded in 1-2
minutes periods and the data acquisition was performed with Clampex software
(pClamp 10, Axon Instruments, Sunnyvalley, USA). Some experiments were recorded
in the presence of bicuculline (20 µM) in the bath perfusion, in order to block
GABAergic currents or DPCPX (1 µM), an A1 receptor antagonist.
The peak amplitude of TS-eEPSC was measured using Clampfit (pClamp 10,
Axon Instruments, Sunnyvalley, USA) and the frequency, amplitude and half-width of
sEPSCs were analyzed using Minianalysis Program (version 5.0, Synaptosoft, Decatur,
USA). The data are expressed as mean ± standard error (SEM) and statistical
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

8
significance (p<0.05) was determined by paired Student t test using the GraphPad Prism
4 software (GraphPad Prism, San Diego, USA).
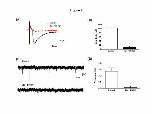
TS-eEPSCs and sEPSCs in the NTS neurons projecting to ventral medulla
Electrical stimulation of the TS often evoked EPSCs in DiI-labeled NTS neurons.
These currents were completely blocked after the addition of 6,7-dinitroquinoxaline-
2,3(1H,4H)-dione (DNQX, 10 µM), a non-NMDA receptor antagonist, and bicuculline
(20 µM), a GABAA receptor antagonist (-104±14 vs -8±1.2 pA, n=8, p<0.05, Figure 2,
panels A and B) in the bath perfusion. sEPSCs in DiI-labeled NTS cells were also
recorded and DNQX and bicuculline were also effective in blocking these events (1.36
± 0.26 vs 0.05 ± 0.03 Hz, n=23, p<0.05, Figure 2, panels C and D).
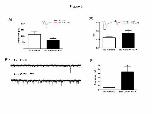
Effect of ATP application on TS-eEPSCs and sEPSCs
Considering our previous data suggesting an interaction between glutamatergic
and purinergic neurotransmission in the NTS (Braga et al., 2007), in the present study
we analyzed the effects of ATP (500 µM) in the bath perfusion on the TS-eEPSCs and
sEPSCs in DiI-labeled NTS neurons. During the experiments the excitatory currents
were isolated by adding bicuculline and 1,3-Dipropyl-8-cyclopentylxanthine (DPCPX,
1 µM), an A1 receptor antagonist, was also added in order to avoid a possible effect of
adenosine generated by ATP break-down.
ATP in the bath perfusion decreased the peak amplitude of TS-eEPSCs in all
recorded Di-labeled NTS neurons (-115.3 ± 24 vs -50 ± 10 pA, n=10, p<0.05, Figure 3,
panel A), suggesting that P2 receptors affect the glutamatergic neurotransmission in
NTS neurons. To analyze whether or not the mechanism by which ATP exerts its effect
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

9
on evoked synaptic transmission involves a decrease in pre-synaptic glutamate release
and/or changes in post-synaptic neuron, we used a paired-pulse protocol. For this
purpose, we analyzed the paired-pulse ratio (PPR) of the amplitudes of two consecutive
TS-eEPSCs (P1 and P2 respectively), with inter-stimulus interval of 100 ms. A change
in the paired-pulse ratio (P2/P1) accompanying a decrease in the first TS-eEPSC peak
amplitude was taken as an indicator for inhibition at a pre-synaptic site (Kline et al.,
2002; Sekizawa et al., 2003; Ohi et al., 2007). ATP increased the PPR in DiI-labeled
neurons (0.6 ± 0.09 vs 0.86 ± 0.17 pA, n=10, p<0.05, Figure 3, panel B), suggesting that
P2 receptors are affecting the excitatory transmission by a pre-synaptic mechanism.
ATP in the bath perfusion increased the frequency of spontaneous synaptic
currents in ~70% of DiI-labeled NTS neurons. In responsive neurons we observed a
significant increase in the frequency of sEPSCs (0.55 ± 0.13 vs 4.4 ± 1.3 Hz, n=8,
p<0.05, Figure 3, panels C and D) and no changes in the amplitude (20 ± 3.4 vs 23
± 1.7 pA) or half-width (2.11 ± 0.37 vs 2.1 ± 0.23 ms) of these currents, suggesting that
activation of P2 receptors increases vesicular glutamate release from pre-synaptic
neurons.
Importance of electrophysiological data for understanding the chemoreflex
neurotransmission in the NTS
Here we evaluated aspects of the neurotransmission in the NTS neurons sending
projections to the ventral medulla, a group of neurons participating in generation of
sympathetic outflow (Seller et al., 1990, Koshiya et al., 1993). We employed
microinjections of the plasma membrane DiI tracer in the ventral aspect of the medulla
for a retrograde labeling of the NTS neurons projecting to this area. Although we aimed
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

10
to reach the pre-motor sympathetic neurons in the RVLM, we assume that the DiI
microinjection was not restricted to the synaptic contact with these neurons and it may
have reached neurons other than those involved in the generation of the sympathetic
activity in the ventral medulla. Therefore, we considered the DiI-labeled NTS cells
generically as ventral medulla-projecting NTS neurons, which include the neurons
sending projections to the RVLM.
Recently, we demonstrated that combined microinjections of glutamatergic and
purinergic antagonists into the commissural NTS blocked the
sympathoexcitatory/pressor response to peripheral chemoreflex activation (Braga et al.,
2007). Therefore, we hypothesized that neurotransmission of the sympathoexcitatory
component of the chemoreflex involves an interaction between L-glutamate and ATP in
NTS and we evaluated this interaction in NTS neurons sending projections to ventral
medulla using electrophysiological approach.
The data of the present study support the concept that L-glutamate is the main
neurotransmitter involved in the synaptic activation of all studied NTS neurons sending
projections to the ventral medulla, since sEPSCs and TS-eEPSCs were abolished after
addition of DNQx, as previously described for NTS neurons (Andresen & Yang, 1990;
Kline et al., 2002; Jin et al., 2004; Accorsi-Mendonça et al., 2007; Kline et al., 2007).
Additionally, after DNQX application we did not observe any residual synaptic event
suggestive of a purinergic synaptic current, as observed in other brain areas, such as
cortex and habenula (Edwards et al., 1992; Pankratov et al., 2007).
The experiments involving ATP were performed in the presence of bicuculine
to block GABAergic currents and DPCPX to block possible effects of adenosine
receptors activation. ATP increased the frequency of the sEPSCs, suggesting that ATP
increases the probability of spontaneous L-glutamate release. The activation of somatic
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

11
and/or pre-synaptic P2X receptors could depolarize the neuron/terminal leading to an
increased firing of the pre-synaptic neurons or an increased probability of release of
glutamatergic synaptic vesicles (Shigetomi & Kato, 2004). On the other hand ATP
inhibited the TS-eEPSC amplitude in Dil-labeled NTS neurons, probably acting pre-
synaptically reducing the release probability of these vesicles due to the observed
increase in the paired-pulse ratio. Therefore, the data of these experimental protocols
shows that in Dil-labeled NTS neurons, ATP has a dual effect on the glutamatergic
neurotransmission in the NTS, being both inhibitory or excitatory, probably by
differential activation of ionotropic (P2X) and metabotropic (P2Y) receptors (Heinrich
et al., 2008). Alternatively we suggest that the increased spontaneous glutamate released
by ATP application can activate pre-synaptic glutamatergic metabotropic receptors
causing the inhibition in the TS-evoked EPSC. A depletion of readily-available vesicles
with consequent reduction in the evoked release cannot be rule out. The findings of the
present study suggest that L-glutamate and ATP are not acting as cotransmitters in the
synaptic transmission in the NTS neurons sending projections to ventral medulla. This
possibility is supported by the fact that in the present study ATP presented only a pre-
synaptic effect, while a cotransmitter, in accordance with the concept described by
Burnstock (2005; 2007), should also activate its post synaptic receptors.
Taking into account a) that only the double antagonism of glutamatergic and
purinergic receptors in the NTS was effective in the blockade of the
sympathoexcitatory/pressor response to chemoreflex activation in awake rats and
WHBP, b) the electrophysiological data showing that in the NTS neurons sending
projections to the ventral medulla, L-glutamate is the main excitatory neurotransmitter
while ATP acts as an neuromodulator, we must considerer several other possibilities in
order to fit together all these findings. First, it is important to note that the
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

12
sympathoexcitatory component of the chemoreflex is not restricted to the NTS-ventral
medulla projections (Figure 4), since there is experimental evidence that other important
areas in the brainstem such as the retrotrapezoid nucleus (Moreira et al, 2006; Takakura
et al., 2006) also participate in the generation of sympathetic and respiratory responses
to chemoreflex activation; second, as illustrated in Figure 4, there is also evidence that
the parabrachial nucleus (Haibara et al., 2002) and A5 area (Koshiya & Guyenet, 1994)
in the pons and the paraventricular nucleus of the hypothalamus (Olivan et al., 2001;
Reddy et al., 2005) participate in the sympathoexcitatory response of the chemoreflex,
but these alternative NTS projections were not yet evaluated using the approach
described in the present study. Therefore, at this time it is not possible to give a
definitive answer to the proposed question of this study about the overall glutamatergic
and purinergic cotransmission at the NTS level. However, the electrophysiological
findings of the present study clearly demonstrated that L-glutamate and ATP are not
acting as cotransmitters in the synaptic transmission in the NTS neurons sending
projections to the ventral medulla.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

13
Acknowledgements:
This work was supported by FAPESP (2004/03285-7) and CNPQ
(471184/2007-1). We thank Rubens F. Melo and Adriana Murashima for their technical
support.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

14
REFERENCES
Accorsi-Mendonca D, Leao RM, Aguiar JF, Varanda WA & Machado BH (2007)
Urethane inhibits the GABAergic neurotransmission in the nucleus of the solitary
tract of rat brain stem slices. Am J Physiol 292(1), R396-402.
Aicher SA, Saravay RH, Cravo S, Jeske I, Morrison SF, Reis DJ & Milner TA (1996)
Monosynaptic projections from the nucleus tractus solitarii to C1 adrenergic neurons
in the rostral ventrolateral medulla: comparison with input from the caudal
ventrolateral medulla. J Comp Neurol 373 (1), 62-75.
Andresen MC & Yang MY (1990) Non-NMDA receptors mediate sensory afferent
synaptic transmission in medial nucleus tractus solitarius.
Am J Physiol 259, H1307-11.
Antunes VR, Bonagamba LG & Machado BH (2005a) Hemodynamic and respiratory
responses to microinjection of ATP into the intermediate and caudal NTS of awake
rats. Brain Res 1032, 85-93.
Antunes VR, Braga VA & Machado BH (2005b) Autonomic and respiratory responses
to microinjection of ATP into the intermediate or caudal nucleus tractus solitarius in
the working heart-brainstem preparation of the rat. Clin Exp Pharmacol Physiol 32,
467-72.
Boscan P & Paton JF (2001) Role of the solitary tract nucleus in mediating nociceptive
evoked cardiorespiratory responses. Auton Neurosci 86(3), 170-82.
Braga VA & Machado BH (2006) Chemoreflex sympathoexcitation was not altered by
the antagonism of glutamate receptors in the commissural nucleus tractus solitarii in
the working heart-brainstem preparation of rats. Exp Physiol 91(3), 551-9.
Braga VA, Soriano RN, Braccialli AL, de Paula PM, Bonagamba LG, Paton JF &
Machado BH (2007) Involvement of L-glutamate and ATP in the neurotransmission
of the sympathoexcitatory component of the chemoreflex in the commissural nucleus
tractus solitarii of awake rats and in the working heart-brainstem preparation. J
Physiol 581, 1129-45.
Burnstock G (2005) Cotransmission. Current Opinion in Pharmacology 4: 47-52.
Burnstock G (2007) Physiology and pathophysiology of purinergic neurotransmission.
Physiol Rev 87(2), 659-797.
Chitravanshi VC & Sapru HN (1995) Chemoreceptor-sensitive neurons in commissural
subnucleus of nucleus tractus solitarius of the rat. Am J Physiol 268, R851-8.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

15
Ciriello J, Schultz CG & Roder S (1994) Collateral axonal projections from
ventrolateral medullary non-catecholaminergic neurons to central nucleus of the
amygdala. Brain Res 663(2), 346-51.
de Paula PM, Antunes VR, Bonagamba LG & Machado BH (2004) Cardiovascular
responses to microinjection of ATP into the nucleus tractus solitarii of awake rats.
Am J Physiol 287(5), R1164-71.
Donoghue S, Felder RB, Jordan D & Spyer KM (1984) The central projections of
carotid baroreceptors and chemoreceptors in the cat: a neurophysiological study. J.
Physiol 347, 397-409.
Edwards FA, Gibb AJ & Colquhoun D (1992) ATP receptor-mediated synaptic currents
in the central nervous system. Nature 359, 144-147.
Finley JC & Katz DM (1992) The central organization of carotid body afferent
projections to the brainstem of the rat. Brain Res 572, 108-16.
Franchini KG & Krieger EM (1993) Cardiovascular response of conscious rats to caroti
body chemoreceptor stimulation by intravenous KCN. J. Auton. Nerv. Syst. 42, 63-
69.
Granata AR (1994) Rostral ventrolateral medulla descending neurons excited by
nucleus tractus solitarii inputs. Brain Res 648, 299-305.
Haibara AS, Bonagamba LG & Machado BH (1999) Sympathoexcitatory
neurotransmission of the chemoreflex in the NTS of awake rats. Am J Physiol 276,
R69-80.
Haibara AS, Colombari E, Chianca DA Jr, Bonagamba LG & Machado BH (1995)
NMDA receptors in NTS are involved in bradycardic but not in pressor response of
chemoreflex. Am J Physiol 269, H1421-7.
Haibara AS, Tamashiro E, Olivan MV, Bonagamba LG, Machado BH (2002)
Involvement of the parabrachial nucleus in the pressor response to chemoreflex
activation in awake rats Auton Neurosci 101(1-2), 60-7.
Heinrich A, Kittel A, Csölle C, Sylvester Vizi E, Sperlágh B (2008) Modulation of
neurotransmitter release by P2X and P2Y receptors in the rat spinal cord.
Neuropharmacology 54(2), 375-86.
Jin YH, Bailey TW, Li BY, Schild JH & Andresen MC (2004) Purinergic and vanilloid
receptor activation releases glutamate from separate cranial afferent terminals in
nucleus tractus solitarius. J Neurosci 24(20), 4709-4717.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

16
Kline DD, Ramirez-Navarro A & Kunze DL (2007) Adaptive depression in synaptic
transmission in the nucleus of the solitary tract after in vivo chronic intermittent
hypoxia: evidence for homeostatic plasticity. J Neurosci 27(17), 4663-73
Kline DD, Takacs KN, Ficker E & Kunze DL (2002) Dopamine modulates synaptic
transmission in the nucleus of the solitary tract, J Neurophysiol 88, 2736–2744.
Koshiya N & Guyenet PG (1994) A5 noradrenergic neurons and the carotid sympathetic
chemoreflex. Am J Physiol 267, R519-26.
Koshiya N, Huangfu D & Guyenet PG (1993) Ventrolateral medulla and sympathetic
chemoreflex in the rat. Brain Res 609(1-2), 174-84.
Machado BH & Bonagamba LG (2005) Antagonism of glutamate receptors in the
intermediate and caudal NTS of awake rats produced no changes in the hypertensive
response to chemoreflex activation. Auton Neurosci 117(1), 25-32.
Machado BH (2001) Neurotransmission of the cardiovascular reflexes in the nucleus
tractus solitarii of awake rats. Ann N Y Acad Sci 940, 179-96.
Machado BH (2004) Chemoreflex and Sympathoexcitation. In Neural Mechanisms of
Cardiovascular Regulation. ed. Dun NJ, Machado BH & Pilowsky PM. pp. 31-58.
Kluwer Academic Publishers, Boston.
Mifflin SW (1992) Arterial chemoreceptor input to nucleus tractus solitarius. Am J
Physiol 263, R368-75.
Moreira TS, Takakura AC, Colombari E & Guyenet PG (2006) Central chemoreceptors
and sympathetic vasomotor outflow. J Physiol 577, 369-86.
Ohi Y, Kato F & Haji A (2007) Codeine presynaptically inhibits the glutamatergic
synaptic transmission in the nucleus tractus solitarius of the guinea pig. Neuroscience
146(3), 1425-33.
Olivan MV, Bonagamba LG & Machado BH (2001) Involvement of the paraventricular
nucleus of the hypothalamus in the pressor response to chemoreflex activation in
awake rats. Brain Res 895(1-2), 167-72.
Pankratov Y, Lalo U, Verkhratsky A & North RA (2007) Quantal release of ATP in
mouse cortex. J Gen Phys 129(3), 257-65.
Paton JF, de Paula PM, Spyer KM, Machado BH & Boscan P (2002) Sensory afferent
selective role of P2 receptors in the nucleus tractus solitarii for mediating the cardiac
component of the peripheral chemoreceptor reflex in rats.
J Physiol 543, 995-1005.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

17
Paton JF, Deuchars J, Li YW & Kasparov S (2001) Properties of solitary tract neurones
responding to peripheral arterial chemoreceptors. Neuroscience 105(1), 231-48.
Reddy MK, Patel KP & Schultz HD (2005) Differential role of the paraventricular
nucleus of the hypothalamus in modulating the sympathoexcitatory component of
peripheral and central chemoreflexes. Am J Physiol 289(3), R789-97.
Ross CA, Ruggiero DA & Reis DJ (1985) Projections from the nucleus tractus solitarii
to the rostral ventrolateral medulla. J Comp Neurol 242, 511-534.
Sekizawa S, Joad JP & Bonham AC (2003) Substance P presynaptically depresses the
transmission of sensory input to bronchopulmonary neurons in the guinea pig
nucleus tractus solitarii, J Physiol 552, 547–559.
Seller H, Konig S & Czachurski J (1990) Chemosensitivity of sympathoexcitatory
neurones in the rostroventrolateral medulla of the cat. Pflugers Arch 416(6), 735-41.
Shigetomi E & Kato F (2004) Action potential-independent release of glutamate by
Ca2+ entry through presynaptic P2X receptors elicits postsynaptic firing in the
brainstem autonomic network. J Neurosci 24(12), 3125-35.
Sun MK, Young BS, Hackett JT & Guyenet PG (1988) Reticulospinal pacemaker
neurons of the rat rostral ventrolateral medulla with putative sympathoexcitatory
function: an intracellular study in vitro. Brain Res 442(2), 229-39.
Takakura AC, Moreira TS, Colombari E, West GH, Stornetta RL & Guyenet PG (2006)
Peripheral chemoreceptor inputs to retrotrapezoid nucleus (RTN) CO2-sensitive
neurons in rats. J Physiol 572, 503-23.
Urbanski RW & Sapru HN (1988) Evidence for a sympathoexcitatory pathway from the
nucleus tractors solitarii to the ventrolateral medullary pressor area. J Auton Nerv
Syst 23,161-174.
Zoccal DB, Simms AE, Bonagamba LG, Braga VA, Pickering AE, Paton JF, Machado
BH (2008) Increased sympathetic outflow in juvenile rats submitted to chronic
intermittent hypoxia correlates with enhanced expiratory activity. J Physiol 586(13),
3253-65.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

18
LEGENDS:
Figure 1. Identification of DiI-labeled NTS neurons
(A): DiI-labeled NTS neuron (arrow) viewed with infrared differential interference
contrast optic (IR-DIC) in the brainstem slice; (B): the same neuron under fluorescence
with an arrow indicating the DiI labeling.
Figure 2. Effect of bicuculine and DNQX on sPSCs and TS-ePSCs in DiI-labeled
NTS neurons
(A): Representative traces of TS- evoked post synaptic currents (ePSCs) from a DiI-
labeled NTS neuron in a control condition and after DNQX (10 µM) and bicuculline (20
µM) in the bath perfusion; (B): Mean peak amplitude of TS-ePSCs from DiI-labeled
NTS neurons (n=8) before and after DNQX and bicuculline in bath perfusion; (C)
Representative traces of spontaneous post synaptic currents (sPSCs) from a DiI-labeled
NTS neuron in a control situation and after DNQX and bicuculline in the bath
perfusion; (D) Mean frequency of sPSCs in DiI-labeled NTS neurons (n=23) before and
after DNQX and bicuculline in bath perfusion. Bic = bicuculline. Holding potential =
-70 mV. * p < 0.05.
Figure 3. Effect of ATP on sEPSCs and TS-eEPSCs in DiI-labeled NTS neurons
(A): Mean peak amplitude of TS-evoked excitatory post-synaptic currents (eEPSCs) in
DiI-labeled NTS neurons (n=10) before and after ATP (500 µM) in bath perfusion.
Inset: Representative traces of TS-eEPSCs before and after ATP application in the bath
perfusion; DPCPX (1 µM), an A1 receptor antagonist used to avoid the effect of
adenosine produced by ATP break-down; (B): Mean PPR (paired pulse ratio) of two
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

19
consecutive eEPSCs (P1 and P2 respectively; time interval: 100 ms) in DiI-labeled NTS
neurons (n=10) before and after ATP in the bath perfusion. Inset: Representative traces
of two TS-eEPSCs before and after ATP application in the bath perfusion, P1 = Peak 1,
P2 = Peak 2; (C): Representative traces of sEPSCs from a DiI-labeled NTS neuron
before and after ATP in the bath perfusion; (D): Frequency of sEPSCs in DiI-labeled
NTS neurons (n=8) before and after ATP in the bath perfusion. Bic = bicuculline.
Holding potential = -70 mV. * p<0.05.
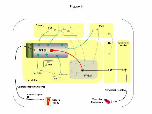
Figure 4. Schematic drawing of sympathoexcitatory pathways of the chemoreflex
in the brainstem, pons and hypothalamus
Lower pO2 in the arterial blood activates peripheral chemoreceptors located in the
carotid body. Experimentally this activation can be induced by intravenous injection of
potassium cyanide (KCN). The afferents of this reflex make the first synaptic contact in
the NTS, via tractus solitarius. NTS neurons sending projections to several other brain
areas are involved in the processing of the chemoreflex, such as paraventricular nucleus
of the hypothalamus (PVN, Olivan et al., 2001; Reddy et al., 2005), parabrachial
nucleus (PBN, Haibara et al., 2002), A5 area (Koshiya & Guyenet, 1994), pre-Bötzinger
nucleus (pre-BötC) and Bötzinger nucleus and retrotrapezoid nucleus (nRT, Moreira et
al., 2006; Takakura et al., 2006). Moreover, there is anatomical and functional evidence
of an important projection from NTS to the rostral ventrolateral medulla (RVLM,
Urbanski & Sapru, 1988; Aicher et al., 1996), in which are located the pre-motor
neurons involved in the generation of the sympathetic outflow (Sun et al., 1988).
Considering the importance of this NTS-RVLM projection in the context of the
sympatoexcitatory response of the chemoreflex, it was explored by using DiI tracer and
an electrophysiological approach. RVLM neurons send direct projections to the
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

20
intermediolateral column of spinal cord (IML), which has connections with the
sympathetic ganglion. Sympathetic post-ganglionic fibers send projections to the end-
organs and release noradrenaline, especially to the resistance vessels, which increases
the vascular resistance and arterial blood pressure. Therefore, the chemoreflex
activation may produce sympathoexcitation by activation of one and/or several neuronal
pathways from the NTS illustrated in this panel. The electrophysiological data on a
possible cotransmission of L-glutamate and ATP described in the present study is
restricted to the sub-population of NTS neurons sending projections to the ventral
medulla.
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (

) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (
) at CAPES - Usage on December 19, 2008 ep.physoc.orgDownloaded from Exp Physiol (
Related Documents





![[1b] Tractus Urinaria 2003](https://static.cupdf.com/doc/110x72/577cd84d1a28ab9e78a0e6b2/1b-tractus-urinaria-2003.jpg)





