Approaching trophic structure in Late Jurassic neritic shelves: A western Tethys example from southern Iberia Federico Olóriz a, ⁎ , Matías Reolid a,b , Francisco J. Rodríguez-Tovar a a Departamento de Estratigrafía y Paleontología, Facultad de Ciencias, Universidad de Granada, Fuentenueva s/n, 18071 Granada, Spain b UMR 5125, Paléoenvironnements and Paléobiosphère, Université Claude Bernard Lyon-1, 2 rue Raphael Dubois, Bât. Geode, F-69622 Villeurbanne Cedex, France Received 5 August 2005; accepted 15 June 2006 Available online 17 August 2006 Abstract The palaeoenvironmental conditions and trophic structure of a mid-outer neritic biota (microfossils, mainly forams, and macroinvertebrate assemblages) have been approached in middle Oxfordian–lowermost Kimmeridgian deposits from the Prebetic Zone (Betic Cordillera) in south-eastern Spain. According to relationships between fossil assemblages and lithofacies, a general seaward trend is identified which displays decreasing sedimentation rates and nutrient inputs, but increasing substrate consistency and presumably depth. Midshelf, terrigenous-rich deposits in the External Prebetic relate to the highest sedimentation rates and nutrient availability. These two parameters correlate with the highest content in vagile-benthic, calcareous perforate, epifaunal forams, as well as with potentially deep infaunal forams and infaunal macroinvertebrates. Outer-shelf lumpy deposits in the Internal Prebetic show the lowest sedimentation rates and nutrient availability and the highest records for macro–micro nektonics and planktics. In contrast, vagile-benthic, calcareous perforate epifaunal and potentially deep infaunal forams are scarcer in the midshelf environments. Colonial encrusting forams, benthic microbial communities and sessile benthic macro-invertebrates increase from the middle to outer shelf. Trophic-analysis structuring through the integration of benthic microbial communities, foraminiferal and macroinvertebrate fossil assemblages makes it possible to interpret: (a) a trophic-level frame composed of producers and primary and secondary consumers; (b) a main trophic-group differentiation in suspension-feeders, detritus-feeders, browsers, grazers, carnivores and scavengers; (c) a preliminary approach to food-chain structure supported by suspension-feeders, deposit-feeders and predators (active prey-selection carnivores); and (d) a food-pyramid model, which takes into account both recorded fossils and envisaged — i.e., ecologically inferred-organisms. © 2006 Elsevier B.V. All rights reserved. Keywords: micro- and macroinvertebrates; trophic relationships; paleoenvironmental features; epicontinental platform; Upper Jurassic 1. Introduction and geological setting The Prebetic Zone (Betic Cordillera, Fig. 1) displays epicontinental shelf deposits accumulated on the South- Iberian palaeomargin during the Mesozoic, in the External and the Internal Prebetic (Jerez-Mir, 1973). Earth-Science Reviews 79 (2006) 101 – 139 www.elsevier.com/locate/earscirev ⁎ Corresponding author. E-mail addresses: [email protected] (F. Olóriz), [email protected] (M. Reolid), [email protected] (F.J. Rodríguez-Tovar). 0012-8252/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.earscirev.2006.06.005

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

9 (2006) 101–139www.elsevier.com/locate/earscirev
Earth-Science Reviews 7
Approaching trophic structure in Late Jurassic neritic shelves:A western Tethys example from southern Iberia
Federico Olóriz a,⁎, Matías Reolid a,b, Francisco J. Rodríguez-Tovar a
a Departamento de Estratigrafía y Paleontología, Facultad de Ciencias, Universidad de Granada, Fuentenueva s/n, 18071 Granada, Spainb UMR 5125, Paléoenvironnements and Paléobiosphère, Université Claude Bernard Lyon-1, 2 rue Raphael Dubois, Bât. Geode, F-69622
Villeurbanne Cedex, France
Received 5 August 2005; accepted 15 June 2006Available online 17 August 2006
Abstract
The palaeoenvironmental conditions and trophic structure of a mid-outer neritic biota (microfossils, mainly forams, andmacroinvertebrate assemblages) have been approached in middle Oxfordian–lowermost Kimmeridgian deposits from the PrebeticZone (Betic Cordillera) in south-eastern Spain. According to relationships between fossil assemblages and lithofacies, a generalseaward trend is identified which displays decreasing sedimentation rates and nutrient inputs, but increasing substrate consistencyand presumably depth.
Midshelf, terrigenous-rich deposits in the External Prebetic relate to the highest sedimentation rates and nutrient availability.These two parameters correlate with the highest content in vagile-benthic, calcareous perforate, epifaunal forams, as well as withpotentially deep infaunal forams and infaunal macroinvertebrates. Outer-shelf lumpy deposits in the Internal Prebetic show thelowest sedimentation rates and nutrient availability and the highest records for macro–micro nektonics and planktics. In contrast,vagile-benthic, calcareous perforate epifaunal and potentially deep infaunal forams are scarcer in the midshelf environments.Colonial encrusting forams, benthic microbial communities and sessile benthic macro-invertebrates increase from the middle toouter shelf.
Trophic-analysis structuring through the integration of benthic microbial communities, foraminiferal and macroinvertebratefossil assemblages makes it possible to interpret: (a) a trophic-level frame composed of producers and primary and secondaryconsumers; (b) a main trophic-group differentiation in suspension-feeders, detritus-feeders, browsers, grazers, carnivores andscavengers; (c) a preliminary approach to food-chain structure supported by suspension-feeders, deposit-feeders and predators(active prey-selection carnivores); and (d) a food-pyramid model, which takes into account both recorded fossils and envisaged —i.e., ecologically inferred-organisms.© 2006 Elsevier B.V. All rights reserved.
Keywords: micro- and macroinvertebrates; trophic relationships; paleoenvironmental features; epicontinental platform; Upper Jurassic
⁎ Corresponding author.E-mail addresses: [email protected] (F. Olóriz), [email protected]
(M. Reolid), [email protected] (F.J. Rodríguez-Tovar).
0012-8252/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.earscirev.2006.06.005
1. Introduction and geological setting
The Prebetic Zone (Betic Cordillera, Fig. 1) displaysepicontinental shelf deposits accumulated on the South-Iberian palaeomargin during the Mesozoic, in theExternal and the Internal Prebetic (Jerez-Mir, 1973).

Fig. 1. Geological sketch of the Prebetic Zone and stratigraphic distribution of Mid-Oxfordian to lowermost Kimmeridgian lithofacies. (A) Location.(B) and (C) geological setting of studied area. (D) Simplified lithofacies stratigraphy and paleogeography.
102 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
These paleogeographical units represent proximal anddistal areas in this neritic, carbonate and carbonate–siliciclastic shelf system. Geographically, outcroppingareas in the Prebetic Zone are divided into the central(Sierra de Cazorla and Sierra de Segura) and eastern(Altos de Chinchilla) sectors. The succession studied(Fig. 1) mainly corresponds to the Middle–UpperOxfordian, and locally ranges into the lowermostKimmeridgian (Planula Zone).
Oxfordian rocks indicate the first hemipelagic–pelagic sedimentation over a discontinuity surface cap-ping white oolitic limestones and dolomites of theEarly–Middle Jurassic shelf that developed in thesouthern margin of Iberia (García-Hernández et al.,1981). The upper boundary of the studied succession is
biostratigraphically recognized in either a marl–lime-stone rhythmite that includes the first Kimmeridgiandeposits or an omission surface or hardground pre-ceding this rhythmite.
The main goal of the present study is to interpretfossil assemblages from the mid-outer shelf in terms ofpalaeocommunity analysis. To this end, macro andmicroinvertebrate assemblages (mainly foraminifera)are characterised and their relation to the lithofacies areexamined. The assemblage composition and distribu-tion enable us to analyse the diversity, lifestyle andfeeding habits, and to interpret the controlling envi-ronmental, eco-sedimentary parameters. Finally, thetrophic structure (food chains, food web and foodpyramid) is approached.
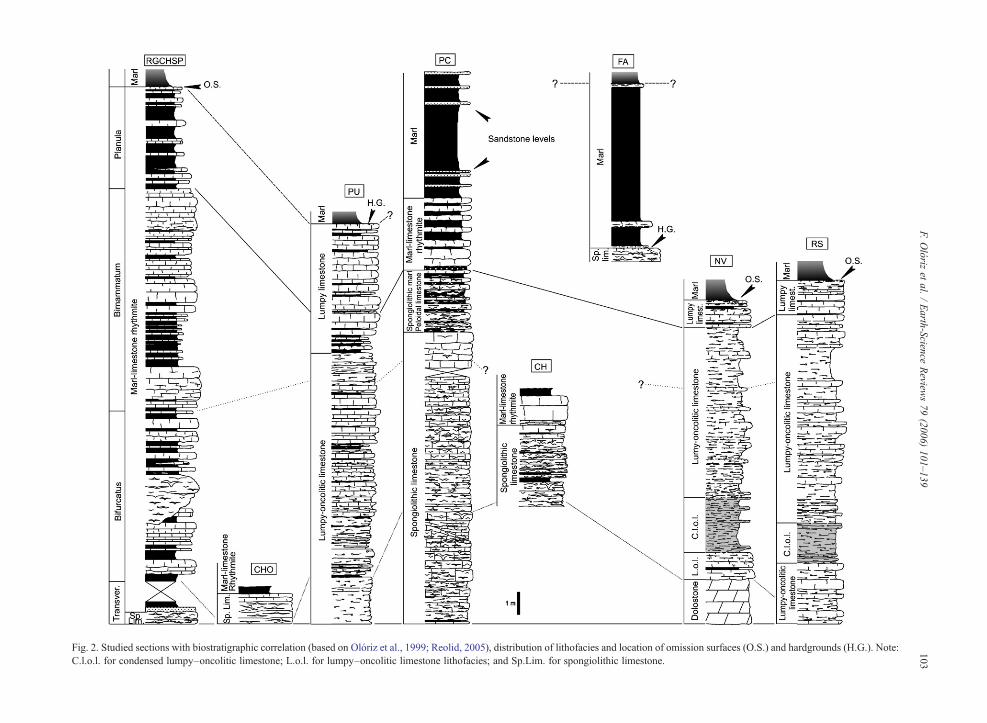
Fig. 2. Studied sections with biostratigraphic correlation (based on Olóriz et al., 1999; Reolid, 2005), distribution of lithofacies and location of omission surfaces (O.S.) and hardgrounds (H.G.). Note:C.l.o.l. for condensed lumpy–oncolitic limestone; L.o.l. for lumpy–oncolitic limestone lithofacies; and Sp.Lim. for spongiolithic limestone.
103F.
Olóriz
etal.
/Earth-Science
Review
s79
(2006)101–139

Fig. 3. Stratigraphic distribution of foraminiferal and macroinvertebrate assemblages of CH section (eastern sector of the External Prebetic), withrepresentation of the different types of spectrum as pie-diagrams (Foraminiferal assemblages: total assemblage, vagile benthics, and sessile benthics;and macroinvertebrate assemblages: general spectrum, ammonoid spectrum and benthos spectrum). Note 1: number close to the pie-diagramscorresponds to number of exemplars. Note 2: Black points for foraminiferal samples taken into account for counting (see text); black stars for samplesize according to the standard for macroinvertebrates (see text); and white stars for sample size below the standard for macroinvertebrates (see text).
104 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
Previous studies of fossil assemblages from theUpper Oxfordian to lowermost Kimmeridgian in thePrebetic Zone have focused on taphonomic aspects ofmacroinvertebrate assemblages (Olóriz et al., 2002a,b,2004a), and on palaeoecological considerations of theforaminiferal assemblages (Olóriz et al., 2002c, 2003a).Recent integrative studies present a preliminary ap-proach to the trophic structure of the local communityinterpreted (Olóriz et al., 2003b; Reolid, 2005).
2. Materials and methods
A study of eight profiles was carried out (Fig. 2), six ofwhich correspond to the External Prebetic (central sector:RGCHSP, CHO, PL; eastern sector: CH, PC, FA) and two
to the Internal Prebetic (NV and RS). Several sponge–microbialite buildups have also been analysed (spongebioherm–microbial lithoherm in Olóriz et al., 2003b).
Fine-resolution ammonite analyses (Olóriz et al.,1999; Reolid, 2005; Olóriz et al., in press), have enableda detailed biostratigraphic characterisation at the zoneand subzone level (even some bio-horizons) from theTransversarium Zone (Middle Oxfordian) to the PlanulaZone (lowermost Kimmeridgian, traditionally upper-most Oxfordian). This biostratigraphic characterisationfits with the standard proposals for southern Submedi-terranean Europe, and allows for a precise correlation atbiozone level for the eight profiles studied (Fig. 2).
The calibration of the absolute time represented inthe stratigraphic interval studied is difficult. Detailed

105F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
biostratigraphic analysis reveals a relative completeness ofthe stratigraphic interval, without biostratigraphicallyrecognized hiatuses. However, absolute calibration isdifficult due to significant variations in the attribution ofgeological ages and durations according to differentproposals. Nevertheless, a tentative calibration can beenvisaged following the most recent proposals. Hardenbolet al. (1998), using the proposals of Gradstein et al. (1994,1995), present a detailed temporal calibration at the level ofammonite zones and subzones for southwestern Europe(Tethyan/Submediterranean). In this proposal the lowerboundary of the Transversarium Zone (Plicatilis/Transver-sarium boundary) is assigned to 157.26 Ma, and the upperboundary of the Planula Zone (Planula/Platynota bound-ary) is located at 154.10 Ma. The interval studied(Transversarium to Planula zones) could therefore havean estimated duration of 3.16 Ma. Moreover, assuming asimilar duration for all of the Oxfordian cronosubzones (19for the Tethyan/Submediterranean domain, in Hardenbol etal., 1998), each would represent a duration of around279000 yr. Hence, the Transversarium–Planula timeinterval, represented by 12 subzones, could last approxi-mately 3.35Ma.On this basis, a time span of 3.16–3.35Macan be considered for the Transversarium–Planula strati-graphic interval.
Four groups of lithofacies are distinguished from theTransversarium (Middle Oxfordian) to the Planula zones(lowermost Kimmeridgian): A lumpy lithofacies group(lumpy limestone, lumpy–oncolitic limestone and con-densed lumpy–oncolitic limestone), a spongiolithiclithofacies group (spongiolithic limestone and spongio-lithic marl and peloidal limestone), a marl–limestonerhythmite lithofacies group and a marl lithofacies group(Olóriz et al., 2002a).
Lumpy limestones refer to nodular-like deposits toavoid any precise reference to Oxfordian typical nodularlimestones in the Prebetic. True nodular limestones in theUpper Jurassic of the Tethys are typically ammoniticorosso facies (i.e. epioceanic, benthos-poor deposits, andnot epicontinental and benthos-rich deposits). Lumpy–oncolitic limestones consist of nodular-like limestonesrich in peloids and bioclasts, oncoids with nubecularidsand lumps (more or less rounded and micritized,carbonate aggregates, finally originated by cementationmainly derived from biotic intractions). Condensedlumpy–oncolitic limestones are nodular-like limestonesrich in ammonoids, oncoids of nubecularids and bioclasts.Lumpy–oncolitic limestones, as well as condensedlumpy–oncolitic limestones, have usually been describedas nodular limestones (i.e., García-Hernández et al.,1981). Spongiolithic limestones are composed of well-stratified beds, characterised by highly abundant, com-
monly adjacent sponges, as a fundamental component ofthe rock. Spongiolithic marl and peloidal limestones arelimestones with irregular stratification surfaces and marls,the latter with abundant sponge fragments and occasionalcarbonate concretions. Marl limestone comprises well-bedded limestones alternating with marls, while marlsrefer to calcareous shales and silts.
In general terms, the lumpy lithofacies group, withmicrofacies characterised by a higher content of oncoidswith nubecularids, is limited palaeogeographically tocomparatively distal areas (the Internal Prebetic). Theremaining lithofacies, spongiolithic limestone, marlylimestone rhythmite and marl lithofacies, show micro-facies with a higher content of detrital non-carbonategrains and peloids. They are registered in the compar-atively proximal sectors (the External Prebetic), whichdisplay a predominance of the spongiolithic lithofaciesgroup in the eastern sector and of the marl–limestonerhythmite lithofacies group in the central sector (Figs.1D and 2).
The study of the microfauna focused on the fora-miniferal assemblages using thin-section analysis. Thistechnique makes it possible to identify and compareassemblages registered in materials with high carbonatecontent and to analyse assemblages of sessile benthicforaminifera. Its main disadvantage is the difficulty in-volved in distinguishing some genera from thin sections,as is the case with some nodosarids (Nodosaria–Den-talina and Lenticulina–Astacolus) and nubeculariids(Nubecularia–Nubeculinella). The study involved 370thin sections obtained from 146 sampling stations, andsome 31000 specimens (Figs. 3–10). Abundance datahave been approached through counting of specimens-per-cm2 in thin sections. In order to characterise theforaminiferal assemblages, three types of spectra wereused (Fig. 3):
(1) spectrum A, which shows the total assemblage,distinguishing between planktics, vagile benthicsand sessile benthics;
(2) spectrum B, which shows the relative compositionin vagile benthic foraminifera: ophthalmidiids,spirillinids, nodosarids, agglutinated forms andother foraminifera, usually minority forms;
(3) spectrum C, which shows the relative composi-tion in sessile benthic foraminifera: Siliceousagglutinated forms, Bullopora, Tubiphytes, andunspecified sessile foraminifera. Colonial en-crusting foraminifera were not included becauseof the difficulty in characterising on thin sectionthe number of individuals in a colony (e.g., Nu-becularia and Nubeculinella). Because of their
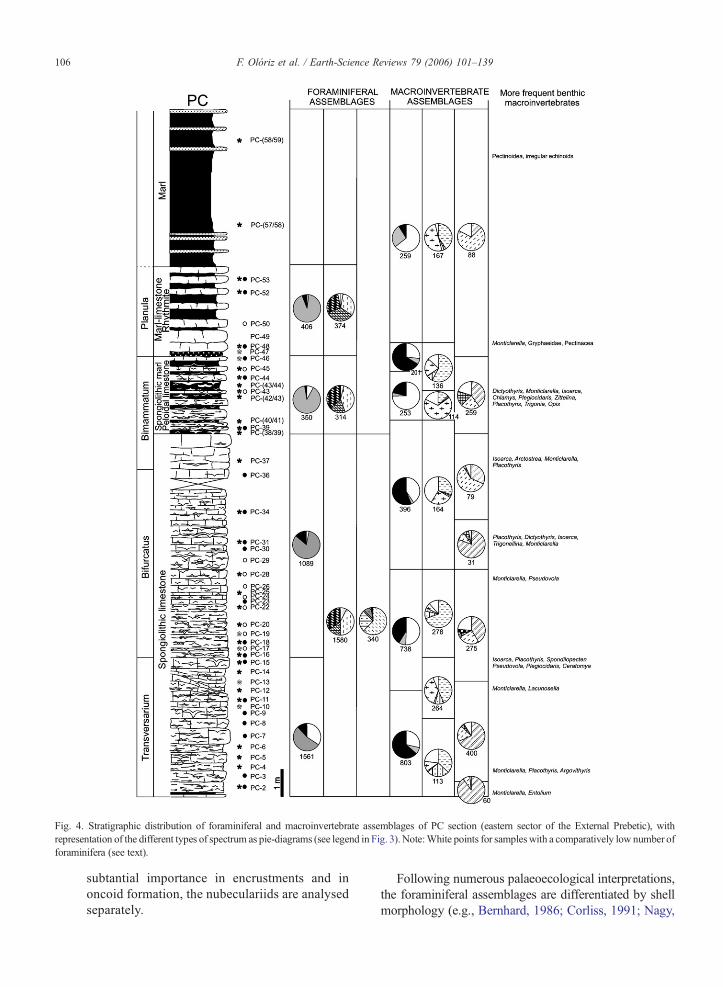
Fig. 4. Stratigraphic distribution of foraminiferal and macroinvertebrate assemblages of PC section (eastern sector of the External Prebetic), withrepresentation of the different types of spectrum as pie-diagrams (see legend in Fig. 3). Note:White points for sampleswith a comparatively low number offoraminifera (see text).
106 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
subtantial importance in encrustments and inoncoid formation, the nubeculariids are analysedseparately.
Following numerous palaeoecological interpretations,the foraminiferal assemblages are differentiated by shellmorphology (e.g., Bernhard, 1986; Corliss, 1991; Nagy,

Fig. 5. Stratigraphic distribution of macroinvertebrate assemblages of FA section (eastern sector of the External Prebetic), with representation of thedifferent types of spectrum as pie-diagrams (see legend in Fig. 3).
107F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
1992; Tyszka, 1994; Nagy et al., 1995), and the benthicforaminiferal assemblages are segregated into threegroups in relation to microhabitat depth in the sediment:
(1) Epifaunal foraminifera, up to 1 cm deep (Corliss,1991); includes all the sessile foraminifera,ophthalmidiids, miliolids, spiral agglutinatedforaminifera (Ammodiscoides, Glomospira andTrochammina) and spiral calcareous foraminifera(spirillinids, Epistomina, Trocholina, etc.), exceptLenticulina.
(2) Shallow infaunal foraminifera for those living indepths of less than 5 cm (Kuhnt et al., 1996), andbetween one and five in this investigation; elon-
Fig. 6. Stratigraphic distribution of foraminiferal and macroinvertebrate asserepresentation of the different types of spectrum as pie-diagrams (see legend
gated calcareous foraminifera (nodosarids such asNodosaria, Dentalina, Planularia, etc.), as wellas elongated agglutinated foraminifera, mainlyuniserial, which in initial stages may be streptos-piral and planispiral (Ammobaculites, Ammomar-ginulina and Haplophragmium).
(3) Ubiquitous foraminifera distributed within a widerange of depths in the substrate; Lenticulina, ahighly ubiquitous genus that tolerates a wide rangeof microhabitats from epifaunal to deep infaunal(Tyszka, 1994); and uniserial (Reophax) and bise-rial (undifferentiated textularids) agglutinated fo-raminifera, that vary between shallow and deepinfaunal microhabitats.
mblages of CHO section (central sector of the External Prebetic), within Fig. 3).
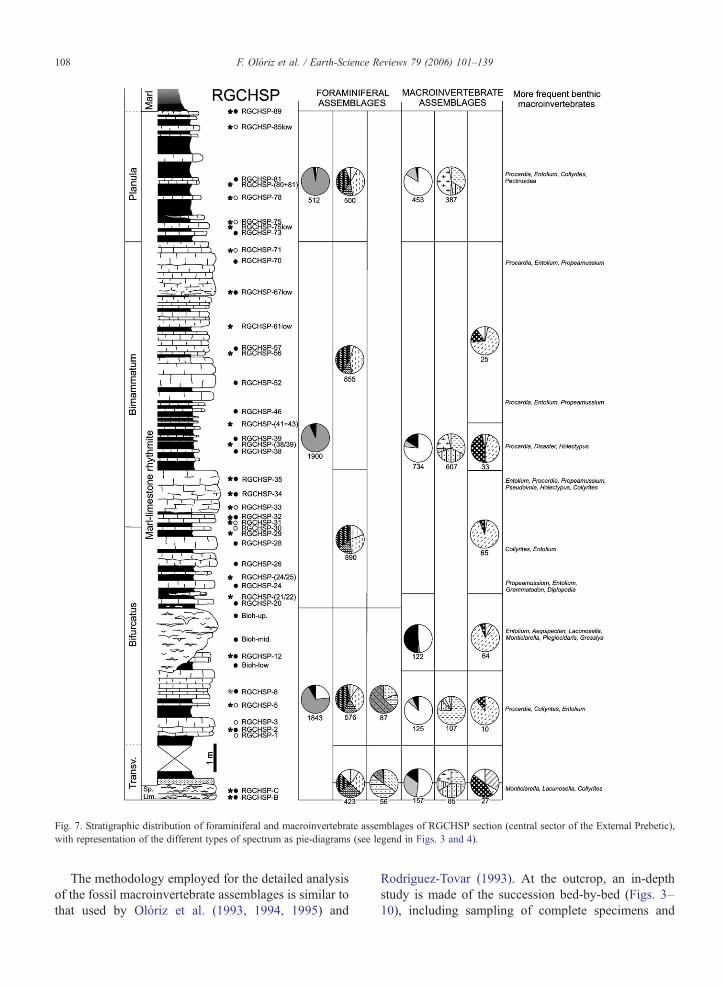
Fig. 7. Stratigraphic distribution of foraminiferal and macroinvertebrate assemblages of RGCHSP section (central sector of the External Prebetic),with representation of the different types of spectrum as pie-diagrams (see legend in Figs. 3 and 4).
108 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
The methodology employed for the detailed analysisof the fossil macroinvertebrate assemblages is similar tothat used by Olóriz et al. (1993, 1994, 1995) and
Rodríguez-Tovar (1993). At the outcrop, an in-depthstudy is made of the succession bed-by-bed (Figs. 3–10), including sampling of complete specimens and

Fig. 8. Stratigraphic distribution of foraminiferal and macroinvertebrate assemblages of PL section (central sector of the External Prebetic showingaffinities to Internal Prebetic), with representation of the different types of spectrum as pie-diagrams (see legend in Figs. 3 and 4).
109F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
fragments, and a preliminary taphonomic analysis.Counting was approached by estimating the number ofspecimens per-rock-volume (taking into account the
particular bed thickness in each case of the fossil-richhorizons sampled) and the time-per-collecting thestandard size-sample (50 specimens) when “normal”
110 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
or fossil-poor horizons were sampled. The commonlyfragmented sponges were excluded from quantitativeanalyses because it is difficult to precisely evaluate thenumber of individuals. Epibionts were excluded due totheir sparse and punctual record. Other remains that arenormally abundant and disarticulated (e.g., crinoids)were excluded because of the impossibility of deter-mining the number of individuals. Thus, sponges, epi-bionts and crinoids were analysed qualitatively.
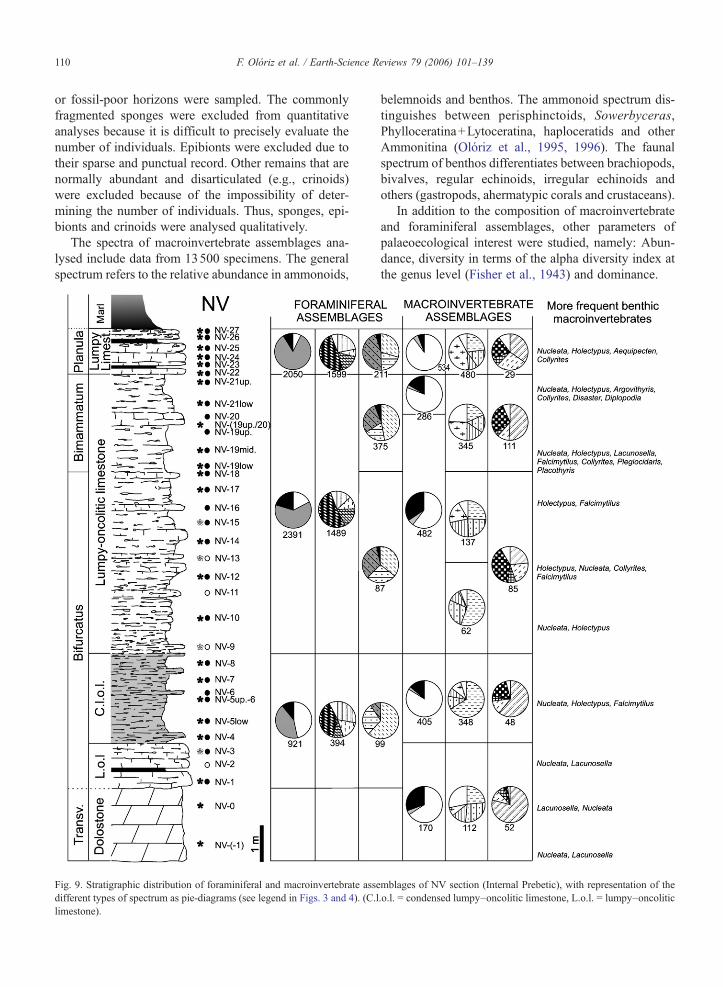
The spectra of macroinvertebrate assemblages ana-lysed include data from 13500 specimens. The generalspectrum refers to the relative abundance in ammonoids,
Fig. 9. Stratigraphic distribution of foraminiferal and macroinvertebrate assedifferent types of spectrum as pie-diagrams (see legend in Figs. 3 and 4). (C.llimestone).
belemnoids and benthos. The ammonoid spectrum dis-tinguishes between perisphinctoids, Sowerbyceras,Phylloceratina+Lytoceratina, haploceratids and otherAmmonitina (Olóriz et al., 1995, 1996). The faunalspectrum of benthos differentiates between brachiopods,bivalves, regular echinoids, irregular echinoids andothers (gastropods, ahermatypic corals and crustaceans).
In addition to the composition of macroinvertebrateand foraminiferal assemblages, other parameters ofpalaeoecological interest were studied, namely: Abun-dance, diversity in terms of the alpha diversity index atthe genus level (Fisher et al., 1943) and dominance.
mblages of NV section (Internal Prebetic), with representation of the.o.l. = condensed lumpy–oncolitic limestone, L.o.l. = lumpy–oncolitic
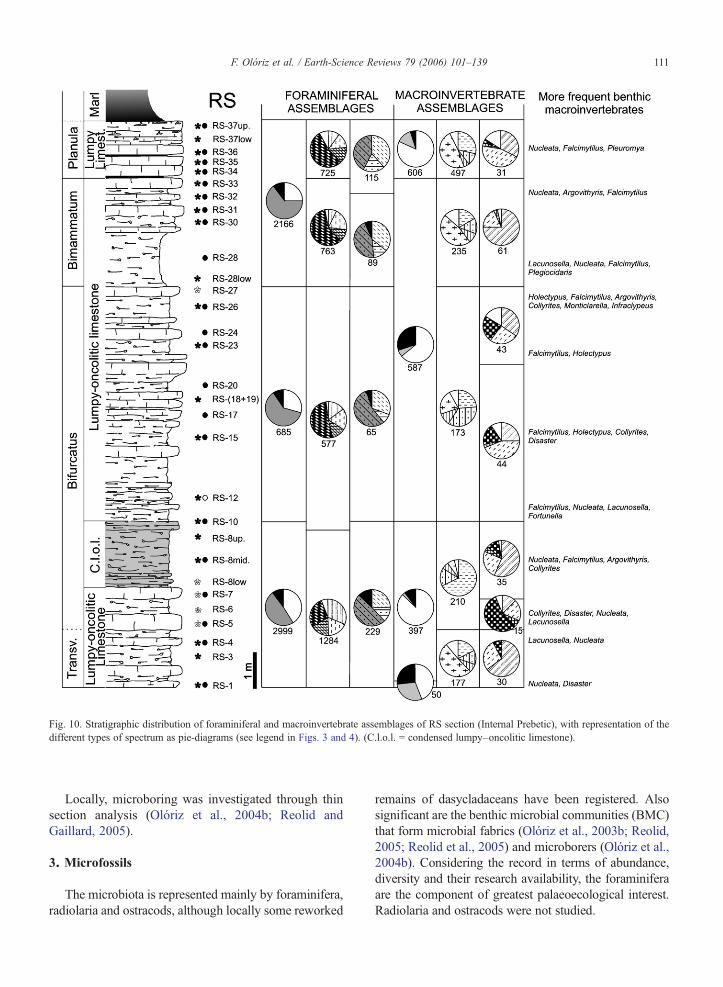
Fig. 10. Stratigraphic distribution of foraminiferal and macroinvertebrate assemblages of RS section (Internal Prebetic), with representation of thedifferent types of spectrum as pie-diagrams (see legend in Figs. 3 and 4). (C.l.o.l. = condensed lumpy–oncolitic limestone).
111F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
Locally, microboring was investigated through thinsection analysis (Olóriz et al., 2004b; Reolid andGaillard, 2005).
3. Microfossils
The microbiota is represented mainly by foraminifera,radiolaria and ostracods, although locally some reworked
remains of dasycladaceans have been registered. Alsosignificant are the benthic microbial communities (BMC)that form microbial fabrics (Olóriz et al., 2003b; Reolid,2005; Reolid et al., 2005) and microborers (Olóriz et al.,2004b). Considering the record in terms of abundance,diversity and their research availability, the foraminiferaare the component of greatest palaeoecological interest.Radiolaria and ostracods were not studied.

112 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
3.1. Foraminiferal assemblages
Foraminifera are mainly benthic (agglutinated,calcareous perforate and porcelanaceous forms) belong-ing to the suborders Textulariina, Lagenina, Spirillinina,Involutinina, Globigerinina, Milionina and Robertinina.Tubiphytes is included within Nubeculariidae, since ithas been interpreted as a nubeculariid in symbiosis withalgae (Flügel, 1981, 1982; Bernier, 1984; Schmid, 1995)or as microbial coating (Leinfelder et al., 1993a).
The averaged foraminiferal assemblage studied in thePrebetic Zone is made up of vagile benthics (68%),followed by planktics (21%) and sessile benthics (11%).Vagile benthic foraminifera are principally agglutinatedforaminifera (41%) and spirillinids (26%). Nodosaridsand porcelanaceous forms (mainly ophthalmidiids) arenormally present to a minor degree. Planktic foraminif-era belong to Globuligerina. Sessile benthic foraminif-era are mainly nubeculariids, and siliceous agglutinatedforms (46%) followed by Tubiphytes (30%) arepredominant among solitary taxa; Bullopora and otherindeterminate foraminifera make up the rest.
3.1.1. Foraminiferal assemblages and lithofaciesAccording to lithofacies, analysis of distribution,
composition, abundance and diversity in foraminiferalassemblages reveal a close relationship (Fig. 11):
(1) Lumpy limestone: Very abundant foraminifera,predominantly vagile benthics (71.5%), followedby planktics (20.5%) and sessile benthics (8%).Predominant among the vagile benthics are theagglutinated forms (47%), mainly Ammobaculitesand to a lesser degree Reophax. Ophthalmidiids,spirillinids and nodosarids (16% each) make up therest. Sessile benthic foraminifera are dominated bysiliceous agglutinated forms (40%) and Tubiphytes(40%). Nubeculariids are abundant. A total of 33genera have been recorded, of which only 12 makeup more than 1%. Individuals belonging to sixgenera (Globuligerina 20.5%, Ammobaculites20.5%, Reophax 12%, Ophthalmidium 11.5%,Spirillina 11.2%, and Lenticulina 6%) represent80% of the total foraminifera quantified. Forbenthic foraminifera, the alpha-index at the genuslevel is 4 in the Internal Prebetic, the epifaunarepresenting 44% and the shallow infauna 31%.
(2) Lumpy–oncolitic limestone: The abundance offoraminifera is variable, with vagile benthicsmaking up 61%, followed by planktics (29.5%)and sessile benthics (9.5%). Vagile benthic fora-minifera are dominated by agglutinated forms
(Ammobaculites and Reophax), while among thesessile benthics, Tubiphytes and siliceous aggluti-nated forms are predominant. Nubeculariids arevery abundant. The foraminiferal assemblage iscomposed of 31 genera, 12N1%, and Globuliger-ina predominates. Individuals belonging to fivegenera (Globuligerina 29.5%, Ammobaculites16.6%, Ophthalmidium 12.5%, Spirillina 11.2%,and Reophax 9.5%) represent 80% of the totalforaminifera quantified. The benthic foraminiferalassemblage presents a genus-level alpha-index of4.2 in the Internal Prebetic and 3.3 in the PL profile(External Prebetic); vagile epifauna is predominantwith an important contribution of sessile epifaunaland shallow infaunal foraminifera, mainly Ammo-baculites (Fig. 11).
(3) Condensed lumpy–oncolitic limestone: High abun-dance of foraminifera; Globuligerina (45%) andvagile benthic foraminifera (41%) are typical, whilesessile forms are subordinate (14%). Vagile benthicsaremainly agglutinated, followed byophthalmidiidsand spirillinids (Fig. 11). Notable among the sessileforaminifera are siliceous agglutinated forms andBullopora. Nubeculariids are very abundant. Theforaminiferal assemblage is composed of 23 generaof which 13 are registered N1%, with a clear pre-dominance of Globuligerina (Fig. 11). Individualsbelonging to seven genera (Globuligerina 45%,Spirillina 8.7%, Ammobaculites 8%, Ophthalmi-dium 8%, Tubiphytes 4.5%, Lenticulina 4.1%, andReophax 4%) represent 80% of the total foraminif-era quantified. The benthic foraminiferal assem-blage has a genus-level alpha-index of 4.4 in theInternal Prebetic; the epifauna is predominant (65%)and there is a notable abundance of sessile forami-nifera and Epistomina.
(4) Spongiolithic limestone: Foraminifera are abun-dant, mainly vagile benthics (65%), followed byplanktic (24%) and sessile benthic (11%). Vagilebenthics are dominated by spirillinids (40%) andthen agglutinated forms, while ophthalmidiids andnodosarids are subordinate. The sessile assem-blage is mainly composed of siliceous agglutinat-ed forms (Fig. 11). Occurrence of nubeculariids ishigh but variable. The total assemblage is com-posed of 31 genera, of which 11N1%. Individualsbelonging to six genera (Spirillina 25.5%, Glo-buligerina 23.5%, Reophax 14%, Ophthalmidium6.8%, Lenticulina 6%, and Ammobaculites 5%)represent 80% of the total foraminifera quantified.Despite the large number of genera identified, thegenus-level alpha-index is 3.5. Benthic epifauna
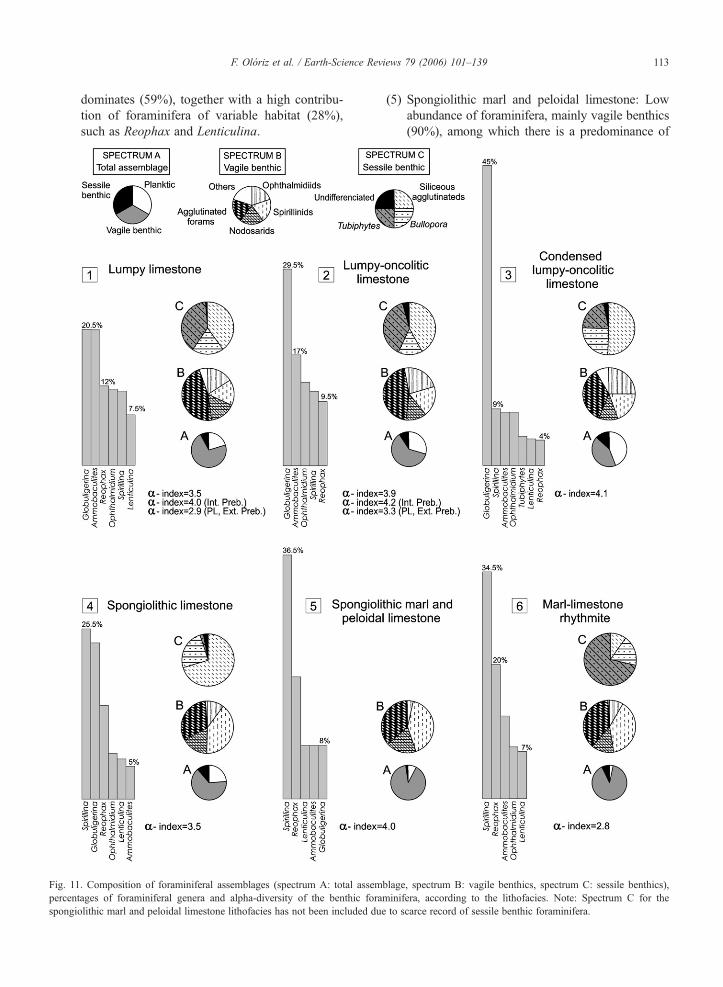
113F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
dominates (59%), together with a high contribu-tion of foraminifera of variable habitat (28%),such as Reophax and Lenticulina.
Fig. 11. Composition of foraminiferal assemblages (spectrum A: total assepercentages of foraminiferal genera and alpha-diversity of the benthic fospongiolithic marl and peloidal limestone lithofacies has not been included d
(5) Spongiolithic marl and peloidal limestone: Lowabundance of foraminifera, mainly vagile benthics(90%), among which there is a predominance of
mblage, spectrum B: vagile benthics, spectrum C: sessile benthics),raminifera, according to the lithofacies. Note: Spectrum C for theue to scarce record of sessile benthic foraminifera.

114 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
spirillinids (41%) and agglutinated forms (37%),mostly Reophax. Planktic foraminifera are scarce(8%) and encrusting foraminifera are even scarcer(2%). Nubeculariids are rare and do not form col-onies. In this lithofacies, 15 genera have beenidentified, of which 12 are N1% of the total record.Individuals belonging to five genera (Spirillina36.6%, Reophax 18.5%, Lenticulina 8%, Ammo-baculites 8%, and Globuligerina 8%) representapproximately 80% of the total foraminifera quan-tified. For benthics, the alpha-index at the genuslevel is 4. There is a large spectrum of micro-habitats, with a predominance of vagile epifauna(47%) followed by foraminifera of variable micro-habitats (32%), such as Reophax and Lenticulina,and finally by shallow infaunal (21%).
(6) Marl–limestone rhythmite: Low abundance of fora-minifera. Predominance of vagile benthics (89%)composed of spirillinids and agglutinated foraminif-era. Most of the sessile benthic foraminifera corres-pond to fragments of Tubiphytes (71.3%), whileother taxa are very scarce outside the buildups. Onlynine of the 26 genera identified are N1% of the totalrecord. Individuals belonging to five genera (Spirilli-na 34.5%, Reophax 20.1%, Ammobaculites 12.5%,Ophthalmidium 7.7%, and Lenticulina 7.1%) rep-resent 80% of the total foraminifera quantified
Fig. 12. Mean composition of foraminiferal andmacroinvertebrate assemblage(total assemblage, vagile benthics, and sessile benthics) and macroinvertebra
(Fig. 11). The genus-level alpha-index for benthicforaminifera is 2.8. Epifaunal, mainly vagile forami-nifera (Spirillina and Ophthalmidium) dominate(52%), while forms with a variable microhabitat(Reophax and Lenticulina) are subordinate (29%).
3.1.2. Areal distribution of the foraminiferal assemblagesThe marked differences between the foraminiferal
assemblages of the External and the Internal Prebeticmakes it possible to identify two typical foraminiferalassemblages (Olóriz et al., 2003a; Reolid, 2005) that areof palaeogeographic significance in relation to aproximal–distal gradient (Fig. 12).
Proximal assemblage: Recorded in spongiolithic,marl–limestone rhythmite and marl lithofacies groups inthe External Prebetic. Characterised by:
(1) Low abundance ofGlobuligerina and nubeculariids.(2) A predominance of vagile foraminifera supported by
spirillinids and, to a lesser extent, by ophthalmidiidsand agglutinated forms. The latter display lowdiversity, are mainly composed of Reophax, andspiral agglutinated forms are almost non-existent.The nodosarids present the highest proportion ofuniserial forms, despite a predominance of spiralforms (Lenticulina).
(3) Low relative diversity of benthic foraminifera.
s for External and Internal Prebetic, with representation of foraminiferaltes spectrums (general, ammonoids and benthos). See legend in Fig. 3.

115F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
Distal assemblage: Recorded in the lumpy lithofaciesgroup in the Internal Prebetic. Distinguished from theproximal assemblage by:
(1) A greater abundance of Globuligerina and nube-culariids (Nubecularia–Nubeculinella).
(2) Vagile foraminifera dominated by uniserial agglu-tinated forms, mainly Ammobaculites followed byReophax and very few spiral agglutinated forms(Ammodiscoides, Glomospira and Trochammina).Ophthalmidium and Epistomina present the great-est abundance in the Prebetic Zone.
(3) Greater diversity of benthic foraminifera.
4. Fossil macroinvertebrates
The recorded assemblage of fossil macroinverte-brates is clearly dominated by ammonoids (56% in theExternal and 78% in the Internal Prebetic; othercephalopods such as belemnoids and nautiloids aresecondary), followed by benthics (30% in the Externaland 16% in the Internal Prebetic, Fig. 12). Among theammonoids, there is a predominance of the perisphinc-toid group (38%), followed by the haploceratids (29%).Among the benthic macroinvertebrates, brachiopodspredominate (47%), followed by bivalves (31% in theExternal and 20% in the Internal Prebetic). Echinoidsmake up 12% in the External Prebetic (7% irregular and5% regular echinoids) and 24% in the Internal Prebetic(20% irregular and 4% regular echinoids). Other ben-thics are scarce. Sponges are mainly recorded in theExternal Prebetic and are characterised by the spongio-lithic lithofacies group.
4.1. Nekto-planktic and demersal macroinvertebrates
Ammonoids are the most abundant component inmacroinvertebrate assemblages. Genera have been iden-tified belonging to the suborders Phylloceratina (19.9%),Lytoceratina (0.7%) and Ammonitina (79.4%). The latterare predominant, especially Perisphinctoidea (38.1%) andHaploceratoidea (29.3%); in Phylloceratina Sowerby-ceras are predominant, while Lytoceratina are very scarce.
The average macroinvertebrate assemblage in thePrebetic Zone is dominated by the perisphinctoid group,followed by haploceratids and Sowerbyceras. The rest ofPhylloceratina presents very low values, while Lytocer-atina are even scarcer (b1%). Towards the Inner Prebetic,an area that is more distal, a decrease is observed in theperisphinctoid group (42% in the External vs. 33% in theInternal Prebetic), while an increase can be seen in thePhylloceratina+Lytoceratina group (3% in the External
vs. 8% in the Internal Prebetic). Important stratigraphicdifferences were observed in lithofacies (Figs. 3–10),which were related to increasing upward proportions ofhaploceratids and a decreasing trend for the perisphinctoidgroup.
The remaining cephalopods are mainly composed ofunequivocal coleoidea (Belemnitida) and, to a lesserextent, nautiloids (Nautilida and Ortoceratida). Belem-noids are more abundant in the External (14%) than inthe Internal Prebetic (5.5%). The abundance of belem-nites varies with the lithofacies (2% in condensedlumpy–oncolitic limestone and 32% in marls; Fig. 13).Nautilids and ortoceratids are very scarce.
4.2. Benthic macroinvertebrates
Sponges are the largest benthics, and are especiallyabundant in spongiolithic limestone lithofacies. Theyare siliceous sponges, with a predominance of dish-shaped (73%) versus other morphotypes (27%), whichare mainly composed of cup-shaped sponges and, morerarely, of tubular, ramose and biconic morphotypes.Most of the sponges belong to Hexactinellida (77%Dictyida and 17% Lychniskida). Demospongia isrepresented by Lithistida (6% of the sponges recorded).
Brachiopod relative abundance is similar in the Ex-ternal (47%) and in the Internal Prebetic (48%, Fig. 12),with a predominance of Terebratulida (66% in the Externaland 78% in the Internal Prebetic) over Rhynchonellida. InTerebratulida, Placothyris (53%) and Dictyothyris (19%)are more abundant in the External Prebetic, and Nucleata(79%) in the Internal Prebetic (Figs. 9 and 10; and PLsection in Fig. 8). In Rhynchonellida,Monticlarella (90%)is predominant in the External Prebetic and Lacunosella(91%) is predominant in the Internal Prebetic.
Bivalves make up 28% of the fossil assemblage andare more abundant in the External Prebetic. A total of 17superfamilies have been identified, among whichMytiloidea, Pectinoidea and Pholadomyoidea are themost abundant. The relative abundance of superfamiliesvaries according to palaeogeography. In the ExternalPrebetic there is a predominance of Pectinoidea [40.3%(Entolium, Pseudovola, Aequipecten, Chlamys, Pro-peamussium and Spondilopecten)], Pholadomyoidea[18.2% (Procardia and Pholadomya)] and Nuculanacea(Isoarca 10.5%). In the Internal Prebetic, Mytiloidea[54.5% (Falcimytilus andMytiloides)] predominate, andto a lesser extent, Pholadomyoidea [11.4% (Procardiaand Pleuromya)] and Pectinoidea (11.4%).
Echinoderms include crinoids, ophiuroids and echi-noids. Disarticulate crinoids are abundant, but are notincluded in the counts. They are more abundant in the
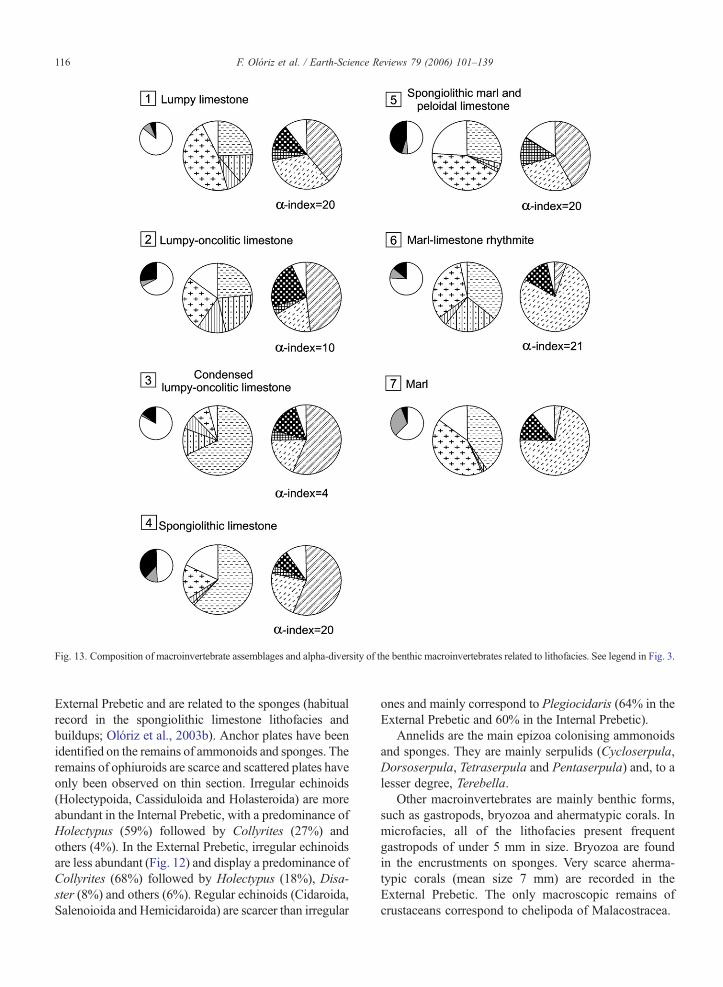
Fig. 13. Composition of macroinvertebrate assemblages and alpha-diversity of the benthic macroinvertebrates related to lithofacies. See legend in Fig. 3.
116 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
External Prebetic and are related to the sponges (habitualrecord in the spongiolithic limestone lithofacies andbuildups; Olóriz et al., 2003b). Anchor plates have beenidentified on the remains of ammonoids and sponges. Theremains of ophiuroids are scarce and scattered plates haveonly been observed on thin section. Irregular echinoids(Holectypoida, Cassiduloida and Holasteroida) are moreabundant in the Internal Prebetic, with a predominance ofHolectypus (59%) followed by Collyrites (27%) andothers (4%). In the External Prebetic, irregular echinoidsare less abundant (Fig. 12) and display a predominance ofCollyrites (68%) followed by Holectypus (18%), Disa-ster (8%) and others (6%). Regular echinoids (Cidaroida,Salenoioida and Hemicidaroida) are scarcer than irregular
ones and mainly correspond to Plegiocidaris (64% in theExternal Prebetic and 60% in the Internal Prebetic).
Annelids are the main epizoa colonising ammonoidsand sponges. They are mainly serpulids (Cycloserpula,Dorsoserpula, Tetraserpula and Pentaserpula) and, to alesser degree, Terebella.
Other macroinvertebrates are mainly benthic forms,such as gastropods, bryozoa and ahermatypic corals. Inmicrofacies, all of the lithofacies present frequentgastropods of under 5 mm in size. Bryozoa are foundin the encrustments on sponges. Very scarce aherma-typic corals (mean size 7 mm) are recorded in theExternal Prebetic. The only macroscopic remains ofcrustaceans correspond to chelipoda of Malacostracea.

117F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
4.2.1. Distribution of benthics by lithofaciesAs has been shown for foraminiferal assemblages,
the compositions of benthic macroinvertebrate assem-blages also vary with the lithofacies (Fig. 13).
(1) Lumpy limestone: Benthics make up 6% of thetotal assemblage (brachiopods 39%, bivalves 33%,irregular echinoids 12% and others 16%). Theabundance of benthics per volume of rock is low,although the diversity is high (alpha-index at thegenus level=20). In brachiopods there is a pre-dominance of Nucleata (35%), Monticlarella(23%) and Placothyris (18%), while the largestproportions of bivalves are represented by Falci-mytilus (25%) and Procardia (17%). Irregularechinoids (Collyrites, Holectypus and Disaster)are more frequent (80%) than regular echinoids.Other benthics (e.g., serpulids, bryozoa) are scarce.
(2) Lumpy–oncolitic limestone: Benthics make up27% (brachiopods 49%, irregular echinoids 21%,bivalves 19%, others 11%). The abundance ofbenthics per volume of rock varies stratigraphicallyfrommedium to high (higher in the Transversariumand Bimammatum zones) and the diversity presentsa genus-level alpha-index=10. In brachiopods theprincipal components are Nucleata (51%) and La-cunosella (34%), while the bivalves present astrong predominance of Mytiloidea (48%) (Figs. 9and 10). The irregular echinoids are mainly Ho-lectypus (55%) and Collyrites (29%). Crinoids,bryozoa, serpulids and Terebella are only abundantin relation to the local appearance of sponges.Serpulids also colonise ammonoid remains.
(3) Condensed lumpy–oncolitic limestone: Benthicsmake up 15% (brachiopods 57%, bivalves 20%,irregular echinoids 15% and others 8%). Althoughthe abundance of fossil macroinvertebrates pervolume of rock is high, the record of benthics isvery low (Fig. 13), as is its diversity (genus-levelalpha-index=4). Brachiopods are representedalmost exclusively by Nucleata (73%), bivalvesby Falcimytilus (83%) and irregular echinoids byHolectypus (55%) and Collyrites (45%). Thepresence of serpulids is limited to the colonisationof ammonoids.
(4) Spongiolithic limestone: There is a clear predom-inance of sponges, and the rest of the benthos totals39% (brachiopods 55%, bivalves 22%, others23%). Abundance and diversity (genus-levelalpha-index=20) are very high in the macroinver-tebrates, but the sponges are not very diverse,taxonomically (Stauroderma, Craticularia, Tre-
madictyon, Cribrospongia, Verrucocoelia) or mor-phologically (mainly dish-shaped sponges). Thebrachiopods are clearly dominated by Monticlar-ella (55%) and Placothyris (22%). Among bi-valves, there is a predominance of Pectinoidea(41%) and Pholadomyoidea (17%), althoughIsoarca striatissima (12%) is notable as a predom-inant species. Echinoids are mainly Collyrites(50%) and Plegiocidaris (28%), while ossiclesand crinoid stalks are common. Abundant ser-pulids and bryozoans exist in association withsponges.
(5) Spongiolithicmarl and peloidal limestone: Benthicsother than spongesmake up 45% (brachiopods 43%and bivalves 29%), and conic and ramose spongesare the most common macroinvertebrates. Regularechinoids are present in the highest proportions(13%, with their radiola being common), whileirregular echinoids are very scarce (Figs. 4 and 13).The abundance of benthics other than sponges isvery high and diversified (genus-level alpha-index=20). Brachiopods are mainly Dictyothyris(33%) andMonticlarella (33%), while bivalves arepredominantly Isoarca (29%) and pectinids (Chla-mys 23% and Propeamussium 6%). Various generaof regular echinoids are represented (mainly Ple-giocidaris, 43%), while ossicles and crinoid stalksare very scarce. Other macroinvertebrates such asserpulids and bryozoa appear in relation with colo-nisations in sponges.
(6) Marl–limestone rhythmite: Benthics constitute14% of the assemblage of fossil macroinverte-brates (bivalves 80%, irregular echinoids 11% andothers 9%). Brachiopods are very scarce (mainlyMonticlarella 38% and Placothyris 35%). Fewcrinoids and radiolae of regular echinoids havebeen recorded in sponge–microbialite buildups.Benthics are not very abundant, but their diversityis very high (genus-level alpha-index=21). Pre-dominant among bivalves are Pectinoidea (39%)and Pholadomyoidea (27%); Procardia (24%),Entolium (14%) and Propeamussium (6%) areespecially significant. Irregular echinoids arerepresented by Collyrites (62%) and to a lesserdegree by Holectypus (19%) and Disaster (14%).Serpulids are scarce and found when colonisingammonoids, but are considerably more abundanton those sponges that constitute buildups. Surfacecounting of macroinvertebrates in favourably ex-posed buildups corroborates the overwhelmingdominance of sponges (75%). Thus, benthos com-position in sponge–microbialite buildups presents
118 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
marked differences with respect to the assemblagedescribed for the surrounding marl–limestonerhythmite, as is shown by: (1) higher spongeabundance, (2) dominance of brachiopods versusbivalves, and (3) higher abundance of epizoa(serpulids) and crinoids.
(7) Marls: Benthics make up 6% of the total assem-blage and are mainly bivalves (72%) and irregularechinoids (12%). Brachiopods are very scarce,while regular echinoids, crinoids and sponges areabsent. Bivalves correspond mainly to Pectinoideaand Pholadomyoidea. The abundance of benthicsper volume of rock is low, and their poor state ofpreservation makes precise taxonomy impossible.Thus, diversity cannot be reliably evaluated.
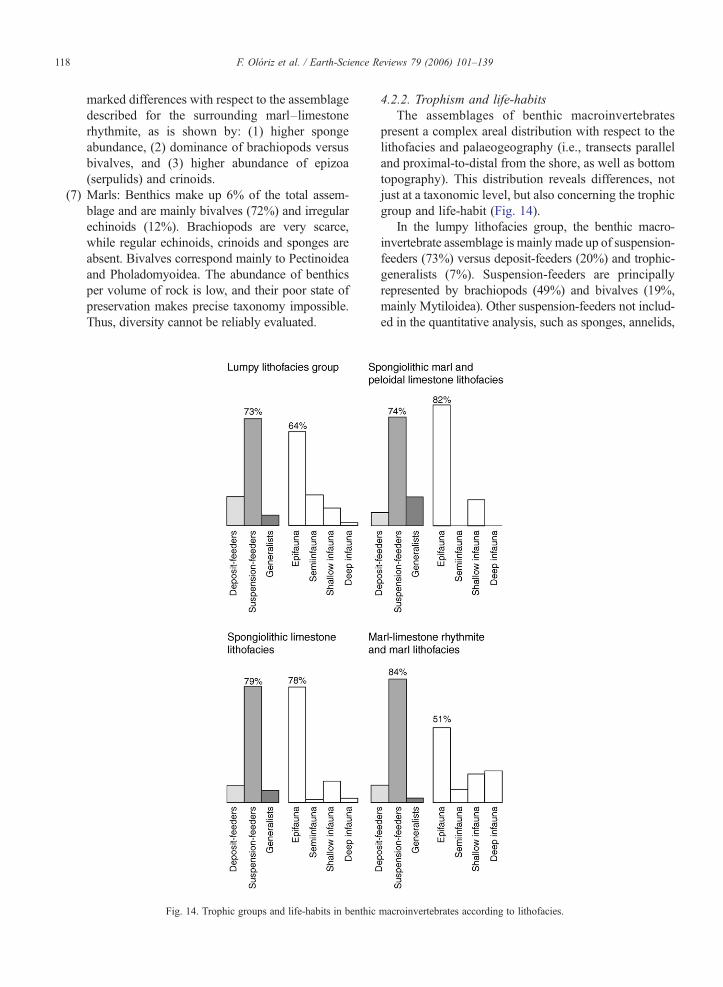
Fig. 14. Trophic groups and life-habits in benthic
4.2.2. Trophism and life-habitsThe assemblages of benthic macroinvertebrates
present a complex areal distribution with respect to thelithofacies and palaeogeography (i.e., transects paralleland proximal-to-distal from the shore, as well as bottomtopography). This distribution reveals differences, notjust at a taxonomic level, but also concerning the trophicgroup and life-habit (Fig. 14).
In the lumpy lithofacies group, the benthic macro-invertebrate assemblage is mainly made up of suspension-feeders (73%) versus deposit-feeders (20%) and trophic-generalists (7%). Suspension-feeders are principallyrepresented by brachiopods (49%) and bivalves (19%,mainly Mytiloidea). Other suspension-feeders not includ-ed in the quantitative analysis, such as sponges, annelids,
macroinvertebrates according to lithofacies.

119F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
bryozoa and crinoids, are scarce. Deposit-feeders areconstituted almost exclusively by irregular echinoids(Holectypus, Collyrites and Disaster) and a few bivalves(Nuculoida). Trophic-generalists are regular echinoids andgastropods (mainly browsers and grazers, but alsocarnivores and scavengers). The epifaunal life-habit(64%, mainly brachiopods) clearly dominates the semi-infaunal one (21%, Holectypus and endobyssate Mytiloi-dea) and the shallow infaunal one (13%, composed ofirregular echinoids and bivalves such as Crassatelloideaand Cardioidea).
In the spongiolithic limestone lithofacies, the pre-dominant trophic group is that of the suspension-feeders(dish-shaped sponges, mainly), which also predominateamong the quantified components (79%, mainlybrachiopods and Pectinoidea and Pholadomyoidea).Deposit-feeders are scarcer (12.5%) and are composedof irregular echinoids and bivalves (Nuculoidea andNuculanoidea). The epifaunal life-habit represents 78%of the total (brachiopods and bivalves, the latter found asboth epibyssally attached Pectinidae and Limidae and asunattached, free-living Entolidae). The shallow infaunallife-habit (15%) is represented by irregular echinoids(Collyrites) and bivalves (Nuculanoidea, Ceratomyoi-dea, Crassatelloidea and Cardioidea). The deep infauna(4%) is made up of bivalves (Pholadomyoidea). Thepredominance of the suspension-feeders and of theepifaunal life-habit is reinforced by the abundance ofsponges, annelids, bryozoa and crinoids; the latter twoare excluded from quantitative analysis but are signif-icant for palaeoecology.
In the spongiolithic marl and peloidal limestonelithofacies, the suspension-feeders also predominate(74%), with a high proportion of brachiopods and alower abundance of bivalves (Pectinoidea and Ostreoi-dea) (21%). This assemblage is characterised by 17%trophic-generalists (regular echinoids 76% and gastro-pods 24%) and lower values of deposit-feeders (9%,represented by I. striatissima). Epifauna reaches 82%,and is made up of brachiopods, regular echinoids andbyssated (Pectinoidea) and cemented bivalves (Ostreoi-dea). Shallow infauna (18%) corresponds to Nucula-noidea (50%) and to a lesser degree to Crassatelloidea,Carditoidea and Trigonioidea. Semi-infauna and deepinfauna are not recorded.
In the lithofacies with the highest marl content(marl–limestone rhythmite lithofacies and marl litho-facies), the macroinvertebrate fossil assemblage isbasically constituted of suspension-feeders (84%), ofwhich most are bivalves (94%) belonging to Pectinoideaand Pholadomyoidea, with scarce brachiopods (6%).There are few deposit-feeders (12%, almost exclusively
irregular echinoids) and trophic-generalists (6%, gastro-pods). The epifaunal life-habit is predominant (51%),and made up of bivalves (83%), byssated and free-livingPectinoidea (Entolium, Propeamussium, Aequipecten,Pseudovola and Camptonectes, among others) and, to alesser degree, Limoidea (Antiquilima). Infaunal life-habits are better registered than in other lithofacies (deepinfauna 22%, shallow infauna 19%, and semi-infauna8%; Fig. 14), with deep infaunal bivalves such as Pro-cardia and Pleuromya (Pholadomyoidea) (22%), andshallow infaunal ones represented by Gresslya (Cer-atomyoidea) and Grammatodon (Trigonioidea), as wellas by irregular echinoids (Collyrites).
5. Taphonomy
A detailed palaeoecological study requires a priortaphonomic analysis of both the foraminiferal assem-blages and those of the macroinvertebrates. This analysishas been performed by the authors (Olóriz et al., 2002a,b,2003a, 2004a; Reolid, 2005). In the case of theforaminifera, Olóriz et al. (2003a) and Reolid (2005)comment on the scant proportion of fragmented speci-mens, both in calcitic and in aragonitic forms (normallyb5%), this proportion being higher for nodosarids, andmainly in Lenticulina. The marl–limestone lithofaciespresents the highest proportion of fragmented foraminif-era, mainly robust forms of Lenticulina and fragments ofTubiphytes, with values of up to 57% at some points.Olóriz et al. (2004b) also noted the presence of specimensof Lenticulina with microboring. Olóriz et al. (2003a)and Reolid (2005) have interpreted the robust forms ofLenticulina as possibly allochthonous because of itsgreater abundance towards more proximal areas, andbecause the lithofacies have a higher terrigenous content.
Taphonomic analyses of macroinvertebrate fossilassemblages (Olóriz et al., 2002a,b, 2004a; Reolid,2005), have revealed a generalised autochthonous–parautochthonous nature. A bed-by-bed control oftaphonomic features was carried out, including state ofpreservation, size of the specimen/fragment, within-bedposition, corrosion, fragmentation, epibionts and encrust-ments, disarticulation, uncoupling and fossil deformation.Particular attention was given to quantification ofcorrasion and fragmentation, which were considered interms of corrasion (Ci) and fragmentation (Fi) indexes(Olóriz et al., 2002a, 2004a; Reolid, 2005).
Ci approaches the degrees of corrasion per sample, asestimated from the individuals it contains:
Ci ¼ ½ðnHCD � 100Þ þ ðnMCD� 50Þ þ ðnLCD� 1Þ�N

120 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
where N is the total number of samples (including thosewithout traces of corrasion), n, the number of samplesshowing a particular corrasion degree, HCD, the highcorrasion degree (worn N60%), MCD, the mediumcorrasion degree (worn 10-to-60%), and LCD, the lowcorrasion degree (worn b10%).
Fi treats the degrees of fragmentation per sample, asmeasured from the individuals it contains:
Fi ¼ ½ðnHFD � 100Þ þ ðnMFD� 50Þ þ ðnLFD� 1Þ�N
where N is the total number of samples (including thosewithout traces of fragmentation), n is the number of indi-viduals showing a particular fragmentation degree, HFD,the high fragmentation degree (very incomplete specimenshowing shape and size deviation), MFD, the mediumfragmentation degree (intermediate cases), and LFD, thelow fragmentation degree (near-complete specimen).
Three taphofacies have been identified: TF-I, TF-IIand TF-III. TF-I, which is found exclusively in thecondensed lumpy–oncolitic limestone lithofacies, is char-acterised by higher values of Ci (51%) and of epibiontsthat mainly affect the larger-sized remains correspondingto ammonoids. The latter may locally present abrasionfacets. However, the benthos presents the same tapho-nomic characteristics as in the background taphofacies,with a slight increase in the disarticulation of brachiopods(14%).
TF-II, which is considered the background tapho-facies, is characterised by the presence of small-sizedremains (b30 mm), a low corrasion index value(Ci≈20%), intermediate values for the fragmentationindex (Fi≈41%), a variable presence of epibionts, thegeneralised disarticulation of epifaunal to shallowinfaunal bivalves (N90%) and a scarcity of brachiopodsand deep infaunal bivalves (b12%, locally greater in thespongiolithic marl–peloidal limestone lithofacies).
TF-III, situated in most of the marly intervals in marl–limestone lithofacies and the marl lithofacies, is distin-guished from the background lithofacies by its lower Ci
(b2%) and by the presence of epibionts, together withhigher values of Fi (86%), related to fragmentation andload deformation.
The taphonomic study highlights the autochthonous–parautochthonous nature of the assemblages of fossilmacroinvertebrates that are registered, in accordance withOlóriz et al. (2004a). These authors analysed Mid- tolower–Upper Oxfordian ammonoid assemblages in thePrebetic Zone and identified taphonic populationscompatible with proximity, and/or coincidence, betweenlife and carcass-setting-areas for neritic ammonites (i.e.,
parautochthonous assemblages). Parautochthony hasbeen interpreted on the basis of relationships betweenpreservation mode, shell size, within-bed position, cor-rasion, fragmentation, and epibionts including encrust-ment. The scarcity of aptychi (b2%) and body chambers(25%) does not distort this image and may be related tolow post-mortem transport of ammonoid shells throughthe water column (Olóriz et al., 1991, 1996, 1997), pos-sibly towards more proximal areas, as was observed byOlóriz et al. (1992) in Lower Kimmeridgian assemblagesof ammonoids in the Prebetic. The alternative hypothesisof selective body-chamber fragmentation through re-working, i.e., forced by physical seabed remobilisation(Reolid, 2005), seems to be less parsimonious. This isdue to the lack of evidence of its close relationship withlithofacies (i.e., energy levels), of shell-breaking in thewater column (e.g., predation, impacts) or of sedimentaryfeatures revealing substrate erosion. Whatever the case,the short limited post-mortem transport did not cause anysignificant contamination of ammonoid assemblages.This is consistent with the scarcity of forms typical ofepioceanic environments (b1% Lytoceratina). Therefore,the ammonoid assemblages recorded are consideredparautochthonous.
6. Environmental control parameters
The interpretation of environmental, eco-sedimenta-ry parameters that control the foraminiferal and macro-invertebrate assemblages is mainly based on the analysisof benthos due to both direct dependence on thesubstrate and favourable condition for evaluating thepotential autochthony, parautochthony or allochthony.However, ammonoid assemblages were potentiallyrelated to seabed conditions via the ecological toleranceand behaviour of organisms on which they fed (e.g.,Gygi, 1999), at least as regards the group of perisphinc-toids. In any case, ammonoids were sensitive to theproximal–distal gradient, although displaying a muchweaker response than benthics, as is to be expected innektonic organisms (compare the composition ofammonite assemblages in the External vs. the InternalPrebetic; Fig. 12).
Taking into account the information obtained in thetaphonomic analysis, a clear relationship exists betweenthe recorded assemblages of foraminifera and benthicmacroinvertebrates, the proximal–distal gradient (Exter-nal and Internal Prebetic) and the lithofacies. Noteworthyamong the parameters that influence the composition anddistribution of the assemblages are the environmentalenergy, the rate of sedimentation, substrate type, nutrientavailability, oxygenation and bathymetry.

121F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
The relative stability in the composition of fossilassemblages was not treated in terms of eco-evolutionarydynamics and related topics (e.g., coordinated stasis; Brettand Baird, 1992, 1995). Except for brachiopods and thevolatile ammonites, genus-level data belonging to rela-tively wide-range taxa gathered from a stratigraphicinterval with an estimated duration of only 3.16–3.35 Ma., make such an approach inappropriate in ourcase-study. In contrast, the context of metapopulationdynamics (e.g., Gilpin and Hanksi, 1991; Wilson, 1992;Mouquet and Loreau, 2003) could be used in theinterpretation of ecostratigraphic events and trends (Olórizet al., 1995) and of the eco-evolutionary dynamics in thefuture.
6.1. Environmental energy
Present data indicate low-to-moderate energy (i.e.,below 50 cm s−1 to 70 cm s−1) in the vicinity of moundsand/or siliceous sponge aggregates at shelf depths, andselective distribution according to current energy (e.g.,Huggett et al., 1986; Mudie et al., 1986; Van Wagoner etal., 1989; Ginn et al., 2000; Krautter et al., 2001). Inaddition, recent interpretations have demonstrated root-ing of hexactinellids in muds at low-energy, nearshoresettings (e.g., Gammon et al., 2000). Since precisebalance between sponge growth and sedimentary rate iscrucial for sponge survival, current energy might becapable of scouring sponge neighbourings (e.g., as lowas 25 cm s−1). Moreover, it provides nutrients, being thatlow-flowing waters have proven disadvantageous forfilter-feeders requiring continuous, nutrient-rich inflows(e.g., Wilkinson and Vacelet, 1979). This scenario iscorroborated by sponge records in carbonate–fine-clastic rhythmites.
Apart from lithologic considerations (e.g., Wieden-mayer, 1980; Pisera, 1991, who also envisaged thepossibility for a deep environment), the occurrence ofhexactinellid sponges in the Prebetic Zone, and especiallyits particular abundance in the External Prebetic, is inter-preted as indicative of low-energy values. This fits withtheir sensitivity to water energy and preference for envi-ronments with very low energy when associated tomicrobes (Bruton and Dixon, 1994). Moreover, the thin,smooth aspect and the small peduncular foramen of thenumerous specimens of Monticlarella, which are pre-dominant in the spongiolithic lithofacies group, may beinterpreted as being related to environments with lowlevels of energy (Fürsich and Hurst, 1974). Additionally,environmental energy during deposition of the spongio-lithic limestone lithofacies must have been low enough toenable the abundant Nuculoidea and Nuculanoidea to
feed upon the organic matter available in the substrate(Delvene, 2000).Most of the infauna, especially bivalves,in the marl–limestone rhythmite and marl lithofacies arealso indicative of low-energy values. Nevertheless, thepredominance of suspension-feeders in all the assem-blages studied (Fig. 14) suggests there was sufficientenergy to maintain the organic matter in suspension or fora continuous supply to be available.
Environmental energy with respect to the lumpylithofacies group was slightly higher than for the otherlithofacies. This interpretation is based on the predom-inance of brachiopods with a thick shell, broad foramenand strong pedicle (Fürsich and Hurst, 1974), and bys-sate semi-infaunal bivalves (Delvene, 2000). Bothadaptations are indicative of a firm attachment to thesubstrate. The development of microbial oncoids withnubeculariids (Reolid et al., 2005) and the abundance oflumps in the lumpy lithofacies group are consistent withthis interpretation.
The appearance of Terebella lapilloides in all of thelithofacies studied is coherent with the interpretation ofgeneral low-energy conditions. This annellid is known tobe absent from high energy environments (Leinfelder etal., 1996; Dupraz and Strasser, 1999), although they mayappear in more energetic environments colonisingcryptic habitats such as cavities and perforations incoral–microbialite reefs (Dupraz and Strasser, 2002).Terebella associated to extant hexactinellid mounds hasbeen reported from a marine shelf under low-to-moderate currents (ca. 44 cm s−1; Huggett et al., 1986;Krautter et al., 2001).
6.2. Sedimentation rate
Low sedimentation rates have been related to envi-ronments in which the growth of present and pastsiliceous sponge aggregates (e.g., Wiedenmayer, 1980;Van Wagoner et al., 1989; Pisera, 1991; Gammon et al.,2000; Krautter et al., 2001) and a close relation betweensediment input and sponge growth have been identified(DFO, 2000). The low-energy context interpreted abovewould also present a low sedimentation rate.
The rate of sedimentation, and especially that ofterrigenous matter, was notably different between litho-facies, on the basis of the assemblages recorded. Thefall in the numbers of mainly colonial sessile forami-nifera (e.g., Nubecularia–Nubeculinella) is related tothe increase in siliciclastics, mainly fine terrigenous,towards more proximal areas. This situation is moreevident in the case of the central sector of the ExternalPrebetic, where the marl–limestone rhythmite lithofa-cies is predominant.
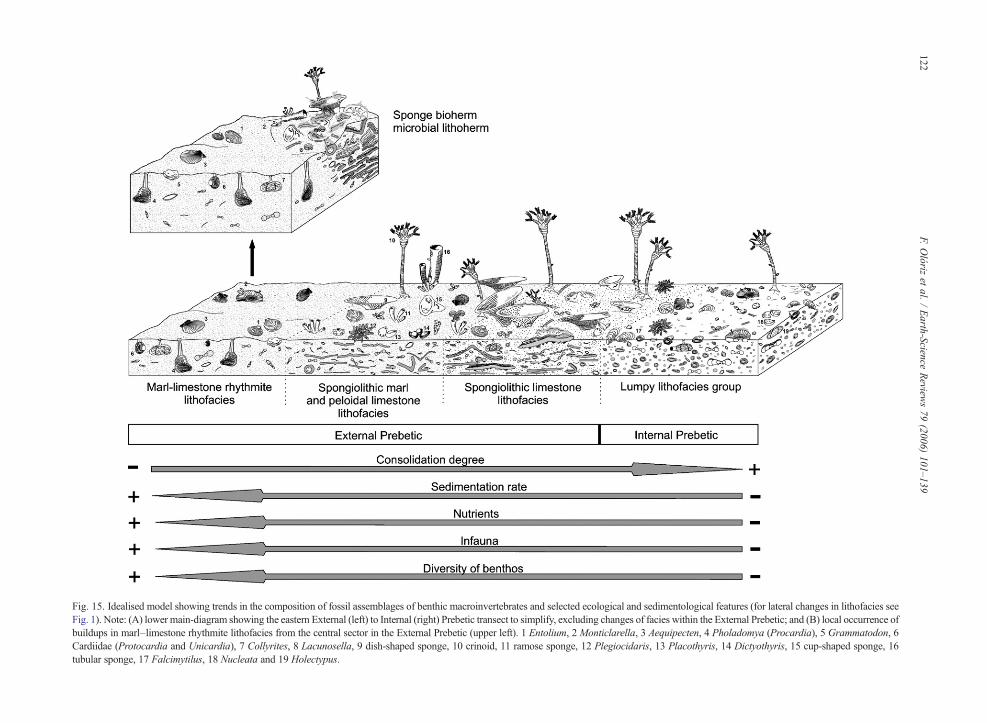
Fig. 15. Idealised model showing trends in the composition of fossil assemblages of benthic macroinvertebrates and selected ecological and sedimentological features (for lateral changes in lithofacies seeFig. 1). Note: (A) lower main-diagram showing the eastern External (left) to Internal (right) Prebetic transect to simplify, excluding changes of facies within the External Prebetic; and (B) local occurrence ofbuildups in marl–limestone rhythmite lithofacies from the central sector in the External Prebetic (upper left). 1 Entolium, 2 Monticlarella, 3 Aequipecten, 4 Pholadomya (Procardia), 5 Grammatodon, 6Cardiidae (Protocardia and Unicardia), 7 Collyrites, 8 Lacunosella, 9 dish-shaped sponge, 10 crinoid, 11 ramose sponge, 12 Plegiocidaris, 13 Placothyris, 14 Dictyothyris, 15 cup-shaped sponge, 16tubular sponge, 17 Falcimytilus, 18 Nucleata and 19 Holectypus.
122F.
Olóriz
etal.
/Earth-Science
Review
s79
(2006)101–139

123F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
With respect to the macroinvertebrates, low rates ofsedimentation favour the development of epifauna,while high rates benefit the infauna and the semi-infauna (Fürsich, 1984; Kidwell, 1985, 1986). Spongeremains in the Oxfordian of the Prebetic Zone indicatea low rate of sedimentation, which would favour thecolonisation and growth of large dish-shaped sponges,the presence of thick colonies of serpulids, bryozoaand sessile foraminifera, among others, and the deve-lopment of microbialites (e.g., Leinfelder et al., 1994;Dupraz and Strasser, 1999; Olóriz et al., 2003b; Reolidet al., 2005). Sponge remains showing the same co-loniser distribution, whatever the sponge orientation,indicate post-mortem colonisation before burial (Oló-riz et al., 2002a, 2003b).
The benthic macroinvertebrate assemblage of moredistal areas showing development of the lumpy litho-facies group (Fig. 15), presents the lowest sedimentationrates. This interpretation is consistent with the predom-inance of sessile epifaunal forms (brachiopods, bivalvesand serpulids). The scarcity of sponges and crinoids, incomparison with the register in the spongiolithic lime-stone and spongiolithic marl and peloidal limestonelithofacies, is perhaps due to the lower availability ofnutrients and may reflect the lack of nutrient-sourcesfrom open-sea currents and waves.
The large geographic extension and homogeneity ofthe spongiolithic limestone lithofacies could be relatedto the constancy of conditions of low sedimentationrates in wide areas of the eastern sector of the ExternalPrebetic. Such an environmental context would enableprolonged colonisation of the seabed by sessile epifau-nal suspension-feeders, mainly dish-shaped sponges.
In the spongiolithic marl and peloidal limestonelithofacies, the reduction in size and occurrence of dish-shaped sponges in favour of conic, ramose or tubularmorphotypes, with narrower oscules, is consistent withan increase in the sedimentation rate in comparativelyproximal areas. According to Trammer (1983), Pisera(1991), Krautter (1995, 1997, 1998) and Leinfelder et al.(1996), hexactinellids with planar morphotypes need alow or null sedimentation rate, while other morphotypeswith smaller upper surface areas intensify the exhalantcurrent and prevent the settling of fine detritic particlesunder conditions in which sedimentation rates increaseor are comparatively high.
The marl–limestone rhythmite lithofacies and marllithofacies reflect a higher rate of terrigenous deposition,which is consistent with their belonging to the mostproximal area in the palaeogeographic context studied(Fig. 15). This is coherent with a reduction in epifaunallevels. Of particular relevance is the virtual absence of
sessile epifaunal forms such as brachiopods, sponges,crinoids, serpulids and sessile foraminifera.
6.3. Substrate
The life-habit of benthic macroinvertebrates is closelyrelated to the degree of consolidation of the substrate andgrain size. Substrates that are more consolidated and/orhave a larger grain size, contain predominantly sessilesuspension-feeders, while opposing characteristics fa-vour the development of infauna (e.g., Fürsich, 1984;Fürsich and Werner, 1986; Fürsich et al., 1991; Copper,1992).
The predominance of sessile suspension-feeder epi-fauna in the lumpy lithofacies group, and in thespongiolithic limestone lithofacies, is indicative of arelatively stable, consistent substrate which was notsignificantly affected by erosive events forcing turbidityin seabed waters. The abundance and diversity of com-paratively selective suspension-feeders such as brachio-pods in these lithofacies are indicative of conditions thatwere more favourable for this group than for bivalves andechinoids. The predominance of terebratulids in thespongiolithic limestone lithofacies and in the lumpy litho-facies group corroborates the existence of a cohesivesubstrate (Almeras and Elmi, 1985), more accentuated inthe latter lithofacies (Internal Prebetic) because of thehigher proportion of terebratulids. In the lumpy lithofaciesgroup and in buildups (Olóriz et al., 2003b; Reolid, 2005),the frequent occurrence of Lacunosella reinforces thisinterpretation, because of its thick shell and welldeveloped foramen; both of these features are related toits attachment in cohesive substrates that are typical ofmoderate-energy environments (Gradinaru and Barbu-lescu, 1994). In this group of lithofacies, the abundance ofendobyssate bivalves belonging to Mytiloidea (Stanley,1972; Aberhan, 1994) is consistent with the type ofsubstrate inferred for Lacunosella.
The macroinvertebrate benthic assemblage recordedin the marl–limestone rhythmite and marl lithofacies isrelated to a less consistent substrate subjected to a higherrate of deposition of fine siliciclastics and potentialturbidity close to the seabed. In these facies, the greaterabundance of shallow and deep infauna (Figs. 14 and15) indicates a substrate that is soft enough to favourbivalves and irregular echinoids. In the marl–limestonerhythmite lithofacies, fine sediment covers hard sur-faces, thus preventing the attachment of encrustingforaminifera and of larvae of sessile macroinvertebrates.However, sponges and other epibenthics proliferatewhen a suitable substrate is locally available, thusproducing growths with sufficient relief to favour the

124 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
development of buildups (Olóriz et al., 2003b; Reolid,2005) (Fig. 15). These cases reveal the role of localbiogenically induced hard substrates that favour thecolonisation of “benthic islands” by vagile and sessilecolonisers, with a localised increase in biomass withrespect to the surrounding muddy bottom (e.g., Zuschinet al., 1999). This effect was recognized long ago forbiogenic mounds, in accordance with stage 1 of themodel proposed by Wilson (1975).
Furthermore, both grain size and the degree ofsubstrate consolidation were fundamental in the distri-bution of endobenthos. When compared to the marl–limestone rhythmite lithofacies and marl lithofacies,cohesion in spongiolithic limestone lithofacies wasfavoured by the abundance of skeletal remains ofmacroinvertebrates, larger grain size and less depositionof fine siliclastics. On the contrary, in the lumpylithofacies group, larger grain size and other associatedfeatures (such as the low sedimentation rate) wereunfavourable for deep infaunal bivalves. However, sucha context did not affect semi-infauna like Holectypus(Smith, 1984) that predominated over other irregularechinoids. In the marl–limestone rhythmite lithofacies,the smaller grain size and the higher rate of finesiliciclastic deposition favoured the abundance of deepand shallow infaunal bivalves and the predominance ofCollyrites over Holectypus among irregular echinoids.
In addition, mounds and patches made up of livingand fossil siliceous sponges have been identified in arelatively large spectrum of substrates (muddy, sandy andhard ones, including shelly bottoms). However, lessfavourable settings are fine-clastic substrates wheretixotropy and potential turbidity can be serious handicapsfor sponge growth and survival (e.g., fromWilkinson andVacelet, 1979 to Krautter et al., 2001). A sedimentarycontext of low-energy and low sedimentation rates, aspreviously interpreted, could be available for spongegrowth even under predominant fine sedimentation,especially if carbonates were present (e.g., at least firmground conditions according to Wiedenmayer, 1980).
6.4. Nutrients and oxygenation
Nutrient availability and type are interpreted throughthe analysis of trophic-group abundance and diversity inbenthic macroinvertebrate assemblages, as well as com-position in foraminiferal assemblages. The predominanceof suspension-feeders indicates the availability of nutrientsthroughout the water column, a fact which has long beeninterpreted as forcing differentiation in ecological behav-iour within a given area (e.g., Walker, 1972). However,palaeogeographic variations in fossil assemblages reveal
differences, with respect to the type of nutrients, betweenmid- and outer-shelf areas (External and Internal Prebetic,respectively). Both distal and proximal foraminiferalassemblages are dominated by epifauna. This reveals arelatively high nutrient availability (e.g., bacteria, organicmatter, dissolved abiotic nutrients) in bottom waters andthe substrate, just above the water–sediment interface(e.g., Walker and Bambach, 1974; Van der Zwaan et al.,1999). According to Turley (2000), both bacteria andforaminifera respond to organic matter deposition by anincrease in biomass and/or activity; in the case offoraminifera, these increases might result from grazingon dense populations of bacteria. Foraminifera wereprobably highly dependent on the availability of nutrients,using microbes as a stable food source (Gradzinski et al.,2004), as is done by certain present-day foraminifera(Bernhard and Bowser, 1992). Sponges could also havebeen favoured by cyanobacteria. The effect of the latter isrecognizable not only in trophic terms, especially inexposed settings (e.g., Wilkinson and Vacelet, 1979), butby facilitating the use of sponges as nutrient-levelindicators (e.g., as regards the level of soluble silicicacid, suspended/resuspended organic matter from conti-nental and/or oceanic sources). Although the completetrophic frame for sponges is not fully understood, it isclearly related to their competition-role (e.g., Dayton et al.,1974; Storr, 1976; Rice et al., 1990; Maldonado et al.,1999; Gammon et al., 2000; Krautter et al., 2001).
In the present study, the deepest layers of the sedimentare interpreted as representing hostile conditions, withreduced oxygenation and, in general, lower content inorganic matter. In this context, vertical movement capacitywould be crucial for feeding and protection against redoxboundary fluctuations, mainly for the microbiota.
6.4.1. Midshelf environment: External PrebeticAssuming that nutrient availability and diversity within
assemblages of benthic macroinvertebrates are related, atleast below certain ecologically modelled thresholds (e.g.,Valentine, 1971), a particular situation represents the case-study belonging to a midshelf environment— i.e., withinupper offshore settings ranging at intermediate waterdepths. Present data from midshelf areas show variableand/or selective productivity (e.g., Angel in Summerhayesand Thorpe, 1998; Sherr et al., 2001), which may bepatchy and higher than in inner and outer settings on theshelf. The factors responsible for this particular context arelocal influences derived from a variable depth range andenvironmental parameters forced by wind driven currents(e.g., Williams and English, 1992; Tester and Steidinger,1997), coastal advection (e.g., Chang et al., 2002; Barth etal., 2005), fronts (e.g., Holt and Raman, 1992; Thorrold,

125F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
1992; Barth et al., 2005), eddies (Wolanski and Sarsenski,1997), upwellings (Giraud and Bertrand, 2003), thermo-cline andCO2 sequestration (Bianchi et al., 2005), and sea-surface temperature gradients (Raman et al., 1998).
According to our data, nutrient availability in themidshelf External Prebetic (from major to trace andorganic nutrients) was favourable for benthos diversity inthis area, whichwas comparatively proximal among thosestudied. In fact, higher but quasi-constant values of alpha-index diversity at the genus level (20–21; Fig. 13) havebeen registered in benthic macroinvertebrate assem-blages, when compared to the record obtained frommore lumpy–oncolitic lithofacies at the outer shelf (loweroffshore settings) where alpha-diversity values were 10and 4 (Fig. 13). This trend of decreasing across-shelfmacrobenthos, before the usual shelf-break peaks, contra-dicts observations from the present (e.g., Day et al., 1971;Boesch, 1979; Neff et al., 1989), but agrees with data andinterpretations from both the present and past (e.g.,Karakassis and Eleitheriou, 1997; Smith et al., 2001).Comparative fine- to mid-scale environmental heteroge-neity, due to longshore advection and/or geostrophicflows and seabed physiography affecting nutrient distri-bution, is thought to fuel benthos diversity in the midshelfstudied (External Prebetic in Figs. 1 and 15).The slightlyhigher values of the alpha-index at the genus level inmidshelf macroinvertebrate assemblages correspond tothe marl–limestone rhythmite (21 vs. 20). This might beindicative of increased habitat differentiation within thesubstrate combined with a positive relation betweennutrient and terrigenous inputs (e.g., Fürsich et al., 1991for Middle–Upper Jurassic examples). The latter coin-cides with a general seaward nutrients decrease when noopen-sea or oceanic inputs exist (e.g., wind and/oroceanic-forced upwellings, waves, and water-massfronts), thus forcing comparative oligotrophy in outerareas on the shelf (e.g., Levinton, 1995). The relationbetween inputs of terrigenous and nutrients, and theirseaward decrease on Late Jurassic epicontinental plat-forms have been well described (Pittet and Gorin, 1997;Pittet and Mattioli, 2002; Bartolini et al., 2003; Olórizet al., 2003b; Olivier et al., 2004; Reolid, 2005). Obser-vations from the present are also in agreement with thistrend, thus indicating nutrient exportation from the innershelf seawards, and deposition at intermediate regions onthe shelf where benthos productivity is favoured (e.g.,MRRI, 1982).
In marl–limestone rhythmite lithofacies deposition offine siliciclastics was higher, and so, presumably, werenutrients. Thus, organic-matter content in the sedimentwas favoured in a context of lower oxygenation within thesubstrate (greyish sediments). These conditions are
inferred on the basis of the dominant sedimentary patternand the resulting type of substrate, which was unfavour-able for sessile epifaunal, suspension-feeder macroinver-tebrates (e.g., sponges, brachiopods, crinoids and byssatebivalves). The scarcity of this kind of suspension-feedersdetermined lower consumption of organic matter from thewater column. Increasing storage of organic matterfavours bacterial activity in the sediment–water interface.This, in turn, influences respiration (e.g., Turley, 2000),thus playing an important role in both organic matterdecay and remineralisation, as also in organic carbonsolubilisation.
The development of bacterial communities in thesediment, favoured by higher contents in both fine sili-clastics and organic matter (Lipps and Valentine, 1970;Copper, 1992), may have increased oxygen consumptionand led to variations in the redox boundary within thesubstrate. Moreover, the small grain size foreseeablyprevented the oxygenation of the substrate, thus favouringa comparatively shallower redox boundary (Fig. 16). Thiscontext would make the development of shallow infaunalforaminifera (e.g., Ammobaculites) more difficult, withrespect to forms such as Reophax, the microhabitat ofwhich shows variable depth related to fluctuations in theMn reduction front (e.g., Van der Zwaan et al., 1999). Thehigh proportion of opportunistic ubiquitous foraminifera(r-type strategists) that are potentially deep infaunal andhave greater mobility, such as Reophax and Lenticulina(Rey et al., 1994; Tyszka, 1994; Jenkins, 2000), is con-sistent with vertical migrations in the first few centimetresof the substrate. These migrations were a response topossible fluctuations in the redox boundary related toorganicmatter input/consumption rates. Thus, the Jurassicopportunist Lenticulina (Tyszka, 1994) could have beendecisive for survival and/or recolonisation of the substrateafter successive fluctuations in the redox boundary. Com-parative mesotrophy landwards in the Prebetic shelf wasinterpreted by Olóriz et al. (2003a) on basis of persistentrelationships between vagile benthic epifaunal, shallowinfaunal and ubiquitous foraminifera, the lower diversityin benthic foraminifera, and the increase in fine clastics ina context of a transgressive–regressive cycle. A similartrend for benthic foraminifera diversity was reported bySnyder et al. (1990) from the present. Reolid (2005)associated mesotrophic conditions with deposition ofmarl–calcareous rhythmite, and variations in nutrientlevels with fluctuations in terrigenous deposition frommore proximal areas. Olivier et al. (2004) performed ajoint analysis of sponge–microbialite reefs and calcareousnannoplankton in equivalent specimens from the LateOxfordian in southern Germany, interpretingmesotrophicconditions for most marly intervals, and comparatively
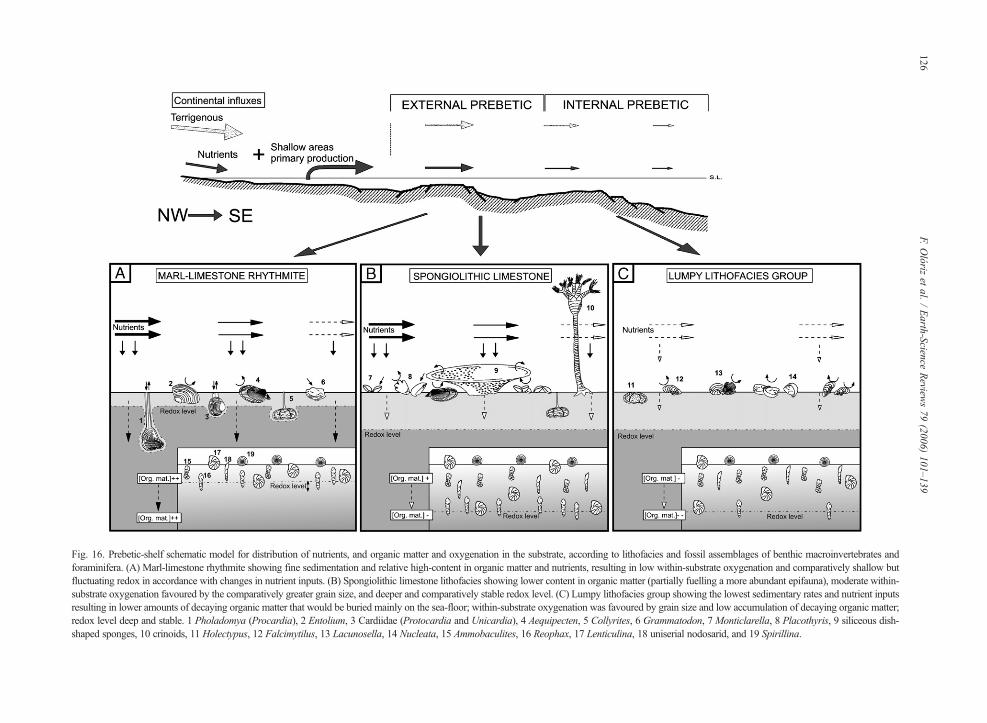
Fig. 16. Prebetic-shelf schematic model for distribution of nutrients, and organic matter and oxygenation in the substrate, according to lithofacies and fossil assemblages of benthic macroinvertebrates andforaminifera. (A) Marl-limestone rhythmite showing fine sedimentation and relative high-content in organic matter and nutrients, resulting in low within-substrate oxygenation and comparatively shallow butfluctuating redox in accordance with changes in nutrient inputs. (B) Spongiolithic limestone lithofacies showing lower content in organic matter (partially fuelling a more abundant epifauna), moderate within-substrate oxygenation favoured by the comparatively greater grain size, and deeper and comparatively stable redox level. (C) Lumpy lithofacies group showing the lowest sedimentary rates and nutrient inputsresulting in lower amounts of decaying organic matter that would be buried mainly on the sea-floor; within-substrate oxygenation was favoured by grain size and low accumulation of decaying organic matter;redox level deep and stable. 1 Pholadomya (Procardia), 2 Entolium, 3 Cardiidae (Protocardia and Unicardia), 4 Aequipecten, 5 Collyrites, 6 Grammatodon, 7Monticlarella, 8 Placothyris, 9 siliceous dish-shaped sponges, 10 crinoids, 11 Holectypus, 12 Falcimytilus, 13 Lacunosella, 14 Nucleata, 15 Ammobaculites, 16 Reophax, 17 Lenticulina, 18 uniserial nodosarid, and 19 Spirillina.
126F.
Olóriz
etal.
/Earth-Science
Review
s79
(2006)101–139

127F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
oligotrophic conditions for most carbonate ones, in whichthe appearance of buildups would be favoured.
The spongiolithic lithofacies group represents sub-strate conditions that are favourable for sponges, which atpresent depend on suspended/resuspended organic matter(Rice et al., 1990) and outcompetition of other micro-organisms (e.g., diatoms from the early Tertiary onwards)for soluble silicic acid (Maldonado et al., 1999). Thelatter arrives from continental sources delivered by rivers(e.g., Storr, 1976; Gammon et al., 2000) or from theocean via upwellings, intermediate-to-shallow, gravita-tional–internal and edge waves, and rip and tidal currents(e.g., “internal tides” in Rice et al., 1990). In the areastudied, the predominance of Hexactinellida overLithistida reflects the nature and source of nutrients.Hexactinellida sponges are filters capable of absorbingdissolved organic carbon (DOC) and colloidal substancesby osmotrophy (Leinfelder et al., 1996; Krautter, 1998);hexactinosidan depends on both direct adsorption andparticulate ingestion and colloidal organic matter (Kraut-ter, 1997). Such nutrients are scarce in more proximalareas due to their consumption by auxotrophers andheterotrophers which favour secondary and tertiaryproductivity in the photic zone. Nevertheless, remainsof phyto- and zooplankton reach relatively deep areas thathave lower energy levels, thus producing enriched DOC.These zones correspond to midshelf areas represented bythe outcrops studied in the External Prebetic, wherevariations in nutrient availability are foreseeable, basedon the well-known adaptation of siliceous sponges (bothDemospongia and Hexactinellida) to these fluctuations(from predominantly oligotrophic to clearly mesotrophicconditions; Leinfelder et al., 1993b, 1994, 1996). Theoutermost sponges could even be supported by low butconstant nutrient levels (e.g., Krautter et al., 2001).However, some authors have related the presence ofsponges, and especially sponge–microbialite consortia,to nutrient-rich environments (e.g., Dayton et al., 1974;Gaillard, 1984; Bruton and Dixon, 1994; Gradinaru andBarbulescu, 1994; Gammon et al., 2000), which in thepresent case-study could reveal patchy nutrient distribu-tion and the episodic existence of midshelf fronts.
Among the foraminifera, the high occurrence ofspirillinids is consistent with nutrient availability, giventhe relation between an abundance of Spirillina on theshelf and the availability of trophic resources (Bou-hamdi, 2000; Bouhamdi et al., 2001). Furthermore,Spirillina is known to prefer seabed surfaces colonisedby sponges, where the input of nutritive particles issignificant (Gaillard, 1984). Thus, spirillinids would beepifaunal grazers indicative of the presence of densebacterial populations related to organic carbon particles,
probably phytodetritus. According to Kitazato (1988),living Spirillina crawls on seaweeds, grazing onepiphitic diatoms and other microalgae.
In accordance with the above, the sea-floor co-lonised by sponges would present nutrient availabilityand high relative productivity (Fig. 16). In this con-text, the presence of shallow (e.g., Ammobaculites,Nodosaria, Dentalina) and potentially deep infaunalforaminifera (Reophax) could be related to a sufficientoxygenation of the seabed (mesotrophic conditions inaccordance with the TROX model of Jorissen et al.,1995), favoured by grain size variability in the sub-strate, as evidenced by microfacies. Sponge persis-tence was favoured by their relatively good adaptationto oxygenation levels from dysoxic (especially duringproductivity cycles; e.g., Krautter et al., 2001) to oxicones (e.g., Gammon et al., 2000). In addition, spongeshave been proven to be excluded only from anoxicwaters (e.g., Leys and Lauzon, 1998).
6.4.2. Outer-shelf environment: Internal PrebeticIn areas that are comparatively distal from the ex-
ternal shelf and the top of the slope (Internal Prebetic),the limited inflow of silicilastics would lead to a loweravailability of nutrients (Fig. 16). This reduction innutrient availability is reflected in the benthic macro-invertebrate assemblage, which presents lower valuesof diversity, in comparison to the record in the mid-shelf, External Prebetic (see above). The scarce evi-dence of siliceous sponges is also indicative of lowerDOC availability and, hence, a lack of marine sourcesfor nutrients (i.e., upwellings and tidal currents, amongothers). The lower occurrence of macroinvertebratebenthos contrasts with that of frequent ammonoids, forwhich the environment was more favourable. Amongforaminifera, the reduced numbers of spirillinids to-wards comparatively distal areas (Fig. 12) may alsoindicate a lower availability of nutrients (increasinglyoligotrophic conditions), which would be consumedmainly on the surface and in the first few centimetres ofthe sediment. This would explain the decrease in thepotentially deep infaunal ubiquitous foraminifera (Reo-phax) with respect to shallow infaunals (Ammobacu-lites). Substrate oxygenation in the lumpy lithofaciesgroup would have been favoured by heterogeneousgrain size and lower clay content. Van der Zwaan et al.(1999) claimed that the arrival of organic matter in theseabed decreases with depth, determining an increase inthe ratio of planktic to benthic foraminifera. Whateverthe case, distality favours the increase in planktics and,thus, the Internal Prebetic presents higher mean valuesfor planktic/benthic foraminifera (27%) than does the

128 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
External Prebetic (15%). This context may also beconsistent with the record of macroinvertebrates, whichreflects a reduction in benthics and a higher content inammonoids (78% in the Internal and 56% in the Ex-ternal Prebetic, Fig. 12).
The abundance of microbial oncoids with nubecular-iids (Reolid et al., 2005) may be interpreted as repre-senting a consortium of microbes and foraminifera welladapted to the oligotrophic conditions prevailing on thesea-floor (e.g., Gradzinski et al., 2004). Nubeculariidsand microbes (cyanobacteria) became closely relatedthrough commensalisms. Biofilms therefore served as asource of nutrition for encrusting foraminifera and assubstrate for attachment. Gradzinski et al. (2004) notedthat cyanobacteria and fungi may have taken advantageof nutrient-rich foraminiferal excretions.
6.5. Bathymetry
Although they may be controversial, evaluations ofshelf depth may be obtained using the general com-position of macroinvertebrate assemblages, along withthe individual analysis of some of their components.
On the basis of shell morphology and septal patterns,various interpretations have been made concerning thewater depth at which different groups of ammonoids lived.Westermann (1996) proposed limit–depth ranges fornekto-planktic ammonoids such as Phylloceratina andLytoceratina (Phylloceras, 65–480 m; Calliphylloceras195–335 m; Holcophylloceras, 150 m; Lytoceras, 60–400 m), and lower values for nektonic and nektobenthicammonoids (Perisphinctidae, 50–120 m; Haploceratidae,90 m; Aspidoceratidae, 25m). Other bathymetric estimateshave been based on the relative content in ammonoidsversus benthics, and among different groups of ammonoids(e.g., Ziegler, 1967; Gygi, 1986, 1999). The topic is bothintricate and controversial, (e.g., Chamberlain et al., 1981;Cecca et al., 1993; Jacobs and Chamberlain, 1996; Maedaand Seilacher, 1996; Olóriz et al., 1996, 2002d, and re-ferences therein). It is difficult to provide precise inter-pretations within a habitat-depth of 50–250 m, which mostprobably was preferred by ammonoids according to recentdata and interpretations (e.g., Henderson, 1984; Ward,1987; Hewitt, 1993, 1996; Olóriz et al., 1996;Westermann,1996; Gygi, 1999; Keupp, 1999, among others).
Fossil assemblage distribution (Fig. 12) is a trait thatcontributes ecological coherence and reliability torelative estimates regarding bathymetry in the PrebeticZone. In the External Prebetic, the mean assemblage ofmacroinvertebrates presented values of 30% in benthosand 56% in ammonoids, with a predominance of theperisphinctoids group. The rest of these macroinverte-
brates were belemnoids. In Ziegler's (1967) pioneeringmodel, the ammonoid assemblages were dominated byperisphinctids at a depth of 40 to 100 m. In the modelproposed by Gygi (1986), the characteristics of themacroinvertebrate assemblage recorded would imply adepth of between 50 and 80 m. The predominance ofDictyida in the sponge assemblages recorded in theExternal Prebetic makes it possible to interpret a mid-shelf environment with depths of around 70 m (seeLeinfelder et al., 1993b) or between 50 and 60 m (seeReitner and Neuweiler, 1995). In the comparativelydistal Internal Prebetic, the average assemblage ofmacroinvertebrates presents fewer benthics (16% anddominated by brachiopods) and more ammonoids (78%,among which the perisphinctoids group and haplocer-atids are close to 30% each). These proportions corres-pond to depths between 80 and 120 m according to theestimates made by Gygi (1986).
Presupposing a landward post-mortem transport forLate Jurassic ammonoid shells in the Prebetic Zone (Olórizet al., 1992; Reolid, 2005), it can be accepted that theammonoids recorded lead to a slight over-evaluation ofdepth. If benthic assemblageswere recorded comparativelyin situ with respect to parautochthonous ammonoidassemblages, the average of 30% benthics recorded in theExternal Prebetic would correspond to a depth of close to60 m, in agreement with Gygi (1986). Even shallowerdepths could be interpreted if the actual incidence ofnecroplanktic ammonoids within macrofossil assemblagesfrom this comparatively proximal area could be accuratelyestablished. These values are consistent with thosecalculated from the taxonomic composition of the sponges,and agree with recent interpretations (Gammon et al.,2000). They are also consistent with direct observations ofsiliceous-sponge mounds (e.g., Leys and Lauzon, 1998;Ginn et al., 2000; Krautter et al., 2001) which contrast withthe previous consensus regarding the interpretation ofdeeper-water environments for facies rich in siliceoussponges (e.g., Wiedenmayer, 1980; James and Macintyre,1985; Pisera, 1991). As regards the Internal Prebeticanalysed, depthwould be somewhat greater (around 80m),in accordance with the above-mentioned ammonoid andbenthos contents, and the possible contamination bynecroplanktic drift from more distal areas. The seawardincrease in distality, and foreseeably in depth, in the studiedsector is confirmed by the greater occurrence of Globuli-gerina, Epistomina and radiolaria in the Internal Prebetic.
7. Trophic analysis
In this study assemblages of macro and micro fossilinvertebrates, including data obtained from structures of

129F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
microbial origin, have been considered. Unlike thetrophic analyses carried out on present-day ecosystemsbased on the local community concept (the set of popu-lations of different species that occupy a given space), theanalysis of palaeocommunities (the set of populationsthat determine recurrent fossil assemblages by the relativeabundances of their components) involves numerouscomplications which derived from: (1) the difficulty indetermining components (e.g., scarcity of records ofremains belonging to organisms lacking a mineralisationcapability, allochthony, time-averaging, conservation);(2) the difficulty in inferring trophic relations betweenorganisms lacking present-day representatives, especiallyif these organisms underwent changes in their ecologicalbehaviour patterns with respect to potential extantanalogs; and (3) the difficulty in establishing the limitsof the palaeocommunity and its functional significance.
In the case studied here, a detailed taphonomic analysisreveals the auchthoctony/parauchthoctony of the fossil
Fig. 17. Simplified trophic structure (food web and food pyramid) for the midearliest Kimmeridgian. Dashed-line boxes indicate organisms with a scaunregistered or not studied (e.g., coccolithophores).
assemblages recorded (Olóriz et al., 2002a, 2004b; Reolid,2005). However, the scarcity of the ichnological recordmakes it difficult to evaluate an important partcorresponding to soft-body biota. The behaviour assumedfor the organisms to which the recorded remains cor-respond, and therefore the interpretation of trophicrelations, has followed hypotheses proposed in numerousstudies. The geographic range of the local palaeocommu-nity interpreted in the studied sector is based on palaeo-geographic terms. At first, we assume a major ecologicalsignificance for adaptation of mainly benthic and nekton–benthic organisms to an epicontinental against an epio-ceanic environment. The case studied here is that of a localpalaeocommunity of mid-outer shelf in which a cleargradation can be distinguished in foraminiferal assem-blages (distal and proximal assemblages) and in benthics(from more condensed lithofacies to those with a greatercontribution of siliciclastics), as well as in the abundanceof cephalopods. These gradients are related to gradual
-outer shelf community in the Prebetic Zone during middle Oxfordian-rce record, and grey boxes, organisms ecologically inferred, either

130 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
changes in environmental parameters, mainly induced bythe distance from the shore and depth. The latter is con-sidered an indirect parameter and one of secondary im-portance, since the range of depths interpreted for theExternal and Internal Prebetic would correspond to thedeep euphotic zone interpreted byGlaub (1994) and Vogelet al. (1999). In fact, the latter usedmicroendolitic ichnoce-nosis to interpret the development of upper Jurassicsponge–microbialite buildups in southernGermanywithintheir shallow euphotic zone III (10% of incident-lightpenetration) and deep euphotic zone (1% of incident-lightpenetration). In theGerman cases, in transparent waters theboundary between these two zones would be situated atabout 50m (Glaub, 1994) and the photic/aphotic boundaryat about 100 m (Vogel et al., 1999).
In this context, the palaeocommunity is based on aconception of the community as multispecific groupingscharacterised by their own dynamics in ecological time,but which do not, strictly speaking, maintain a taxonomiccomposition that is constant in either space (within high-order ecological bounds) or time.
7.1. Major trophic levels and trophic groups
In any community two major trophic groups arenormally distinguished in terms of their basic ecologicalrole: producers or consumers (Fig. 17); the latter aresubdivided into diverse levels and include scavengers,decomposers and transformers.
7.1.1. ProducersAutotrophic organisms in the shelf environment are
normally represented by photoautotrophs. These organ-isms are scarce in the fossil assemblages recorded, andare probably included in the BMC (cyanobacteria) andin the scarce and probably allochthonous remains ofdasycladacean algae (Terquemella), thus placing thestudy area seaward with respect to main-productiveshelf areas (i.e., inner shelf). Bertling and Insalaco(1998) suggested that microbial mats were importantprimary producers because of their partly cyanobacterialorigin. However, the bacterial component might beinterpreted as being mainly decomposers despite theproduction of carbonate. Olóriz et al. (2004b) analysedmicroboring in Upper Jurassic deposits from the InternalPrebetic and proposed, as a tentative approach, thepresence of microborers belonging to chlorophyta andfungi, and secondarily to cyanobacteria. Thus, the exis-tence of some producers was envisaged as being in-cluded in the microborers guild (chlorophyta andcyanobacteria), although their significance may besecondary. There is no doubt that most of the producers
correspond to coastal phytoplankton (dinoflagellatesand coccolithophorids) and photosynthetic algae that arenot the object of the present study.
7.1.2. ConsumersThese are heterotrophic organisms inwhich it is possible
to distinguish between primary and secondary consumers.Strictly speaking, such a distinction cannot be made in thepalaeocommunity recorded, because there are numerousmicrophagous consumers that may feed on phyto- andzooplankton and particles of organic matter that is colloidalor in suspension, and on organisms of under 100 μm (bio-seston). Moreover, the lowest-level secondary consumerscannot be identified from the record obtained. Gooday et al.(1992) and Gooday and Hughes (2002) highlighted theimportant role of foraminifera in the cycle of organicmatter, although they also noted that these protists have ahighly mobile and pervasive web of pseudopods and mayfeed voraciously at different trophic levels.
(1) Herbivores: Potential nannoplankton would beassimilated by nannoherbivores (s. Lipps andValentine, 1970), mainly planktic protozoa in-cluding radiolaria andGlobuligerina. Microfaunalbrowsers and grazers would mainly correspond toforaminifera (Spirillinidae, Nodosaridae, Ophthal-midiidae and Epistominidae) and small gastro-pods. At a macroscopic scale were regularechinoid omnivores with preferent herbivory(herbivorous preferences in Bertling and Insalaco,1998) and gastropods.
(2) Suspension-feeders: Feeders mainly on phytoplank-ton and to a lesser extent on zooplankton and otherseston particles, including the detritus of primaryproducers. In the recorded assemblages, the suspen-sion-feeders are the best-represented trophic group,with predominance of active suspension-feeders(e.g., sponges, brachiopods, bivalves, serpulids,bryozoa and ostracods) over passive ones (crinoids,ophiuroids and sessile benthic foraminifera), al-though according toBottjer andAusich (1986) somesponges and brachiopods may be active or passivesuspension-feeders. The passive behaviour s. str. is arather equivocal image for sponges, since at leastthey change orientation to be favoured by currentflows. Both epifaunal suspension-feeders (Terebra-tuloidea, Terebratelloidea, Rhynchonelloidea, Myti-loidea, Pectinoidea, among others) and infaunalsuspension-feeders (Pholadomyoidea) have beenrecorded, the latter being less abundant.
(3) Deposit-feeders: These preferentially exploit theorganic matter in the substrate (organic detritus, and

131F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
bacteria and fungi associated as coatings; Walkerand Bambach, 1974), together with meiofauna. Inthe assemblages recorded they were mainly re-presented by infaunal and semi-infaunal organismsand by epifauna to a lesser degree. Foraminifera areimportant at a microscopic level (Lipps andValentine, 1970), both agglutinated (Lituolidae,Hormosinidae and Textularidae, among others)and calcareous (Spirillinidae, Nodosaridae,Ophthalmidiidae and Hauerinidae) (Nagy, 1992;Tyszka, 1994), and to a lesser degree ostracods (notstudied). The main macroinvertebrate deposit-feeders were irregular echinoids (Holectypoida,Holasteroida andCassiduloida) and bivalves (Nucu-loida, especially I. striatissima). Ophiuroids werealso present, but it is difficult to estimate theirabundance. The rare presence of Chondrites, Pla-nolites andThalassinoides (Reolid, 2005) completesthe record of other sediment-feeder organismslacking a body-fossil record.
(4) Carnivores: Animals which combine behaviour aspredators and scavengers to a variable extent. Theymay capture their prey without persecution (e.g.,ahermatypic corals) or by means of a more or lessactive persecution or search (presumably someammonoids, belemnoids and crustaceans, ambushpredation included). Carnivores, together withsuspension-feeders, make up the trophic group pre-senting the best record and the greatest diversity. Ata microscopic level, nannoherbivores (in the case-study planktic protozoa represented by Globuli-gerina and radiolaria) were preyed upon by smallcarnivores, zooplankters and possibly chaetog-naths, pteropods and others (Lipps and Valentine,1970), or by their potential equivalents. All of theseorganisms have very low probabilities of preserva-tion. Ahermatypic corals are microcarnivores ofzooplankton and passive filters. Among themacroinvertebrates, there is a significant predom-inance of ammonoids and belemnoids; nautiloidsand gastropods are a minority. Although naticidshave not been recorded, there are some shells ofbivalves and brachiopods with characteristic bor-ing. The record of some chelipeds of malacostraceaconfirms the occurrence of crustaceans, but theirproportions cannot be estimated due to their scantfossilisation potential. Foreseeably, the componentsof the carnivore trophic group were also scavengers(e.g., cephalopods, crustaceans). Additionally,necrophagy cannot be ruled out by the occurrenceof some benthic, deposit-feeder foraminifera (e.g.,Reophax and Ammobaculites as bacterial sca-
vengers s. Nagy, 1992; Tyszka, 1994), regularechinoids and ophiuroids.
7.1.3. DecomposersSaprophagous bacteria and fungi degrade organic
matter and decompose it into nutrients and secondarymetabolites that favour nutrient cycling. The former areprobably the most important agents in the decomposi-tion of labile phytodetritus, both in the water columnand on the seabed (Turley et al., 1995). Fundamental inall ecosystems, their presence in the assemblagesrecorded must be inferred ecologically. In addition totheir role as normal symbionts and complementarynutrient-sources (for sponge sheltering and/or nutrient-sources, see Schmidt, 1970; Weissenfels, 1974, 1976;Wilkinson and Vacelet, 1979), bacteria have long beenenvisaged as involved in biogenic decay, promotingsponge preservation through accelerated carbonateprecipitation (i.e., darker than enclosing sediment“lime mummies” of the German school of research onSchwabian Jurassic Sponge Reefs and Mounds).Gaillard (1983) attributed the precipitation of carbonatesin the frame of spicules to putrefaction processes inwhich decomposers would be involved. This saproph-agous-derived carbonate is characterised by its darkercolour in relation to its organic origin, a feature that hasbeen confirmed by the darker colouration localised inreticular webs of spicules. Reitner and Schumann-Kindel (1997) and Schumann-Kindel et al. (1997)showed the importance of sulphate-reducing bacteria(SRB) in degrading sponge tissues and how they controlthe calcification of these tissues when alkalinityincreases (automicrite). Fungal activity was envisagedfrom microboring analysis conducted on Upper Jurassicmaterials in the Prebetic Zone (Olóriz et al., 2004b).
7.2. Trophic groups, food chains and food web
In order to approach trophic structure we followedCopper (1988) in terms of food chains, food web andfood pyramid. However, we assume any allusion to foodchains as a theoretical complement, and therefore weavoid mentioning precise trophic links.
7.2.1. Food chains based on nutrients in suspensionSuspension-feeders constitute the best represented
trophic group in the mid-outer shelf palaeocommunityinterpreted in this study. Primary and export productionsgenerated by phytoplankton, possibly dinoflagellates,coccolithophores and bacteria, maintained the lowermostlevels of food chain that included suspension-feeders.Thesemicroorganisms constituted themain food source for

132 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
suspension-feeders; the incorporation of zooplankton(radiolaria, Globuligerina and probably larvae of macro-invertebrates) and seston for nutrition of some suspension-feeders cannot be discounted. The food source ofbrachiopods and bryozoa was mainly composed ofphytoplankton (Fürsich and Hurst, 1974; Suchanek andLevinton, 1974; Stanton andNelson, 1980), while bivalvespossibly fed on phytoplankton, although they were notselective, and so may have included free bacteria andmicrofauna in their diet (Stanton andNelson, 1980), aswellas other basic and complex nutrients. Infaunal suspension-feeders such as Pholadomyoidea obtained most of theirfood from organic detritus retained close to the water–sediment interface. As usual in sponge aggregates, wheresponge filtering through multiple sieving can capture 80%of particles in the bacteria-size range (i.e., 0.1–0.2 μm),sponges were the main non-selective suspension-feeders inthe spongiolithic lithofacies group. Lithistida were activefilters that fed on nannoplankton,mainly free bacteria in thewater column (Leinfelder et al., 1996; Krautter, 1998),while the supposedly less effective Hexactinellida (Dic-tyida and Lychniskida) were able to absorb DOC andcolloidal substances by osmotrophy (Leinfelder et al.,1996; Krautter, 1998), although recent research indicates ahigh potential for particle ingestion (e.g., Wyeth, 1999).Sessile benthic foraminifera (Nubecularidae, Polymorphi-nidae and agglutinated forms) may have been passivesuspension-feeders of phytoplankton. Finally, although it isdifficult to be precise, a possible secondary contribution bymicrophagous ammonoidswithin this food chain cannot bediscounted.
Suspension-feeders would have been integrated intoseveral food chains and, therefore, in the envisaged foodweb, would have contributed to maintaining trophiclevels based on predation (carnivores) as well as onscavenging. In the fossil assemblages recorded, carni-vores and scavengers were represented by cephalopods,gastropods, echinoderms and possibly crustaceans.Larvae of the macroinvertebrates constituted a foodsource for numerous predators. Barbulescu et al. (1997)observed that ahermatypic corals, of microcarnivorehabit, fed on bivalve larvae. Despite the absence of fishand reptiles from the record, their existence is eco-logically highly likely, thus representing successivehigher levels, most probably, of selective predators.
7.2.2. Food chains based on substrate nutrients otherthan living plants and animals
In the registered fossil assemblages, more or lessselective depositivorous animals included ostracods,foraminifera, irregular echinoids, bivalves (Nuculoideaand Nuculanoidea) and ophiuroids. All fed on organic
matter accumulated in the substrate as particles derivedfrom the decomposition of soft tissues, and on the abun-dant bacteria related to organic coating of sedimentaryparticles (Levinton, 1982; Copper, 1992). Ostracods andforaminiferawere perhaps themain sediment-feeders, andwere doubtless consumed, in turn, by larger sediment-feeders (e.g., irregular echinoids) that ingested largevolumes of sediment without particle selection (Lipps andValentine, 1970; Stanton and Nelson, 1980; Smith, 1984).Among the macroinvertebrate deposit-feeders, the pres-ence of worms and other organisms with a low preserva-tion potential cannot be discounted; they possibly madeup some of the rare bioturbations recorded (Reolid, 2005).
Predators of infaunal sediment-feeder macroinverte-brates possibly included gastropods and crustaceans.Presumably, the predators acted indiscriminately orselectively, on epifaunal deposit-feeders and suspension-feeders (see above), and facultative scavengers such ascephalopods (e.g., ammonoids) might occasionally havebeen included due to their selective foraging. Scavengersworking on the end-products of different detritus-feederbased food chains would then have been integrated intothe food web before the action of decomposers.
7.2.3. Food chains based on phytobenthosRepresentation of browsers and grazers is lower in the
assemblages recorded. This fact is related to the scantregister of producers; the only recognized ones areencrusting BMC, possibly cyanobacteria, together withchlorophyta and endolithic cyanobacteria as potentialmakers ofmicroboring.Gaillard (1983) noted that encrust-ments produced by cyanobacteria (or by BMC) are fairlyinedible, but at least sponges should be taken into account(see above). However, Gradzinski et al. (2004) have pro-posed that microbes and organic biofilms served as a foodsource for encrusting foraminifers (nubeculariids and To-lypammina) inmicrobial–foraminiferal oncoids from con-densed deposits during Early–Middle Jurassic times in theTatra Mountains region (Poland). In reference to the ParisBasin, Bertling and Insalaco (1998) have interpreted anOxfordian reefal foodweb strongly sustained bymicrobialmats (cyanobacteria). In addition, recent endolithic algae,fungi and cyanobacteria served as food for grazers such asgastropods and echinoids in cold waters from the Van-couver Island during the past millennium (Young andNelson, 1988). Foreseeably, other producers were non-encrusting BMC and perhaps other types of algal growths,although their development could not have been verynoticeable, judging from the alternative outcompetition bysponges, i.e., algal growth favoured in more nearshoresettings, which do not crop out in the studied area. Theestimated depths would make it more difficult for the

133F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
significant development of phytobenthos, assuming arelatively low penetration of light (comparable to shalloweuphotic zone III to deep euphotic zone s. Vogel et al.,1999) determined by the level of seston concentration.Neagu et al. (1997) proposed that the abundance of herbi-vores might be correlated with the presence of vegetablecoverage, which could in turn be indicative of a shallowphotic zone. Thus, the scarcity of registered herbivoreswould confirm the localisation of the deposits studiedseaward of the proximal shallow photic zone (inner shelf),and is compatible with the relative transparency envisagedfor the water column. The trophic group of microherbi-vores is well represented by abundant foraminifera (spiril-linids, nodosarids and ophthalmidiids), but among theherbivoremacroinvertebrates, only regular echinoids (Ple-giocidaris) and some gastropods (e.g., Amberleyidae andMetriomphalus) have been recorded. Given the preserva-tion of thin shells and even of some that were originallyaragonitic (e.g., ammmonoids), the scarcity of herbivoremacroinvertebrates does not seem to be the result ofpreservation, but may be related to the absence or scarcityof benthic algae, since the abundance of encrustments byBMC did not influence the growth of herbivores (grazersand browsers). The presence of other herbivores (fish)cannot be discounted, but this may not be significant.
As in the case of the other trophic chains that areenvisaged, the incidence of rather unspecialized carni-vores and scavengers sustained by the herbivores isassumed, and would thus be integrated into the trophic orfood web.
7.3. Approaching the food-web and the food-pyramidmodel
After having identified the components of differenttrophic groups, and having interpreted their trophicrelations in a simplified manner, on the basis of thedifferentiation of envisaged food chains, a food-webmodel is proposed that integrates food chains and demon-strates the relations between producers, consumers anddecomposers that make up a local community through theinterpreted palaeocommunity (Fig. 17). Thus, the trophicor food pyramid model proposed assigns organisms in thepalaeocommunity to different trophic levels, whilerecognizing the limitations implicit in selective preserva-tion, the limited information available on nutritionalpreferences of the organisms that gave rise to the as-semblages recorded, and the fact that a given organismmay belong to various trophic levels, either at the sametime or during its ontogenetic development. The structureof present-day food pyramids is founded upon theirefficiency in transferring energy between trophic levels,
which may range from 10 to 20% (Stanton and Nelson,1980), thus imposing limitations on the length of foodchains and on the number of trophic levels. However, theeffectiveness of energy transfer between different trophiclevels may have been different in the past, taking intoaccount the possibility of changes in ecological behaviour.Another fact that makes it more difficult to describe atrophic pyramid in quantitative terms is the variation in theassemblage recorded (the ammonoid : Benthos ratio) inrelation to palaeogeography and stratigraphy; this factorwould at least lead to its areal subdivision (comparativelyproximal and distal areas) for a givenmoment. Similarly, itis difficult to calculate the biomass represented byrecorded remains (both macro and microscopic) and,evenmore so, to extrapolate such information in terms of atotal biomass (including non-recorded organisms, with orwithout a mineralised skeleton). With these limitations inmind, a simplified image is proposed of a trophic structurebased on the local palaeocommunity which is interpretedby assigning taxa to trophic levels, despite the impossi-bility of characterising them in quantitative terms. In thefood pyramid model for the local palaeocommunity of themid-outer shelf of the Prebetic during the middleOxfordian–early Kimmeridgian, the following trophiclevels have been distinguished:
Level 1: Producers (procarionts, phytoplankton and
phytobenthics) such as cyanobacteria, dasyclada-cean algae, dinoflagellates or coccolithophores;partially recorded and inferred ecologically.Level 2: Primary consumers, mainly microphagous
deposit-feeders (e.g., irregular echinoids, gastro-pods), suspension-feeders (e.g., sponges, brachio-pods, bivalves, bryozoa, serpulids), and herbivores(regular echinoids, gastropods, fishes) to a lesserdegree. Microphagous ammonoids could belong tothis trophic level. At the microscopic level, primaryconsumers would be represented by zooplankton(e.g., radiolaria, Globuligerina and macroinverte-brate larvae), ostracods and benthic foraminifera;recorded and ecologically inferred organisms.Level 3, 4 and 5: Secondary consumers.(1) Level 3: Small predators and necrophagous
organisms, both included in the carnivore trophicgroup (gastropods, ahermatypic corals, macro-invertebrate larvae and cephalopods such assmall ammonites and belemnitids); recordedorganisms.
(2) Level 4: Intermediate-level predators (carni-vores) (cephalopods such as mid-size ammo-nites, small fishes and crustaceans); recorded andecologically inferred organisms.

134 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
(3) Level 5: Higher-level predators and ‘topcarnivores’ (large ammonites, larger fishesand reptiles); ecologically inferred organisms.
8. Conclusions
The integrated study of microfossils (mainly forami-nifera) andmacroinvertebrate fossil assemblages, as wellas their relationships with lithofacies in the South Iberianepicontinental palaeomargin, the Prebetic Zone, makes itpossible to interpret palaeoecological structuring in amid-outer shelf environment during Mid-Oxfordian toearliest Kimmeridgian times. The interpretation of waterenergy, sedimentation rate, substrate type, nutrientavailability, oxygenation and bathymetry, indicates ageneral seaward trend, and displays both a decreasingsedimentation rate and nutrient input and an increasingsubstrate consistency and, presumably, depth.
The marl–limestone rhythmite and marl lithofaciesdominate in the midshelf, central sector of the ExternalPrebetic. They represent the highest values of sedimen-tation rate and nutrient availability, accompanied by thehighest abundance of epibenthic foraminifera (spirilli-nids), potentially deep infaunal foraminifera (Reophax,Lenticulina) and infaunal macroinvertebrates mainlycomposed of bivalves and echinoids (Procardia, Col-lyrites, Holectypus).
The midshelf spongiolitic lithofacies group is typicalin the eastern sector of the External Prebetic, showingintermediate values in sedimentation rates and nutrientlevels comparable to other midshelf areas studied (seeabove). There is lower abundance in both epibenthicforaminifera (spirillinids) and potentially deep infaunalforaminifera (Reophax, Lenticulina). Sessil benthicmacroinvertebrates mainly composed of siliceoussponges (Hexactinellids) and associated fauna are typical.
The lumpy lithofacies group deposited in more distalareas of the outer shelf show: (1) the lowest values insedimentation rates and nutrients; (2) the highest recordfor nektonic and planktic organisms (ammonoids, Glo-buligerina and radiolaria); and (3) scarcer spirillinids andpotentially deep infaunal foraminifera (Reophax, Lenti-culina), as well as increasing nubeculariids, BMC andsessile benthic macroinvertebrates mainly composed ofbrachiopods (Nucleata, Lacunosella) and secondarybivalves (Falcimytilus) and echinoids (Holectipus).
Lithofacies and fossil assemblages indicate a higherenvironmental heterogeneity at the midshelf and acomparatively homogeneous outer shelf. Fossil assem-blages are consistent with comparatively minor cross-shelf community differentiation forced by environmentalparameters mainly related to shore distance and nutrient
availability. No evidence of persistent ecological influ-ence from the outer shelf landwards has been found.
On the basis of a trophic-analysis approach whichintegrates BMC, foraminiferal assemblages and macro-invertebrate fossil assemblages we propose: (1) trophic-level and main trophic-group differentiation includingproducers and consumers, the latter being suspension-feeders, detritus-feeders, browsers, grazers, carnivoresand scavengers; (2) their preliminary inclusion in foodchains supported by suspension-feeders, deposit-feedersand predators (active prey-selection carnivores); and (3)a food-web integration within a pyramid–food model,considering both recorded fossils and envisaged, i.e.,ecologically inferred, organisms.
Acknowledgement
This research was financed by Project BTE2001-3029(CICYT) and the EMMI group (RNM-178, Junta deAndalucía). M. Reolid is grateful to the Universidad deGranada for a research fellowship (Ayuda Puente andPostdoctoral grant at Universitetet i Oslo and UniversitéClaude-Bernard Lyon1). The authors are also indebted toE. Mattioli (Univ. Lyon, France) and Carlton E. Brett(Univ. Cincinnati, USA) for their valuable comments andsuggestions regarding an early version of this paper.Editing of the Englishmanuscript was done byM.Bettini.
References
Aberhan, M., 1994. Guild structure and evolution of Mesozoic benthicshelf communities. Palaios 9, 516–545.
Almeras, Y., Elmi, S., 1985. Le contrôle des peuplements de brachio-podes: comparison des dominées du Jurassique et de l'actuel.Annales Societé Géologique du Nord 104, 127–140.
Barbulescu, A., Gradinaru, E., Stoica, I., 1997. Paleoecology of earlyBathonian molluscan faunas in the Codlea area (South Carpatians,Romania). Acta Palaeontogica Romaniae 1, 45–56.
Barth, J.A., Pierce, S.D., Castelao, R.M., 2005. Time-dependent, wind-driven flow over a shallow midshelf submarine bank. Journal ofGeophysical Research 110, C10S05. doi:10.1029/2004JC002761.
Bartolini, A., Pittet, B., Mattioli, E., Hunziker, J.C., 2003. Shallow-platform paleoenvironmental conditions recorded in deep-shelfsediments: C and O stable isotopes in Upper Jurassic sections ofsouthern Germany (Oxfordian–Kimmeridgian). Sedimentary Ge-ology 160, 107–130.
Bernhard, J.M., 1986. Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic, organic rich deposits: JurassicthroughHolocene. Journal of Foraminiferal Research 16, 207–215.
Bernhard, J.M., Bowser, S.S., 1992. Bacterial biofilms as a trophicresource of certain foraminifera. Marine Ecology. Progress Series(Halstenbek) 83, 263–272.
Bernier, P., 1984. Les formations carbonatées du Kimméridgien et duPortlandien dans le Jura méridional. Stratigraphie, micropaléontolo-gie, sédimentologie. Documents des Laboratoires de Géologie Lyon92, 1–803.

135F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
Bertling, M., Insalaco, E., 1998. Late Jurassic coral/microbial reefs fromthe northern Paris Basin — facies, palaeoecology and paleobiogeo-graphy. Palaeogeography, Palaeoclimatology, Palaeoecology 139,139–175.
Bianchi, A.A., Bianucci, L., Piola, A.R., Ruiz Pino, D., Schloss, I.,Poisson, A., Balestrini, C.F., 2005. Vertical stratification and air–sea CO2 fluxes in the Patagonian shelf. Journal of GeophysicalResearch 110, C07003. doi:10.1029/2004JC002488.
Boesch, D.F., 1979. Benthic ecological studies: macrobenthos. SpecialReport in Applied Marine Science and Ocean Engineering 194,Chapter 6, for BLM Contracts AA550-CT6-62. 301 pp.
Bottjer, D.J., Ausich, W.I., 1986. Phanerozoic development of tieringin soft substrata suspension feeding communities. Paleobiology12, 400–420.
Bouhamdi, A., 2000. Composition, distribution et évolution despeuplements de foraminifères benthiques de la plate-forme aubassin. Oxfordien moyen du Sud-Est de la France. Documents desLaboratoires de Géologie 151, 1–213.
Bouhamdi,A.,Gaillard,C.,Ruget,C., 2001. Spirillines versus agglutinants:Impact du flux organique et intérêt paléoenvironnemental (Oxfordienmoyen du Sud-Est de la France). Geobios 34, 267–277.
Brett, C.E., Baird, G.C., 1992. Coordinate stasis and evolutionaryecology of Silurian–Devonian marine biotas in the AppalachianBasin. Geological Society of America 24 (7), 139 (Abstracts withPrograms).
Brett, C.E., Baird, G.C., 1995. Coordinate stasis and evolutionaryecology of Silurian to Middle Devonian faunas in the AppalachianBasin. In: Erwin, D.H., Antsey, R.L. (Eds.), New Approaches toSpeciation in the Fossil Record. Columbia Univ. Press, pp. 285–315.
Bruton, F.R., Dixon, O.A., 1994. Siliceous sponge–microbe bioticassociations and their recurrence through the Phanerozoic as reefmound constructors. Palaios 9, 370–387.
Cecca, F., Fözy, I., Wierbowski, A., 1993. Ammonites du Tithonieninférieur de la Téthys occidentale: analyse des espectres fauniques.In: Elmi, S., Almeras, Y. (Eds.), 3ème Symposium InternationalCéphalopodes Actueles et Fossiles, Symposium Frederic Roman,vol. 15. Geobios, MS, pp. 39–48.
Chamberlain, J.A., Ward, P.D., Weaver, J.S., 1981. Post-mortemascent of Nautilus shells: implications for cephalopods paleobio-geography. Paleobiology 7, 494–509.
Chang, G.C., Dickey, T.D., Schafield, O.M., Weidemann, A.D.,Boss, E., Pegan, W.S., Moline, M.A., Glenn, S.M., 2002. Near-shore physical processes and bio-optical properties in the NewYork Bight. Journal of Geophysical Research 107 (C9), 3133.doi:10.1029/2001JC001018.
Copper, P., 1988. Paleoecology, paleosystems, paleocommunities.Geoscience Canada, Paleoscene 10, 199–208.
Copper, P., 1992. Organisms and carbonate substrates in marineenvironments. Geoscience Canada, Paleoscene 14, 97–112.
Corliss, B.H., 1991. Morphology and microhabitat preferences ofbenthic foraminifera from the northwest Atlantic Ocean. MarineMicropaleontology 17, 195–236.
Day, J.H., Field, J.G., Montgomery, M.P., 1971. The use of numericalmethods to determine the distribution of the benthic fauna across thecontinental shelf of North Carolina. Journal of Animal Ecology 40,93–123.
Dayton, P.K., Robillard, G.A., Paine, R.T., Dayton, L.B., 1974.Biological accommodation in the benthic community of McMurdoSound, Antarctica. Ecological Monographs 44, 105–128.
Delvene, G., 2000. Los bivalvos del Jurásico Medio and Superior en laCordillera Ibérica (España): sistemática y paleoecología. PhDThesis, Universidad de Zaragoza, Zaragoza.
DFO, 2000. Hexactinellid sponge reefs on the British Columbiacontinental shelf: geological and biological structure. DFO PacificRegion Habitat Status Report 2000/02.
Dupraz, C., Strasser, A., 1999. Microbialites and micro-encrusters inshallow coral bioherms (Middle to Late Oxfordian, Swiss JuraMountains). Facies 40, 101–130.
Dupraz, C., Strasser, A., 2002. Nutritional modes in coral–microbialitereefs (Jurassic, Oxfordian, Switzerland): evolution of trophic structureas a response to environmental change. Palaios 17, 449–471.
Fisher, R.A., Corbet, A.S., Williams, C.B., 1943. The relationsbetween the number of species and the number of individuals in aramdom sample of an animal population. Journal of AnimalEcology 12, 42–58.
Flügel, E., 1981. “Tubiphyten” aus dem fränkischen Malm. Geolo-gische Blatter für Nordost-Bayern 31, 126–142.
Flügel, E., 1982. Microfacies Analysis of Limestones. Springer-Verlag, Berlin–Heidelberg–New York.
Fürsich, F.T., 1984. Palaeoecology of boreal invertebrate faunas fromthe Upper Jurassic of Central East Greenland. Palaeogeography,Palaeoclimatology, Palaeoecology 48, 309–364.
Fürsich, F.T., Hurst, J.M., 1974. Environmental factors determiningthe distribution of brachiopods. Palaeontology 17, 879–900.
Fürsich, F.T., Werner, W., 1986. Benthic associations and theirenvironmental significance in the Lusitanian Basin (UpperJurassic, Portugal). Neues Jahrbuch für Geologie und Paläontolo-gie Abhandlungen 172, 271–329.
Fürsich, F.T., Oschmann, W., Jaitly, A.K., Singh, I.B., 1991. Faunalresponse to transgressive–regressive cycles: example from theJurassic of western India. Palaeogeography, Palaeoclimatology,Palaeoecology 85, 149–159.
Gaillard, C., 1983. Les biohermes à spongiaires et leur environmentdans l´Oxfordian du Jura meridional. Documents des Laboratoiresde Géologie Lyon 90, 1–515.
Gaillard, C., 1984. Bioconstructions jurassiques. Mémoires BureauRecherche Géologie Mineralogie France 125, 276–281.
Gammon, P.R., James, N.P., Pisera, A., 2000. Eocene spiculites andspongiolites in southwestern Australia: not deep, not polar, butshallow and warm. Geology 28, 855–858.
García-Hernández, M., López-Garrido, A.C., Olóriz, F., 1981. Étudedes calcaires noduleux du Jurassique superieur de la ZonePrébétique (Cordillères Bétiques, SE de l´Espagne). In: Farinacci,A., Elmi, S. (Eds.), Proceedings Rosso Ammonitico Symposium,Tecnoscienza, Roma, pp. 419–434.
Gilpin, M.E., Hanksi, I., 1991. Metapopulation Dynamics: Empiricaland Theoretical Investigations. Academic Press, London, UK.
Ginn, B.K., Logan, A., Thomas, M.L.H., 2000. Sponge ecology onsublittoral hard substrates in a high current velocity area. Estuarine,Coastal and Shelf Science 50, 403–414.
Giraud, X., Bertrand, Ph., 2003. Interpretation of the nitrogen isotopicsignal variations in the Mauritanian upwelling with a 2D physical–biogeochemical model. Global Biogeochemical Cycles 17 (2),1059. doi:10.1029/2002GB001951.
Glaub, I., 1994. Mikrobohrspuren in ausgewählten Ablagerungsräu-men des europäischen Jura und der Unterkreide (Klassifikationund Palökologie). Courier Forschungs-Institutt Senckenberg 174,1–324.
Gooday, A., Hughes, J.A., 2002. Foraminifera associated withphytodetritus deposits at a bathyal site in the northern RockhallTrough (NEAtlantic): seasonal contrasts and a comparison of stainedand dead assemblages. Marine Micropaleontology 46, 83–110.
Gooday, A., Levin, L., Linke, P., Heeger, T., 1992. The role of benthicforaminifera in deep-sea food webs in carbon cycling. In: Rowe, G.T.,

136 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
Pariente, V. (Eds.), Deep-sea Food Chains and the Global CarbonCycle. Kluwer Academic Publishers, The Netherlands, pp. 63–91.
Gradinaru, E., Barbulescu, A., 1994. Upper Jurassic brachiopod faunasof central and north Dobrogea (Romania): biostratigraphy, paleo-ecology and paleobiogeography. Jahrbuch der Geologischen Bun-densanstalt 137, 43–84.
Gradstein, F.M., Agterberg, F.P., Ogg, J.G., Hardenbol, J., van Veen,P., Thierry, J., Huang, Z., 1994. A Mesozoic time scale. Journal ofGeophysical Research 99, 24.051–24.074.
Gradstein, F.M., Agterberg, F.P., Ogg, J.G., Hardenbol, J., van Veen, P.,Thierry, J., Huang, Z., 1995. ATriassic, Jurassic and Cretaceous timescale. In: Berggren, W.A., Kent, D.V., Aubry, M.-P., Hardenbol, J.(Eds.), Geochronology, Time Scales and Global StratigraphicCorrelation. SEPM Special Publication, vol. 54, pp. 95–126.
Gradzinski, M., Tyszka, J., Uchman, A., Jach, R., 2004. Largemicrobial–foraminiferal oncoids from condensed Lower–MiddleJurassic deposits: a case study from the Tatra Mountains, Poland.Palaeogeography, Palaeoclimatology, Palaeoecology 213, 133–151.
Gygi, R.A., 1986. Eustatic sea level changes of the Oxfordian (LateJurassic) and their effect documented in sediments and fossilassemblages of an epicontinental sea. Eclogae geologicaeHelvetiae 79, 455–491.
Gygi, R.A., 1999. Ammonite ecology in Late Jurassic time in northernSwitzerland. Eclogae geologicae Helvetiae 92, 129–137.
Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., De Gracianski, P.C.,Vail, P.R., 1998. Mesozoic and Cenozoic sequence chronostrati-graphic framework of European basins. In: De Gracianski, P.C.,Hardenbol, J., Jacquin, T., Vail, P.R. (Eds.), Mesozoic and CenozoicSequence Stratigraphy of European Basins. SEPM Special Publica-tion, vol. 60, pp. 1–8. charts.
Henderson, R.A., 1984. A muscle attachment proposal for septalfunction in Mesozoic ammonites. Palaeontology 27, 461–486.
Hewitt, R.A., 1993. Relation of shell strength to evolution in theAmmonoidea. In: House, M.R. (Ed.), The Ammonoidea: Envi-ronment, Ecology, and Evolutionary Change. Clarendon Press,Oxford, pp. 35–56.
Hewitt, R.A., 1996. Architecture and strength of the ammonoid shell.In: Landman, N.H., Tanabe, K., Davis, R.A. (Eds.), AmmonoidPaleobiology. Topics in Geobiology, vol. 13. Plenum Press, NewYork, pp. 297–343.
Holt, T.R., Raman, S., 1992. Three-dimensional mean and turbulencestructure of a coastal front influenced by the gulf stream. MonthlyWeather Review 120 (1), 17–39.
Huggett, W.S., Thomson, R.E., Woodward, M.J., Douglas, A.N.,1986. Data record of current observations, Queen Charlotte Sound,1977, vol. 19. Institute of Ocean Sciences, Sidney, B.C. 197 pp.
Jacobs, D.K., Chamberlain, J.A., 1996. Buoyancy and hydrodynamicsin ammonoids. In: Landman, N.H., Tanabe, K., Davis, R.A. (Eds.),Ammonoid Paleobiology. Topics in Geobiology, vol. 13. PlenumPress, New York, pp. 169–224.
James, N.P., Macintyre, I.G., 1985. Carbonate depositional environ-ments. Modern and Ancient. Part I: Reefs — Zonation, Depo-sitional Facies, Diagenesis. Colorado School of Mines Quarterly,vol. 80, p. 70.
Jenkins, C.D., 2000. The ecological significance of foraminifera in theKimmeridgian of SouthernEngland. In: Hart,M.B., Kaminski,M.A.,Smart, C.W. (Eds.), Proceedings of the Fifth International Workshopon Agglutinated Foraminifera. Grzybowski Foundation SpecialPublication, vol. 7, pp. 167–178.
Jerez-Mir, L., 1973. Geología de la Zona Prebética en la transversal deElche de la Sierra y sectores adyacentes (provincias de Albacete yMurcia). PhD Thesis, Universidad de Granada, Granada.
Jorissen, F.J., De Stigter, H.C., Widmark, J.G.V., 1995. A conceptualmodel explaining benthic foraminiferal habitats. Marine Micropa-leontology 26, 3–15.
Karakassis, I., Eleitheriou, A., 1997. The continental shelf of Crete:structure of macrobenthic communities. Marine Ecology ProgressSeries 160, 185–196.
Keupp, H., 1999. Injuries — a key to understanding life modes ofammonoids. In: Histon, K. (Ed.), V International Symposium‘Cephalopods — Present and Past. Berichte der GeologischenBundesanstal, vol. 46, p. 56.
Kidwell, S.M., 1985. Palaeobiological and sedimentological implica-tions of fossil concentrations. Letters to Nature 318, 457–460.
Kidwell, S.M., 1986. Models for fossil concentrations: paleobiologicimplications. Paleobiology 12, 6–24.
Kitazato, H., 1988. Ecology of benthic foraminifera in the tidal zone ofa rocky shore. Revue Paléobiologie, Special Volume 2, 815–825.
Krautter, M., 1995. Kieselschwämme als potentielle indikatoren fürsedimentationsrate und Nährstoffagebot am beispiel der Oxford—schawammkalke von Spanien. Profil 8, 281–304.
Krautter, M., 1997. Aspekte zur paläökologie postpaläozoischerkieselschwämme. Profil 11, 199–324.
Krautter, M., 1998. Ecology of siliceous sponges: application to theenvironmental interpretation of the Upper Jurassic sponge facies(Oxfordian) fromSpain. Cuadernos deGeología Ibérica 24, 223–239.
Krautter, M., Conway, K.W., Barrie, J.V., Neuweiler, M., 2001. Discoveryof a “living dinosaur”: globally unique modern Hexactinellid spongereefs off British Columbia, Canada. Facies 44, 265–282.
Kuhnt, W., Moullade, M., Kaminski, M.A., 1996. Ecological structuringand evolution of deep sea agglutinated foraminifera — a review.Revue de Micropaléontologie 39, 271–281.
Leinfelder, R.R., Nose, M., Schmid, D.U., Werner, W., 1993a. Microbialcrust of the Late Jurassic: composition, palaeoecological signifi-cance and importance in reef constructions. Facies 29, 195–230.
Leinfelder, R.R., Krautter, M., Nose, M., Ramalho, M.M., Werner, W.,1993b. Siliceous sponge facies from the Upper Jurassic ofPortugal. Neues Jahrbuch für Geologie und Paläontologie,Abhandlungen 189, 199–254.
Leinfelder, R.R., Krautter, M., Laternser, R., Nose, M., Schmid, D.U.,Schweigert, G., Werner, W., Keupp, H., Brugger, H., Herrmann,R., Rehfeld-Kiefer, U., Schroeder, J.H., Reinhold, C., Koch, R.,Zeiss, A., Schweizer, V., Christmann, H., Menges, G., Luterbacher,H., 1994. The origin of Jurassic reefs: current research develop-ments and results. Facies 31, 1–56.
Leinfelder, R.R., Werner, W., Nose, M., Schmid, D.U., Krautter, M.,Laternser, R., Takacs, M., Hartmann, D., 1996. Paleoecology,growth parameters and dynamics of coral, sponge and microbolitereefs from the Late Jurassic. Göttinger Arbeiten zur Geologie undPaläontogie 2, 227–248.
Levinton, J.S., 1982. Marine Ecology. Prentice Hall, New Jersey.Levinton, J.S., 1995. Marine biology. Function, Biodiversity, Ecology.
Oxford University Press, New York. 420 pp.Leys, S.P., Lauzon, N.R.J., 1998. Hexactinellid sponge ecology:
growth rates and seasonality in deep water sponges. Journal ofExperimental Marine Biology and Ecology 230, 111–119.
Lipps, J.H., Valentine, J.W., 1970. The role of the foraminifera in thetrophic structure of marine communities. Lethaia 3, 279–286.
Maeda, H., Seilacher, A., 1996. Ammonoid taphonomy. In: Landman,N.H., Tanabe, K., Davis, R.A. (Eds.), Ammonoid Paleobiology.Topics inGeobiology, vol. 13. PlenumPress,NewYork, pp. 543–578.
Maldonado, M., Carmona, M.C., Uriz, M.J., Cruzado, A., 1999.Decline in Mesozoic reef-building sponges explained by siliconlimitation. Nature 401, 785–788.

137F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
Marine Resources Research Institute (MRRI), 1982. South AtlanticOCS Area Living Marine Resources Study, Year II. Vols. I–III.Final Report submitted by MRRI (S.C. Wildlife and MarineResources Department) under Contract AA551-CT1-18 to Miner-als Management Service, Washington, D.C.
Mouquet, N., Loreau, M., 2003. Community patterns in source sinkmetacommunities. American Naturalist 162, 544–557.
Mudie, P.J., Dabros, M.J., Redden, A., 1986. ISIS field report 1986:ice island sampling and investigations of sediments. GeologicalSurvey of Canada, open File 1950.
Nagy, J., 1992. Environmental significance of foraminiferal mor-phogroups in Jurassic North Sea deltas. Palaeogeography,Palaeoclimatology, Palaeoecology 95, 111–134.
Nagy, J., Gradstein, F.M., Kaminski, M.A., Holbourn, A.E., 1995.Foraminiferal morphogroups, paleoenvironments and new taxafrom Jurassic to Cretaceous strata of Thakkhola, Nepal. Grzybow-ski Foundation Special Publication 3, 181–209.
Neagu, T., Dragastan, O., Csiki, Z., 1997. Early Cretaceous shelfpaleocommunities of Cernavoda (south Dobrogea, SE Romania).Acta Paleontologica Romaniae 1, 28–36.
Neff, J.M., Bothner, M.H., Maciolek, N.J., Grassle, J.F., 1989. Impactsof exploratory drilling for oil and gas on the benthic environmentof Georges Bank. Marine Environmental Research 27, 77–114.
Olivier, N., Pittet, B., Mattioli, E., 2004. Palaeoenvironmental control onsponge–microbialite reefs and contemporaneous deep-shelf marl–limestone deposition (Late Oxfordian, southern Germany). Palaeo-geography, Palaeoclimatology, Palaeoecology 212, 233–263.
Olóriz, F., Marques, B., Rodríguez-Tovar, F.J., 1991. Eustatism andfaunal associations. Examples from the South Iberian Marginduring the Late Jurassic (Oxfordian–Kimmeridgian). Eclogaegeologicae Helvetiae 84, 83–106.
Olóriz, F., Rodríguez-Tovar, F.J., Marques, B., 1992. Asociacionesfósiles y medio deposicional. Proximalidad y profundidad dedepósito de la ritmita del Kimmeridgiense inferior (Zona Platynota)en el Prebético central. Revista de la Sociedad Geológica de España5, 89–99.
Olóriz, F., Rodríguez-Tovar, F.J., Marques, B., Caracuel, J.E., 1993.Ecostratigraphy and sequence stratigraphy in high frequency sealevel fluctuations: examples from Jurassic macroinvertebrateassemblages. Palaeogeography, Palaeoclimatology, Palaeoecology101, 131–145.
Olóriz, F., Rodríguez-Tovar, F.J., Caracuel, J.E., 1994. Faunalassemblages, ecostratigraphy and high resolution sequencestratigraphy. In: Johnson, S.D. (Ed.), High Resolution SequenceStratigraphy, Innovation and Application, Liverpool, pp. 198–203.
Olóriz, F., Caracuel, J., Rodríguez-Tovar, F.J., 1995. Using ecostrati-graphic trends in sequence stratigraphy. In: Haq, B.U. (Ed.),Sequence Stratigraphy and Depositional Response to Eustatic,Tectonic and Climatic Forcing. Kluwer Academic Publisher,Dordrecht, pp. 59–85.
Olóriz, F., Caracuel, J.E., Ruiz-Heras, J.J., Rodríguez-Tovar, F.J.,Marques, B., 1996. Ecostratigraphic approaches, sequence stratig-raphy proposals and block tectonics: examples from epioceanicswell areas in south and east Iberia. Palaeogeography, Palaeocli-matology, Palaeoecology 121, 273–295.
Olóriz, F., Villaseñor, A.B., Gonzalez-Arreola, C., 1997. Factorscontrolling Upper Jurassic ammonite assemblages in north-centralMexico. Lethaia 30, 337–351.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 1999. Fine-resolutionammonite biostratigraphy at the Rio Gazas-Chorro II section inSierra de Cazorla (Prebetic Zone, Jaén province, southern Spain).Profil 16, 83–94.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 2002a. Fossil assem-blages, lithofacies and taphofacies for interpreting depositionaldynamics in epicontinental Oxfordian (Prebetic Zone, BeticCordillera, southern Spain). Palaeogeography, Palaeoclimatology,Palaeoecology 185, 53–75.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 2002b. Taphonomicfeatures in Upper Oxfordian ammonite assemblages (BifurcatusZone) from the Navalperal section (Internal Prebetic, BeticCordillera). In: de Renzi, M., Pardo, M.V., Belinchón, M., Pe-ñalver, E., Montoya, P., Márquez-Aliaga, A. (Eds.), Currenttopics on taphonomy and fossilization, Universidad de Valen-cias, pp. 215–222.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 2002c. Approachingmicrobiotas, macroinvertebrate assemblages and the palaeoenvir-onmental evolution during the Mid–Late Oxfordian in the PrebeticZone (Betic Cordillera, southern Spain). Documents des Labor-atoires de Géologie Lyon 156, 179–180.
Olóriz, F., Palmqvist, P., Pérez-Claros, J.A., 2002d. Morphostructuralconstraints and phylogenetic overprint on sutural frilling in LateJurassic ammonites. Lethaia 35, 158–168.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 2003a. Palaeogeographicand stratigraphic distribution of Mid–Late Oxfordian foraminiferalassemblages in the Prebetic Zone (Betic Cordillera, southernSpain). Geobios 36, 733–747.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 2003b. A Late Jurassiccarbonate ramp colonized by sponges and benthic microbialcommunities (External Prebetic, Southern Spain). Palaios 18,528–545.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 2004a. Taphonomy ofammonite assemblages from the Middle–Upper Oxfordian(Transversarium?–Bifurcatus Zones) in the Internal Prebetic(Betic Cordillera, Southern Spain): Taphonic populations andtaphofacies for ecostratigraphic interpretations. Rivista Italiana diPaleontologia e Stratigraphia 110, 239–248.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., 2004b. Microboring andtaphonomy in Middle Oxfordian to lowermost Kimmeridgian(Upper Jurassic) from the Prebetic Zone (southern Iberia).Palaeogeography, Palaeoclimatology, Palaeoecology 212, 181–197.
Olóriz, F., Reolid, M., Rodríguez-Tovar, F.J., in press. Taphonomy offossil macroinvertebrate assemblages for improving ecostratigra-phy: Shifting eco-sedimentary conditions from carbonate tocarbonate–siliciclastic shelf during the early Late Jurassic(Prebetic Zone, southern Spain). Geobios.
Pisera, A., 1991. Upper Jurassic sponge megafacies in Spain: preli-minary report. In: Reitner, J., Keupp, H. (Eds.), Fossil and RecentSponges. Springer-Verlag, Berlin-Heidelberg, pp. 486–497.
Pittet, B., Gorin, G.E., 1997. Distribution of sedimentary organicmatterin amixed carbonate–siliciclastic platform environment: Oxfordianof the Swiss Jura Mountains. Sedimentology 44, 915–937.
Pittet, B., Mattioli, E., 2002. The carbonate signal and calcareousnannofossil distribution in an Upper Jurassic section (Balingen–Tieringen, Late Oxfordian, southern Germany). Palaeogeography,Palaeoclimatology, Paleoecology 179, 71–96.
Raman, S., Reddy, N.C., Niyogi, D.S., 1998. Mesoscale analysis of aCarolina coastal front. Boundary Layer Meteorology 86, 125–145.
Reitner, J., Neuweiler, F., 1995. Mud mounds: a polygenetic spectrumof fine-grained carbonate buildups. Facies 32, 1–70.
Reitner, J., Schumann-Kindel, G., 1997. Pyrite in mineralized spongetissue — product of sulfate reducing sponge related bacteria?Facies 36, 272–276.
Reolid, M., 2005. Dinámica eco-sedimentaria durante el Oxfordiensemedio-Kimmeridgiense temprano en la Zona Prebética:

138 F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
interpretación ecoestratigráfica y secuencial. PhD Thesis, Universi-dad de Granada, Granada.
Reolid, M., Gaillard, C., 2005. Microencrusters and bioclastsinteractions from Upper Jurassic spongiolithic limestones (Exter-nal Prebetic, Southern Spain). In: Martinell, J., Domènech, R.(Eds.), Abstract volume 2nd International Meeting Taphos'05,pp. 127–128.
Reolid, M., Gaillard, C., Olóriz, F., Rodríguez-Tovar, F.J., 2005.Microbial encrustations from the Middle Oxfordian–earliestKimmeridgian lithofacies in the Prebetic Zone (Betic Cordillera,southern Spain): characterization, distribution and controllingfactors. Facies 50, 529–543.
Rey, J., Bonnet, L., Cubaynes, R., Qajoun, A., Ruget, C., 1994.Sequence stratigraphy and biological signals: statistical studies ofbenthic foraminifera from Liassic series. Paleogeography, Paleo-climatology, Palaeoecology 111, 149–171.
Rice, L., Thurston, M.H., New, A.L., 1990. Dense aggregations of ahexactinellid sponge, Pheronema carpentieri, in the porcupineSeabight (northeast Atlantic Ocean), and possible causes. Progressin Oceanography 24, 179–196.
Rodríguez-Tovar, F.J., 1993. Evolución sedimentaria y ecoestratigrá-fica en plataformas epicontinentales del margen Sudibérico duranteel Kimmeridgiense inferior. PhD Thesis, Universidad de Granada,Granada.
Schmid, D.U., 1995. “Tubiphytes” morronensis: eine fakultativinkrustierende foraminifere mit endosymbiontischen algen. Profil8, 305–317.
Schmidt, I., 1970. Phagocytose et pinocytose chez les Spongillidae.Zeitschrift fur vergleichende Physiologie 66, 398–420.
Schumann-Kindel, G., Bergbauer, M., Manz, W., Szewzyk, U., Reitner,J., 1997. Aerobic and anaerobic microorganims in modern sponges:a possible relationship to fossilization-processes. In: Neuweiler, F.,Reitner, J., Monty, C. (Eds.), Biosedimentology of MicrobialBuildups IGCP project No. 380. Facies, vol. 36, pp. 268–272.
Sherr, E.B., Sherr, B.F., Cowles, T.J., 2001. Mesoscale variability inbacterial activity in the Northeast Pacific Ocean off Oregon, USA.Aquatic Microbial Ecology 25, 21–30.
Smith, A., 1984. Echinoid palaeobiology. Special Topics in Palaeon-tology. Allen and Unwin Ltd, London.
Smith, A.B., Gale, A.S., Monks, N.E.A., 2001. Sea-level change androck-record bias in the Cretaceous: a problem for extinction andbiodiversity studies. Paleobiology 27, 241–253.
Snyder, S.W., Hale, W.R., Kontrovitz, M., 1990. Distributionalpatterns of modern benthic Foraminifera on the WashingtonContinental Shelf. Micropaleontology 36, 245–258.
Stanley, S.M., 1972. Functional morphology and evolution of bysallyattached bivalve mollusks. Journal of Paleontology 46, 165–212.
Stanton Jr., R.J., Nelson, P.C., 1980. Reconstruction of the trophic webin paleontology: community structure in the Stone City Formation(Middle Eocene, Texas). Journal of Paleontology 54, 118–135.
Storr, J.F., 1976. Ecological factors controlling sponge distribution inthe Gulf of Mexico and the resulting zonation. In: Harrison, F.W.,Cowden, R.R. (Eds.), Aspects of Sponge Biology. AcademicPress, New York, pp. 261–282.
Suchanek, T.H., Levinton, J., 1974. Articulate brachiopod food.Journal of Paleontology 48, 1–5.
Summerhayes, C.P., Thorpe, S.A., 1998. Oceanography. An IllustratedGuide. Mason Publishing, London, Southampton OceanographyCentre. 352 pp.
Tester, P.A., Steidinger, K.A., 1997. Gymnodinium breve red tideblooms: Initiation, transport, and consequences circulation ofsurface. Limnology and Oceanography 42, 1039–1051.
Thorrold, S.R., 1992. Coupling of hydrography and pre-settlement reeffish distributions in the centralGreatBarrierReefLagoon. Proceedings7th International Coral Reefs Symposium, vol. 1, pp. 629–635.
Trammer, J., 1983. The relation of the morphological type of a sponge towater turbulence. Speculation in Science and Technology 6, 143–146.
Turley, C.M., 2000. Bacteria in the cold deep-sea benthic boundarylayer and sediment–water interface of the NE Atlantic. FEMSMicrobiology Ecology 33, 89–99.
Turley, C.M., Lochte, K., Lampitt, R.S., 1995. Transformation ofbiological particles during sedimentation in the northeasternAtlantic. Philosophical Transactions of the Royal Society ofLondon 348A, 179–189.
Tyszka, J., 1994. Response of Middle Jurassic benthic foraminiferalmorphogroups to dysoxic/anoxic conditions in the PieninyKlippen Basin, Polish Carpathians. Palaeogeography, Palaeocli-matology, Palaeoecology 110, 55–81.
Valentine, J.W., 1971. Resource supply and species diversity patterns.Lethaia 4, 51–61.
Van der Zwaan, G.J., Duijnstee, I.A.P., Den Dulk, M., Ernst, S.R.,Jannink, N.T., Kouwenhoven, T.J., 1999. Benthic foraminifers:proxies or problem? A review of paleoecological concepts. Earth-Science Reviews 46, 213–236.
Van Wagoner, N.A., Mudie, P.J., Cole, F.E., Daborn, G., 1989.Siliceous sponge communities, biological zonation, and recent sea-level change on the Arctic margin: ice island results. CanadianJournal of Earth Sciences 26, 2341–2355.
Vogel, K., Balog, S.J., Bundschuh, M., Gektidis, M., Glaub, I., Krut-schinna, J., Radtke, G., 1999. Bathymetrical studies in fossil reefs, withmicroendoliths as paleoecological indicators. Profil 16, 181–191.
Walker, K.R., 1972. Trophic analysis: a method for studying thefunction of ancient communities. Journal of Paleontology 46,82–93.
Walker, K.R., Bambach, R.K., 1974. Feeding by benthic invertebrates:classification and terminology for paleoecological analysis.Lethaia 7, 67–78.
Ward, P.D., 1987. The Natural History of Nautilus. Allen and Unwin,London. 267 pp.
Weissenfels, N., 1974. Bau und Funktion des SüsswasserschwammsEphydatia fluviatilis. I Das Nephridialsystem der Pinacocyten.Cytobiologie 8, 269–288.
Weissenfels, N., 1976. Bau und Funktion des SüsswasserschwammsEphydatia fluviatilis L. (Porifera) III, Nahrungsaufnahme, Ver-dauung und Defakation. Zoomorphologie 85, 73–88.
Westermann, G.E.G., 1996. Ammonoid life and habitat. In: Landman,N., Tanabe, K., Davis, R.A. (Eds.), Ammonoid Paleobiology. Topicsin Geobiology, vol. 13. Plenum Press, New York, pp. 607–707.
Wiedenmayer, F., 1980. 2 siliceous sponges. In: Hartman, W.D.,Wendt, J.W., Wiedenmayer, F. (Eds.), Living and Fossil Sponges-Notes for a Short Course, Sedimenta, vol. VIII, pp. 55–85.
Wilkinson, C.R., Vacelet, J., 1979. Transplantation of marine spongesto different conditions of light and current. Journal of ExperimentalMarine Biology and Ecology 37, 91–104.
Williams, D.McB., English, S., 1992. Distribution of fish larvaearound a coral reef: direct detection of a meso-scale, multi-specificpatch? Continental Shelf Research 12, 923–937.
Wilson, J.L., 1975. Carbonate Facies in Geologic History. SpringerVerlag, New York–Heidelberg–Berlin.
Wilson, D.S., 1992. Complex interactions in metacommunities, withimplications for biodiversity and higher levels of selection.Ecology 73, 1984–2000.
Wolanski, E., Sarsenski, J., 1997. Larvae dispersion in coral reefs andmangroves. American Scientist 85, 236–243.

139F. Olóriz et al. / Earth-Science Reviews 79 (2006) 101–139
Wyeth, R.C., 1999. Video and electron microscopy of particle feedingin sandwich cultures of the hexactinellid sponge, Rhabdocalyptusdawsoni. Invertebrate Biology 118, 236–242.
Young, H.R., Nelson, C.S., 1988. Endolithic biodegradation of cool-water skeletal carbonates on Scott shelf northwestern VancouverIsland, Canada. Sedimentary Geology 60, 251–267.
Ziegler, B., 1967. Ammoniten-Ökologie am beispiel des Oberjura.Geologische Rundschau 56, 439–464.
Zuschin, M., Stachowitsch, M., Pervesler, P., Kollmann, H., 1999.Structural features and taphonomic pathways of a high-biomassepifauna in the northern Gulf of Trieste, Adriatic Sea. Lethaia 32,299–317.
Related Documents











