ORIGINAL PAPER Antipredator responses of koomal (Trichosurus vulpecula hypoleucus) against introduced and native predators Jennyffer Cruz & Duncan R. Sutherland & Dean P. Anderson & Alistair S. Glen & Paul J. de Tores & Luke K.-P. Leung Received: 3 September 2012 / Revised: 3 April 2013 / Accepted: 13 May 2013 / Published online: 31 May 2013 # Springer-Verlag Berlin Heidelberg 2013 Abstract Antipredator behavior studies generally assess prey responses to single predator species although most real systems contain multiple species. In multi-predator environments prey ideally use antipredator responses that are effective against all predator species, although responses may only be effective against one predator and counterproductive for another. Multi-predator sys- tems may also include introduced predators that the prey did not co-evolve with, so the prey may either fail to recognize their threat (level 1 naiveté), use ineffective responses (level 2 naiveté) or succumb to their superior hunting ability (level 3 naiveté). We analyzed microhab- itat selection of an Australian marsupial (koomal, Trichosurus vulpecula hypoleucus) when faced with spa- tiotemporal differences in the activity/density levels of one native (chuditch, Dasyurus geoffroii) and two intro- duced predators (red fox, Vulpes vulpes; feral cat, Felis catus). From this, we inferred whether koomal recog- nized introduced predators as a threat, and whether they minimized predation risk by either staying close to trees and/or using open or dense microhabitats. Koomal remained close to escape trees regardless of the predator species present, or activity/density levels, suggesting koomal employ this behavior as a first line of defense. Koomal shifted to dense cover only under high risk scenarios (i.e., with multiple predator species present at high densities). When predation risk was low, koomal used open microhabitats, which likely provided benefits not associated with predator avoidance. Koomal did not exhibit level 1 naiveté, although further studies are required to determine if they exhibit higher levels of naiveté (2–3) against foxes and cats. Keywords Brushtail possum . Conditional logistic regression . Habitat selection . Predator–prey interactions . Spool-and-line tracking Introduction Prey species avoid predation using a variety of antipredator responses including vigilance, shifts in habitat use, decreased movement, concealment and grouping. The choice of antipredator behavior depends on prey attributes such as body size and predator attributes such as their hunting techniques (Kats and Dill 1998). For example, small mammals typically prefer dense vegetation cover in response to increased preda- tion risk (e.g., Dickman 1992; Morris and Davidson 2000), whereas larger species usually employ vigilance (e.g., Apfelbach et al. 2005; Lung and Childress 2007) or increased grouping (e.g., Banks 2001). Prey avoiding ambush predators often rely on open habitats (e.g., Korpimäki et al. 1996), alter Communicated by E. Korpimäki Electronic supplementary material The online version of this article (doi:10.1007/s00265-013-1561-2) contains supplementary material, which is available to authorized users. J. Cruz (*) : L. K.<P. Leung School of Agriculture and Food Sciences, University of Queensland, Gatton, QLD 4343, Australia e-mail: [email protected] J. Cruz : D. R. Sutherland : A. S. Glen Invasive Animals Cooperative Research Centre, University of Canberra, Kirinari St, Bruce, ACT 2617, Australia J. Cruz : D. R. Sutherland : A. S. Glen : P. J. de Tores Department of Environment and Conservation, Dwellingup Research Centre, Banksiadale Rd, Dwellingup, WA 6213, Australia J. Cruz : D. P. Anderson : A. S. Glen Landcare Research, PO Box 40, Lincoln 7640, New Zealand D. R. Sutherland Research Department, Phillip Island Nature Parks, PO Box 97, Cowes 3922, Australia Behav Ecol Sociobiol (2013) 67:1329–1338 DOI 10.1007/s00265-013-1561-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Antipredator responses of koomal (Trichosurus vulpeculahypoleucus) against introduced and native predators
Jennyffer Cruz & Duncan R. Sutherland & Dean P. Anderson &
Alistair S. Glen & Paul J. de Tores & Luke K.-P. Leung
Received: 3 September 2012 /Revised: 3 April 2013 /Accepted: 13 May 2013 /Published online: 31 May 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Antipredator behavior studies generally assessprey responses to single predator species although mostreal systems contain multiple species. In multi-predatorenvironments prey ideally use antipredator responsesthat are effective against all predator species, althoughresponses may only be effective against one predatorand counterproductive for another. Multi-predator sys-tems may also include introduced predators that theprey did not co-evolve with, so the prey may either failto recognize their threat (level 1 naiveté), use ineffectiveresponses (level 2 naiveté) or succumb to their superiorhunting ability (level 3 naiveté). We analyzed microhab-itat selection of an Australian marsupial (koomal,Trichosurus vulpecula hypoleucus) when faced with spa-tiotemporal differences in the activity/density levels of
one native (chuditch, Dasyurus geoffroii) and two intro-duced predators (red fox, Vulpes vulpes; feral cat, Feliscatus). From this, we inferred whether koomal recog-nized introduced predators as a threat, and whether theyminimized predation risk by either staying close to treesand/or using open or dense microhabitats. Koomalremained close to escape trees regardless of the predatorspecies present, or activity/density levels, suggestingkoomal employ this behavior as a first line of defense.Koomal shifted to dense cover only under high riskscenarios (i.e., with multiple predator species presentat high densities). When predation risk was low, koomalused open microhabitats, which likely provided benefitsnot associated with predator avoidance. Koomal did notexhibit level 1 naiveté, although further studies arerequired to determine if they exhibit higher levels ofnaiveté (2–3) against foxes and cats.
Keywords Brushtail possum . Conditional logisticregression . Habitat selection . Predator–prey interactions .
Spool-and-line tracking
Introduction
Prey species avoid predation using a variety of antipredatorresponses including vigilance, shifts in habitat use, decreasedmovement, concealment and grouping. The choice ofantipredator behavior depends on prey attributes such as bodysize and predator attributes such as their hunting techniques(Kats and Dill 1998). For example, small mammals typicallyprefer dense vegetation cover in response to increased preda-tion risk (e.g., Dickman 1992; Morris and Davidson 2000),whereas larger species usually employ vigilance (e.g.,Apfelbach et al. 2005; Lung and Childress 2007) or increasedgrouping (e.g., Banks 2001). Prey avoiding ambush predatorsoften rely on open habitats (e.g., Korpimäki et al. 1996), alter
Communicated by E. Korpimäki
Electronic supplementary material The online version of this article(doi:10.1007/s00265-013-1561-2) contains supplementary material,which is available to authorized users.
J. Cruz (*) : L. K.<P. LeungSchool of Agriculture and Food Sciences,University of Queensland, Gatton, QLD 4343, Australiae-mail: [email protected]
J. Cruz :D. R. Sutherland :A. S. GlenInvasive Animals Cooperative Research Centre,University of Canberra, Kirinari St,Bruce, ACT 2617, Australia
J. Cruz :D. R. Sutherland :A. S. Glen : P. J. de ToresDepartment of Environment and Conservation,Dwellingup Research Centre, Banksiadale Rd,Dwellingup, WA 6213, Australia
J. Cruz :D. P. Anderson :A. S. GlenLandcare Research, PO Box 40, Lincoln 7640, New Zealand
D. R. SutherlandResearch Department, Phillip Island Nature Parks, PO Box 97,Cowes 3922, Australia
Behav Ecol Sociobiol (2013) 67:1329–1338DOI 10.1007/s00265-013-1561-2

their activity levels, and/or reduce their travel distances (e.g.,Desy et al. 1990; Fischhoff et al. 2007).
Predator avoidance becomes complicated in multi-predator systems. Predators present may be morphologicallyor behaviorally similar, or prey may effectively employsimilar antipredator strategies against them, so that the pred-ators can be classified as part of the same ‘archetype’ (Coxand Lima 2006). Alternatively, the system may includepredators that utilize widely different hunting techniques(belonging to different archetypes), so that the strategy foravoiding one predator may increase vulnerability to another(Kotler et al. 1993; Sih et al. 1998). For instance, field voles(Microts agrestis) shift their habitat to open areas in thepresence of least weasels (Mustela nivalis) alone; andselect greater cover and decrease their mobility in thepresence of kestrels (Falco tinnunculus) alone(Korpimäki et al. 1996). However, when both predatorsare present, voles are forced into cover, despite thegreater risk of predation from weasels in this habitat. Whileit is widely recognized that most prey species live in multi-predator environments (Lima 1992) antipredator behaviorstudies often assess responses to single predator species (butsee: Dickman 1992;Willems and Hill 2009; Morosinotto et al.2010, for some notable exceptions), which can lead to spuri-ous conclusions.
Predator–prey interactions are further complicated ifprey are naïve to introduced predators they did not co-evolve with (Cox and Lima 2006). For a prey species toavoid a predator, it needs to recognize that predator as athreat. Prey species can display naiveté to introducedpredators by either failing to recognize them as a threat(level 1 naiveté); recognizing them, but using inappro-priate antipredator responses against them (level 2naiveté) or; recognizing them, responding appropriately,but still succumbing to their superior predation (level 3naiveté, Banks and Dickman 2007).
Native Australian mammals have suffered severe impactsdue to the introduction of the red fox (Vulpes vulpes) and theferal cat (Felis catus) (Dickman 1996). This has been partlyattributed to different levels of naiveté by native mammalsto these introduced predators (Cox and Lima 2006; Russelland Banks 2007). Several field studies have assessed thenaiveté of Australian species towards foxes with mixed re-sults (see supplementary material). For example, variousnative species, such as the fawn-footed melomys (Melomyscervinipes) recognised fox odors (Hayes et al. 2006), butothers, including the brown antechinus (Antechinusstuartii), appeared not to respond to fox cues (Russell andBanks 2007). Some species, such as the common brushtailpossum (Thrichosurus vulpecula vulpecula), altered theirbehavior in response to increased fox density (Gresser1996; Pickett et al. 2005), and showed mixed responses tofox odors (Russell and Banks 2005; Nernesian et al. 2012).
Behavioral responses of native species to feral cats havereceived less attention (but see Blumstein et al. 2000;Jones et al. 2004 for captive trials). Furthermore, whilefoxes, cats and native predators generally occur sympatri-cally posing a combination of risks for prey, most studieshave investigated prey responses to each predator inisolation.
This study investigated microhabitat selection by a nativeAustralian marsupial, the koomal (Trichosurus vulpeculahypoleucus), while subject to various levels of predationrisk from two introduced predators (the 3.5–8 kg red foxand the 2.5–6.5 kg feral cat), and one native predator (the 1–2 kg chuditch, Dasyurus geoffroii) in the wild. The koomalis one of the smaller (1–2 kg) subspecies of the nocturnalcommon brushtail possum that inhabit the southwest ofAustralia (Kerle et al. 1991). Cats, foxes and chuditch areknown to consume T. vulpecula (Jones and Coman 1981;Glen et al. 2006, 2010b), and foxes have been shown tolimit koomal populations (Kinnear et al. 2002). Chuditchco-evolved with koomal, are good tree climbers, and haveacute sight and smell (Serena et al. 1991). Feral cats areambush and stalk predators with excellent night visionand good climbing skills (Murray et al. 1995; May andNorton 1996) and have co-existed with koomal since the1840–1850 s (Abbott 2002). Foxes are more recent in-vaders to the southwest of Australia, co-existing withkoomal since the 1930s (Dickman 1996). Foxes rely onsmell and sight to hunt, and make use of open areas torun down their prey (Murray et al. 1995; May and Norton1996).
We examined microhabitat selection by koomal (i.e.,microhabitat use in greater proportion to its availability)in association with spatiotemporal changes in theactivity/density levels of foxes, cats and chuditch. Weassumed that increased selection for a particular micro-habitat would reflect an antipredator behavior if it oc-curred in association with increased predatoractivity/density. We hypothesized that koomal could se-lect three possible microhabitats to minimize their pre-dation risk: (a) remain close to trees which provide anescape route from predators; (b) select ground areaswith dense visual cover, which provide concealmentfrom predators; or (c) select open microhabitats thatfacilitate early detection of approaching predators. Ifmicrohabitats were selected to minimize predation riskthen use of (b) and (c) would be mutually exclusive,while (a) may be used in conjunction with either (b) or(c). We could not predict which microhabitat(s) would beselected by koomal in association with a particular predatorspecies as this depends on two unknowns: (1) the archetypethe predator species belongs to; and (2) the level of naiveté (ifany) that is displayed by koomal in response to the introducedpredators.
1330 Behav Ecol Sociobiol (2013) 67:1329–1338

Methods
Study area and data collection
The antipredator responses use by koomal were assessed atthree geographically independent sites located in the north-ern jarrah forest of Western Australia: Jarrahdale, 116°11′E,32°19′S; George (about 70 km further south), 116°22′E,32°53′S; and Stene (about 100 km south of Jarrahdale),116°31′E, 33°9′S. Sites were selected based on the presenceof koomal, from a survey of 11 sites known to have hadextant populations in 1999 (P. de Tores, unpublished data).The jarrah forest is a dry sclerophyll forest dominated byEucalyptus marginata and Corymbia calophylla, at canopylevel, although E. wandoo, E. rudis and E. patens are alsofound in smaller proportions (Whitford and Stoneman2004). The forest experiences a Mediterranean climate withrainfall gradient from 635 to 1250 mm from east to west(Whitford and Stoneman 2004). The jarrah forest is managedfor conservation, recreation, timber harvesting and mining(Whitford and Williams 2002). Over 290,000 ha of the forestare currently baited to control foxes (Orell 2004). During thestudy, fox baiting was conducted five times a year over an areaencompassing Jarrahdale and George sites (but not Stene)using Probait® sausage style baits each containing 3 mg ofthe toxin 1080 (sodium fluoroacetate).
Koomal were captured at each site during five consecu-tive night-trapping sessions every season, from November2007 to November 2008, using wire cage traps (600×200×200 mm; Sheffield Wire Products, Welshpool, WesternAustralia), as part of a concurrent study examining koomaldemographics (Cruz 2012). Traps were baited with a mix-ture of rolled oats, peanut butter, honey and sardines. Trapswere set at each site using a trapping web arrangement(Lukacs et al. 2005) consisting of eight lines radiating froma center point in cardinal directions, with 12 trap points perline spaced 20 m apart, except for the first trap which was10 m from the center. Individuals were also trapped alongroad networks surrounding the trapping web using 50 cagetraps spaced every 200 m. The total area trapped at each sitehad roughly a 2-km radius. On initial capture, koomal werelightly restrained inside a dark cloth bag and marked with asubcutaneously implanted Passive Integrated Transponder(PIT) tag and a numbered ear-tag.
Determining microhabitat selection by koomal requiredan assessment of whether koomal used (a) close distances totrees, and/or (b) dense cover, or (c) open microhabitat ingreater proportion to the availability of these microhabitatfeatures. We assessed microhabitat use by koomal withspool-and-line tracking. Spool-and-line tracking involvesattaching a spool of thread to an animal, which unravelsfrom the inside as the animal moves away from the point ofrelease, leaving a comprehensive record of the animal’s path
(Miles et al. 1981). Spool-and-line tracking was chosen forits low disturbance to an individual’s behavior, its ease ofuse, low cost, and ability to provide fine-scale detail ofanimal movements in three dimensions (Miles et al. 1981;Key and Woods 1996), making it ideal for assessing habitatuse of a shy semi-arboreal animal. One to four adult koomalwere fitted with a spool device (Sutherland et al. 2008) onlyon the last day of each trapping session to avoid individualsbeing recaptured before the spool unraveled completely. Weassumed that the behavior of trapped individuals was thesame regardless of the number of times they were captured.We initially used 1,000-m-long spools, but changed to 500-m-long spools because we found individuals did not coverdistances greater than 500 m during a single night.
Spool lines were followed by an individual observer theday following the release of each individual koomal. Oncereleased, individuals retreated to a nearby tree or hollow log.Our observations suggested that individuals rarely (if ever)left their initial refuge before nightfall. Thus, we character-ized the level of visual cover and distance to the nearest‘escape’ trees (±0.1 m) used by koomal at points along thespool (referred to as used points), starting at 10 m from theinitial refuge and continuing at 10-m intervals until the endof the thread. When individuals climbed a tree, we contin-ued sampling 10 m from where they returned to the ground(which was often a different from the tree they climbedinitially). The location of each used point was recordedusing a handheld GPS (Garmin etrex®H, ±3–10 m).Escape trees were defined as trees at least 20 mm in diam-eter at breast height over bark, and taller than 2 m. Visualcover was estimated using the Quadrant Cover Method(Glen et al. 2010a), which assesses on a scale of 0–100 thelevel of visual cover provided to an animal by vegetationand other obstacles within a designated radius (in this case,1 m) surrounding the sampling point.
The methodology for assessing available microhabitatrequired careful consideration. There are four general de-signs for sampling available habitat however, not all designsare applicable at microhabitat scales (Manly et al. 2002).Designs I and II measure available habitat at the populationlevel and are therefore inappropriate for microhabitat selec-tion. Design III measures available habitat specific to eachindividual, usually by sampling random points within eachindividual’s home range. However, the entire home range ofan individual does not represent the microhabitat availableto it on a single night if the animal is less mobile, with alarge or poorly defined home range (Compton et al. 2002), asis the case for koomal (home range: male=31 ha, female=16 ha; Cruz et al. 2012). Design IV measures available habitatspecific to each point used by an individual and ideally re-quires multiple control points to be sampled at varying dis-tances surrounding each used (spool) point, which is verylabor intensive.
Behav Ecol Sociobiol (2013) 67:1329–1338 1331

In this study, a design IV approach would have greatlylimited the number of individuals that could be tracked eachseason since each spool took between 1 and 5 h to follow.Consequently, we devised a novel methodology under aDesign III framework to sample the surrounding microhabitatavailable to each individual on the night it was fitted with aspool as follows. Once a spool line was surveyed, its trajectorywas mapped using ArcGIS (version 9.3; ESRI, USA) todetermine the number, length and location of control transectsby firstly drawing four quadrants around the entire spool linewith the start of the spool line representing the center (Fig. 1).The distance from the center to the furthest spool point in eachquadrant represented the length of the control lines. Thenumber of control lines in each quadrant was determined sothat they had a combined number of points (termed ‘availablepoints’) equal to or more than the number of used (spool)points in that quadrant. Available points were sampled at 10-mintervals. The direction of the control transects followed ran-domly generated compass bearings within each quadrant. Nocontrol transects were placed in quadrants that the spool linedid not enter. The microhabitat characteristics along thesecontrol transects were surveyed using the same methods asfor the spool lines.
We sampled predators during the same seasons that koomalwere fitted with spool devices to associate the spatiotemporalchanges in microhabitat selection by koomal to the spatiotem-poral differences in predator activity/density. Chuditch weretrapped for 5 consecutive days every season using 50 cagetraps spaced 200 m apart along existing road networks andbaited with rotting chicken pieces. Individuals were markedon first capture as for koomal. Density estimates were derivedusing the spatially explicit capture–recapture (SECR) models
in program DENSITY 4.4 (Efford 2009). In seasons withinsufficient recaptures to estimate density using SECR, den-sity was estimated as the number of individuals known to bealive divided by the effective trapping area using an 820 mbuffer as calculated in DENSITY 4.4. The buffer was themean maximum distance moved by recaptured individualsacross all sites and seasons.
Unlike chuditch, introduced foxes and cats are not easilytrapped, so we used their activity as a surrogate of theseasonal predation risk they posed to koomal. At each site,foxes and cats were monitored for 5 consecutive days everyseason, from December 2007 to December 2008, using asand plot network of 12 ‘active’ plots (with a meat luresecured to the middle of a 75-cm-long PVC pipe (insidediameter, 240 mm) fitted with doubled sided sticky tapeacross each entrance (attached to a rubber band at approx-imately 12 cm in depth and height) which was used as a haircollection device in an unpublished study), alternating with13 ‘passive’ plots (no lure or hair collection device) alongexisting tracks surrounding the area where koomal weretrapped. Sand plots were 1 m wide and spanned the widthof the track, and were separated by minimum distances of500 m. Plots were monitored daily for hair or footprintswhich could be attributed to either a fox or a cat, for 6consecutive days every season. Their seasonal activity wasthen calculated using a modified General Index (GI) fromEngeman (2005), which replaced the need to count individ-ual prints on each sand plot (termed incursions in Engeman2005) with a binary measure of activity for each sand plot(i.e., whether predator presence was detected or not). Thus:
GI ¼ 1
D
XDd¼1
xdsd;
where D is the total number of monitoring days, xd is thenumber of plots where activity was recorded on the dth day,and sd is the number of plots monitored on the dth day. Thedifference in area sizes between study sites creates varyingsampling effort, which is often corrected by incorporatingan area term into the density equation (e.g., Zellweger-Fischer et al. 2011). We corrected for varying study sitesizes by dividing the seasonal GI by the area of the sandplot network (using a convex polygon with a 500-m buffer)to obtain a seasonal index of predator activity at each site,which was used as a surrogate of the predation risk posed byeach species.
Modeling koomal’s antipredator responses
A multi-stage Bayesian approach was used for its flexibilityto tailor models to suit the data rather than having to modifythe data to fit frequentist statistical frameworks (Clark andLaDeau 2006). In the first stage of the model, we used a
Fig. 1 Schematic diagram of how available habitat (control lines) wasdetermined for each spool line (bold line). The distance from the startof the spool line (centre of the quadrants) to the furthest spool point ineach quadrant (dashed line) determined the length of the control lines.The total number of available points (grey circles) had to at least equalthe number of spool points (black circles) in each quadrant. Thedirection of the control lines (grey lines) followed randomly generatedcompass bearings
1332 Behav Ecol Sociobiol (2013) 67:1329–1338

resource selection framework (Manly et al. 2002) to modelthe event (zi,j*,s,t) that an individual koomal i selected tomove to a used point j* among a set (J) of available points atsite s and spooling event t. The relative probability of theobserved event (i.e., movement to a used point) was condi-tioned on available points (J) and the used point’s locationin relation to other used points in the set (N):
Pr zi; j�;s;t���J ;Nh i
¼exp μi; j�;s;t
� �X
j∈ Jexp μi; j;s;t
� � ;
where μij,s,t is the relative suitability of point j. This is a case-controlled logistic-regression likelihood (Hosmer andLemeshow 2000), which accounts for repeated sampling ofindividual koomal and incorporates used points as j* (the cases)and j≠j* as the available points for each spool. The linearprediction of μi,j,s,t is a function of environmental covariatesin site s.
μi; j;s;t ¼ β1;i;s;tTreeDist j þ β2;i;s;tCover j þ β3Neighbor j;
where TreeDistj is the distance from point j to the nearest escapetree, Coverj is the measure of visual cover, Neighborj is a binaryvariable indicating whether point j was neighboring anotherused point from the same spool, and β1,i,s,t, β2,i,s,t and β3 are theassociated coefficients for each covariate. The variableNeighborj is included to control for the inherent autocorrelationin the relative probability of selection due to identifying usedlocations every 10 m along a spool line (Fig. 1). The coefficientβ3 accounts for the strength of this spatial dependence in themodel. The coefficients β1,i,s,t and β2,i,s,t quantify the relativestrength of selection for or against escape trees and cover,respectively, and were expected to change with varying preda-tor activities/densities across sites s and over spooling events t.By examining how these coefficients differed over space andtime in relation to predator activity/density levels, we madeinferences on koomal’s habitat selection for: (a) close distancesto escape trees, and/or (b) dense or (c) open cover. The β1,i,s,tand β2,i,s,t were modeled as latent coefficients in the secondstage of the overall model:
β1;i;s;t ¼ α0 þ α1Foxs;t þ α2Cats;t þ α3Chuditchs;t þ gs þ εi;tβ2;i;s;t ¼ δ0 þ δ1Foxs;t þ δ2Cats;t þ δ3Chuditchs;t þ ls þ ei;t
εi;t e Normal 0; σ2� �
ei;t e Normal 0;π2� �
;
where Foxs,t, Cats,t, and Chuditchs,t denote the predationactivity/density covariates; α0–3 and δ0–3 are the interceptsand covariate coefficients; gs and ls are random site effects;and εi,t and ei,t are over-dispersion terms. The predator covar-iates were scaled to have a mean of zero and a standarddeviation (SD) of one to facilitate direct comparisons of theireffects. A sum-to-zero constraint was imposed on random siteeffects (gs and ls), which were included to account for inter-site
variability not included in the model (Gillies et al. 2006;Duchesne et al. 2010). The over-dispersion terms (εi,t and ei,t)were assumed to be independent and identically distributedwith variance σ2 and π2, respectively.
Bayesian inference was derived from Markov chainMonte Carlo analysis by iteratively updating prior parame-ters in the first- and second-stage models. The global modelfor the posterior distributions for unknowns given covariatesX was:
p β;α; δ; g; l;σ;π���X� �
∝∏Ii¼1∏
Tt¼1Pr zi; j;s;t
���J ;Nh i�Normal β1;i;s;t
���X 0αþ gs;σ
2� �
�Normal β2;i;s;t
���X 0δ þ ls; p2
� ��∏A
a¼1Normal αa
���0; 1000� ��∏D
d¼1Normal δd���0; 1000� �
�∏3s¼1Normal γs
���0; 1000� ��∏3
s¼1Normal λs
���0; 1000� ��InverseGamma σ2
���0:1; 0:1� ��InverseGamma π2
���0:1; 0:1� �
The posteriors of first stage coefficients (β1,i,s,t, β2,i,s,t,and β3) were estimated using a Metropolis algorithm fol-lowing Clark (2007, pp 175–177). All second-stage coeffi-cients were sampled directly from the conditional posteriors(Clark 2007).
The contributions of the predator covariates and siterandom intercepts to overall model fit were examined bycomparing models using the Deviance Information Criterion(DIC), (Spiegelhalter et al. 2002). Biologically plausiblemodels were compared from a set that included the globalmodel (incorporating all variables) and reduced models thatexcluded non-significant variables (with Bayesian credibleintervals [CIs] overlapping zero; Thomas et al. 2006).Models with a DIC difference (ΔDIC) of less than 2 fromthe lowest DIC model were strongly supported, whilemodels with ΔDIC of more than ten had little support(Clark 2007). Models were fitted in R software (version2.11.1, http://cran.r-project.org) using mvnorm and CODApackages (Carlin et al. 2006).
Model convergence was assessed with Geweke–Brooksplots (Geweke 1992) and Heidelberger and Welch (1983)statistics, calculated for each covariate. Convergence wasachieved relatively quickly, so the first 20,000 iterationswere discarded as a burn in period for all models.Posterior summaries were thus taken from further 100,000iterations, thinned by 10, for a total of 10,000 iterations.Using every 10th iteration adequately minimized autocorre-lation based on autocorrelation lag plots.
Behav Ecol Sociobiol (2013) 67:1329–1338 1333
Results
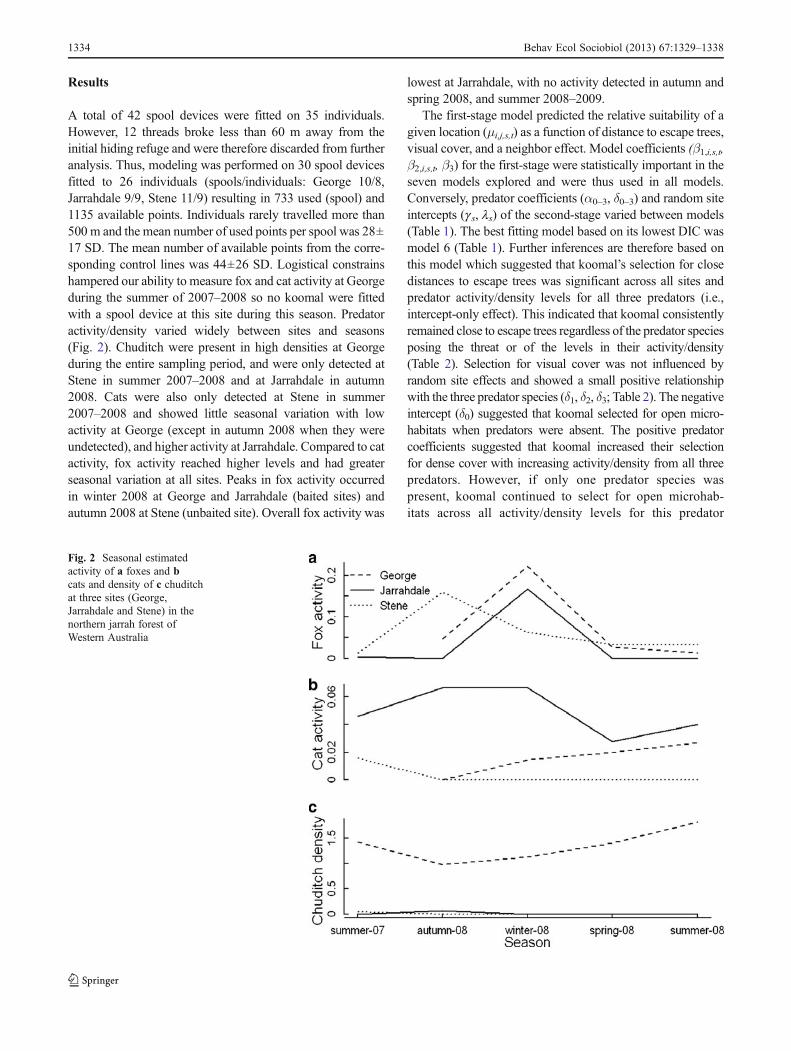
A total of 42 spool devices were fitted on 35 individuals.However, 12 threads broke less than 60 m away from theinitial hiding refuge and were therefore discarded from furtheranalysis. Thus, modeling was performed on 30 spool devicesfitted to 26 individuals (spools/individuals: George 10/8,Jarrahdale 9/9, Stene 11/9) resulting in 733 used (spool) and1135 available points. Individuals rarely travelled more than500 m and the mean number of used points per spool was 28±17 SD. The mean number of available points from the corre-sponding control lines was 44±26 SD. Logistical constrainshampered our ability to measure fox and cat activity at Georgeduring the summer of 2007–2008 so no koomal were fittedwith a spool device at this site during this season. Predatoractivity/density varied widely between sites and seasons(Fig. 2). Chuditch were present in high densities at Georgeduring the entire sampling period, and were only detected atStene in summer 2007–2008 and at Jarrahdale in autumn2008. Cats were also only detected at Stene in summer2007–2008 and showed little seasonal variation with lowactivity at George (except in autumn 2008 when they wereundetected), and higher activity at Jarrahdale. Compared to catactivity, fox activity reached higher levels and had greaterseasonal variation at all sites. Peaks in fox activity occurredin winter 2008 at George and Jarrahdale (baited sites) andautumn 2008 at Stene (unbaited site). Overall fox activity was
lowest at Jarrahdale, with no activity detected in autumn andspring 2008, and summer 2008–2009.
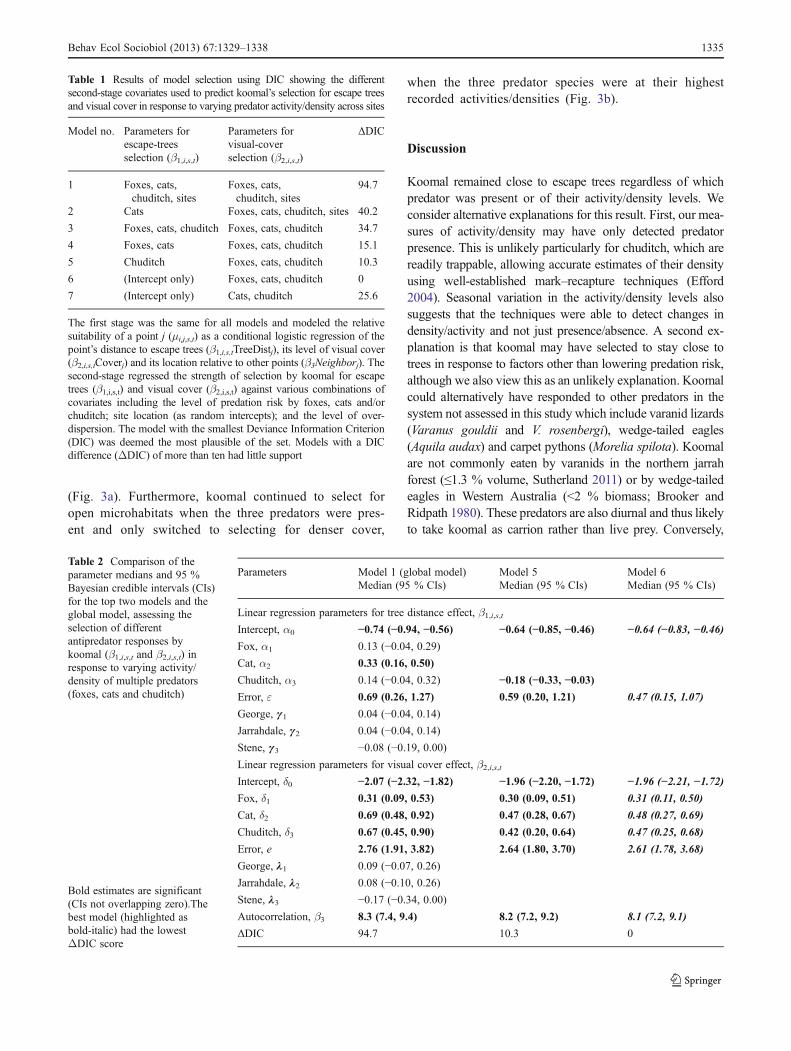
The first-stage model predicted the relative suitability of agiven location (μi,j,s,t) as a function of distance to escape trees,visual cover, and a neighbor effect. Model coefficients (β1,i,s,t,β2,i,s,t, β3) for the first-stage were statistically important in theseven models explored and were thus used in all models.Conversely, predator coefficients (α0–3, δ0–3) and random siteintercepts (gs, ls) of the second-stage varied between models(Table 1). The best fitting model based on its lowest DIC wasmodel 6 (Table 1). Further inferences are therefore based onthis model which suggested that koomal’s selection for closedistances to escape trees was significant across all sites andpredator activity/density levels for all three predators (i.e.,intercept-only effect). This indicated that koomal consistentlyremained close to escape trees regardless of the predator speciesposing the threat or of the levels in their activity/density(Table 2). Selection for visual cover was not influenced byrandom site effects and showed a small positive relationshipwith the three predator species (δ1, δ2, δ3; Table 2). The negativeintercept (δ0) suggested that koomal selected for open micro-habitats when predators were absent. The positive predatorcoefficients suggested that koomal increased their selectionfor dense cover with increasing activity/density from all threepredators. However, if only one predator species waspresent, koomal continued to select for open microhab-itats across all activity/density levels for this predator
Fig. 2 Seasonal estimatedactivity of a foxes and bcats and density of c chuditchat three sites (George,Jarrahdale and Stene) in thenorthern jarrah forest ofWestern Australia
1334 Behav Ecol Sociobiol (2013) 67:1329–1338
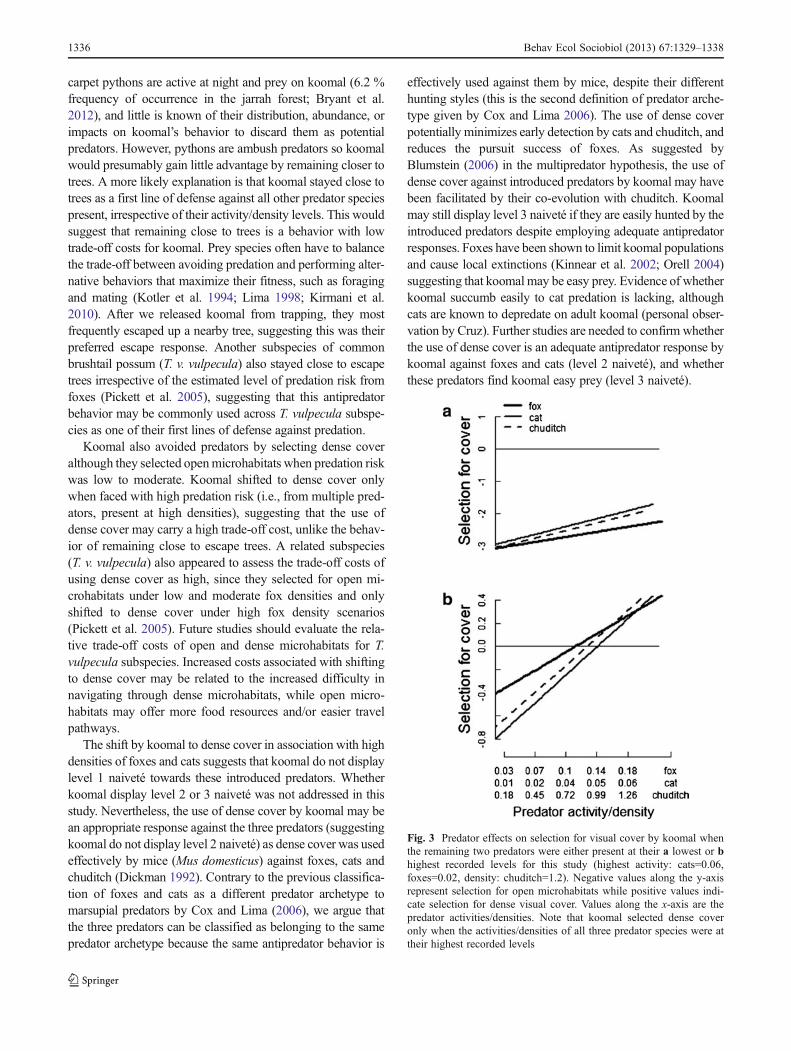
(Fig. 3a). Furthermore, koomal continued to select foropen microhabitats when the three predators were pres-ent and only switched to selecting for denser cover,
when the three predator species were at their highestrecorded activities/densities (Fig. 3b).
Discussion
Koomal remained close to escape trees regardless of whichpredator was present or of their activity/density levels. Weconsider alternative explanations for this result. First, our mea-sures of activity/density may have only detected predatorpresence. This is unlikely particularly for chuditch, which arereadily trappable, allowing accurate estimates of their densityusing well-established mark–recapture techniques (Efford2004). Seasonal variation in the activity/density levels alsosuggests that the techniques were able to detect changes indensity/activity and not just presence/absence. A second ex-planation is that koomal may have selected to stay close totrees in response to factors other than lowering predation risk,although we also view this as an unlikely explanation. Koomalcould alternatively have responded to other predators in thesystem not assessed in this study which include varanid lizards(Varanus gouldii and V. rosenbergi), wedge-tailed eagles(Aquila audax) and carpet pythons (Morelia spilota). Koomalare not commonly eaten by varanids in the northern jarrahforest (≤1.3 % volume, Sutherland 2011) or by wedge-tailedeagles in Western Australia (<2 % biomass; Brooker andRidpath 1980). These predators are also diurnal and thus likelyto take koomal as carrion rather than live prey. Conversely,
Table 1 Results of model selection using DIC showing the differentsecond-stage covariates used to predict koomal’s selection for escape treesand visual cover in response to varying predator activity/density across sites
Model no. Parameters forescape-treesselection (β1,i,s,t)
Parameters forvisual-coverselection (β2,i,s,t)
ΔDIC
1 Foxes, cats,chuditch, sites
Foxes, cats,chuditch, sites
94.7
2 Cats Foxes, cats, chuditch, sites 40.2
3 Foxes, cats, chuditch Foxes, cats, chuditch 34.7
4 Foxes, cats Foxes, cats, chuditch 15.1
5 Chuditch Foxes, cats, chuditch 10.3
6 (Intercept only) Foxes, cats, chuditch 0
7 (Intercept only) Cats, chuditch 25.6
The first stage was the same for all models and modeled the relativesuitability of a point j (μi,j,s,t) as a conditional logistic regression of thepoint’s distance to escape trees (β1,i,s,tTreeDistj), its level of visual cover(β2,i,s,tCoverj) and its location relative to other points (β3Neighborj). Thesecond-stage regressed the strength of selection by koomal for escapetrees (β1,i,s,t) and visual cover (β2,i,s,t) against various combinations ofcovariates including the level of predation risk by foxes, cats and/orchuditch; site location (as random intercepts); and the level of over-dispersion. The model with the smallest Deviance Information Criterion(DIC) was deemed the most plausible of the set. Models with a DICdifference (ΔDIC) of more than ten had little support
Table 2 Comparison of theparameter medians and 95 %Bayesian credible intervals (CIs)for the top two models and theglobal model, assessing theselection of differentantipredator responses bykoomal (β1,i,s,t and β2,i,s,t) inresponse to varying activity/density of multiple predators(foxes, cats and chuditch)
Bold estimates are significant(CIs not overlapping zero).Thebest model (highlighted asbold-italic) had the lowestΔDIC score
Parameters Model 1 (global model) Model 5 Model 6Median (95 % CIs) Median (95 % CIs) Median (95 % CIs)
Linear regression parameters for tree distance effect, β1,i,s,tIntercept, α0 −0.74 (−0.94, −0.56) −0.64 (−0.85, −0.46) −0.64 (−0.83, −0.46)Fox, α1 0.13 (−0.04, 0.29)
Cat, α2 0.33 (0.16, 0.50)
Chuditch, α3 0.14 (−0.04, 0.32) −0.18 (−0.33, −0.03)Error, ε 0.69 (0.26, 1.27) 0.59 (0.20, 1.21) 0.47 (0.15, 1.07)
George, g1 0.04 (−0.04, 0.14)
Jarrahdale, g2 0.04 (−0.04, 0.14)
Stene, g3 −0.08 (−0.19, 0.00)
Linear regression parameters for visual cover effect, β2,i,s,tIntercept, δ0 −2.07 (−2.32, −1.82) −1.96 (−2.20, −1.72) −1.96 (−2.21, −1.72)Fox, δ1 0.31 (0.09, 0.53) 0.30 (0.09, 0.51) 0.31 (0.11, 0.50)
Cat, δ2 0.69 (0.48, 0.92) 0.47 (0.28, 0.67) 0.48 (0.27, 0.69)
Chuditch, δ3 0.67 (0.45, 0.90) 0.42 (0.20, 0.64) 0.47 (0.25, 0.68)
Error, e 2.76 (1.91, 3.82) 2.64 (1.80, 3.70) 2.61 (1.78, 3.68)
George, l1 0.09 (−0.07, 0.26)
Jarrahdale, l2 0.08 (−0.10, 0.26)
Stene, l3 −0.17 (−0.34, 0.00)
Autocorrelation, β3 8.3 (7.4, 9.4) 8.2 (7.2, 9.2) 8.1 (7.2, 9.1)
ΔDIC 94.7 10.3 0
Behav Ecol Sociobiol (2013) 67:1329–1338 1335
carpet pythons are active at night and prey on koomal (6.2 %frequency of occurrence in the jarrah forest; Bryant et al.2012), and little is known of their distribution, abundance, orimpacts on koomal’s behavior to discard them as potentialpredators. However, pythons are ambush predators so koomalwould presumably gain little advantage by remaining closer totrees. A more likely explanation is that koomal stayed close totrees as a first line of defense against all other predator speciespresent, irrespective of their activity/density levels. This wouldsuggest that remaining close to trees is a behavior with lowtrade-off costs for koomal. Prey species often have to balancethe trade-off between avoiding predation and performing alter-native behaviors that maximize their fitness, such as foragingand mating (Kotler et al. 1994; Lima 1998; Kirmani et al.2010). After we released koomal from trapping, they mostfrequently escaped up a nearby tree, suggesting this was theirpreferred escape response. Another subspecies of commonbrushtail possum (T. v. vulpecula) also stayed close to escapetrees irrespective of the estimated level of predation risk fromfoxes (Pickett et al. 2005), suggesting that this antipredatorbehavior may be commonly used across T. vulpecula subspe-cies as one of their first lines of defense against predation.
Koomal also avoided predators by selecting dense coveralthough they selected openmicrohabitats when predation riskwas low to moderate. Koomal shifted to dense cover onlywhen faced with high predation risk (i.e., from multiple pred-ators, present at high densities), suggesting that the use ofdense cover may carry a high trade-off cost, unlike the behav-ior of remaining close to escape trees. A related subspecies(T. v. vulpecula) also appeared to assess the trade-off costs ofusing dense cover as high, since they selected for open mi-crohabitats under low and moderate fox densities and onlyshifted to dense cover under high fox density scenarios(Pickett et al. 2005). Future studies should evaluate the rela-tive trade-off costs of open and dense microhabitats for T.vulpecula subspecies. Increased costs associated with shiftingto dense cover may be related to the increased difficulty innavigating through dense microhabitats, while open micro-habitats may offer more food resources and/or easier travelpathways.
The shift by koomal to dense cover in association with highdensities of foxes and cats suggests that koomal do not displaylevel 1 naiveté towards these introduced predators. Whetherkoomal display level 2 or 3 naiveté was not addressed in thisstudy. Nevertheless, the use of dense cover by koomal may bean appropriate response against the three predators (suggestingkoomal do not display level 2 naiveté) as dense cover was usedeffectively by mice (Mus domesticus) against foxes, cats andchuditch (Dickman 1992). Contrary to the previous classifica-tion of foxes and cats as a different predator archetype tomarsupial predators by Cox and Lima (2006), we argue thatthe three predators can be classified as belonging to the samepredator archetype because the same antipredator behavior is
effectively used against them by mice, despite their differenthunting styles (this is the second definition of predator arche-type given by Cox and Lima 2006). The use of dense coverpotentially minimizes early detection by cats and chuditch, andreduces the pursuit success of foxes. As suggested byBlumstein (2006) in the multipredator hypothesis, the use ofdense cover against introduced predators by koomal may havebeen facilitated by their co-evolution with chuditch. Koomalmay still display level 3 naiveté if they are easily hunted by theintroduced predators despite employing adequate antipredatorresponses. Foxes have been shown to limit koomal populationsand cause local extinctions (Kinnear et al. 2002; Orell 2004)suggesting that koomal may be easy prey. Evidence of whetherkoomal succumb easily to cat predation is lacking, althoughcats are known to depredate on adult koomal (personal obser-vation by Cruz). Further studies are needed to confirm whetherthe use of dense cover is an adequate antipredator response bykoomal against foxes and cats (level 2 naiveté), and whetherthese predators find koomal easy prey (level 3 naiveté).
Fig. 3 Predator effects on selection for visual cover by koomal whenthe remaining two predators were either present at their a lowest or bhighest recorded levels for this study (highest activity: cats=0.06,foxes=0.02, density: chuditch=1.2). Negative values along the y-axisrepresent selection for open microhabitats while positive values indi-cate selection for dense visual cover. Values along the x-axis are thepredator activities/densities. Note that koomal selected dense coveronly when the activities/densities of all three predator species were attheir highest recorded levels
1336 Behav Ecol Sociobiol (2013) 67:1329–1338

Most antipredator studies assess single behaviorsand/or single predators even though most vertebrateprey must employ multiple antipredator responsesagainst multiple predators. Here, we evaluated theantipredator behaviors used by prey in multi-predatorsystems using multi-stage conditional logistic models.The use of a resource selection framework (Manly etal. 2002) as the first stage of the model makes thetechnique particularly useful for studies assessingantipredator behaviors involving resource selection inresponse to one or multiple predators. In this study,the multi-stage approach allowed us to partition theeffects of different predators on selection by prey forvisual cover and distance to trees. This framework alsofacilitated the inclusion of random effects to account foradditional site-specific variability and a ‘neighbor’ effectto control for the autocorrelation inherent in the sam-pling technique, and also common to other techniquessuch as radio-tracking. Bayesian multi-stage, conditionallogistic regression is a flexible and powerful tool forevaluating habitat choices by prey living in multi-predatorenvironments.
Overall, results from this study indicate that koomalremained close to escape trees and used dense cover toavoid foxes, cats and chuditch. Remaining close to treesappeared to carry a low trade-off cost for koomal and maybe used as a first line of defense against all predator speciesand activity/density levels. Conversely, the use of densecover probably carried high trade-off costs, as koomal onlyused this avoidance behavior under high risk scenarios (i.e.,with multiple predator species present at high densities).When predation risk was low, koomal appeared to preferthe use of open microhabitats instead, which likelypresented koomal with benefits not associated with predatoravoidance, such as easier travel pathways and/or food re-sources. Results from this study also indicate that koomaldid not exhibit level 1 naiveté against foxes or cats althoughfurther studies are required to determine if they exhibithigher levels of naiveté (2–3) against these introducedpredators.
Acknowledgments J. Cruz was supported by an AustralianPostgraduate Award and an Invasive Animals CRC (IA-CRC)Scholarship. D. Sutherland and A. Glen were supported by IA-CRCPostdoctoral Fellowships. Research funds were provided by theWestern Australian Department of Environment and Conservation(DEC) and the IA-CRC. Procedures were carried out in accordancewith animal ethics permit CAEC/54/2006 (DEC). The authors wish tothank S. Garretson, R. Hill, L. Strümpher, DEC volunteers and inparticular L. Bloomfield, for technical assistance as well as R.P.Pech, A.D. Latham and M.C. Latham for useful contributions to themanuscript prior to submission.
Ethical standards We hereby declare that the present study complieswith the ethical standards of animal research in Australia.
References
Abbott I (2002) Origin and spread of the cat, Felis catus, on mainlandAustralia, with a discussion of the magnitude of its early impacton native fauna. Wildlife Res 29:51–74
Apfelbach R, Blanchard CD, Blanchard RJ, Hayes RA, McGregor IS(2005) The effects of predator odors in mammalian prey species: areview of field and laboratory studies. Neurosci Biobehav R29:1123–1144
Banks PB (2001) Predation-sensitive grouping and habitat use byeastern grey kangaroos: a field experiment. Anim Behav61:1013–1021
Banks PB, Dickman CR (2007) Alien predation and the effects ofmultiple levels of prey naiveté. Trends Ecol Evol 22:229–230
Blumstein DT (2006) The multipredator hypothesis and the evolu-tionary persistence of antipredator behavior. Ethology 112:209–217
Blumstein DT, Daniel JC, Griffin AS, Evans CS (2000) Insular tammarwallabies (Macropus eugenii) respond to visual but not acousticcues from predators. Behav Ecol 11:528–535
Brooker MG, Ridpath MG (1980) The diet of the wedge-tailed eagle,Aquila audax, in Western Australia. Aust Wildl Res 7:433–452
Bryant GL, de Tores PJ, Warren KW, Fleming PA (2012) Does bodysize influence thermal biology and therefore diet of a python(Morelia spilota imbricata)? Aust Ecol 37:583–591
Carlin BP, Clark JS, Gelfand AE (2006) Elements of hierarchicalBayesian inference. In: Clark JS, Gelfand AE (eds) Hierarchicalmodelling for the environmental sciences: statistical methods andapplications. Oxford University Press, New York, pp 3–24
Clark JS (2007) Models for ecological data: an introduction. PrincetonUniversity Press, New Jersey
Clark JS, LaDeau S (2006) Synthesizing ecological experiments andobservational data with hierarchical Bayes. In: Clark JS, GelfandAE (eds) Hierarchical modelling for the environmental sciences:statistical methods and applications. Oxford Biology, New York,pp 41–58
Compton BW, Rhymer JM, McCollough M (2002) Habitat selectionby wood turtles (Clemmys insculpta): an application of pairedlogistic regression. Ecology 83:833–843
Cox JG, Lima SL (2006) Naiveté and an aquatic–terrestrial dichotomy inthe effects of introduced predators. Trends Ecol Evol 21:674–680
Cruz J (2012) Ecology of the koomal (Trichosurus vulpecula hypoleucus)in the northern jarrah forest in relation to predation and resourceavailability. PhD dissertation, University of Queensland
Cruz J, Sutherland DR, Leung LK-P (2012) Den use, home range andterritoriality of the koomal (Trichosurus vulpecula hypoleucus)and implications for current forest management strategies. Aust JZool 60:141–151
Desy EA, Batzli GO, Liu J (1990) Effects of food and predation onbehaviour of prairie voles: a field experiment. Oikos 58:159–168
Dickman CR (1992) Predation and habitat shift in the house mouse,Mus domesticus. Ecology 73:313–322
Dickman CR (1996) Impact of exotic generalist predators on the nativefauna of Australia. Wildlife Biol 2:185–195
Duchesne T, Fortin D, Courbin N (2010) Mixed conditional logisticregression for habitat selection studies. J Anim Ecol 79:548–555
Efford M (2004) Density estimation in live-trapping studies. Oikos106:598–610
Efford MG (2009) Density 4.4: software for spatially explicit capture–recapture. Department of Zoology, University of Otago, Dunedin
Engeman RM (2005) Indexing principles and a widely applicableparadigm for indexing animal populations. Wildl Res 32:203–210
Fischhoff IR, Sundaresan SR, Cordingley J, Rubenstein DI (2007)Habitat use and movements of plains zebra (Equus burchelli) inresponse to predation danger from lions. Behav Ecol 18:725–729
Behav Ecol Sociobiol (2013) 67:1329–1338 1337

Geweke J (1992) Evaluating the accuracy of sampling-based ap-proaches to calculating posterior moments. In: Bernardo JM,Berger JO, Dawid AP, Smith AFM (eds) Bayesian statistics 4.Oxford University Press, London, pp 169–194
Gillies CS, Hebblewhite M, Nielsen S, Krawchuk MA, Aldridge CL,Frair JL, Saher DJ, Stevens CE, Jerde CL (2006) Application ofrandom effects to the study of resource selection by animals. JAnim Ecol 75:887–898
Glen AS, Fay AR, Dickman CR (2006) Diets of sympatric red foxesVulpes vulpes and wild dogs Canis lupus in the Northern RiverRegion, New South Wales. Aust Mammal 28:101–104
Glen AS, Sutherland DR, Cruz J (2010a) An improved method ofmicrohabitat assessment relevant to predation risk. Ecol Res25:311–314
Glen AS, Wayne A, Maxwell M, Cruz J (2010b) Comparative diets ofchuditch, a threatened marsupial carnivore, in the northern andsouthern jarrah forests, Western Australia. J Zool 282:276–283
Gresser S (1996) Anti-predator behaviour in the common brushtailpossum. B.Sc. (Hons) dissertation, University of Sydney
Hayes RA, Nahrung HF, Wilson JC (2006) The response of nativeAustralian rodents to predator odours varies seasonally: a by-product of life history variation? Anim Behav 71:1307–1314
Heidelberger P, Welch PD (1983) Simulation run length control in thepresence of an initial transient. Oper Res 31:1109–1144
Hosmer DW, Lemeshow S (2000) Applied logistic regression, 2nd edn.John Wiley & Sons, New York
Jones E, Coman BJ (1981) Ecology of the feral cat, Felis catus (L.), inSouth-Eastern Australia: I. Diet. Aust Wildl Res 8:537–547
Jones ME, Smith GC, Jones SM (2004) Is anti-predator behaviour inTasmanian eastern quolls (Dasyurus viverrinus) effective againstintroduced predators? Anim Conserv 7:155–160
Kats LB, Dill LM (1998) The scent of death: chemosensory assessmentof predation risk by prey animals. Ecoscience 5:361–394
Kerle JA, McKay GM, Sharman GB (1991) A systematic analysis ofbrushtail possum, Trichosurus vulpecula (Kerr, 1792) (Marsupialia:Phalangeridae). Aust J Zool 39:313–331
Key GE, Woods RD (1996) Spool and line studies on the behaviouralecology of rats (Rattus spp) in the Galapagos Islands. Can J Zool74:733–737
Kinnear JE, Sumner NR, Onus ML (2002) The red fox inAustralia—an exotic predator turned biocontrol agent. BiolConserv 108:335–359
Kirmani SN, Banks PB, McArthur C (2010) Integrating the costs ofplant toxins and predation risk in foraging decisions of a mam-malian herbivore. Oecologia 164:349–356
Korpimäki E, Koivunen V, Hakkarainen H (1996) Microhabitat useand behavior of voles under weasel and raptor predation risk:predator facilitation? Behav Ecol 7:30–34
Kotler BP, Brown JS, Slotow RH, Goodfriend WL, Michal S (1993)The influence of snakes on the foraging behavior of gerbils. Oikos67:309–316
Kotler BP, Brown JS, Mitchell WA (1994) The role of predation inshaping the behaviour, morphology and community organisationof desert rodents. Aust J Zool 42:449–466
Lima SL (1992) Life in a multi-predator environment: some consider-ations for anti-predatory vigilance. Ann Zool Fenn 29:217–226
Lima SL (1998) Nonlethal effects in the ecology of predator–preyinteractions. Bioscience 48:25–34
Lukacs PM, Anderson DR, Burnham KP (2005) Evaluation oftrapping-web designs. Wildl Res 32:103–110
Lung MA, Childress MJ (2007) The influence of conspecifics andpredation risk on the vigilance of elk (Cervus elaphus) inYellowstone National Park. Behav Ecol 18:12–20
Manly B, McDonald L, Thomas D, McDonald T, Erickson W (2002)Resource selection by animals: statistical design and analysis forfield studies, 2nd edn. Kluwer, Boston
May SA, Norton TW (1996) Influence of fragmentation and distur-bance on the potential impact of feral predators on native fauna inAustralian forest ecosystems. Wildl Res 23:387–400
Miles MA, Desouza AA, Povoa MM (1981) Mammal tracking andnest location in brazilian forest with an improved spool-and-linedevice. J Zool 195:331–347
Morosinotto C, Thomson RL, Korpimäki E (2010) Habitat selection asan antipredator behaviour in a multi-predator landscape: all ene-mies are not equal. J Anim Ecol 79:327–333
Morris DW, Davidson DL (2000) Optimally foraging mice match patchuse with habitat differences in fitness. Ecology 81:2061–2066
Murray DL, Boutin S, O'Donoghue M, Nams VO (1995) Huntingbehaviour of a sympatric felid and canid in relation to vegetativecover. Anim Behav 50:1203–1210
Nernesian CL, Banks PB, McArthur C (2012) Behavioural responsesto indirect and direct predator cues by a mammalian herbivore, thecommon brushtail possum. Behav Ecol Sociobiol 66:47–55
Orell P (2004) Fauna monitoring and staff training: Western Shieldreview — February 2003. Conserv Sci WA 5:51–95
Pickett KN, Hik DS, Newsome AE, Pech RP (2005) The influence ofpredation risk on foraging behaviour of brushtail possums inAustralian woodlands. WildlRes 32:121–130
Russell BG, Banks PB (2005) Responses of four Critical Weight Range(CWR) marsupials to the odours of native and introduced preda-tors. Aust Zool 33:217–222
Russell BG, Banks PB (2007) Do Australian small mammals respondto native and introduced predator odours? Aust Ecol 32:277–286
Serena M, Soderquist TR, Morris K (1991) The chuditch (Dasyurusgeoffroii). Department of Conservation and LandManagement, Como
Sih A, Englund G, Wooster D (1998) Emergent impacts of multiplepredators on prey. Trends Ecol Evol 13:350–355
Spiegelhalter DJ, Best NG, Carlin BP, van der Linde A (2002)Bayesian measures of model complexity and fit. J R Stat Soc B64:583–616
Sutherland DR (2011) Dietary niche overlap and size partitioning insympatric varanid lizards. Herpetologica 67:42–49
Sutherland DR, Glen AS, Cruz J (2008) An alternative spool-and-linedevice for medium sized animals. Aust Mammal 30:89–90
Thomas DL, Johnson D, Griffith B (2006) A bayesian random effectsdiscrete-choice model for resource selection: population-levelselection inference. J Wildl Manag 70:404–412
Whitford KR, Stoneman G (2004) Management of tree hollows in thejarrah Eucalyptus marginata forest of Western Australia. In:Lunney D (ed) Conservation of Australia's forest fauna. RoyalZoological Society of New South Wales, Mosman, pp 807–829
Whitford KR, Williams MR (2002) Hollows in jarrah (Eucalyptusmarginata) and marri (Corymbia calophylla) trees: II. Selecting treesto retain for hollow dependent fauna. For Ecol Manag 160:215–232
Willems EP, Hill RA (2009) Predator-specific landscapes of fear andresource distribution: effects on spatial range use. Ecology90:546–555
Zellweger-Fischer J, Kéry M, Pasinelli G (2011) Population trends ofbrown hares in Switzerland: the role of land-use and ecologicalcompensation areas. Biol Conserv 144:1364–1373
1338 Behav Ecol Sociobiol (2013) 67:1329–1338
Related Documents











