Archives of Virology 80, 171--181 (1984) Archives of Virology © by Springer-Verlag 1984 Appearance and Quantification of Infectious Hematopoietic Necrosis Virus in Female Sockeye Salmon (Oncorhynchus nerka) l)uring Their Spawninq Migration By D. MULCAHY, C. K. JE~ES, and R. PASCHO U.S. Fish & Wildlife Service, N~tional Fishery Research Center, Naval Station, Seattle, Washington, U.S.A. With 3 Figures Accepted November 1, 1983 Summary The incidence and amount of infectious hematopoietic necrosis (IHN) virus was determined in 10 organs and body fluids from each of 100 female sockeye salmon (Oncorhynchus nerka) before, during, and after their spawning migration into freshwater. Virus was found in high concentrations only in fish sampled during and after spawning. Infection rates increased from nil to 100 percent within 2 weeks. In spawning fish, incidences of IttN virus were high in all organs and fluids except brain and serum, and the highest concentrations were in the pyloric caeca and lower gut. Immediately before spawning, IHN virus was found most frequently in the gills, less frequently in the pyloric caeca and spleen, and rarely in other organs. Introduction Infectious hematopoietic necrosis (IHN) is a viral disease of salmonid fishes of countries on the h~orth Pacific rim. Although mortality due to the disease is most apparent in fish cultured in hatcheries, at least one serious epizootic has been documented in feral fish (12). The disease and the rhabdovirus causing it have been included in recent reviews of fish viruses (6, 10). Because salmon are anadromous, and reservoirs of b~ection other than the fish themselves are apparently lacking, survival of IHN virus in a salmon population depends on a close association of the virus with the life cycle of its host. Virtually every population of sockeye salmon (Onco-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Archives of Virology 80, 171--181 (1984) Archives of Virology © by Springer-Verlag 1984
Appearance and Quantification of Infectious Hematopoietic Necrosis Virus in Female Sockeye Salmon (Oncorhynchus nerka)
l)uring Their Spawninq Migration
By
D. MULCAHY, C. K. JE~ES, and R. PASCHO U.S. Fish & Wildlife Service,
N~tional Fishery Research Center, Naval Station, Seattle, Washington, U.S.A.
With 3 Figures
Accepted November 1, 1983
Summary
The incidence and amount of infectious hematopoietic necrosis (IHN) virus was determined in 10 organs and body fluids from each of 100 female sockeye salmon (Oncorhynchus nerka) before, during, and after their spawning migration into freshwater. Virus was found in high concentrations only in fish sampled during and after spawning. Infection rates increased from nil to 100 percent within 2 weeks. In spawning fish, incidences of I t tN virus were high in all organs and fluids except brain and serum, and the highest concentrations were in the pyloric caeca and lower gut. Immediately before spawning, IHN virus was found most frequently in the gills, less frequently in the pyloric caeca and spleen, and rarely in other organs.
Introduction
Infectious hematopoietic necrosis (IHN) is a viral disease of salmonid fishes of countries on the h~orth Pacific rim. Although mortality due to the disease is most apparent in fish cultured in hatcheries, at least one serious epizootic has been documented in feral fish (12). The disease and the rhabdovirus causing it have been included in recent reviews of fish viruses (6, 10).
Because salmon are anadromous, and reservoirs of b~ection other than the fish themselves are apparently lacking, survival of IHN virus in a salmon population depends on a close association of the virus with the life cycle of its host. Virtually every population of sockeye salmon (Onco-

t 72 D. Mru'LCAHY, C. K. JENES, and l~. P iscr Io :
rhynchus nerka) examined has been found to include fish with I H N virus and infection rat~s in adults range from a few percent up to 100 percent. Adult sockeye salmon migrate from the ocean into freshwater to spawn, spending up to several months in lakes and rivers attaining sexual matura- tion. The eggs are deposited in the gravel of suitable rivers and streams, and the adults die soon after spawning. Sockeye salmon fry remain in the gravel of the streambed until the yolk sac is absorbed and then emerge from the gravel and immediately migrate to a nursery lake. The fry live in the lake for at least a year before migrating to saltwater. The immature fish remain in the ocean for 3 to 5 years before returning to spawn in the same streams in which they were hatched. There is little overlap of the stages of the life cycle, due to differences in preferred habitat of fish at each stage, making horizontal transmission of the disease between year- classes difficult.
Infectious virus typically can be isolated only during two stages of the sockeye salmon's life cycle: for the first few months after emergence of fry from the gravel (and then only during an epizootic), and when adult fish spawn. After an epizootic, I H N virus enters a latent state in surviving fish and cannot be isolated (1). The reappearance of the virus during spawning is necessary for transmission of the virus with the eggs to the next gener- ation. Protocols for the examination of fish stocks for I H N virus recognize the nature of this pathogen and specify ovarian fluid (OF) as the preferred sample (5, 7). Ovarian or cavity fluid is the viscous liquid associated with salmon eggs after ovulation and can be obtained only immediately before, during, or after spawning. However, I H N virus cannot always be detected in OF from carrier fish (13). As judged by the presence of virus in OF, the incidence of infection increases as the fish mature and spawn, and sampling OF from spent fish offers the best chance for detecting the virus. In a previous s tudy (8), the virus was found to be widely distributed in organs and fluids sampled from infected sockeye salmon captured on the spawning grounds late in the spawning season. In the present study, we sampled sockeye salmon as the population migrated from saltwater to the spawning grounds to determine how soon II-IN virus could be detected and the concentration of virus in specific organs and fluids.
Materials and Methods
F i s h
A total of 100 adult female sockeye salmon were captured at four locations and times of the 1980 brood year during their spa~ l ing migration from Puget, Sound to the Cedar River, Washington. Marine-phase fish were chosen at random aboard ship from those caught by purse seine during commercial fishing in Puget Sound. Fish were obtained by angling in Lake Washington, gill netting at the mouth of the Cedar River, and beach seining in a spawning area of the river. Additional fish were taken from a

I H N Virus in M i g r a t i n g Sockeye Sa lmon 173
h a t c h e r y f i sh -ho ld ing a rea in a s ide -channe l of t h e r iver . F i sh were d e s i g n a t e d as be ing in p r e s p a w n i n g c o n d i t i o n w h e n OF was l ack ing a n d eggs could no t be expressed b y pressure on t he a b d o m e n ; i n s p a w n i n g cond i t i on w h e n O F a n d eggs could be expressed ; a n d s p e n t w h e n on ly a few eggs r e m a i n e d in t he f ish (8).
Sampling Organs and Fluids Procedures for t a k i n g samples for v i rus de t ec t ion a n d q u a n t i f i c a t i o n were desc r ibed
in de t a i l b y MULCAH¥ et al. (8). W h e n f ish were c a p t u r e d to d e t e r m i n e t he in fec t ion ra te , only a n OF sample was t aken . The d e v e l o p m e n t of t h e in fec t ion in i n d i v i d u a l m i g r a t i n g f ish on a n o rgan level was d e t e r m i n e d f rom v i r a l q u a n t i f i c a t i o n of OF, blood, organs , a n d eggs of i n d i v i d u a l spec imens t a k e n in a t i m e sequence. B lood a n d O F were t a k e n a t col lec t ion s i tes a n d earcasses were iced for t r a n s p o r t to t he l abo ra to ry . Samples of eggs, k idneys , spleen, pylor ic caeca, lower gut , l iver, b ra in , a n d gills were t h e n t a k e n a n d p laced in se rum-f ree cell cu l tu re f lu id (MEM) a d j u s t e d to p H 7.2 w i t h t r i s buf fe r c o n t a i n i n g g e n t a m y c i n su l fa te (80 ~xg/mL), penic i l l in (40 I U / m L ) , s t r e p t o m y c i n (40 ~g/ mL) a n d a m p h o t e r i c i n B (2.5 ~g/mL).
Virus Quantification Virus levels were m e a s u r e d b y a s t a n d a r d i z e d p l aque assay (3) on t he E P C cell
l ine in e ight-wel l p la tes ( @5218, L ux Scient i f ic Corp.). The t i ssue samples were d i lu t ed 1 :10 w i t h M E N a n d mechan ieMly homogenized . H o m o g e n a t e s were c lar i f ied b y c e n t r i f u g a t i o n a t 2000 x g for 10 m i n u t e s . Rep l i ca te l - d a y old cell mono laye r s were i n o c u l a t e d w i t h 0. I m L pe r well of u n d i l u t e d sera a n d O F c u d w i t h scr im log10 d i lu t ions to 10 -a (sera) or 10-5 (OF). Organ h o m o g e n a t e s were p laced on cell cu l tures as homoge- nized, a n d w i t h f ive scr im log10 d i lu t ions . Cul tures were f ixed a n d s t a i n e d a f t e r ineu- b a t i o n a t 15 ° C for 7 days . Levels of v i r n s were ea lcu la ted as p l a q u e - f o r m i n g - u n i t s (PFU) pe r g r a m of t i ssue or per mi l l i l i t re of fluid. Mean levels were ca lcu la ted a f t e r va lues d e t e r m i n e d for i n d i v i d u a l samples were t r a n s f o r m e d to log values .
S e r u m f rom each f ish was t e s t ed for t he presence of n e u t r a l i z i n g a c t i v i t y aga in s t I H N vi rus . Because of t he t ox i c i t y of s a lmon se rum for t h e ceil l ine used, sera were d i l u t ed to 1 : 32 a n d I : 64 w i t h M E M before t he tes t . A 1980 iso la te of t he Cedar lg iver s t r a i n of I N N v i ru s passed twice in E P C cells was d i lu t ed to g ive 100 P F U per well in con t ro l wells. S e r u m f rom n o r m a l s t ee thead t r o u t (Salmo gairdneri) a t s imi la r d i lu t ions , a n d M E M were used as nega t i ve controls . E q u a l vo lumes of each d i l u t i on of f ish s e rum a n d d i lu t ed v i ru s were mixed , a n d occas ional ly s t i r red whi le be ing i n c u b a t e d at, r oom t e m p e r a t u r e for one hour . Q u a d r u p l i c a t e wells were i n o c u l a t e d w i t h 0.1 m L a.liquots of t h e s e rum-v i r u s m i x t u r e a t each of t he se rum di lu t ions . Resu l t s w i t h t h e sockeye sera were expressed as pe rcen t of the level of v i rus in controls . Samples t h a t were tox ic were e l imina ted .
Statistics
Corre la t ions in levels of v i rus be tween d i f fe ren t o rgans a n d f luids f rom s p a w n i n g sockeye sMmon were eMcula ted b y Pea r son ' s r (a =~ 0.01). The nul l h y p o t h e s i s was t h a t t he obse rva t i ons were d r a w n f rom a p o p u l a t i o n in wh ich t h e eo r re l a t ion of t he v i rus levels b e t w e e n organs a n d f luids was zero.
Results
Detection o/ the Virus
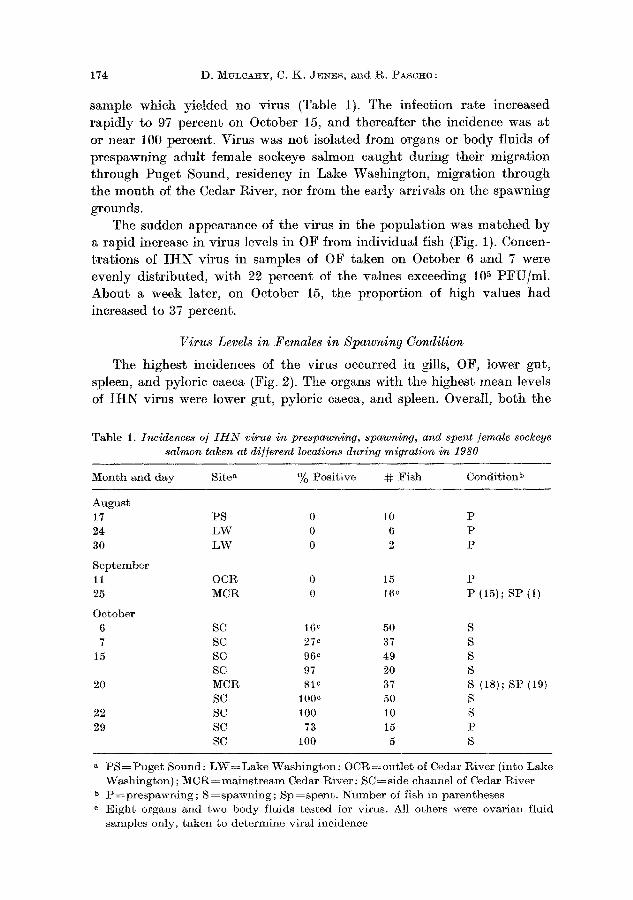
V i r u s w a s f i r s t d e t e c t e d i n 16 p e r c e n t of t h e O F s a m p l e s t a k e n f r o m
f i s h h e l d i n t h e s i d e - c h a n n e l o n O c t o b e r 6, 11 d a y s a f t e r t h e p r e v i o u s
174 D. MULeAH¥, C. K. JENES, and t%. PASCHO:
sample which yielded no virus (Table 1). The infection ra te increased
rapidly to 97 percent on October 15, and thereaf ter the incidence was at
or near I00 percent. Virus was not isolated from organs or b o d y fluids of
prespawning adul t female sockeye salmon caught during their migrat ion th rough Puge t Sound, residency in Lake Washington, migrat ion th rough
the m o u t h of the Cedar River, nor f rom the early arrivals on the spawning
grounds. The sudden appearance of the virus in the popula t ion was matched b y
a rapid increase in virus levels in OF f rom individual fish (Fig. 1). Concen-
t ra t ions of I H N virus in samples of OF taken on October 6 and 7 were evenly distr ibuted, with 22 percent of the values exceeding 105 PFU/ml .
Abou t a week later, on October 15, the propor t ion of high values had
increased to 37 percent.
Virus Levels in Females in Spawning Condition
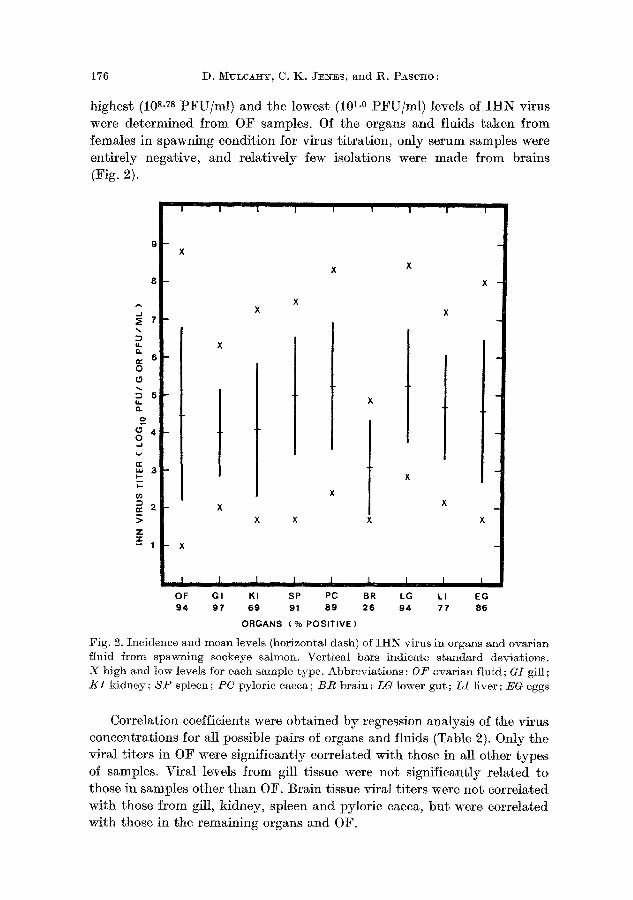
The highest incidences of the virus occurred in gills, OF, lower gut, spleen, and pytoric caeca (Fig. 2). The organs with the highest mean levels
of I I t N virus were lower gut, pyloric caeca, and spleen. Overall, bo th the
Table 1. Incidences o] I H N virus in prespawning, spawning, and spent ]emale sockeye salmon taken at diJ]erent locations during migration in 1980
Month and day Site a % Positive @ Fish Condition b
August 17 24 30
September 11 25
October
PS 0 10 P LW 0 6 P LW 0 2 P
OCR 0 15 P MCR 0 16 c P (15); SP (1)
6 SC 16 c 50 S 7 SC 27 c 37 S
15 SC 96 c 49 S SC 97 20 S
20 MCR 81 e 37 S (18); SP (19) SC 100~ 50 S
22 SC 100 10 S 29 SC 73 15 P
SC 100 5 S
PS = Puget Sound; L~¥ := Lake V~Tashington; OCR == outlet of Cedar River (into Lake Washington); MCR=mainstream Cedar River; SC=side channel of Cedar River
b p =prespawning ; S = spawning; Sp = spent. Number of fish in parentheses c Eight organs and two body fluids tested for virus. All others were ovarian fluid
samples only, taken to determine virM incidence
I H N V i r u s i n M i g r a t i n g S o c k e y e S a h n o n I75
11 t 6-7 OCTOBER 1980 21% INFECTED
0 1 2 3 4 5 6 7 8
.f. ¢n ,T
r,.. tu
z
2 0 -
15
10
r | o 1 2
IIIIIIIIII 15 OCTOBER 1980 97% INFECTED
I I I I I
3 4 5 6 7 8 9 P F U / M L (LOG10)
I i
l O
F i g . 1. F r e q u e n c y d i s t r i b u t i o n of I H N v i r u s l eve l s i n o v a r i a n f l u i d s a m p l e s t a k e n to
d e t e r m i n e v i r M i n c i d e n c e i n s p a w n i n g s o c k e y e s M m o n h e l d i n s i d e c h a n n e l of t h e
C e d a r R i v e r w h e n t h e v i r u s w a s f i r s t d e t e c t e d i n t h e p o p u l a t i o n ( top) a n d a b o u t 9 d a y s l a t e r ( b o t t o m )
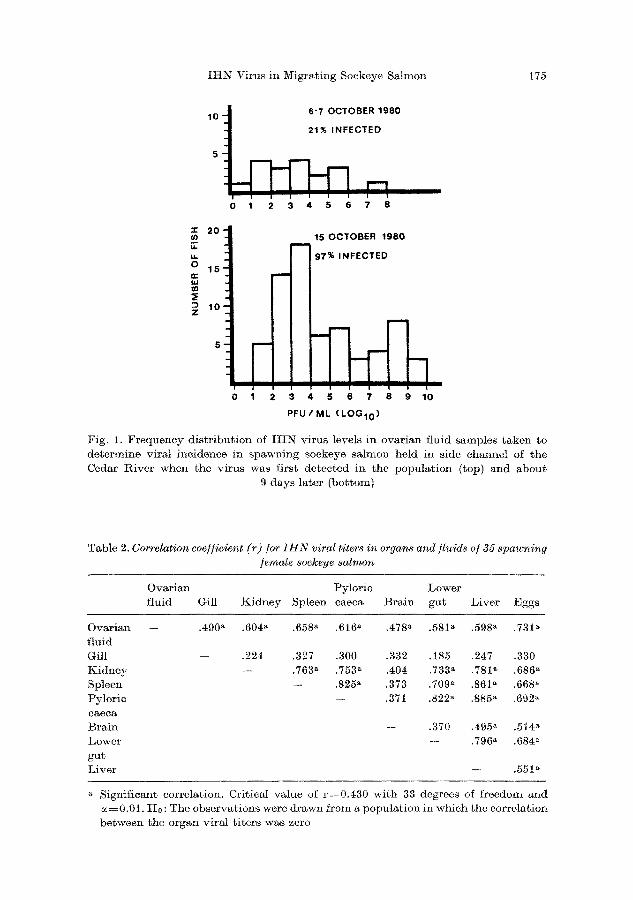
T a b l e 2. Correlation coe//icient (r) /or I H N viral titers in organs and fluids o/35 spawning /create sockeye salmon
O v a r i a n P y l o r i c L o w e r
f l u i d Gil l K i d n e y S p l e e n c a e c a B r a i n g u t L i v e r E g g s
- - .490 a .604 a .658 a .616 a ,478 a .581 a .598 a .731 ~
- - .224 .327 .300 .332 .185 .247 .330
- - .763 a .753 a .404 .733 ~ .781 a .686 ~ - - .825 ~ .373 .709 a .861 ~ .668 a
- - .371 .822 ~ .885 ~ .692 a
Ovarian
f l u i d
Gil l K i d n e y
S p l e e n P y l o r i e
c a e c a
B r a i n
L o w e r
g u t L i v e r
.370 .495 ~ .514 a
- - ,796 a .684a
.551~
Significant correlation. Critical value of r=0.430 with 33 degrees of freedom and
~0.01. H0 : The observations wore drawn from a population in which the correlation
between the organ viral titers was zero
176 D . M V L C A ~ Y , C. K . J:ESrES, a n d ~ . P A s c ~ o :
highest (10 s-Ts PFU/ml ) and the lowest (10 I-0 PF U / mt ) levels of I H N virus were de te rmined f rom OF samples. Of the organs and fluids t aken f rom females in spawning condit ion for virus t i t ra t ion, only serum samples were ent i re ly negative, and relat ively few isolations were made f rom brains (Fig. 2).
8
.J
,,,,,
u. o,. ,y 6 0 0
D 5 tJ. o.
2 O 4 O ,J
er Lu 3
~2 D >
Z "v - - 1
I I I I I ! I II |
t
X
X X
X X X
X X X X
X
I i I i I I I I l OF GI KI SP PC BR LG LI EG 9 4 9 7 6 9 9 1 8 9 2 6 9 4 7 7 8 6
ORGANS ( % POSIT IVE)
Fig. 2. Incidence and mean levels (horizontM dash) of IHN virus in organs and ovarian fluid from spawning sockeye salmon. Vertical bars indicate standard deviations. X high and low levels for each sample type. Abbreviations: 0 F ovarian fluid; GI gill; /~I kidney; S P spleen; P C pylorie caeca; B R brain; LG lower gut; L I liver; EG eggs
Correlat ion coefficients were obta ined b y regression analysis of the virus concentra t ions for all possible pairs of organs and fluids (Table 2). Only the viral t i ters in OF were significantly correlated wi th those in M1 other types of samples. Viral levels f rom gill tissue were not significantly re la ted to those in samples o ther t han OF. Bra in tissue viral t i ters were not corre la ted wi th those f rom gill, kidney, spleen and pylorie caeca, b u t were correla ted with those in the remaining organs and OF.
INN Virus in Migrating Sockeye Salmon 177
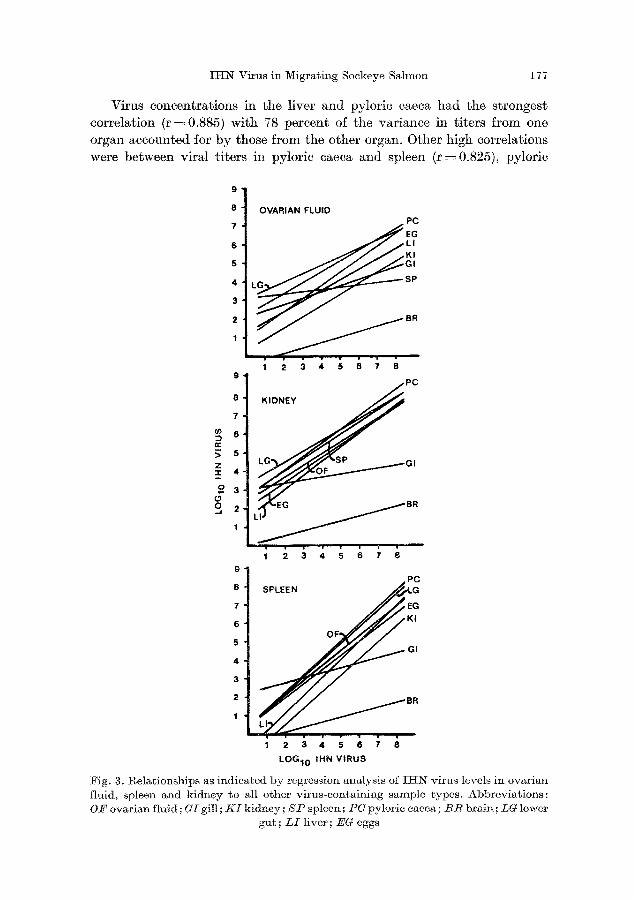
Virus concentrations in the liver and pylorie caeca had the strongest correlation (r =0.885) with 78 percent of the variance in titers from one organ accounted for by those from the other organ. Other high correlations were between viral titers in pylorie caeca and spleen (r=0.825), pylorie
9
8 OVARIAN FLUID 7 PC
4 3 L ~ SP
1 2 3 4 5 6 7 8 9 PC
7 8 K I D N E y
e ~
s
9 PC
8 SPLEE N ~ G
7 EG
6 KI
OF 5 GI 4
3
2 BR
1
1 2 3 4 5 6 7 8
LOG10 IHN VIRUS
Fig. g. l~elationships as indicated by regression analysis o2 II-tN virus levels in ovarian fluid, spleen and kidney to all other virus-containing sample types. Abbreviations: O/~ ovarian fluid; GI gill ; K I kidney; S P spleen; P U pyloric caeca; B R brain; LG lower
gut; L I liver; EO eggs

178 ]). MVLeA~Y, C. K. JENES, and R. PAscJ~o:
caeca and lower gut (r = 0.822), and liver and lower gut (r--~ 0.796), with
68, 68, and 63 percent, respectively, of the var iance accounted for. Kidney,
spleen, and OF are the most common types of samples used for the de- tect ion of I H N virus. When the regressions of virus ti ters for all possible
combinat ions of organs and fluids with these three types of samples were compared, the slopes of the lines were similar except for those for gill and
brain (Fig. 3).
Virus i n Fema le s i n P r e s p a w n i n g Condi t ion
Virus was isolated from 11 of 15 (73 percent) females in prespawning
condition, collected two weeks after I H N virus was detected in high incidence in spawning females. The length of t ime the prespawning females
were present on the spawning grounds was unknown. Mean virus levels of organs were lower t h a n those of spawning fish (Table 3) and no virus was
isolated from serum, lower gut or liver. Virus was isolated from 91 percent of the gill s a m p l e s - - t h e only infected organ in six f i s h - - a n d gills had the
highest mean concentra t ion (103-7G PFU/g) . The gills of one fish failed to
show virus, a l though virus was present in the spleen and pyloric caeca. Overall, I H N virus was isolated f rom four samples of pytoric caeca (36 per-
cent), three of spleen (27 percent) and one each (9 percent) of kidney, brain,
and eggs.
Table 3. Incidences and levels of I H N virus in organs (P t fU/g) and fluids ( P F U / m l ) from naturally infected prespawning female sockeye salmon. Total number of fish sampled was 15, of which 11 were positive for 1HN virus. Only the virus-positive fish are shown. Means are geometric means ]or the positive fish. No virus was isolated from any sample
of serum, lower gut, or liver
Organ (PFU/ml or PFU/g)
Pyloric Fish number Gill Kidney Spleen caeca Brain Eggs
1 3 .0 x 10 2 0 0 0 0 0
2 5.1 × 10 z 0 0 0 0 0 3 1 .0 x 10 2 0 0 0 0 0
4 3 .6 x 10 ~ 0 0 2 .0 × 10 e 0 0
5 5 . 5 x 10 5 0 0 0 0 0
6 4.0 × 10 ~ 3.5 × 10 2 1.3 × 10 s 0 0 0 7 3.5 x 10 ~ 0 0 0 0 0 8 6.0X 10 ~ 0 0 1.4× 10 4 0 4.0x 10 ~ 9 3.5x t0 s 0 1.OX 10 ~ 2.5X 101 5.0x 10 ~ 0
10 0 0 1.1X t0 s 1.0× 10 s 0 0 11 4.1 x 10 4 0 0 0 0 0
~o positive 91 9 27 36 9 9 Mean 5.8XlO s 3.5×10 2 5.2X10 ~ 5.1x10 ~ 5.0x10 ~ 4.0X10 2

IHN Virus in Migrating Sockeye Salmon 179
Serum Neutralizing Antibodies
There was an average 18 percent reduction in plaque counts for all of the sera tested at a 1 : 32 dilution. No serum neutralized all 100 PFU of the virus. Sera from 7 (8 percent) of 87 fish caused a 50 percent plaque reduction of the challenge virus. The virus neutralizing activity of the sera from 13 of the fish in the s tudy was not determined due to missing samples, excessive toxicity, or contamination.
Discussion
The inapparent IHI~ virus infection in sockeye salmon is not activated until immediately before spawning, when the fish are senile and their immune mechanisms are greatly weakened. By only appearing in the animM at a time to be transmitted vertically, the virus :is not exposed to host resistance mechanisms that might eliminate the infection from the indi- vidual animal and the population.
The incidence of infection in spawning or spent fish taken early in the spawning season increased rapidly, approaching I00 percent within about 2 weeks of the first isolation of virus. In a population with a lower overall incidence there would probably be a similar increase in infection rate in spawning and spent fish as the run progresses.
Virus was present in all organs and fluids, except serum, from fish in spawning condition. The development of virus throughout the body may have been due to viremia, but low levels ( ~ 101 PFU/ml) of virus in serum might not be detectable because of the dilution required to avoid toxicity of serum for cell cultures.
The incidence of IHI~ virus was very low in prespawning female fish captured early in the Cedar River sockeye spawning migration. In an earlier study (8), a high incidence of infection was demonstrated in pre- spawning sockeye salmon taken late in their spawning migration. The difference in results obtained in these two studies, done at opposite extremes of the migration period, suggests that there is an increase in virus incidence in the population or a more rapid development of the infection in individual fish as the run progresses. This increase might be due to changing physio- logical or environmental conditions or to greater horizontal transmission of the virus between fish late in the run (when the density of fish is highest and more virus is being released into the water).
The gills are the portal of entry for waterborne fish rhabdovirus infec- tions such as IHN (9), spring viremia of carp (2), and viral hemorrhagic septicemia (4). The high incidence of infection and virus concentrations we found in gill samples in the present study suggests horizontal transmission may be an important factor in achieving the 100 percent incidence in the population. However, the virus levels in most of the gill samples were below the i05 PFU/g level apparently required for the transit of the virus

180 D. Mb~CA~Y, C. K. JElls , and R. PASCI40:
from the gill into the internal organs (9). Also, regression analysis indicated no significant relation between virus levels in the gill and those in all other tissue sampled, except OF. Although waterborne virus released by other fish may be the source of infection for the gitl tissue, it does not appear to be responsible for the systemic infection. Vertical transmission, with a life- long carrier state providing an endogenous source of virus, seems to be the best explanation for the presence of I t IN virus in spawning sockeye salmon.
Serological screening of spawning fish as a means of detecting virus carriers does not appear to be possible because very few of the fish examined showed significant serological evidence of I H N virus infection as judged by serum neutralization tests. Acute II-IN with high mortali ty occurs in juvenile fish, and the virus is then latent until spawning occurs years later. Antibody induced by the primary infection may not persist until sexual maturation. When I H N virus appears during spawning, the fish are senile and immunologically incompetent.
We at tempted to trace the possible site of latency by sampling organs from prespawning fish as the latent infection is reactivated. However, it is difficult to obtain prespawning fish in which the virus is just reappearing. By the time the tests to determine the presence of the virus in the popu- lation are completed, the infection has become systemic in most of the fish, and virus can be recovered from most of the organs. We found five pro- spawning females in which organs other than gills contained I H N virus. The titers in all organs were similar but very low compared with those fish in spawing condition. There is no single organ in which the virus clearly first appears, but I H N virus was found in spleen and pyloric caecae most often.
Testing for the possible site of viral latency by examining correlation coefficients and regression analyses of virus titers in various organs is limited by the assumption that the titers in two organs, one infected by virus released by another, increase at a similar rate. The data presented here suggest that is so, but it is also possible that viral growth in the first organ that supports viral replication may not match that in secondarily infected organs.
IHN virus could be isolated from eggs of only one of the prespawning fish, indicating that eggs are not infected as they develop. Rather than in ovo growth of the virus, development of adequate levels of II-IN virus in OF of carrier fish as they mature to spawning condition, may be necessary for the vertical transmission of IIIN virus.
Because IHN virus cannot be detected in marine-phase sockeye salmon, the shipping of fresh or frozen ocean-caught fish is not a likely source of infection. Early investigators suspected that the feeding of salmon viscera to hatchery fish was a likely source of infection (ll). Our results suggest that offal from ocean-caught fish would not be a likely source of IHN virus.

I H N Virus in Migrating Sockeye Salmon 181
Acknowledgments We thank Kevin Ba:uersfeld and Dick Allen, YVashington Department of Fisheries
for assistance and access to the sockeye salmon population used in this study.
References
1. AMEND, D. F. : Detection and transmission of infectious hematopoietic necrosis virus in rainbow trout. J. WildI. Dis. l l , 471--478 (1975).
2. BAUDOUY, A. M., DA~IYO:N ~, M., h{iERLE, G. : Vir6mie printanigre de Ia carpe. Rdsultats de contaminations exp6rimentales effeetu6es au printemps. Ann. 1%eeh. Vet. 11, 245---249 (1980).
3. BUR:KE, J . A., M u s c a t , D.: Plaquing procedure for infectious hematopoietic necrosis virus. Appl. Environ. Microbiol. 39, 872--876 (1980).
4. CttILNONCZYK, S,, MONGE, D. : Rainbow trout gill pillar cells: demonstration of inert particle phagocytosis and involvement in viral infection. J . 1%eticulocndothel. Soe. 28, 327--332 (1980).
5. Fisheries and Marine Service, Department of Fisheries and the Environment : Fish Health Protection 1%egulations. Manual of Compliance. Misc. Spee. Publ. No. 31, 32 pp. Ottawa: 1976.
6. McALLIS~ER, P. E. : Fish viruses and viral infections. In : F~AE~.~5-CoNRaT, H., WAGNER, 1%. 1%. (eds.), Comprehensive Virology, Vol. 14, 401--470. New York: Plenum 1979.
7. McDANIEL, D.: Procedures for the detection and identification of certain fish pathogens. American Fisheries Society, Fish Health Section, 118 pp. ~¥ashing- ton, D. C.: 1979.
8. Mu~cAHY, D., BURKE, J. , PaSCHO, R., JENES, C. K. : Pathogenesis of infectious hematopoietic necrosis virus in spawning sockeye salmon (Oncorhynchus nerlca), Can. J. Fish. Aquat. Sei. 39, t144-- 1149 (1982).
9. Mm;caI-IY, D., Pasc~o, 1%. J . , JENES, C. K. : Detection of infectious hemato- poietie necrosis virus in river water and demonstration of waterborne transmission. J. Fish. Dis. 6, 321--330 (1983).
t0. PmcI~R, K. S., FRYEr, J . L.: The viral diseases of fish: A review through 1978. Par t I : Diseases of proven viral, etiology. CRC CAt. l%ev. Micro. 7, 287--364 (1980).
i1. Vy~rsoN, S. ~¥., GUENT~ER, 1~. W., 1%UCKER, 1%. ~ . : Virus disease of sockeye sM- mon. Inter im report, 36 pp. U.S. Fish Wildl. Serv. Spec. Sci. l%ept.--Fisheries No. 138. Washington, D.C.: 1954.
12. WHmla~S, I. V., A~fEXD, D. F. : A natural cpizootic of infectious hematopoietic necrosis in fry of sockeye salmon (Oncorhynchus nerka) at, Chilko Lake, British Columbia. J. Fish. l~es. Board Can. 33, 1564~--1567 (1976).
13. WINGFIELn, W. It . , C~aN, 15. D. : Studies on the Sacramento River chinook disease and its causative agent. In : SNIESZKO, S. F. (ed.), A Symposium on Disease of Fishes and Shellfishes (Special Publication No. 5), 307--318. Washington, D. C. : American Fisheries Society 1970.
Authors' address: Dr. D. M~rLCA:uY, National Fishery 1%esearch Center, Seattle Bldg. 204, Naval Suppor~ Activity, Seattle, WA 98115, U.S.A.
1%eceived June 8, 1983
13 Arch. Virol. 80/2--3
Related Documents











