Printed in the United States of America, 2014 ISBN : 978-1-63315-205-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Printed in the United States of America, 2014 ISBN : 978-1-63315-205-2

Printed in the United States of America, 2014 ISBN : 978-1-63315-205-2
Short Views on Insect Biochemistry and Molecular Biology Vol.(2), October 2014
Insect Pest Management
through Biochemical and
Molecular Approaches
© 2014
Section VII

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
595 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
Application of RNAi toward insecticide resistance management
Fang Zhu1,2*, Yingjun Cui3, Douglas B. Walsh1, Laura C. Lavine2
1 Irrigated Agriculture Research and Extension Center, Washington State University, Prosser, WA, USA 2 Department of Entomology, Washington State University, Pullman, WA, USA
3 Department of Entomology, University of Kentucky, Lexington, KY, USA
Abstract
Insecticide resistance is a major challenge for effective and sustainable Integrated Pest Management (IPM). Recent advances in genomic and post-genomic approaches not only dramatically improve our understanding of new mechanisms with regard to insecticide resistance but also provide potential tactics to manage pest populations. In this review, we focus on the latest insights on using RNA interference (RNAi) in combination with existing strategies to investigate molecular mechanisms underlying insect adaptation to insecticides. Importantly, we outline the potential for RNAi as a new alternative for pest control.
Key words: RNA interference, environmental RNAi, double-stranded RNA, insecticide resistance, adaptation, xenobiotic, detoxification, IPM alternative
*For Correspondence Email: [email protected] (Fang Zhu)
1. Introduction
Insects, as the most evolutionary successful metazoan group, have made a huge impact on human society. Insect and other arthropod pests cause severe damage to crops, forest, and livestock production in addition to propagating deadly diseases (1). Over the past 60 years, chemical control of arthropod populations remains the most common and successful pest management strategy (2). However, insects evolve robust capacities to cope with these chemical stresses which makes pest control a constant challenge (3, 4).
Vol. (1), 00 – 00, 2009 IN
TE
RN
AT
IONAL BOOK
MIS
SIO
N
Short Views on Insect Biochemistry and Molecular Biology Invited Review
Vol. (2) 595 – 619, 2014
Chapter –27

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
596 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
Overview
1. Introduction2. RNAi Approach
2a) RNAi machinery2b) RNAi delivery
3. Factors in Insecticide Resistance3a) Cytochrome P450s (CYPs) and NADPH Cytochrome P450 reductases (CPRs)3b) Hydrolases3c) Glutathione S-transferases (GSTs)3d) Cuticular proteins3e) ATP-binding cassette (ABC) transporters3f) Cadherins and aminopeptidases3g) Symbionts3h) Regulation of xenobiotic detoxification
4. Conclusion and Future Directions5. Acknowledgements6. References
Currently the initiation of thousands of arthropod genome projects (1, 5) and the availability of next-generation sequencing technologies (6, 7) combined with post- genomic approaches such as RNA interference (RNAi) provide crucial opportunities to increase our understanding of almost every branch of insect science (8-18). These new opportunities also facilitate the investigation of the genomic architecture and genetic machinery underlying the rapid adaptation of arthropod pests to chemical stresses from host plant alleochemicals (19-25), biochemicals (26, 27), and synthetic insecticides (4, 28-30). Furtherrmore, RNAi, which targets and knocks down the expression of genes in a species-specific manner, holds great potential to manage pest populations and thereby protect crops and forests, and reduce the spread of vector-borne diseases (31, 32).
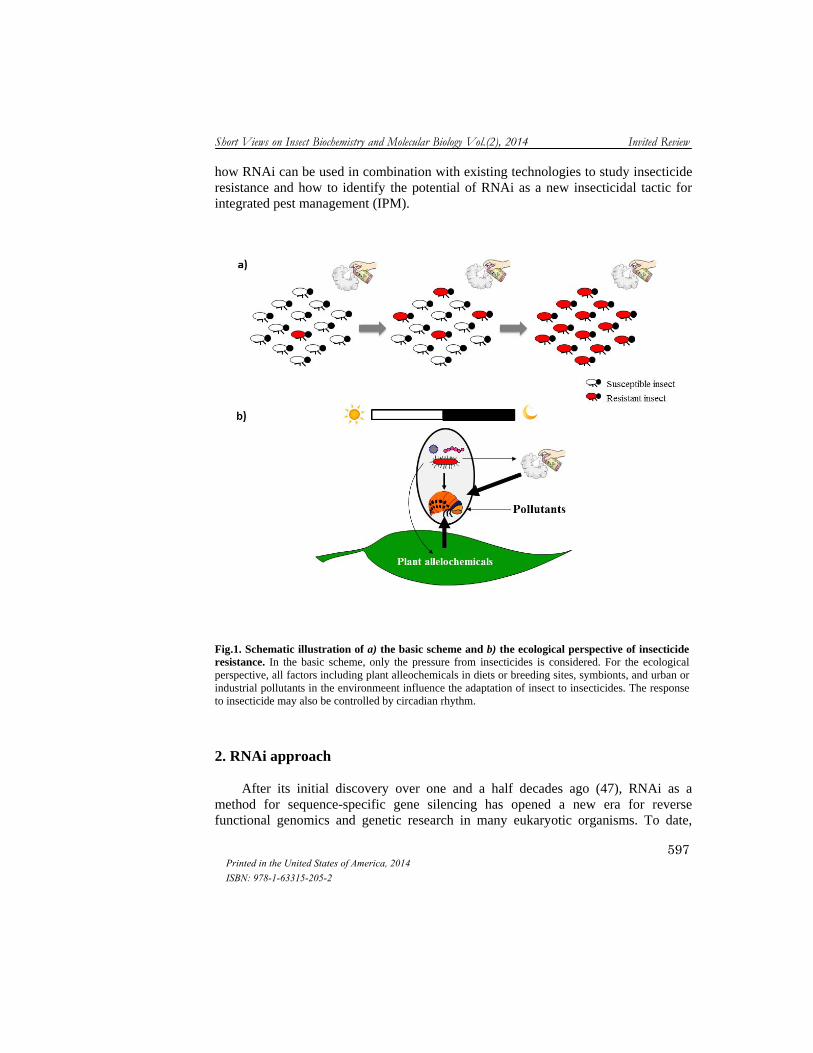
Insecticide resistance is an example of rapid and efficient adaptation through selection. The extensive application of insecticides accelerates the accumulation of resistance associated factors in survivors (Fig. 1a). Recent studies suggest that arthropod detoxification pathways found to mediate insecticide resistance may also interfere with such factors as plant alleochemicals (25, 33-35), symbionts (36-38), urban or industrial pollutants (39, 40), and circadian rhythms (41, 42) (Fig. 1b). Taken together, these results argue that studies on the molecular basis of adaptive traits are of theoretical and applied importance in understanding the evolution of insecticide resistance and devising the most effective and sustainable resistance management strategies. During the past decade, with the advance of genomic and post-genomic technologies there have been many studies uncovering the genes, pathways, and mechanisms responsible for insecticide resistance (4, 42-46). Our review focuses on
Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
597 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
how RNAi can be used in combination with existing technologies to study insecticide resistance and how to identify the potential of RNAi as a new insecticidal tactic for integrated pest management (IPM).
Fig.1. Schematic illustration of a) the basic scheme and b) the ecological perspective of insecticide resistance. In the basic scheme, only the pressure from insecticides is considered. For the ecological perspective, all factors including plant alleochemicals in diets or breeding sites, symbionts, and urban or industrial pollutants in the environmeent influence the adaptation of insect to insecticides. The response to insecticide may also be controlled by circadian rhythm.
2. RNAi approach
After its initial discovery over one and a half decades ago (47), RNAi as a method for sequence-specific gene silencing has opened a new era for reverse functional genomics and genetic research in many eukaryotic organisms. To date,

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
598 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
double-stranded RNA (dsRNA) mediated loss-of-function technique has been broadly exploited in the basic research of non-model insect species. Moreover, the remarkable systemic nature of this mechanism in insects makes RNAi-based insecticides an exciting new IPM alternative for agricultural pest control (31). Here, we review the current knowledge of the mechanism of RNAi mainly, dsRNA uptake and delivery, which are critical concepts underlying the practical application of RNAi for both basic and applied insect research.
2a) RNAi machinery
The RNAi pathway initiates when insect cells are exposed to dsRNA to trigger the post-transcriptional degradation of target mRNA sequences (Fig. 2a). The dsRNA molecules can be introduced into the cell through microinjection, transfection or expression from endogenous genes (48). dsRNA can also move into the cells from transporters SID1/2 or endocytosis machinery. The long dsRNA precursors are processed by an RNase III enzyme called Dicer into 21-25 nt-long short interference RNA (siRNA). With the assistance of dsRNA-binding motif proteins (dsRBMs) the siRNAs assemble with the RNase H enzyme Argonaute protein (AGO) into the multi-protein RNA-induced silencing complex (RISC), where one strand is destroyed and the other “guide” strand is maintained (49). Then the loaded AGO is guided by the single strand siRNA to homologous mRNAs containing complementary sequence, ultimately resulting in the degradation of the target mRNAs. The core components of the RNAi machinery are evolutionarily conserved among insect species. Typically insect genomes encode two Dicer-like proteins, Dic-1 and Dic-2. Dic-1 functions in the conversion of pre-miRNA to miRNA, whereas Dic-2 is in charge of processing long dsRNA into siRNAs (50, 51). Additionally, other core components of the RNAi machinery such as Ago 1 and 2, dsRBMs have also been observed in fruit fly, honey bee, red flour beetle, silkworm and Colorado potato beetle (51, 52).
In plants and some animals including many insects, the locally initiated RNAi spreads to remote tissues with dsRNA movement from cell to cell which triggers systemic RNAi (Fig. 2b). Insects can also generate silencing signals in response to environmentally encountered dsRNA, termed environmental RNAi (48). During environmental RNAi, the dsRNAs are taken up through the intestinal lumen while feeding followed by exportation of the dsRNA from the intestinal cells and then importation into cells of other tissues and initiation of the silencing signals in distinct tissues (Fig. 2b). This remarkable system of RNAi in insects paves the way for the potential use of RNAi as a method of pest control. SID-1 and SID-2 were originally identified in Ceanorhabditis elegans with roles in systemic and environmental RNAi (53, 54). SID-1 functions as a broadly conserved, multi-pass transmembrane protein allowing dsRNA, siRNA or other RNAi signaling to spread throughout the organism. However, SID-1 is not required for ingested dsRNA to pass through the gut lumen

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
599 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
(54). In C. elegans SID-2 is an intestinal luminal transmembrane protein and is necessary for endocytosis of dsRNA from the gut lumen (54, 55). In Drosophila S2 cells, two scavenger receptors, SR-CI and Eater were suggested to play a vital role in SID-independent dsRNA uptake (56, 57). While the core components of the RNAi machinery are conserved among different insect species, the molecular mechanism underpinning systemic spread of silencing signals vary substantially across insect taxa. For example, a single SID-1 homolog has been detected in Lepidoptera, Coleoptera, Hymenoptera, and Hemiptera but not in Diptera (58). In honey bee, SID-1 seems to be required for systemic RNAi (59). Although there were three SID-1-like (SIL) genes found in red flour beetle and silkworm, systemic RNAi in silkworm is extremely poor and is basically non-functional (51). The link between the presence of SID-1 or SIL in insect genomes and systemic RNAi remains an unanswered question in molecular biology.
A focal factor for amplifying silencing signals or siRNA and sustaining systemic RNAi in plants and C. elegans is a RNA-dependent RNA polymerase (RdRp) (60, 61). Since there is no evidence of RdRp in insects, it has been suggested that an unknown mechanism may be responsible for the systemic RNAi in insects (31, 58, 62).
Fig. 2. The process of RNAi signaling. a) RNAi process within a cell. Only the key components are illustrated; b) Environmental RNAi in insects. The different shape of circles filled with pink or green colors symbolize dissimilar type of membrane transporters.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
600 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
2b) RNAi delivery
The method of introducing dsRNA into the insect cells or the insect body directly affects the efficiency and convenience of RNAi delivery. To date, there are many tactics described and applied in RNAi experiments, including injection (63, 64), ingestion (with in vivo and in vitro synthesized dsRNA) (65, 66), transgenic plants (22, 67, 68), nanoparticle RNAi (13, 69), soaking and transfection (57, 59, 70), and topical application (71).
By far, injection of dsRNA through a microinjection system is the most common and efficient RNAi delivery mode. Injection of dsRNA has been extensively used to knockdown target genes in all life stages across various insect and arthropod taxa (70, 72-74). The site of microinjection is critical for the success of delivery. Typically it has been suggested to inject through locations where the needle is easier to insert and causes minimal damage to the insect body, such as the posterior end of the egg (75), the lateral side of the thorax (adult) (8), the suture joining the ventral mesothorax and metathorax (nymph) (76), the lateral side of the abdomen (larva or adult) (45, 64), and the spermalege of the abdomen (adult) mimicking traumatic insemination (4, 77). In addition, different species show distinct sensitivity levels to systemic RNAi (78). Other factors that influence the efficacy of systemic RNAi are: the amount of dsRNA injected into the insect, the type of tissue, and the target gene (67, 77-79). The primary limitations of RNAi by injection are that this technique can only be used in the laboratory, is labor intensive, and introduces mechanical damage either from the needle itself or from the process of immobilization (72, 73). Probably the most important drawback of dsRNA microinjection for gene silencing is that because of the apparent lack of RdRps in insects, siRNA amplification and systemic RNAi effects are restricted; therefore to sustain persistent gene silencing, dsRNA must be continuously provided until the desired effect is attained (31, 62).
To increase the efficiency of RNAi target selection and utilize RNAi for crop protection against insect pests, a more convenient delivery method must be achieved. Ingestion of dsRNA allows insects to automatically and constantly take up dsRNA from their diet and this technique (feeding RNAi) has been developed for several insects (31, 80-82). In many feeding RNAi studies, the dsRNA was produced as a result of either in vitro synthesis through enzymatic reverse transcription (65, 80, 81, 83) or in vivo synthesis by a special Escherichia coli strain (84). The specific bacterialstrain is deficient in RNAase III, an enzyme that degrades dsRNAs in the normal bacterial cell, named HT115 (DE3) (84). This method was originally developed in C. elegans for a rapid and effective genome-wide analysis of gene functions (85, 86). Subsequently, it was adopted in insect research largely due to its cost effectiveness and the potential usage in the field for pest population control and crop protection (66, 87-90).

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
601 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
For effective management of herbivorous insects, especially coleopteron, lepidopteron as well as phloem-sucking hemipteran pests, dsRNA can be synthesized through genetically modified (GM) host plants. The feasibility of expressing hairpin dsRNAs in plants to silence genes from insects has been demonstrated by recent studies in Coleoptera (67), Lepidoptera (22, 68, 91), and Hemiptera (92, 93). The plants can also be infected by Agrobacterium tumefaciens virus transformed by engineered viral vectors carrying the target gene sequence of an insect pest, which provides a rapid transgenic RNAi effect within a single generation during the spread of viruses to deliver RNAi signals throughout the plant (94-96). These technologies hold a great promise for developing species-specific, environmentally friendly RNAi-based insecticides for crop protection against insects.
The efficacy of ingested RNAi varies among insect species as a result of the variation in sensitivity of different species to systemic RNAi and the variation in gut environment of many insect species (72, 73, 97). It remains an enormous technical challenge to achieve stable dsRNA delivery in some insect species, such as mosquito, honey bee, and many species of Lepidoptera (70, 72). To increase the stability of dsRNA or siRNA and enhance cellular uptake, a chitosan/dsRNA based nanoparticle has been used in mosquito feeding RNAi which led to successful gene silencing of two chitin synthase genes (69). More recently, Li-Byarlay and colleagues coupled siRNA of DNA methyltransferase 3 to perfluocarbon nanoparticles and delivered siRNA by an aerosol application which resulted in decreased genomic methylation levels in honey bees (13).
3. Factors in insecticide resistance
The physiological and biochemical mechanisms of insecticide resistance are known to evolve along several trajectories (Fig. 3) (4, 98, 99). Behavioral resistance and decreased cuticular penetration allows insects avoiding lethal effects of insecticides as they come in contact with or consume insecticides (99-101). Once the insecticides enter the organism, enhanced metabolic detoxification, sequestration, or excretion of the toxins may take place in resistant insects before the insecticides ever reach their target sites (4, 98). Resistant insects are also likely to evolve target site insensitivity mechanisms which reduce or eliminate the binding affinity of insecticides to their target proteins (98, 102, 103). Commonly, a phenomenon of insecticide resistance is that these mechanisms operate simultaneously in resistant insect populations to contribute the overall resistance (4, 101, 104-109).
Investigation of insecticide resistance mediated genes or pathways through dsRNA related sequence-specific gene silencing is of theoretical and applied importance. For example, RNAi as a simple loss-of-function technique provides the possibility to assess the role of a particular gene or detoxification pathway in

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
602 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
insecticide resistance. Once these genes and signaling pathways are identified, they can then be used as a target of RNAi-based strategies for the control of insect pests. Many factors from detoxification pathways to symbionts are known to be involved in the development of insecticide resistance (Fig. 1b). Each of these factors is expected to have a high potential to serve as RNAi targets for pest control. Some insect Cytochrome P450s (CYPs) that play a predominant role in insecticide resistance could serve as RNAi-based insecticide targets themselves. For instance, the house fly gene CYP6D1 is primarily responsible for pyrethroid resistance in the highly resistant LPR strain (110), a single Drosophila CYP, CYP6G1 is primarily responsible for resistance to DDT (43), CYP6BQ9 in the red flour beetle is a major contributor of deltamethrin resistance in the QTC279 strain (45), and a chimeric CYP337B3 in the Australian Helicoverpa armigera confers a 42-fold resistance to fenvalerate (111). In addition to these promising potential RNAi targets, transcription factors that universally regulate the transcription of many detoxification genes may serve as even more robust targets (46). In this review we discuss the most current discoveries in several gene families and signaling pathways as well as other biotic factors related to insecticide resistance that could be used as targets of RNAi.
Fig. 3. Major physiological and biochemical mechanisms of insecticide resistance. The thickness of the arrow represents the concentration of insecticides.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
603 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
3a) Cytochrome P450s (CYPs) and NADPH Cytochrome P450 reductases (CPRs)
As significant players in our biosphere, CYPs constitute one of the largest and oldest gene superfamilies that are found in virtually all aerobic organisms (112). Insects typically contain tens to more than one hundred individual P450 genes in their genomes (113-116). Over-expression of CYPs contributes to the resistance and cross-resistance of animal cells to a broad range of xenobiotics by virtue of their extraordinarily flexible substrate specificity and catalytic versatility (35, 117). There are two hallmarks of CYPs associated with insecticide detoxification, constitutive overexpression and induction in insecticide resistant strains (107, 118, 119). In a classic study performed by Mao et al., the in planta RNAi of the gossypol induced CYP6AE14 caused decreased tolerance of gossypol in the cotton bollworm (68). In a follow-up study, five more gossypol induced CYPs were suggested to contribute tolerance to deltamethrin in cotton bollworm (22). Plant-derived RNAi of CYP9A14 led to increased sensitivity of larva to deltamethrin, indicating that the cotton bolloworm might take advantage of the plant alleochemicals (gossypol) in their diet to detoxify synthetic insecticides (22). Knockdown of CYP6BG1 in a larval diamondback moth population resistant to permethrin caused a reduction in total P450 activity in the midgut of larvae; clearly implicating the involvement of CYP in permethrin resistance in this moth (120). A brain-specific CYP, CYP6BQ9 in the red flour beetle has been demonstrated to be responsible for the majority of extremely high deltamethrin resistance to beetles of QTC279 strain supported by evidence from RNAi, microarray data, transgenic overexpression in D. melanogaster, and deltmethrin metabolism analysis (45). In another study, after silencing two new deltamethrin induced CYPs, CYP408B1 and CYP409A1, the mortality of oriental migratory locust exposed to deltamethrin significantly increased, suggesting the involvement of these two CYPs in deltamethrin resistance (121). A recent study has shown that the RNAi silencing of four highly expressed CYPs in pyrethroid resistant bed bugs simultaneously caused significantly more susceptible to pyrethroid insecticide, confirming the CYP mediated detoxification was a key mechanism of resistance in bed bugs (4).
As the required electron donor for all CYPs, insect CPRs play a vital role in metabolism-based insecticide resistance. Knocking down the CPR in an organism would effectively disable all CYPs in this organism since the CPR is required for electrons transferring from Nicotinamide Adenine Dinucleotide Phosphate (NADPH) to the P450 heme center; therefore it is an exciting potential target for the development of an RNAi-based insecticide (77, 122). dsRNA silencing of CPR had been achieved in Anopheles gambiae (122), Cimex lectularius (77), and Helicoverpa armigera (123), resulting in the increased susceptibility of these insects to permethrin, deltamethrin, and fenvalerate, respectively. Applying these results from the laboratory to the field is an exciting prospect for future control efforts.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
604 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
3b) Hydrolases
Hydrolases or esterases including carboxylesterases and phosphotriester hydrolases represent a large variety of enzymes present in most organisms (124). Hydrolases are involved in the metabolism of organophosphates, carbamates, pyrethroids, and juvenile hormone (102, 124, 125). There are at least two mechanisms in hydrolase-mediated detoxification: non-elevated and elevated esterase-based mechanisms. Non-elevated esterase-based mechanisms have a mutation in carboxylesterase which causes the loss of its carboxylesterase activity but a gain of hydrolase activity. Elevated esterase-based mechanisms exhibit increased carboxyl -esterase activity through gene amplification, protecting insects by binding and sequestering insecticides (98, 102). Increased carboxylesterase activity has been shown to be the major detoxification mechanism responsible for malathion resistance in field populations of the oriental migratory locust (126). RNAi silencing of two highly expressed caboxylesterase genes, LmCarE9 and LmCarE25, in these malathion resistant locusts significantly increased mortality when exposed to malathion (126). These same caboxylesterases were also suggested to play a possible role in locust resistance to chlorpyrifos (127). In another study that used plant-mediated RNAi, a carboxylesterase, Cbe E4 reduced the tolerance of wheat aphids to the organophosphate pesticide phoxim (128). These examples show that hydrolases are also a potential candidate target for RNAi mediated pest control.
3c) Glutathione S-transferases (GSTs)
The GSTs are a superfamily of detoxification enzymes that act by conjugating the reduced glutathione (GSH) to insecticides, making them more soluble and thus easier to excrete (102, 129). With a broad substrate range to an extremely wide variety of xenobiotics, elevated levels of GSTs have been linked in detoxification of insecticides in resistant insects (129, 130). The loss-of-function approach provides additional evidence for GSTs’ function. For instance, in the mosquito Aedes aegypti, the partial knockdown of two GSTs, GSTe7 and GSTe2, led to increased susceptibility to deltamethrin, but not DDT (131). In the oriental migratory locust, RNAi silencing technology combined with biochemistry studies revealed that a sigma GST LmGSTs3 was involved in carbaryl detoxification (132). Two GSTs identified from one rice planthopper were induced by multiple insecticides in a resistant strain (133). After silencing the expression of these two GSTs, planthoppers increased their sensitivity to chlorpyrifos (133). Most recently, two more GSTs were specifically silenced from the migratory locust which resulted in increased mortality of insects when exposed to discriminating doses of malathion and carbaryl (ImGSTs5) or chlorpyrifos and carbaryl (ImGSTu1) (134). Clearly, from the examples given, GSTs are likely to play important roles in insecticide resistance.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
605 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
3d) Cuticular proteins
Cuticular proteins are the major components of insect cuticle serving as the first line of defense to insecticides. Almost 2% of protein coding genes in insects are cuticular proteins. It has been reported that cuticle thickening is associated with pyrethroid resistance in the mosquito Anopheles funestus (135). Further evidence that cuticular proteins are involved in insecticide resistance comes from the Colorado potato beetle; mRNA levels of three putative cuticular proteins were higher in azinphosmethyl resistant Colorado potato beetles than in susceptible beetles (136). Recent transcriptomic analyses also illustrate the pervasiveness of the link between up regulation of multiple genes encoding cuticular proteins with insecticide resistance (29, 137). Moreover, in mosquito larvae, the presence of pollutants can affect the expression of genes encoding cuticular proteins (39, 138). Besides cuticular proteins, any other xenobiotic responsible genes involved in cuticle formation and dynamics are also expected to play key roles in cuticle-mediated insecticide resistance and potentially serve as a candidate target for RNAi mediated pest control (99, 139).
3e) ATP-binding cassette (ABC) transporters
ABC transporters constitute one of the largest classes of transporters that are responsible for the ATP-powered translocation of many substrates across membranes (140). These substrates are numerous and include ions, sugars, amino acids, vitamins, peptides, polysaccharides, hormones, lipids and xenobiotics (141). In insects, ABC transporters have been shown to be involved in insecticide resistance in several studies (141-145). ABC transporters are considered to act during the last phase of the detoxification process to transport polar compounds or conjugates out of cells (143). Next-generation sequencing and microarray studies have revealed that the up-regulation of many ABC transporters is clearly associated with insecticide resistance (5, 29, 108, 109, 146-149). Functional studies of ABC transporters also support their role in insecticide detoxification. In the human body louse the silencing of one ivermectin-induced ABC transporter PhABCC4, increased the susceptibility of female lice to ivermectin (150). In A. aegypti, the up-regulation of an ABC transporter was correlated with organophosphate treatment in larvae and when knocked down with RNAi, was found to increase the susceptibility of larvae to the insecticide (151). A more recent study showed that four ABC transporters, Abc8-11 are up-regulated in pyrethroid resistant bed bug populations (4). When two of these ABC transporters were silenced in the resistant bed bug populations, the dsRNA treated individuals were more susceptible to pyrethroids (4). ABC transporters are also considered to be involved in the transport of plant secondary compounds (152-155) or function as a pacemaker for sequestration of plant toxins (156). These discoveries implicate ABC transporters as key regulators in insecticide resistance and support their involvement in cross-resistance between plant alleochemicals and synthetic insecticides. Recent

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
606 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
genetic studies in four Lepidopteran species revealed that the ABC transporter ABCC2 plays a key role in the insertion of the pre-pore structure into the membrane of the midgut epithelial cells as a receptor for Bacillus thuringiensis (Bt) Cry1A toxins (143, 157, 158). The non-synonymous substitutions on ABCC2 confer high levels of resistance to Cry1A toxins thereby threatening the extensive usage of transgenic Bt crops (142, 159, 160). ABC transporters are outstanding candidates for RNAi based pest control strategies due to the diversity of subtypes and their functions in insecticide detoxification.
3f) Cadherin and aminopeptidases
As biopesticides, Bt toxins have been used for nearly 60 years in the control of agriculturally, medically, and economically important insect pests (161). The first Bt transgenic crops were commercialized in 1996, and since then, more than 1 billion acres have been planted with Bt crops worldwide (27). Understanding the mechanisms of Bt resistance is fundamental in global monitoring and enhancing the sustainable usage of Bt crops in agricultural pest control. Careful management practices have been recommended to decrease the time that insects are likely to develop resistance to Bt toxins. Nevertheless, Bt resistance can occur at any step during the mode of action of Bt toxins, including solubilization, proteolytic processing, passage through the peritrophic membrane, receptor binding, membrane insertion, pore formation, and osmotic lysis of midgut cells (162, 163). Among these, the most common mechanism of Bt resistance is the disruption of Bt toxin binding to midgut receptors (162, 164). dsRNA mediated silencing of midgut cadherin or aminopeptidase genes in Spodoptera litura (165), Manduca sexta (166), Helicoverpa armigera (167), and Ostrinia nubilalis (168) resulted in increased resistance to Bt toxins. Further investigations towards the role of cadherin or aminopeptidase in Bt toxicity have been achieved at the genetic, biochemical and molecular levels. Discovering the functions of these genes is critical in providing new approaches to modify Bt toxins to be more effective (166), revealing the possible receptor-associated Bt resistance, and serving as outstanding candidates for potential RNAi strategies of pest control (168).
3g) Symbionts
The beneficial associations between insects and symbiotic microbes are suggested to confer considerable fitness benefits to hosts (169). For example, the bacterial symbiont of aphids, Buchnera aphidicola, confers resistance to parasitoid attack by causing high mortality of parasitoid larvae (170). Another example is that the bacteria found in the oral secretions of Colorado potato beetle which have been shown to be responsible for the suppression of host plant defenses (171). Recently two studies have illustrated the role of symbionts in the development of insecticide resistance (37, 172). One discovery revealed that infection in the midgut with an

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
607 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
insecticide-degrading bacterial symbiont directly contributed to a rapid acquisition of fenitrothion (organophosphate) resistance in stinkbugs (37). Similarly, Patil and co-workers showed that mutualistic gut symbiotic bacteria in the mosquito A. stephensi can degrade Bt toxicity which facilitates the development of Bt resistance (172). Other studies have also suggested the link between the amounts of microbial symbionts and the level of host insecticide susceptibility that supports the potential roles these symbionts have in insecticide resistance (38, 173, 174). To further understand the role of these symbionts in insecticide resistance, RNAi approaches targeting the lethal genes of symbiotic microbes within the insect host would reveal their role in resistance (175). Furthermore, RNAi mediated manipulation of symbionts that cause insecticide resistance is an exciting alternative target for enhancing specific pest management (51, 62).
3h) Regulation of xenobiotic detoxification
In order to survive in harsh environment, insects have evolved complex and unique regulatory responses by multiple detoxification genes to cope with the continual threat of chemical stresses in their environments (46, 176). Deciphering the genetic machinery underpinning regulation of xenobiotic detoxification will facilitate the development of new pest control strategies (177). Studies in house fly have found that the up-regulation of metabolic detoxification genes in pyrethroid resistance is regulated by trans-acting factors and/or cis-acting elements (107, 110, 178, 179). Pyrethroid resistance in the LPR house fly strain is due to the overexpression of a single P450, CYP6D1 (178, 180-182). It has been proposed that two factors, one nearly completely dominant factor on autosome 1 and one incompletely recessive factor on autosome 2, contribute to the high level expression of CYP6D1 in the LPR house flies (183). Genetic studies revealed that pyrethroid resistance in house fly strain ALHF is linked to autosomes 2, 3, and 5, with a minor role from autosome 1 (184, 185). Additionally, multiply up regulated P450s CYP6A5, CYP6A5v2, CYP6A36, CYP4D4v2, and CYP6A38 are located on autosome 5 and the regulation of CYP6A36 was linked to autosomes 1 and 2 (118, 186-188). Recent transcriptomal linkage analysis suggests that the co-up-regulation of detoxification genes involved in pyrethroid resistance in the house fly may be regulated by G-protein-coupled receptors (GPCRs: rhodopsin and arrestin), protein kinase/phosphatases and proteases (107, 179). This hypothesis is supported by a current functional study on a Culex pipiens pallens GPCR arrestin (189). With evidence from RNAi experiments, overexpressed arrestin in the deltamethrin resistant strain was demonstrated to regulate the insecticide resistance-associated CYP gene (189).
Insect nuclear receptors also have important functional roles in the regulation of xenobiotic gene induction (190). In the genetic model, D. melanogaster, the ortholog of the human SXR and CAR xenobiotic receptors, DHR96, participates in the control

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
608 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
of phenobarbital-regulated genes (191). However, only 10% of the phenobarbital- regulated genes in the wild-type flies respond to phenobarbital through DHR96, suggesting the existence of extra trans-acting factors (46). Recent studies illustrate that the evolutionary conserved Cap’n’collar isoform-C (CncC)/Keap1 pathway is necessary and sufficient for xenobiotic-induced transcription as well as constitutive overexpression of a wide range of detoxification genes in insecticide resistant Drosophila strains (46, 177). The CncC isoforms of the Cnc locus were also identified from the genome of two-spotted spider mite (35) and the transcriptome of Colorado potato beetle (Zhu et al., unpublished data). Further studies on the regulation of CncC/Keap1 pathway in these arthropod species should reveal whether CncC plays a general role in the xenobiotic responses within agricultural pests. Understanding biochemical detoxification pathway functions and their roles in insecticide resistance have also recently been shown to be influenced by circadian rhythms (41, 42, 192, 193) (Fig. 1b). If insecticide resistance genes and proteins are fluctuating according to a circadian clock, this has important implications for pest control. While this is a relatively new area, it is important to note that circadian rhythms should be taken into account when identifying and testing candidate targets for RNAi mediated pest control strategies.
4. Conclusion and future directions
Insecticide resistance is the most significant challenge facing sustainable IPM efforts around the world. Insects have been adapting to their environments for over 450 million years which prepare them could evolve resistance to every chemicals that human beings have discovered over the past 60 years, no matter how unique the mode of action of the insecticide is (72, 194). On one hand, RNAi technology has served as a robust functional genomics tool for investigating molecular mechanisms underlying insect adaptation to a diversity of insecticides. As a new insecticidal tactic, RNAi mediated pest control has the distinct advantage of being designed to work specifically with existing pest management approaches. For example, the feasibility of incorporating carefully designed dsRNA targets (including focal genes resulting in insecticide resistance and lethal or sublethal genes) by RNAi would facilitate multi-dimensional pest management strategies through integrating the RNAi-based insecticide with insecticidal compounds already in use and/or Bt crops in a complementary pattern to manipulate a single or multiple target pests (195,196). On the other hand, the resistance to RNAi itself is an issue of concern. Although recent study provided evidence that RNAi resistance due to mutations in the target sequences is nearly impossible (197), the resistance development through other routes such as single nucleoitide polymorphisms on genes coding for proteins related to the intricate RNAi pathway is not unlikely (198).

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
609 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
Indeed, RNAi as a pest management alternative is already ongoing. RNAi-based GM crops have been developed in the laboratory for three major crops: corn (67), cotton (68, 91), and rice (93). The field risk assessment, environmental fate of dsRNA evaluation, and RNAi effect variation trials are on the way or have been conducted (32, 198, 200). Compared with RNAi-based GM crops, using bacteria to synthesize and deliver dsRNA also holds promise to manage agricultural pests especially in non-major crops, such as vegetables and fruits (201) or urban pests as insecticidal baits (65, 89). Due to the high efficiency and low cost, feeding RNAi through bacteria synthesis dsRNA could become not only a tool for genome-wide RNAi screening for insecticide targets in the laboratory but also a promising IPM alternative for pest control under greenhouse or field conditions (Fig. 4). Unlike discovering a new synthetic insecticide or developing an additional GM crop, it is technologically flexible and relatively effortless, allowing for rapid adjustment to the components of the dsRNA cocktail as the efficiency declines or resistance develops. In addition, delivery methods of bacterially synthesized dsRNA could also utilize existing chemical spray equipment and would not require additional specialized equipment.
Fig. 4. Prospective procedure of RNAi application in laboratory (Stage I), semi-field and field (Stage II).

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
610 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
Although the RNAi approach is a sequence-specific gene knockdown procedure and as such was predicted to have little to no non-target effects, the question remains as to how risky this might be on a large, agricultural scale. Many studies have shown that unintentional off-target gene silencing in target cells and gene silencing in non-target organisms occur more commonly than expected (67, 202-204). Moreover, the physiological basis of RNAi and the genomics and physiologies of potentially exposed non-target organisms remain largely unknown. To fill these huge knowledge gaps, future research should focus on developing more desirable sequence and species specific gene silencing to enhance our ability to assess the activity spectrum of RNAi across all animals that are likely to be impacted (32, 205). Finally, understanding the environmental risks of RNAi crops impact on food webs in agroecosystems and continuously seeking candidate gene targets are critical focuses for RNAi mediated pest control research.
5. Acknowledgements
The authors are grateful to Dr. Raman Chandrasekar (Kansas State University, USA) for encouraging us to write this review. Our special thanks to Mr. Timothy W. Moural (Washington State University, USA) for editing the manuscript. We also appreciate all valuable suggestions and critical comments from three reviewers.
6. References
1. Robinson, G.E., Hackett, K.J., Purcell-Miramontes, M., Brown, S.J., Evans, J.D., et al. (2011) Creating a buzz aboutinsect genomes. Science 331: 1386.
2. Enserink, M., Hines, P.J., Vignieri, S.N., Wigginton, N.S., Yeston, J.S. (2013) Smarter pest control. The pesticideparadox. Introduction. Science 341: 728-729.
3. Alyokhin, A., Baker, M., Mota-Sanchez, D., Dively, G., Grafius, E. (2008) Colorado potato beetle resistance toinsecticides. Am. J. Potato Res. 85: 395-413.
4. Zhu, F., Gujar, H., Gordon, J.R., Haynes, K.F., Potter, M.F., et al. (2013) Bed bugs evolved unique adaptive strategyto resist pyrethroid insecticides. Sci. Rep. 3: 1456.
5. You, M., Yue, Z., He, W., Yang, X., Yang, G. et al. (2013) A heterozygous moth genome provides insights intoherbivory and detoxification. Nat. Genet. 45: 220-225.
6. Hawkins, R.D., Hon, G.C., Ren, B. (2010) Next-generation genomics: an integrative approach. Nat. Rev. Genet. 11: 476-486.
7. Metzker, M.L. (2010) Sequencing technologies—the next generation. Nat. Rev. Genet. 11: 31-46. 8. Zhou, X, Oi, F.M., Scharf, M.E. (2006) Social exploitation of hexamerin: RNAi reveals a major caste-regulatory factor
in termites. Proc. Natl. Acad. Sci. USA 103: 4499-4504. 9. Nelson, C.M., Ihle, K.E., Fondrk, M.K., Page, R.E., Amdam, G.V. (2007) The gene vitellogenin has multiple
coordinating effects on social organization. PLoS Biol. 5: 673-677. 10. Kamakura, M. (2011) Royalactin induces queen differentiation in honeybees. Nature 473: 478-483. 11. Emlen, D.J., Warren, I.A., Johns, A., Dworkin, I., Lavine, L.C. (2012) A mechanism of extreme growth and reliable
signaling in sexually selected ornaments and weapons. Science 337: 860-864.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
611 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
12. Howlett, N., Dauber, K.L., Shukla, A., Morton, B., Glendinning, J.I., et al. (2012) Identification of chemosensoryreceptor genes in Manduca sexta and knockdown by RNA interference. BMC Genomics 13: 211.
13. Li-Byarlay, H., Li Y., Stroud, H., Feng, S., Newman, T.C., et al. (2013) RNA interference knockdown of DNAmethyl-transferase 3 affects gene alternative splicing in the honey bee. Proc. Natl. Acad. Sci. USA 110: 12750-12755.
14. Zhu, F., Xu, P., Barbosa, R.M.R., Choo, Y.M., Leal, W.S. (2013) RNAi-based demonstration of direct link betweenspecific odorant receptors and mosquito oviposition behavior. Insect Biochem. Mol. Biol. 43: 916-923.
15. Zhu, Q., Arakane, Y., Beeman, R.W., Kramer, K.J., Muthukrishnan, S. (2008) Functional specialization among insect chitinase family genes revealed by RNA interference. Proc. Natl. Acad. Sci. USA 105: 6650-6655.
16. Ma, Z., Guo, W., Guo, X., Wang, X., Kang, L. (2011) Modulation of behavioral phase changes of the migratory locust by the catecholamine metabolic pathway. Proc. Natl. Acad. Sci. USA 108: 3882-3887.
17. Wu, R., Wu, Z., Wang, X., Yang, P., Yu, D., et al. (2012) Metabolomic analysis reveals that carnitines are keyregulatory metabolites in phase transition of the locusts. Proc. Natl. Acad. Sci. USA 109: 3259-3263.
18. Sheng, Z., Xu, J., Bai, H., Zhu, F., Palli, S.R. (2011) Juvenile hormone regulates vitellogenin gene expression through insulin-like peptide sgnaling pathway in the red flour beetle, Tribolium castaneum. J. Biol. Chem. 286: 41924-419236.
19. Pauchet, Y., Wilkinson, P., Vogel, H., Nelson, D.R., Reynolds, S.E., et al. (2010) Pyrosequencing the Manduca sextalarval midgut transcriptome: messages for digestion, detoxification and defence. Insect Mol. Biol. 19: 61-75.
20. Lopez, M.F., Cano-Ramirez, C., Cesar-Ayala, A.K., Ruiz, E.A., Zuniga, G. (2013) Diversity and expression of P450genes from Dendroctonus valens LeConte (Curculionidae: Scolytinae) in response to different kairomones. InsectBiochem. Mol. Biol. 43: 417-432.
21. Cano-Ramírez, C., López, M.F., Cesar-Ayala, A.K., Pineda-Martínez, V., Sullivan, B.T., et al. (2013) Isolation andexpression of cytochrome P450 genes in the antenna and gut of pine beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) following exposure to host monoterpenes. Gene 520: 47-63.
22. Tao, X., Xue, X., Huang, Y., Chen, X., Mao, Y. (2012) Gossypol-enhanced P450 gene pool contributes to cottonbollworm tolerance to a pyrethroid insecticide. Mol. Ecol. 21: 4371-485.
23. Claudianos, C., Ranson, H., Johnson, R.M., Biswas, S., Schuler, M.A., et al. (2006) A deficit of detoxificationenzymes: pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 15: 615-636.
24. Li, X., Schuler, M.A., Berenbaum, M.R. (2007) Molecular mechanisms of metabolic resistance to synthetic andnatural xenobiotics. Annu. Rev. Entomol. 52: 231-253.
25. Despres, L., David, J.P., Gallet, C. (2007) The evolutionary ecology of insect resistance to plant chemicals. TrendsEcol. Evol. 22: 298-307.
26. Paris, M., Melodelimaa, C., Coissac, E., Tetreau, G., Reynaud, S., et al. (2012) Transcription profiling of resistance toBt toxins in the mosquito Aedes aegypti using next-generation sequencing. J. Invertebr. Pathol. 109: 201-208.
27. Tabashnik, B.E., Brevault, T., Carriere, Y. (2013) Insect resistance to Bt crops: lessons from the first billion acres. Nat. Biotechnol. 31: 510-521.
28. Adelman, Z.N., Kilcullen, K.A., Koganemaru, R., Anderson, M.A.E., Anderson, T.D., et al. (2011) Deep sequencingof pyrethroid-resistant bed bugs reveals multiple mechanisms of resistance within a single population. PLoS One 6:e26228.
29. Mamidala, P., Wijeratne, A.J., Wijeratne, S., Kornacker, K., Sudhamalla, B., et al. (2012) RNA-Seq and moleculardocking reveal multi-level pesticide resistance in the bed bug. BMC Genomics 13: 6.
30. Pavlidi, N., Dermauw, W., Rombauts, S., Chrisargiris, A., Van Leeuwen, T., Vontas, J. (2013) Analysis of the olivefruit fly Bactrocera oleae transcriptome and phylogenetic classification of the major detoxification gene families.PLoS One 8: e66533.
31. Price, D.R., Gatehouse, J.A. (2008) RNAi-mediated crop protection against insects. Trends Biotechnol. 26: 393-400. 32. Lundgren, J.G., Duan, J.J. (2013) RNAi-based insecticidal crops: potential effects on nontarget species. Bioscience 63:
657-665. 33. Gordon, H.T. (1961) Nutritional factors in insect resistance to chemicals. Annu. Rev. Entomol. 6: 27-254. 34. Kim, C.H., Muturi, E.J. (2012) Relationship between leaf litter identity, expression of cytochrome P450 genes and life
history traits of Aedes aegypti and Aedes albopictus. Acta Tropica. 122: 94-100.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
612 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
35. Dermauw, W., Wybouw, N., Rombauts, S., Menten, B., Vontas, J., et al. (2013) A link between host plant adaptation and pesticide resistance in the polyphagous spider mite Tetranychus urticae. Proc. Natl. Acad. Sci. USA 110:E113-E22
36. Ghanim, M., Kontsedalov, S. (2009) Susceptibility to insecticides in the Q biotype of Bemisia tabaci is correlated withbacterial symbiont densities. Pest Manag. Sci. 65: 939-942.
37. Kikuchi, Y., Hayatsu, M., Hosokawa, T., Nagayama, A., Tago, K., et al. (2012) Symbiont-mediated insecticideresistance. Proc. Natl. Acad. Sci. USA 109: 8618-8622.
38. Pan, H., Chu, D., Liu, B., Xie, W., Wang, S., et al. (2013). Relative amount of symbionts in insect hosts changes withhost-plant adaptation and insecticide resistance. Environ. Entomol. 42: 74-78.
39. David, J.P., Coissac, E., Melodelima, C., Poupardin, R., Riaz, M.A., et al. (2010) Transcriptome response to pollutants and insecticides in the dengue vector Aedes aegypti using next-generation sequencing technology. BMC Genomics 11:216.
40. David, J.P., Ismail, H.M., Chandor-Proust, A., Paine, M.J. (2013) Role of cytochrome P450s in insecticide resistance: impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos. Trans. R. Soc. Lond. B Biol. Sci. 368: 20120429.
41. Hooven, L.A., Sherman, K.A., Butcher, S., Giebultowicz, J.M. (2009) Does the clock make the poison? Circadianvariation in response to pesticides. PloS One 4: e6469.
42. Beaver, L.M., Hooven, L.A., Butcher, S.M., Krishnan, N., Sherman, K.A., et al. (2010) Circadian clock regulatesresponse to pesticides in Drosophila via conserved Pdp1 pathway. Toxicol. Sci. 115: 513-520.
43. Daborn, P.J., Yen, J.L., Bogwitz, M.R., Le Goff, G., Feil, E., et al. (2002) A single p450 allele associated withinsecticide resistance in Drosophila. Science 297: 2253-2256.
44. David, J.P., Strode, C., Vontas, J., Nikou, D., Vaughan, A., et al. (2005) The Anopheles gambiae detoxification chip: a highly specific microarray to study metabolic-based insecticide resistance in malaria vectors. Proc. Natl. Acad. Sci.USA 102: 4080-4084.
45. Zhu, F., Parthasarathy, R., Bai, H., Woithe, K., Kaussmann, M., et al. (2010) A brain-specific cytochrome P450responsible for the majority of deltamethrin resistance in the QTC279 strain of Tribolium castaneum. Proc. Natl.Acad. Sci. USA 107: 8557-8562.
46. Misra, J.R., Horner, M.A., Lam, G., Thummel, C.S. (2011) Transcriptional regulation of xenobiotic detoxification inDrosophila. Genes Dev. 25: 1796-1806.
47. Fire, A., Xu, S., Montgomery, M.K., Kostas, S.K., Driver, S.E., et al. (1998) Potent and specific genetic interferenceby double-stranded RNA in Caenorhabditis elegans. Nature 391: 806-811.
48. Whangbo, J.S., Hunter, C.P. (2008) Environmental RNA interference. Trends Genet. 24: 297-305. 49. Siomi, H., Siomi, M.C. (2009) On the road to reading the RNA-interference code. Nature 457: 396-404. 50. Tomoyasu, Y., Miller, S.C., Tomita, S., Schoppmeier, M., Grossmann, D., et al. (2008) Exploring systemic RNA
interference in insects: a genome-wide survey for RNAi genes in Tribolium. Genome Biol. 9: R10. 51. Aronstein, K., Oppert, B., Lorenzen, M.D. (2011) RNAi in agriculturally-important arthropods. In RNA processing,
ed. P Grabowski, InTech-Open Access, pp. 157-80. 52. Swevers, L., Huvenne, H., Menschaert, G., Kontogiannatos, D., Kourti, A., et al. (2013) Colorado potato beetle
(Coleoptera) gut transcriptome analysis: expression of RNA interference-related genes. Insect Mol. Biol. 22: 668-684. 53. Winston, W.M., Molodowitch, C., Hunter, C.P. (2002) Systemic RNAi in C. elegans requires the putative
transmembrane protein SID-1. Science 295: 2456-2459. 54. Winston, W.M., Sutherlin, M., Wright, A.J., Feinberg, E.H., Hunter, C.P. (2007) Caenorhabditis elegans SID-2 is
required for environmental RNA interference. Proc. Natl. Acad. Sci. USA 104: 10565-10570. 55. McEwan, D.L., Weisman, A.S., Hunter, C.P. (2012) Uptake of extracellular double-stranded RNA by SID-2. Mol. Cell
47: 746-754. 56. Saleh, M.C., van Rij, R.P., Hekele, A., Gillis, A., Foley, E., et al. (2006) The endocytic pathway mediates cell entry of
dsRNA to induce RNAi silencing. Nat. Cell Biol. 8: 793-802. 57. Ulvila, J., Parikka, M., Kleino, A., Sormunen, R., Ezekowitz, R.A., et al. (2006) Double-stranded RNA is internalized
by scavenger receptor-mediated endocytosis in Drosophila S2 cells. J. Biol. Chem. 281: 14370-14375. 58. Gordon, K.H., Waterhouse P.M. (2007) RNAi for insect-proof plants. Nat Biotechnol 25: 1231-1232.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
613 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
59. Aronstein, K., Pankew, T., Saldivar, E. (2006) SID-1 is implicated in systemic gene silencing in the honey bee. J.Apicult. Res. 45: 20-24.
60. Sijen, T., Fleenor, J., Simmer, F., Thijssen, K.L., Parrish, S., et al. (2001) On the role of RNA amplification indsRNA-triggered gene silencing. Cell 107: 465-476.
61. Hamilton, A., Voinnet, O., Chappell, L., Baulcombe, D. (2002) Two classes of short interfering RNA in RNAsilencing. EMBO J. 21: 4671-4679.
62. Barnard, A., Nijhof, A.M., Fick, W., Stutzer, C., Maritz-Olivier, C. (2012) RNAi in arthropods: insight into themachinery and applications for understanding the pathogen-vector interface. Genes 3: 702-741.
63. Tan, A., Palli, SR. (2008) Identifiation and characterization of nuclear receptors from the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 38: 430-439.
64. Martin, D., Maestro, O., Cruz, J., Mane-Padros, D., Belles, X. (2006) RNAi studies reveal a conserved role for RXR inmolting in the cockroach Blattella germanica. J. Insect Physiol. 52: 410-416.
65. Zhou, X., Wheeler, M.M., Oi, F.M., Scharf, M.E. (2008) RNA interference in the termite Reticulitermes flavipes through ingestion of double-stranded RNA. Insect Biochem. Mol. Biol. 38: 805-815.
66. Zhu, F., Xu, J., Palli, R., Ferguson, J., Palli, S.R. (2011) Ingested RNA interference for managing the populations ofthe Colorado potato beetle, Leptinotarsa decemlineata. Pest Manag. Sci. 67: 175-182.
67. Baum, J.A., Bogaert, T., Clinton, W., Heck, G.R., Feldmann, P., et al. (2007) Control of coleopteran insect peststhrough RNA interference. Nat. Biotechnol. 25: 1322-1326.
68. Mao, Y., Cai, W., Wang, J., Hong, G., Tao, X., et al. (2007) Silencing a cotton bollworm P450 monooxygenase geneby plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 25: 1307-1313.
69. Zhang, X., Zhang, J., Zhu, K.Y. (2010) Chitosan/double-stranded RNA nanoparticle-mediated RNA interference tosilence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambiae). Insect Mol. Biol. 19: 683-693.
70. Terenius, O., Papanicolaou, A., Garbutt, J.S., Eleftherianos, I., Huvenne, H., et al. (2011) RNA interference inLepidoptera: an overview of successful and unsuccessful studies and implications for experimental design. J. InsectPhysiol. 57: 231-245.
71. Pridgeon, J.W., Zhao, L., Becnel, J.J., Strickman, D.A., Clark, G.G., et al. (2008) Topically applied AaeIAP1double-stranded RNA kills female adults of Aedes aegypti. J. Med. Entomol. 45: 414-420.
72. Scott, J.G., Michel, K., Bartholomay, L.C., Siegfried, B.D., Hunter, W.B., et al. (2013) Towards the elements ofsuccessful insect RNAi. J. Insect Physiol. 59: 1212-1221.
73. Yu, N., Christiaens, O., Liu, J., Niu, J., Cappelle, K., et al. (2013) Delivery of dsRNA for RNAi in insects: anoverview and future directions. Insect Sci. 20: 4-14.
74. Zhang, H., Li, H., Miao, X. (2013) Feasibility, limitation and possible solutions of RNAi-based technology for insectpest control. Insect Sci. 20: 15-30.
75. Brown, S.J., Mahaffey, J.P., Lorenzen, M.D., Denell, R.E., Mahaffey, J.W. (1999) Using RNAi to investigateorthologous homeotic gene function during development of distantly related insects. Evol. Dev. 1: 11-15.
76. Mutti, N.S., Park, Y., Reese, J.C., Reeck, G.R. (2006) RNAi knockdown of a salivary transcript leading to lethality inthe pea aphid, Acyrthosiphon pisum. J. Insect Sci. 6: 1-7.
77. Zhu, F., Sams, S., Moural, T., Haynes, K.F., Potter, M.F., et al. (2012) RNA interference of NADPH-cytochrome P450 reductase results in reduced insecticide resistance in the bed bug, Cimex lectularius. PLoS One 7: e31037.
78. Belles, X. (2010) Beyond Drosophila: RNAi in vivo and functional genomics in insects. Annu. Rev. Entomol. 55: 111-128.
79. Ghanim, M., Kliot, A. (2013) RNA interference and its potential for developing new control methods against insectpests. In Advanced Technologies for Managing Insect Pests, ed. I Ishaaya, SR Palli, AR Horowitz, Springer, NewYork, pp.179-193.
80. Araujo, R.N., Santos, A., Pinto, F.S., Gontijo, N.F., Lehane, M.J., et al. (2006) RNA interference of the salivary gland nitrophorin 2 in the triatomine bug Rhodnius prolixus (Hemiptera: Reduviidae) by dsRNA ingestion or injection.Insect Biochem. Mol. Biol. 36: 683-693.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
614 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
81. Turner, C.T., Davy, M.W., MacDiarmid, R.M., Plummer, K.M., Birch, N.P., et al. (2006) RNA interference in thelight brown apple moth, Epiphyas postvittana (Walker) induced by double-stranded RNA feeding. Insect Mol. Biol. 15: 383-391.
82. Huvenne, H., Smagghe, G. (2010) Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: a review. J. Insect Physiol. 56: 227-235.
83. Upadhyay, S.K., Chandrashekar, K., Thakur, N., Verma, P.C., Borgio, J.F., et al. (2011) RNA interference for thecontrol of whiteflies (Bemisia tabaci) by oral route. J. Biosci. 36: 153-161.
84. Kamath, R.S., Ahringer, J. (2003) Genome-wide RNAi screening in Caenorhabditis elegans. Methods 30: 313-321. 85. Timmons, L., Fire, A. (1998) Specific interference by ingested dsRNA. Nature 395: 854. 86. Timmons, L., Court, D.L., Fire, A. (2001). Ingestion of bacterially expressed dsRNAs can produce specific and potent
genetic interference in Caenorhabditis elegans. Gene 263: 103-112. 87. Tian, H., Peng, H., Yao, Q., Chen, H., Xie, Q., et al. (2009) Developmental control of a lepidopteran pest Spodoptera
exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PLoS One 4: e6225. 88. Li, X., Zhang, M., Zhang, H. (2011) RNA interference of four genes in adult Bactrocera dorsalis by feeding their
dsRNAs. PLoS One 6: e17788. 89. Ratzka, C., Gross, R., Feldhaar, H. (2013) Systemic gene knockdown in Camponotus floridanus workers by feeding of
dsRNA. Insectes Soci. 60: 475-484. 90. Zhou, L., Jia, S., Wan, P., Kong, Y., Guo, W., et al. (2013). RNA interference of a putative
S-adenosyl-L-homocysteine hydrolase gene affects larval performance in Leptinotarsa decemlineata (Say). J. InsectPhysiol. 59: 1049-1056.
91. Mao, Y., Tao, X., Xue, X., Wang, L., Chen X. (2011) Cotton plants expressing CYP6AE14 double-stranded RNAshow enhanced resistance to bollworms. Transgenic Res. 20: 665-673.
92. Pitino, M., Coleman, A.D., Maffei, M.E., Ridout, C.J., Hogenhout, S.A. (2011) Silencing of aphid genes by dsRNAfeeding from plants. PLoS One 6: e25709.
93. Zha, W., Peng, X., Chen, R., Du, B., Zhu, L., He, G. (2011) Knockdown of midgut genes by dsRNA-transgenicplant-mediated RNA interference in the hemipteran insect Nilaparvata lugens. PLoS One 6: e20504.
94. Burch-Smith, T.M., Anderson, J.C., Martin, G.B., Dinesh-Kumar, S.P. (2004) Applications and advantages ofvirus-induced gene silencing for gene function studies in plants. Plant J. 39: 734-746.
95. Kumar, P., Pandit, S.S., Baldwin, I.T. (2012) Tobacco rattle virus vector: A rapid and transient means of silencingmanduca sexta genes by plant mediated RNA interference. PLoS One 7: e31347.
96. Swevers, L., Vanden Broeck, J., Smagghe, G. (2013) The possible impact of persistent virus infection on the functionof the RNAi machinery in insects: a hypothesis. Front. Physiol. 4: 319.
97. Swevers, L., Smagghe, G. (2012) Use of RNAi for control of insect crop pests. In Arthropod-Plant Interactions: Novel Insights and Approaches for IPM, ed. G Smagghe, I Diaz, Springer, Netherlands, pp. 177-97.
98. Liu, N., Zhu, F., Xu, Q., Pridgeon, Y., Gao, X.W. (2006) Behavioral change, physiological modification, andmetabolic detoxification: mechanisms of insecticide resistance. Acta Entomologica Sinica 48: 672-680.
99. Nkya, T.E., Akhouayri, I., Kisinza, W., David, J.P. (2013) Impact of environment on mosquito response to pyrethroid insecticides: facts, evidences and prospects. Insect Biochem. Mol. Biol. 43: 407-416.
100. Wada-Katsumata, A., Silverman, J., Schal, C. (2013) Changes in taste neurons support the emergence of an adaptive behavior in cockroaches. Science 340: 972-975.
101. Strycharz, J.P., Lao, A., Li, H.M., Qiu, X.H., Lee, S.H., et al. 2013. Resistance in the highly DDT-resistant 91-R strain of Drosophila melanogaster involves decreased penetration, increased metabolism, and direct excretion. Pestic. Biochem. Phys. 107: 207-217.
102. Hemingway, J., Hawkes, N.J., McCarroll, L., Ranson, H. (2004) The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 34: 653-665.
103. Dong, K., Du, Y., Rinkevich, F., Nomura, Y., Xu, P., et al. (2014) Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 50: 1-17.
104. Scott, J.G., Georghiou, G.P. (1986) Mechanisms responsible for high-levels of permethrin resistance in the housefly. Pestic. Sci. 17: 195-206.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
615 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
105. Liu, N., Liu, H., Zhu, F., Zhang, L. (2007) Differential expression of genes in pyrethroid resistant and susceptible mosquitoes, Culex quinquefasciatus (S.). Gene 394: 61-68.
106. Awolola, T.S., Oduola, O.A., Strode, C., Koekemoer, L.L., Brooke, B., et al. (2009) Evidence of multiple pyrethroid resistance mechanisms in the malaria vector Anopheles gambiae sensu stricto from Nigeria. Trans. R. Soc. Trop. Med. Hyg. 103: 1139-1145.
107. Liu, N. (2011) Pyrethroid resistance in insects: genes, mechanisms, and regulation. In Insecticides- Advances in Integrated Pest Management, ed. F Perveen, InTech-Open Access, pp. 457-68.
108. Bariami, V., Jones, C.M., Poupardin, R., Vontas, J., Ranson, H. (2012) Gene amplification, ABC transporters and cytochrome P450s: unraveling the molecular basis of pyrethroid resistance in the dengue vector, Aedes aegypti. PLoS Negl. Trop. Dis. 6: e1692.
109. Jones, C.M., Toe, H.K., Sanou, A., Namountougou, M., Hughes, A., et al. (2012) Additional selection for insecticide resistance in urban malaria vectors: DDT resistance in Anopheles arabiensis from Bobo-Dioulasso, Burkina Faso. PLoS One 7: e45995.
110. Liu, N., Scott, J.G. (1998) Increased transcription of CYP6D1 causes cytochrome P450-mediated insecticide resistance in house fly. Insect Biochem. Mol. Biol. 28: 531-535.
111. Joussen, N., Agnolet, S., Lorenz, S., Schone, S.E., Ellinger, R., et al. (2012) Resistance of Australian Helicoverpa armigera to fenvalerate is due to the chimeric P450 enzyme CYP337B3. Proc. Natl. Acad. Sci. USA 109: 15206-15211.
112. Nelson, D.R. (2013) A world of cytochrome P450s. Philos. Trans. R. Soc. Lond. B Biol. Sci. 368: 20120430. 113. Zhu, F., Moural, T.W., Shah, K., Palli, S.R. (2013) Integrated analysis of cytochrome P450 gene superfamily in the red
flour beetle, Tribolium castaneum. BMC Genomics 14: 174. 114. Yang, T., Liu, N. (2011) Genome analysis of cytochrome P450s and their expression profiles in insecticide resistant
mosquitoes, Culex quinquefasciatus. PloS One 6: e29418. 115. Yan, L., Yang, P., Jiang, F., Cui, N., Ma, E., et al. (2012) Transcriptomic and phylogenetic analysis of Culex pipiens
quinquefasciatus for three detoxification gene families. BMC Genomics 13: 609. 116. Feyereisen, R. (2012) Insect CYP genes and P450 enzymes. In Insect molecular biology and biochemistry, ed. LI
Gilbert, Elsevier Academic Press, pp. 236-316. 117. Schuler, M.A. (1996) The role of cytochrome P450 monooxygenases in plant-insect interactions. Plant Physiol. 112:
1411-1419. 118. Zhu, F., Li, T, Zhang, L., Liu, N. (2008) Co-up-regulation of three P450 genes in response to permethrin exposure in
permethrin resistant house flies, Musca domestica. BMC Physiol. 8: 18. 119. Liu, N., Li, T., Reid, W.R., Yang, T., Zhang, L. (2011) Multiple cytochrome P450 genes: their constitutive
overexpression and permethrin induction in insecticide resistant mosquitoes, Culex quinquefasciatus. PLoS One 6: e23403.
120. Bautista, M.A., Miyata, T., Miura, K., Tanaka, T. (2009) RNA interference-mediated knockdown of a cytochrome P450, CYP6BG1, from the diamondback moth, Plutella xylostella, reduces larval resistance to permethrin. Insect Biochem. Mol. Biol. 39: 38-46.
121. Guo, Y., Zhang, J., Yu, R., Zhu, K., Guo, Y., et al. (2012) Identification of two new cytochrome P450 genes and RNA interference to evaluate their roles in detoxification of commonly used insecticides in Locusta migratoria. Chemosphere 87: 709-717.
122. Lycett, G.J., McLaughlin, L.A., Ranson, H., Hemingway, J., Kafatos, F.C., et al. (2006) Anopheles gambiae P450 reductase is highly expressed in oenocytes and in vivo knockdown increases permethrin susceptibility. Insect Mol. Biol. 15: 321-327.
123. Tang, T., Zhao, C., Feng, X., Liu, X., Qiu, L. (2012) Knockdown of several components of cytochrome P450 enzyme systems by RNA interference enhances the susceptibility of Helicoverpa armigera to fenvalerate. Pest Manag. Sci. 68: 1501-1511.
124. Scott, J.G. (1991). Insecticide resistance in insects. In Hanbook of Pest Management, ed. D Pimental, CRC Press, Boca Raton, pp. 663-77.
125. Ranson, H., Claudianos, C., Ortelli, F., Abgrall, C., Hemingway, J., et al. (2002) Evolution of supergene families associated with insecticide resistance. Science 298: 179-181.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
616 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
126. Zhang, J., Zhang, J., Yang, M., Jia, Q., Guo, Y., et al. (2011) Genomics-based approaches to screening carboxylesterase-like genes potentially involved in malathion resistance in oriental migratory locust (Locusta migratoria manilensis). Pest Manag. Sci. 67: 183-190.
127. Zhang, J., Li, D., Ge, P., Yang, M., Guo, Y., et al. (2013) RNA interference revealed the roles of two carboxylesterase genes in insecticide detoxification in Locusta migratoria. Chemosphere 93: 1207-1215.
128. Xu, L., Duan, X., Lv, Y., Zhang, X., Nie, Z., et al. (2013) Silencing of an aphid carboxylesterase gene by use of plant-mediated RNAi impairs Sitobion avenae tolerance of Phoxim insecticides. Transgenic Res. DOI 10.1007/s11248-013-9765-9.
129. Enayati, A.A., Ranson, H., Hemingway, J. (2005) Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 14: 3-8.
130. Rauch, N., Nauen, R. (2004) Characterization and molecular cloning of a glutathione S-transferase from the whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Biochem. Mol. Biol. 34: 321-329.
131. Lumjuan, N., Rajatileka, S., Changsom, D., Wicheer, J., Leelapat, P., et al. (2011) The role of the Aedes aegypti Epsilon glutathione transferases in conferring resistance to DDT and pyrethroid insecticides. Insect Biochem. Mol. Biol. 41: 203-209.
132. Qin, G., Jia, M., Liu, T., Zhang, X., Guo, Y., et al. (2012) Heterologous expression and characterization of a sigma glutathione S-transferase involved in carbaryl detoxification from oriental migratory locust, Locusta migratoria manilensis (Meyen). J. Insect Physiol. 58: 220-227.
133. Zhou, W., Liang, Q., Xu, Y., Gurr, G., Bao, Y., et al. (2013) Genomic insights into the glutathione S-transferase gene family of two rice planthoppers, Nilaparvata lugens (Stål) and Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). PLoS One 8: e56604.
134. Qin, G., Jia, M., Liu, T., Zhang, X., Guo, Y., et al. (2013) Characterization and functional analysis of four glutathione S-transferases from the migratory locust, Locusta migratoria. PLoS One 8: e58410.
135. Wood, O.R., Hanrahan, S., Coetzee, M., Koekmoer, L.L., Brooke, B.D. (2010) Cuticle thickening assoicated with pyrethroid resistance in the major malaria vector Anopheles funestus. Parasites & Vectors 3: 1-7.
136. Zhang, J., Goyer, C., Pelletier, Y. (2008) Environmental stresses induce the expression of putative glycine-rich insect cuticular protein genes in adult Leptinotarsa decemlineata (Say). Insect Mol. Biol. 17: 209-216.
137. Bonizzoni, M., Afrane, Y., Dunn, WA., Atieli, F.K., Zhou, G., et al. (2012) Comparative transcriptome analyses of deltamethrin-resistant and -susceptible Anopheles gambiae mosquitoes from Kenya by RNA-Seq. PLoS One 7: e44607.
138. Riaz, M.A., Chandor-Proust, A., Dauphin-Villemant, C., Poupardin, R., Jones, C.M., et al. (2013) Molecular mechanisms associated with increased tolerance to the neonicotinoid insecticide imidacloprid in the dengue vector Aedes aegypti. Aquat. Toxicol. 126: 326-337.
139. Zhang, J., Zhu, K.Y. (2006) Characterization of a chitin synthase cDNA and its increased mRNA level associated with decreased chitin synthesis in Anopheles quadrimaculatus exposed to diflubenzuron. Insect Biochem. Mol. Biol. 36: 712-725.
140. Rees, D.C., Johnson, E., Lewinson, O. (2009) ABC transporters: the power to change. Nat. Rev. Mol. Cell Biol. 10: 218-227.
141. Labbe, R., Caveney, S., Donly, C. (2011) Genetic analysis of the xenobiotic resistance-associated ABC gene subfamilies of the Lepidoptera. Insect Mol. Biol. 20: 243-256.
142. Gahan, L.J., Pauchet, Y., Vogel, H., Heckel, D.G. (2010) An ABC transporter mutation is correlated with insect resistance to Bacillus thuringiensis Cry1Ac toxin. PLoS Genet. 6(12): e1001248.
143. Dermauw, W., Van Leeuwen, T. (2013) The ABC gene family in arthropods: comparative genomics and role in insecticide transport and resistance. Insect Biochem. Mol. Biol. 45:89-110.
144. Luo, L., Sun, Y., Yang, L., Huang, S., Wu, Y. (2013) Avermectin induces P-glycoprotein expression in S2 cells via the calcium/calmodulin/NF-kappaB pathway. Chem. Biol. Interact. 203: 430-439.
145. Luo, L., Sun, Y., Wu, Y. (2013) Abamectin resistance in Drosophila is related to increased expression of P-glycoprotein via the dEGFR and dAkt pathways. Insect Biochem. Mol. Biol. 43: 627-634.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
617 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
146. Pedra, J.H., McIntyre, L.M., Scharf, M.E., Pittendrigh, B.R. (2004) Genome-wide transcription profile of field- and laboratory-selected dichlorodiphenyltrichloroethane (DDT)-resistant Drosophila. Proc. Natl. Acad. Sci. USA 101: 7034-7039.
147. Bao, Y., Li, B., Liu, Z., Xue, J., Zhu, Z., et al. (2010) Triazophos up-regulated gene expression in the female brown planthopper, Nilaparvata lugens. J. Insect Physiol. 56: 1087-1094.
148. Silva, A.X., Jander, G., Samaniego, H., Ramsey, J.S., Figueroa, C.C. (2012) Insecticide resistance mechanisms in the green peach aphid Myzus persicae (Hemiptera: Aphididae) I: A transcriptomic survey. PloS One 7: e36366.
149. Yang, N., Xie, W., Jones, C.M., Bass, C., Jiao X., et al. (2013) Transcriptome profiling of the whitefly Bemisia tabaci reveals stage-specific gene expression signatures for thiamethoxam resistance. Insect Mol. Biol. 22: 485-496.
150. Yoon, K.S., Strycharz, J.P., Baek, J.H., Sun, W., Kim, J.H., et al. (2011) Brief exposures of human body lice to sublethal amounts of ivermectin over-transcribes detoxification genes involved in tolerance. Insect. Mol. Biol. 20: 687-699.
151. Figueira-Mansur, J., Ferreira-Pereira, A., Mansur, J.F., Franco, T.A., Alvarenga, E.S., et al. (2013) Silencing of P-glycoprotein increases mortality in temephos-treated Aedes aegypti larvae. Insect. Mol. Biol. 22:648-658.
152. Aurade, R.M., Jayalakshmi, S.K., Sreeramulu, K. (2010) Modulatory effects of natural curcuminoids on P-glycoprotein ATPase of insecticide-resistant pest Helicoverpa armigera (Lepidopetera: Noctuidae). J. Membr. Biol. 236: 271-278.
153. Aurade, R.M., Jayalakshmi, S.K., Sreeramulu, K. (2010) P-glycoprotein ATPase from the resistant pest, Helicoverpa armigera: Purification, characterization and effect of various insecticides on its transport function. Bba-Biomembranes 1798: 1135-1143.
154. Petschenka, G., Pick, C., Wagschal, V., Dobler, S. (2013) Functional evidence for physiological mechanisms to circumvent neurotoxicity of cardenolides in an adapted and a non-adapted hawk-moth species. Proc. Biol. Sci. 280: 20123089.
155. Robert, J.A., Pitt, C., Bonnett, T.R., Yuen, M.M., Keeling, C.I., et al. (2013) Disentangling detoxification: gene expression analysis of feeding mountain pine beetle illuminates molecular-level host chemical defense detoxification mechanisms. PLoS One 8: e77777.
156. Strauss, A.S., Peters, S., Boland, W., Burse, A. (2013) ABC transporter functions as a pacemaker for sequestration of plant glucosides in leaf beetles. eLife 2: e01096.
157. Heckel, D.G. (2012) Learning the ABCs of Bt: ABC transporters and insect resistance to Bacillus thuringiensis provide clues to a crucial step in toxin mode of action. Pestic. Biochem. Phys. 104: 103-110.
158. Tanaka, S., Miyamoto, K., Noda, H., Jurat-Fuentes, J.L., Yoshizawa, Y., et al. (2013) The ATP-binding cassette transporter subfamily C member 2 in Bombyx mori larvae is a functional receptor for Cry toxins from Bacillus thuringiensis. FEBS J. 280: 1782-94.
159. Baxter, S.W., Badenes-Perez, F.R., Morrison, A., Vogel, H., Crickmore, N., et al. (2011) Parallel evolution of Bacillus thuringiensis toxin resistance in Lepidoptera. Genetics 189: 675-679.
160. Atsumi, S., Miyamoto, K., Yamamoto, K., Narukawa, J., Kawai, S., et al. (2012) Single amino acid mutation in an ATP-binding cassette transporter gene causes resistance to Bt toxin Cry1Ab in the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 109: E1591-8.
161. Schnepf, E., Crickmore, N., Van Rie, J., Lereclus, D., Baum, J., et al. (1998) Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. R 62: 775-806.
162. Ferre, J., Van Rie, J. (2002) Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 47: 501-533.
163. Griffitts, J.S., Aroian, R.V. (2005) Many roads to resistance: how invertebrates adapt to Bt toxins. Bioessays 27: 614-624.
164. Bravo, A., Soberon, M. (2008) How to cope with insect resistance to Bt toxins? Trends Biotechnol. 26: 573-579. 165. Rajagopal, R., Sivakumar, S., Agrawal, N., Malhotra, P., Bhatnagar, R.K. (2002) Silencing of midgut aminopeptidase
N of Spodoptera litura by double-stranded RNA establishes its role as Bacillus thuringiensis toxin receptor. J. Biol. Chem. 277: 46849-46851.
166. Soberon, M., Pardo-Lopez, L., Lopez, I., Gomez, I., Tabashnik, B.E., et al. (2007) Engineering modified Bt toxins to counter insect resistance. Science 318: 1640-1642.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
618 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
167. Sivakumar, S., Rajagopal, R., Venkatesh, G.R., Srivastava, A., Bhatnagar, R.K. (2007) Knockdown of aminopeptidase-N from Helicoverpa armigera larvae and in transfected Sf21 cells by RNA interference reveals its functional interaction with Bacillus thuringiensis insecticidal protein Cry1Ac. J. Biol. Chem. 282: 7312-7319.
168. Khajuria, C., Buschman, L.L., Chen, M.S., Siegfried, B.D., Zhu, K.Y. (2011) Identification of a novel aminopeptidase P-like gene (OnAPP) possibly involved in Bt toxicity and resistance in a major corn pest (Ostrinia nubilalis). PLoS One 6: e23983.
169. Moran, N.A.(2006) Symbiosis. Curr. Biol. 16: R866-71. 170. Oliver, K.M., Russell, J.A., Moran, N.A., Hunter, M.S. (2003) Facultative bacterial symbionts in aphids confer
resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 100: 1803-1807. 171. Chung, S.H., Rosa, C., Scully, E.D., Peiffer, M., Tooker, J.F., et al. (2013) Herbivore exploits orally secreted bacteria
to suppress plant defenses. Proc. Natl. Acad. Sci. USA 110: 15728-15733. 172. Patil, C.D., Borase, H.P., Salunke, B.K., Patil, S.V. (2013) Alteration in Bacillus thuringiensis toxicity by curing gut
flora: novel approach for mosquito resistance management. Parasitol. Res. 112: 3283-3288. 173. Hernandez-Martinez, P., Naseri, B., Navarro-Cerrillo, G., Escriche, B., Ferre, J., et al. (2010) Increase in midgut
microbiota load induces an apparent immune priming and increases tolerance to Bacillus thuringiensis. Environ. Microbiol. 12: 2730-2737.
174. Xia, X., Zheng, D., Zhong, H., Qin, B., Gurr, G., et al. (2013) DNA sequencing reveals the midgut microbiota of diamondback moth, Plutella xylostella (L.) and a possible relationship with insecticide resistance. PloS One 8: e68852.
175. Itakura, S., Murayama, S., Kamata, Y., Tanaka, H., Enoki, A. (2009) RNA interference in symbiotic protists of the termite Coptotermes formosanus (Isoptera: Rhinotermitidae) through ingestion of siRNA by the host termite. Sociobiology 54: 77-87.
176. Perry, T., Batterham, P., Daborn, P.J. (2011) The biology of insecticidal activity and resistance. Insect Biochem. Mol. Biol. 41: 411-422.
177. Misra, J.R., Lam, G., Thummel, C.S. (2013) Constitutive activation of the Nrf2/Keap1 pathway in insecticide-resistant strains of Drosophila. Insect Biochem. Mol. Biol. 43: 1116-1124.
178. Liu, N., Scott, J.G. (1996) Genetic analysis of factors controlling high-level expression of cytochrome P450, CYP6D1, cytochrome b5, P450 reductase, and monooxygenase activities in LPR house flies, Musca domestica. Biochem. Genet. 34: 133-148.
179. Li, M., Reid, W.R., Zhang, L., Scott, J.G., Gao, X., et al. (2013) A whole transcriptomal linkage analysis of gene co-regulation in insecticide resistant house flies, Musca domestica. BMC Genomics 14: 803.
180. Liu, N., Scott, J.G. (1995) Genetics of resistance to pyrethroid insecticides in the house fly, Musca domestica. Pestic. Biochem. Phys. 52: 116-124.
181. Tomita, T., Liu, N., Smith, F.F., Sridhar, P., Scott, J.G. (1995) Molecular mechanisms involved in increased expression of a cytochrome P450 responsible for pyrethroid resistance in the housefly, Musca domestica. Insect Mol. Biol. 4: 135-140.
182. Zhang, M., Scott, J.G. (1996) Cytochrome b5 is essential for cytochrome P450 6D1-mediated cypermethrin resistance in LPR house flies. Pestic. Biochem. Phys. 55: 150-156.
183. Liu, N., Scott, J.G. (1997) Inheritance of CYP6D1-mediated pyrethroid resistance in house fly (Diptera: Muscidae). J. Econ. Entomol. 90: 1478-14781.
184. Liu, N., Yue, X. (2001) Genetics of pyrethroid resistance in a strain (ALHF) of house flies (Diptera: Muscidae). Pestic. Biochem. Phys. 70: 151-158.
185. Tian, L., Cao, C.W., He, L., Li, M., Zhang, L., et al. (2011) Autosomal interactions and mechanisms of pyrethroid resistance in house flies, Musca domestica. Int. J. Biol. Sci. 7: 902-911.
186. Cohen, M.B., Feyereisen, R. (1995) A cluster of cytochrome P450 genes of the CYP6 family in the house fly. DNA Cell Biol. 14: 73-82.
187. Zhu, F., Liu, N. (2008) Differential expression of CYP6A5 and CYP6A5v2 in pyrethroid-resistant house flies, Musca domestica. Arch. Insect Biochem. Physiol. 67: 107-119.
188. Zhu, F., Feng, J., Zhang, L., Liu, N. (2008) Characterization of two novel cytochrome P450 genes in insecticide-resistant house-flies. Insect Mol. Biol. 17: 27-37.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
619 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
189. Sun, Y., Zou, P., Yu, X., Chen, C., Yu, J., et al. (2012) Functional characterization of an arrestin gene on insecticide resistance of Culex pipiens pallens. Parasit. Vectors 5: 134.
190. Fahrbach, S.E., Smagghe, G., Velarde, R.A. (2012) Insect nuclear receptors. Annu. Rev. Entomol. 57: 83-106. 191. King-Jones, K., Horner, M.A., Lam, G., Thummel, C.S. (2006) The DHR96 nuclear receptor regulates xenobiotic
responses in Drosophila. Cell Metab. 4: 37-48. 192. Seay, D.J., Thummel, C.S. (2011) The circadian clock, light, and cryptochrome regulate feeding and metabolism in
Drosophila. J. Biol. Rhythms. 26: 497-506. 193. Hamby, K.A., Kwok, R.S., Zalom, F.G., Chiu, J.C. (2013) Integrating circadian activity and gene expression profiles
to predict chronotoxicity of Drosophila suzukii response to insecticides. PLoS One 8: e68472. 194. Ffrench-Constant, R.H. (2013) The molecular genetics of insecticide resistance. Genetics 194: 807-815. 195. Ochoa-Campuzano, C., Martínez-Ramírez, A.C., Contreras, E., Rausell, C., Real, M.D. (2013) Prohibitin, an essential
protein for Colorado potato beetle larval viability, is relevant to Bacillus thuringiensis Cry3Aa toxicity. Pestic. Biochem. Phys. 107: 299-308.
196. Gu, L., Knipple, D.C. (2013) Recent advances in RNA interference research in insects: Implications for future insect pest management strategies. Crop Prot. 45: 36-40.
197. Bachman, P.M., Bolognesi, R., Moar, W.J., Mueller, G.M., Paradise, M.S., et al. (2013) Characterization of the spectrum of insecticidal activity of a double-stranded RNA with targeted activity against Western Corn Rootworm (Diabrotica virgifera virgifera LeConte). Transgenic Res. 22: 1207-1222.
198. Palli, S.R. (2014) RNA interference in Colorado potato beetle: steps toward development of dsRNA as a commercial insecticide. Curr. Opin. Insect Sci. 3 DOI: 10.1016/j. cois. 2014. 09. 011 (in press).
199. Dubelman, S., Fishcher, J., Zapata, F., Huizinga, K., Jiang, C., et al. (2014) Environmental fate of double-stranded RNA in the agricultural soils. PLoS One 9: e93155.
200. Chu, C., Sun, W., Spencer, J.L., Pittendrigh, B.R., Seufferheld, M.J. (2014) Differential effects of RNAi treatments on field populations of the western corn rootworm. Pestic. Biochem. Phys. 110: 1-6.
201. Shelton, A.M. (2012) Genetically engineered vegetables expressing proteins from Bacillus thuringiensis for insect resistance: successes, disappointments, challenges and ways to move forward. GM Crops Food 3: 175-183.
202. Qiu, S., Adema, C.M., Lane, T. (2005) A computational study of off-target effects of RNA interference. Nucleic Acids Res. 33: 1834-1847.
203. Davidson, B.L., McCray, P.B., Jr. (2011) Current prospects for RNA interference-based therapies. Nat. Rev. Genet. 12: 329-240.
204. Zhang, L., Hou, D., Chen, X., Li, D., Zhu, L., et al. (2012) Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA. Cell Res. 22: 107-126.
205. Mohr, S.E., Perrimon, N. (2012) RNAi screening: new approaches, understandings, and organisms. Wiley Interdiscip. Rev. RNA 3: 145-158.
Article History: Received 15th April 2014; Revised 19th June 2014 and Accepted on 25th June 2014; and Published 30th October 2014.
Reviewed by: Hua Bai, Brown University, USA. Raman Chandrasekar, Kansas State University, USA.
Cite this article: Zhu F, Cui Y, Walsh DB, Lavine LC. (2014) Application of RNA interference toward insecticide resistance management. In Short Views on Insect Biochemistry and Molecular Biology, eds. R. Chandrasekar, B.K. Tyagi, Z.Z., Gui, G.R., Reeck. International Book Mission, Academic Publisher, Manhattan, USA. Chapter -27, Vol.(2), pp 595-619.

Short Views on Insect Biochemistry and Molecular Biology Vol.(2), 2014 Invited Review
620 Printed in the United States of America, 2014 ISBN: 978-1-63315-205-2
Insect Bioinformatics
© 2014
Section VIII

Related Documents











