Antipsychotic Treatment of Acute Paranoid Schizophrenia Patients with Olanzapine Results in Altered Glycosylation of Serum Glycoproteins Jayne E. Telford, † Jonathan Bones, † Ciara McManus, † Radka Saldova, † Gwen Manning, ‡ Margaret Doherty, † F. Markus Leweke, §,∥ Matthias Rothermundt, ⊥ Paul C. Guest, # Hassan Rahmoune, # Sabine Bahn, # and Pauline M. Rudd* ,† † NIBRT Dublin-Oxford Glycobiology Laboratory, National Institute for Bioprocessing Research and Training, Foster Avenue, Mount Merrion, Co., Dublin, Ireland ‡ Conway Institute, University College Dublin, Belfield, Dublin 4, Ireland § Department of Psychiatry and Psychotherapy, Central Institute of Mental Health, Medical Faculty Mannheim, Heidelberg University, Mannheim, Germany ∥ Department of Psychiatry and Psychotherapy, University of Cologne, Cologne, Germany ⊥ Department of Psychiatry, University of Muenster, Muenster, Germany # Department of Chemical Engineering and Biotechnology, University of Cambridge, Tennis Court Road, Cambridge, United Kingdom * S Supporting Information ABSTRACT: Atypical antipsychotic drugs, such as olanza- pine, have been shown to alleviate the positive, negative and, to a lesser degree, the cognitive symptoms of schizophrenia in many patients. However, the detailed mechanisms of action of these drugs have yet to be elucidated. We have carried out the first investigation aimed at evaluating the effects of olanzapine treatment on the glycosylation of serum proteins in schizophrenia patients. Olanzapine treatment resulted in increased levels of a disialylated biantennary glycan and reduced levels of a number of disialylated bi- and triantennary glycans on whole serum glycoproteins. These changes were not observed on a low-abundance serum protein fraction. α1 acid glycoprotein was identified as a carrier of some of the detected altered oligosaccharides. In addition, glycan analysis of haptoglobin, transferrin, and α1 antitrypsin reported similar findings, although these changes did not reach significance. Exoglycosidase digestion analysis showed that olanzapine treatment increased galactosylation and sialylation of whole serum proteins, suggesting increased activity of specific galactosyltransferases and increased availability of galactose residues for sialylation. Taken together, these findings indicate that olanzapine treatment results in altered glycosylation of serum proteins. KEYWORDS: glycosylation, schizophrenia, olanzapine, α1 acid glycoprotein ■ INTRODUCTION Schizophrenia is a complex debilitating neuropsychiatric disorder that affects 1% of the population and is manifested by positive symptoms such as hallucinations and delusions, negative symptoms including apathy and loss of motivation, and cognitive symptoms such as memory impairment. The pathophysiology of the disease is not understood. However, aberrant dopaminergic and glutamatergic neurotransmission are thought to be associated with the disease, along with neurodevelopmental alterations. To gain a deeper under- standing of the physiology of the disease and to identify disease biomarkers, numerous proteomic profiling studies of serum, plasma, cerebrospinal fluid (CSF) and urine from schizophrenic patients and many disease-specific proteomic differences have been performed. Decreased apolipoprotein A1 levels were found in serum, brain, CSF and liver from schizophrenic patients. 1 Levin et al. reported that the concentrations of 10 proteins, including transferrin, α2-HS glycoprotein and a number of apolipoproteins, were altered in serum from schizophrenic patients compared to healthy controls. 2 Other studies have shown that acute phase reactants including haptoglobin, hemopexin, fibrinogen, complement component 3 and 4, α1 acid glycoprotein (AGP) and α1 Received: March 6, 2012 Published: May 17, 2012 Article pubs.acs.org/jpr © 2012 American Chemical Society 3743 dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−3752

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Antipsychotic Treatment of Acute Paranoid Schizophrenia Patientswith Olanzapine Results in Altered Glycosylation of SerumGlycoproteinsJayne E. Telford,† Jonathan Bones,† Ciara McManus,† Radka Saldova,† Gwen Manning,‡
Margaret Doherty,† F. Markus Leweke,§,∥ Matthias Rothermundt,⊥ Paul C. Guest,# Hassan Rahmoune,#
Sabine Bahn,# and Pauline M. Rudd*,†
†NIBRT Dublin-Oxford Glycobiology Laboratory, National Institute for Bioprocessing Research and Training, Foster Avenue, MountMerrion, Co., Dublin, Ireland‡Conway Institute, University College Dublin, Belfield, Dublin 4, Ireland§Department of Psychiatry and Psychotherapy, Central Institute of Mental Health, Medical Faculty Mannheim, HeidelbergUniversity, Mannheim, Germany∥Department of Psychiatry and Psychotherapy, University of Cologne, Cologne, Germany⊥Department of Psychiatry, University of Muenster, Muenster, Germany#Department of Chemical Engineering and Biotechnology, University of Cambridge, Tennis Court Road, Cambridge, UnitedKingdom
*S Supporting Information
ABSTRACT: Atypical antipsychotic drugs, such as olanza-pine, have been shown to alleviate the positive, negative and,to a lesser degree, the cognitive symptoms of schizophrenia inmany patients. However, the detailed mechanisms of action ofthese drugs have yet to be elucidated. We have carried out thefirst investigation aimed at evaluating the effects of olanzapinetreatment on the glycosylation of serum proteins inschizophrenia patients. Olanzapine treatment resulted inincreased levels of a disialylated biantennary glycan andreduced levels of a number of disialylated bi- and triantennaryglycans on whole serum glycoproteins. These changes werenot observed on a low-abundance serum protein fraction. α1acid glycoprotein was identified as a carrier of some of the detected altered oligosaccharides. In addition, glycan analysis ofhaptoglobin, transferrin, and α1 antitrypsin reported similar findings, although these changes did not reach significance.Exoglycosidase digestion analysis showed that olanzapine treatment increased galactosylation and sialylation of whole serumproteins, suggesting increased activity of specific galactosyltransferases and increased availability of galactose residues forsialylation. Taken together, these findings indicate that olanzapine treatment results in altered glycosylation of serum proteins.
KEYWORDS: glycosylation, schizophrenia, olanzapine, α1 acid glycoprotein
■ INTRODUCTIONSchizophrenia is a complex debilitating neuropsychiatricdisorder that affects 1% of the population and is manifestedby positive symptoms such as hallucinations and delusions,negative symptoms including apathy and loss of motivation,and cognitive symptoms such as memory impairment. Thepathophysiology of the disease is not understood. However,aberrant dopaminergic and glutamatergic neurotransmission arethought to be associated with the disease, along withneurodevelopmental alterations. To gain a deeper under-standing of the physiology of the disease and to identifydisease biomarkers, numerous proteomic profiling studies ofserum, plasma, cerebrospinal fluid (CSF) and urine from
schizophrenic patients and many disease-specific proteomicdifferences have been performed. Decreased apolipoprotein A1levels were found in serum, brain, CSF and liver fromschizophrenic patients.1 Levin et al. reported that theconcentrations of 10 proteins, including transferrin, α2-HSglycoprotein and a number of apolipoproteins, were altered inserum from schizophrenic patients compared to healthycontrols.2 Other studies have shown that acute phase reactantsincluding haptoglobin, hemopexin, fibrinogen, complementcomponent 3 and 4, α1 acid glycoprotein (AGP) and α1
Received: March 6, 2012Published: May 17, 2012
Article
pubs.acs.org/jpr
© 2012 American Chemical Society 3743 dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−3752

antitrypsin were increased in plasma from schizophrenicpatients, as determined by laser nephelometry3 and matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry analysis.4 The concentration of acutephase proteins increases dramatically in inflammation, andthere are many reports of increased inflammation inschizophrenia.5−8
Studies focusing on alterations in post-translational mod-ifications, such as glycosylation, have also been reported inschizophrenia.9−12 Glycosylation is the most abundant post-translational modification and plays crucial roles in cellularprocesses, such as cell signaling, cellular recognition processesand protein folding. Oligosaccharides, synthesized in the roughendoplasmic reticulum and in the Golgi apparatus through theaction of glycosidases and glycosyltransferases, are highlyheterogeneous structures that can have variable lengths andcomplexities. Glycan changes, including altered levels ofA3F1G3S3 and A4G4LacS4, have previously been found inserum samples from schizophrenia patients when compared tohealthy subjects and may therefore have potential as biomarkersfor the disease.9 However, there are no studies to date thatinvestigate the effects of antipsychotic drug treatment on theglycosylation of serum proteins in schizophrenia. A recent studyfrom our laboratory highlighted the importance of consideringthe effects of medication on glycosylation with glycan changesreported as a result of treatment with nonsteroidal anti-inflammatory drugs and oral contraceptives.13
Atypical antipsychotics such as olanzapine are widely used inthe treatment of schizophrenia. Olanzapine has been shown tohave beneficial effects on the positive, negative and, to a lesserextent, the cognitive symptoms of schizophrenia in manypatients.14−16 Previous studies suggest that olanzapine hasenhanced therapeutic properties when compared with otherantipsychotic treatments. Olanzapine was more effective atreducing positive and negative symptoms, depression andagitation compared to typical antipsychotics such as haloper-idol.17 A more recent study showed that olanzapine treatmentresulted in greater improvements in the general mental state ofpatients compared with other atypical drugs such aripiprazole,risperidone, ziprasidone and quetiapine.18 Alvarez et al. foundthat olanzapine reduced negative symptoms more effectivelythan risperidone in schizophrenia patients.19 The mode ofaction of olanzapine is not completely understood. However, itis known to bind with high affinity to serotonin, dopamine,histamine and adrenergic α1 receptors.20,21 After dosing,olanzapine is extensively distributed throughout the body andshows 93% binding to plasma proteins, primarily albumin andAGP.22 Studies have been carried out on the effects ofolanzapine on gene expression23,24 and on protein levels,25,26
but the effects of olanzapine on post-translational modificationssuch as glycosylation have not been investigated to date.In the present study, we have investigated the glycosylation
profile of glycoproteins in serum from acute paranoidschizophrenia patients before and after 6 weeks of treatmentwith olanzapine by using high-throughput hydrophilic inter-action chromatography (HILIC)-based glycoanalytical technol-ogy. Subsequently, we attempted to identify the proteinsbearing the identified changes. In addition, we usedexoglycosidase digestion analyses and weak anion exchangechromatography to determine which stages of the glycanprocessing pathway were potentially affected.
■ EXPERIMENTAL SECTIONSample Selection and Collection
Blood samples were collected from acute paranoid schizo-phrenia patients (pretreatment) and after 6 weeks of treatmentwith olanzapine (post-treatment) at the Department ofPsychiatry and Psychotherapy of the University of Cologneunder protocols for sample collection and analysis approved bythe Ethics Committee of the University of Cologne. Informedconsent was provided in writing by all participants, and clinicalinvestigations were conducted according to the principlesexpressed in the Declaration of Helsinki. Diagnosis wasdetermined according to the Diagnostic and Statistical Manualof Mental Disorders, fourth edition (DSM-IV) and theInternational Statistical Classification of Diseases and RelatedHealth Problems 10th Revision (ICD 10) by experiencedpsychiatrists using the Structured Clinical Interview for DSM-IV (SCID). The subjects included 23 antipsychotic-naivepatients diagnosed with acute paranoid schizophrenia (DSM-IV 295.3). The group consisted of 7 females and 16 males withan age of 30.3 ± 10.5 years and body mass index (BMI) of 22.6± 3.1 kg/m2, and included 6 smokers and 17 nonsmokers.Blood samples were obtained by venous puncture and collectedinto S-Monovette 7.5 mL serum tubes (Sarstedt; Numbrecht,Germany). The blood was kept at room temperature for 2 h toallow a consistent time for clot formation, followed bycentrifugation at 4000g for 5 min to pellet clotted materialand other cellular debris. The supernatants (sera) werecollected and stored at −80 °C in Low Binding Eppendorftubes (Hamburg, Germany) until analysis.
Serum Depletion with MARS-14 Column
Serum samples were depleted of the 14 most abundant proteins(albumin, IgG, transferrin, fibrinogen, IgA, haptoglobin, α1antitrypsin, α2 macroglobulin, IgM, apolipoprotein AI, AGP,complement C3, apolipoprotein AII and transthyretin) usingthe Multiple Affinity Removal System (MARS) Human 14(Agilent Technologies; Wilmington, DE, USA). Samples (40μL) were diluted with 120 μL of MARS Buffer A andcentrifuged at 16000g for 1 min through a 0.22 μm spin filter.The filtered samples were injected onto the MARS-14 columnaccording to the manufacturer’s instructions using a 1200 LCsystem (Agilent Technologies). The unbound (depleted)serum fractions were collected and stored at −80 °C.Preparation and Analysis of N-Glycans
N-Glycans were released, derivatized and analyzed using high-throughput HILIC fluorescence-based glycoanalytical technol-ogy as described previously by Royle et al.27 Glycan structureswere assigned using GlycoBase (http://glycobase.nibrt.ie). Asrequired, specific exoglycosidase digestions were used in theassignment of glycan structures and in the further investigationof the effects of olanzapine on glycosylation as outlined inRoyle et al.27 Arthrobacter ureafaciens sialidase (ABS), bovinetestes β-galactosidase (BTG), bovine kidney α-fucosidase(BKF), almond meal α-fucosidase (AMF), β-N-acetylglucosa-minidase cloned from Streptococcus pneumonia, expressed inEscherichia coli (GUH) were used in these experiments. Theglycan nomenclature applied is illustrated in Figure 1.28
Isolation of Proteins by Two-Dimensional GelElectrophoresis (2DE)
The protein concentrations of whole serum samples from 6patients before and after treatment with olanzapine weredetermined using the method of Bradford29 with bovine serum
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523744

albumin as a reference standard. 2DE was performed asdescribed in Bones et al.30 using pH 3−7 nonlinear (NL) forisoelectric focusing followed by sodium dodecylsulfatepolyacrylamide gel electrophoresis (SDS-PAGE) on 12% gels.The protein load was 1 mg in all instances. Protein spots werevisualized by Coomassie Brilliant Blue staining, and the spotswere manually excised from the gels. The migration positions ofthe proteins of interest were cross-validated with the SwissProt2D PAGE Human plasma reference gel to confirm proteinidentifications (http://world-2dpage.expasy.org/swiss-2dpage/). Glycans were released from proteins in the excised gel pieceswith PNGase F and were prepared for glycan analysis byHILIC-fluorescence as described above. Chromatographicpeaks were assigned with the same numbers as those in thewhole serum profile allowing comparisons to be made.
MALDI-TOF Mass Spectrometry Identification of IsolatedProteins
Following removal of glycans, the gel pieces were prepared forMALDI-TOF mass spectrometry (MS) analysis. In-geldigestion was carried out overnight by incubation with trypsin(Promega; Madison, WI, USA) at 37 °C. The supernatantswere harvested and further extraction was carried out bywashing the gel pieces with 70% acetonitrile containing 4%formic acid. The subsequent supernatants were added to thepreviously collected supernatants, dried down and resuspendedin 2 μL of 1% formic acid. The samples were analyzed using a4800 plus MALDI TOF/TOF Analyzer (Applied Biosystems;Foster City, CA, USA). Peptide masses were acquired over arange from 800 to 4000 m/z, with a focus mass of 2000 m/z.MS spectra were summed from 2500 shots using an Nd:YAGlaser (355 nm, 200 Hz). Automated plate calibration wasperformed using five peptide standards (masses 900−2400 m/z; Applied Biosystems) to update the instrument default masscalibration, which was applied to all MS and MS/MS spectra. Amaximum of 12 precursors per sample well with a signal-to-noise ratio of >20 were selected automatically for subsequentfragmentation by collision induced dissociation, and MS/MSspectra were summed from 4000 laser shots. Spectra wereprocessed and analyzed by the Global Protein ServerWorkstation (Version 3.6, Applied Biosystems) and searchedusing MASCOT (Matrix Science; London, U.K.) against theUniProt knowledgebase with the human taxonomic filterspecified (release 2011_01; http://ftp.ebi.ac.uk/pub/databases). The search parameters included a maximum ofone missed cleavage, a maximum of two variable post-translational modifications (oxidation of methionines anddeamidation of asparagines), a precursor tolerance of 100
Figure 1. The glycan symbols, linkages, and linkage positioninformation. The Oxford nomenclature was used in this paper torepresent glycan structures.
Figure 2. Olanzapine treatment significantly altered the relative percentage areas of HILIC peaks 16 and 20 from whole serum proteins but did notalter the relative percentage areas of peaks in the HILIC profile of low-abundance serum proteins in schizophrenia samples. Typical HILICchromatograms for (a) whole serum N-glycans and the assigned peaks (1−24) and (b) low-abundance serum protein N-glycans and the assignedpeaks. The relative percentage areas of peaks pretreatment and post-treatment are shown in black and gray, respectively, for whole serum (c) and forlow-abundance serum protein fraction (d). The glycans contained in the peaks 16 and 20 are illustrated. GLM repeated measures ANOVA withposthoc paired sample t-tests were performed. Significance levels of posthoc tests were corrected for multiple comparisons (α/n), with p-values ≤0.0022 found to be significant and shown with ***. Samples were analyzed in triplicate, and results are expressed as mean ± SE (error bars).
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523745

ppm, MS/MS tolerance of 0.25 Da and peptide charge of +1.Peptides were filtered for subsequent identification and analysiswith a confidence interval of 95% for top ranked proteins usingthe Mowse scoring algorithm.
ELISA Determination of AGP Concentration
An AssayMax quantitative competitive enzyme-linked immu-nosorbent assay (ELISA) kit for human AGP (EG5001-1) waspurchased from AssayPro (St Charles, MO, USA). Serialdilutions of AGP standard were carried out to generate astandard curve. Bradford protein assays29 were used todetermine protein concentrations of serum samples, and allsamples were adjusted to 70 mg/mL using phosphate bufferedsaline. Serum samples were diluted 1:1000 with the mix diluent,and all samples and standards were incubated for 2 h withbiotinylated AGP on a 96-well plate, precoated with polyclonalantibody specific for human AGP. The wells were washed andincubated subsequently with streptavidin-peroxidase conjugatefor 30 min. The wells were washed again, and chromogensubstrate was added, followed by addition of the stop solution.The absorbance values were determined on a Perkin-ElmerVictor X3 multilabel plate reader at 450 nm. The results wereanalyzed by regression analysis using four-parameter logisticcurve-fit.
Statistical Analysis
General linear model (GLM), repeated measures ANOVA testswere carried out to determine if there were significantdifferences between the glycosylation profiles of serum frompatients before and after olanzapine treatment using SPSSstatistical analysis software version 18 (Dublin, Ireland). Whensignificant interactions were found, posthoc paired sample t-tests were performed to identify peaks that were alteredsignificantly after treatment. In experiments that tested theeffects of olanzapine treatment on glycosylation of whole serumproteins and low-abundance serum proteins (n = 23), thesignificance levels of posthoc tests were corrected for multiplecomparisons (α/n; 0.05/23 = 0.0022). Therefore, only p-values≤ 0.0022 were considered to show significant differences inthese studies. The significance levels of the paired sample t-testson glycosylation of isolated high-abundance proteins and onthe levels of branching, galactosylation and sialylation in wholeserum were not corrected for multiple comparisons, as only 6patients were considered in these tests. A paired sample t-testwas carried out on the AGP ELISA results.
■ RESULTS
Effects of Olanzapine Treatment on Serum Glycome
HILIC fluorescence was used to analyze released N-glycanpools from proteins in whole serum obtained from patientsbefore and after treatment with olanzapine (Figure 2). Thisfigure shows that two glycan peaks were altered significantly inresponse to olanzapine treatment, on the basis of averagerelative peak area measurements (n = 23). Peak 16 wasincreased from 32.7 ± 0.6% to 35.1 ± 0.7% (increase of 7.3%; p= 0.0018), and peak 20 was decreased from 2.17 ± 0.07% to1.98 ± 0.06% (decrease of 8.8%; p = 0.0016). Glycanassignments were made with the aid of exoglycosidasedigestions as described by Royle et al.27 and comparisonswith GlycoBase (Supporting Information Table S1). Theglycan identified in peak 16 was A2G2S2, a disialylateddigalactosylated biantennary structure that is the mostabundant glycan in human serum. Peak 20 was found to
contain three disialylated bi- and triantennary glycans, A3G3S2,A2F1G2S2 and A3BG3S2.The same analysis was also performed on the low-abundance
serum protein fraction of serum, obtained after the depletionstep, and peaks were assigned with the equivalent numbers asthose in whole serum. This showed that olanzapine had noeffect on the relative size of any of the 15 peaks detected, as allp-values were higher than 0.0022 (Figure 2). However, peaks16 and 20 did show the same directional changes as those seenin whole serum (peak 16 was increased and peak 20 wasdecreased after treatment). Characterization of the glycans ineach of the peaks showed that peak 16 contained the mostabundant glycan in serum, A2G2S2, and peak 20 containedA3G3S2, A2F1G2S2 and A3BG3S2 (Supporting InformationTable S1).
Identification of Serum Proteins Associated withOlanzapine-Induced Glycan Changes
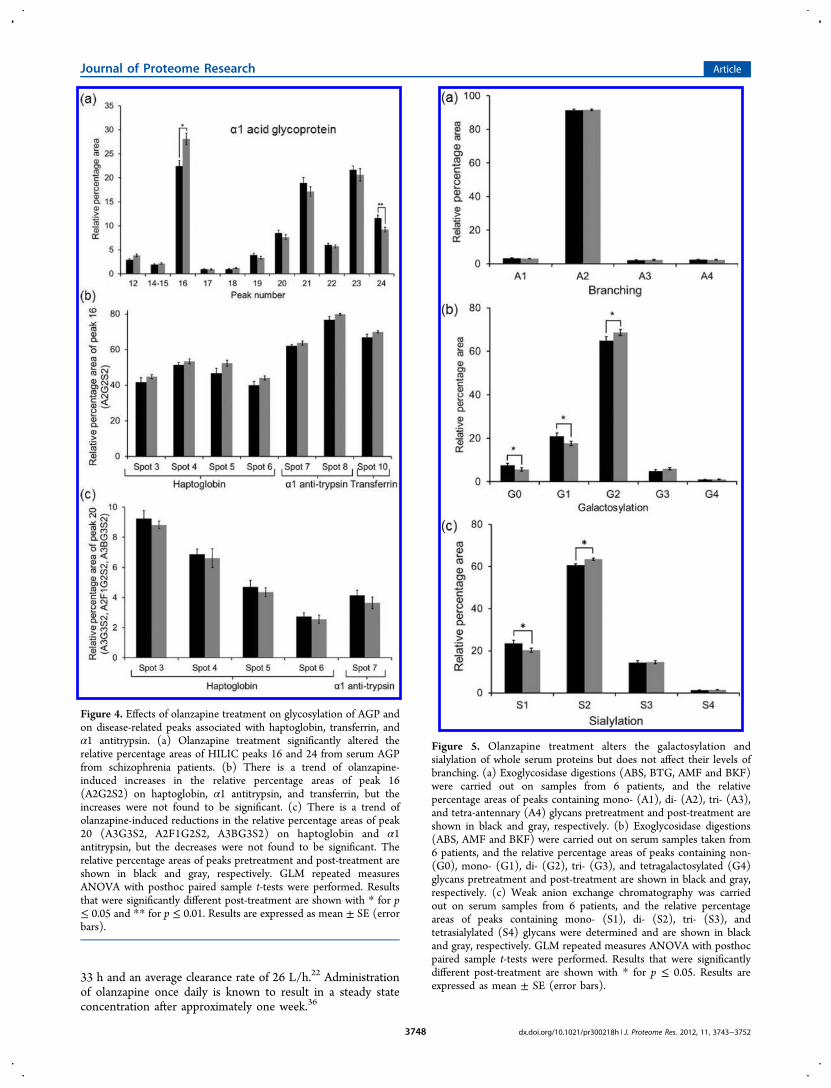
The subsequent phase of the study was aimed at identificationof serum proteins that were associated with the olanzapine-induced glycan changes. 2DE of whole serum from 6 patientsbefore and after treatment was performed as described in theExperimental Section. These 6 samples were chosen as theyshowed the highest increase in A2G2S2 in whole serum afterthe olanzapine treatment. Protein spots corresponding to 4glycoproteins (AGP, haptoglobin β-chain, transferrin and α1antitrypsin) were isolated using a Gilson p1000 pipet and tipsand the SwissProt 2D PAGE Human plasma image as areference guide (Figure 3). AGP was selected because it hasbeen shown that olanzapine can bind to this protein.22 Inaddition, AGP and the other selected proteins (haptoglobin β-chain, α1 antitrypsin, transferrin and α1 antitrypsin) are knownto carry A2G2S2, and some also carry A3G3S2, A2F1G2S2 andA3BG3S2.30−33 MALDI-TOF analysis was performed on theisolated protein spots to confirm the protein identifications.The resulting data showed that spot 1 contained AGP, spots 2−6 contained haptoglobin β-chain, spots 7−9 contained α1antitrypsin and spot 10 contained transferrin (Table 1). Theisolated proteins were subjected to glycan analysis, andexamples of the HILIC-fluorescence profiles are shown inFigure 3. Peak numbers were assigned as above for wholeserum to allow direct comparisons. The low intensity HILIC-fluorescence profiles from analysis of spots 2 and 9 from somesamples resulted in numbers that were too small for statisticalanalyses. However, analysis of the AGP spots resulted in asignificantly different post-treatment profile (Figure 4). Theresults show that peak 16 increased from 22.4 ± 1.2% to 28.1 ±1.2% (p = 0.027) and peak 24 decreased from 11.6 ± 0.6% to9.2 ± 0.5% (p = 0.007) with treatment. Peak 20 was notsignificantly altered as seen above for the whole serum analysis,although the same directional change was observed with adecrease in peak area. Analysis of the glycan structuresassociated with AGP showed that peak 16 contained A2G2S2as above, peak 20 contained A3G3S2, and peak 24 containedmultiple tetra-antennary structures, including A4G4S4,A4F1G4S4 and A4F2G4S4 (Supporting Information TableS1). Even though olanzapine did not significantly alter theglycosylation of haptoglobin, α1 antitrypsin and transferrin, thesame directional changes were observed for peak 16 (increased)and peak 20 (decreased) as found in the case of whole serumproteins (Figures 2 and 4).
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523746

Effects of Olanzapine Treatment on the Concentration ofAGP in Serum Samples
To determine whether AGP levels were altered after olanzapinetreatment, we performed an ELISA analysis to determine theconcentrations of this protein in the serum samples fromschizophrenia patients before and after treatment witholanzapine. This showed that the concentration of AGP wasnot significantly altered by the treatment with a pretreatment
level of 0.50 ± 0.02 mg/mL and a post-treatment level of 0.44± 0.03 mg/mL (p = 0.069).Effects of Olanzapine Treatment on Branching,Galactosylation and Sialylation of Whole Serum Glycans
Glycans are synthesized by the actions of a number ofglycosyltransferases and glycosidases in the endoplasmicreticulum and the Golgi apparatus, which bring about removaland addition of monosaccharides to the trimannosyl core in astepwise manner. To investigate the olanzapine-induced glycanalterations found in whole serum, experiments were carried outon specific sections of the glycan processing pathway todetermine if the identified changes were due to alteredbranching, galactosylation and/or sialylation. Analysis ofsamples after specific exoglycosidase digestions examined thedegree of glycan branching and the levels of galactosylation.These analyses showed that the olanzapine treatment did notaffect the levels of mono- (A1), di- (A2), tri- (A3) and tetra-antennary (A4) glycans, although significant alterations wereidentified in the degree of galactosylation (Figure 5).Specifically, the levels of non- (G0) and monogalactosylated(G1) glycans were significantly lower after treatment (p = 0.025and 0.027, respectively), whereas the levels of digalactosylated(G2) glycans were significantly increased (p = 0.022). G0 andG1 forms decreased from 7.5 ± 1.1% to 5.6 ± 0.7% and from20.9 ± 1.6% to 17.6 ± 1.1%, respectively, and G2 increasedfrom 64.9 ± 1.8% to 68.7 ± 1.4%. Similarly, the levels ofsialylated glycans were affected by olanzapine treatment withmonosialylated (S1) glycans decreased from 23.5 ± 1.5% to20.3 ± 1.0% (p = 0.038), and disialylated (S2) glycansincreased from 60.7 ± 0.6% to 63.5 ± 0.5% (p = 0.040) (Figure5).
■ DISCUSSIONThis is the first study to investigate the effects of antipsychotictreatment on serum glycosylation profiles in schizophreniapatients. Serum samples were obtained from paranoidschizophrenia patients before and after 6 weeks of treatmentwith olanzapine. This protocol served to minimize differencesin factors such as age, smoking, BMI and cholesterol levels,which have been reported previously to affect the glycosylationof proteins.34,35 The pharmacokinetics of olanzapine showlinearity and are dose-proportional, with an average half-life of
Figure 3. Isolation of high-abundance serum proteins by 2DE andtypical HILIC profiles. (a) 2DE separation of whole serum from adrug-naive schizophrenia patient. High-abundance serum glycopro-teins, AGP (spot 1), haptoglobin β-chain (spots 2−6), α1 antitrypsin(spots 7−9), and transferrin (spot 10), were excised from the gels andwere prepared for N-glycan analysis and for identification of proteinsby MALDI-TOF/TOF mass spectrometry. (b) Typical HILIC profilesfor AGP, haptoglobin β-chain, α1 antitrypsin and transferrin, with theassigned peak numbers shown for AGP and relevant peaks illustratedfor haptoglobin β-chain, α1 antitrypsin, and transferrin.
Table 1. Identification of the Peptides in Excised Gel Spots 1to 10 by MALDI-TOF/TOF Mass Spectrometry Analysis
spot identificationaccessionnumber
proteinion score
sequencecoveragea (%)
peptidesmatcheda
1 AGP P02763 122 23 32 haptoglobin β-
chainP00738 124 6 2
3 haptoglobin β-chain
P00738 407 20 8
4 haptoglobin β-chain
P00738 459 20 8
5 haptoglobin β-chain
P00738 380 15 6
6 haptoglobin β-chain
P00738 442 17 7
7 α1 antitrypsin P01009 96 9 38 α1 antitrypsin P01009 482 27 89 α1 antitrypsin P01009 594 31 1010 transferrin P02787 491 21 12
aCombined results from identical gel spots from up to 12 samples.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523747
33 h and an average clearance rate of 26 L/h.22 Administrationof olanzapine once daily is known to result in a steady stateconcentration after approximately one week.36
Figure 4. Effects of olanzapine treatment on glycosylation of AGP andon disease-related peaks associated with haptoglobin, transferrin, andα1 antitrypsin. (a) Olanzapine treatment significantly altered therelative percentage areas of HILIC peaks 16 and 24 from serum AGPfrom schizophrenia patients. (b) There is a trend of olanzapine-induced increases in the relative percentage areas of peak 16(A2G2S2) on haptoglobin, α1 antitrypsin, and transferrin, but theincreases were not found to be significant. (c) There is a trend ofolanzapine-induced reductions in the relative percentage areas of peak20 (A3G3S2, A2F1G2S2, A3BG3S2) on haptoglobin and α1antitrypsin, but the decreases were not found to be significant. Therelative percentage areas of peaks pretreatment and post-treatment areshown in black and gray, respectively. GLM repeated measuresANOVA with posthoc paired sample t-tests were performed. Resultsthat were significantly different post-treatment are shown with * for p≤ 0.05 and ** for p ≤ 0.01. Results are expressed as mean ± SE (errorbars).
Figure 5. Olanzapine treatment alters the galactosylation andsialylation of whole serum proteins but does not affect their levels ofbranching. (a) Exoglycosidase digestions (ABS, BTG, AMF and BKF)were carried out on samples from 6 patients, and the relativepercentage areas of peaks containing mono- (A1), di- (A2), tri- (A3),and tetra-antennary (A4) glycans pretreatment and post-treatment areshown in black and gray, respectively. (b) Exoglycosidase digestions(ABS, AMF and BKF) were carried out on serum samples taken from6 patients, and the relative percentage areas of peaks containing non-(G0), mono- (G1), di- (G2), tri- (G3), and tetragalactosylated (G4)glycans pretreatment and post-treatment are shown in black and gray,respectively. (c) Weak anion exchange chromatography was carriedout on serum samples from 6 patients, and the relative percentageareas of peaks containing mono- (S1), di- (S2), tri- (S3), andtetrasialylated (S4) glycans were determined and are shown in blackand gray, respectively. GLM repeated measures ANOVA with posthocpaired sample t-tests were performed. Results that were significantlydifferent post-treatment are shown with * for p ≤ 0.05. Results areexpressed as mean ± SE (error bars).
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523748

The olanzapine treatment in the current study brought aboutalterations in the glycosylation profile of whole serumglycoproteins. The levels of the disialylated biantennary glycan,A2G2S2, were significantly increased, whereas other disialylatedbi- and triantennary oligosaccharides, A3G3S2, A2F1G2S2 andA3BG3S2, were significantly decreased post-treatment.A2G2S2 is the most abundant glycan in human serum and isexpressed on the surface of many glycoproteins, including anumber of abundant serum proteins such as haptoglobin,transferrin, AGP, α1 antitrypsin and IgG. A2F1G2S2 containsthe sialyl lewis X (sLex) epitope, which is a ligand for theselectins (E-selectin, P-selectin and L-selectin) and is involvedin the process of leukocyte extravasation and possibly inmetastasis, making it an interesting target in cancerresearch.30,31,37 In a previous study, we showed that the levelsof another sLex-containing glycan, A3F1G3S3, were altered inserum from schizophrenia patients compared to healthycontrols.9 However, sLex-carrying glycans did not appear tobe altered by the olanzapine treatment in the current study.In the current study, we identified alterations on whole
serum glycoproteins but did not identify any significantalterations in glycosylation profiles through analysis of animmunodepleted serum fraction, comprising proteins of lowerabundance. This suggested that olanzapine treatment onlyaffects glycosylation of the most abundant serum proteins.Alternatively, it is also possible that significant changes in thelow-abundance fraction were not detected because oflimitations in the sensitivity of the method. This is consistentwith the finding that increased levels of A2G2S2 and decreasedA3G3S2, A2F1G2S2 and A3BG3S2 were observed in the low-abundance protein fraction, although these alterations did reachthe level of significance.Four high-abundance serum proteins were purified from 2DE
gels to determine whether any of these could account for theobserved changes in the glycosylation profile in whole serum.This included selection of AGP based on studies that haveshown that olanzapine can bind to this protein.22 In addition,AGP and the other selected proteins (haptoglobin β-chain,transferrin and α1 antitrypsin) are known to carry A2G2S2, andsome also carry A3G3S2, A2F1G2S2 and A3BG3S2.30−33 TheIPG strips used for the 2DE gels had a pH range from 3 to 7,which was chosen to maximize detection of targeted sialylatedglycoproteins AGP, haptoglobin β-chain, transferrin and α1antitrypsin. This analysis revealed that AGP carried significantlyaltered glycans, with increased levels of A2G2S2 and reducedlevels of tetra-antennary glycans. However, nonsignificantchanges were also observed in the levels of other glycanstructures on haptoglobin β-chain, transferrin and α1antitrypsin. These findings suggested that the olanzapine-induced glycan alterations result from modification of specificaspects of the glycan processing machinery (e.g., branching,galactosylation and/or sialylation) rather than solely as a resultof altered glycans on AGP.AGP is a 41−43 kDa acute phase protein that has 5 sites of
N-glycosylation,38 and the oligosaccharides account for 40% ofthe molecular weight. Alterations in microheterogeneity ofAGP glycosylation can occur in diseases such as in pancreaticcancer,39 ovarian cancer,32 inflammation40−42 and diabetes,41
and can result in modification of certain immunomodulatoryand binding properties of the protein.43 Such alterations inAGP glycosylation can have downstream effects on biologicalpathways. For example, it has been shown that the ability ofAGP to inhibit lymphocyte proliferation is affected by its
glycosylation state, with the presence of branched glycansleading to more effective inhibition.44 Also, the nonsialylatedform of AGP has been shown to have greater plateletaggregation inhibitory capacity than other forms.45
AGP has the ability to bind to several basic and neutraldrugs,46 such as olanzapine. The drug interaction sites on AGPoverlap spatially, and amino acids such as tryptophan, tyrosine,lysine and histidine are crucial for binding.47 Previous studieshave suggested that the glycans carried on AGP are notinvolved in the binding of drugs to the protein, with reportsshowing that the removal of glycan structures such as sialicacids from AGP did not reduce the binding of drugs to thisprotein.48−50 This is also supported by the finding that arecombinant form of AGP containing high mannose glycanshad similar binding capacity as human AGP, even though theglycans are different.51 These findings suggest that it is unlikelythat the binding of olanzapine to AGP is responsible for theolanzapine-induced glycan alterations found in the currentstudy. In addition, the changes were not likely to be due tochanges in overall AGP levels, as we found that this protein wasnot significantly altered by the olanzapine treatment. Instead,these findings suggest that the observed changes were due toalterations in the glycan processing pathway.To confirm this, we examined the effects of olanzapine on
specific stages of glycan processing, including branching,galactosylation and sialylation. Extension of oligosaccharidechains takes place in the Golgi apparatus where glycosyl-transferases facilitate transfer of mononucleotide sugars ontocarbohydrate backbone structures in a specific order. Theexisting carbohydrate structure can affect and determine theposition and addition of further mononucleotides to the chain.Exoglycosidase digestion analyses showed that olanzapine didnot alter the degree of branching (A1−A4) of the whole serumglycans, suggesting that there was no effect on the N-acetylglucosaminyltransferase enzymes that add N-acetylglucos-amine (GlcNAc) residues to core mannose residues or on theexpression of the genes encoding these enzymes. However, theanalyses identified alterations in galactosylation of glycans. Thelevels of G2 glycans were increased post-treatment, whereas thelevels of G0 and G1 glycans were decreased. This increase indigalactosylated structures could occur as a result of increasedexpression of genes that encode for specific galactosyltrans-ferases, or it could result from increased activation of theenzymes. In addition, other factors that can affect the level ofgalactosylation include the availability of the UDP-α-galactosesubstrate, the availability of cofactors required by the enzymes,and the residence times of proteins in the Golgi apparatus andendoplasmic reticulum where glycosylation processing takesplace. A previous study showed that olanzapine treatmentincreased the expression of the B4GALT1 gene in the liver ofschizophrenia patients.23 This gene encodes for β1,4-galactosyltransferase I (Gal-T1), which adds galactose fromUDP-α-galactose to a GlcNAc residue at the nonreducing endof a carbohydrate chain in a β1−4-linkage. Gal-T1 is a type IItransmembrane protein that is localized to the trans-Golginetwork and has a secondary function of converting α-lactalbumin and glucose to lactose in the presence of α-lactalbumin.52 Increased expression and activity of this enzymecould lead to increased levels of galactosylation of GlcNAcresidues in glycans.Sialylation occurs on galactose residues at the nonreducing
ends of glycans. Weak anion exchange chromatography wasused in the current study to determine the levels of mono-
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523749

(S1), di- (S2), tri- (S3), and tetrasialylated (S4) glycans inwhole serum samples. This analysis showed that the levels of S2glycans were increased post-treatment and the levels of S1glycans were decreased. The concurrent increase of S2 and G2suggests that sialylation levels may have been increased as aresult of increased levels of digalactosylated glycans available forsialylation. Taken together, these results suggest that olanzapinealters the overall glycosylation of whole serum proteins bymodifying the levels of galactosylation and sialylation ofglycans.The source of the olanzapine-induced glycan alterations in
the current study remains unclear; however, two possiblesources are discussed here. One possible source could berelated to olanzapine-induced changes to the central nervoussystem (CNS). Approximately 80% of proteins in the CSForiginate from the blood and 20% from the CNS.53 There isbidirectional transfer of proteins from the plasma to CSF bysize dependent diffusion through the blood CSF barrier.Because the concentration of proteins in the CSF is much lowerthan that of plasma, it is unlikely that many proteins transferfrom the CSF to the plasma, suggesting that the olanzapine-induced changes observed in the current study in serum maynot come about as a result of olanzapine-induced alterations tothe CNS, as these effects would most likely not be evident onserum proteins.A more likely possible source of the glycan alterations is that
olanzapine acts on the periphery, altering the activity of hepaticenzymes, resulting in altered glycosylation of proteins,particularly acute phase proteins. Olanzapine is primarilymetabolized in the liver to its inactive compound bycytochrome P450 CYP1A2, a monooxygenase that is locatedin the endoplasmic reticulum.22 Cytochrome P450 enzymes areconcentrated in the liver and are also present in lower levels intissues of other organs including lungs and kidneys. Previousstudies have investigated the effects of olanzapine on the levelsof liver enzymes such as alkaline phosphatase, alanineaminotransferase and aspartate aminotransferase during treat-ment with the drug. Olanzapine-induced elevations in liverenzymes in a transient, asymptomatic and dose-independentmanner were reported in a number of patients.54−56 To a lesserdegree, increased liver enzymes in a considerable and enduringmanner were also reported in a small number of patients.57
Acute phase proteins such as AGP, haptoglobin and α1antitrypsin are produced in the liver, with their productiondramatically increasing during inflammatory processes. Glyco-sylation of these acute phase proteins takes places in theendoplasmic reticulum and Golgi apparatus in hepatic cells bythe actions of glycosyltransferases and glycosidases. Because ofthe reported effects of olanzapine on hepatic enzymes, it maybe possible that the olanzapine-induced glycosylation alter-ations reported in the current study may occur as a result ofaltered activity of hepatic glycosylation processing enzymes.The current study examined the glycosylation profiles of
serum samples from schizophrenia patients who had beentreated with olanzapine. Because it is possible that the drug-induced alterations may occur as a result of alterations tohepatic enzyme function as discussed above, it is possible thatthese reported drug-induced glycan alterations could be seen ina population of healthy controls if treated with olanzapine inthe same manner as the schizophrenia patients. This providesscope for a future study to examine this further.Such induced changes by olanzapine to the glycosylation of
biologically active glycoproteins may influence their immuno-
genicity, their involvement in cell−cell interactions andfunctions and/or their clearance rate. A previous study showedthat the glycans on the surface of AGP regulated the clearancerate of the protein with high mannose structures resulting inthe fastest rate of clearance and complex-type glycans showingthe slowest clearance rate.58
Previous studies have shown that the levels of A2G2S2 onAGP are increased in acute inflammation and the levels of tri-and tetra-antennary glycans are increased in chronic inflamma-tion.40−42 There is evidence of inflammation occurring inschizophrenia,5−7,59,60 but there are conflicting reports onwhether olanzapine has pro- or anti-inflammatory effects.61,62
Sugino et al. showed that olanzapine decreased the levels ofTNFα and IL-6 and increased the levels of anti-inflammatoryIL-10,62 implying a drug-induced decrease in inflammation.However, Meyer et al. reported an olanzapine-inducedincreased level of c-reactive protein, a marker of inflamma-tion.63
■ CONCLUSIONSThis study has shown that olanzapine treatment of schizo-phrenia patients resulted in changes in the glycosylationmachinery associated with the biosynthesis of abundantserum proteins. Specifically, olanzapine appeared to affect theextent of digalactosylation and disialylation of serum proteins.As glycosylation impacts on many important cellular processesincluding cellular recognition, protein function, proteinstability, immunogenicity and protein clearance rates, olanza-pine-induced glycosylation changes may induce a number ofdownstream effects.
■ ASSOCIATED CONTENT*S Supporting Information
Table S1: Predominant glycan structures that are present inwhole serum, low-abundance serum protein fraction, and AGPand the GU values of the peaks in each sample. This material isavailable free of charge via the Internet at http://pubs.acs.org.
■ AUTHOR INFORMATIONCorresponding Author
*Tel.: +353-12158142. Fax: +353-12158116. E-mail: [email protected]
The authors declare no competing financial interest.
■ ACKNOWLEDGMENTSThis work was supported by the European Commission underthe Seventh Framework Programme (FP7) SchizDX [GrantNo. 223427]. R.S. acknowledges the European Union SeventhFramework Programme (FP7) under Grant Agreement No.260600 (GlycoHIT).
■ ABBREVIATIONSA1, A2, A3, A4, mono-, bi-, tri-, tetra-antennary glycans; 2AB, 2-amino benzamide; ABS, Arthrobacter ureafaciens sialidase; AGP,α1 acid glycoprotein; AMF, almond meal α-fucosidase; BKF,bovine kidney α-fucosidase; BTG, bovine testes β-galactosi-dase; CSF, cerebrospinal fluid; 2DE, 2D electrophoresis; G0,G1, G2, G3, G4, non-, mono-, di-, tri-, tetragalactosylatedglycans; GalT1, β1,4-galactosyltransferase I; GlcNAc, N-acetylglucosamine; GLM, general linear model; GU, glucose
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523750

units; GUH, β-N-acetylglucosaminidase cloned from Strepto-coccus pneumonia, expressed in Escherichia coli; HILIC,hydrophilic interaction liquid chromatography; MALDI-TOF,matrix-assisted laser desorption/ionization time-of-flight massspectrometry; S1, S2, S3, S4, mono-, di-, tri-, tetrasialylatedglycans; sLex, sialyl-Lewis X
■ REFERENCES(1) Huang, J. T.; Wang, L.; Prabakaran, S.; Wengenroth, M.;Lockstone, H. E.; Koethe, D.; Gerth, C. W.; Gross, S.; Schreiber, D.;Lilley, K.; Wayland, M.; Oxley, D.; Leweke, F. M.; Bahn, S.Independent protein-profiling studies show a decrease in apolipopro-tein A1 levels in schizophrenic CSF, brain and peripheral tissues. Mol.Psychiatry 2008, 12, 1118−1128.(2) Levin, Y.; Wang, L.; Schwarz, E.; Koethe, D.; Leweke, F. M.;Bahn, S. Global proteomic profiling reveals altered proteomicsignature in schizophrenia serum. Mol. Psychiatry 2010, 15, 1088−1100.(3) Maes, M.; Delange, J.; Ranjan, R.; Meltzer, H. Y.; Desnyder, R.;Cooremans, W.; Scharpe, S. Acute phase proteins in schizophrenia,mania and major depression: modulation by psychotropic drugs.Psychiatry Res. 1997, 66, 1−11.(4) Yang, Y.; Wan, C.; Li, H.; Zhu, H.; La, Y.; Xi, Z.; Chen, Y.; Jiang,L.; Feng, G.; He, L. Altered levels of acute phase proteins in theplasma of patients with schizophrenia. Anal. Chem. 2006, 78, 3571−3576.(5) Lin, A.; Kenis, G.; Bignotti, S.; Tura, G. J.; De Jong, R.; Bosmans,E.; Pioli, R.; Altamura, C.; Scharpe, S.; Maes, M. The inflammatoryresponse system in treatment-resistant schizophrenia: Increased seruminterleukin-6. Schizophr. Res. 1998, 32, 9−15.(6) Abdeljaber, M. H.; Nair, M. P.; Schork, M. A.; Schwartz, S. A.Depressed natural killer cell activity in schizophrenic patients.Immunol. Invest. 1994, 23, 259−268.(7) Radewicz, K.; Garey, L. J.; Gentleman, S. M.; Reynolds, R.Increase in HLA-DR immunoreactive microglia in frontal andtemporal cortex of chronic schizophrenics. J. Neuropathol. Exp. Neurol.2000, 59, 137−150.(8) Schwarz, E.; Guest, P. C.; Rahmoune, H.; Harris, L. W.; Wang, L.;Leweke, F. M.; Rothermundt, M.; Bogerts, B.; Koethe, D.; Kranaster,L.; Ohrmann, P.; Suslow, T.; McAllister, G.; Spain, M.; Barnes, A.; vanBeveren, N. J.; Baron-Cohen, S.; Steiner, J.; Torrey, F. E.; Yolken, R.H.; Bahn, S. Identification of a biological signature for schizophrenia inserum. Mol. Psychiatry 2011, 1−9.(9) Stanta, J. L.; Saldova, R.; Struwe, W. B.; Byrne, J. C.; Leweke, F.M.; Rothermund, M.; Rahmoune, H.; Guest, P. C.; Bahn, S.; Rudd, P.M. Identification of N-glycosylation changes in the CSF and serum ofpatients with schizophrenia. J. Proteome. Res. 2010, 9, 4476−4489.(10) Bauer, D.; Haroutunian, V.; Meador-Woodruff, J. H.;McCullumsmith, R. E. Abnormal glycosylation of EAAT1 andEAAT2 in prefrontal cortex of elderly patients with schizophrenia.Schizophr. Res. 2010, 117, 92−98.(11) Isomura, R.; Kitajima, K.; Sato, C. Structural and functionalimpairments of polysialic acid by a mutated polysialyltransferase foundin schizophrenia. J. Biol. Chem. 2011, 286, 21535−21545.(12) Fukuda, T.; Hashimoto, H.; Okayasu, N.; Kameyama, A.; Onogi,H.; Nakagawasai, O.; Nakazawa, T.; Kurosawa, T.; Hao, Y.; Isaji, T.;Tadano, T.; Narimatsu, H.; Taniguchi, N.; Gu, J. α1,6-Fucosyltransfer-ase-deficient mice exhibit multiple behavioural abnormalities asso-ciated with a schizophrenia-like phenotype. J. Biol. Chem. 2011, 286,18434−18443.(13) Saldova, R.; Huffmann, J. E.; Adamczyk, B.; Muzinic, A.; Kattla,J. J.; Pucic, M.; Novokmet, M.; Abrahams, J. L.; Hayward, C.; Rudan,I.; Wild, S. H.; Wright, A.; Polasek, O.; Lauc, G.; Campbell, H.;Wilson, J. F.; Rudd, P. M. Association of medication and the humanplasma N-glycome. J. Proteome Res. 2012, 11, 1821−1831.(14) Smith, R. C.; Infante, M.; Singh, A.; Khandat, A. The effects ofolanzapine on neurocognitive functioning in medication-refractoryschizophrenia. Int. J. Neuropsychopharmacol. 2001, 4, 239−250.
(15) Guo, X.; Zhai, J.; Wei, Q.; Twamley, E. W.; Jin, H.; Fang, M.;Hu, M.; Zhao, J. Neurocognitive effects of first- and second-generationantipsychotic drugs in early-stage schizophrenia: A naturalistic 12-month follow-up study. Neurosci. Lett. 2011, 503, 414−146.(16) Bhana, N.; Foster, R. H.; Olney, R.; Plosker, G. L. Olanzapine:an updated review of its use in the management of schizophrenia.Drugs 2001, 61, 111−161.(17) Bobes, J.; Gibert, J.; Ciudad, A.; Alvarez, E.; Canas, F.; Carrasco,J. L.; Gascon, J.; Gomez, J. C.; Guttierrez, M. Safety and effectivenessof olanzapine versus conventional antipsychotics in the acutetreatment of first episode schizophrenic inpatients. Prog. Neuro-psychopharmacol. Biol. Psychiatry 2003, 27, 473−481.(18) Komossa, K.; Rummel-Kluge, C.; Hunger, H.; Schmid, F.;Schwarz, S.; Duggan, L.; Kissling, W.; Leucht, S. Olanzapine versusother atypical antipsychotics for schizophrenia. Cochrane Database Syst.Rev. 2010, 17 (3), CD006654.(19) Alvarez, E.; Ciudad, A.; Olivares, J. M.; Bousono, M.; Gomez, J.C. A randomized 1-year follow-up study of olanzapine and risperidonein the treatment of outpatients with schizophrenia. J. Clin.Psychopharmacol. 2006, 26, 238−249.(20) Bymaster, F. P.; Calligaro, D. O.; Falcone, J. F.; Marsh, R. D.;Moore, N. A.; Tye, N. C.; Seeman, P.; Wong, D. T. Radioreceptorbinding profile of the atypical antipsychotic olanzapine. Neuro-psychopharmacology 1996, 14, 87−96.(21) Meltzer, H. Y.; Massey, B. W. The role of serotonin receptors inthe action of atypical antipsychotic drugs. Curr. Opin. Pharmacol. 2011,11, 59−67.(22) Callaghan, J. T.; Bergstrom, R. F.; Ptak, L. R.; Beasley, C. M.Olanzapine. Pharmacokinetic and pharmacodynamic profile. Clin.Pharmacokinet. 1999, 37, 177−193.(23) Choi, K. H.; Higgs, B. W.; Weis, S.; Song, J.; Llenos, I. C.;Dulay, J. R.; Yolken, R. H.; Webster, M. J. Effects of typical andatypical antipsychotic drugs on gene expression profiles in the liver ofschizophrenic patients. BMC Psychiatry 2009, 9, 57−72.(24) Chiba, S.; Hashimoto, R.; Hattori, S.; Yohda, M.; Lipska, B.;Weinberger, D. R.; Kunugi, H. Effect of antipsychotic drugs on DISC1and dysbindin expression in mouse frontal cortex and hippocampus. J.Neural Transm. 2006, 113, 1337−1346.(25) Chan, M. K.; Tsang, T. M.; Harris, L. W.; Guest, P. C.; Holmes,E.; Bahn, S. Evidence for disease and antipsychotic medication effectsin post-mortem brain from schizophrenia patients. Mol. Psychiatry2011, 16, 1189−1202.(26) Ma, D.; Chan, M. K.; Lockstone, H. E.; Pietsch, S. R.; Jones, D.N. C.; Cilia, J.; Hill, M. D.; Robbins, M. J.; Benzel, I. M.; Umrania, Y.;Guest, P. C.; Levin, Y.; Maycox, P. R.; Bahn, S. Antipsychotictreatment alters protein expression associated with presynapticfunction and nervous system development in rat frontal cortex. J.Proteome Res. 2009, 8, 3284−3297.(27) Royle, L.; Campbell, M. P.; Radcliffe, C. M.; White, D. M.;Harvey, D. J.; Abrahams, J. L.; Kim, Y. G.; Henry, G. W.; Shadick, N.A.; Weinblatt, M. E.; Lee, D. M.; Rudd, P. M.; Dwek, R. A. HPLC-based analysis of serum N-glycans on a 96-well plate platform withdedicated database software. Anal. Biochem. 2008, 376, 1−12.(28) Harvey, D. J.; Merry, A. H.; Royle, L.; Campbell, M. P.; Dwek,R. A.; Rudd, P. M. Proposal for a standard system for drawingstructural diagrams of N- and O-linked carbohydrate and relatedcompounds. Proteomics 2009, 9, 3796−3801.(29) Bradford, M. M. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing the principleof protein-dye binding. Anal. Biochem. 1976, 72, 248−254.(30) Bones, J.; Byrne, J. C.; O’Donoghue, N.; McManus, C.; Scaife,C.; Boissin, H.; Nastase, A.; Rudd, P. M. Glycomic and glycoproteomicanalysis of serum from patients with stomach cancer reveals potentialmarkers arising from host defense mechanisms. J. Proteome Res. 2011,10, 1246−1265.(31) Saldova, R.; Wormald, M. R.; Dwek, R. A.; Rudd, P. M.Glycosylation changes on serum glycoproteins in ovarian cancer maycontribute to disease pathogenesis. Dis. Markers 2008, 25, 219−232.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523751

(32) Saldova, R.; Royle, L.; Radcliffe, C. M.; Hamid, U. M.; Evans, R.;Arnold, J. N.; Banks, R. E.; Hutson, R.; Harvey, D. J.; Antrobus, R.;Petrescu, S. M.; Dwek, R. A.; Rudd, P. M. Ovarian cancer is associatedwith changes in glycosylation in both acute-phase proteins and IgG.Glycobiology 2007, 17, 1344−1356.(33) ones, J.; Mittermayr, S.; O’Donoghue, N.; Guttman, A.; Rudd, P.M. Ultra performance liquid chromatographic profiling of serum N-glycans for fast efficient identification of cancer associated alterationsin glycosylation. Anal. Chem. 2010, 82, 10208−10215.(34) Knezevic,́ A.; Polasek, O.; Gornik, O.; Rudan, I.; Campbell, H.;Hayward, C.; Wright, A.; Kolcic, I.; O’Donoghue, N.; Bones, J.; Rudd,P. M.; Lauc, G. Variability, heritability and environmental determi-nants of human plasma N-glycome. J. Proteome Res. 2009, 8, 694−701.(35) Knezevic, A.; Gornik, O.; Polasek, O.; Pucic, M.; Redzic, I.;Novokmet, M.; Rudd, P. M.; Wright, A. F.; Campbell, H.; Rudan, I.;Lauc, G. Effects of aging, body mass index, plasma lipid profiles andsmoking on human plasma. Glycobiology 2010, 8, 959−969.(36) Sharif, Z. A. Pharmacokinetics, metabolism and drug-druginteractions of atypical anti-psychotics in special populations. J. Clin.Psychiatry Suppl. 2003, 6, 22−25.(37) Abd Hamid, U. M.; Royle, L.; Saldova, R.; Radcliffe, C. M.;Harvey, D. J.; Storr, S. J.; Pardo, M.; Androbus, R.; Chapman, C. J.;Zitzmann, N.; Robertson, J. F.; Dwek, R. A.; Rudd, P. M. A strategy toreveal potential glycan markers from serum glycoproteins associatedwith breast cancer progression. Glycobiology 2008, 18, 1105−1118.(38) Yoshima, H.; Matsumoto, A.; Mizuochi, T.; Kawasaki, T.;Kobata, A. Comparative study of the carbohydrate moieties of rat andhuman plasma alpa1 acid glycoproteins. J. Biol. Chem. 1981, 256,8476−8484.(39) Sarrats, A.; Saldova, R.; Pla, E.; Fort, E.; Harvey, D. J.; Struwe,W. B.; de Llorens, R.; Rudd, P. M.; Peracaula, R. Glycosylation of liveracute-phase proteins in pancreatic cancer and chronic pancreatitis.Proteomics: Clin. Appl. 2010, 4, 1−17.(40) de Graaf, T. W.; van der Stelt, M. E.; Anbergen, M. G.; van Dijk,W. Inflammation-induced expression of sialyl Lewis X-containingglycan structures on alpha 1-acid glycoprotein (orosomucoid) inhuman sera. J. Exp. Med. 1993, 177, 657−666.(41) Higai, K.; Azuma, Y.; Aoki, Y.; Matsumoto, K. Alteredglycosylation of α1 acid glycoprotein in patients with inflammationand diabetes mellitus. Clin. Chim. Acta 2003, 329, 117−125.(42) Higai, K.; Aoki, Y.; Azuma, Y.; Matsumoto, K. Glycosylation ofsite-specific glycans of α1 acid glycoprotein and alterations in acuteand chronic inflammation. Biochim. Biophys. Acta 2005, 1725, 128−135.(43) Cecilia, F.; Pocacqua, V. The acute phase protein α1 acidglycoprotein: A model for altered glycosylation during diseases. Curr.Protein Pept. Sci. 2007, 8, 91−108.(44) Costello, M.; Fiedel, B. A.; Gewurz, H. Inhibition of plateletaggregation by native and desialised alpha-1 acid glycoprotein. Nature1979, 281, 677−678.(45) Pos, O.; Oostendorp, R. A.; van der Stelt, M. E.; Scheper, R. J.;Van Dijk, W. Con A-nonreactive human alpha 1-acid glycoprotein(AGP) is more effective in modulation of lymphocyte proliferationthan Con A-reactive AGP serum variants. Inflammation 1990, 14,133−141.(46) Kremer, J. M.; Wilting, J.; Janssen, L. H. Drug binding to humanalfa-1-acid glycoprotein in health and disease. Pharmacol. Rev. 1988,40, 1−47.(47) Otagiri, M. A molecular functional study on the interactions ofdrugs with plasma proteins. Drug Metab. Pharmokinet. 2005, 20, 309−323.(48) Friedman, M. L.; Wermeling, J. R.; Halsall, H. B. The influenceof N-acetylneuraminic acid on the properties of human orosomucoid.Biochem. J. 1986, 236, 149−153.(49) Aubert, J. P.; Loucheux-Lefebvre, M. H. Conformational studyof alpha1-acid glycoprotein. Arch. Biochem. Biophys. 1976, 175, 400−409.
(50) Schmid, K.; Burlingame, R. W.; Paulson, J. C.; Sperandio, K.The relationship between the carbohydrate units and the secondarystructure of alpha1 acid glycoprotein. Fed. Proc. 1978, 37, 1298.(51) Nishi, K.; Fukunaga, N.; Otagiri, M. Construction of expressionsystem for human alpha1-acid glycoprotein in Pichia Pastoris andevaluation of its drug-binding properties. Drug Metab. Dispos. 2004, 32,1069−1074.(52) Brodbeck, U.; Denton, W. L.; Tanahashi, N.; Ebner, K. E. Theisolation and identification of the B protein of lactose synthetase asalpha-lactalbumin. J. Biol. Chem. 1967, 242, 1391−1397.(53) Regeniter, A.; Kuhle, J.; Mehling, M.; Moller, H.; Wurster, U.;Freidank, H.; Siede, W. H. A modern approach to CSF analysis:pathophysiology, clinical application, proof of concept and laboratoryreporting. Clin. Neurol. Neurosurg. 2009, 111, 313−318.(54) Conley, R. R.; Meltzer, H. Y. Adverse events related toolanzapine. J. Clin. Psychiatry 2000, 61, 26−29.(55) Mouradian-Stamatiadis, L.; Dumortier, G.; Januel, D.; Delmas,B. A.; Cabaret, W. Liver function tests during treatment withantipsychotics drugs: a case series of 23 patients. Prog. Neuro-psychopharmacol. 2002, 26, 1409−1411.(56) Pae, C. U.; Lim, H. K.; Kim, T. S.; Kim, J. J.; Lee, C. U.; Lee, S.J.; Lee, C.; Paik, I. H. Naturalistic observation on the hepatic enzymechanges in patients treated with either risperidone or olanzapine alone.Int. Clin. Psychopharmacol. 2005, 20, 173−176.(57) Dumortier, G.; Cabaret, W.; Stamatiadis, L.; Saba, G.;Benadhira, R.; Rocamora, J. F.; Aubriot-Delmas, B.; Glikman, J.;Januel, D. Hepatic tolerance of atypical antipsychotic drugs. Encephale2002, 28, 542−551.(58) Gross, V.; Steube, K.; Tran-Thi, T.; Haussinger, D.; Legler, G.;Decker, K.; Heinrich, P. C.; Gerok, W. The role of N-glycosylation forthe plasma clearance of rat liver secretory glycoproteins. Eur. J.Biochem. 1987, 162, 83−88.(59) Bayer, T. A.; Buslei, R; Havas, L; Falkai, P Evidence foractivation of microglia in patients with psychiatric illnesses. Neurosci.Let. 1999, 271, 126−128.(60) Zhang, X. Y.; Zhou, D. F.; Cao, L. Y.; Zhang, P. Y.; Wu, G. Y.;Shen, Y. C. Changes in serum interleukin-2, -6, and -8 levels beforeand during treatment with risperidone and haloperidol: Relationshipto outcome in schizophrenia. J. Clin. Psychiatry 2004, 65, 940−947.(61) Hou, Y.; Wu, C. F.; Yang, J. Y.; He, X.; Bi, X. L.; Yu, L.; Guo, T.Effects of clozapine, olanzapine and haloperidol on nitric oxideproduction by lipopolysaccharide-activated N9 cells. Prog. Neuro-psychopharmacol. Biol. Psychiatry 2006, 30, 1523−1528.(62) Sugino, H.; Futamura, T.; Mitsumoto, Y.; Maeda, K.; Marunaka,Y. Atypical antipsychotics suppress production of proinflammatorycytokines and up-regulate interleukin-10 in lipopolysaccharide-treatedmice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 303−307.(63) Meyer, J. M.; McEvoy, J. P.; Davis, V. G.; Goff, D. C.; Nasrallah,H. A.; Davis, S. M.; Hsiao, J. K.; Swartz, M. S.; Stroup, T. S.;Lieberman, J. A. Inflammatory Markers in Schizophrenia: Comparingantipsychotic effects in Phase 1 of the clinical antipsychotic trials ofintervention effectiveness study. Biol. Psychiatry 2009, 66, 1013−1022.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr300218h | J. Proteome Res. 2012, 11, 3743−37523752
Related Documents











