Limnologica 51 (2015) 1–7 Contents lists available at ScienceDirect Limnologica jo ur nal ho me page: www.elsevier.com/locate/limno Aquatic prey subsidies to riparian spiders in a stream with different land use types Bonny Krell ∗ , Nina Röder, Moritz Link, René Gergs, Martin H. Entling, Ralf B. Schäfer Institute for Environmental Science, University of Koblenz-Landau, Campus Landau, Fortstrasse 7, 76829 Landau, Germany a r t i c l e i n f o Article history: Received 23 April 2014 Received in revised form 13 October 2014 Accepted 13 October 2014 Available online 25 October 2014 Keywords: River Aquatic-terrestrial linkage Emergence Food web Stable isotope Land use a b s t r a c t Land use related habitat degradation in freshwater ecosystems has considerably increased over the past decades, resulting in effects on the aquatic and the riparian communities. Previous studies, mainly in undisturbed systems, have shown that aquatic emergent insects contribute substantially to the diet of riparian predators. To evaluate the effect of land use on aquatic prey subsidies of riparian spiders, we performed a longitudinal study from June to August 2012 along a first order stream (Rhineland-Palatinate, Germany) covering three land use types: forest, meadow and vineyard. We determined the contribution of aquatic and terrestrial resources to the diet of web-weaving (Tetragnathidae spp.) and ground-dwelling (Pardosa sp.) riparian spiders using stable isotope analyses of aquatic emergent insects and terrestrial arthropods. The contribution of aquatic and terrestrial sources differed between Tetragnathidae spp. and Pardosa sp. as well as among land use types. Tetragnathidae spp. consumed 80–100% of aquatic insects in the meadows and 45–65% in the forest and vineyards. Pardosa sp. consumed 5–15% of aquatic insects in the forest, whereas the proportions of aquatic and terrestrial sources were approximately 50% in the meadow and vineyard. Thus, aquatic emergent insects are an important subsidy to riparian spiders and land use is likely to affect the proportion of aquatic sources in the spider diet. © 2014 Elsevier GmbH. All rights reserved. Introduction Freshwater ecosystems are facing a broad range of environmen- tal pressures from anthropogenic land use. Habitat degradation, toxic pollution, eutrophication, flow modifications and species invasion have considerably increased over the past decades due to Anthropogenic land use (e.g. agriculture, urbanization, etc.) and have been reported to reduce freshwater biodiversity (MEA, 2005; Dudgeon et al., 2006). In the riparian zone, aquatic and terrestrial ecosystems are linked through fluxes of energy and nutrients, also termed subsidies (Rowe and Richardson, 2001; Baxter et al., 2005; Post et al., 2007; Richardson et al., 2010). Studying the linkages between these ecosystems is crucial to understand how environ- mental impacts on one ecosystem can propagate to the adjacent ecosystem (Knight et al., 2005; Paetzold et al., 2011). Aquatic- terrestrial food chains or webs are crucial connections and may offer a pathway of propagation of environmental impact across these ecosystem boundaries (Polis et al., 1997). Whereas many studies examined the role of allochthonous inputs from terrestrial systems (e.g. leaf litter input) to aquatic food webs, aquatic prey ∗ Corresponding author. Tel.: +49 0 6341 280 31835. E-mail address: [email protected] (B. Krell). subsidies to riparian predators as well as terrestrial prey input to streams have only recently received increasing attention (Baxter et al., 2005; Hering and Plachter, 1997; Nakano and Murakami, 2001; Collier et al., 2002). Environmental impacts on aquatic orga- nisms may propagate to the adjacent riparian zone via different paths, two of them are: (1) Potential export of metals and/or organic contaminants by emergent insects to riparian predators. Evidence for the potential risk for higher terrestrial trophic levels from biomagnification of aquatic contaminants has been provided by (Cristol et al., 2008, Walters et al., 2008 and Raikow et al., 2011). (2) The loss of aquatic biodiversity, especially the loss of emer- gent aquatic insects, can affect the adjacent riparian zone through reduced food availability to riparian predators. Previous studies have shown that aquatic emergent insects can contribute substantially to the diet of riparian predators, such as arthropods, birds, bats, lizards and spiders (Sabo and Power, 2002; Sanzone et al., 2003; Kato et al., 2004; Briers et al., 2005; Paetzold et al., 2005; Marczak and Richardson, 2007). Moreover, experimental studies have shown that a reduction in the flux of emergent insects can affect distribution, growth and density of terrestrial predators (Nakano et al., 1999; Kato et al., 2003; Paetzold and Tockner, 2005; Marczak and Richardson, 2007; Kolb et al., 2012). However, these studies have mainly been performed in undisturbed ecosystems (Baxter et al., 2005). Paetzold et al. http://dx.doi.org/10.1016/j.limno.2014.10.001 0075-9511/© 2014 Elsevier GmbH. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Aw
BI
a
ARRAA
KRAEFSL
I
ttithDetPbmetotss
h0
Limnologica 51 (2015) 1–7
Contents lists available at ScienceDirect
Limnologica
jo ur nal ho me page: www.elsev ier .com/ locate / l imno
quatic prey subsidies to riparian spiders in a streamith different land use types
onny Krell ∗, Nina Röder, Moritz Link, René Gergs, Martin H. Entling, Ralf B. Schäfernstitute for Environmental Science, University of Koblenz-Landau, Campus Landau, Fortstrasse 7, 76829 Landau, Germany
r t i c l e i n f o
rticle history:eceived 23 April 2014eceived in revised form 13 October 2014ccepted 13 October 2014vailable online 25 October 2014
eywords:iverquatic-terrestrial linkagemergence
a b s t r a c t
Land use related habitat degradation in freshwater ecosystems has considerably increased over the pastdecades, resulting in effects on the aquatic and the riparian communities. Previous studies, mainly inundisturbed systems, have shown that aquatic emergent insects contribute substantially to the diet ofriparian predators. To evaluate the effect of land use on aquatic prey subsidies of riparian spiders, weperformed a longitudinal study from June to August 2012 along a first order stream (Rhineland-Palatinate,Germany) covering three land use types: forest, meadow and vineyard. We determined the contribution ofaquatic and terrestrial resources to the diet of web-weaving (Tetragnathidae spp.) and ground-dwelling(Pardosa sp.) riparian spiders using stable isotope analyses of aquatic emergent insects and terrestrialarthropods. The contribution of aquatic and terrestrial sources differed between Tetragnathidae spp. and
ood webtable isotopeand use
Pardosa sp. as well as among land use types. Tetragnathidae spp. consumed 80–100% of aquatic insectsin the meadows and 45–65% in the forest and vineyards. Pardosa sp. consumed 5–15% of aquatic insectsin the forest, whereas the proportions of aquatic and terrestrial sources were approximately 50% in themeadow and vineyard. Thus, aquatic emergent insects are an important subsidy to riparian spiders andland use is likely to affect the proportion of aquatic sources in the spider diet.
© 2014 Elsevier GmbH. All rights reserved.
ntroduction
Freshwater ecosystems are facing a broad range of environmen-al pressures from anthropogenic land use. Habitat degradation,oxic pollution, eutrophication, flow modifications and speciesnvasion have considerably increased over the past decades dueo Anthropogenic land use (e.g. agriculture, urbanization, etc.) andave been reported to reduce freshwater biodiversity (MEA, 2005;udgeon et al., 2006). In the riparian zone, aquatic and terrestrialcosystems are linked through fluxes of energy and nutrients, alsoermed subsidies (Rowe and Richardson, 2001; Baxter et al., 2005;ost et al., 2007; Richardson et al., 2010). Studying the linkagesetween these ecosystems is crucial to understand how environ-ental impacts on one ecosystem can propagate to the adjacent
cosystem (Knight et al., 2005; Paetzold et al., 2011). Aquatic-errestrial food chains or webs are crucial connections and mayffer a pathway of propagation of environmental impact across
hese ecosystem boundaries (Polis et al., 1997). Whereas manytudies examined the role of allochthonous inputs from terrestrialystems (e.g. leaf litter input) to aquatic food webs, aquatic prey∗ Corresponding author. Tel.: +49 0 6341 280 31835.E-mail address: [email protected] (B. Krell).
ttp://dx.doi.org/10.1016/j.limno.2014.10.001075-9511/© 2014 Elsevier GmbH. All rights reserved.
subsidies to riparian predators as well as terrestrial prey input tostreams have only recently received increasing attention (Baxteret al., 2005; Hering and Plachter, 1997; Nakano and Murakami,2001; Collier et al., 2002). Environmental impacts on aquatic orga-nisms may propagate to the adjacent riparian zone via differentpaths, two of them are: (1) Potential export of metals and/ororganic contaminants by emergent insects to riparian predators.Evidence for the potential risk for higher terrestrial trophic levelsfrom biomagnification of aquatic contaminants has been providedby (Cristol et al., 2008, Walters et al., 2008 and Raikow et al., 2011).(2) The loss of aquatic biodiversity, especially the loss of emer-gent aquatic insects, can affect the adjacent riparian zone throughreduced food availability to riparian predators.
Previous studies have shown that aquatic emergent insects cancontribute substantially to the diet of riparian predators, suchas arthropods, birds, bats, lizards and spiders (Sabo and Power,2002; Sanzone et al., 2003; Kato et al., 2004; Briers et al., 2005;Paetzold et al., 2005; Marczak and Richardson, 2007). Moreover,experimental studies have shown that a reduction in the fluxof emergent insects can affect distribution, growth and density
of terrestrial predators (Nakano et al., 1999; Kato et al., 2003;Paetzold and Tockner, 2005; Marczak and Richardson, 2007; Kolbet al., 2012). However, these studies have mainly been performedin undisturbed ecosystems (Baxter et al., 2005). Paetzold et al.
2 B. Krell et al. / Limnologica 51 (2015) 1–7
Table 1Environmental characterization of the survey sites considering different physico-chemical, riparian vegetation and habitat parameters.
Parameter Forest Meadow Vineyard
Channel (mean cm) Width 0.6 ± 0.2 0.5 ± 0.1 0.5 ± 0Depth 10 ± 5 15 ± 10 15 ± 10
PhysicochemicalParameter
Velocity (ms−1) 0.25 ± 0.05 0.3 ± 0.05 0.3 ± 0.06Temperature (◦C) 14.01 ± 1.5 14.1 ± 1.59 14.8 ± 1.56pH 7.59 ± 0.04 7.65 ± 0.04 7.67 ± 0.07O2 (% sat) 94.3 ± 1.15 95.1 ± 0.83 94 ± 0.84Conductivity (�S cm−1) 111 ± 3.48 117 ± 5.92 144 ± 4.27
Substratum (%) Boulder 5 5 0Cobble 10 5 0Gravel 10 10 0Pebble 15 10 10Sand 60 70 90
Vegetation (%) Emersed Macrophytes 0 20 60Trees 90 20 20Scrubs 0 30 40
(psalrtem
fbsamettfElafTi
M
S
wWwtmtsteEvssia
Gravel
Shade (%)
Woody debris (%)
2011) provided first evidence that stream pollution can controlopulations and community structure of terrestrial predators viaustained alterations in aquatic subsidies. Moreover, land use canlso strongly affect terrestrial subsidies to aquatic ecosystems (i.e.eaf litter and prey input) (Baxter et al., 2005). A reduction in ter-estrial subsidies may propagate along the aquatic food web and inurn lead to changes in the availability of aquatic subsidy (Nakanot al., 1999). Hence, land use may affect aquatic prey subsidies viaultiple paths.To examine how changes in freshwater ecosystems resulting
rom anthropogenic land use may manifest across the ecosystemoundary, we evaluated the effect of land use on aquatic prey sub-idies to riparian spiders. We performed a longitudinal study along
first order stream covering three different land use types: forest,eadows and vineyard. Following the method outlined in (Briers
t al., 2005) we aimed to use a 15N tracer for a better discrimina-ion of the dietary sources of spiders. We hypothesized (1) thathe amount of aquatic subsidy to spiders would decrease fromorest to vineyards, due to a loss of sensitive aquatic insects (e.g.phemeropterans, Plecopterans and Trichopterans) resulting fromand use related stressors such as pollution and habitat degradationnd (2) that the proportion of aquatic subsidies would be higheror the web-weaving than the ground-dwelling spiders, becauseetragnatha build their webs over the stream to catch aquaticnsects and is therefore more specialized on these (see Section 2.2).
ethods
election of study sites
The study sites were located at the Hainbach, a first order streamithin the Rhine river catchment, in Rhineland-Palatinate (South-estern, Germany). The Hainbach is about 10–15 cm deep, 0.5 mide, and had a mean current velocity of 0.3 ± 0.06 m s−1 during
he study period. Temperatures ranged from 11 to 16 ◦C and theean discharge was approximately 0.015 m3 s−1. These charac-
eristics were similar at all survey sites. The sampling sites wereelected to represent three different land use types. From upstreamo downstream the sites are: a relatively undisturbed mixed for-st site (N49◦14′34.3′′; E008◦02′42.4′′), a meadow (N49◦14′18.4′′;008◦03′0.00′′) with occasional mowing or sheep grazing and aineyard site (N49◦14′09.0′′; E008◦03′58.1′′). The distance between
ites was about 1 km (Fig. 1). The forest site had a heterogeneousubstrate with riffle and pool sections. The meadow site was sim-lar to it in substrate coverage but channelized and had lowermount of woody debris (Table 1). The vineyard site was strongly10 50 4088 12 9015 5 0
channelized and the stream substrate was relatively homogeneousconsisting of sand, with 60% and 20% cover of emersed macrophytesand scrubs, respectively. Regarding the riparian zone, the forestbank had almost no slope and was composed mainly by a layer ofleaves, whereas meadow and vineyard had a steep slope due to thechannelization and were similar in vegetation, mainly grasses andscrubs (Table 1). Pesticides, predominantly fungicides and herbi-cides, were applied at the vineyard site but we have no informationon compounds and amounts. Fishes have been rarely spotted at allsites.
Selection of spider species
Orbweb tetragnathid (Tetragnatha sp. and Metellina sp.) andground-dwelling lycosid spiders (Pardosa sp.) were selected as ter-restrial predators as they are common riparian spiders and werepresent in the riparian zone of the three land-use types. In addi-tion, they have contrasting prey capture techniques. Tetragnathidsuse orb webs that are anchored to debris or vegetation protrud-ing from the water to intercept flying insects that emerge from thestream (Forster and Forster, 2005). By contrast, ground dwellinglycosid spiders hunt in the riparian zone and their prey consistsof a wide range of invertebrates including terrestrial coleopterans,lepidopterans, orthopterans and hemipterans as well as aquatic insectimagos (Nyffeler, 1999).
Physical-chemical and habitat parameter
Physical-chemical data was sampled every 10 days (beforetracer was replaced) using a WTW measuring kit (WTW Multi 340iSet, Wissenschaftlich-Technische Werkstätten GmbH, Weilheim).
Habitat parameter such as substrate description, shading, per-centage of reach covered by macrophytes and the structuralcomposition of riparian zones were performed once during the thefield study. The description was based on a visual estimation overthe length of the surveyed stream reach (75 m) using diagrams fromthe Rapid Bioassessment Methodology for Rivers and Streams (EPA,2003) as guides.
Tracer addition
To improve discrimination of aquatic and terrestrial prey
according to their isotope signature, we enriched the stream with15N at the three study sites. The tracer addition was adapted from(Briers et al., 2005) to the discharge of our stream and set to approx-imately 0.013 g of 15NH4Cl (Cambridge Isotope Laboratories, Inc.
nologi
AaMg
F
wetaGvdwec−
asoo‘1sun
t
(pstttwtuwww(
S
ewddircattof(adrT
B. Krell et al. / Lim
ndover, USA) per day. The tracer was released into the stream as anqueous solution of 15NH4Cl for a period of twelve weeks betweenay and August 2012, which is the period of highest aquatic emer-
ence in the study region, using a Marriott bottle (Moore, 2004).
aunal sampling
Aquatic emergent insects were continuously sampled startingeek 2 after tracer addition started (May, 2012) using aquatic
mergence traps with a basal area of 0.25 m2, which were emp-ied twice a week. Insects were trapped in bottles containing anqueous solution of 1% (vol.) TWEEN® 80 (Carl Roth, Karlsruhe,ermany), a non-ionic detergent, and 5% (vol.) of NaCl (for preser-ation purposes). Three traps were located each at 20, 40 and 60 mownstream of the tracer injection point and three control trapsere located 10 m upstream. Samples were rinsed with water to
liminate traces of salt, identified to family level using a stereomi-roscope (Leica M10) in the laboratory, and afterwards frozen at20 ◦C.
Sampling of terrestrial arthropods and spiders was performedt week 4, 8 and 12 after tracer addition started within 1 m from thetream edge and within 2 m up and downstream from the positionf the emergence traps. We handpicked 3–5 individuals of juveniler female spiders of Pardosa sp. and tetragnathids. In addition, five
live’ (i.e. without trapping liquid) pitfall traps were placed within m from the stream edges to catch additional individuals of Pardosap. with daily removal of the trapped spiders. These traps were setp one day before spider collection and in case that an insufficientumber of spiders were caught, left for another day.
Spiders where individually kept in vials for twenty four hours,o allow gut clearance.
Terrestrial arthropods were sampled using a suction samplermodified Stihl SH 86; Stihl, Waiblingen, Germany). For each sam-le (3 at each location of the emergence traps), a 0.28 m2 plasticampling cylinder (20 cm height) was driven 1 cm in the groundo prevent the escape of crawling arthropods. Then the vegeta-ion, litter and soil surface were vacuumed for 1 minute withinhe sampling cylinder. Arthropods were collected in a gauze bagithin the nozzle of the sampler. Samples were transferred to plas-
ic bags, transported to the lab and frozen at −20 ◦C until sortingnder a stereomicroscope (Leica M10). All captured arthropodsere counted and identified at family or genus level. Organismsere dried at 60 ◦C for 3 days to reach constant dry weight. After-ards dry body mass was determined using an analytic balance
XA105 DualRange, Mettler Toledo).
table isotope analysis (SIA)
Single organisms were used for stable isotope analysis when-ver size allowed. Regarding aquatic organisms, only chironomidsere available in sufficient numbers to perform the SIA. The abun-ance of other taxa was too low to be included in SIA. This is likelyue to temporal differences in emergence peaks. Seven to fifteen
ndividuals of chironomids were pooled to provide sufficient mate-ial for analysis. We performed stable isotope analyses for severalhironomid samples for the weeks 4, 8 and 12 for all distances atll sites. The results showed that the tracer did not enrich and thathe stable isotope signatures were similar for the distances fromracer. Thus, we performed the complete stable isotope analysisnly for a consistent set of samples (week 12 and distance of 20 mrom tracer) that yielded to the highest sample size for all categoriesaquatic, terrestrial and spiders). All samples were dried, ground,
nd weighed into tin cups (0.3–0.7 mg). Terrestrial organisms wereivided in two categories, crawling and flying organisms. Threeeplicates of all taxa were analyzed. SIA was conducted using ahermo Finnigan Delta Advantage Isotope Ratio Mass Spectrometerca 51 (2015) 1–7 3
connected to a Flash HT elemental analyzer. Stable carbon andnitrogen values are presented in conventional ı notation ofı = Rsamples/Rstandard − 1 and units of per mille (‰) in relation to theinternational reference standards V-PDB (Vienna PeeDee Belem-nite) for carbon and atmospheric N2 for nitrogen. Repeated analysesof an internal standard typically resulted in an accuracy (±1standard deviation (SD)) of 0.12‰ and 0.05‰ for carbon and nitro-gen, respectively.
Data analysis
Statistical analysis was conducted using the statistical soft-ware R (R Development Core Team, 2012) and the SIAR mixingmodel (Version 4.0) (Parnell et al., 2010), which follows a Bayesianapproach to estimate the probability distribution of prey contri-bution to a consumer diet (in our case spiders). Stable Isotopesignatures of resources (i.e. aquatic and terrestrial invertebrates)were significantly different (t-test, p < 0.001). This is a prerequisiteto run SIAR mixing models properly (i.e. to be able to discriminatebetween food sources). We used trophic enrichment factors (TEF’s)and uncertainties for terrestrial consumers as given in (McCutchanet al., 2003), i.e. 0.5 ± 0.19 ‰ for �13C and 2.3 ± 0.24 ‰ for ı15N. SIARmixing models for tetragnathid spiders at the forest site were runonly with two sources (aquatic and terrestrial flying invertebrates)because the crawling terrestrial invertebrates were mainly com-posed of soil invertebrates such as collembolans, which are unlikelyto get caught in tetragnathid webs (Nyffeler, 1999). Stable isotopemixing models might exhibit uncertainties regarding prey orga-nisms that have not been sampled or were in too low number forSI analysis.
Pairwise t-tests with bonferoni correction were used to checkfor potential differences in the biomass of aquatic emergence(mg m−2 d−1, 12 emergence traps per site) and of terrestrial arthro-pods (mg 0.25 m−2, 3 suction samples per site) between sites.Prior to community analysis, the mean abundance of single taxaof the terrestrial and aquatic emergent community compositionwas computed per site using the R package Vegan (Version 2.0-8). The similarity of aquatic emergent and terrestrial communitycomposition among sites was calculated using the Bray–Curtisdissimilarity metric analysis after square root transformation, todecrease the weight of the highly abundant chironomids or collem-bolans (Legendre and Gallagher, 2001).
Results and discussion
Tracer enrichment
To increase the difference in the isotope signature betweenaquatic and terrestrial sources, we added the 15N tracer to thethree stream sites. In contrast to (Briers et al., 2005) a mean iso-tope enrichment of only 2.1 ± 1.4‰ in the stable isotope signatureof aquatic emergent chironomids was detected across the threemonth experimental run, instead of the expected 200‰. This isapproximately one trophic level (2.3 ± 0.24), based on TEF’s for ter-restrial consumers (McCutchan et al., 2003) and therefore withinthe range of natural variation of aquatic and terrestrial sources. Weattribute this finding to a too low tracer addition. This may haveresulted from (1) using the mean of the reported stream dischargerange from Briers et al. (2005) in our concentration calculation or(2) a lower incorporation of nitrogen in the aquatic food web inour stream. In fact, a follow-up experiment with a 30 times higher
tracer concentration at one of the sites yielded an isotope enrich-ment of up to 400‰ (unpublished results). Overall, this calls formore studies for identification of factors influencing isotope enrich-ment.
4 B. Krell et al. / Limnologica 51 (2015) 1–7
t from
tifatgfwaollt�c(
nrimt
i1twtr
C
b(
Composition of spider diet
Spiders are generalist predators and known to perform beston mixed diets (Nyffeler, 1999; Toft, 1999; Amalin et al., 1999).
Table 2Aquatic emergent families at the forest, meadow and vineyard site with total abun-dance (ind m−2 day−1) at each site.
Order Family Forest Meadow Vineyard
Ephemeroptera Baetidae 0 0 1.6Trichoptera Hydropsychidae 0 0 0.6Trichoptera Polycentropodidae 0.3 0 0Trichoptera Psychodidae 0.31 0 0.6Trichoptera Sericostomatidae 0 0.31 0
Fig. 1. Map of study sites represented by the circles. The colors represen
The isotope signature was most distinct between aquatic anderrestrial insects in the meadows, whereas differences were minorn the forest site with respect to �13C. This can be explained by dif-erences in the importance of allochthonous (i.e. terrestrial) andutochthonous (i.e. aquatic) organic carbon in the stream ecosys-em. Unshaded stream sections such as within meadows havereater primary production, whereas shaded sections like in theorest rely stronger on allochthonous detritus (Webster, 2007). Thisill be reflected in an isotopic signal closer to terrestrial C signal in
quatic organisms emerging from shaded sites. The higher amountf �15N at the vineyard site can be attributed to the agriculturalandscape, which is a dominant source of phosphorus and nitrogenoading to aquatic ecosystems. (Vander Zanden et al., 2005) foundhat agricultural and urban nitrogen inputs were linked to elevated15N in the biota. Likewise, a positive relationship between agri-ultural land use and stream nitrate �15N has been reported byHarrington et al., 1998).
Nevertheless, at all survey sites the natural stable isotope sig-atures allowed for differentiation between aquatic and terrestrialesources. For instance, despite a similar �13C signal, the organismsn the forest were distinct based on �15N. These distinct signals per-
itted the determination of the relative contribution of aquatic anderrestrial organisms to the spider diet (Fig. 2).
Stable isotope signatures of crawling and flying terrestrialnvertebrates differed at the forest site in �15N values by at least.45 ‰, whereas the stable isotope signatures were overlapping inhe meadow and vineyard site. Therefore, the terrestrial sourcesere pooled (i.e. the mean value and standard deviation of the
errestrial signals was used) for the meadow and vineyard siteesulting in 2 sources (aquatic and combined terrestrial).
omposition of resources
There were no significant differences between theiomass of total aquatic emergence among land use types0.83 ± 1.91; 1.21 ± 1.73 and 2.97 ± 4.37 mg m−2 day−1 for forest,
the darkest to lightest grey, Forest, Urban, Meadow and Vineyard areas.
meadow and vineyard, respectively) (all p > 0.28, t-test). Yetthe biomass of chironomids was higher in the vineyard(0.44 ± 0.25 mg m−2 day−1) than in the meadow and forest(0.13 ± 0.22 and 0.14 ± 0.29 mg m−2 day−1, respectively), (p < 0.03).The higher biomass of chironomids in the vineyards coincided withthe absence of Trichopterans resulting overall in a similar biomassas the other sites (Table 2). The mean biomass of terrestrial preyin the vineyards was up to two-fold higher (221.6 ± 93.9 mg m−2)than in the other sites (118.6 ± 34.3 and 143.5 ± 67.3 mg m−2
in the forest and meadow, respectively), though this differencewas not statistically significant (all p > 0.36, t-test). The terrestrialarthropod communities were more similar between meadow andvineyard (Bray–Curtis dissimilarity of 0.62) than between forestmeadow (0.87) or vineyard (0.93), respectively. The aquatic emer-gent community in the vineyard differed more strongly from boththe meadows and forest communities (Bray–Curtis dissimilarityof 0.51 and 0.52, respectively) than the latter two between eachother (Bray–Curtis dissimilarity 0.3).
Diptera Chironomidae 15.7 14.3 112.7Diptera Dolichopodidae 0.3 2 2Diptera Empididae 0 1 0Diptera Simuliidae 0 0.31 2.3
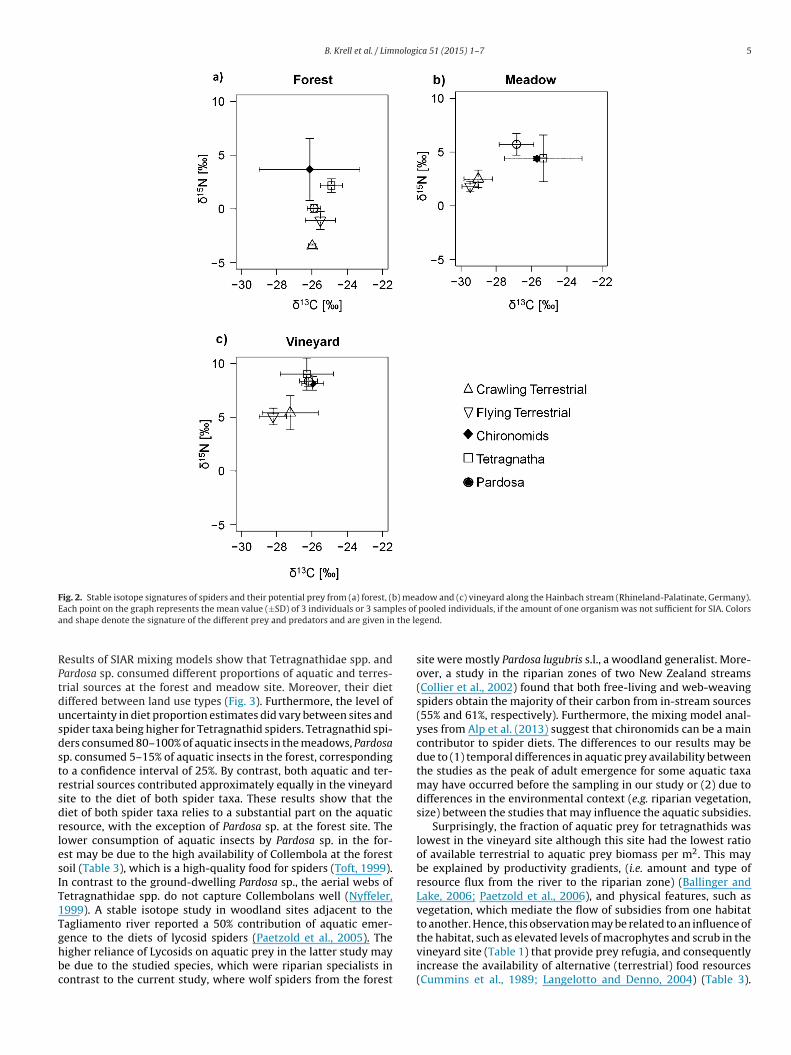
B. Krell et al. / Limnologica 51 (2015) 1–7 5
Fig. 2. Stable isotope signatures of spiders and their potential prey from (a) forest, (b) meadow and (c) vineyard along the Hainbach stream (Rhineland-Palatinate, Germany).E les of
a the le
RPtdusdstrsdrlesIT1Tghbc
ach point on the graph represents the mean value (±SD) of 3 individuals or 3 sampnd shape denote the signature of the different prey and predators and are given in
esults of SIAR mixing models show that Tetragnathidae spp. andardosa sp. consumed different proportions of aquatic and terres-rial sources at the forest and meadow site. Moreover, their dietiffered between land use types (Fig. 3). Furthermore, the level ofncertainty in diet proportion estimates did vary between sites andpider taxa being higher for Tetragnathid spiders. Tetragnathid spi-ers consumed 80–100% of aquatic insects in the meadows, Pardosap. consumed 5–15% of aquatic insects in the forest, correspondingo a confidence interval of 25%. By contrast, both aquatic and ter-estrial sources contributed approximately equally in the vineyardite to the diet of both spider taxa. These results show that theiet of both spider taxa relies to a substantial part on the aquaticesource, with the exception of Pardosa sp. at the forest site. Theower consumption of aquatic insects by Pardosa sp. in the for-st may be due to the high availability of Collembola at the forestoil (Table 3), which is a high-quality food for spiders (Toft, 1999).n contrast to the ground-dwelling Pardosa sp., the aerial webs ofetragnathidae spp. do not capture Collembolans well (Nyffeler,999). A stable isotope study in woodland sites adjacent to theagliamento river reported a 50% contribution of aquatic emer-
ence to the diets of lycosid spiders (Paetzold et al., 2005). Theigher reliance of Lycosids on aquatic prey in the latter study maye due to the studied species, which were riparian specialists inontrast to the current study, where wolf spiders from the forestpooled individuals, if the amount of one organism was not sufficient for SIA. Colorsgend.
site were mostly Pardosa lugubris s.l., a woodland generalist. More-over, a study in the riparian zones of two New Zealand streams(Collier et al., 2002) found that both free-living and web-weavingspiders obtain the majority of their carbon from in-stream sources(55% and 61%, respectively). Furthermore, the mixing model anal-yses from Alp et al. (2013) suggest that chironomids can be a maincontributor to spider diets. The differences to our results may bedue to (1) temporal differences in aquatic prey availability betweenthe studies as the peak of adult emergence for some aquatic taxamay have occurred before the sampling in our study or (2) due todifferences in the environmental context (e.g. riparian vegetation,size) between the studies that may influence the aquatic subsidies.
Surprisingly, the fraction of aquatic prey for tetragnathids waslowest in the vineyard site although this site had the lowest ratioof available terrestrial to aquatic prey biomass per m2. This maybe explained by productivity gradients, (i.e. amount and type ofresource flux from the river to the riparian zone) (Ballinger andLake, 2006; Paetzold et al., 2006), and physical features, such asvegetation, which mediate the flow of subsidies from one habitatto another. Hence, this observation may be related to an influence of
the habitat, such as elevated levels of macrophytes and scrub in thevineyard site (Table 1) that provide prey refugia, and consequentlyincrease the availability of alternative (terrestrial) food resources(Cummins et al., 1989; Langelotto and Denno, 2004) (Table 3).
6 B. Krell et al. / Limnologica 51 (2015) 1–7
F (b,) reo adow
Ivpsn
TTna
ig. 3. Proportions of prey groups in the diet of Tetragnathidae (a,) and Pardosa sp.f 95, 75 and 25% for the ingested prey source at each of the survey sites: forest, me
n fact, another study demonstrated the relevance of supralitoral
egetation for the production of both aquatic and terrestrial arthro-ods (Romanuk and Levings, 2003). However, because of lack ofpatial replication and a potential location effect, our study doesot permit definite conclusions regarding the influence of land use.able 3errestrial arthropod taxa at the forest, meadow and vineyard site. Shown are taxo-omic level of identification (order, family), and the total abundance (ind 0.75 m-2)t each site.
Order Family Forest Meadow Vineyard
Pseudoscorpiones Neobisiidae 9 0 2Ixodida Ixodidae 16 12 11Oribatida 35 140 0Isopoda Platyarthridae 4 0 0Isopoda Trichoniscidae 5 3 4Diplopoda Chordeumatidae 1 0 45Diplopoda Julidae 2 0 0Collembola 388 0 0Ensifera Gryllidae 0 0 1Hemiptera Cicadellidae 0 154 0Hemiptera Miridae 0 3 0Hemiptera Nabidae 0 5 5Coleoptera Byturidae 1 0 0Coleoptera Carabidae 0 0 6Coleoptera Chrysomelidae 0 6 9Coleoptera Coccinellidae 0 0 4Coleoptera Curculionidae 0 0 12Coleoptera Endomychidae 1 0 0Coleoptera Staphilinidae 0 1 6Coleoptera Tenebrionidae 2 0 0Hymenoptera Evaniidae 2 0 0Hymenoptera Formicidae 5 29 66Hymenoptera Proctotrupidae 1 0 0Diptera Lonchopteridae 0 2 2Diptera Rhinophoridae 2 0 0Diptera Sepsidae 1 0 3
spectively. The Boxplots represent the probability densities in credibility intervals and vineyard.
Overall, both spiders relied substantially on aquatic emergence,except for Pardosa sp. in the forest. This suggests that the aquaticsubsidy is more relevant for them in anthropogenically alteredenvironments. Yet, the riparian zone is also directly influenced byland use and future studies should disentangle direct (i.e. changesin aquatic communities) and indirect (i.e. changes in riparian veg-etation resulting in community changes) influences of land use onthe coupling of aquatic and terrestrial food webs.
Acknowledgments
We are grateful to Mr. and Ms. Hundemer for permission to sam-ple on their property. Funding for the study and BK was providedby the Ministry for Education, Science, and Culture (MBWWK) ofRhineland-Palatinate (Project AufLand).
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.limno.2014.10.001.
References
Alp, M., Peckarsky, B.L., Bernasconi, S.M., Robinson, C.T., 2013. Shifts in isotopicsignatures of animals with complex life-cycles can complicate conclusions oncross-boundary trophic links. Aquat. Sci. 75 (4), 595–606.
Amalin, D.M., Reiskind, J., McSorley, R., Pena, J., 1999. Survival of the hunting spi-der Hibana velox (Araneae, Anyphaenidae), raised on different artificial diets. J.
Arachnol. 27, 692–696.Ballinger, A., Lake, P.S., 2006. Energy and nutrient fluxes from rivers and streamsinto terrestrial food webs. Mar. Freshwater Res. 57, 15–28.
Baxter, C.V., Fausch, K.D., Saunders, W.C., 2005. Tangled webs: reciprocal flows ofinvertebrate prey link streams and riparian zones. Freshwater Biol. 50, 201–220.

nologi
B
C
C
C
D
E
F
H
H
K
K
K
K
L
L
M
M
M
M
N
B. Krell et al. / Lim
riers, R.A., Cariss, H.M., Geoghegan, R., Gee, J.H.R., 2005. The lateral extent of the sub-sidy from an upland stream to riparian lycosid spiders. Ecography 28, 165–170.
ollier, K.J., Bury, S., Gibbs, M., 2002. A stable isotope study of linkages betweenstream and terrestrial food webs through spider predation. Freshwater Biol. 47,1651–1659.
ristol, D.A., Brasso, R.L., Condon, A.M., Fovargue, R.E., Friedmann, S.L., Hallinger,K.K., Monroe, A.P., White, A.E., 2008. The movement of aquatic mercury throughterrestrial food webs. Science 320, 335.
ummins, K.W., Wilzbach, M.A., Gates, D.M., Perry, J.P., Taliaferro, W.B., 1989.Shredders and riparian vegetation: leaf litter that falls into streams influencescommunities of stream invertebrates. BioScience 39, 24–30.
udgeon, D., Arthington, A.H., Gessner, M.O., Kawabata, Z.I., Knowler, D.J., Lev-eque, C., Naiman, R.J., Prieur-Richard, A.H., Soto, D., Stiassny, M.L.J., Sullivan,C.A., 2006. Freshwater biodiversity: importance, threats, status and conservationchallenges. Biol. Rev. 81, 163–182.
PA, 2003. Guideline for Environmental Management—Rapid BioassessmentMethodology for Rivers and Streams. EPA, Victoria, Australia.
orster, R.R., Forster, L.M., 2005. Spiders of New Zealand and Their Worldwide Kin.University of Otago Press in association with Otago Museum, Dunedin, NewZeland.
arrington, R.R., Kennedy, B.P., Chamberlain, C.P., Blum, J.D., Folt, C.L., 1998. 15Nenrichment in agricultural catchments: field patterns and applications to track-ing Atlantic salmon (Salmo salar). Chem. Geol. 147, 281–294.
ering, D., Plachter, H., 1997. Riparian ground beetles (Coeloptera, Carabidae) prey-ing on aquatic invertebrates: a feeding strategy in alpine floodplains. Oecologia111 (2), 261–270.
ato, C., Iwata, T., Nakano, S., Kishi, D., 2003. Dynamics of aquatic insect flux affectsdistribution of riparian web-building spiders. Oikos 103, 113–120.
ato, C., Iwata, T., Wada, E., 2004. Prey use by web-building spiders: stable isotopeanalyses of trophic flow at a forest-stream ecotone. Ecol. Res. 19 (6), 633–643.
night, T.M., McCoy, M.W., Chase, J.M., McCoy, K.A., Holt, R.D., 2005. Trophic cascadesacross ecosystems. Nature 437, 880–883.
olb, G.S., Jerling, L., Hamback, P.A., 2012. The impact of Cormorants on plant-arthropod food webs on their Nesting Islands. Ecosystems 13, 353–366.
angelotto, G.A., Denno, R.F., 2004. Responses of invertebrate natural enemies tocomplex-structured habitats: a meta-analytical synthesis. Oecologia 139, 1–10.
egendre, P., Gallagher, E.D., 2001. Ecologically meaningful transformations for ordi-nation of species data. Oecologia 129, 271–280.
arczak, L.B., Richardson, J.S., 2007. Spiders and subsidies: results from the riparianzone of a coastal temperate rainforest. J. Anim. Ecol. 76, 687–694.
cCutchan, J.H.J., Lewis, W.M.J., Kendall, C., McGrath, C.C., 2003. Variation in trophicshift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 102, 378–390.
EA, 2005. Ecosystems and Human Well-being: Synthesis. Millennium EcossystemAssessment, Washington, DC.
oore, R.D., 2004. Construction of a Mariotte Bottle for constant-rate injection intosmall streams. Watersheat Manage. Bull. 8, 15–16.
akano, S., Miyasaka, H., Kuhara, N., 1999. Terrestrial–aquatic linkages: riparianarthropod inputs alter trophic cascades in a stream food web. Ecology 80,2435–2441.
ca 51 (2015) 1–7 7
Nakano, S., Murakami, M., 2001. Reciprocal subsidies: dynamic interdependencebetween terrestrial and aquatic food webs. PNAS 98, 166–170.
Nyffeler, M., 1999. Prey selection of spiders in the field. J. Arachnol. 27, 317–324.Paetzold, A., Bernet, J.F., Tockner, K., 2006. Consumer-specific responses to river-
ine subsidy pulses in a riparian arthropod assemblage. Freshwater Biol. 51,1103–1115.
Paetzold, A., Schubert, C.J., Tockner, K., 2005. Aquatic–terrestrial linkages along abraided river: Riparian arthropods feeding on aquatic insects. Ecosystems 8,748–759.
Paetzold, A., Smith, M., Warren, P.H., Maltby, L., 2011. Environmental impact propa-gated by cross-system subsidy: chronic stream pollution controls riparian spiderpopulations. Ecology 92, 1711–1716.
Paetzold, A., Tockner, K., 2005. Effects of riparian arthropod predation on the biomassand abundance of aquatic insect emergence. J. N. Am. Benthol. Soc. 24, 395–402.
Parnell, A.C., Inger, R., Bearhop, S., Jackson, A.L., 2010. Source partitioning using stableisotopes: coping with too much variation. PLoS One 5, e9672.
Polis, G.A., Anderson, W.B., Holt, R.D., 1997. Toward an integration of landscape andfood web ecology: the dynamic of spatial subsidized food webs. Annu. Rev. Ecol.Evol. Syst. 28, 289–316.
Post, D.M., Doyle, M.W., Sabo, J.L., Finlay, J.C., 2007. The problem of boundariesin defining ecosystems: a potential landmine for uniting geomorphology andecology. Geomorohology 89, 111–126.
R Development Core Team, 2012. R: A Language and Environment for StatisticalComputing, Reference Index Version 2.15.2. R Foundation for Statistical Com-puting, Vienna, Austria.
Raikow, D.F., Walters, D.M., Fritz, K.M., Mills, M.A., 2011. The distance that contam-inated aquatic subsidies etend into lake riparian zones. Ecol. Appl. 21, 983–990.
Richardson, J.S., Zhang, Y., Marczak, L.B., 2010. Resource subsidies across the land-freshwater interface and responses in recipient communities. River Res. Appl.26, 55–66.
Romanuk, T.N., Levings, C.D., 2003. Associations between arthropods and thesupralittoral ecotone: dependence of aquatic and terrestrial taxa on riparianvegetation. Environ. Entomol. 32, 1343–1353.
Rowe, L., Richardson, J.S., 2001. Community responses to experimental food deple-tion: resource tracking by stream invertebrates. Oecologia 129, 473–480.
Sabo, J.L., Power, M.E., 2002. Numerical response of lizards to aquatic insects andshort-term consequences for terrestrial prey. Ecology 83, 3023–3036.
Sanzone, D.M., Meyer, J.L., Marti, E., Gardiner, E.P., Tank, J.L., Grimm, N.B., 2003. Car-bon and nitrogen transfer from a desertstream to riparian predators. Oecologia134, 238–250.
Toft, S., 1999. Prey choice and spider fitness. J. Arachnol. 27, 301–307.Vander Zanden, M.J., Vadeboncoeur, Y., Diebel, M.W., Jeppesen, E., 2005. Primary
consumer stable nitrogen isotopes as indicators of nutrient source. Environ. Sci.Technol. 39, 7509–7515.
Walters, D.M., Fritz, K.M., Otter, R.R., 2008. The dark side of subsidies: adultstream insect export organic contaminants to riparian predators. Ecol. Appl. 18,1835–1841.
Webster, J.R., 2007. Spiraling down the river continuum: stream ecology and theU-shaped curve. J. N. Am. Benthol. Soc. 26, 375–389.
Related Documents











