ORIGINAL PAPER Aortic coarctation hypertension induces fibroblast growth factor-2 immunoreactivity in the stimulated nucleus tractus solitarii Debora Rejane Fior-Chadi Tatiana Cristina Nogueira Varella Jessica Ruivo Maximino Gerson Chadi Received: 28 February 2007 / Accepted: 16 May 2007 / Published online: 12 June 2007 ȑ Springer Science+Business Media B.V. 2007 Abstract The actions of neurotrophic factors i.e. basic fibroblast growth factor (bFGF, FGF-2) to neurons are related not only to neuronal development and maintenance but also to synaptic plasticity regarding neurotransmission. We analyzed here the levels of FGF-2 immunoreactivity in the nucleus tractus solitarii (NTS) of Wistar Kyoto rats in response to alterations of neuronal activity promoted by the stimulation of the baroreceptor reflex following an aortic coarctation-induced-hypertension. The FGF-2 immunore- activity (IR) was found in the cytoplasm of the neurons and in the nuclei of the glial cells in the NTS. A large number of NTS neurons expressed FOS immunoreactivity 4 h after coarctation, as an indication of neuronal activity. Stereol- ogical methods showed an increased number of FGF-2 immunoreactive (ir) neuronal profiles (90%) and glial profiles (149%) in the NTS of the 72 h aortic coarctated rats. 1-week later, FGF-2 ir neurons were still increased (54%) but no change was found in the number of FGF-2 ir glial profiles. The double immunoperoxidase method revealed that the majority of the FGF-2 ir glial cells was glial fibrillary acidic protein (GFAP) positive astrocytes. GFAP immunohistochemistry showed an astroglial reaction at 72 h time-interval (55%) but not 1 week after stimula- tion. The number of the cresyl violet positive neurons and OX42 ir profiles (marker of activated microglia) in the NTS of coarctated rats were not different from control by 1 week and 1 month after the surgery, indicating a lack of NTS injury in this period following coarctation hypertension. FGF-2 may be an important neurotrophic factor in areas involved in the control of blood pressure. The increased FGF-2 IR in the NTS cells following neuronal stimulation may represent trophic and plastic adaptive responses in this nucleus in an autocrine/paracrine fashion. Keywords Fibroblast growth factor-2 Á Coarctation hypertension Á Nucleus tractus solitarii Á Immunohistochemistry Á Astrocytes Á Neuronal stimulation Introduction Basic fibroblast growth factor (bFGF, FGF-2) is a mitogenic protein with actions on multiple cell types such as fibro- blasts, nerve cells and glial cells (Baird and Klagsbrun 1991). FGF-2 has also been described as a potent angio- genic factor (Gospodarowicz 1990; Puumala et al. 1990). FGF-2 plays a role in the survival and plasticity of neurons in many brain regions of adult rats (Walicke 1988; Chadi et al. 1993a). Much has been learned about the trophic properties of FGF-2 on neuronal survival from in vitro and in vivo studies, however the role of this growth factor in response to altera- tions of neuronal activity has been poorly investigated. It has been suggested that the expression of FGF-2 in the brain of adult animals changes in response to an excessive neuronal excitation as observed following electroconvulsive shock (Follesa et al. 1994) and after convulsive seizures (Riva et al. 1992) or after physical exercise (Gomez-Pinilla et al. 1997). Several peptide growth factors have been postulated to be important in the initiation and progression of vascular growth D. R. Fior-Chadi Á T. C. N. Varella Department of Physiology, Institute of Biosciences, University of Sa ˜o Paulo, Sao Paulo 05508-900, Brazil J. R. Maximino Á G. Chadi (&) Department of Neurology, University of Sa ˜o Paulo School of Medicine, Av. Dr. Arnaldo 455, 2nd Floor, Room 2119, Sao Paulo 01246-903, Brazil e-mail: [email protected] 123 J Mol Hist (2007) 38:285–294 DOI 10.1007/s10735-007-9101-x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Aortic coarctation hypertension induces fibroblast growth factor-2immunoreactivity in the stimulated nucleus tractus solitarii
Debora Rejane Fior-Chadi ÆTatiana Cristina Nogueira Varella ÆJessica Ruivo Maximino Æ Gerson Chadi
Received: 28 February 2007 / Accepted: 16 May 2007 / Published online: 12 June 2007
� Springer Science+Business Media B.V. 2007
Abstract The actions of neurotrophic factors i.e. basic
fibroblast growth factor (bFGF, FGF-2) to neurons are
related not only to neuronal development and maintenance
but also to synaptic plasticity regarding neurotransmission.
We analyzed here the levels of FGF-2 immunoreactivity in
the nucleus tractus solitarii (NTS) of Wistar Kyoto rats in
response to alterations of neuronal activity promoted by the
stimulation of the baroreceptor reflex following an aortic
coarctation-induced-hypertension. The FGF-2 immunore-
activity (IR) was found in the cytoplasm of the neurons and
in the nuclei of the glial cells in the NTS. A large number of
NTS neurons expressed FOS immunoreactivity 4 h after
coarctation, as an indication of neuronal activity. Stereol-
ogical methods showed an increased number of FGF-2
immunoreactive (ir) neuronal profiles (90%) and glial
profiles (149%) in the NTS of the 72 h aortic coarctated
rats. 1-week later, FGF-2 ir neurons were still increased
(54%) but no change was found in the number of FGF-2 ir
glial profiles. The double immunoperoxidase method
revealed that the majority of the FGF-2 ir glial cells was
glial fibrillary acidic protein (GFAP) positive astrocytes.
GFAP immunohistochemistry showed an astroglial reaction
at 72 h time-interval (55%) but not 1 week after stimula-
tion. The number of the cresyl violet positive neurons and
OX42 ir profiles (marker of activated microglia) in the NTS
of coarctated rats were not different from control by 1 week
and 1 month after the surgery, indicating a lack of NTS
injury in this period following coarctation hypertension.
FGF-2 may be an important neurotrophic factor in areas
involved in the control of blood pressure. The increased
FGF-2 IR in the NTS cells following neuronal stimulation
may represent trophic and plastic adaptive responses in this
nucleus in an autocrine/paracrine fashion.
Keywords Fibroblast growth factor-2 �Coarctation hypertension � Nucleus tractus solitarii �Immunohistochemistry � Astrocytes � Neuronal stimulation
Introduction
Basic fibroblast growth factor (bFGF, FGF-2) is a mitogenic
protein with actions on multiple cell types such as fibro-
blasts, nerve cells and glial cells (Baird and Klagsbrun
1991). FGF-2 has also been described as a potent angio-
genic factor (Gospodarowicz 1990; Puumala et al. 1990).
FGF-2 plays a role in the survival and plasticity of neurons
in many brain regions of adult rats (Walicke 1988; Chadi
et al. 1993a).
Much has been learned about the trophic properties of
FGF-2 on neuronal survival from in vitro and in vivo studies,
however the role of this growth factor in response to altera-
tions of neuronal activity has been poorly investigated. It has
been suggested that the expression of FGF-2 in the brain of
adult animals changes in response to an excessive neuronal
excitation as observed following electroconvulsive shock
(Follesa et al. 1994) and after convulsive seizures (Riva et al.
1992) or after physical exercise (Gomez-Pinilla et al. 1997).
Several peptide growth factors have been postulated to be
important in the initiation and progression of vascular growth
D. R. Fior-Chadi � T. C. N. Varella
Department of Physiology, Institute of Biosciences,
University of Sao Paulo, Sao Paulo 05508-900, Brazil
J. R. Maximino � G. Chadi (&)
Department of Neurology, University of Sao Paulo School of
Medicine, Av. Dr. Arnaldo 455, 2nd Floor, Room 2119,
Sao Paulo 01246-903, Brazil
e-mail: [email protected]
123
J Mol Hist (2007) 38:285–294
DOI 10.1007/s10735-007-9101-x

following different types of experimental hypertension
(Saltis et al. 1993; Zettler and Rush 1993; Anwar and
Delafontaine 1994), however little information is available
on the involvement of neurotrophic factors in the central
control blood pressure areas.
The subdiaphragmatic aortic coarctation is an experi-
mental model used to induce a rapid and sustained hyper-
tension in rats (Krieger 1970; Michelini et al. 1992). When
arterial pressure raises there is an increased afferent activity
directed to the nucleus tractus solitarii (NTS) (Mifflin et al.
1988; Langhorst et al. 1992; Li and Dampney 1992; Rogers
et al. 1993; Li and Dampney 1994), the primary site of
integration of the baroreceptor reflex (Ciriello and Calaresu
1981; Spyer 1981; Ross et al. 1985; Chapleau et al. 1995).
The NTS, in turn, outputs to efferent neurons that provide the
parasympathetic (Loewy and Burton 1978; Nosaka et al.
1979) and sympathetic (Soato and Krieger 1974; Cravo et al.
1991; Rogers et al. 1993) innervations of the cardiovascular
system. The aortic coarctation model of induced hyperten-
sion was employed in this study since it promotes a moderate
elevation in the level of arterial pressure leading to neuronal
activation in the NTS without neuronal damage. Stroke and
expression of neurotrophic factors have been observed only
in ageing stroke-prone spontaneously hypertensive rats that
show mean arterial pressure around 240 mmHg (Lin et al.
1997) but not in other models of less severe inducible
hypertension.
In the present work the coarctation hypertension
approach was used to produce a sustained increase of neu-
ronal activity in the NTS, without neuronal damage, in
order to evaluate whether this activation may modify the
FGF-2 immunoreactivity in the NTS cells of the Wistar
Kyoto rats.
Methods
Animal treatment
Specific pathogen-free adult male Wistar Kyoto rats from
the University of Sao Paulo (USP) with body weight (b.w.)
of 250 g were used in the experiments. The animals were
kept under standardized lighting conditions (lights on at
7:00 a.m. and off at 7:00 p.m.), at a constant temperature
of 23�C and with free access to food pellets and tap water.
The rats were submitted to a sham-operation or to a sub-
diaphragmatic aortic coarctation as previously described
(Krieger 1970). Briefly, median laparotomy was performed
under chloral hydrate (7%, Merck, Germany, 350 mg/Kg/
b.w., i.p) anaesthesia and the abdominal aorta was isolated.
A cotton thread was used to constrict the aorta immediately
below the diaphragm and the extent of narrowing was
limited by a hypodermic needle of 0.7 or 0.8 mm in
diameter depending on rat weight. In the sham-operation
the aorta was exposed and isolated in an identical manner
but it was not tied. The animals were sacrificed 72 h
(n = 11), 1 week (n = 8) and 1 month (n = 9) after the
surgery. In the day before killing, the rats from groups of
72 h and 1 week were submitted to arterial pressure
recording. Rats were then anaesthetized with ether (Sigma,
USA) and a tygon catheter filled with heparinized (500 UI/
ml, Liquemine, Roche) saline was placed in the ascending
aorta through the brachial artery. The other end of the
cannula was slipped beneath the skin and exteriorized on
the back of the neck for arterial pressure recording. After a
20 min stabilization period, the arterial pressure was re-
corded during 15 min in the freely moving rats by means of
a transducer connected to a Recorder (Narco Bio-Systems,
USA). Finally, a set of sham-operated (n = 5) and aortic-
coarctated (n = 5) rats were sacrificed 4 h after surgery in
order to evaluate FOS protein expression in the NTS
neuronal cells, as a marker of neuronal activity in this
nucleus. Those animals were not submitted to arterial
pressure recording.
Immunohistochemical procedures
After blood pressure recording the rats were deeply
anaesthetized with chloral hydrate 10% (420 mg/kg, b.w.,
i.p.) and killed by a perfusion through a cannula inserted in
the ascending aorta with 50 ml of isotonic saline at room
temperature followed by 350 ml of fixation fluid (4�C)
during 6 min. The fixative (Zamboni and De Martino 1969)
consisted of 4% (w/v) paraformaldehyde and 0.2% (v/v)
picric acid in 0.1 M phosphate buffer (pH 6.9). The brains
were dissected out and kept in the fixative solution for
90 min. The fixed brains were washed in 10% sucrose
dissolved in 0.1 M phosphate buffered saline (PBS) for
2 days, frozen in dry ice and stored at –70�C freezer.
Coronal adjacent serial 50 lm thick sections were obtained
in a cryostat (Leica, CM 3000, Germany) from the NTS
region, from rostro-caudal level –13.40 mm to –14.30 mm
according to the atlas of Paxinos and Watson (1986). The
sections were sampled systematically during sectioning.
Five series in a rostro-caudal order including every fifth
section were taken to immunohistochemistry.
The immunoreactivities were detected using avidin-
biotin peroxidase technique (Hsu et al. 1981). Floating
sections were washed 2 · 10 min in PBS (0.1 M, pH 7.4)
at room temperature and incubated with normal goat serum
(NGS) 5% for 30 min at room temperature under shaking.
Following, the sections from the series were incubated for
48 h at 4�C under shaking with the rabbit polyclonal FGF-
2 antiserum (Gonzalez et al. 1990) (diluted 1:800), a rabbit
policlonal glial fibrillary acidic protein (GFAP, a marker
for astrocytes, Dakopatts, Denmark, diluted 1:200) or also
286 J Mol Hist (2007) 38:285–294
123

with a mouse monoclonal antibody against the OX42 (a
marker for microglia, Harlan, England, diluted 1:600). The
GFAP and OX42 immunoreactivities were counterstained
with cresyl violet to allow interalia the demonstration of
the nuclei in the astrocytes and microglial profiles as well
as the nucleoli and the Nissl substance in the neuronal
profiles. Sections from rats sacrificed 4 h after surgery
were incubated with a sheep polyclonal anti-FOS antibody
(Affinity, UK) as described above. The antibodies were
diluted in PBS containing 0.5% Triton X-100 (Sigma) and
1% bovine serum albumin (Sigma). The detection of the
antibodies was achieved by the indirect immunoperoxidase
method (ABC) (Hsu et al. 1981) using the avidin-biotin
peroxidase technique as previously described (Chadi et al.
1993a, b). After washing in PBS (3 · 10 min), the sections
were incubated with a biotinylated goat anti-rabbit, or
horse anti-mouse or rabbit anti-goat immunoglobulins
(diluted 1:200, Vector, USA) for 1 h. In a third step the
sections were washed in PBS and incubated with avidin-
biotin peroxidase complex (both diluted 1:100, Vectastain,
Vector) during 45 min. The staining was performed using
0.03% of 3, 3¢ diaminobenzidine tetrahydrochloride (DAB,
Sigma) as a chromogen and 0.05% (v/v) of H2O2 (Sigma)
during 8 min, which gave a brownish colour to the
immunoreaction. For standardization of the immunohisto-
chemical procedure we have used a dilution of the primary
antibody and a concentration of DAB far from saturation
and an incubation time adjusted so that the darkest
elements in the brain sections were below saturation (Zoli
et al. 1990). The FGF-2 antiserum used is a well charac-
terized polyclonal antiserum raised against the n terminal
(residues 1–24) of the synthetic peptide of bovine FGF-2
(1–146) (Gonzalez et al. 1990). This antiserum does not
recognize acidic FGF (FGF-1, cross reactivity less than
1%) (Gonzalez et al. 1990). The OX42 antibody employed
in the present analysis recognizes the complement receptor
type 3 (CR3), an immunologically important surface mol-
ecule of mononuclear phagocytes and it is considered a
well established tool to label resting and activated micro-
glia in the central nervous system (Ling et al. 1990). As
control, sections were incubated overnight at 4�C with the
FGF-2 antiserum pre-incubated with human recombinant
FGF-2 (50 lg/ml, for 24 h at 4�C). To further analyze the
specificity of the immunostainings, sections were incubated
with the solvent of the primary and secondary antibodies or
with the solvent of the avidin-biotin solution and processed
at the same time with the experimental sections.
Double immunolabeling
The two-colour immunoperoxidase method was employed
in a series of sections for the simultaneously detection of
the FGF-2 and the GFAP immunoreactivities. The FGF-2
immunoreactivity was firstly demonstrated as described
above. Following the DAB reaction, the sections were
rinsed several times in PBS and incubated during 48 h in a
humidified chamber with the rabbit polyclonal antiserum
against GFAP, as described above (1:400). After several
rinses in PBS, the sections were incubated with biotiny-
lated goat anti-rabbit immunoglobulins (1:200, Vector) for
1 h at room temperature and with avidin-biotin peroxidase
couplex (both diluted 1:100, Vectastain, Vector) for
45 min at room temperature. The staining was performed
using 4-chloro naphtol 0.05% (Sigma) as a chromogen and
0.05% (v/v) of H2O2 (Sigma) during 10 min. This proce-
dure gave brownish and bluish colours to the FGF-2 and
GFAP immunoreactions, respectively.
Quantitative stereological analysis
The FGF-2, GFAP and OX42 immunoreactive cell profiles
and the neurons stained by cresil violet were quantified by
stereological analysis. The optical fractionator (West and
Gundersen 1990; Chadi et al. 1993a; Janson and Moller
1993) was used to estimate the number of FGF-2, GFAP
and OX42 immunoreactive profiles in the sampled volume
of the NTS, bilaterally.
As described, every fifth section was systematically
sampled (f1 = 5). The series of sections immunostained to
the above described antisera were analyzed using a CAST-
system (Computer Assisted Stereological Toolbox). Briefly:
an Olympus BX50 microscope (Olympus, Denmark) was
interfaced with a computer (IBM 330-P75, USA) and a
colour video camera (JAL 2040, Protec, Japan), both linked
to a colour video monitor (G70, IBM). The GRID software
package (Interactivision, Silkeborg, Denmark) was used to
generate sampling (counting) frames as an overlay image to
the microscopic image on the monitor as well as to control
the motorized X-Y stage (Lang, Huttenberg, FRG). A
microcator (MT12, Heidenhain, FRG) was linked to the
microscope to monitor movements in a vertical (Z) direction.
The border of a square, of an area of 75 lm2, representing the
sampled region to be quantified was outlined on both sides
of the NTS using a 4 · objective. The step rates were entered
(50 lm), after which the program created series of uniformly
sampled fields of vision throughout the entire delimitated
region. For counting the cells, a 100 · oil-immersion
objective with the numerical aperture 1.4 was used.
A counting frame (with a known area, aframe) was created by
the GRID software, and by knowing the step-rates it was
possible to calculate the second sampled fraction (f2):
(X step-length�Y step-length)/aframe). The sampling volume
(disector) in the Z-axis extended 9 lm deep (height of the
disector) after excluding the parts of the section close to the
slide and coverslip (3 lm). Also the total thickness of the
section was measured, giving the third sampling fraction (f3):
J Mol Hist (2007) 38:285–294 287
123

height of the section/height of the disector. After having
counted all cells (P
Q–) fulfilling the criteria for being
sampled, the total number of cells (Ntotal) in the sampled
region was estimated: Ntotal =P
Q–�f1�f2�f3The coefficient of error (CE) for the estimated total
number of immunoreactive particles in the sample volume
in the NTS of each animal was calculated (Gundersen and
Jensen 1987; Pakkenberg and Gundersen 1988; West et al.
1991).
The cytoplasmatic and nuclear localization of the FGF-2
immunoreactivity (Chadi et al. 1993b; Cintra et al. 1991)
were taken into account in the discrimination of the neu-
ronal and glial FGF-2 cell profiles (see the results).
Statistical analysis
The statistical analysis was performed using the non-
parametric two-tailed Mann-Whitney U-test (Hollander
and Wolfe 1973). The numbers represent the mean ±
S.E.M.
Results
Blood pressure recording
The subdiaphragmatic aortic coarctation resulted in a sus-
tained moderate hypertensive response observed at 72 h
and 1 week following the surgery. 72-h after the surgery
the arterial pressure was 158 ± 2.4 mmHg in the coarctated
rats (n = 5) and 109 ± 6.9 mmHg in the sham operated rats
(n = 6) (P < 0.05), according to the Mann-Whitney
U-test). 1 week after the surgery the mean arterial pressure
was 156 ± 2.4 mmHg in the coarctated rats (n = 4) and
114 ± 1.9 mmHg in the sham operated rats (n = 4)
(P < 0.05, according to the Mann-Whitney U-test). Arterial
pressure was not recorded after 1 week since it is known
that the arterial pressure is maintained constant after this
period (Giuliano and Brezenoff 1987; also unpublished
observations of our laboratory).
Qualitative analysis of the FGF-2 immunoreactivity
The FGF-2 immunoreactivity was seen in the nucleus of
glial cells and in the cytoplasm of neurons in all extent of
the analyzed regions in the NTS (Fig. 1), area postrema,
dorsal motor nucleus of the vagus and other brain stem
nuclei. The FGF-2 immunoreactivity was also observed in
some fibres within the NTS (Fig. 1).
The aortic coarctation increased the intensity and den-
sity of the FGF-2 immunoreactive cell profiles in the NTS
of the 72 h (Fig. 1) and 1 week operated rats.
Quantitative analysis
The stereological method employing the optical fraction-
ator demonstrated an increased number of the cytoplasmic
FGF-2 immunoreactive neuronal profiles (90 ± 23% of the
control group, n = 5) and nuclear FGF-2 immunoreactive
glial profiles (149 ± 11% of the control group, n = 5) on
both sides of the NTS of the coarctated rats at the 72 h
time-interval (Fig. 2A). 1 week after the coarctation the
Fig. 1 Microphotographs of coronal sections showing the FGF-2
immunoreactivity in the nucleus tractus solitarii (NTS) of the sham-
operated (control) (A, B) and the 72 h aortic coarctated (C, D) rats. Band D represent higher magnification of areas depicted from the
nucleus tractus solitarii of A and C, respectively. The FGF-2
immunoreactivity is present in the cytoplasm of neurons (arrows)
and in the nuclei of glial cells (arrowheads) of the NTS. The FGF-2
immunoreactivity is also observed in some neuronal fibres in the NTS
(open arrow). The aortic coarctation increases the FGF-2 immuno-
reactivity in the neurons and glial cells 72 h after the surgery.
Bars = 100 lm (A, C) and 10 lm (B, D)
288 J Mol Hist (2007) 38:285–294
123
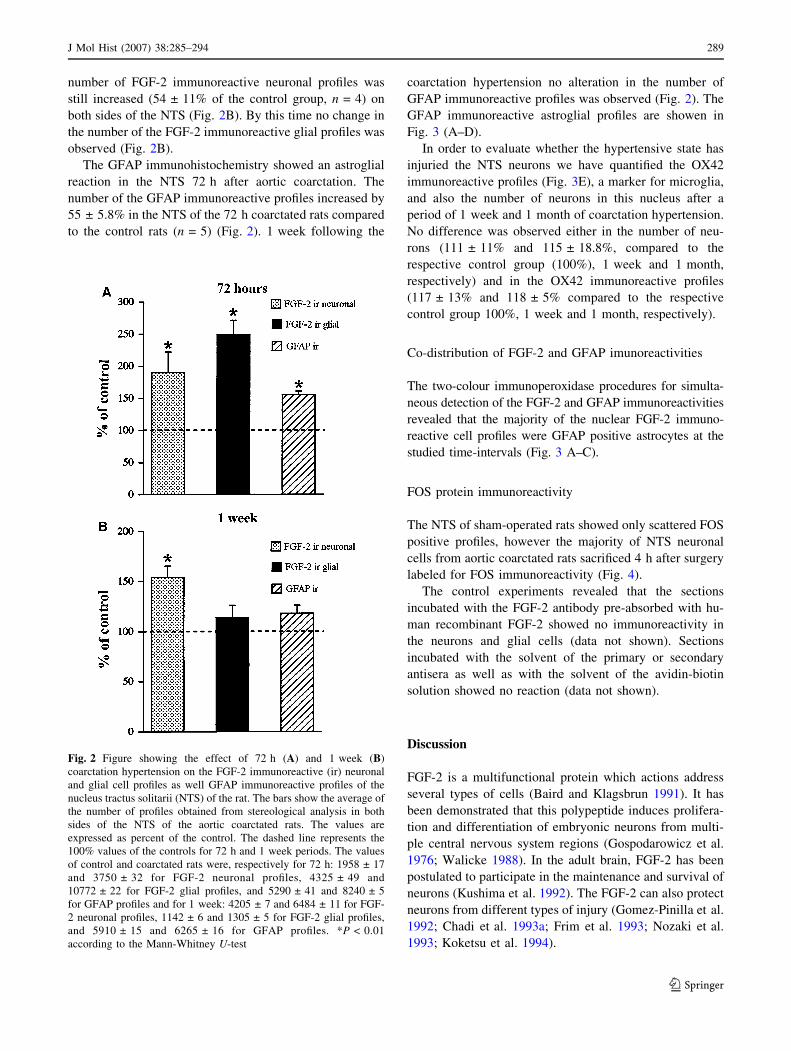
number of FGF-2 immunoreactive neuronal profiles was
still increased (54 ± 11% of the control group, n = 4) on
both sides of the NTS (Fig. 2B). By this time no change in
the number of the FGF-2 immunoreactive glial profiles was
observed (Fig. 2B).
The GFAP immunohistochemistry showed an astroglial
reaction in the NTS 72 h after aortic coarctation. The
number of the GFAP immunoreactive profiles increased by
55 ± 5.8% in the NTS of the 72 h coarctated rats compared
to the control rats (n = 5) (Fig. 2). 1 week following the
coarctation hypertension no alteration in the number of
GFAP immunoreactive profiles was observed (Fig. 2). The
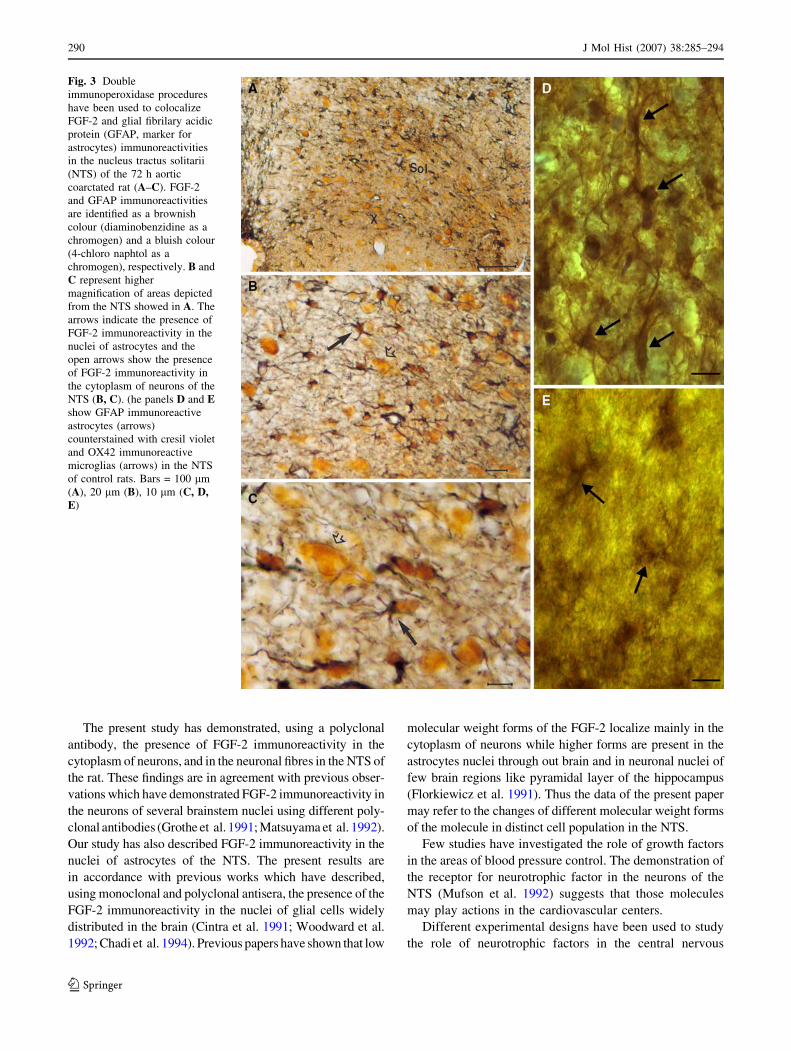
GFAP immunoreactive astroglial profiles are showen in
Fig. 3 (A–D).
In order to evaluate whether the hypertensive state has
injuried the NTS neurons we have quantified the OX42
immunoreactive profiles (Fig. 3E), a marker for microglia,
and also the number of neurons in this nucleus after a
period of 1 week and 1 month of coarctation hypertension.
No difference was observed either in the number of neu-
rons (111 ± 11% and 115 ± 18.8%, compared to the
respective control group (100%), 1 week and 1 month,
respectively) and in the OX42 immunoreactive profiles
(117 ± 13% and 118 ± 5% compared to the respective
control group 100%, 1 week and 1 month, respectively).
Co-distribution of FGF-2 and GFAP imunoreactivities
The two-colour immunoperoxidase procedures for simulta-
neous detection of the FGF-2 and GFAP immunoreactivities
revealed that the majority of the nuclear FGF-2 immuno-
reactive cell profiles were GFAP positive astrocytes at the
studied time-intervals (Fig. 3 A–C).
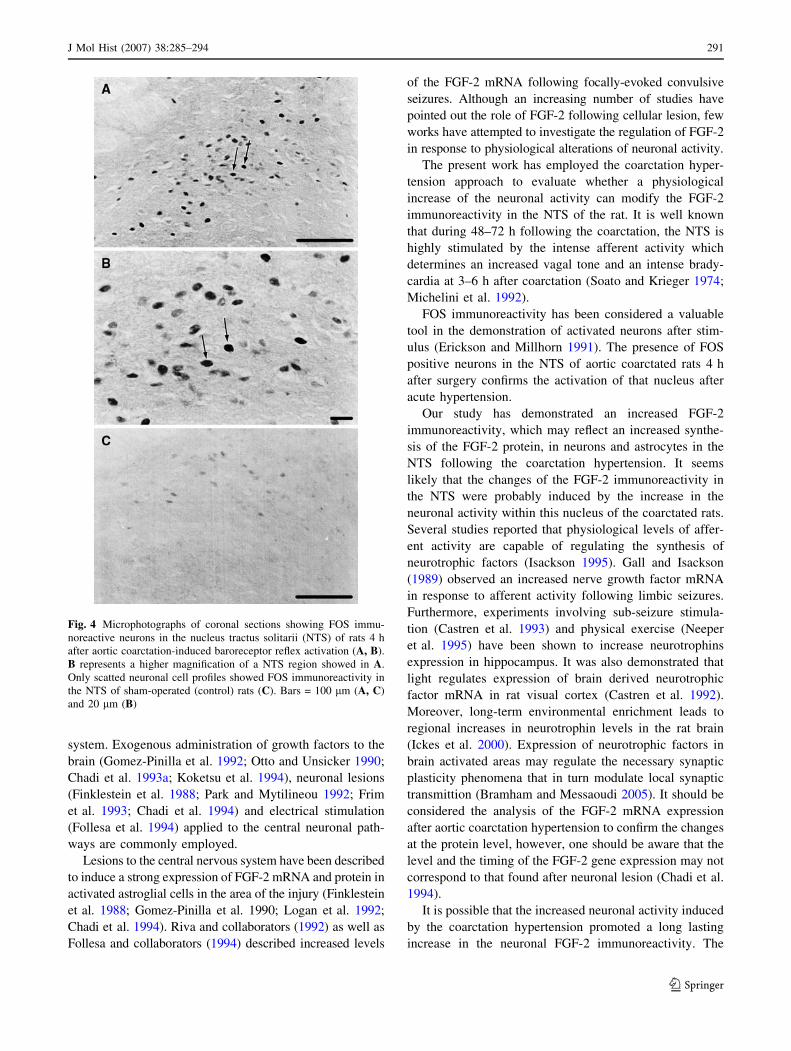
FOS protein immunoreactivity
The NTS of sham-operated rats showed only scattered FOS
positive profiles, however the majority of NTS neuronal
cells from aortic coarctated rats sacrificed 4 h after surgery
labeled for FOS immunoreactivity (Fig. 4).
The control experiments revealed that the sections
incubated with the FGF-2 antibody pre-absorbed with hu-
man recombinant FGF-2 showed no immunoreactivity in
the neurons and glial cells (data not shown). Sections
incubated with the solvent of the primary or secondary
antisera as well as with the solvent of the avidin-biotin
solution showed no reaction (data not shown).
Discussion
FGF-2 is a multifunctional protein which actions address
several types of cells (Baird and Klagsbrun 1991). It has
been demonstrated that this polypeptide induces prolifera-
tion and differentiation of embryonic neurons from multi-
ple central nervous system regions (Gospodarowicz et al.
1976; Walicke 1988). In the adult brain, FGF-2 has been
postulated to participate in the maintenance and survival of
neurons (Kushima et al. 1992). The FGF-2 can also protect
neurons from different types of injury (Gomez-Pinilla et al.
1992; Chadi et al. 1993a; Frim et al. 1993; Nozaki et al.
1993; Koketsu et al. 1994).
Fig. 2 Figure showing the effect of 72 h (A) and 1 week (B)
coarctation hypertension on the FGF-2 immunoreactive (ir) neuronal
and glial cell profiles as well GFAP immunoreactive profiles of the
nucleus tractus solitarii (NTS) of the rat. The bars show the average of
the number of profiles obtained from stereological analysis in both
sides of the NTS of the aortic coarctated rats. The values are
expressed as percent of the control. The dashed line represents the
100% values of the controls for 72 h and 1 week periods. The values
of control and coarctated rats were, respectively for 72 h: 1958 ± 17
and 3750 ± 32 for FGF-2 neuronal profiles, 4325 ± 49 and
10772 ± 22 for FGF-2 glial profiles, and 5290 ± 41 and 8240 ± 5
for GFAP profiles and for 1 week: 4205 ± 7 and 6484 ± 11 for FGF-
2 neuronal profiles, 1142 ± 6 and 1305 ± 5 for FGF-2 glial profiles,
and 5910 ± 15 and 6265 ± 16 for GFAP profiles. *P < 0.01
according to the Mann-Whitney U-test
J Mol Hist (2007) 38:285–294 289
123
The present study has demonstrated, using a polyclonal
antibody, the presence of FGF-2 immunoreactivity in the
cytoplasm of neurons, and in the neuronal fibres in the NTS of
the rat. These findings are in agreement with previous obser-
vations which have demonstrated FGF-2 immunoreactivity in
the neurons of several brainstem nuclei using different poly-
clonal antibodies (Grothe et al. 1991; Matsuyama et al. 1992).
Our study has also described FGF-2 immunoreactivity in the
nuclei of astrocytes of the NTS. The present results are
in accordance with previous works which have described,
using monoclonal and polyclonal antisera, the presence of the
FGF-2 immunoreactivity in the nuclei of glial cells widely
distributed in the brain (Cintra et al. 1991; Woodward et al.
1992; Chadi et al. 1994). Previous papers have shown that low
molecular weight forms of the FGF-2 localize mainly in the
cytoplasm of neurons while higher forms are present in the
astrocytes nuclei through out brain and in neuronal nuclei of
few brain regions like pyramidal layer of the hippocampus
(Florkiewicz et al. 1991). Thus the data of the present paper
may refer to the changes of different molecular weight forms
of the molecule in distinct cell population in the NTS.
Few studies have investigated the role of growth factors
in the areas of blood pressure control. The demonstration of
the receptor for neurotrophic factor in the neurons of the
NTS (Mufson et al. 1992) suggests that those molecules
may play actions in the cardiovascular centers.
Different experimental designs have been used to study
the role of neurotrophic factors in the central nervous
Fig. 3 Double
immunoperoxidase procedures
have been used to colocalize
FGF-2 and glial fibrilary acidic
protein (GFAP, marker for
astrocytes) immunoreactivities
in the nucleus tractus solitarii
(NTS) of the 72 h aortic
coarctated rat (A–C). FGF-2
and GFAP immunoreactivities
are identified as a brownish
colour (diaminobenzidine as a
chromogen) and a bluish colour
(4-chloro naphtol as a
chromogen), respectively. B and
C represent higher
magnification of areas depicted
from the NTS showed in A. The
arrows indicate the presence of
FGF-2 immunoreactivity in the
nuclei of astrocytes and the
open arrows show the presence
of FGF-2 immunoreactivity in
the cytoplasm of neurons of the
NTS (B, C). (he panels D and Eshow GFAP immunoreactive
astrocytes (arrows)
counterstained with cresil violet
and OX42 immunoreactive
microglias (arrows) in the NTS
of control rats. Bars = 100 lm
(A), 20 lm (B), 10 lm (C, D,E)
290 J Mol Hist (2007) 38:285–294
123
system. Exogenous administration of growth factors to the
brain (Gomez-Pinilla et al. 1992; Otto and Unsicker 1990;
Chadi et al. 1993a; Koketsu et al. 1994), neuronal lesions
(Finklestein et al. 1988; Park and Mytilineou 1992; Frim
et al. 1993; Chadi et al. 1994) and electrical stimulation
(Follesa et al. 1994) applied to the central neuronal path-
ways are commonly employed.
Lesions to the central nervous system have been described
to induce a strong expression of FGF-2 mRNA and protein in
activated astroglial cells in the area of the injury (Finklestein
et al. 1988; Gomez-Pinilla et al. 1990; Logan et al. 1992;
Chadi et al. 1994). Riva and collaborators (1992) as well as
Follesa and collaborators (1994) described increased levels
of the FGF-2 mRNA following focally-evoked convulsive
seizures. Although an increasing number of studies have
pointed out the role of FGF-2 following cellular lesion, few
works have attempted to investigate the regulation of FGF-2
in response to physiological alterations of neuronal activity.
The present work has employed the coarctation hyper-
tension approach to evaluate whether a physiological
increase of the neuronal activity can modify the FGF-2
immunoreactivity in the NTS of the rat. It is well known
that during 48–72 h following the coarctation, the NTS is
highly stimulated by the intense afferent activity which
determines an increased vagal tone and an intense brady-
cardia at 3–6 h after coarctation (Soato and Krieger 1974;
Michelini et al. 1992).
FOS immunoreactivity has been considered a valuable
tool in the demonstration of activated neurons after stim-
ulus (Erickson and Millhorn 1991). The presence of FOS
positive neurons in the NTS of aortic coarctated rats 4 h
after surgery confirms the activation of that nucleus after
acute hypertension.
Our study has demonstrated an increased FGF-2
immunoreactivity, which may reflect an increased synthe-
sis of the FGF-2 protein, in neurons and astrocytes in the
NTS following the coarctation hypertension. It seems
likely that the changes of the FGF-2 immunoreactivity in
the NTS were probably induced by the increase in the
neuronal activity within this nucleus of the coarctated rats.
Several studies reported that physiological levels of affer-
ent activity are capable of regulating the synthesis of
neurotrophic factors (Isackson 1995). Gall and Isackson
(1989) observed an increased nerve growth factor mRNA
in response to afferent activity following limbic seizures.
Furthermore, experiments involving sub-seizure stimula-
tion (Castren et al. 1993) and physical exercise (Neeper
et al. 1995) have been shown to increase neurotrophins
expression in hippocampus. It was also demonstrated that
light regulates expression of brain derived neurotrophic
factor mRNA in rat visual cortex (Castren et al. 1992).
Moreover, long-term environmental enrichment leads to
regional increases in neurotrophin levels in the rat brain
(Ickes et al. 2000). Expression of neurotrophic factors in
brain activated areas may regulate the necessary synaptic
plasticity phenomena that in turn modulate local synaptic
transmittion (Bramham and Messaoudi 2005). It should be
considered the analysis of the FGF-2 mRNA expression
after aortic coarctation hypertension to confirm the changes
at the protein level, however, one should be aware that the
level and the timing of the FGF-2 gene expression may not
correspond to that found after neuronal lesion (Chadi et al.
1994).
It is possible that the increased neuronal activity induced
by the coarctation hypertension promoted a long lasting
increase in the neuronal FGF-2 immunoreactivity. The
Fig. 4 Microphotographs of coronal sections showing FOS immu-
noreactive neurons in the nucleus tractus solitarii (NTS) of rats 4 h
after aortic coarctation-induced baroreceptor reflex activation (A, B).
B represents a higher magnification of a NTS region showed in A.
Only scatted neuronal cell profiles showed FOS immunoreactivity in
the NTS of sham-operated (control) rats (C). Bars = 100 lm (A, C)
and 20 lm (B)
J Mol Hist (2007) 38:285–294 291
123

neuronal FGF-2 might modulate autocrine trophic and
plastic actions in the NTS neurons. Furthermore, the tran-
sient enhancement of the glial FGF-2 immunoreactivity in
the NTS could be due to the neuronal-glial interaction in
the NTS following the coarctation hypertension in a para-
crine fashion.
In our experiments slight activation of astrocytes was
observed at 72 h time-interval, following the aortic coarcta-
tion, as seen by the increased GFAP immunoreactivity.
Previous studies have demonstrated that a physiological
increased neuronal activity can activate brain astrocytes.
Repeated but kindled seizures applied in one side of the
hippocampus resulted in bilateral increase of the GFAP
mRNA synthesis (Steward et al. 1991). Moreover, short
period of spontaneous physical exercise is sufficient to induce
astroglial activation in rat hippocampus (Gomez-Pinilla et al.
1997). In the present work, it is possible that the coarctation
hypertension has led to a slight and short-lasting activation of
NTS astrocytes to transiently synthesize glial FGF-2 which
may act as a paracrine factor to NTS neurons. It has been
described the role of astroglial FGF-2 in the trophic state of
neurons of several parts of the brain (Engele and Bohn 1991;
Chadi et al. 1994).
The signals triggering the changes in the trophic/plastic
state in the NTS after coarctation hypertension were not
identified. The proto oncogene c-fos was suggested to
participate in the regulation of neurotrophic factor (Morgan
and Curran 1991) and was described to be expressed in the
NTS in response to stimulation of afferent fibres (Erickson
and Millhorn 1991; Rutherfurd et al. 1992) which is in line
with present observation of FOS expression in the NTS
neurons 4 h after aortic coarctation. It should be mentioned
that the majority of NTS neurons of stimulated rats in the
present analysis expressed FOS protein which indicates
that the up-regulation of the FGF-2 protein may have
occurred in those cells.
Recent studies have indicated that the excitatory amino
acid glutamate is the neurotransmitter of the baroreceptor
primary afferent fibres (Guyenet et al. 1987; Pawloski-Dahm
and Gordon 1992). Lawrence and Jarrot (1994) observed
increases of glutamate in microdialysis lisates in the NTS of
phenylephrine-induced hypertensive rats. Moreover, previous
work has demonstrated that low doses of glutamate induce
FGF-2 mRNA in neurons and astrocytes in culture (Pechan
et al. 1993). Thus, increased FGF-2 immunoreactivity
observed in the stimulated NTS may be due to the increased
release of glutamate from the baroreceptor terminals which, in
turn, may have stimulated the FGF-2 synthesis in the NTS
cells, underlining the role of the neurotrophic factor in events
related to neuroplasticity in the central areas of blood pressure
control.
Furthermore, it is known that the glutamate can become
neurotoxic by persistently NMDA receptor activation
(Novelli et al. 1988). In this way the synthesis and local
release of the FGF-2 following coarctation hypertension
might be able to protect the NTS neurons from excitotox-
icity (Cheng et al. 1993; Mattson et al. 1993).
However, it is unlikely that neurotoxicity may have
taken place following coarctation hypertension since we
have not observed any alteration in the OX42 immunore-
activity 1 week and 1 month following the aortic coarcta-
tion. Furthermore, it is postulated that the degree of
degeneration of the injured neurons is related to the
intensity of the microglial reaction (Giulian and Robertson
1990; Giulian 1993). Moreover, we have not found changes
in the number of NTS neurons up to 1 month of aortic
coarctation.
Thus, there is no indication of neuronal lesion of stimu-
lated NTS in the present model and the upregulated FGF-2
may be involved in local neuronal plasticity (Isackson 1995)
and changes in neurotransmitter phenotype (Otto and
Unsicker 1990) may represent a trophic adaptive response in
this nucleus.
In conclusion the presence of the FGF-2 immunoreactivity
in the neurons and astrocytes of the NTS indicates that the
FGF-2 may be an important neurotrophic factor in the areas
involved in blood pressure control. The increased FGF-2
immunoreactivity in the NTS following an increased neu-
ronal activity induced by the coarctation hypertension may
represent a neurotrophic/neuroplastic adaptive response in
this nucleus.
Acknowledgements This work was supported by grants from
Fundacao de Amparo a Pesquisa do Estado de Sao Paulo (FAPESP
98/13122-5; 99/01319-1) and Conselho Nacional para o Desen-
volvimento Cientıfico e Tecnologico (CNPq: 521004/97-1; 311797/
2006-7), Brasil. We are grateful to Professor Kjell Fuxe, Karolinska
Institute, Stockholm, Sweden, for valuable discussion and to Dr.
Andrew Baird Whittier Institute, La Jolla, USA for the generous gift
of the FGF-2 antibody. Varella, T.C.N. was a FAPESP undergraduate
trainee fellow (96/9387-8).
References
Anwar A, Delafontaine P (1994) Hypertension increases insulin-like
growth factor binding protein-4 mRNA levels in rat aorta.
Hypertension 24:679–685
Baird A, Klagsbrun M (1991) The fibroblast growth factor family: An
overview. In: Baird A, Klagsbrun M (Eds) The fibroblast growth
factor family: Ann N Y Acad Sci pp xi
Bramham CR, Messaoudi E (2005) BDNF function in adult synaptic
plasticity: the synaptic consolidation hypothesis. Prog Neurobiol
76:99–125
Castren E, Pitkanen M, Sirvio J, Parsadanian A, Lindholm D,
Thoenen H, Riekkinen PJ (1993) The induction of LTP increases
BDNF and NGF mRNA but decreases NT-3 mRNA in the
dentate gyrus. Neuroreport 4:895–898
Castren E, Zafra F, Thoenen H, Lindholm D (1992) Light regulates
expression of brain-derived neurotrophic factor mRNA in rat
visual cortex. Proc Natl Acad Sci USA 89:9444–9448
292 J Mol Hist (2007) 38:285–294
123

Chadi G, Cao Y, Pettersson RF, Fuxe K (1994) Temporal and spatial
increase of astroglial basic fibroblast growth factor synthesis
after 6-hydroxydopamine-induced degeneration of the nigrostri-
atal dopamine neurons. Neuroscience 61:891–910
Chadi G, Moller A, Rosen L, Janson AM, Agnati LA, Goldstein M,
Ogren SO, Pettersson RF, Fuxe K (1993a) Protective actions of
human recombinant basic fibroblast growth factor on MPTP-
lesioned nigrostriatal dopamine neurons after intraventricular
infusion. Exp Brain Res 97:145–158
Chadi G, Rosen L, Cintra A, Tinner B, Zoli M, Pettersson RF, Fuxe K
(1993b) Corticosterone increases FGF-2 (bFGF) immunoreac-
tivity in the substantia nigra of the rat. Neuroreport 4:783–786
Chapleau MW, Cunningham JT, Sullivan MJ, Wachtel RE, Abboud
FM (1995) Structural versus functional modulation of the arterial
baroreflex. Hypertension 26:341–347
Cheng B, McMahon DG, Mattson MP (1993) Modulation of calcium
current, intracellular calcium levels and cell survival by glucose
deprivation and growth factors in hippocampal neurons. Brain
Res 607:275–285
Cintra A, Cao YH, Oellig C, Tinner B, Bortolotti F, Goldstein M,
Pettersson RF, Fuxe K (1991) Basic FGF is present in
dopaminergic neurons of the ventral midbrain of the rat.
Neuroreport 2:597–600
Ciriello J, Calaresu FR (1981) Projections from buffer nerves to the
nucleus of the solitary tract: an anatomical and electrophysio-
logical study in the cat. J Auton Nerv Syst 3:299–310
Cravo SL, Morrison SF, Reis DJ (1991) Differentiation of two
cardiovascular regions within caudal ventrolateral medulla. Am J
Physiol 261:R985–994
Engele J, Bohn MC (1991) The neurotrophic effects of fibroblast
growth factors on dopaminergic neurons in vitro are mediated by
mesencephalic glia. J Neurosci 11:3070–3078
Erickson JT, Millhorn DE (1991) Fos-like protein is induced in
neurons of the medulla oblongata after stimulation of the
carotid sinus nerve in awake and anesthetized rats. Brain Res
567:11–24
Finklestein SP, Apostolides PJ, Caday CG, Prosser J, Philips MF,
Klagsbrun M (1988) Increased basic fibroblast growth factor
(bFGF) immunoreactivity at the site of focal brain wounds. Brain
Res 460:253–259
Florkiewicz RZ, Baird A, Gonzalez AM (1991) Multiple forms of
bFGF: differential nuclear and cell surface localization. Growth
Factors 4:265–275
Follesa P, Gale K, Mocchetti I (1994) Regional and temporal pattern
of expression of nerve growth factor and basic fibroblast growth
factor mRNA in rat brain following electroconvulsive shock.
Exp Neurol 127:37–44
Frim DM, Uhler TA, Short MP, Ezzedine ZD, Klagsbrun M,
Breakefield XO, Isacson O (1993) Effects of biologically
delivered NGF, BDNF and bFGF on striatal excitotoxic lesions.
Neuroreport 4:367–370
Gall CM, Isackson PJ (1989) Limbic seizures increase neuronal
production of messenger RNA for nerve growth factor. Science
245:758–761
Giulian D (1993) Reactive glia as rivals in regulating neuronal
survival. Glia 7:102–110
Giulian D, Robertson C (1990) Inhibition of mononuclear phagocytes
reduces ischemic injury in the spinal cord. Ann Neurol 27:33–42
Giuliano R, Brezenoff HE (1987) Increased central cholinergic
activity in rat models of hypertension. J Cardiovasc Pharmacol
10:113–122
Gomez-Pinilla F, Lee JW, Cotman CW (1992) Basic FGF in adult rat
brain: cellular distribution and response to entorhinal lesion and
fimbria-fornix transection. J Neurosci 12:345–355
Gomez-Pinilla F, Dao L, So V (1997) Physical exercise induces FGF-
2 and its mRNA in the hippocampus. Brain Res 764:1–8
Gonzalez AM, Buscaglia M, Ong M, Baird A (1990) Distribution of
basic fibroblast growth factor in the 18-day rat fetus: localization
in the basement membranes of diverse tissues. J Cell Biol
110:753–765
Gospodarowicz D (1990) Fibroblast growth factor and its involve-
ment in developmental processes. Curr Top Dev Biol 24:57–93
Gospodarowicz D, Moran J, Braun D, Birdwell C (1976) Clonal
growth of bovine vascular endothelial cells: fibroblast growth
factor as a survival agent. Proc Natl Acad Sci USA 73:4120–
4124
Grothe C, Zachmann K, Unsicker K (1991) Basic FGF-like immu-
noreactivity in the developing and adult rat brainstem. J Comp
Neurol 305:328–336
Gundersen HJ, Jensen EB (1987) The efficiency of systematic
sampling in stereology and its prediction. J Microsc 147:229–263
Guyenet PG, Filtz TM, Donaldson SR (1987) Role of excitatory
amino acids in rat vagal and sympathetic baroreflexes. Brain Res
407:272–284
Hollander M, Wolfe DA (1973) Non-parametric statistical methods.
Wiley, New York
Hsu SM, Raine L, Fanger H (1981) Use of avidin-biotin-peroxidase
complex (ABC) in immunoperoxidase techniques: a comparison
between ABC and unlabeled antibody (PAP) procedures.
J Histochem Cytochem 29:577–580
Ickes BR, Pham TM, Sanders LA, Albeck DS, Mohammed AH,
Granholm AC (2000) Long-term environmental enrichment
leads to regional increases in neurotrophin levels in rat brain.
Exp Neurol 164:45–52
Isackson PJ (1995) Trophic factor response to neuronal stimuli or
injury. Curr Opin Neurobiol 5:350–357
Janson AM, Moller A (1993) Chronic nicotine treatment counteracts
nigral cell loss induced by a partial mesodiencephalic hemitran-
section: an analysis of the total number and mean volume of
neurons and glia in substantia nigra of the male rat. Neuroscience
57:931–941
Koketsu N, Berlove DJ, Moskowitz MA, Kowall NW, Caday CG,
Finklestein SP (1994) Pretreatment with intraventricular basic
fibroblast growth factor decreases infarct size following focal
cerebral ischemia in rats. Ann Neurol 35:451–457
Krieger EM (1970) Time course of baroreceptor resetting in acute
hypertension. Am J Physiol 218:486–490
Kushima Y, Nishio C, Nonomura T, Hatanaka H (1992) Effects of
nerve growth factor and basic fibroblast growth factor on
survival of cultured septal cholinergic neurons from adult rats.
Brain Res 598:264–270
Langhorst P, Lambertz M, Kluge W, Rittweger J (1992) Different
modes of dampening influence from baroreceptors are deter-
mined by the functional organization of the NTS neuronal
network. J Auton Nerv Syst 41:141–156
Lawrence AJ, Jarrott B (1994) L-glutamate as a neurotransmitter at
baroreceptor afferents: evidence from in vivo microdialysis.
Neuroscience 58:585–591
Li YW, Dampney RA (1992) Expression of c-fos protein in the
medulla oblongata of conscious rabbits in response to barore-
ceptor activation. Neurosci Lett 144:70–74
Li YW, Dampney RA (1994) Expression of Fos-like protein in brain
following sustained hypertension and hypotension in conscious
rabbits. Neuroscience 61:613–634
Lin TN, Wong YP, Chen JJ, Cheng JT, Yu SF, Sun SH, Chi SI, Chai
CY (1997) Elevated basic fibroblast growth factor levels in
stroke-prone spontaneously hypertensive rats. Neuroscience
76:557–570
Ling EA, Kaur LC, Yick TY, Wong WC (1990) Immunocytochem-
ical localization of CR3 complement receptors with OX-42 in
amoeboid microglia in postnatal rats. Anat Embryol (Berl)
182:481–486
J Mol Hist (2007) 38:285–294 293
123

Loewy AD, Burton H (1978) Nuclei of the solitary tract: efferent
projections to the lower brain stem and spinal cord of the cat.
J Comp Neurol 181:421–449
Logan A, Frautschy SA, Gonzalez AM, Baird A (1992) A time course
for the focal elevation of synthesis of basic fibroblast growth
factor and one of its high-affinity receptors (flg) following a
localized cortical brain injury. J Neurosci 12:3828–3837
Matsuyama A, Iwata H, Okumura N, Yoshida S, Imaizumi K, Lee Y,
Shiraishi S, Shiosaka S (1992) Localization of basic fibroblast
growth factor-like immunoreactivity in the rat brain. Brain Res
587:49–65
Mattson MP, Zhang Y, Bose S (1993) Growth factors prevent
mitochondrial dysfunction, loss of calcium homeostasis, and cell
injury, but not ATP depletion in hippocampal neurons deprived
of glucose. Exp Neurol 121:1–13
Michelini LC, de Oliveira M, dos Santos M (1992) Baroreceptor
reflex control of heart rate during development of coarctation
hypertension. Hypertension 19:II159–163
Mifflin SW, Spyer KM, Withington-Wray DJ (1988) Baroreceptor
inputs to the nucleus tractus solitarius in the cat: postsynaptic
actions and the influence of respiration. J Physiol 399:349–367
Morgan JI, Curran T (1991) Proto-oncogene transcription factors and
epilepsy. Trends Pharmacol Sci 12:343–349
Mufson EJ, Brashers-Krug T, Kordower JH (1992) p75 nerve growth
factor receptor immunoreactivity in the human brainstem and
spinal cord. Brain Res 589:115–123
Neeper SA, Gomez-Pinilla F, Choi J, Cotman C (1995) Exercise and
brain neurotrophins. Nature 373:109
Nosaka S, Yamamoto T, Yasunaga K (1979) Localization of vagal
cardioinhibitory preganglionic neurons with rat brain stem.
J Comp Neurol 186:79–92
Novelli A, Reilly JA, Lysko PG, Henneberry RC (1988) Glutamate
becomes neurotoxic via the N-methyl-D-aspartate receptor when
intracellular energy levels are reduced. Brain Res 451:205–212
Nozaki K, Finklestein SP, Beal MF (1993) Basic fibroblast growth
factor protects against hypoxia-ischemia and NMDA neurotox-
icity in neonatal rats. J Cereb Blood Flow Metab 13:221–228
Otto D, Unsicker K (1990) Basic FGF reverses chemical and
morphological deficits in the nigrostriatal system of MPTP-
treated mice. J Neurosci 10:1912–1921
Pakkenberg B, Gundersen HJ (1988) Total number of neurons and
glial cells in human brain nuclei estimated by the disector and
the fractionator. J Microsc 150:1–20
Park TH, Mytilineou C (1992) Protection from 1-methyl-4-phen-
ylpyridinium (MPP+) toxicity and stimulation of regrowth
of MPP(+)-damaged dopaminergic fibers by treatment of
mesencephalic cultures with EGF and basic FGF. Brain Res
599:83–97
Pawloski-Dahm C, Gordon FJ (1992) Evidence for a kynurenate-
insensitive glutamate receptor in nucleus tractus solitarii. Am J
Physiol 262:H1611–1615
Paxinos G, Watson C (1986) The rat brain: in stereotaxic coordinates.
Harcourt Brace Jovanovich, San Diego
Pechan PA, Chowdhury K, Gerdes W, Seifert W (1993) Glutamate
induces the growth factors NGF, bFGF, the receptor FGF-R1 and
c-fos mRNA expression in rat astrocyte culture. Neurosci Lett
153:111–114
Puumala M, Anderson RE, Meyer FB (1990) Intraventricular infusion
of HBGF-2 promotes cerebral angiogenesis in Wistar rat. Brain
Res 534:283–286
Riva MA, Gale K, Mocchetti I (1992) Basic fibroblast growth factor
mRNA increases in specific brain regions following convulsive
seizures. Brain Res Mol Brain Res 15:311–318
Rogers RF, Paton JF, Schwaber JS (1993) NTS neuronal responses to
arterial pressure and pressure changes in the rat. Am J Physiol
265:R1355–1368
Ross CA, Ruggiero DA, Reis DJ (1985) Projections from the nucleus
tractus solitarii to the rostral ventrolateral medulla. J Comp
Neurol 242:511–534
Rutherfurd SD, Widdop RE, Sannajust F, Louis WJ, Gundlach AL
(1992) Expression of c-fos and NGFI-A messenger RNA in the
medulla oblongata of the anaesthetized rat following stimulation
of vagal and cardiovascular afferents. Brain Res Mol Brain Res
13:301–312
Saltis J, Agrotis A, Bobik A (1993) Differences in growth charac-
teristics of vascular smooth muscle from spontaneously hyper-
tensive and Wistar-Kyoto rats are growth factor dependent.
J Hypertens 11:629–637
Soato GG, Krieger EM (1974) Heart rate after acute hypertension in
the rat. Am J Physiol 227:1389–1393
Spyer KM (1981) Neural organisation and control of the baroreceptor
reflex. Rev Physiol Biochem Pharmacol 88:24–124
Steward O, Torre ER, Tomasulo R, Lothman E (1991) Neuronal
activity up-regulates astroglial gene expression. Proc Natl Acad
Sci USA 88:6819–6823
Walicke PA (1988) Basic and acidic fibroblast growth factors have
trophic effects on neurons from multiple CNS regions.
J Neurosci 8:2618–2627
West MJ, Gundersen HJ (1990) Unbiased stereological estimation of
the number of neurons in the human hippocampus. J Comp
Neurol 296:1–22
West MJ, Slomianka L, Gundersen HJ (1991) Unbiased stereological
estimation of the total number of neurons in thesubdivisions of
the rat hippocampus using the optical fractionator. Anat Rec
231:482–497
Woodward WR, Nishi R, Meshul CK, Williams TE, Coulombe M,
Eckenstein FP (1992) Nuclear and cytoplasmic localization of
basic fibroblast growth factor in astrocytes and CA2 hippocam-
pal neurons. J Neurosci 12:142–152
Zamboni I, De Martino C (1969) Buffered picric acid formaldehyde: a
new rapid fixative for electron microscopy. J Cell Biol 35:148A
Zettler C, Rush RA (1993) Elevated concentrations of nerve growth
factor in heart and mesenteric arteries of spontaneously hyper-
tensive rats. Brain Res 614:15–20
Zoli M, Pich EM, Cimino M, Lombardelli G, Peruzzi G, Fuxe K,
Agnati LF, Cattabeni F (1990) Morphometrical and microdensi-
tometrical studies on peptide- and tyrosine hydroxylase-like
immunoreactivities in the forebrain of rats prenatally exposed to
methylazoxymethanol acetate. Brain Res Dev Brain Res 51:
45–61
294 J Mol Hist (2007) 38:285–294
123
Related Documents











